

Abstract
Severe acute respiratory syndrome coronavirus type 2 (SARS‐CoV‐2) causes coronavirus disease 2019 (COVID‐19), characterised by high levels of inflammation and oxidative stress (OS). Oxidative stress induces oxidative damage to lipids, proteins, and DNA, causing tissue damage. Both inflammation and OS contribute to multi‐organ failure in severe cases. Magnesium (Mg2+) regulates many processes, including antioxidant and anti‐inflammatory responses, as well as the proper functioning of other micronutrients such as vitamin D. In addition, Mg2+ participates as a second signalling messenger in the activation of T cells. Therefore, Mg2+ deficiency can cause immunodeficiency, exaggerated acute inflammatory response, decreased antioxidant response, and OS. Supplementation with Mg2+ has an anti‐inflammatory response by reducing the levels of nuclear factor kappa B (NF‐κB), interleukin (IL) ‐6, and tumor necrosis factor alpha. Furthermore, Mg2+ supplementation improves mitochondrial function and increases the antioxidant glutathione (GSH) content, reducing OS. Therefore, Mg2+ supplementation is a potential way to reduce inflammation and OS, strengthening the immune system to manage COVID‐19. This narrative review will address Mg2+ deficiency associated with a worse disease prognosis, Mg2+ supplementation as a potent antioxidant and anti‐inflammatory therapy during and after COVID‐19 disease, and suggest that randomised controlled trials are indicated.
Keywords: COVID‐19, inflammation, magnesium deficiency, oxidative stress, post‐COVID‐19 manifestations, SARS‐CoV‐2
Abbreviations
- 4‐HNE
4‐hydroxynonenal
- ACE2
angiotensin‐converting enzyme 2
- Ang II
angiotensin II
- ATP
adenosine triphosphate
- CAT
catalase
- COPD
chronic obstructive pulmonary disease
- COVID‐19
coronavirus disease 2019
- CRP
C‐reactive protein
- E protein
envelope protein
- eNOS
endothelial nitric oxide synthase
- ETC
electron transport chain
- GCL
γ‐glutamyl‐cysteine ligase
- GCSF
granulocyte‐colony stimulating factor
- GGT
γ‐glutamyl‐transpeptidase
- GPx
glutathione peroxidase
- GR
glutathione reductase
- GS
glutathione synthetase
- GSH
glutathione
- GST
glutathione S‐transferase
- H2O2
hydrogen peroxide
- ICU
intensive care unit
- IFN‐γ
interferon‐gamma
- IL
interleukin
- IP‐10
interferon‐γ‐inducible protein 10
- M protein
membrane protein
- MagT1
magnesium transporter 1
- MCP‐1
monocyte chemoattractant protein 1
- MDA
malondialdehyde
- Mg2+
magnesium
- MgSO4
magnesium sulphate
- MIP‐1A
macrophage inflammatory proteins
- N protein
nucleocapsid protein
- NF‐κB
nuclear factor kappa B
- NK
natural killer
- NKG2D
NK activator receptor
- non‐ICU
non‐intensive care unit
- NOXs
NADPH oxidases
- Nrf2
nuclear factor erythroid 2‐related factor 2
- nsps
non‐structural proteins
- O2
oxygen
- O2 ·−
superoxide radical
- OH
hydroxyl radical
- ORF
open reading frame
- OS
oxidative stress
- PAF
platelet‐activating factor
- pp
polyprotein
- RBD
receptor‐binding domain
- ROS
reactive oxygen species
- RTC
replication and transcription complex
- S protein
spike glycoprotein
- SARS
severe acute respiratory syndrome
- SARS‐CoV‐2
severe acute respiratory syndrome coronavirus type 2
- SOD
superoxide dismutase
- TCR
T cell receptor
- TF
tissue factor
- TMPRSS2
transmembrane protease serine 2
- TNF‐α
tumour necrosis factor‐alpha
- VDBP
vitamin D binding protein
- VDR
vitamin D receptor
- XMEN
X‐linked immunodeficiency with Mg2+ deficiency, Epstein‐Barr virus infection, and neoplasia
- XO
xanthine oxidase
1. INTRODUCTION
SARS‐CoV‐2 produces coronavirus disease 2019 (COVID‐19), which has caused more than 4.6 million deaths worldwide. 1 COVID‐19 is associated with inflammation and oxidative stress (OS) conditions, inducing respiratory and cardiac complications, such as respiratory insufficiency and arrhythmias. In addition, COVID‐19 patients also may submit minor complications like dry cough, fever, fatigue, sore throat, and diarrhoea. 2 , 3 , 4
Scientific publications have been shown that nutritional status and nutrition habits are relevant in developing different comorbidities associated with higher mortality in COVID‐19. 5 , 6 , 7 In this sense, magnesium (Mg2+) deficiency causes low‐grade chronic inflammation and OS. 8 , 9 , 10 It has led to the hypothesis that Mg2+ supplementation might improve the severity of COVID‐19 disease, 11 , 12 including a possible intervention of Mg2+ in SARS‐CoV‐2 infection by inhibiting the activity of proteases required for protein S cleavage. 12 Therefore, this narrative review focuses on the effect of Mg2+ to prevent developing severe COVID‐19 from the perspective of Mg2+ deficiency, which could contribute to an antioxidant/anti‐inflammatory development in COVID‐19 and post‐COVID‐19 manifestations. We also proposed that Mg2+ supplementation like a potential therapy might ameliorate COVID‐19 effects and post‐COVID‐19 sequels. Our exhaustive review of the scientific literature was conducted in the ‘PubMed databases’. Search keyword terms included all possible combinations, abbreviations, and synonyms between ‘magnesium’, ‘immune system’, ‘magnesium deficiency’, ‘magnesium supplementation’, ‘oxidative stress’, ‘inflammation’, ‘COVID‐19’, ‘SARS‐CoV‐2’, ‘thrombosis’, ‘post‐COVID‐19 manifestations’. We also considered the publication date from March 1971 to October 2021.
2. SARS‐CoV‐2 VIRAL LIFE CYCLE
The SARS‐CoV‐2 virus has been identified as a single‐stranded RNA‐enveloped (positive sense), spherical or pleomorphic beta coronavirus of the Coronaviridae family. SARS‐CoV‐2 infects lung and intestinal epithelial cells via the angiotensin‐converting enzyme 2 (ACE2) receptor, causing mild to moderate upper respiratory and gastrointestinal infections (Figure 1). 2 , 13 , 14 SARS‐CoV‐2 can also bind to the central nervous system cells by alternative receptor CD147, expressed in high levels in the brain, producing neurological symptoms such as headaches, vision changes, dizziness, ataxia, or impaired consciousness. 15
FIGURE 1.
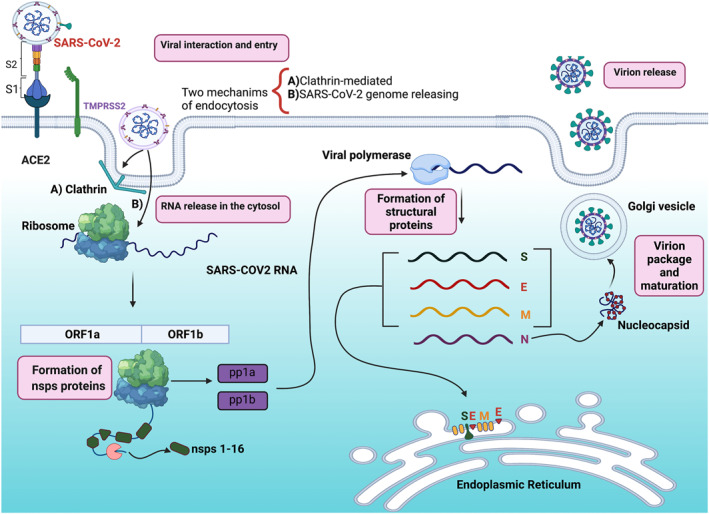
The viral life cycle of severe acute respiratory syndrome coronavirus type 2 (SARS‐COV‐2). SAR‐COV‐2 interacts with angiotensin‐converting enzyme 2 (ACE2) receptor and then the transmembrane protease serine 2 (TMPRSS2) and proprotein convertase furin primes S protein for entry into target cells. Two endocytosis mechanisms are known that SARS‐CoV‐2 uses for entrance to the cell: (a) clathrin‐mediated and (b) the releasing direct of its genome into the cytosol. Both mechanisms permit the viral genome to reach the cytosol, and once released, SARS‐CoV‐2 is translated into two open reading frames (ORF): ORF1a and ORF1b, promoting the expression of non‐structural proteins (nsps) 1–16, and the polyprotein 1a (pp1a) and pp1b. The latter allows the replication of the viral structural proteins: spike (S), envelope (e), the membrane (M), and the nucleocapsid (N). S, E, and M form the viral capsid, and N organises the nucleocapsid. Finally, the virion is packaged and released outer the infected cell. The figure was created with BioRender
SARS‐CoV‐2 virion has an outer surface with 24 to 40 spike glycoproteins (S protein; divided into S1 and S2), which fuse with different human cells such as nasal cavity cells. 16 The latter bidding is performed by the receptor‐binding domain (RBD) in the protein S1 subunit, which binds specifically at ACE2 (Figure 1). 17 ACE2 is the predominant host cell receptor, and this is the critical protein for SARS‐CoV‐2 to invade susceptible cells. 14 , 18 , 19 Moreover, SARS‐CoV‐2 employs the transmembrane protease serine 2 (TMPRSS2) and the proprotein convertase furin (host cell proteases) to primes the S protein, triggering viral envelope fusion with the host cell membrane. 20 , 21 Then, the virus enters basal cells by activating different endocytosis pathways, such as clathrin‐dependent endocytosis, or directly releasing the SARS‐CoV‐2 genome into the cytosol. 22 Both routes allow the viral genome to reach the cytosol, where the SARS‐CoV‐2 RNA genome unwraps from its viral envelope to translate the viral polyproteins (pp). 17 , 22
In the cytosol, the SARS‐CoV‐2 RNA genome is translated in two large open reading frames (ORF), ORF1a and ORF1b, inducing the expression of the individual non‐structural proteins (nsps) and polyprotein 1a (pp1a) and 1b (pp1ab). 17 , 23 The nsps comprise the viral replication and transcription complex (RTC) that includes RNA‐ processing. The nsps also reorganise the host membranes, where the SARS‐CoV‐2 RNA will replicate and structural viral proteins will be expressed. 23 Once the viral genome has been amplified, nucleocapsid proteins (N protein) encapsulate it, where membrane proteins (M protein) and envelope proteins (E protein) ensure SARS‐CoV‐2 incorporation in the viral particle during the assembly process (Figure 1). Finally, virions are secreted from the infected cell by exocytosis to attach to another cell surface. 17 , 24
3. INFLAMMATION, OXIDATIVE STRESS, AND COVID‐19
During SARS‐CoV‐2 viral life cycle, SARS‐CoV‐2 is exposed to the innate defence system, developing pronounced inflammation and, in acute cases, severe acute respiratory syndrome (SARS) or multi‐organ failure, called in general terms, COVID‐19. 25 , 26 During COVID‐19 and its pronounced inflammation increase the secretion of interleukin (IL) ‐1β, IL‐4, IL‐10, interferon‐gamma (IFN‐γ), interferon‐γ‐inducible protein 10 (IP‐10), and monocyte chemoattractant protein 1 (MCP‐1). 27 , 28 Furthermore, COVID‐19 produces several nuclear factor kappa B (NF‐κB)‐mediated cytokines, including IL‐6 and IL‐8 (Figure 2). 29 COVID‐19 also induces elevated plasma levels of pro‐inflammatory cytokines (tumour necrosis factor‐alpha (TNF‐α), IL‐2, IL‐6, and IL‐1β). 28 , 30 In addition, COVID‐19 patients from the intensive care unit (ICU) show elevated levels of IL‐7, IL‐10, MCP1, granulocyte‐colony stimulating factor (GCSF), IP‐10, and macrophage inflammatory proteins (MIP‐1A). 27 The latter promotes hyper‐inflammation, hyperpyrexia, and organ failure. 31 Organ failure results in respiratory failure, acute cardiac complications, respiratory distress syndrome, organ dysfunction, septic shock, and in critical cases, causes death. Therefore, organ failure spreads mortality risk. 32
FIGURE 2.
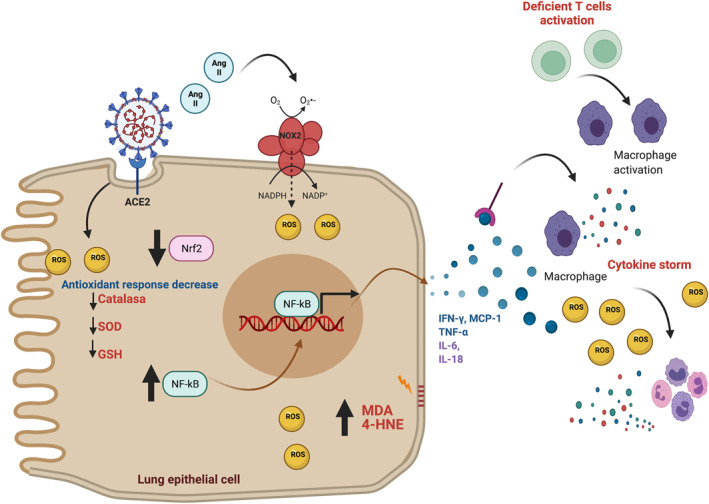
Inflammation and reactive oxygen species (ROS) overproduction during coronavirus disease 2019 (COVID‐19) infection. The interaction between angiotensin convertase enzyme 2 (ACE2) receptor and severe acute respiratory syndrome coronavirus type 2 (SARS‐COV‐2) generates ROS through angiotensin II (Ang II) because the latter stimulates NADPH oxidase 2 (NOX2). Moreover, the antioxidant response decreases through SARS‐CoV‐2 infection by lessening catalase, superoxide dismutase (SOD), and glutathione (GSH). ROS overproduction oxidises lipids in the cell membranes, generating the products of lipid peroxidation malondialdehyde (MDA) and 4‐hydroxinonenal (4‐HNE), which are increased In COVID‐19 deceased patients. On the other hand, SARS‐CoV‐2 activates the nuclear factor kappa B (NF‐κB), inducing the secretion of several cytokines and chemokines that include interferon‐gamma (IFN‐γ), tumoral necrosis factor‐alpha (TNF‐α), interleukin (IL) 6 (IL‐6), IL‐18, and monocyte chemoattractant protein 1 (MCP‐1). The latter and the deficient inactivation of T cells prompt macrophages activation, inducing the production of other cytokines, triggering cytokine storm accompanied by ROS overproduction. The figure was created with BioRender
T cell response in developing protective immunity is essential in regulating inflammation. For example, an adequate T cell response reduces the overactivation of the inflammatory response (Figure 2). On the other hand, the suppression or deficient T cell activity increases the burden on macrophages and monocytes, exacerbating the inflammatory process, distinctive of COVID‐19 (Figure 2). 33 , 34 , 35 Laboratory results showed decreased T helper lymphocytes (CD3+, CD4+) and suppressor T lymphocytes (CD3+, CD8+). These cells control infections and prevent overactivity of the immune system and uncontrolled virus infection. 30 Following the latter, Zhang et al. 26 reported an impaired immune response related to deficit T cell function in patients infected with SARS‐CoV‐2. Since there is an impaired T cell function, the inflammatory process exacerbates inflammation with an uncontrolled increase in levels of pro‐inflammatory cytokines and chemokines (cytokine storm), producing multi‐organ failure due to tissue damage. Therefore, the cytokine storm is associated with COVID‐19 severity. 35
Inflammation also produces reactive oxygen species (ROS), and if these ROS are not attenuated, OS is triggered, inducing oxidative damage to proteins, lipids, and DNA. Regarding lipids, its oxidation results in lipids radicals, such as malondialdehyde (MDA) and 4‐hydroxynonenal (4‐HNE), 36 which are highly reactive, causing DNA damage (Figure 2). The latter induces cell cycle arrest to permit DNA repair and proteostasis; however, if oxidative damage persists, apoptosis cell death is promoted. 37 A study in deceased COVID‐19 patients showed elevated 4‐HNE levels in the lungs, associated with lethal outcomes, suggesting that deceased COVID‐19 patients have a critical failure of the antioxidant response. 38 Furthermore, the MDA levels are increased in ICU and non‐intensive care unit (non‐ICU) patients, compared with healthy groups. 39 , 40 Therefore, these works suggest a close relationship between antioxidant response and COVID‐19 severity.
ROS overproduction during SARS‐CoV‐2 infection has been attributed to NADPH oxidases (NOXs) activation, principally NOX2. It has been shown that NOX2 is upregulated during COVID‐19 infection. 41 Supporting the latter, SARS‐CoV‐2 S protein together IL‐6 activate NOX2, producing high ROS levels in endothelial cells (Figure 2). 42 Moreover, NOX2 is stimulated by angiotensin II (Ang II), which plasma levels are elevated in COVID‐19. 43 Indeed, the upregulation of Ang II has been associated with the overstimulation of NOXs and the consequent production of ROS. 44 , 45
Although some antioxidant enzymes such as catalase (CAT) and superoxide dismutase (SOD) increase at the onset of COVID‐19 disease, suppressing the overproduction of ROS and OS, 39 in progressive stages of COVID‐19, these enzymes significantly decrease along with plasma levels of vitamins (A, C, E), glutathione (GSH), as well as cofactors of antioxidant enzymes like manganese and copper. 40 These data suggest that COVID‐19 patients are prone to a deficient antioxidant system during the progression of COVID‐19.
In conclusion, although the antioxidant and anti‐inflammatory responses are present at the beginning of SARS‐CoV‐2 infection, both are downregulated during COVID‐19 development, resulting in OS and inflammation. Therefore, the uncontrolled inflammatory and insufficient antioxidant response related to OS may be the principal mechanism in multi‐organ failure during COVID‐19.
4. MAGNESIUM, INFLAMMATION AND OXIDATIVE STRESS
Around 600 enzymes require Mg2+ as a cofactor, while other 200 enzymes need Mg2+ as an activator to realise their functions (Figure 3). 46 Thus, Mg2+ is crucial for energetic metabolism, protein, and amino acid synthesis, and maintenance of the electrical potential in tissues and cell membranes. 47 , 48 Mg2+ also participates in bone mineralisation, muscle relaxation, and neurotransmission (Figure 3). 49 In addition, Mg2+ regulates lipid composition, stabilising the cellular membrane and reducing its fluidity and permeability. 50 , 51 Furthermore, Mg2+ is also involved in most reactions in which adenosine triphosphate (ATP) functions as a cofactor. For example, ATP‐Mg2+ complexes are required for the activity of glycolytic enzymes such as hexokinase, phosphofructokinase, aldolase, phosphoglycerate kinase, and pyruvate kinase (Figure 3). 52
FIGURE 3.
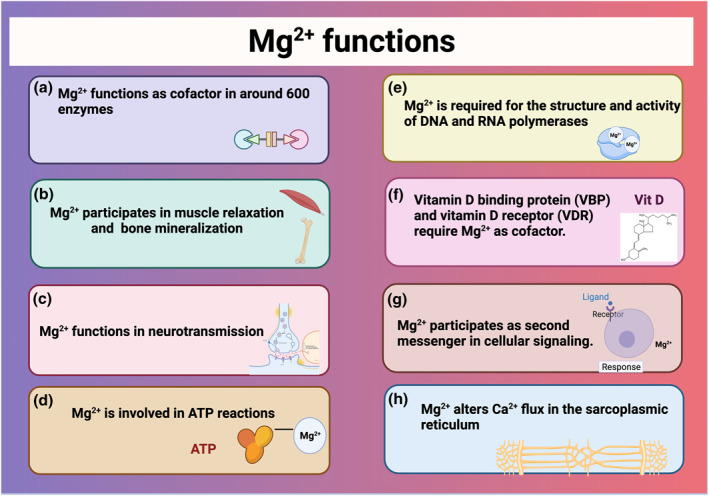
Functions of magnesium (Mg2+). In the figure, the roles of Mg2+ have been listed (A‐H)
Mg2+ is required for the structure and activity of DNA and RNA polymerases since it contains 2 Mg2+ binding sites, essential for conformational changes of the enzymes during catalytic reactions. 53 Vitamin D and vitamin D, enzymes responsible for vitamin D metabolism, require Mg2+ as a cofactor to bind to vitamin D (Figure 3). Mg2+ is also necessary for 25‐hydroxylation of vitamin D in the liver and 1α‐hydroxylation in the kidneys. 54 Mg2+ participates in different cell signal pathways, functioning as a second messenger. 55 , 56 Finally, Mg2+ alters Ca2+ flux in the sarcoplasmic reticulum, which modifies the permeability of the protons in the mitochondrial membrane, altering oxidative phosphorylation (Figure 3). 57
4.1. Magnesium and immune system
As described above, Mg2+ is involved in essential enzymatic reactions in the cells, including immune response. Mg2+ has a closer relationship in adaptative immunity, related to cellular signalling and immunomodulatory pathways. 52 , 56 , 58 Mg2+ has been described as a second signalling messenger in T cells, promoting their activation. 56 , 59 In individuals with X‐linked immunodeficiency with Mg2+ deficiency, Epstein‐Barr virus infection, and neoplasia (XMEN), the magnesium transporter 1 (MagT1) is downregulated in immune T cells. 56 Since MagT1 is essential for T cell receptor (TCR) stimulation and T cell activation, its downregulation is related to immunosuppression in XMEN patients. 60
The reduction of free intracellular Mg2+ causes defective expression of the natural killer (NK) activator receptor (NKG2D) on CD8+ T and NK cells, decreasing their cytolytic responses (Figure 4). 60 NK and CD8+ T cells' functions are essential for controlling viral infections because these cells induce apoptotic cell‐infected death, a regulated programed cell death that does not induce inflammation, and defects in this type of cell death might cause excessive viral load. 61 Since these cells are decreased, the innate immune cells such as macrophages and neutrophils are activated to control the infection, promoting exacerbated immune response by triggering cytokine storm (Figure 4). 60 , 61 In this understanding, strengthening NKs and T cells activation through Mg2+ supplementation could be associated with a better prognosis of COVID‐19. In contrast, Mg2+ deficiency may promote inflammation due to the deficient activation of cytolytic response in CD8+ T cells and NK cells. 56 , 59 , 60
FIGURE 4.
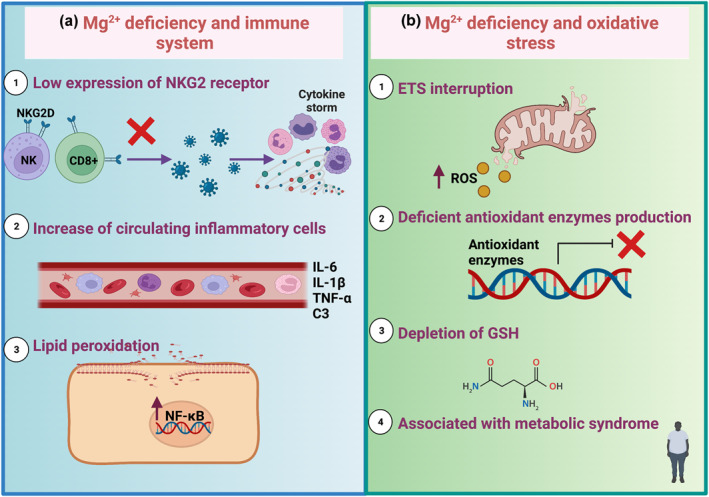
Magnesium (Mg2+), immune system, and oxidative stress (OS). (a) Mg2+ deficiency related to the immune system produces the (1) low expression of the natural killer (NK) activator receptor (NKG2D) on the T CD8+ cells, promoting a mild or no anti‐inflammatory response to viruses. Consequently, a cytokine storm is triggered. (2) Moreover, circulating inflammatory cells augment, inducing the production of interleukin (IL) 6 (IL‐6), IL‐1β, tumoral factor‐alpha (TNF‐α), and complement system C3. (3) Mg2+ deficiency induces lipid peroxidation, which activates nuclear factor kappa B (NF‐κB). Mg2+ also induces oxidative stress by promoting (1) electron transfer system (ETS) interruption, generating reactive oxygen species (ROS) production. Furthermore, (2) the production of antioxidant enzymes is deficient, and (3) glutathione (GSH) is depleted because of anormal Mg2+ levels. (4) Mg2+ deficiency is also associated with metabolic syndrome and low‐grade chronic inflammation, such as obesity, diabetes, and cardiovascular diseases
Different preclinic studies have also demonstrated that Mg2+ deficiency leads to exaggerated acute inflammatory response, such as increased circulating pro‐inflammatory cytokines (IL‐1β, IL‐6, TNF‐α), leucocytosis, increased plasma levels of complement component C3, and the marked elevation of circulating substance P, especially after immune stress (Figure 4). 9 , 62 , 63 , 64 , 65 , 66 Mg2+ deficiency induces the Novo synthesis of ceramide and lipid peroxidation, activating NF‐κB. 67 , 68 NF‐κB is involved in the transcription of inflammatory genes such as cytokines (IL‐1β and TNF‐α). 61 , 69 , 70 , 71 The latter means that Mg2+ deficiency can be related to inflammation by itself.
Imbalance Mg2+ levels are a decisive factor in mortality in COVID‐19 cases. A retrospective study by Alamdari et al. 3 found that older patients of Teheran Iran with hypomagnesaemia have a higher risk of mortality due to COVID‐19. Moreover, Quilliot et al. 72 found an association between Mg2+ and COVID‐19. This group conducted a cohort study, demonstrating low serum Mg2+ concentrations in hospitalised COVID‐19 adult patients. The serum concentrations of Mg2+ revealed that 73.7% of the patients had hypomagnesaemia, showing that most of the patients are located in serious to severe cases. 72 These studies reveal that hypomagnesaemia is more frequent in patients with COVID‐19, possibly associated with the severity of the disease. In this way, the severity of the COVID‐19 disease might be strongly related to the pro‐inflammatory state in Mg2+ deficiency patients.
Sugimoto et al. 73 reported that Mg2+ therapy during inflammatory states decreases NF‐κB, IL‐6, and TNF‐α. A similar beneficial effect is observed in the inflammatory marker C reactive protein (CRP), which levels are decreased. 74 Moreover, optimal Mg2+ status enhances vitamin D functionality that regulates inflammation by promoting an anti‐inflammatory effect. 75 , 76 Although the molecular mechanism of the relationship between Mg2+ and inflammation is poorly described, even more in COVID‐19, it is clear that during Mg2+ supplementation decreases inflammation markers.
Mg2+ could also indirectly be involved in the immune response by modulating the gut microbiome. For instance, it has been reported that diets containing Mg2+ can change microbiome composition. 77 , 78 , 79 , 80 , 81 , 82 , 83 In contrast, Mg2+ deficiency can lead to dysbiosis. 77 , 79 Dysbiosis refers to quantitative or qualitative changes in the composition of the normal microbiota that causes a microbial imbalance, playing an essential role in susceptibility to infectious diseases. 84 Many studies have associated altered gut microbiome with the severity of COVID‐19, producing dysbiosis 84 , 85 , 86 , 87 , 88 , 89 while restoring Mg2+ levels could benefit the diversity and health of the gut microbiome. Moreover, the use of probiotics and prebiotics for preventing and treating COVID‐19 could modulate the gut microbiome. 84 , 88 , 89 Therefore, Mg2+ usage could be able to mitigate systemic inflammation by regulating microbiome, preventing dysbiosis. 77 , 90 , 91 , 92
4.2. Magnesium and oxidative stress
Aerobic living systems use oxygen (O2) for many processes. For instance, in mitochondria, to produce energy in the form of ATP, O2 is the final electron acceptor in the electron transport chain (ETC). 93 However, electron leakage reduces O2 to superoxide radical (O2 ·−). It is the most reactive radical during this process. O2 ·− also be produced in cytosol by different enzymes such as NOXs or xanthine oxidase (XO) or in the endoplasmic reticulum by cytochrome p450 reductase or cytochrome b5. 36 The antioxidant system can reduce O2 ·− into hydrogen peroxide (H2O2) by different enzymes such as SOD or NOX4. 36 H2O2, in turn, is reduced to H2O by CAT, glutathione peroxidase (GPx), or glutathione reductase (GR), among others. O2 ·− and H2O2 can react together or with some metals in Fenton and Haber‐Weiss reaction inducing hydroxyl radical (•OH), a highly reactive ROS that generates different biomolecules damage. 36 Thus, these ROS have been maintained at low concentrations in a redox balance. Low ROS concentrations are associated with cellular signalling; however, if redox balance cannot be maintained, ROS levels increase induces OS and oxidative damage. 36 , 93 Therefore, the establishment of cellular antioxidant responses must be fast and efficient to neutralise the potential oxidant effects of ROS.
Low consumption and the consequent deficient status of Mg2+ have been associated with antioxidant response decrease, and, in consequence, OS increase (Figure 4). 8 , 58 , 94 , 95 Moreover, in a state of intracellular Mg2+ deficiency, a reverse flow of Mg2+ in the mitochondria is induced, promoting the ETS interruption and mitochondrial decoupling, which increases ROS production. 96 Adding to this increase of ROS, the low availability of Mg2+ decreases the production of antioxidant enzymes, leading to OS. 97 , 98 Mg2+ participates as an indirect antioxidant due to being a cofactor of enzymes that produce GSH, mitigating the effects of OS in the stabilisation of the cell membrane (Figure 4). 99 , 100 Also, during OS and proinflammatory states, the treatment with organic Mg2+ salt such as magnesium isoglycyrrhizinate and magnesium lithospermate B induces activation of nuclear factor erythroid 2‐related factor 2 (Nrf2). 101 , 102 , 103 This transcriptional factor modulates gene transcriptions and protein expressions of antioxidant enzymes such as CAT, SOD, and GPx. 101 , 104 Evidence shows that Mg2+ deficiency is associated with increased OS and cytokine due to antioxidant defences decrease in different cardiovascular diseases such as hypertension. 10 , 94 , 105 , 106 Furthermore, moderate or subclinical Mg2+ deficiency prepares phagocytic cells to release pro‐inflammatory cytokines that lead to chronic oxidative and inflammatory stress. 107 Therefore, Mg2+ deficient status reduces antioxidant defence and increases inflammation, producing OS.
Regarding GSH, a nonprotein tripeptide conformed by glycine, cysteine, and glutamate, is an essential and the highest ubiquitous intracellular antioxidant. 108 , 109 Therefore, it has multiple important biological functions, including maintaining the normal cellular oxidation‐reduction state, cell signalling, and antioxidant functions due to reducing particular ROS such as H2O2. 110 Also, GSH can maximise the activity of the other antioxidants such as vitamin C through glutathione S‐transferase (GST). GST catalyses the glutathione‐dependent reduction of dehydroascorbate, restoring their antioxidant function. 111 , 112 It has been shown that GSH depletion frequently occurs due to insufficient Mg2+ concentrations in acute deficiency. 94 , 105 , 106 That is, some enzymes that catalyse the biosynthesis reaction of GSH and maintain its correct operation require Mg2+. For example, γ‐glutamyl‐cysteine ligase (GCL) and glutathione synthetase (GS) are enzymes completely dependent on ATP, requiring Mg2+ as a cofactor. 105 , 113 , 114 Also, γ‐glutamyl‐transpeptidase (GGT) uses Mg2+ as an activator of this enzyme. 115 , 116 , 117 A study published by Mohammadi et al. 118 reported that magnesium sulphate (MgSO4) significantly decreased oxidative damage caused by hypoxia in mouse brains. In addition, Mg2+ supplementation improves mitochondrial function and increases the content of GSH in mitochondria. 118 , 119 Also, MgSO4 was effective as a treatment for preeclampsia, significantly promoting GSH production and thus suppressing ROS production. 120 As described in this section, Mg2+ deficiency represents a risk factor for maintaining an optimal oxidation‐reduction state, leading to OS development. Thus, chronic Mg2+ deficiency has severe oxidative implications such as lipid peroxidation, causing general cellular dysfunction and even cell apoptosis associated with inflammation and OS. 94 , 121
Mg2+ deficiency is frequently associated with is a strong relationship between OS and metabolic syndrome, associated with low‐grade chronic inflammation, such as obesity, diabetes, and cardiovascular diseases (Figure 4). 8 , 58 For instance, the increase in lipid peroxidation and OS was observed in a study of obese women with Mg2+ deficient diets, which presented low Mg2+ concentration in erythrocytes. 122
Because Mg2+ has multiple functions in the body, its deficiency has been related to chronic inflammatory and OS, which can compromise the immune response, inducing individuals more prone to infection such as SARS‐CoV2. Thus, nutritional supplementation may strengthen the immune system to manage COVID‐19.
5. MAGNESIUM CONSUMPTION DURING COVID‐19 AND POST‐COVID‐19 DISEASE
In addition to inflammatory and OS conditions, low Mg2+ consumption is associated with a higher incidence of diabetes and cardiovascular diseases. 96 , 123 , 124 , 125 , 126 Both diseases are associated with a worse prognosis in COVID‐19, according to the meta‐analysis published by Gold et al. 127 Furthermore, COVID‐19 patients with serum concentrations of Mg2+ ≤0.75 mM (low concentrations) are the most frequent hospital admissions. 3 , 72 Considering the above, we suggest that the frequent consumption of foods high in Mg2+ might prevent severe COVID‐19 symptoms. The diet can provide the primary source of Mg2+. 128 For instance, whole‐grain cereals (Mg2+ in pericarp) are considered the best dietary source of Mg2+; even part of the observed benefit with whole‐grain cereals intake is due to Mg2+. 129 , 130 Also, leafy‐greens foods such as chard, spinach, purslane (due to chlorophyll), and nuts are good sources of Mg2+. Some other foods consumed more frequently also have high levels of Mg2+, such as dark chocolate, black beans, soy nuts, and some other seeds (Table 1). 131 , 132
TABLE 1.
Foods and their Mg2+ content
| Food group | Food | Amount of Mg2+ (mg/100 g raw food) |
|---|---|---|
| Nut and seed | Pumpkin seeds | 550 |
| Chia seeds | 392 | |
| Brazil nuts | 376 | |
| Sesame seeds | 345 | |
| Cashew nuts | 292 | |
| Almond nuts | 270 | |
| Peanuts | 168 | |
| Legumes | Black beans (mature seeds) | 171 |
| Soy nuts | 145 | |
| Lentils | 47 | |
| Chickpeas | 45 | |
| Green peas | 33 | |
| Whole grain cereals | Oats | 138 |
| Natural puffed wheat | 133 | |
| Post shredded wheat | 132 | |
| Leafy greens | Chard | 81 |
| Spinach | 79 | |
| Purslane | 68 | |
| Kale | 33 | |
| Turnip greens | 31 | |
| Watercress | 21 | |
| Green cabbage | 12 | |
| Vegetables | Okra | 57 |
| Nopal | 52 | |
| Artichoke | 42 | |
| Sweet corn | 37 | |
| Acorn squash | 32 | |
| Potato | 23 | |
| Broccoli | 21 | |
| Summer squash | 18 | |
| Red cabbage | 16 | |
| Asparagus | 14 | |
| Turnip | 11 | |
| Fatty fish | Mackerel | 60 |
| Tuna | 35 | |
| Pink salmon fish | 27 | |
| Halibut | 23 | |
| Fruits | Avocado | 29 |
| Banana | 27 | |
| Guava | 22 | |
| Papaya | 21 | |
| Fig | 17 | |
| Kiwi fruit | 16 | |
| Cantaloupe | 13 | |
| Berries | 11 | |
| Grapefruit | 9 | |
| Others | Dark chocolate | 132 |
| Whole milk | 12 | |
| Liquid yogurt | 12 | |
| Whole egg | 12 |
Source: Collected data from U.S. Department of Agriculture‐FoodData central. 133
Kopf et al. 134 reported that the correct daily intake of fruit, vegetables, and whole grains significantly decreased levels in some inflammatory markers such as the lipopolysaccharide‐binding protein, TNF‐α, and, IL‐6. Therefore, the latter suggestion points out that increasing Mg2+ intake through food has favourable effects. Moreover, the severity of COVID‐19 cases are related to respiratory insufficiency or acute respiratory distress syndrome (indicator: low O2 saturation); coagulopathies (indicator: elevated D‐dimer, prothrombin time prolongation); inflammation (indicator: elevated CRP (an indicator of active acute inflammation), IL‐6); multi‐organ damage: liver damage (indicator: hypoalbuminemia), and lymphopenia. 3 , 72 , 135 Thus, these symptoms can be reduced by Mg2+ supplementation. For instance, it has been observed that concomitant supplementation of vitamin D, vitamin B12, and Mg2+ in COVID‐19 patients can decrease ICU admission incidence and O2 therapy requirements. 11
5.1. Respiratory insufficiency and magnesium supplementation
Inflammation markers in COVID‐19 severity could be directly related to serum Mg2+ levels. Chronic obstructive pulmonary disease (COPD) and COVID‐19 show high levels of systemic inflammatory markers, such as CRP, leucocytes, IL‐6, IL‐8, and fibrinogen. Both diseases also present the same mechanisms in endothelial dysfunction, including vascular inflammation and OS. Furthermore, both COPD and COVID‐19 reduce mediators that promote vasodilation such as endothelial nitric oxide synthase (eNOS). 28 , 30 , 38 , 136 , 137 , 138 , 139 , 140 , 141 It has been demonstrated that COPD severity was associated with serum Mg2+ deficiency and a worse quality of life in COPD patients. 142 In line with this, Mukerji et al. 143 demonstrated that intravenous adjuvant therapy with MgSO4 improves bronchodilator therapy in acute exacerbations of COPD. Additionally, intravenous MgSO4 as adjuvant therapy benefits moderate to severe acute asthma, a disease with high levels of inflammation and OS, by acting as a bronchodilator and improving pulmonary functions. 144 , 145 , 146 Since COPD, asthma and COVID‐19 present systemic inflammation, it is suggested that Mg2+ supplementation might enhance lung function in cases of respiratory failure in COVID‐19. Together these shreds of evidence suggest that complementary therapy with Mg2+ reduces the symptoms of inflammatory respiratory diseases and improves respiratory functions. Therefore, Mg2+ may be a potential therapy in cases of COVID‐19 respiratory failure due to exacerbated inflammation, preventing their development at severe.
Another aspect that may be related to respiratory failure is diaphragm dysfunction, implying a partial or complete diaphragm function loss. 147 , 148 McCool and Tzelepis 149 mentioned that ‘Diaphragmatic dysfunction is an underdiagnosed cause of dyspnoea.’ In this sense, a possible cause of respiratory failure in COVID‐19 may be diaphragm dysfunction; however, this area has been poorly explored. Interestingly, it has been reported that prolonged mechanical ventilation causes diaphragm dysfunction. 150 However, van Steveninck and Imming 151 reported a case of diaphragm dysfunction before intubation and mechanical ventilation in a COVID‐19 patient. This suggests that diaphragm dysfunction without mechanical ventilation or intubations may also be related to others processes, such as sepsis due to SARS‐CoV‐2. Sepsis is a massive inflammatory response characterised by OS, cytokines, and mitochondrial dysfunction. 152 , 153 In a prospective study, Demoule et al. 154 reported that sepsis contributes to the development of diaphragmatic dysfunction in ICU admissions. It has been shown that elevated TNF‐α and IL‐6 in diaphragm dysfunction is strongly associated with OS, driving muscle contractile dysfunction and atrophy. 155 , 156 , 157 Whidden et al. 155 demonstrated the use of Trolox (a potent antioxidant) attenuated diaphragmatic contractile dysfunction and prevented skeletal muscle loss, concluding that preventing OS could be an optimal strategy to avert diaphragm weakness. On the other hand, Jiang et al. 158 reported that MgSO4 protects against diaphragm dysfunction caused by sepsis, attenuating the loss of diaphragm force by reducing inflammation through TLR4/NF‐κB pathway inhibition. In this understanding, if diaphragm dysfunction caused by sepsis contributes to respiratory failure in COVID‐19, Mg2+ therapy could attenuate OS and inflammation, avoiding respiratory insufficiency.
5.2. Coagulopathies, inflammation, and OS: magnesium supplementation
During COVID‐19 infection, fibrinogen and circulating D‐dimer levels are elevated, inducing hypercoagulation. 159 Coagulation is a consequence of innate and adaptative immunity activation. It has been shown that inflammation‐induced coagulation is featured by tissue factor (TF) activation and upregulation of coagulant pathways, promoting thrombin production. 160 Although the mechanism by which a COVID‐19 induced coagulopathy development is not yet fully understood, it is supposed to have started mainly by TF activation. 61 , 161 , 162 TF is expressed in monocytes and endothelial cells in response to proinflammatory cytokines (i.e., TNF‐α, IL‐1, IL‐8, IL‐6, and MCP‐1), leading to the production of fibrinogen, fibrin, and thrombin. In turn, thrombin stimulates the production of the coagulation factors V, VIII, IX, and XII, resulting in thrombus formation. Increased plasma levels of D‐dimer are frequently used as an indicator of coagulation. Moreover, it has been demonstrated that high plasma levels of D‐dimer are related to coagulopathies in COVID‐19. 72 , 161 , 163 , 164 Thus, D‐dimer has been proposed to evaluate the severity of lung injury in COVID‐19. 161 , 162 Certainly, elevated D‐dimer levels have been associated with NF‐κB activation in COVID‐19 patients. 13 , 61 , 165 , 166 NF‐κB is the common link between inflammatory and thrombotic processes by increasing cytokines (TNF‐α, IL‐6 and MCP‐1), which activates the expression of TF, the main trigger of the coagulation cascade. 61 , 71 , 167 It has been shown that Mg2+ deficiency is associated with hypercoagulability and is partly mediated by excessive inflammation related to high levels of NF‐κB. 168 Interestingly, Mg2+ supplementation reduces NF‐κB expression and its activation, 73 , 101 which suggests that Mg2+ supplementation decreases cytokine production, deactivating the coagulopathy pathway. Moreover, in vitro studies show that Mg2+ inhibits clotting factors (prothrombin, thrombin, V, VII, IX) and reduces clotting blood. 169 , 170 , 171 Also, fibrin clots density and lysis time decreased with increasing Mg2+ concentrations. 170 , 172 Some authors also consider platelet‐activating factor (PAF) as key in generating microthrombosis in COVID‐19. 12 , 173 PAF has a central role in inflammation due to stimulating the activation of NF‐κB. Furthermore, it has been shown that the reduction of extracellular Mg2+ results in a rapid increase in active PAF, inducing the NF‐κB activation. 174 , 175 The above indicates a strong association between the Mg2+ deficiency and coagulopathies, where Mg2+ supplementation may be a potential therapy in coagulopathies associated with COVID‐19.
5.3. Post COVID‐19 manifestations
Lasting effects of illness have been reported post‐COVID‐19, scientific community named as ‘Long‐COVID’ to refer to the post‐COVID conditions or sequels. In a study, Kamal et al. 176 collected post‐COVID‐19 symptoms and diseases from COVID‐19 survivors, and the most common symptom was fatigue (72.8%). Other manifestations were mental disorders [anxiety (38%), dementia (28.6%), depression (28.6%), obsessive‐compulsive disorder (4.9%)], pain [joints pain (31.4%), continuous headache (28.9%), chest pain (28.9%)], dyspnoea (28.2%), blurred vision (17.1%), tinnitus (16.7%), and intermittent fever (11.1%). 176 Mandal et al. 177 reported in a post‐COVID‐19 follow‐up that 71.9% of the patients present cough, breathlessness, and fatigue. Carfi et al. 178 and Cares‐Marambio et al. 179 also reported that the main symptoms post‐COVID‐19 are fatigue, dyspnoea, chest pain, cough, psychological distress, and cognitive dysfunction. 176 , 177 , 178 , 179 The authors concluded that the severity of post‐COVID‐19 manifestations are related to the severity of COVID‐19. 176 , 177
According to a meta‐analysis by Iqbal et al. 180 the most prevalent symptoms that persist beyond 12 weeks in post‐COVID‐19 syndrome were fatigue, dyspnoea and sleep disturbance. Thus, fatigue is the most frequent manifestation post‐COVID‐19. Halpin et al. 181 reported that the fatigue in post‐COVID‐19 patients was not associated with old age, since no differences of age was found on patients in the ICU. The latter indicates that fatigue is not related to age, as might be thought. Chronic fatigue syndrome, which could be related to COVID‐19 fatigue, is characterised by low ATP levels and lipid peroxidation, indicating OS presence. Castro‐Marrero et al. 182 demonstrated that OS is related to mitochondrial dysfunction. Also, Myhill et al. 183 observed a strong association between mitochondrial dysfunction and severity of chronic fatigue syndrome through the diagnostic tool ‘ATP profile’ that helped differentiate fatigue due to energy wastage or cellular respiration disfunction. The mitochondrial provides energy, and when there exists mitochondrial dysfunction apart from low ATP levels, another consequence is the increase of ROS. In a preclinic study, Liu et al. 119 demonstrated that in diabetic mice with mitochondrial dysfunction, the mitochondrial function was improved with dietary Mg2+ supplementation. They proposed that the mechanism that justifies the mitochondrial function improvement is the alteration of mitochondrial Ca2+ homoeostasis. Since fatigue is related to mitochondrial dysfunction, Mg2+ supplementation could be a therapy for post‐COVID fatigue by improving this mitochondrial function.
Dyspnoea is the second most frequent symptom reported in post‐COVID‐19 patients. 176 , 177 , 178 , 179 , 180 Farr et al. 184 reported that patients who required mechanical ventilation during COVID‐19 have a high prevalence of diaphragm dysfunction. They suggested that diaphragm dysfunction significantly contributes to dyspnoea in post‐COVID‐19 patients. Persistent dyspnoea could be a consequence of respiratory tissue damage caused by OS since the COVID‐19 patients presented high levels of 4‐HNE. 38 , 185 Thus, we suggest that dyspnoea could be attenuated with Mg2+ supplementation since Mg2+ antioxidant and anti‐inflammation's functions.
6. CONCLUSIONS
COVID‐19 disease has been considered as an inflammatory disease in which OS occurs, leading to multi‐organ failure mainly due to vascular epithelial damage. Some antioxidant and anti‐inflammatory treatments are used to reduce damage, among them is Mg2+. Despite the little evidence of Mg2+ supplementation in COVID‐19 patients, the data indicate that Mg2+ supplementation decreases inflammation and OS in acute COVID‐19 cases, avoiding the progression of the disease. In addition, the incorporation of Mg2+ in the diet can prevent pro‐inflammatory and ‐OS, situations that make people susceptible to infections and states of exacerbated inflammation. Moreover, Mg2+ supplementation in post‐COVID‐19 may avoid fatigue and dyspnoea associated with inflammation and OS in post‐COVID‐19 patients. Randomised controlled trials to evaluate the effects of Mg2+ supplementation in COVID‐19 patients on coagulopathies, inflammatory and oxidative markers would be an excellent way to investigate the direct impact of Mg2+ on the disease.
CONFLICT OF INTEREST
All authors declare no conflict of interest.
AUTHOR CONTRIBUTIONS
Yalith Lyzet Arancibia‐Hernández, José Pedraza‐Chaverri, and Alfredo Cruz‐Gregorio: conceptualisation; investigation; writing‐original draft and writing – review and editing. Ana Karina Aranda‐Rivera: figures, writing – review, and editing.
ACKNOWLEDGEMENTS
We gratefully acknowledge to the Postdoctoral Grant's Programme from the Consejo Nacional de Ciencia y Tecnología (CONACyT) for the postdoctoral fellow position to Alfredo Cruz‐Gregorio. Ana Karina Aranda‐Rivera. Wants to thank CONACyT for supporting the doctoral scholarship that she is receiving. This research was partially supported by the CONACyT México, Grants Numbers A1‐S‐7495; by the Programa de Apoyo a Proyectos de Investigación e Innovación Tecnológica (PAPIIT), Grant Numbers IN202219 and IN200922 of the Universidad Nacional Autónoma de México (UNAM), and by the Programa de Apoyo a la Investigación y el Posgrado (PAIP), Grant Number 5000‐9105 of the UNAM.
Arancibia‐Hernández YL, Aranda‐Rivera AK, Cruz‐Gregorio A, Pedraza‐Chaverri J. Antioxidant/anti‐inflammatory effect of Mg2+ in coronavirus disease 2019 (COVID‐19). Rev Med Virol. 2022;32(5):e2348. 10.1002/rmv.2348
Contributor Information
Alfredo Cruz‐Gregorio, Email: cruzgalfredo@comunidad.unam.mx.
José Pedraza‐Chaverri, Email: pedraza@unam.mx.
DATA AVAILABILITY STATEMENT
Data is openly available in a public repository that issues datasets with https://pubmed.ncbi.nlm.nih.gov/.
REFERENCES
- 1. COVID‐19 Map. Johns Hopkins Coronavirus Resource Center. Accessed August 21, 2021. https://coronavirus.jhu.edu/map.html [Google Scholar]
- 2. Yang P, Wang X. COVID‐19: a new challenge for human beings. Cell Mol Immunol. 2020;17(5):555‐557. 10.1038/s41423-020-0407-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Alamdari NM, Afaghi S, Rahimi FS, et al. Mortality risk factors among hospitalized COVID‐19 patients in a referral center in Iran. Tohoku J Exp Med. 2020;252(1):73‐84. 10.1620/tjem.252.73 [DOI] [PubMed] [Google Scholar]
- 4.de; las Heras N, Martín Giménez VM, Ferder L, Manucha W, Lahera V. Implications of oxidative stress and potential role of mitochondrial dysfunction in COVID‐19: therapeutic effects of vitamin D. Antioxidants. 2020;9(9):897. 10.3390/antiox9090897 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Dizdar O, Baspınar O, Kocer D, et al. Nutritional risk, micronutrient status and clinical outcomes: a prospective observational study in an infectious disease clinic. Nutrients. 2016;8(3):124. 10.3390/nu8030124 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Calder P, Carr A, Gombart A, Eggersdorfer M. Optimal nutritional status for a well‐functioning immune system is an important factor to protect against viral infections. Nutrients. 2020;12(4):1181. 10.3390/nu12041181 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Gombart AF, Pierre A, Maggini S. A review of micronutrients and the immune system–working in harmony to reduce the risk of infection. Nutrients. 2020;12(1):236. 10.3390/nu12010236 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Nielsen FH. Magnesium, inflammation, and obesity in chronic disease: nutrition Reviews©. Nutr Rev. 2010;68(6):333‐340. 10.1111/j.1753-4887.2010.00293.x [DOI] [PubMed] [Google Scholar]
- 9. Maier JA, Castiglioni S, Locatelli L, Zocchi M, Mazur A. Magnesium and inflammation: advances and perspectives. Seminars Cell & Dev Biol. 2021;115:37‐44. 10.1016/j.semcdb.2020.11.002 [DOI] [PubMed] [Google Scholar]
- 10. Blache D, Devaux S, Joubert O, et al. Long‐term moderate magnesium‐deficient diet shows relationships between blood pressure, inflammation and oxidant stress defense in aging rats. Free Radic Biol Med. 2006;41(2):277‐284. 10.1016/j.freeradbiomed.2006.04.008 [DOI] [PubMed] [Google Scholar]
- 11. Tan CW, Ho LP, Kalimuddin S, et al. Cohort study to evaluate the effect of vitamin D, magnesium, and vitamin B12 in combination on progression to severe outcomes in older patients with coronavirus (COVID‐19). Nutrition. 2020;79:111017‐111080. 10.1016/j.nut.2020.111017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Trapani V, Rosanoff A, Baniasadi S, et al. The relevance of magnesium homeostasis in COVID‐19. Eur J Nutr. 2022;61(2):625‐636. 10.1007/s00394-021-02704-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Wallace TC. Combating COVID‐19 and building immune resilience: a potential role for magnesium nutrition? J Am Coll Nutr. 2020;39(8):685‐693. 10.1080/07315724.2020.1785971 [DOI] [PubMed] [Google Scholar]
- 14. Iotti S, Wolf F, Mazur A, Maier JA. The COVID‐19 pandemic: is there a role for magnesium? Hypotheses and perspectives. Magnesium Res. 2020;33(2):21‐27. 10.1684/mrh.2020.0465 [DOI] [PubMed] [Google Scholar]
- 15. Qiao J, Li W, Bao J, et al. The expression of SARS‐CoV‐2 receptor ACE2 and CD147, and protease TMPRSS2 in human and mouse brain cells and mouse brain tissues. Biochem Biophys Res Commun. 2020;533(4):867‐871. 10.1016/j.bbrc.2020.09.042 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Ke Z, Oton J, Qu K, et al. Structures and distributions of SARS‐CoV‐2 spike proteins on intact virions. Nature. 2020;588(7838):498‐502. 10.1038/s41586-020-2665-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. V’kovski P, Kratzel A, Steiner S, Stalder H, Thiel V. Coronavirus biology and replication: implications for SARS‐CoV‐2. Nat Rev Microbiol. 2021;19(3):155‐170. 10.1038/s41579-020-00468-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Xu H, Zhong L, Deng J, et al. High expression of ACE2 receptor of 2019‐nCoV on the epithelial cells of oral mucosa. Int J Oral Sci. 2020;12(1):8. 10.1038/s41368-020-0074-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Ciaglia E, Vecchione C, Puca AA. COVID‐19 infection and circulating ACE2 levels: protective role in women and children. Front Pediatr. 2020;8:206. 10.3389/fped.2020.00206 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Hoffmann M, Kleine‐Weber H, Schroeder S, et al. SARS‐CoV‐2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell. 2020;181(2):271‐280.e8. 10.1016/j.cell.2020.02.052 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Coutard B, Valle C, de Lamballerie X, Canard B, Seidah NG, Decroly E. The spike glycoprotein of the new coronavirus 2019‐nCoV contains a furin‐like cleavage site absent in CoV of the same clade. Antivir Res. 2020;176:104742. 10.1016/j.antiviral.2020.104742 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Bayati A, Kumar R, Francis V, McPherson PS. SARS‐CoV‐2 infects cells after viral entry via clathrin‐mediated endocytosis. J Biol Chem. 2021;296:100306. 10.1016/j.jbc.2021.100306 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Hartenian E, Nandakumar D, Lari A, Ly M, Tucker JM, Glaunsinger BA. The molecular virology of coronaviruses. J Biol Chem. 2020;295(37):12910‐12934. 10.1074/jbc.REV120.013930 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Klein S, Cortese M, Winter SL, et al. SARS‐CoV‐2 Structure and Replication Characterized by in Situ Cryo‐Electron Tomography; 2020. 10.1101/2020.06.23.167064 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Wong CK, Lam CWK, Wu AKL, et al. Plasma inflammatory cytokines and chemokines in severe acute respiratory syndrome. Clin Exp Immunol. 2004;136(1):95‐103. 10.1111/j.1365-2249.2004.02415.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Zhang Q, Meng Y, Wang K, et al. Inflammation and antiviral immune response associated with severe progression of COVID‐19. Front Immunol. 2021;12:135. 10.3389/fimmu.2021.631226 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Huang C, Wang Y, Li X, et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet. 2020;395(10223):497‐506. 10.1016/S0140-6736(20)30183-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Han H, Ma Q, Li C, et al. Profiling serum cytokines in COVID‐19 patients reveals IL‐6 and IL‐10 are disease severity predictors. Emerg Microbes Infect. 2020;9(1):1123‐1130. 10.1080/22221751.2020.1770129 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Yoshikawa T, Hill T, Li K, Peters CJ, Tseng CTK. Severe acute respiratory syndrome (SARS) coronavirus‐induced lung epithelial cytokines exacerbate SARS pathogenesis by modulating intrinsic functions of monocyte‐derived macrophages and dendritic cells. J Virol. 2009;83(7):3039‐3048. 10.1128/JVI.01792-08 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Qin C, Zhou L, Hu Z, et al. Dysregulation of immune response in patients with coronavirus 2019 (COVID‐19) in Wuhan, China. Clin Infect Dis. 2020;71(15):762‐768. 10.1093/cid/ciaa248 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Fu Y, Cheng Y, Wu Y. Understanding SARS‐CoV‐2‐mediated inflammatory responses: from mechanisms to potential therapeutic tools. Virol Sin. 2020;35(3):266‐271. 10.1007/s12250-020-00207-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Mehta P, McAuley DF, Brown M, Sanchez E, Tattersall RS, Manson JJ. COVID‐19: consider cytokine storm syndromes and immunosuppression. Lancet. 2020;395(10229):1033‐1034. 10.1016/S0140-6736(20)30628-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Merad M, Martin JC. Pathological inflammation in patients with COVID‐19: a key role for monocytes and macrophages. Nat Rev Immunol. May. 2020;6:1‐362. Published online. 10.1038/s41577-020-0331-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Payen D, Cravat M, Maadadi H, et al. A longitudinal study of immune cells in severe COVID‐19 patients. Front Immunol. 2020;11:580250. 10.3389/fimmu.2020.580250 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Liao M, Liu Y, Yuan J, et al. Single‐cell landscape of bronchoalveolar immune cells in patients with COVID‐19. Nat Med. 2020;26(6):842‐844. 10.1038/s41591-020-0901-9 [DOI] [PubMed] [Google Scholar]
- 36. Valko M, Leibfritz D, Moncol J, Cronin MTD, Mazur M, Telser J. Free radicals and antioxidants in normal physiological functions and human disease. Int J Biochem Cell Biol. 2007;39(1):44‐84. 10.1016/j.biocel.2006.07.001 [DOI] [PubMed] [Google Scholar]
- 37. Park J, Min JS, Kim B, et al. Mitochondrial ROS govern the LPS‐induced pro‐inflammatory response in microglia cells by regulating MAPK and NF‐κB pathways. Neurosci Lett. 2015;584:191‐196. 10.1016/j.neulet.2014.10.016 [DOI] [PubMed] [Google Scholar]
- 38. Žarković N, Orehovec B, Milković L, et al. Preliminary findings on the association of the lipid peroxidation product 4‐hydroxynonenal with the lethal outcome of aggressive COVID‐19. Antioxidants. 2021;10(9):1341. 10.3390/antiox10091341 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Mehri F, Rahbar AH, Ghane ET, Souri B, Esfahani M. The comparison of oxidative markers between Covid‐19 patients and healthy subjects. Arch Med Res. June 7, 2021. Published online. 10.1016/j.arcmed.2021.06.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Muhammad Y, Kani YA, Iliya S, et al. Deficiency of antioxidants and increased oxidative stress in COVID‐19 patients: a cross‐sectional comparative study in Jigawa, Northwestern Nigeria. SAGE Open Med. 2021;9:2050312121991246. 10.1177/2050312121991246 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Violi F, Oliva A, Cangemi R, et al. Nox2 activation in covid‐19. Redox Biol. 2020;36:101655. 10.1016/j.redox.2020.101655 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Youn JY, Zhang Y, Wu Y, Cannesson M, Cai H. Therapeutic application of estrogen for COVID‐19: attenuation of SARS‐CoV‐2 spike protein and IL‐6 stimulated, ACE2‐dependent NOX2 activation, ROS production and MCP‐1 upregulation in endothelial cells. Redox Biol. 2021;46:102099. 10.1016/j.redox.2021.102099 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Liu Y, Yang Y, Zhang C, et al. Clinical and biochemical indexes from 2019‐nCoV infected patients linked to viral loads and lung injury. Sci China Life Sci. 2020;63(3):364‐374. 10.1007/s11427-020-1643-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Dikalov SI, Nazarewicz RR. Angiotensin II‐induced production of mitochondrial reactive oxygen species: potential mechanisms and relevance for cardiovascular disease. Antioxid Redox Signal. 2013;19(10):1085‐1094. 10.1089/ars.2012.4604 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Daiber A. Redox signaling (cross‐talk) from and to mitochondria involves mitochondrial pores and reactive oxygen species. Biochimica Biophysica Acta (BBA) ‐ Bioenergetics. 2010;1797(6‐7):897‐906. 10.1016/j.bbabio.2010.01.032 [DOI] [PubMed] [Google Scholar]
- 46.de; Baaij JHF, Hoenderop JGJ, Bindels RJM. Regulation of magnesium balance: lessons learned from human genetic disease. Clin Kidney J. 2012;5(Suppl 1):i15‐i24. 10.1093/ndtplus/sfr164 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Crosby V, Elin RJ, Twycross R, Mihalyo M, Wilcock A. Magnesium. J Pain Symptom Manag. 2013;45(1):137‐144. 10.1016/j.jpainsymman.2012.10.005 [DOI] [PubMed] [Google Scholar]
- 48. Jahnen‐Dechent W, Ketteler M. Magnesium basics. Clin Kidney J. 2012;5(Suppl 1):i3‐i14. 10.1093/ndtplus/sfr163 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Vormann J. Magnesium: nutrition and homoeostasis. AIMS Public Health. 2016;3(2):329‐340. 10.3934/publichealth.2016.2.329 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Bara M, Guiet‐Bara A, Durlach J. Regulation of sodium and potassium pathways by magnesium in cell membranes. Magnes Res. 1993;6(2):167‐177. [PubMed] [Google Scholar]
- 51. Mahfouz MM, Smith TL, Kummerow FA. Changes in phosphofipid composition and calcium flux. Biochim Biophys Acta. 1989;1006(1):75‐83. 10.1016/0005-2760(89)90325-1 [DOI] [PubMed] [Google Scholar]
- 52.de; Baaij JHF, Hoenderop JGJ, Bindels RJM. Magnesium in man: implications for health and disease. Physiol Rev. 2015;95(1):1‐46. 10.1152/physrev.00012.2014 [DOI] [PubMed] [Google Scholar]
- 53. Yang L, Arora K, Beard WA, Wilson SH, Schlick T. Critical role of magnesium ions in DNA polymerase β’s closing and active site assembly. J Am Chem Soc. 2004;126(27):8441‐8453. 10.1021/ja049412o [DOI] [PubMed] [Google Scholar]
- 54. Uwitonze AM, Razzaque MS. Role of magnesium in vitamin D activation and function. J Am Osteopath Assoc. 2018;118(3):181‐189. 10.7556/jaoa.2018.037 [DOI] [PubMed] [Google Scholar]
- 55. Stangherlin A, O’Neill JS. Signal transduction: magnesium manifests as a second messenger. Curr Biol. 2018;28(24):R1403‐R1405. 10.1016/j.cub.2018.11.003 [DOI] [PubMed] [Google Scholar]
- 56. Li FY, Chaigne‐Delalande B, Kanellopoulou C, et al. Second messenger role for Mg2+ revealed by human T‐cell immunodeficiency. Nature. 2011;475(7357):471‐476. 10.1038/nature10246 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Rodríguez‐Zavala JS, Moreno‐Sánchez R. Modulation of oxidative phosphorylation by Mg2+ in rat heart mitochondria. J Biol Chem. 1998;273(14):7850‐7855. 10.1074/jbc.273.14.7850 [DOI] [PubMed] [Google Scholar]
- 58. Shahi A, Aslani S, Ataollahi M, Mahmoudi M. The role of magnesium in different inflammatory diseases. Inflammopharmacol. 2019;27(4):649‐661. 10.1007/s10787-019-00603-7 [DOI] [PubMed] [Google Scholar]
- 59. Brandao K, Deason‐Towne F, Perraud AL, Schmitz C. The role of Mg2+ in immune cells. Immunol Res. 2013;55(1):261‐269. 10.1007/s12026-012-8371-x [DOI] [PubMed] [Google Scholar]
- 60. Chaigne‐Delalande B, Li FY, O’Connor GM, et al. Mg2+ regulates cytotoxic functions of NK and CD8 T cells in chronic EBV infection through NKG2D. Science. 2013;341(6142):186‐191. 10.1126/science.1240094 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. DiNicolantonio JJ, O’Keefe JH. Magnesium and vitamin D deficiency as a potential cause of immune dysfunction, cytokine storm and disseminated intravascular coagulation. Mo Med. 2021;118(1):68‐73. [PMC free article] [PubMed] [Google Scholar]
- 62. Rude RK, Gruber HE, Wei LY, Frausto A, Mills BG. Magnesium deficiency: effect on bone and mineral metabolism in the mouse. Calcif Tissue Int. 2003;72(1):32‐41. 10.1007/s00223-001-1091-1 [DOI] [PubMed] [Google Scholar]
- 63. Castiglioni S, Cazzaniga A, Locatelli L, Maier JA. Burning magnesium, a sparkle in acute inflammation: gleams from experimental models. Magnesium Res. 2017;30(1):8‐15. 10.1684/mrh.2017.0418 [DOI] [PubMed] [Google Scholar]
- 64. Malpuech‐Brugère C, Nowacki W, Daveau M, et al. Inflammatory response following acute magnesium deficiency in the rat. Biochimica Biophysica Acta (BBA) ‐ Mol Basis Dis. 2000;1501(2):91‐98. 10.1016/S0925-4439(00)00018-1 [DOI] [PubMed] [Google Scholar]
- 65. Weglicki WB, Mak IT, Chmielinska JJ, Tejero‐Taldo MI, Komarov AM, Kramer JH. The role of magnesium deficiency in cardiovascular and intestinal inflammation. Magnes Res. 2010;23(4):S199‐S206. 10.1684/mrh.2010.0218 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66. Bussière FI, Tridon A, Zimowska W, Mazur A, Rayssiguier Y. Increase in complement component C3 is an early response to experimental magnesium deficiency in rats. Life Sci. 2003;73(4):499‐507. 10.1016/S0024-3205(03)00291-1 [DOI] [PubMed] [Google Scholar]
- 67. Altura BM, Shah NC, Shah G, et al. Short‐term magnesium deficiency upregulates ceramide synthase in cardiovascular tissues and cells: cross‐talk among cytokines, Mg2+, NF‐κB, and de novo ceramide. Am J Physiology‐Heart Circulatory Physiology. 2012;302(1):H319‐H332. 10.1152/ajpheart.00453.2011 [DOI] [PubMed] [Google Scholar]
- 68. Altura BM, Gebrewold A, Zhang A, Altura BT. Low extracellular magnesium ions induce lipid peroxidation and activation of nuclear factor‐kappa B in canine cerebral vascular smooth muscle: possible relation to traumatic brain injury and strokes. Neurosci Lett. 2003;341(3):189‐192. 10.1016/S0304-3940(03)00134-4 [DOI] [PubMed] [Google Scholar]
- 69. Mitchell S, Vargas J, Hoffmann A. Signaling via the NFκB system. WIREs Syst Biol Med. 2016;8(3):227‐241. 10.1002/wsbm.1331 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70. Zhang FX, Kirschning CJ, Mancinelli R, et al. Bacterial lipopolysaccharide activates nuclear factor‐κB through interleukin‐1 signaling mediators in cultured human dermal endothelial cells and mononuclear phagocytes. J Biol Chem. 1999;274(12):7611‐7614. 10.1074/jbc.274.12.7611 [DOI] [PubMed] [Google Scholar]
- 71. Mussbacher M, Salzmann M, Brostjan C, et al. Cell type‐specific roles of NF‐κB linking inflammation and thrombosis. Front Immunol. 2019;10:85. 10.3389/fimmu.2019.00085 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72. Quilliot D, Bonsack O, Jaussaud R, Mazur A. Dysmagnesemia in Covid‐19 cohort patients: prevalence and associated factors. Magnesium Res. 2020;33(4):114‐122. 10.1684/mrh.2021.0476 [DOI] [PubMed] [Google Scholar]
- 73. Sugimoto J, Romani AM, Valentin‐Torres AM, et al. Magnesium decreases inflammatory cytokine production: a novel innate immunomodulatory mechanism. J Immunol. 2012;188(12):6338‐6346. 10.4049/jimmunol.1101765 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74. Simental‐Mendia LE, Sahebkar A, Rodriguez‐Moran M, Zambrano‐Galvan G, Guerrero‐Romero F. Effect of magnesium supplementation on plasma C‐reactive protein concentrations: a systematic review and meta‐analysis of randomized controlled trials. CPD. 2017;23(31). 10.2174/1381612823666170525153605 [DOI] [PubMed] [Google Scholar]
- 75. Dai Q, Zhu X, Manson JE, et al. Magnesium status and supplementation influence vitamin D status and metabolism: results from a randomized trial. Am J Clin Nutr. 2018;108(6):1249‐1258. 10.1093/ajcn/nqy274 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76. Reddy P, Edwards LR. Magnesium supplementation in vitamin D deficiency. Am J Ther. 2019;26(1):e124‐e132. 10.1097/MJT.0000000000000538 [DOI] [PubMed] [Google Scholar]
- 77. Pachikian BD, Neyrinck AM, Deldicque L, et al. Changes in intestinal bifidobacteria levels are associated with the inflammatory response in magnesium‐deficient mice. J Nutr. 2010;140(3):509‐514. 10.3945/jn.109.117374 [DOI] [PubMed] [Google Scholar]
- 78. García‐Legorreta A, Soriano‐Pérez LA, Flores‐Buendía AM, et al. Effect of dietary magnesium content on intestinal microbiota of rats. Nutrients. 2020;12(9):2889. 10.3390/nu12092889 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79. Gommers LMM, Ederveen THA, Wijst J, et al. Low gut microbiota diversity and dietary magnesium intake are associated with the development of PPI‐induced hypomagnesemia. FASEB j. 2019;33(10):11235‐11246. 10.1096/fj.201900839R [DOI] [PubMed] [Google Scholar]
- 80. Jørgensen BP, Winther G, Kihl P, et al. Dietary magnesium deficiency affects gut microbiota and anxiety‐like behaviour in C57BL/6N mice. Acta Neuropsychiatr. 2015;27(5):307‐311. 10.1017/neu.2015.10 [DOI] [PubMed] [Google Scholar]
- 81. Winther G, Pyndt Jørgensen BM, Elfving B, et al. Dietary magnesium deficiency alters gut microbiota and leads to depressive‐like behaviour. Acta Neuropsychiatr. 2015;27(3):168‐176. 10.1017/neu.2015.7 [DOI] [PubMed] [Google Scholar]
- 82. Barnich N, Rodrigues M, Sauvanet P, et al. Beneficial effects of natural mineral waters on intestinal inflammation and the mucosa‐associated microbiota. Int J Mol Sci. 2021;22(9):4336. 10.3390/ijms22094336 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83. Crowley EK, Long‐Smith CM, Murphy A, et al. Dietary supplementation with a magnesium‐rich marine mineral blend enhances the diversity of gastrointestinal microbiota. Mar Drugs. 2018;16(6):216. 10.3390/md16060216 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84. Olaimat AN, Aolymat I, Al‐Holy M, et al. The potential application of probiotics and prebiotics for the prevention and treatment of COVID‐19. npj Sci Food. 2020;4(1):17. 10.1038/s41538-020-00078-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85. Yeoh YK, Zuo T, Lui GCY, et al. Gut microbiota composition reflects disease severity and dysfunctional immune responses in patients with COVID‐19. Gut. 2021;70(4):698‐706. 10.1136/gutjnl-2020-323020 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86. Kim HS. Do an altered gut microbiota and an associated leaky gut affect COVID‐19 severity? mBio. 2021;12(1):e03022‐20. 10.1128/mBio.03022-20 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87. Hirayama M, Nishiwaki H, Hamaguchi T, et al. Intestinal Collinsella may mitigate infection and exacerbation of COVID‐19 by producing ursodeoxycholate. PLoS One. 2021;16(11):e0260451. 10.1371/journal.pone.0260451 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88. Gasmi A, Tippairote T, Mujawdiya PK, et al. The microbiota‐mediated dietary and nutritional interventions for COVID‐19. Clin Immunol. 2021;226:108725. 10.1016/j.clim.2021.108725 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89. Walton GE, Gibson GR, Hunter KA. Mechanisms linking the human gut microbiome to prophylactic and treatment strategies for COVID‐19. Br J Nutr. October 2020;126:1‐227. Published online. 10.1017/S0007114520003980 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90. Del Chierico F, Trapani V, Petito V, et al. Dietary magnesium alleviates experimental murine colitis through modulation of gut microbiota. Nutrients. 2021;13(12):4188. 10.3390/nu13124188 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91. Zhao J, Zhang Qli, Shen Jhua, Wang K, Liu J. Magnesium lithospermate B improves the gut microbiome and bile acid metabolic profiles in a mouse model of diabetic nephropathy. Acta Pharmacol Sin. 2019;40(4):507‐513. 10.1038/s41401-018-0029-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92. Liu C, Cheng Y, Guo Y, Qian H. Magnesium‐L‐threonate alleviate colonic inflammation and memory impairment in chronic‐plus‐binge alcohol feeding mice. Brain Res Bull. 2021;174:184‐193. 10.1016/j.brainresbull.2021.06.009 [DOI] [PubMed] [Google Scholar]
- 93. Holmström KM, Finkel T. Cellular mechanisms and physiological consequences of redox‐dependent signalling. Nat Rev Mol Cell Biol. 2014;15(6):411‐421. 10.1038/nrm3801 [DOI] [PubMed] [Google Scholar]
- 94. Wiles ME, Wagner TL, Weglicki WB. Effect of acute magnesium deficiency (MgD) on aortic endothelial cell (EC) oxidant production. Life Sci. 1996;60(3):221‐236. 10.1016/S0024-3205(96)00619-4 [DOI] [PubMed] [Google Scholar]
- 95. Nielsen FH, Milne DB, Klevay LM, Gallagher S, Johnson L. Dietary magnesium deficiency induces heart rhythm changes, impairs glucose tolerance, and decreases serum cholesterol in post menopausal women. J Am Coll Nutr. 2007;26(2):121‐132. 10.1080/07315724.2007.10719593 [DOI] [PubMed] [Google Scholar]
- 96. Liu M, Dudley SC. Magnesium, oxidative stress, inflammation, and cardiovascular disease. Antioxidants. 2020;9(10):907. 10.3390/antiox9100907 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97. Touyz RM. Transient receptor potential melastatin 6 and 7 channels, magnesium transport, and vascular biology: implications in hypertension. Am J Physiology‐Heart Circulatory Physiology. 2008;294(3):H1103‐H1118. 10.1152/ajpheart.00903.2007 [DOI] [PubMed] [Google Scholar]
- 98. Beltrán‐García J, Osca‐Verdegal R, Pallardó FV, et al. Oxidative stress and inflammation in COVID‐19‐associated sepsis: the potential role of anti‐oxidant therapy in avoiding disease progression. Antioxidants. 2020;9(10):936. 10.3390/antiox9100936 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99. Abad C, Vargas FR, Zoltan T, et al. Magnesium sulfate affords protection against oxidative damage during severe preeclampsia. Placenta. 2015;36(2):179‐185. 10.1016/j.placenta.2014.11.008 [DOI] [PubMed] [Google Scholar]
- 100. Morais JBS, Severo JS, Santos LRdos, et al. Role of magnesium in oxidative stress in individuals with obesity. Biol Trace Elem Res. 2017;176(1):20‐26. 10.1007/s12011-016-0793-1 [DOI] [PubMed] [Google Scholar]
- 101. Orhan C, Er B, Deeh PBD, et al. Different sources of dietary magnesium supplementation reduces oxidative stress by regulation Nrf2 and NF‐κB signaling pathways in high‐fat diet rats. Biol Trace Elem Res. 2021;199(11):4162‐4170. 10.1007/s12011-020-02526-9 [DOI] [PubMed] [Google Scholar]
- 102. Gao F, Limeng J, Xi C, et al. Magnesium lithospermate B protects the endothelium from inflammation‐induced dysfunction through activation of Nrf2 pathway. Acta Pharmacol Sin. 2019;40(7):867‐878. 10.1038/s41401-018-0189-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103. Tan QY, Hu Q, Zhu SN, et al. Licorice root extract and magnesium isoglycyrrhizinate protect against triptolide‐induced hepatotoxicity via up‐regulation of the Nrf2 pathway. Drug Deliv 2018;25(1):1213‐1223. 10.1080/10717544.2018.1472676 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104. Abreu CC, Cardozo LFMF, Stockler‐Pinto MB, et al. Does resistance exercise performed during dialysis modulate Nrf2 and NF‐κB in patients with chronic kidney disease? Life Sci. 2017;188:192‐197. 10.1016/j.lfs.2017.09.007 [DOI] [PubMed] [Google Scholar]
- 105. Minnich V, Smith MB, Brauner MJ, Majerus PW. Glutathione biosynthesis in human erythrocytes: I. Identification of the enzymes of glutathione synthesis in hemolysates. J Clin Invest. 1971;50(3):507‐513. 10.1172/JCI106519 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106. Hsu JM, Rubenstein B, Paleker AG. Role of magnesium in glutathione metabolism of rat erythrocytes. J Nutr. 1982;112(3):488‐496. 10.1093/jn/112.3.488 [DOI] [PubMed] [Google Scholar]
- 107. Nielsen FH. Dietary magnesium and chronic disease. Adv Chronic Kidney Dis. 2018;25(3):230‐235. 10.1053/j.ackd.2017.11.005 [DOI] [PubMed] [Google Scholar]
- 108. Franco R, Cidlowski JA. Apoptosis and glutathione: beyond an antioxidant. Cell Death Differ. 2009;16(10):1303‐1314. 10.1038/cdd.2009.107 [DOI] [PubMed] [Google Scholar]
- 109. Oestreicher J, Morgan B. Glutathione: subcellular distribution and membrane transport. Biochem Cell Biol. 2019;97(3):270‐289. 10.1139/bcb-2018-0189 [DOI] [PubMed] [Google Scholar]
- 110. Sies H, Jones DP. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nat Rev Mol Cell Biol. 2020;21(7):363‐383. 10.1038/s41580-020-0230-3 [DOI] [PubMed] [Google Scholar]
- 111. Riso P, Del Bo’ C, Vendrame S, et al. Modulation of plasma antioxidant levels, glutathione S‐transferase activity and DNA damage in smokers following a single portion of broccoli: a pilot study. J Sci Food Agric. 2014;94(3):522‐528. 10.1002/jsfa.6283 [DOI] [PubMed] [Google Scholar]
- 112. Schmuck EM, Board PG, Whitbread AK, et al. Characterization of the monomethylarsonate reductase and dehydroascorbate reductase activities of Omega class glutathione transferase variants: implications for arsenic metabolism and the age‐at‐onset of Alzheimer’s and Parkinson’s diseases. Pharmacogenetics Genomics. 2005;15(7):493‐501. 10.1097/01.fpc.0000165725.81559.e3 [DOI] [PubMed] [Google Scholar]
- 113. BRENDA ‐ Information on EC 6.3.2.2 ‐ Glutamate‐Cysteine Ligase. Accessed October 2, 2021. https://www.brenda‐enzymes.org/enzyme.php?ecno=6.3.2.2 [Google Scholar]
- 114. BRENDA ‐ Information on EC 6.3.2.3 ‐ Glutathione Synthase. Accessed October 2, 2021. https://www.brenda‐enzymes.org/enzyme.php?ecno=6.3.2.3 [Google Scholar]
- 115. Zhang H, Jay Forman H, Choi J. γ‐Glutamyl transpeptidase in glutathione biosynthesis. In: Methods in Enzymology. 401. Elsevier; 2005:468‐483. 10.1016/S0076-6879(05)01028-1 [DOI] [PubMed] [Google Scholar]
- 116. Ige AO, Adewoye EO, Makinde EO. Oral magnesium potentiates glutathione activity in experimental diabetic rats. Int J Diab Res. 2016;5(2):21‐25. [Google Scholar]
- 117. BRENDA ‐ Information on EC 2.3.2.2 ‐ Gamma‐Glutamyltransferase. Accessed October 2, 2021. https://www.brenda‐enzymes.org/enzyme.php?ecno=2.3.2.2#GENERAL%20INFORMATION [Google Scholar]
- 118. Mohammadi H, Shamshirian A, Eslami S, Shamshirian D, Ebrahimzadeh MA. Magnesium sulfate attenuates lethality and oxidative damage induced by different models of hypoxia in mice. In: Bunevicius A, ed. BioMed Res Int. 2020; 2020:1‐8. 10.1155/2020/2624734 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119. Liu M, Jeong EM, Liu H, et al. Magnesium supplementation improves diabetic mitochondrial and cardiac diastolic function. JCI Insight. 2019;4(1):e123182. 10.1172/jci.insight.123182 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120. Kawasaki K, Kondoh E, Chigusa Y, et al. Metabolomic profiles of placenta in preeclampsia. Hypertension. 2019;73(3):671‐679. 10.1161/HYPERTENSIONAHA.118.12389 [DOI] [PubMed] [Google Scholar]
- 121. Martin H, Uring‐Lambert B, Adrian M, et al. Effects of long‐term dietary intake of magnesium on oxidative stress, apoptosis and ageing in rat liver. Magnes Res. 2008;21(2):124‐130. [PubMed] [Google Scholar]
- 122. Morais JBS, Severo JS, de Oliveira ARS, et al. Magnesium status and its association with oxidative stress in obese women. Biol Trace Elem Res. 2017;175(2):306‐311. 10.1007/s12011-016-0797-x [DOI] [PubMed] [Google Scholar]
- 123. Fang X, Han H, Li M, et al. Dose‐response relationship between dietary magnesium intake and risk of type 2 diabetes mellitus: a systematic review and meta‐regression analysis of prospective cohort studies. Nutrients. 2016;8(11):739. 10.3390/nu8110739 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124. Abbott RD, Ando F, Masaki KH, et al. Dietary magnesium intake and the future risk of coronary heart disease (The Honolulu Heart Program). Am J Cardiol. 2003;92(6):665‐669. 10.1016/S0002-9149(03)00819-1 [DOI] [PubMed] [Google Scholar]
- 125. Fang X, Wang K, Han D, et al. Dietary magnesium intake and the risk of cardiovascular disease, type 2 diabetes, and all‐cause mortality: a dose–response meta‐analysis of prospective cohort studies. BMC Med. 2016;14:210. 10.1186/s12916-016-0742-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126. Piuri G, Zocchi M, Della Porta M, et al. Magnesium in obesity, metabolic syndrome, and type 2 diabetes. Nutrients. 2021;13(2):320. 10.3390/nu13020320 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127. Gold MS, Sehayek D, Gabrielli S, Zhang X, McCusker C, Ben‐Shoshan M. COVID‐19 and comorbidities: a systematic review and meta‐analysis. PGM (Postgrad Med). 2020;132(8):749‐755. 10.1080/00325481.2020.1786964 [DOI] [PubMed] [Google Scholar]
- 128. Al Alawi AM, Majoni SW, Falhammar H. Magnesium and human health: perspectives and research directions. Int J Endocrinol. 2018;2018:9041694‐17. 10.1155/2018/9041694 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129. McCarty MF. Magnesium may mediate the favorable impact of whole grains on insulin sensitivity by acting as a mild calcium antagonist. Med Hypotheses. 2005;64(3):619‐627. 10.1016/j.mehy.2003.10.034 [DOI] [PubMed] [Google Scholar]
- 130. Fung TT, Hu FB, Pereira MA, et al. Whole‐grain intake and the risk of type 2 diabetes: a prospective study in men. Am J Clin Nutr. 2002;76(3):535‐540. 10.1093/ajcn/76.3.535 [DOI] [PubMed] [Google Scholar]
- 131. Glasdam SM, Glasdam S, Peters GH. The importance of magnesium in the human body: a systematic literature review. Adv Clin Chem. 2016;73:169‐193. 10.1016/bs.acc.2015.10.002 [DOI] [PubMed] [Google Scholar]
- 132. Arshad MS, Khan U, Sadiq A, et al. Coronavirus disease (COVID‐19) and immunity booster green foods: a mini review. Food Sci Nutr. 2020;8(8):3971‐3976. 10.1002/fsn3.1719 [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 133. FoodData Central. Accessed October 29, 2021. https://fdc.nal.usda.gov/ [Google Scholar]
- 134. Kopf JC, Suhr MJ, Clarke J, et al. Role of whole grains versus fruits and vegetables in reducing subclinical inflammation and promoting gastrointestinal health in individuals affected by overweight and obesity: a randomized controlled trial. Nutr J. 2018;17(1):72. 10.1186/s12937-018-0381-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135. Yoo EH, Chang SH, Song D‐Y, et al. Comprehensive laboratory data analysis to predict the clinical severity of coronavirus disease 2019 in 1,952 patients in Daegu, Korea. Annals Lab Med. 2022;42(1):24‐35. 10.3343/alm.2022.42.1.24 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136. Guimarães LMF, Rossini CVT, Lameu C. Implications of SARS‐Cov‐2 infection on eNOS and iNOS activity: consequences for the respiratory and vascular systems. Nitric Oxide. 2021;111‐112:64‐71. 10.1016/j.niox.2021.04.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137. Su B, Liu T, Fan H, et al. Inflammatory markers and the risk of chronic obstructive pulmonary disease: a systematic review and meta‐analysis. PLOS ONE. 2016;11(4):e0150586. 10.1371/journal.pone.0150586 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138. Yang Q, Shigemura N, Underwood MJ, et al. NO and EDHF pathways in pulmonary arteries and veins are impaired in COPD patients. Vasc Pharmacol. 2012;57(2):113‐118. 10.1016/j.vph.2012.05.004 [DOI] [PubMed] [Google Scholar]
- 139. Rahman I, van Schadewijk AAM, Crowther AJL, et al. 4‐Hydroxy‐2‐Nonenal, a specific lipid peroxidation product, is elevated in lungs of patients with chronic obstructive pulmonary disease. Am J Respir Crit Care Med. 2002;166(4):490‐495. 10.1164/rccm.2110101 [DOI] [PubMed] [Google Scholar]
- 140. Pasini AMF, Ferrari M, Stranieri C, et al. Nrf2 expression is increased in peripheral blood mononuclear cells derived from mild–moderate ex‐smoker COPD patients with persistent oxidative stress. COPD. 2016;11(1):1733‐1743. 10.2147/COPD.S102218 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141. Yamada K, Asai K, Nagayasu F, et al. Impaired nuclear factor erythroid 2‐related factor 2 expression increases apoptosis of airway epithelial cells in patients with chronic obstructive pulmonary disease due to cigarette smoking. BMC Pulm Med. 2016;16:27. 10.1186/s12890-016-0189-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142. Hashim Ali Hussein S, Nielsen LP, Konow Bøgebjerg Dolberg M, Dahl R. Serum magnesium and not vitamin D is associated with better QoL in COPD: a cross‐sectional study. Respir Med. 2015;109(6):727‐733. 10.1016/j.rmed.2015.03.005 [DOI] [PubMed] [Google Scholar]
- 143. Mukerji S, Shahpuri B, Clayton‐Smith B, et al. Intravenous magnesium sulphate as an adjuvant therapy in acute exacerbations of chronic obstructive pulmonary disease: a single centre, randomised, double‐blinded, parallel group, placebo‐ controlled trial: a pilot study. Randomized Controlled Trial. 2015;128(1425):9. [PubMed] [Google Scholar]
- 144. Irazuzta JE, Chiriboga N. Magnesium sulfate infusion for acute asthma in the emergency department. J Pediatr. 2017;93:19‐25. 10.1016/j.jped.2017.06.002 [DOI] [PubMed] [Google Scholar]
- 145. Cheuk D, Chau T, Lee S. A meta‐analysis on intravenous magnesium sulphate for treating acute asthma. Arch Dis Child. 2005;90(1):74‐77. 10.1136/adc.2004.050005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146. Shan Z, Rong Y, Yang W, et al. Intravenous and nebulized magnesium sulfate for treating acute asthma in adults and children: a systematic review and meta‐analysis. Respir Med. 2013;107(3):321‐330. 10.1016/j.rmed.2012.12.001 [DOI] [PubMed] [Google Scholar]
- 147. Dubé BP, Dres M. Diaphragm dysfunction: diagnostic approaches and management strategies. J Clin Med. 2016;5(12):113. 10.3390/jcm5120113 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148. Supinski GS, Morris PE, Dhar S, Callahan LA. Diaphragm dysfunction in critical illness. Chest. 2018;153(4):1040‐1051. 10.1016/j.chest.2017.08.1157 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149. McCool FD, Tzelepis GE. Dysfunction of the diaphragm. N. Engl J Med. 2012;366(10):932‐942. 10.1056/NEJMra1007236 [DOI] [PubMed] [Google Scholar]
- 150. Hermans G, Agten A, Testelmans D, Decramer M, Gayan‐Ramirez G. Increased duration of mechanical ventilation is associated with decreased diaphragmatic force: a prospective observational study. Crit Care. 2010;14(4):R127. 10.1186/cc9094 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.van; Steveninck AL, Imming LM. Diaphragm dysfunction prior to intubation in a patient with Covid‐19 pneumonia; assessment by point of care ultrasound and potential implications for patient monitoring. Respiratory Med Case Reports. 2020;31:101284. 10.1016/j.rmcr.2020.101284 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152. Galley HF, Lowes DA, Allen L, Cameron G, Aucott LS, Webster NR. Melatonin as a potential therapy for sepsis: a phase I dose escalation study and an ex vivo whole blood model under conditions of sepsis. J Pineal Res. 2014;56(4):427‐438. 10.1111/jpi.12134 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153. Arnalich F, Garcia‐Palomero E, López J, et al. Predictive value of nuclear factor κB activity and plasma cytokine levels in patients with sepsis. Infect Immun. 2000;68(4):1942‐1945. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154. Demoule A, Jung B, Prodanovic H, et al. Diaphragm dysfunction on admission to the intensive care unit. Prevalence, risk factors, and prognostic impact—a prospective study. Am J Respir Crit Care Med. 2013;188(2):213‐219. 10.1164/rccm.201209-1668OC [DOI] [PubMed] [Google Scholar]
- 155. Whidden MA, Smuder AJ, Wu M, Hudson MB, Nelson WB, Powers SK. Oxidative stress is required for mechanical ventilation‐induced protease activation in the diaphragm. J Appl Physiology. 2010;108(5):1376‐1382. 10.1152/japplphysiol.00098.2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156. Janssen SPM, Gayan‐Ramirez G, Van Den Bergh A, et al. Interleukin‐6 causes myocardial failure and skeletal muscle atrophy in rats. Circulation. 2005;111(8):996‐1005. 10.1161/01.CIR.0000156469.96135.0D [DOI] [PubMed] [Google Scholar]
- 157. Li X, Moody MR, Engel D, et al. Cardiac‐specific overexpression of tumor necrosis factor‐α causes oxidative stress and contractile dysfunction in mouse diaphragm. Circulation. 2000;102(14):1690‐1696. 10.1161/01.CIR.102.14.1690 [DOI] [PubMed] [Google Scholar]
- 158. Jiang J, Chen Q, Chen X, Li J, Li S, Yang B. Magnesium sulfate ameliorates sepsis‐induced diaphragm dysfunction in rats via inhibiting HMGB1/TLR4/NF‐κB pathway. Neuroreport. 2020;31(12):902‐908. 10.1097/WNR.0000000000001478 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159. Panigada M, Bottino N, Tagliabue P, et al. Hypercoagulability of COVID‐19 patients in intensive care unit: a report of thromboelastography findings and other parameters of hemostasis. J Thrombosis Haemostasis. 2020;18(7):1738‐1742. 10.1111/jth.14850 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160. Levi M, van der Poll T. Inflammation and coagulation. Crit Care Med. 2010;38:S26‐S34. 10.1097/CCM.0b013e3181c98d21 [DOI] [PubMed] [Google Scholar]
- 161. Asakura H, Ogawa H. COVID‐19‐associated coagulopathy and disseminated intravascular coagulation. Int J Hematol. 2021;113(1):45‐57. 10.1007/s12185-020-03029-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162. Iba T, Levy JH, Levi M, Thachil J. Coagulopathy in COVID‐19. J Thrombosis Haemostasis. 2020;18(9):2103‐2109. 10.1111/jth.14975 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163. Tang N, Li D, Wang X, Sun Z. Abnormal coagulation parameters are associated with poor prognosis in patients with novel coronavirus pneumonia. J Thromb Haemost. 2020;18(4):844‐847. 10.1111/jth.14768 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164. Lau FH, Majumder R, Torabi R, et al. Vitamin D Insufficiency Is Prevalent in Severe COVID‐19; 2020. Infectious Diseases (except HIV/AIDS). 10.1101/2020.04.24.20075838 [DOI] [Google Scholar]
- 165. Hariharan A, Hakeem AR, Radhakrishnan S, Reddy MS, Rela M. The role and therapeutic potential of NF‐kappa‐B pathway in severe COVID‐19 patients. Inflammopharmacol. 2021;29(1):91‐100. 10.1007/s10787-020-00773-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166. Hirano T, Murakami M. COVID‐19: a new virus, but a familiar receptor and cytokine release syndrome. Immunity. 2020;52(5):731‐733. 10.1016/j.immuni.2020.04.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167. Spronk HMH, Govers‐Riemslag JWP, ten Cate H. The blood coagulation system as a molecular machine. BioEssays. 2003;25(12):1220‐1228. 10.1002/bies.10360 [DOI] [PubMed] [Google Scholar]
- 168. Gromova OA, Torshin IY, Kobalava ZD, et al. Deficit of magnesium and states of hypercoagulation: intellectual analysis of data obtained from a sample of patients aged 18‐50 years from medical and preventive facilities in Russia. Kardiologiia. 2018;58(4):22‐35. 10.18087/cardio.2018.4.10106 [DOI] [PubMed] [Google Scholar]
- 169. Mannuß S, Schuff‐Werner P, Dreißiger K, Kohlschein P. Magnesium sulfate as an alternative in vitro anticoagulant for the measurement of platelet parameters? Am J Clin Pathology. 2016;145(6):806‐814. 10.1093/ajcp/aqw066 [DOI] [PubMed] [Google Scholar]
- 170. Sobczak AIS, Phoenix FA, Pitt SJ, Ajjan RA, Stewart AJ. Reduced plasma magnesium levels in type‐1 diabetes associate with prothrombotic changes in fibrin clotting and fibrinolysis. Thromb Haemost. 2020;120(02):243‐252. 10.1055/s-0039-3402808 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171. Vadivel K, Agah S, Messer AS, et al. Structural and functional studies of γ‐carboxyglutamic acid domains of factor VIIa and activated protein C: role of magnesium at physiological calcium. J Mol Biol. 2013;425(11):1961‐1981. 10.1016/j.jmb.2013.02.017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172. Jankun J, Skrzypczak‐Jankun E, Lipinski B. Experimental immunology Complex function of magnesium in blood clot formation and lysis. cejoi. 2013;2:149‐153. 10.5114/ceji.2013.35203 [DOI] [Google Scholar]
- 173. Demopoulos C, Antonopoulou S, Theoharides TC. COVID‐19, microthromboses, inflammation, and platelet activating factor. BioFactors. 2020;46(6):927‐933. 10.1002/biof.1696 [DOI] [PubMed] [Google Scholar]
- 174. Altura BM, Li W, Zhang A, et al. The expression of platelet‐activating factor is induced by low extracellular Mg2+ in aortic, cerebral and neonatal coronary vascular smooth muscle; cross talk with ceramide production, NF–kB and proto‐oncogenes: possible links to atherogenesis and sudden cardiac death in children and infants, and aging; hypothesis, review and viewpoint. IJCRR. Publ online March. 2016;31:47‐67. 10.19070/2470-4563-1600011 [DOI] [Google Scholar]
- 175. Altura B, Gebrewold A, Shah G, Shah N, Altura B. Potential roles of magnesium deficiency in inflammation and atherogenesis: importance and cross‐talk of platelet‐activating factor and ceramide. J Clin Exp Cardiolog. 2016;07(03). 10.4172/2155-9880.1000427 [DOI] [Google Scholar]
- 176. Kamal M, Abo Omirah M, Hussein A, Saeed H. Assessment and characterisation of post‐COVID‐19 manifestations. Int J Clin Pract. 2021;75(3):e13746. 10.1111/ijcp.13746 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177. Mandal S, Barnett J, Brill SE, et al. ‘Long‐COVID’: a cross‐sectional study of persisting symptoms, biomarker and imaging abnormalities following hospitalisation for COVID‐19. Thorax. 2021;76(4):396‐398. 10.1136/thoraxjnl-2020-215818 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178. Carfì A, Bernabei R, Landi F. For the gemelli against COVID‐19 post‐acute care study group. Persistent symptoms in patients after acute COVID‐19. JAMA. 2020;324(6):603‐605. 10.1001/jama.2020.12603 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179. Cares‐Marambio K, Montenegro‐Jiménez Y, Torres‐Castro R, et al. Prevalence of potential respiratory symptoms in survivors of hospital admission after coronavirus disease 2019 (COVID‐19): a systematic review and meta‐analysis. Chron Respir Dis. 2021;18:14799731211002240. 10.1177/14799731211002240 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180. Iqbal FM, Lam K, Sounderajah V, Clarke JM, Ashrafian H, Darzi A. Characteristics and predictors of acute and chronic post‐COVID syndrome: a systematic review and meta‐analysis. EClinicalMedicine. 2021;36:100899. 10.1016/j.eclinm.2021.100899 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181. Halpin SJ, McIvor C, Whyatt G, et al. Postdischarge symptoms and rehabilitation needs in survivors of COVID‐19 infection: a cross‐sectional evaluation. J Med Virology. 2021;93(2):1013‐1022. 10.1002/jmv.26368 [DOI] [PubMed] [Google Scholar]
- 182. Castro‐Marrero J, Cordero MD, Sáez‐Francas N, et al. Could mitochondrial dysfunction Be a differentiating marker between chronic fatigue syndrome and fibromyalgia? Antioxidants Redox Signal. 2013;19(15):1855‐1860. 10.1089/ars.2013.5346 [DOI] [PubMed] [Google Scholar]
- 183. Myhill S, Booth NE, McLaren‐Howard J. Chronic fatigue syndrome and mitochondrial dysfunction. Int J Clin Exp Med. 2009;2(1):1‐16. [PMC free article] [PubMed] [Google Scholar]
- 184. Farr E, Wolfe AR, Deshmukh S, et al. Diaphragm dysfunction in severe COVID‐19 as determined by neuromuscular ultrasound. Annals Clin & Trans Neurol. 2021;8(8):1745‐1749. 10.1002/acn3.51416 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 185. Satturwar S, Fowkes M, Farver C, et al. Postmortem findings associated with SARS‐CoV‐2: systematic review and meta‐analysis. Am J Surg Pathology. 2021;45(5):587‐603. 10.1097/PAS.0000000000001650 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Data is openly available in a public repository that issues datasets with https://pubmed.ncbi.nlm.nih.gov/.