

Abstract
Aflatoxins count to the most toxic known mycotoxins and are a threat to food safety especially in regions with a warm and humid climate. Contaminated food reaches consumers globally due to international trade, leading to stringent regulatory limits of aflatoxins in food. While the formation of aflatoxin (AF) B1 by the filamentous fungus Aspergillus flavus is well investigated, less is known about the formation kinetics of its precursors and further aflatoxins. In this study, autoclaved maize kernels were inoculated with A. flavus and incubated at 25 °C for up to 10 days. Aflatoxins and precursors were analyzed by a validated UHPLC-MS method. Additional to AFB1 and AFB2, AFM1 and AFM2 were detected, confirming the ability of the formation of M-group aflatoxins on cereals by A. flavus. The measured relative levels of AFB2, AFM1, and AFM2 on maize compared to the level of AFB1 (mean of days 5, 7, and 10 of incubation) were 3.3%, 1.5%, and 0.2%, respectively. The occurrence and kinetics of the measured aflatoxins and their precursors sterigmatocystin, O-methylsterigmatocystin, 11-hydroxy-O-methylsterigmatocystin, aspertoxin, and 11-hydroxyaspertoxin (group 1) as well as of dihydrosterigmatocystin and dihydro-O-methylsterigmatocystin (group 2) supported the so far postulated biosynthetic pathway. Remarkable high levels of O-methylsterigmatocystin and aspertoxin (17.4% and 4.9% compared to AFB1) were found, raising the question about the toxicological relevance of these intermediates. In conclusion, based on the study results, the monitoring of O-methylsterigmatocystin and aspertoxin as well as M-group aflatoxins in food is recommended.
Supplementary information
The online version contains supplementary material available at 10.1007/s12550-022-00452-4.
Keywords: Aspergillus flavus, Aflatoxins, Aspertoxin, O-Methylsterigmatocystin, Food safety
Introduction
Aflatoxins (AFs) are secondary metabolites of filamentous fungi, especially produced by Aspergillus species like A. flavus, A. minisclerotigenes, and A. parasiticus, which count to the most serious fungal contaminants of food and feed (Coppock et al. 2018). AFs comprise different compounds, including aflatoxin B1 (AFB1), B2 (AFB2), G1 (AFG1), G2 (AFG2), M1 (AFM1), and M2 (AFM2) (IARC 2012). Main sources of intake of B- and G-group AFs are contaminated maize, peanuts, tree nuts, and dried fruits (Taniwaki et al. 2018). AFM1 and AFM2 have been considered until now mainly as hydroxylation products of AFB1 and AFB2, formed enzymatically in the liver of lactating dairy cows being fed with AFs contaminated feed (Min et al. 2021). Thus, AFM1 and AFM2 can especially be found as contaminants in milk and dairy products (Mohammed et al. 2016; Min et al. 2021). Little attention has been paid to limited older findings that M-/GM-group AFs can also be produced by A. flavus and A. parasiticus on/in laboratory media (Ramachandra Pai et al. 1975; Dutton et al. 1985; Yabe et al. 2012). Additionally, AFM1 has already been found a few times in cereals like maize and Perl millet (Matumba et al. 2015a; Abdallah et al. 2017; Houissa et al. 2019) as well as dried fruits like figs (Sulyok et al. 2020).
According to the International Agency for Research on Cancer (IARC), AFB1, AFG1, and AFM1 are carcinogenic with sufficient evidence in experimental animals. Limited evidence for carcinogenicity in experimental animals exists for AFB2 and inadequate evidence for AFG2. AFs in general are classified as group 1 carcinogens, due to the sufficient evidence for their carcinogenicity in humans (IARC 2012). AFB1, AFG1, and AFM1 are considered as pro-carcinogens. An enzymatic bioactivation by cytochrome P450 monooxygenases in the liver at the double bond at the 8,9-position in the furan ring to aflatoxin (AF)-8,9-epoxide is necessary for the carcinogenic and toxic activity (Dohnal et al. 2014; EFSA 2020). These epoxides can then form adducts with macromolecules like proteins or DNA, preferably at N7-position of the DNA guanine bases. Compared to AFB1, AFG1 has a reduced capability to intercalate into the DNA due to the less planar δ-lactone ring in its structure (Raney et al. 1990). Particularly, the AF-N7-guanine adduct formation of codon 249 of the p53 tumor suppressor gene is significant, which leads frequently to a missense mutation of this gene. The prevalence of this mutation is associated with the occurrence of hepatocellular carcinoma, as the liver is the main target tissue of AFs (Soini et al. 1996; Kucukcakan and Hayrulai-Musliu 2015).
The biosynthesis pathway of B- and G-group AFs in A. flavus is well elucidated. The necessary genes are grouped together in a gene cluster comprising nearly 80 kb, which is located on chromosome 3 of its genome (Yu 2012; Caceres et al. 2020). This was confirmed by whole genome sequencing of the strain used in the current study (Schamann et al. 2022). The biosynthesis pathway starts with hexanoate units from acetyl-CoA and malonyl-CoA, which are converted via intermediates to sterigmatocystin (ST) in case of the biosynthesis of AFB1 and to dihydrosterigmatocystin (DHST) in the biosynthesis pathway of AFB2. ST and DHST are then methylated to O-methylsterigmatocystin (OMST) and dihydro-OMST (DHOMST), respectively, further hydroxylated to 11-hydroxy-OMST (HOMST) and dihydro-HOMST, respectively, and then metabolized to AFB1 and AFB2, respectively, via intermediate steps (Yu 2012; Caceres et al. 2020). Additional to AFs, some of these AF precursors were investigated for their toxicological potential and revealed to have genotoxic properties (Theumer et al. 2018; Gauthier et al. 2020).
Conflicting hypotheses existed on the biosynthetic relationship between B-/G-group and M-/GM-group AFs (Biollaz et al. 1970; Dutton et al. 1985). However, this uncertainty seemed to be clarified by the postulation of a pathway of the formation of M-/GM-group AFs from OMST and DHOMST by Yabe et al. (2012). According to this, OMST and DHOMST are hydroxylated to aspertoxin (ASP) and dihydro-ASP, respectively, and further hydroxylated to 11-hydroxy-ASP (HASP) and dihydro-HASP, respectively. Both reactions are catalyzed by the enzyme OrdA. Starting from HASP and dihydro-HASP, AFM1 and AFM2 as well as AFGM1 and AFGM2 are formed via intermediates. The enzyme OrdA, which is a monooxygenase belonging to the cytochrome P450 family, is also involved in the biosynthetic pathway of AFB1 (Yabe et al. 2012). The postulated pathway of formation of M-/GM-group AFs would also explain the biosynthetic pathway of ASP, which has received hardly any attention compared to the other AFs so far (Benkerroum 2020). ASP was first isolated and described in 1968 (Rodricks et al. 1968a, b; Waiss et al. 1968). Adverse effects were found in developing chicken embryos, in which beak malformations, hemorrhage from umbilical vessels, edema, and loss of muscle tone were observed (LD50 of 0.7 µg/egg compared to the LD50 of 0.025 µg/egg for AFB1 in this study) (Rodricks et al. 1968a). Furthermore, low acute toxicity was reported in zebra fish larvae (LD50 of 6.6 mg/mL), showing 1/20 the acute toxicity of AFB1 (Abedi and Scott 1969).
The aim of the study was to elucidate the importance of M-AFs by analyzing the formation of AFs and their precursors synthesized by A. flavus (strain MRI19) on maize kernels as one of the most important staple foods in the world. Additionally, the formation kinetics of these compounds was investigated.
Methods
Reference compounds, chemicals, and reagents
The following reference standards were purchased: aflatoxin B1 (AFB1, > 99%), aflatoxin B2 (AFB2, > 99%), aflatoxin G1 (AFG1, > 99%), aflatoxin G2 (AFG2, > 99%), aflatoxin M1 (AFM1, > 98%), and sterigmatocystin (ST, > 99%) all solved in acetonitrile from Sigma-Aldrich Chemie GmbH (Taufkirchen, Germany), aflatoxin M2 (AFM2) in acetonitrile (> 98%) and aflatoxicol (AFL, > 99%) from Cfm Oskar Tropitzsch GmbH (Marktredwitz, Germany), and O-methylsterigmatocystin (OMST, > 95%) from Cayman Chemical (Ann Arbor, MI, USA). Of these standards, two standard mixtures were generated: standard mixture 1 (containing 640 nmol/L of AFB1, AFB2, AFG1, and AFG2 in acetonitrile) and standard mixture 2 (containing 640 nmol/L of AFM1, AFM2, ST, and AFL as well as 608 nmol/L of OMST in acetonitrile). All further chemicals and solvents were of analytical grade. Deionized water was obtained from an in-house ultrapure water system (LaboStar, Erlangen, Germany).
Fungal strain and growth conditions
The strain A. flavus MRI19 (Schamann et al. 2022) of the culture collection of the Max Rubner-Institut was used for the experiments. The strain was originally isolated from tiger nuts, which were grown in the surrounding of Valencia in Spain. For the generation of spores, the fungus was cultivated on MG agar (malt extract [Carl Roth, Karlsruhe, Germany] 17 g/L, glucose [Carl Roth, Karlsruhe, Germany] 5 g/L, agar [Agar–Agar Kobe I; Carl Roth, Karlsruhe, Germany] 16 g/L) at 25 °C. A spore suspension was prepared using Tween-80/NaCl-mixture (NaCl [Carl Roth, Karlsruhe, Germany] 9 g/L, Tween-80 [Serva, Heidelberg, Germany] 1 g/L, agar 1 g/L). Spores were counted using a Thoma cell counting chamber (Paul Marienfeld GmbH & Co. KG, Lauda-Königshofen, Germany) and were diluted to obtain 1.0 × 104 spores per mL. Autoclaved (15 min, 121 °C, 200 kPa) maize kernels (packaged popcorn maize kernels from local supermarket) were used as growth substrate. Eight grams (± 0.1 g) of maize was weighed out per petri dish (Ø = 5.5 cm) and were moistened with 1.8 mL of sterile deionized water. The maize kernels were gently stirred with the aid of a sterile pipette tip for achieving a uniform humidification. After an incubation of 24 h at 25 °C, the kernels were inoculated with 1.5 mL of the spore suspension and again stirred with a pipette tip for achieving a uniform distribution of spores. Then, the inoculated kernels were incubated for up to 10 days at 25 °C. As control, the maize kernels of eight petri dishes were neither moistened with sterile water nor inoculated with spores (control 1). Of these, the kernels of six petri dishes were used for the validation experiment of the analytical method. The kernels of further two petri dishes were moistened with 1.8 mL of sterile deionized water, and 24 h later with 1.5 mL of Tween-80/NaCl mixture without fungal spores (control 2).
Sampling
On the day of inoculation (day 0), the samples of control 1 and 2 (see “Fungal strain and growth conditions”) were taken. Additionally, the first samples of inoculated maize kernels were taken directly after the inoculation (control 3). The further sampling was performed 1, 2, 3, 4, 5, 7, and 10 days after the inoculation. At the beginning, the empty petri dishes were marked to divide them into three equal parts. Maize kernels, which were located within one of these thirds, were determined to be one biological sample. At each sampling time, samples of six biological replicates in total were taken from two different petri dishes and were stored at −20 °C until the mycotoxin extraction.
Mycotoxin extraction
For the mycotoxin extraction, the maize kernels were first homogenized with a ball mill (MM400, Retsch, Haan, Germany). For this, the grinding jars filled with the maize kernels of a sample and one grinding ball were pre-cooled in liquid nitrogen. Then, the kernels were homogenized for 1 min at 30 Hz. The toxin extraction was performed according to the DIN EN ISO norm 16050:2011 with some modifications (DIN 2011). For the extraction, 1 mL of methanol:water (70:30, v:v) and 40 mg of NaCl were added to 200 mg ± 2 mg of ground maize and the samples were shaken on a rotary shaker (VXR basic Vibrax®, IKA, Staufen im Breisgau, Germany) for 2 min at 2,500 rpm. The samples were centrifuged for 5 min at 16,200 × g at room temperature. The extract was transferred to a new tube and the extraction was repeated with 1 mL methanol:water (70:30, v:v). After the second centrifugation, the transferred extracts were combined. Then, the maize samples were centrifuged again for 3 min at 16,200 × g without adding further extracting agent to enable the transfer of the remains of methanol:water. The extract was vortexed and filtered through a 0.2-µm PTFE filter (Puradisc-13, Whatman™, Merck, Darmstadt, Germany). For the UHPLC-MS measurements, 100 µL of the filtrate was diluted with 100 µL of methanol:water (70:30, v:v) to receive concentrations of target analytes within the calibrated range.
Validation of the analytical method
The analytical method for the quantitation of AFs and precursors was validated for selectivity, accuracy, precision, linearity, limit of quantitation, recovery, and matrix effect. For this, maize kernels of six petri dishes of control 1 (see “Fungal strain and growth conditions”) were homogenized as described in “Mycotoxin extraction”. Of each of these six petri dishes, one maize sample was extracted as a blank sample, following the extraction protocol described above. Additionally, of petri dish no. 1, six maize aliquots were used as pre-extract samples and further six aliquots as post-extract samples (each aliquot 200 mg ± 1 mg). For each pre-extract sample, 525 µL of standard mixtures 1 and 2 (see “Reference compounds, chemicals, and reagents”), respectively, were merged, evaporated with nitrogen, and re-dissolved in 100 µL of methanol:water (70:30, v:v). Each pre-extract sample was spiked with 100 µL of these re-dissolved standard mixtures (content of AFs in µg/kg maize: AFB1: 499.6, AFB2: 502.9, AFG1: 525.2, AFG2: 528.5, AFM1: 525.2, AFM2: 528.5, ST: 518.9, OMST: 514.2, AFL: 502.9) directly after the addition of the first milliliter of methanol:water (70:30, v:v) at the beginning of the toxin extraction. Then, the extraction protocol of the spiked samples was followed as described above. The post-extract samples were directly extracted and spiked after the sample workup. For this, instead of the final 1:1 dilution, 100 µL of the extracted samples was mixed with each 25 µL of standard mixtures 1 and 2 (containing 640 nmol/L of each target compound except for OMST [608 nmol/L]), respectively, and 50 µL of methanol:water (70:30, v:v). Additionally, six “solvent-spiked” samples were prepared. For this, 150 µL of methanol:water (70:30, v:v) was spiked with 25 µL of standard mixtures 1 and 2, respectively.
The final concentration of the target compounds in the injected pre-extract (in case of 100% recovery), post-extract as well as solvent-spiked samples was 80 nmol/L, except for OMST (76 nmol/L). The recovery of each compound was calculated by dividing the mean value of the measured peak areas of the target compounds in the pre-extract samples by the mean value in the post-extract samples. The accuracy of the measured analyte levels in the pre-extract samples was calculated by dividing the mean value of the measured concentrations of the target compounds in the pre-extract samples by the nominal concentration of 80 nmol/L (76 nmol/L for OMST) and correcting it for the individual recoveries of the compounds. The matrix effect was calculated by dividing the mean value of the measured peak areas of the target compounds in the post-extract samples by the mean value in the solvent-spiked samples. In this context, values below 100% indicate a signal suppression, whereas values above 100% a signal enhancement.
UHPLC-MS analysis
The samples were measured on a 1290 Infinity LC system (Agilent Technologies, Waldbronn, Germany) consisting of a pump (G4220A), an autosampler (G4226A) a column oven, and a DAD (G4212A) coupled with a Triple TOF 5600 mass spectrometer (AB Sciex, Darmstadt, Germany). The separation was performed on a Waters Cortecs UPLC C18 column (2.1 mm × 150 mm, 1.6 µm; Waters, Eschborn, Germany) equipped with a pre-column (Security Guard Ultra UHPLC C18; Phenomenex, Aschaffenburg, Germany). Aqueous ammonium acetate buffer (10 mmol/L) was used as eluent A and methanol as eluent B at a flow rate of 0.25 mL per min. A gradient with the following elution profile was performed: 0.0–1.0 min isocratic with 30% B, 1.0–13.0 min from 30 to 42% B, 13.0–23.0 min from 42 to 77% B, 23.0–23.5 min from 77 to 95% B, 23.5–26.5 min isocratic with 95% B, 26.5–27.0 min from 95 to 30% B, and 27.0–37.0 min isocratic with 30% B. The column oven was set at 45 °C, and the injection volume was 1 µL. The DAD recorded from 200 to 600 nm operating with a sampling rate of 2.5 Hz. Measurements of the MS were performed in the positive ESI mode, selecting the following ionization source conditions: curtain gas 35 psi, ion spray voltage 5,500 V, ion source gas-1 50 psi, ion source gas-2 60 psi, and ion source gas-2 temperature 550 °C. The declustering potential was set to 120 V. The MS full scans were recorded from m/z 100–1000 with an accumulation time of 100 ms, and a collision energy voltage of 10 V. The MS/MS spectra were recorded in the high sensitivity mode from m/z 50–1000 with an accumulation time of 25 ms, a collision energy voltage of 35 V, and a collision energy spread of 15 V.
Analytes were identified by retention time, accurate mass, and isotope pattern. Extracted ion chromatograms (XIC) based on the accurate mass of the molecular ions of the compounds (5 mDa extraction width) were used to monitor and quantify the analytes (Table 1). For the quantitation of the target compounds, two mixtures of standard solutions (standard mixtures 1 and 2; see “Reference compounds, chemicals, and reagents”) were measured in four different concentrations (AFB1, AFB2, AFG1, AFG2, AFM1, AFM2, ST, and AFL: 640 nmol/L, 160 nmol/L, 40 nmol/L, 10 nmol/L; OMST: 608 nmol/L, 152 nmol/L, 38 nmol/L, 9.5 nmol/L) at the beginning and at the end of each measuring day to obtain a standard curve. The working standard solutions were renewed every measuring day. The standard solutions measured on the same day as the samples were used for data quantification.
Table 1.
Analyte specific parameters of UHPLC-MS analysis, showing the retention time as well as the monitored ion species and accurate mass. Additionally, MS/MS data of analytes are displayed (precursor ion and some major product ions)
| Analyte | Retention time [min] | Monitored accurate mass [Da] | Ion species | Precursor ion (MS/MS analysis) [m/z] | Product ions (MS/MS analysis)a [m/z] |
|---|---|---|---|---|---|
| AFB1 | 13.2 | 313.07066 ± 0.00250 | [M + H]+ | 313.1 | 285.1 (31), 284.1 (11), 270.1 (18), 269.0 (14), 241.0 (17), 214.1 (10) |
| AFB2 | 11.3 | 315.08631 ± 0.00250 | [M + H]+ | 315.1 | 297.1 (5), 287.1 (21), 271.1 (5), 259.1 (19), 243.1 (5), 203.1 (3) |
| AFG1 | 9.7 | 329.06558 ± 0.00250 | [M + H]+ | 329.1 | 311.1 (24), 283.1 (18), 255.1 (13), 243.1 (32), 215.1 (14), 214.1 (13) |
| AFG2 | 8.1 | 331.08123 ± 0.00250 | [M + H]+ | 331.1 | 313.1 (16), 303.1 (6), 285.1 (7), 257.1 (6), 245.1 (9), 217.1 (4) |
| AFM1 | 8.5 | 329.06558 ± 0.00250 | [M + H]+ | 329.1 | 301.1 (28), 273.1 (97), 259.1 (41), 258.1 (13), 255.1 (11), 229.0 (22) |
| AFM2 | 6.7 | 331.08123 ± 0.00250 | [M + H]+ | 331.1 | 313.1 (31), 285.1 (40), 273.1 (100), 259.1 (41), 257.1 (16), 229.0 (13) |
| ST | 22.7 | 325.07066 ± 0.00250 | [M + H]+ | 325.1 | 310.0 (98), 309.0 (6), 297.1 (7), 282.1 (16), 281.0 (63), 253.0 (5) |
| OMST | 20.7 | 339.08631 ± 0.00250 | [M + H]+ | 339.1 | 324.1 (28), 311.1 (7), 306.1 (38), 295.1 (24), 278.1 (15), 277.1 (18) |
| HOMST | 14.8 | 355.08123 ± 0.00250 | [M + H]+ | 355.1 | 340.1 (2), 327.1 (20), 299.1 (54), 285.1 (30), 266.1 (20), 255.1 (12)b |
| ASP | 16.4 | 355.08123 ± 0.00250 | [M + H]+ | 355.1 | 340.1 (36), 327.1 (5), 322.0 (66), 294.1 (16), 293.0 (19), 266.1 (5)b |
| HASP | 13.6 | 371.07614 ± 0.00250 | [M + H]+ | 371.1 | 343.1 (20), 315.1 (42), 282.1 (35), 281.0 (9), 301.1 (22), 300.1 (20)b |
| DHST | 22.1 | 327.08631 ± 0.00250 | [M + H]+ | 327.1 | 312.1 (14), 299.1 (10), 284.1 (7), 283.1 (7), 271.1 (10), 99.0 (10) |
| DHOMST | 19.7 | 341.10196 ± 0.00250 | [M + H]+ | 341.1 | 326.1 (11), 313.1 (4), 308.1 (5), 297.1 (13), 285.1 (10), 280.1 (6) |
| AFL | 17.3 | 297.07575 ± 0.00250 | [M-H2O + H]+ | 297.1 | 281.1 (28), 269.1 (38), 268.1 (31), 254.1 (20), 241.1 (20), 225.1 (19) |
AFB1 aflatoxin B1, AFB2 aflatoxin B2, AFG1 aflatoxin G1, AFG2 aflatoxin G2, AFM1 aflatoxin M1, AFM2 aflatoxin M2, ST sterigmatocystin, OMST O-methylsterigmatocystin, HOMST 11-hydroxy-O-methylsterigmatocystin, ASP aspertoxin, HASP 11-hydroxyaspertoxin, DHST dihydrosterigmatocystin, DHOMST dihydro-O-methylsterigmatocystin, AFL aflatoxicol
aIntensities of product ions are indicated in brackets in percentage
bPresented product ions include only ions of the revised MS/MS spectrum of the target analytes without ions of polysiloxanes
Data analysis
For data analysis, the software MultiQuant 3.0.2 and PeakView 2.2 (AB Sciex, Darmstadt, Germany) were used. Quantification of the analytes was based on standard curves, for which linear regression with a weighting of 1/x2 was applied. Since standards were not commercially available for all target analytes, the compound which is structurally most similar to the target analyte was used for its quantification. Thus, ASP, HOMST, HASP, and DHOMST were semi-quantified based on the standard of OMST and DHST on the standard of ST. The lowest concentration of the standard curve (10 nmol/L) was set as limit of quantitation (LOQ) of all target compounds. At this concentration, the signal to noise value was 8 for AFL, between 26 and 40 for AFB1, AFB2, AFM1, and AFM2, and between 60 and 70 for AFG1, AFG2, ST, and OMST. Concentrations measured below 10 nmol/L as well as the absence of a compound in a sample were indicated as “ < LOQ” in the results. For calculating mean values, “ < LOQ” was set to 0.0 mg/kg maize. Samples containing target compounds in a higher concentration than the highest point of the standard curve (640 nmol/L) were diluted appropriately with methanol:water (70:30, v:v) to get them within the calibrated range. Then, these samples were repeatedly measured. For each target compound specifically, the dilution level, at which this compound was in the calibrated range, was used for data analysis. Statistical analyses were performed using SPSS Statistics 26 (IBM, Armonk, NY, USA).
Results
Mycotoxin identification
The analytes AFB1, AFB2, AFG1, AFG2, AFM1, AFM2, ST, OMST, and AFL were identified by comparison of their retention times, accurate masses in MS spectra and MS/MS spectra with those of the reference compounds. Additionally, MS/MS spectra were verified with spectra in the literature (Plattner et al. 1984; Uka et al. 2019). Furthermore, the inoculated maize samples were checked for the occurrence of the following compounds based on their accurate masses (5 mDa extraction width): AFGM1, AFGM2, ASP, HASP, HOMST, DHST, DHOMST, dihydro-ASP, dihydro-HASP, and dihydro-HOMST. Of these, a peak in the respective mass trace was detected in the inoculated maize samples for the following analytes: ASP, HASP, HOMST, DHST, and DHOMST. The MS/MS spectra of the putative ASP, DHST, and DHOMST signals (Table 1) were verified by comparison with those published by Uka et al. (2019). No MS/MS spectra were found in literature for HASP and HOMST. The measured accurate masses and isotope ratios fit to the theoretical calculated values (HASP: measured m/z 371.0792, +8.2 ppm mass error; HOMST: measured m/z 355.0811, −0.4 ppm mass error). The recorded MS/MS spectra of these two compounds were examined carefully (Table 1). In the MS/MS spectrum of the supposed HASP (hydroxylated ASP), some fragment ions with a m/z difference for oxygen (15.995 u) compared to fragment ions in the spectrum of ASP were recorded (e.g., m/z 343.083 and 282.051 compared to 327.086 and 266.056, respectively) (Table 1), which affirmed the putative identification as HASP. In the case of the supposed HOMST, an analog observation was made, namely fragment ions having a m/z difference for oxygen (15.995 u) to fragment ions in the spectrum of OMST (m/z 340.058 and 327.086 compared to 324.063 and 311.091, respectively) (Table 1).
It should be mentioned that a continuous and stable background noise of polysiloxanes (m/z 371.10 and 355.07) was detected. Such polysiloxane interferences were already described in a previous work (Keller et al. 2008). This polysiloxane background noise led to mixed MS/MS spectra of ASP, HOMST, and HASP, since the precursor ions in MS/MS analysis were selected by a non-high resolving quadrupole. The product ions of the MS/MS spectra of the polysiloxanes were subtracted from the mixed spectra to obtain the pure MS/MS spectra of the target analytes. Only product ions of these subtracted MS/MS spectra are listed in Table 1. However, this background noise was not relevant for the quantification of the target compounds, due to high mass resolution in MS full scan analysis.
Validation
None of the target compounds was detected in the blank samples. For calibration curves, the best fit line was obtained by linear regression applying a weighting of 1/x2. The correlation coefficient of all analytes indicated the quality of the calibration curves and was ≥ 0.9963. Recoveries of the analytes after extraction from maize exhibited values between 85.9 and 94.3%. Thus, the measured concentrations of the analytes in the study samples were corrected for these recoveries. Matrix effect values between 97.6 and 99.1% were measured, except for AFM2 (93.9%) and OMST (90.0%). Recovery-corrected accuracies and intra-day precision of analytes in spiked maize were 93.0–104.1% and 0.4–3.7%, respectively. The correlation coefficients, recoveries of the extraction process, matrix effect and the precision as well as accuracies corrected for the recoveries of the pre-extract samples of the included standards are listed in Table 2.
Table 2.
Results of the validation experiment for mycotoxin analysis in maize by UHPLC-MS with a spiking level of the injected samples of 80 nmol/L for each analyte except for OMST (76 nmol/L). The correlation coefficients (R), recoveries of the extraction process (n = 6), matrix effect (n = 6), and the intra-day precision (n = 6) as well as the recovery-corrected accuracies (n = 6) are listed
| Analyte | Correlation coefficient (R) | Recovery [%] | Matrix effect [%] | Precision [%] | Accuracya [%] |
|---|---|---|---|---|---|
| AFB1 | 0.9968 | 89.0 | 98.3 | 2.3 | 104.1 |
| AFB2 | 0.9963 | 88.0 | 99.1 | 3.2 | 103.7 |
| AFG1 | 0.9969 | 88.4 | 97.7 | 1.7 | 101.9 |
| AFG2 | 0.9964 | 89.4 | 98.4 | 0.9 | 102.7 |
| AFM1 | 0.9974 | 90.1 | 98.8 | 1.7 | 100.3 |
| AFM2 | 0.9970 | 90.9 | 93.9 | 2.6 | 95.5 |
| ST | 0.9972 | 85.9 | 97.6 | 1.4 | 99.6 |
| OMST | 0.9967 | 88.6 | 90.0 | 0.4 | 93.0 |
| AFL | 0.9970 | 94.3 | 99.0 | 3.7 | 97.4 |
AFB1 aflatoxin B1, AFB2 aflatoxin B2, AFG1 aflatoxin G1, AFG2 aflatoxin G2, AFM1 aflatoxin M1, AFM2 aflatoxin M2, ST sterigmatocystin, OMST O-methylsterigmatocystin, AFL aflatoxicol
aAccuracies corrected for analyte-specific recoveries
Kinetics of AFs, their precursors, and the metabolization product AFL on maize
In our pre-experiments to this study (data not shown), an untargeted analysis of AF metabolites produced by A. flavus MRI19 on potato dextrose agar (PDA) was performed using UHPLC-MS. In addition to the formation of AFM1 (1–2% of the produced AFB1), high peaks of ASP and OMST were observed in this pre-experiment. The current study was performed among others to analyze, if these compounds were also produced in high levels on food like maize.
For this, autoclaved maize kernels were inoculated with a spore suspension of A. flavus MRI19 and were incubated for up to 10 days. The following AFs were detected in the samples: AFB1, AFB2, AFM1, AFM2, and AFL. Additionally, the samples were checked for metabolites of the last steps of the AF biosynthesis and the following precursors were detected: ST, DHST, OMST, DHOMST, HOMST, ASP, and HASP. Chemical structures of the detected compounds are shown in Fig. 1. Measured levels of AFs and their precursors on maize samples are listed in Table S1 and are illustrated in Figs. 2, 3 and 4. The Pearson correlation coefficients between selected target compounds were calculated and are listed in Table 3. In the following, the results are presented in detail for each analyte group.
Fig. 1.

Chemical structure of aflatoxins (AFs) and precursors detected in the study. The last steps of the AF group 1 pathway for the formation of AFB1 and AFM1 and the AF group 2 pathway for the formation of AFB2 and AFM2 are shown. The pathway for the biosynthesis of B-group AFs is based on Yu (2012) and Caceres et al. (2020), and the pathway for the biosynthesis of M-group AFs is postulated by Yabe et al. (2012). The carbons were numbered regarding Pfeiffer et al. (2014)
Fig. 2.

Line diagram showing the formation of aflatoxin B1 (AFB1), aflatoxin B2 (AFB2), aflatoxin M1 (AFM1), aflatoxin M2 (AFM2), aspertoxin (ASP), 11-hydroxyaspertoxin (HASP), and 11-hydroxy-O-methylsterigmatocystin (HOMST) by A. flavus on autoclaved maize kernels over the incubation time of 10 days. Data is given as arithmetic mean ± standard deviation of six biological samples in milligrams per kilogram of maize
Fig. 3.
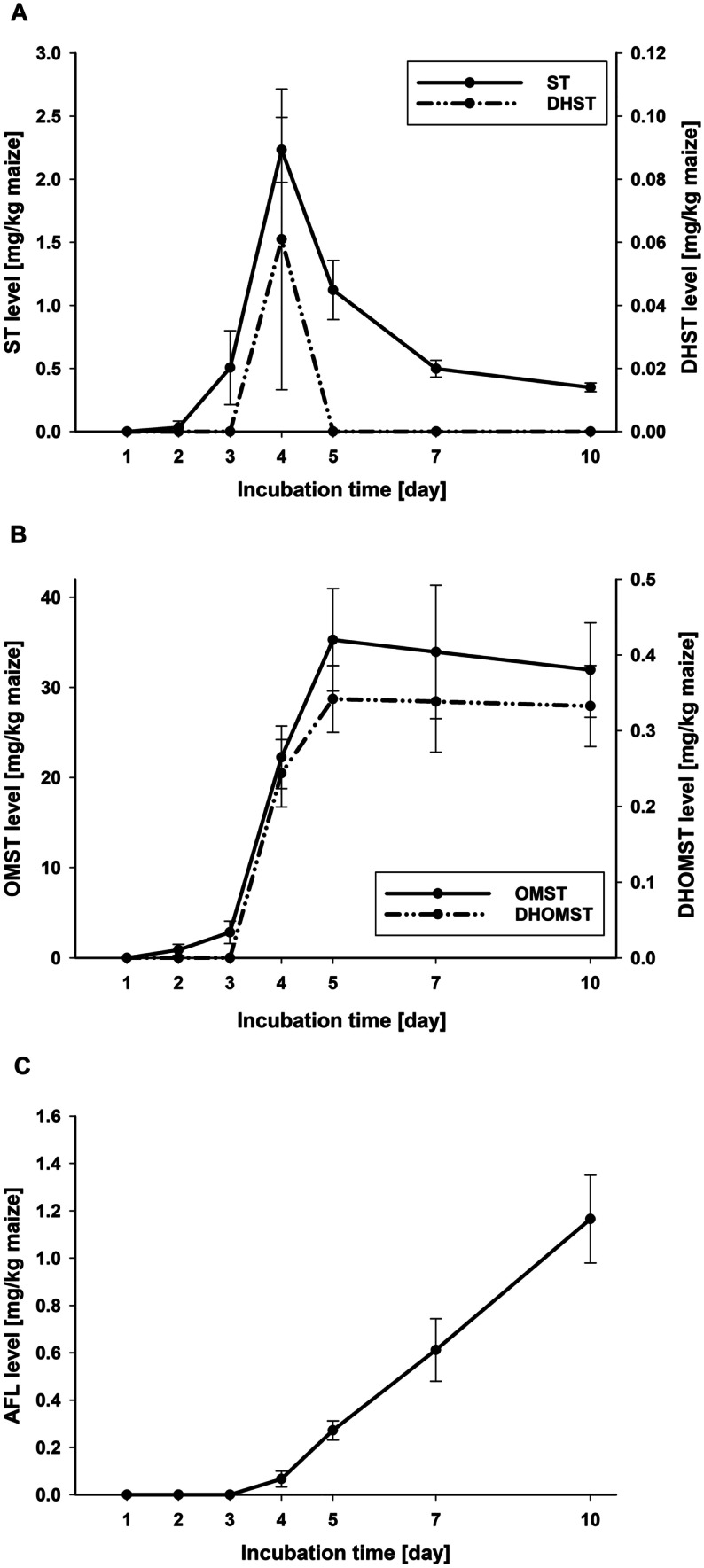
Line diagram showing the formation of the aflatoxin precursors sterigmatocystin (ST, primary y-axis) and dihydrosterigmatocystin (DHST, secondary y-axis) (A), as well as O-methylsterigmatocystin (OMST, primary y-axis) and dihydro-O-methylsterigmatocystin (DHOMST, secondary y-axis) (B), and of aflatoxicol (AFL) as metabolization product of aflatoxin B1 (AFB1) (C) by A. flavus on autoclaved maize kernels over the incubation time of 10 days. Data is given as arithmetic mean ± standard deviation of six biological samples in milligrams per kilogram of maize
Fig. 4.

Pie chart showing the relative levels of aflatoxins and precursors related to the sum of all detected analytes presenting the arithmetic mean over days 5, 7, and 10 (n = 18). AFB1, aflatoxin B1; AFB2, aflatoxin B2; AFM1, aflatoxin M1; AFM2, aflatoxin M2; ASP, aspertoxin; DHOMST, dihydro-O-methylsterigmatocystin; HASP, 11-hydroxyaspertoxin; HOMST, 11-hydroxy-O-methylsterigmatocystin; ST, sterigmatocystin; OMST, O-methylsterigmatocystin; AFL, aflatoxicol
Table 3.
Pearson correlation coefficients of associations between aflatoxin/precursor levels produced by A. flavus on autoclaved maize kernels from days 3 to 10 of incubation
| Analytes (group 1 pathway)a | Pearson correlation | Analytes (group 2 pathway)b | Pearson correlation | Analytes (corresponding metabolites) | Pearson correlation | |||
|---|---|---|---|---|---|---|---|---|
| r | p value | r | p value | r | p value | |||
| ST-OMST | 0.046 | 0.810 | DHST-DHOMST | 0.015 | 0.936 | ST-DHST | 0.781 | < 0.001* |
| OMST-HOMST | 0.865 | < 0.001* | AFB2-AFM2 | 0.919 | < 0.001* | OMST-DHOMST | 0.981 | < 0.001* |
| OMST-ASP | 0.863 | < 0.001* | AFB1-AFB2 | 0.984 | < 0.001* | |||
| HOMST-AFB1 | 0.977 | < 0.001* | AFM1-AFM2 | 0.916 | < 0.001* | |||
| ASP-HASP | 0.943 | < 0.001* | ||||||
| HASP-AFM1 | 0.987 | < 0.001* | ||||||
| AFB1-AFM1 | 0.996 | < 0.001* | ||||||
*Significant correlation (p < 0.001)
Analytes are divided as follows:
aGroup 1 aflatoxin pathway: ST, sterigmatocystin; OMST, O-methylsterigmatocystin; HOMST, 11-hydroxy-O-methylsterigmatocystin; ASP, aspertoxin; HASP, 11-hydroxyaspertoxin; AFB1, aflatoxin B1; AFM1, aflatoxin M1
bGroup 2 aflatoxin pathway: DHST, dihydrosterigmatocystin; DHOMST, dihydro-O-methylsterigmatocystin; AFB2, aflatoxin B2; AFM2, aflatoxin M2
Group 1 AFs
The first compound of the monitored pathway of group 1 AFs (AFB1, AFG1, AFM1) analyzed in this study is ST, which was detected in low levels from day 2 on of the incubation. A clear maximum in its formation (2.2 mg/kg maize) was detected on day 4 with a subsequent decrease to 0.35 mg ST/kg maize on day 10 (Fig. 3A, Table S1) due to its methylation to OMST. OMST was measured first on day 2 in low levels, increased strongly from days 3 to 5 and peaked on day 5 (35.3 mg/kg maize). Then, it decreased slightly until day 10 (32.0 mg/kg maize) (Fig. 3B, Table S1). OMST is hydroxylated to HOMST and, according to Yabe et al. (2012), also to ASP. Both compounds showed an increasing formation from the first quantification on day 2 for ASP and day 4 for HOMST to day 10 (Fig. 2, Table S1). From day 4, the detected level of ASP was about threefold higher than that of HOMST within the respective day. HOMST is further metabolized to AFB1, which was already detected after 2 days of incubation. AFB1 was the main metabolite of A. flavus in this study, being produced by far in the highest level compared to the other monitored metabolites. It showed a continuous increase over time to 269.0 mg/kg maize on day 10 (Fig. 2, Table S1). According to Yabe et al. (2012), ASP is hydroxylated to HASP and further to AFM1, which were both quantified the first time on day 3 and showed a very comparable, well correlated (r = 0.987) increase over time to 3.2 and 4.1 mg/kg maize for HASP and AFM1, respectively, after 10 days of incubation (Fig. 2, Tables 3 and S1). The formation of AFM1 correlated also very well with the AFB1 formation (r = 0.996; Table 3). AFG1 was not detected on maize samples inoculated with A. flavus MRI19 at any day of incubation.
Group 2 AFs
A lower number of compounds was detected of the group 2 AF pathway (AFB2, AFG2, AFM2). Only in a few samples DHST was measured in levels above the LOQ. DHST showed like the group 1 analog ST a peak in its formation on day 4 of incubation, after which it decreased to levels under the LOQ in all samples on days 5, 7, and 10 (Fig. 3A, Table S1). DHOMST was detected from day 4 on and increased to day 5, after which it showed a slight decrease until day 10. The kinetics of DHOMST was correlative to that of the group 1 analog OMST (Fig. 3B), showing a high association (r = 0.981; Table 3), although the produced level of DHOMST was only 1/100 of that of OMST. As end products of the pathway, AFB2 and AFM2 were detected. AFB2, which was quantified first on day 3, showed a continuous increase to 10.2 mg/kg maize on day 10 (Fig. 2, Table S1). Its kinetics was comparable to that of AFB1 and correlated very well (r = 0.984; Table 3). AFM2 was first measured in levels above the LOQ on day 5. Its formation increased linearly to 0.7 mg/kg maize on day 10 (Fig. 2, Table S1). AFG2 was not detected on maize samples inoculated with A. flavus MRI19 at any day of incubation.
Metabolization product AFL
Maize samples inoculated with A. flavus MRI19 were checked for AFL, a hydroxylated metabolization product of AFB1. AFL was detected the first time above the LOQ on day 4 of incubation and showed a continuous increase. After 10 days of incubation, 1.2 mg of AFL was measured per kilogram of maize (Fig. 3C, Table S1).
Mycotoxin profile on maize
For the visualization of the profile of the measured AFs and their precursors on maize, the relative level of each analyte related to the sum of all target compounds was calculated. For this, the mean levels of all detected analytes (AFB1, AFB2, AFM1, AFM2, ST, DHST, OMST, DHOMST, HOMST, ASP, HASP, and AFL) were summed up separately for each day of incubation (days 5, 7, and 10). Then, the ratio between the mean produced level of each compound and the corresponding sum was calculated separately for each day (5, 7, and 10). Next, the relative levels of each analyte were averaged over days 5, 7, and 10 of incubation. The means of these three days were calculated to obtain a rather general illustration of the formation of the target compounds within the investigated incubation period, since not all compounds had their formation peak on the same day. Furthermore, the early days of incubation were omitted, since not all compounds were detected on the same day of incubation the first time. Results are shown as pie chart in Fig. 4. The highest proportions were contributed by AFB1 (76.6%), OMST (13.2%), and ASP (3.8%). When including only the four AFs AFB1, AFB2, AFM1, and AFM2 in this calculation, a total of 218.8 mg AFs per kilogram of maize (mean of days 5, 7, and 10) was produced, of which 95.3% belonged to AFB1, 3.2% to AFB2, 1.4% to AFM1, and 0.2% to AFM2. Since AFB1 is the main metabolite in this study, analog ratios related to the level of AFB1 instead of the sum of the levels of all analytes were further calculated. Again, the mean formation of the compounds on days 5, 7, and 10 was considered. AFB2, AFM1, and AFM2 were measured to 3.3%, 1.5%, and 0.2% of AFB1, respectively. ST and AFL were detected in low levels of 0.4% and 0.3% of AFB1, whereas OMST and ASP were measured in relatively high levels of 17.4% and 4.9% compared to AFB1.
Discussion
The aim of the study was to analyze the formation kinetics of AFs and their precursors by A. flavus on maize. In general, the proposed synthesis pathway of AFs could be supported, since most of its compounds were detected. The kinetics of ST formation with a first increase and a subsequent decline in the ST level indicated a quick conversion to OMST. Thus, ST contributed quantitatively only marginally to the toxicological profile of the fungus. A rapid conversion of ST to OMST was already described by Rank et al. (2011). Yogendrarajah et al. (2015) reported low concentrations of ST compared to those of OMST and AFB1 as well, when analyzing the mycotoxin profile of a variety of A. flavus and A. parasiticus strains on malt extract agar by LC–MS/MS. However, other fungal species like A. nidulans cannot further transform ST to OMST (Brown et al. 1996; Chen et al. 2016), which could lead to relatively high ST levels in food infected by such species. In contrast to ST, OMST was detected in high levels on day 7 and 10 in the current study, which may suggest that its formation is higher than the further metabolization. Yogendrarajah et al. (2015) confirmed the formation of high levels of OMST by different strains. The measured ratios of OMST to AFB1 reported by Yogendrarajah et al. (2015) were mostly in a comparable range of that measured in the current study. Due to the high OMST formation, the question about its toxicological relevance raised, since it has the same structural element (double bond in the furan ring), which is responsible for the genotoxicity of AFB1. Little is so far known about the toxicity of OMST. When checking its genotoxicity using a hepatocyte primary culture/DNA repair test, a genotoxic effect was found due to its positive reaction for DNA repair (dose 10−4, 10−5, 10−6 M) (Mori et al. 1986). In contrast, no genotoxicity was observed for OMST in a recent cell study using on the one hand two human cell lines with bioactivation capabilities (HepG2 hepatoblastoma cells, LS-174 T epithelial colorectal adenocarcinoma cells) and on the other hand one human cell line with poor bioactivation capabilities (ACHN renal cell adenocarcinoma cells) (Theumer et al. 2018). However, in this study, cytotoxic effects were observed at the highest OMST concentration (100 µmol/L). Furthermore, analyzing the mutagenicity in an Ames test using Salmonella typhimurium with and without metabolic activation showed no mutagenic effects of OMST (0.1, 1, 10, and 100 µg/testing plate) in contrast to ST as well as AFB1 (Wehner et al. 1978). Thus, considering the measured levels of OMST in the current study as well as the lack of consistent data, further toxicological analyses of OMST would be necessary.
For the formation of AFB1, OMST is hydroxylated to HOMST by the enzyme OrdA (Udwary et al. 2002). Beyond that, the relatively low detected levels of HOMST compared to AFB1 (approx. 1:60) suggested that the further conversion of HOMST proceeded again relatively rapidly. Via further intermediates, which were not analyzed in this study, AFB1 was formed by far in the highest levels compared to all other measured metabolites. Since AF formation can be expected to increase above day 10 of incubation, the ratios of AFs and their precursors (Fig. 4) are expected to change with prolonged incubation.
The pathway of the formation of B-group AFs is well elucidated. However, less is known about the biosynthetic relationship between B-/G-group and M-/GM-group AFs. Yabe et al. (2012) postulated that M-group AFs were formed from OMST via ASP followed by HASP and further intermediates. Unexpectedly, ASP was the analyte being measured in this study in the third highest level of all analyzed metabolites. It must be noticed that no reference compound of ASP was commercially available and that ASP was quantified using the calibration curve of OMST. For this reason, the actual levels of ASP in the study samples may differ from the values reported here. Surprisingly, hardly any research was published about ASP so far, although its toxicity was already shown in developing chicken embryos in the first description of the compound (Rodricks et al. 1968a). As far as we know, this is the first report on the detection of ASP in food and on the analysis of its kinetics, which is probably due to the fact that samples were simply not tested for ASP in the past. Therefore, monitoring its presence in food is highly relevant, considering the measured levels in the current study. In addition, the toxicity of ASP needs to be investigated, as the compound (like OMST and AFB1) carries the toxicologically relevant double bond in the furan ring.
The end product of this branch of the postulated AF pathway is AFM1. In the current study, the detected level of AFM1 was 1.5% of that of AFB1. A comparable AFM1 formation of 1/100–1/200 of that of AFB1 was already described (Nakazato et al. 1991). It was suggested that the higher formation of AFB1 compared to AFM1 may be due to the higher affinity of the enzyme OrdA for the hydroxylation of OMST at the 11-carbon compared to the 12c-carbon. This would lead to the preferred formation of HOMST instead of ASP, since the same enzyme seems to be responsible for these two hydroxylations (Yabe et al. 2012). The detection of AFM1 in food like maize was already reported a few times. It has been speculated that insects metabolized AFB1 to AFM1 after ingesting AFB1 contaminated grains (Matumba et al. 2015a; Abdallah et al. 2017; Getachew et al. 2018). However, the current study demonstrated that A. flavus can produce AFM1 on maize, as a possible transformation by maize enzymes was highly likely suppressed by autoclaving the maize kernels. Furthermore, AFM1 was also detected in A. flavus cultured on PDA medium (data not shown). G-group as well as GM-group aflatoxins were not detected in the current study. This is not surprising, since A. flavus usually does not produce these aflatoxins in contrast to, for instance, A. parasiticus (Ehrlich et al. 2004).
Compared to group 1 AFs (AFB1, AFM1), low levels of group 2 AFs (AFB2, AFM2) were formed in this study, as reported in most studies analyzing AF formation (Kensler et al. 2011; Matumba et al. 2015b; Ting et al. 2020). Analogies in the kinetics of the corresponding compounds of group 1 and 2 AFs and precursors could be observed, which was already described for AFB1 and AFB2 as well as AFM1 and AFM2 (Nakazato et al. 1991).
In the current study, AFL was firstly measured two days later as AFB1 (day 4 vs. day 2), which suggested that AFL is a metabolization product of AFB1 (Detroy and Hesseltine 1970). Interestingly, a slightly different shape of formation kinetics of AFL compared to that of AFB1 was observed. The AFL formation seemed to start slightly exponential (days 3 to 5), followed by nearly linear formation kinetics from days 5 to 10 (Fig. 3C). In contrast, AFB1 formation seemed to show a slightly flattening curve from day 5 on (Fig. 2). It has to be noticed that more data points are required for a more conclusive interpretation of the shape of the curves. Although the difference between the kinetics of AFL and AFB1 formation is small and requires further investigation, this is in line with observations reported by Nakazota et al. (1991): The formation of B- and M-group AFs by most of the analyzed A. flavus strains investigated increased until day 15 of incubation and decreased then until day 20, whereas AFL increased beyond day 15. Furthermore, the authors described that the formation of AFB1 and AFL did not correlate (Nakazato et al. 1991).
In the EU, maximum levels for AFs were defined by the Commission Regulation No. 1881/2006, which was complemented by the Regulations 165/2010 and 1058/2012. Maximum levels for AFM1 exist only for milk, milk-based products, infant formulae, and dietary foods. Additionally, maximum levels were set for AFB1 as well as the sum of AFB1, AFB2, AFG1, and AFG2 for a variety of cereals, nuts, dried fruits, and spices (EU 2006, 2010, 2012). Hardly any report of the occurrence of AFM2 in foods other than milk and dairy products exists. This may be due to the fact that M-group AFs have so far been monitored almost exclusively in this food group. For instance, the data considered for the risk assessment of the EFSA included mainly the food categories milk and dairy products, animal and vegetable fats and oils, food for infants, and snacks, whereas M-group AFs were hardly controlled in studies analyzing for example cereals or nuts (EFSA 2020). However, since the current study clearly demonstrates that AFM1 and AFM2 can occur in food beside milk and dairy products, the monitoring of these AFs would be necessary in the same food categories regularly monitored for B- and G-group AFs. If it was confirmed that M-group AFs were frequently found in these food categories as well, it would be necessary to discuss, whether M-group AFs should be included in the EU sum maximum level of AFs (currently sum of AFB1, AFB2, AFG1, and AFG2) for different foodstuffs. The inclusion of M-group aflatoxins in this sum level should be considered, especially when keeping in mind that the carcinogenicity of AFM1 is better confirmed than that of AFB2 and AFG2 (IARC 2012). For AFM2, the toxicity is still relatively unknown and, thus, it was not included in the risk assessment of AFs in food of the EFSA (EFSA 2020). However, we support the opinion of the EFSA that further data on AFM2 are needed (EFSA 2020). Beside the evaluation of M-group AFs, it should be considered in further studies, in how far toxicologically relevant precursors, like versicolorin A, contribute to health risk. After a subsequent risk assessment, it would be necessary to examine, whether such precursors must also be regulated in food.
To the best of our knowledge, this study is the first detailed description of the kinetics of precursors of AFs on food (maize kernels). However, further toxicological relevant precursors of AFs are known (e.g., versicolorin A) (Theumer et al. 2018; Gauthier et al. 2020), which were not monitored in this study, since a valid semi-quantification of these compounds was not fully achievable due to lack of the (structural related) reference standards. Further studies should be conducted to elucidate the quantitative relevance of these toxins in food. Due to the laborious sample preparation, the application of a validated method, and the inclusion of six biological replicates per sampling day, it was possible to show the biological variation rather than the technical variation. This can be demonstrated by the relatively low standard deviations. Unfortunately, standards were not commercially available for all analyzed compounds. Thus, it was necessary to estimate the concentration of those compounds based on the available standards of closely related analytes. However, this has no influence on the shape of the kinetics of these compounds. Furthermore, it should be mentioned that autoclaved maize kernels were used, which might alter the maize surface structure and might facilitate the infection of the fungus compared to unprocessed maize. Additionally, this did not fully represent the common situation of stored maize. However, the autoclavation of the maize kernels was necessary to inactivate other microorganisms and to suppress the activity of maize enzymes.
In conclusion, the study showed the formation of B- and M-group AFs and precursors like ST, OMST, and ASP on maize being produced by A. flavus. The kinetics of the detected compounds was described in detail in the present study. The results indicate that these compounds could possibly be found in contaminated food in relevant levels. Therefore, the monitoring of the occurrence of M-group AFs, ASP, and OMST in food as well as further investigation of the toxicological potency of ASP and OMST are suggested in order to enable and/or improve a risk assessment of these compounds.
Supplementary information
Below is the link to the electronic supplementary material.
Abbreviations
- AF
Aflatoxin
- AFB1
Aflatoxin B1
- AFB2
Aflatoxin B2
- AFG1
Aflatoxin G1
- AFG2
Aflatoxin G2
- AFM1
Aflatoxin M1
- AFM2
Aflatoxin M2
- ST
Sterigmatocystin
- OMST
O-Methylsterigmatocystin
- HOMST
11-Hydroxy-O-methylsterigmatocystin
- ASP
Aspertoxin
- HASP
11-Hydroxyaspertoxin
- DHST
Dihydrosterigmatocystin
- DHOMST
Dihydro-O-methylsterigmatocystin
- AFL
Aflatoxicol
- LOQ
Limit of quantitation
Funding
Open Access funding enabled and organized by Projekt DEAL. The project is being funded by the Federal Ministry of Food and Agriculture (BLE) under the reference AflaZ 2816PROC11.
Declarations
Conflict of interest
The authors declare no competing interests.
Footnotes
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- Abdallah MF, Girgin G, Baydar T, Krska R, Sulyok M. Occurrence of multiple mycotoxins and other fungal metabolites in animal feed and maize samples from Egypt using LC-MS/MS. J Sci Food Agric. 2017;97:4419–4428. doi: 10.1002/jsfa.8293. [DOI] [PubMed] [Google Scholar]
- Abedi ZH, Scott PM. Detection of toxicity of aflatoxins, sterigmatocystin, and other fungal toxins by lethal action on zebra fish larvae. J Assoc off Anal Chem. 1969;52:963–969. doi: 10.1093/jaoac/52.5.963. [DOI] [Google Scholar]
- Benkerroum N. Aflatoxins: producing-molds, structure, health issues and incidence in southeast Asian and sub-Saharan African countries. Int J Environ Res Public Health. 2020;17:1215. doi: 10.3390/ijerph17041215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Biollaz M, Büchi G, Milne G. Biosynthesis of the aflatoxins. J Am Chem Soc. 1970;92:1035–1043. doi: 10.1021/ja00707a050. [DOI] [PubMed] [Google Scholar]
- Brown DW, Yu JH, Kelkar HS, Fernandes M, Nesbitt TC, Keller NP, Adams TH, Leonard TJ. Twenty-five coregulated transcripts define a sterigmatocystin gene cluster in Aspergillus nidulans. PNAS. 1996;93:1418–1422. doi: 10.1073/pnas.93.4.1418. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caceres I, Al Khoury A, El Khoury R, Lorber S, Oswald IP, El Khoury A, Atoui A, Puel O, Bailly JD. Aflatoxin biosynthesis and genetic regulation: a review. Toxins. 2020;12:150. doi: 10.3390/toxins12030150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen AJ, Frisvad JC, Sun BD, Varga J, Kocsubé S, Dijksterhuis J, Kim DH, Hong SB, Houbraken J, Samson RA. Aspergillus section Nidulantes (formerly Emericella): polyphasic taxonomy, chemistry and biology. Stud Mycol. 2016;84:1–118. doi: 10.1016/j.simyco.2016.10.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coppock RW, Christian RG, Jacobsen BJ (2018) Aflatoxins. In: Gupta RC (ed) Veterinary toxicology. Basic and clinical principles, 3rd edn. Academic Press, Amsterdam, p 983–994
- Detroy RW, Hesseltine CW. Aflatoxicol: structure of a new transformation product of aflatoxin B1. Can J Biochem. 1970;48:830–832. doi: 10.1139/o70-130. [DOI] [PubMed] [Google Scholar]
- DIN - Deutsches Institut für Normung eV (2011) Foodstuff - determination of aflatoxin B1, and the total content of aflatoxins B1, B2, G1 and G2 in cereals, nuts and derived products - high performance liquid chromatographic method (ISO 16050:2003); German version EN ISO 16050:2011
- Dohnal V, Wu Q, Kuca K. Metabolism of aflatoxins: key enzymes and interindividual as well as interspecies differences. Arch Toxicol. 2014;88:1635–1644. doi: 10.1007/s00204-014-1312-9. [DOI] [PubMed] [Google Scholar]
- Dutton MF, Ehrlich K, Bennett JW. Biosynthetic relationship among aflatoxins B1, B2, M1, and M2. Appl Environ Microbiol. 1985;49:1392–1395. doi: 10.1128/aem.49.6.1392-1395.1985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- EFSA – European Food Safety Authority, Panel on Contaminants in the Food Chain (2020) Scientific opinion – Risk assessment of aflatoxins in food. EFSA J 18:6040. Available from: 10.2903/j.efsa.2020.6040
- Ehrlich KC, Chang PK, Yu J, Cotty PJ. Aflatoxin biosynthesis cluster gene cypA is required for G aflatoxin formation. Appl Environ Microbiol. 2004;70:6518–6524. doi: 10.1128/AEM.70.11.6518-6524.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- EU – European Commission (2006) Commission regulation (EC) No 1881/2006 of 19 December 2006 setting maximum levels for certain contaminants in foodstuffs. Off J Eur Union L 364:5–24. Last consolidated version available from: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A02006R1881-20180319
- EU – European Commission (2010) Commission regulation (EC) No 165/2010 of 26 February 2010 amending regulation (EC) No 1881/2006 setting maximum levels for certain contaminants in foodstuffs as regards aflatoxins. Off J Eur Union L 50:8–12. Available from: https://eur-lex.europa.eu/legal-content/EN/ALL/?uri=CELEX%3A32010R0165
- EU – European Commission (2012) Commission regulation (EC) No 1058/2012 of 12 November 2012 amending regulation (EC) No 1881/2006 as regards maximum levels for aflatoxins in dried figs. Off J Eur Union L 313:14–15. Available from: https://eur-lex.europa.eu/legal-content/EN/ALL/?uri=CELEX%3A32012R1058
- Gauthier T, Duarte-Hospital C, Vignard J, Boutet-Robinet E, Sulyok M, Snini SP, Alassane-Kpembi I, Lippi Y, Puel S, Oswald IP, Puel O. Versicolorin A, a precursor in aflatoxins biosynthesis, is a food contaminant toxic for human intestinal cells. Environ Int. 2020;137:105568. doi: 10.1016/j.envint.2020.105568. [DOI] [PubMed] [Google Scholar]
- Getachew A, Chala A, Hofgaard IS, Brurberg MB, Sulyok M, Tronsmo AM. Multimycotoxin and fungal analysis of maize grains from south and southwestern Ethiopia. Food Addit Contam: B Surveill. 2018;11:64–74. doi: 10.1080/19393210.2017.1408698. [DOI] [PubMed] [Google Scholar]
- Houissa H, Lasram S, Sulyok M, Šarkanj B, Fontana A, Strub C, Krska R, Schorr-Galindo S, Ghorbel A. Multimycotoxin LC-MS/MS analysis in pearl millet (Pennisetum glaucum) from Tunisia. Food Control. 2019;106:106738. doi: 10.1016/j.foodcont.2019.106738. [DOI] [Google Scholar]
- IARC – International Agency for Research on Cancer (2012) Aflatoxins. In: Chemical agents and related occupations, Vol. 100F: a review of human carcinogens, p 225–248. Available from: https://monographs.iarc.who.int/wp-content/uploads/2018/06/mono100F.pdf
- Keller BO, Sui J, Young AB, Whittal RM. Interferences and contaminants encountered in modern mass spectrometry. Anal Chim Acta. 2008;627:71–81. doi: 10.1016/j.aca.2008.04.043. [DOI] [PubMed] [Google Scholar]
- Kensler TW, Roebuck BD, Wogan GN, Groopman JD. Aflatoxin: a 50-year odyssey of mechanistic and translational toxicology. Toxicol Sci. 2011;120(Suppl 1):S28–48. doi: 10.1093/toxsci/kfq283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kucukcakan B, Hayrulai-Musliu Z. Challenging role of dietary aflatoxin B1 exposure and hepatitis B infection on risk of hepatocellular carcinoma. Open Access Maced J Med Sci. 2015;3:363–369. doi: 10.3889/oamjms.2015.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matumba L, Sulyok M, Monjerezi M, Biswick T, Krska R. Fungal metabolites diversity in maize and associated human dietary exposures relate to micro-climatic patterns in Malawi. World Mycotoxin J. 2015;8:269–282. doi: 10.3920/WMJ2014.1773. [DOI] [Google Scholar]
- Matumba L, Sulyok M, Njoroge SMC, Njumbe Ediage E, Van Poucke C, De Saeger S, Krska R. Uncommon occurrence ratios of aflatoxin B1, B2, G1, and G2 in maize and groundnuts from Malawi. Mycotoxin Res. 2015;31:57–62. doi: 10.1007/s12550-014-0209-z. [DOI] [PubMed] [Google Scholar]
- Min L, Fink-Gremmels J, Li D, Tong X, Tang J, Nan X, Yu Z, Chen W, Wang G. An overview of aflatoxin B1 biotransformation and aflatoxin M1 secretion in lactating dairy cows. Anim Nutr. 2021;7:42–48. doi: 10.1016/j.aninu.2020.11.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mohammed S, Munissi JJE, Nyandoro SS. Aflatoxin M1 in raw milk and aflatoxin B1 in feed from household cows in Singida, Tanzania. Food Addit Contam: B Surveill. 2016;9:85–90. doi: 10.1080/19393210.2015.1137361. [DOI] [PubMed] [Google Scholar]
- Mori H, Sugie S, Yoshimi N, Kitamura J, Niwa M, Hamasaki T, Kawai K. Genotoxic effects of a variety of sterigmatocystin-related compounds in the hepatocyte/DNA-repair test and the Salmonella microsome assay. Mutat Res Lett. 1986;173:217–222. doi: 10.1016/0165-7992(86)90039-4. [DOI] [PubMed] [Google Scholar]
- Nakazato M, Morozumi S, Saito K, Fujinuma K, Nishima T, Kasai N (1991) Production of aflatoxins and aflatoxicols by Aspergillus flavus and Aspergillus parasiticus and metabolism of aflatoxin B1 by aflatoxin-non-producing Aspergillus flavus 37:107–116. 10.1248/jhs1956.37.107
- Pfeiffer E, Fleck SC, Metzler M. Catechol formation: a novel pathway in the metabolism of sterigmatocystin and 11-methoxysterigmatocystin. Chem Res Toxicol. 2014;27:2093–2099. doi: 10.1021/tx500308k. [DOI] [PubMed] [Google Scholar]
- Plattner RD, Bennett GA, Stubblefield RD. Identification of aflatoxins by quadrupole mass spectrometry/mass spectrometry. J Assoc off Anal Chem. 1984;67:734–738. doi: 10.1093/jaoac/67.4.734. [DOI] [PubMed] [Google Scholar]
- Ramachandra Pai M, Jayanthi Bai N, Venkitasubramanian TA. Production of aflatoxin M in a liquid medium. Appl Microbiol. 1975;29:850–851. doi: 10.1128/am.29.6.850-851.1975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raney KD, Gopalakrishnan S, Byrd S, Stone MP, Harris TM. Alteration of the aflatoxin cyclopentenone ring to a delta-lactone reduces intercalation with DNA and decreases formation of guanine N7 adducts by aflatoxin epoxides. Chem Res Toxicol. 1990;3:254–261. doi: 10.1021/tx00015a011. [DOI] [PubMed] [Google Scholar]
- Rank C, Nielsen KF, Larsen TO, Varga J, Samson RA, Frisvad JC. Distribution of sterigmatocystin in filamentous fungi. Fungal Microbiol. 2011;115:406–420. doi: 10.1016/j.funbio.2011.02.013. [DOI] [PubMed] [Google Scholar]
- Rodricks JV, Henery-Logan KR, Campbell AD, Stoloff L, Jaqueline Verrett M. Isolation of a new toxin from cultures of Aspergillus flavus. Nature. 1968;217:668. doi: 10.1038/217668a0. [DOI] [PubMed] [Google Scholar]
- Rodricks JV, Lustig E, Campbell AD, Stoloff L, Henery-Logan KR. Aspertoxin, a hydroxy derivative of O-methylsterigmatocystin from aflatoxin-producing cultures of Aspergillus flavus. Tetrahedron Lett. 1968;9:2975–2978. doi: 10.1016/S0040-4039(00)89626-4. [DOI] [PubMed] [Google Scholar]
- Schamann A, Geisen R, Schmidt-Heydt M. Draft genome sequence of an aflatoxin-producing Aspergillus flavus strain isolated from food. Microbiol Resour Announc. 2022;11:e00894–e921. doi: 10.1128/MRA.00894-21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soini Y, Chia SC, Bennett WP, Groopman JD, Wang JS, DeBenedetti VM, Cawley H, Welsh JA, Hansen C, Bergasa NV, Jones EA, DiBisceglie AM, Trivers GE, Sandoval CA, Calderon IE, Munoz Espinosa LE, Harris CC. An aflatoxin-associated mutational hotspot at codon 249 in the p53 tumor suppressor gene occurs in hepatocellular carcinomas from Mexico. Carcinogenesis. 1996;17:1007–1012. doi: 10.1093/carcin/17.5.1007. [DOI] [PubMed] [Google Scholar]
- Sulyok M, Krska R, Senyuva H. Profiles of fungal metabolites including regulated mycotoxins in individual dried Turkish figs by LC-MS/MS. Mycotoxin Res. 2020;36:381–387. doi: 10.1007/s12550-020-00398-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taniwaki MH, Pitt JI, Magan N. Aspergillus species and mycotoxins: occurrence and importance in major food commodities. Curr Opin Food Sci. 2018;23:38–43. doi: 10.1016/j.cofs.2018.05.008. [DOI] [Google Scholar]
- Theumer MG, Henneb Y, Khoury L, Snini SP, Tadrist S, Canlet C, Puel O, Oswald IP, Audebert M. Genotoxicity of aflatoxins and their precursors in human cells. Toxicol Lett. 2018;287:100–107. doi: 10.1016/j.toxlet.2018.02.007. [DOI] [PubMed] [Google Scholar]
- Ting WTE, Chang CH, Szonyi B, Gizachew D. Growth and aflatoxin B1, B2, G1, and G2 production by Aspergillus flavus and Aspergillus parasiticus on ground flax seeds (Linum usitatissimum) J Food Prot. 2020;83:975–983. doi: 10.4315/JFP-19-539. [DOI] [PubMed] [Google Scholar]
- Udwary DW, Casillas LK, Townsend CA. Synthesis of 11-hydroxyl O-methylsterigmatocystin and the role of a cytochrome P-450 in the final step of aflatoxin biosynthesis. J Am Chem Soc. 2002;124:5294–5303. doi: 10.1021/ja012185v. [DOI] [PubMed] [Google Scholar]
- Uka V, Moore GG, Arroyo-Manzanares N, Nebija D, De Saeger S, Diana Di Mavungu J. Secondary metabolite dereplication and phylogenetic analysis identify various emerging mycotoxins and reveal the high intra-species diversity in Aspergillus flavus. Front Microbiol. 2019;10:667. doi: 10.3389/fmicb.2019.00667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Waiss AC, Wiley M, Black DR, Lundin RE. 3-Hydroxy-6,7-dimethoxydifuroxanthone - a new metabolite from Aspergillus flavus. Tetrahedron Lett. 1968;9:3207–3210. doi: 10.1016/S0040-4039(00)89525-8. [DOI] [Google Scholar]
- Wehner FC, Thiel PG, van Rensburg SJ, Demasius IP. Mutagenicity to Salmonella typhimurium of some Aspergillus and Penicillium mycotoxins. Mutat Res Genet Toxicol. 1978;58:193–203. doi: 10.1016/0165-1218(78)90009-5. [DOI] [PubMed] [Google Scholar]
- Yabe K, Chihaya N, Hatabayashi H, Kito M, Hoshino S, Zeng H, Cai J, Nakajima H. Production of M-/GM-group aflatoxins catalyzed by the OrdA enzyme in aflatoxin biosynthesis. Fungal Genet Biol. 2012;49:744–754. doi: 10.1016/j.fgb.2012.06.011. [DOI] [PubMed] [Google Scholar]
- Yogendrarajah P, Devlieghere F, Njumbe Ediage E, Jacxsens L, De Meulenaer B, De Saeger S. Toxigenic potentiality of Aspergillus flavus and Aspergillus parasiticus strains isolated from black pepper assessed by an LC-MS/MS based multi-mycotoxin method. Food Microbiol. 2015;52:185–196. doi: 10.1016/j.fm.2015.07.016. [DOI] [PubMed] [Google Scholar]
- Yu J. Current understanding on aflatoxin biosynthesis and future perspective in reducing aflatoxin contamination. Toxins. 2012;4:1024–1057. doi: 10.3390/toxins4111024. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.