

Abstract
Dengue is one of the most prevalent mosquito-borne diseases in the world, affecting an estimated 390 million people each year, according to models. For the last two decades, efforts to develop safe and effective vaccines to prevent dengue virus (DENV) infections have faced several challenges, mostly related to the complexity of conducting long-term studies to evaluate vaccine efficacy and safety to rule out the risk of vaccine-induced DHS/DSS, particularly in children. At least seven DENV vaccines have undergone different phases of clinical trials; however, only three of them (Dengvaxia®, TV003, and TAK-003) have showed promising results, and are addressed in detail in this review in terms of their molecular design, efficacy, and immunogenicity. Safety-related challenges during DENV vaccine development are also discussed.
Key Points
| Dengue vaccine development has been challenging because of the need to provide protection against all four dengue serotypes to avoid potentially causing antibody-dependent enhancement in further infections. |
| Denvaxia® is currently the only licenced vaccine, but phase III clinical trials with two other vaccines, TV-003/TV-005 and TAK-003, are currently ongoing, with promising results. |
Introduction
Dengue fever represents a great burden for the public health systems worldwide and is considered the most prevalent mosquito-borne disease in tropical and subtropical regions of the world [1, 2], rapidly expanding every year due to several factors such as climate change, deforestation associated with uncontrolled urbanization, overpopulation, and the emergence of mosquitoes resistant to common insecticides, amongst others [3–5]. Dengue fever is caused by dengue virus (DENV), which is classified under the genus Flavivirus of the family Flaviviridae and is transmitted by mosquitoes from the Aedes genus, mostly Aedes aegypti and Aedes albopictus. Four genetically distinct DENV serotypes (DENV-1, DENV-2, DENV-3, and DENV-4) have been reported to co-circulate amongst humans worldwide [6, 7]. Dengue is characterized by a wide spectrum of clinical manifestations ranging from a mild febrile illness to severe dengue, increasing the risk of developing dengue hemorrhagic fever (DHF) and dengue shock syndrome (DSS). A secondary DENV infection (e.g., exposure to a heterotypic serotype) is the greatest risk factor for serious diseases due to the phenomenon of antibody-dependent enhancement (ADE) [8]. Briefly, cross-reactive antibodies generated after exposure to the first DENV serotype combine with the second DENV serotype to create infectious immune complexes that enter Fc-receptor-bearing cells. As a result, the number of infected cells and the amount of virus produced per cell increases. This is therefore imperative that DENV vaccines protect against infection from all four serotypes to avoid ADE.
At least seven DENV vaccines based on different platforms including live attenuated viruses, inactivated viruses, chimeric live attenuated viruses, DNA, and recombinant proteins have been developed and are currently undergoing different phases of clinical trials (Table 1) or are under preclinical investigation [6]. In this review we discuss recent progress of the three most advanced DENV vaccines (Dengvaxia®, TV003/TV005, and TAK-003) (Table 2), focusing on the molecular characteristics of each vaccine (Fig. 1) and the available clinical data on their efficacy and immunogenicity. Finally, we emphasize the safety-related challenges associated with the risk of vaccine-induced DHS/DSS.
Table 1.
Candidate dengue vaccines in phase I or phase II clinical trials
| Candidate | Platform | Phase/stage | References |
|---|---|---|---|
| TDEN-LAV (WRAIR/GSK) | Live-attenuated | Phase II (Discontinued) | [9] |
| TDENV-PIV (WRAIR/FioCruz/GSK) | Inactivated adjuvanted | Phase I (No recent reports) | [10] |
| D1ME100/TVDV (NMRC) | DNA vaccine | Phase I (No recent updates) | [11] |
| V180 (DEN-80E) (Merck/NIAD) | Recombinant (subunit) | Phase I (Published 2019) | [12] |
| DENV-1-LVHC | Live-attenuated | Phase I (Published 2021) | Clinicaltrials.gov [13] |
Table 2.
Dengue vaccines that have reach phase III or have been licensed
| Vaccine | Manufacturer | Platform | Efficacy | Comments | References |
|---|---|---|---|---|---|
| Licensed | |||||
|
CYT-TDV Dengvaxia® |
Sanofi Pasteur | YFV ∆30 backbone | 25–59% | Increases hospitalizations in seronegative vaccinees | [7] |
| Phase III | |||||
|
TAK-003 (DENVax) |
Takeda/Inviragen | Attenuated DENV-2 backbone for the four serotypes | 73.3–85.3% | Well tolerated in adolescents and children | [14] |
|
LATV TV003/TV005 |
NIAD/Butantan/Merck | DENV-1,3,4 ∆30 and rDENV2/4 ∆30 | Not yet released | Single dose | [15] |
Fig. 1.
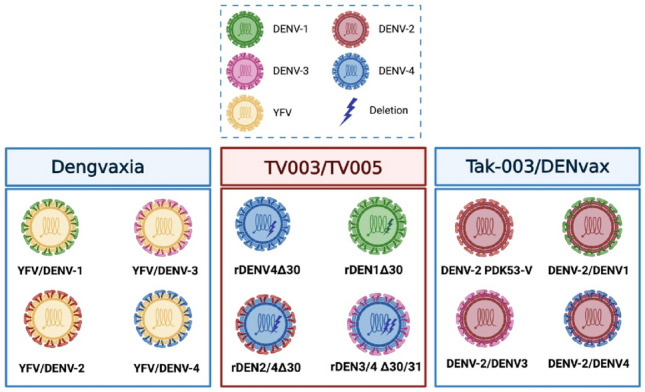
Molecular design of the anti-dengue virus (DENV) vaccines in advanced stages of clinical development. A Dengvaxia® is based on a yellow fever backbone in which the pre-membrane (prM) and envelope (E) genes of YFV have been replaced by the homologous genes from each one of the four DENV serotypes [16, 17]. B TV003/TV005 was constructed by a deletion of 30 nucleotides (172–143) in the TL2 stem-loop of the 3′-UTR of DENV-4 and DENV-1 (rDEN4∆30 and rDEN1∆30), DENV-2 and DENV-3 components were constructed from the rDEN4∆30 backbone [21, 22]. C Tak-003/DENVax is based on a live-attenuated DENV-2 strain (PDK-53-V) in which the pre-membrane (prM) and envelope (E) genes of YFV have been replaced by the homologous genes from each one of the four DENV serotypes [27]
For this review we systematically searched PubMed, Web of Science, and clinicaltrial.gov to obtain results on specific topics such as vaccines, trials as well as safety on dengue vaccines, and immunopathogenesis. We selected the most appropriate bibliography for this document.
Development and Preclinical Evaluation of Dengue Virus (DENV) Vaccines
Dengvaxia®
Initially developed in the early 2000s by the National Institutes of Health (NIH), the University of St. Louis, and Acambis Inc, and subsequently licensed by Sanofi Pasteur, this vaccine takes advantage of the ChimeriVax™ technology. Based on a vaccine strain (17D) of yellow fever virus (YFV) in which the pre-membrane (prM) and envelope (E) genes of YFV have been replaced by the homologous genes from each one of the four DENV serotypes derived from DENV isolates obtained in Thailand and Indonesia between 1978 and 1988, this technology enabled the generation of four chimeric YF-DEN viruses that were used in the formulation of a tetravalent DENV vaccine (ChimeriVax™ DENV 1-4) [16, 17].
Preclinical evaluation of the safety and immunogenicity of the tetravalent ChimeriVax™ DENV 1-4 vaccine showed a reduced neurovirulence profile in mice compared to the parental YFV vaccine strain (YF-VAX) [18]. Also, neurovirulence tests performed in Macaca fascicularis confirmed that the tetravalent ChimeriVax™ DENV 1-4 vaccine was significantly less neurovirulent than the parental YF-VAX strain [19]. In addition, the vaccine generated seroconversion and strong neutralizing antibody responses against all four DENV serotypes following one administration of either a high or a low dose of the vaccine in cynomolgus macaques and limited viremias compared to the parental DENV strains. Interestingly, challenge studies revealed that 92% of the vaccinated monkeys were protected against a challenge with wild-type DENV 1-4 [20].
TV003/TV005
The development of the live attenuated DENV vaccines called TV003/TV005 began in 1996 at the Laboratory of Infectious Diseases (LID) of the National Institute of Allergy and Infectious Diseases (NIAID). Considering the importance of the untranslated regions (UTRs) for the replication of DENV genome, the initial attenuation strategy focused on deleting 30 contiguous nucleotides (172-143) in the TL2 stem-loop from the 3´-UTR of DENV-4 (rDEN4∆30) [20]. A mutant lacking the same homologous genomic region was also constructed for DENV-1 (rDEN1∆30). Both mutants displayed an attenuated phenotype as demonstrated by their reduced infectivity, and exhibited their capacity to induce strong neutralizing antibody responses in rhesus macaques that correlated with the protection when challenged with wild types of DENV-1 and DENV-4 [21].
Further efforts to achieve a tetravalent DENV vaccine led to the generation of the attenuated DENV-2 component by using the backbone of rDEN4∆30 to generate two attenuated DENV4-DENV2 chimeric viruses in which the membrane and envelope genes (rDEN2/4 ∆30 (ME)), or the capsid, membrane, and envelope genes (rDENV2/4 ∆30 (CME)) of DENV-4 were replaced with the homologous genes of DENV-2 [21, 22]. Preclinical evaluation of the two chimeras showed that both display a highly attenuated phenotype in SCID-HuH-7 mice, and rhesus macaques in which the chimerization and the ∆ 30 deletion was additive rendering the virus not infectious for monkeys [21]. A chimeric DENV3-DENV4 virus containing the original 30 nt deletion at the 3′-UTR was generated (rDEN3 ∆30 (ME)) and further modified by introducing a non-continuous deletion of 31 nt (258-228) (rDEN3 ∆30/31). Preclinical evaluation of the mutant virus rDENV3 ∆30/31 in non-human primates revealed the desirable safety, undetectable viremia, and strong neutralizing antibody responses, which were sufficient to protect the vaccinated monkeys when challenged with wild-type DENV-3 [23].
TAK-003 (DENVax)
The development of the DENVax vaccine started in the late 1980s, when researchers from the Mahidol University in Bangkok, Thailand, isolated a DENV-2 strain (DENV-2 16681) from the serum of a patient with dengue hemorrhagic fever. The attenuation of the DENV-2 16681 strain by 53 serial passages in primary dog kidney cells (PDK cells) lead to the obtention of the DENV-2 PDK-53-V strain, which in contrast to the parental DENV-2 PDK-53 strain, which has attenuation-related mutations in the 5´UTR and NS1 gene, possesses an additional non-synonymous mutation in the NS3 gene. The DENV-2 PDK53-V strain displays reduced neurovirulence in suckling mice and lower replication rates in C6/36 cells [24–26], and was further used as the backbone to generate the DENVax vaccines.
The vaccine strains for DENV-1, DENV-3, and DENV-4 used to formulate tetravalent DENVax vaccine were generated by replacing the pre-membrane (prM) and envelope (E) genes from the DENV-2 PDK53-V strain, with the prM and E genes from wild-type DENV strains [27]. The chimeric DENVax viruses displayed “small plaque” and temperature-sensitive phenotypes when replicated in LLC-MK2 cells, in contrast to the parental wild-type strains. Tetravalent formulations containing the four DENVax vaccine strains (DENV 1-4) demonstrated reduced neurovirulence profiles in newborn ICR mice and were shown to be immunogenic in AG129 knockout mice, inducing high neutralizing antibody titers (1:320–1:2560) against the four DENV serotypes [27]. Preclinical evaluation of the tetravalent DENVax vaccine in cynomolgus macaques (Macaca fascicularis) showed a good vaccine safety profile and was well tolerated when administered by the subcutaneous route, while inducing protection against the four DENV serotypes (measured by the protection against viremia) after two immunizations with a high dose scheme of a 5:5:5:5 formulation of each DENVax strain (DENVax 1-4) [28].
Clinical Evaluation of DENV Vaccines
To date, Dengvaxia® is the only licensed DENV vaccine, yet phase III clinical trials with the TV-003/TV-005 and TAK-003 are currently ongoing with promising results. Nevertheless, as we discussed, differences between the age and serostatus of the vaccinees have been shown to have a direct impact on vaccine efficacy and safety. Most of the safety concerns regarding the DENV vaccines derive from phase III pediatric clinical trials to date, only Dengvaxia® and TAK003 (DENVax) have been tested in children, with mixed results (Fig. 2).
Fig. 2.
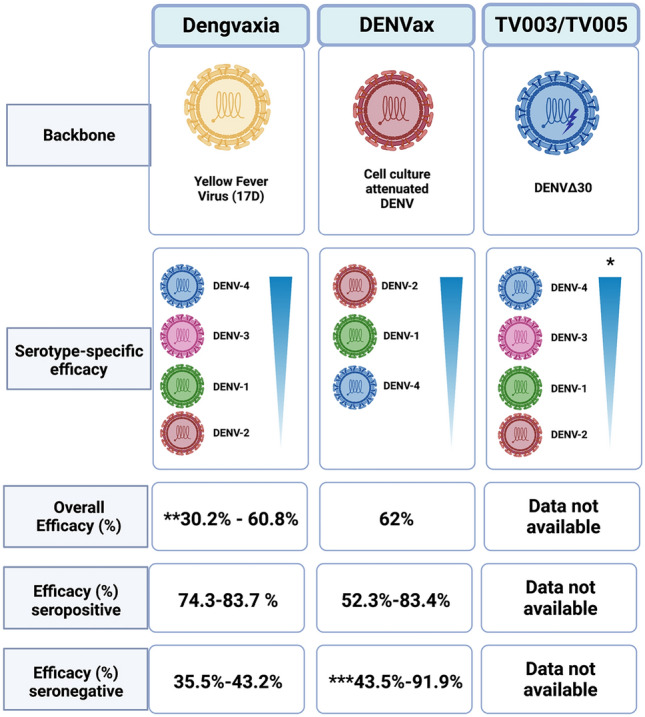
Overview of the efficacy trials of anti-dengue virus (DENV) vaccines in children in Latin America and Asia. Phase III clinical trials have been conducted for Dengvaxia and DENVax with mixed results. TV003/TV005 is currently undergoing phase III clinical trials. *The seroconversion rates for TV003/TV005 observed in phase II clinical trials are illustrated. **The lower value of the efficacy range depicted corresponds to the efficacy observed during the phase IIb trial conducted in Thailand. ***DENVax was only efficacious against DENV-1 and DENV-2 in seronegative individuals [38, 86]
Dengvaxia®
Early phase I and phase II clinical trials to evaluate the safety, immunogenicity, and reactogenicity of the tetravalent presentation of Dengvaxia® (TDV) in healthy adults, between 18 and 45 years, were carried out in USA, the Philippines, Australia, Mexico, Vietnam, Singapore, and India; some of these studies also included pediatric populations aged between 2 and 18 years [29].
The primary safety evaluation of the Dengvaxia® vaccine in adults showed that it was safe and well tolerated in a three-dose regimen at 0, 3–4, and 12 months, inducing mild to moderate transient local and systemic adverse events such as injection site pain, headache, malaise, and low-grade fever amongst others, with no vaccine-related severe adverse events (SAEs) reported. Assessment of the vaccine-induced cellular responses revealed that the vaccine did not induce the release of proinflammatory cytokines (IFN-γ, IL-1β, IL-6, IL-8, IL-10, IL-12p70), while inducing DENV serotype specific T-helper responses [30, 31].
The CD8 responses against the NS3 protein of the YFV-17D strain used as the backbone for the ChimeriVax™ viruses were observed [32]. In individuals with pre-existing DENV immunity, the vaccine induced broader neutralizing antibody responses and boosted specific CD8+ responses against DENV non-structural proteins, particularly NS3, which are not elicited in naïve individuals. Even though most of the phase III clinical trials to evaluate the efficacy of Dengvaxia® have been conducted in the pediatric population (< 18 years), a randomized phase III clinical trial to evaluate the lot-to-lot consistency of the vaccine in healthy adults in Australia [33] revealed that even though naïve individuals developed neutralizing antibodies against all four DENV serotypes after receiving one dose of the vaccine, the only serotype-specific neutralizing antibodies produced were against DENV-4 [34]. Instead, DENV 1-3 were neutralized by cross-reactive antibodies, revealing the immunodominance of the DENV-4 component of the vaccine [34]. Moreover, differential rates of viral replication of each vaccine component were observed by RT-PCR in dengue-naïve individuals after receiving one vaccine dose: DENV-4 (44%), DENV-3 (12%), DENV-1 (7%), and DENV-2 (0%), which may pose a risk for seronegative individuals to develop vaccine-enhanced disease when infected with DENV-2.
The most informative study regarding the efficacy and safety of Dengvaxia® in the pediatric population was conducted in 35,000 children aged between 2 and 16 years. The study revealed that the efficacy was age dependent, ranging from 65% in children > 9 years to 45% in children 9 years or younger [35]. Worryingly, children under 9 years showed a tendency to develop severe dengue after immunization following natural exposure to the virus, an effect that was particularly seen in dengue-naïve children [36]. Pediatric studies to evaluate the efficacy of Dengvaxia® have shown that efficacy varies between DENV serotypes and age groups. Results from a phase IIb trial conducted in Thailand to evaluate the efficacy of Dengvaxia® in a cohort of 4002 children between 4 and 11 years revealed that vaccine efficacy was higher against DENV-4 and DENV-3 (100% and 75.3%, respectively) than for DENV-1 and DENV-2 (55.6% and 9.2%, respectively) [37].
In larger pediatric phase III trials conducted in five dengue endemic Latin America countries in a cohort of 20,869 healthy children between 9 and 16 years of age, efficacy against DENV-3 and DENV-4 was higher (74.0% and 77.0%, respectively) than for DENV-1 and DENV-2 (50.3% and 42.3%, respectively), confirming that the vaccine efficacy varies between serotypes and that it is age dependent [38]. The latter was later confirmed by a larger phase III clinical trial to evaluate the efficacy of Dengvaxia® in a cohort of healthy children from Latin America and Asia aged between 2 to 16 years. Results from this study revealed that the pooled rates of efficacy for symptomatic dengue in a follow-up period of 25 months were higher for children older than 9 years (65.6%) than for children under 9 years (44.6%). Moreover, the pooled relative risk of dengue requiring hospitalization was higher for children under 9 years of age (1.58) than for children aged over 9 years (0.5) [39].
Further trials have suggested that the observer age-dependent risk of dengue requiring hospitalization in children under 9 years of age might be related to pre-existing anti-DENV immunity. A cohort study that analyzed data from three different efficacy trials revealed a higher incidence of hospitalization due to virologically confirmed dengue (VCD) for seronegative vaccinees aged between 2 and 16 years (3.06%) than for seronegative controls (1.87%), a trend that was also observed for seronegative vaccinees aged between 9 and 16 years (1.57% in seronegative individuals vs. 1.09% in seropositive individuals) [36].
Despite the high vaccine efficacy observed with Dengvaxia® in different clinical trials, recent mathematical models using data of more than 800,000 children vaccinated in the Philippines estimate there will be more than 1000 hospitalizations due to severe dengue in a period of 4 years post-vaccination, in both seronegative and seropositive individuals [40]. These models along with the high rates of hospitalization observed in children under 9 years of age, that were wrongly considered as vaccine failure cases, highlight the importance of conducting enhanced phase IV surveillance studies in children vaccinated with Dengvaxia® for a better assessment of the effectiveness of the vaccine in dengue endemic countries, even before the approval and deployment of the vaccine.
TV003/TV005
During the early phases of its clinical development, several phase I clinical trials were conducted using monovalent formulations of the DENVax vaccine candidates to evaluate the safety profile, replication capacity of the individual DENVax viruses, and transmissibility from vaccinated individuals to Toxorhynchites splendens mosquitoes [41–45].
After these initial studies, six monovalent DENVax vaccine candidates were selected for further clinical evaluation formulated as five tetravalent mixtures (TV-001 to TV-005). Two vaccine candidates (TV003 and TV005) formulated with rDEN1D30 rDEN2/4D30, rDEN3D30/31, and rDEN4D30, but with different amounts of the rDEN2/4D30 component (103 PFUs/mL in TV003 and 104 PFUs/mL in TV005) were selected for further clinical evaluation after inducing the most balanced neutralizing antibody responses against the four DENV serotypes [46].
The safety and immunogenicity of TV003 and TV005, evaluated in two randomized placebo-controlled trials [47, 48] in flavivirus-naïve subjects, revealed that both TV003 and TV005 were well tolerated and showed a good safety profile, with low-grade rash reported as the most frequent adverse event in 76% of the participants in both groups. A single dose of TV003 induced balanced neutralizing antibody responses against the four DENV serotypes with seroconversion rates between 64% (DENV-2) and 100% (DENV-4), while the specific response against DENV-2 improved with the TV005 vaccine (84%) after a single dose.
The lower proportion of participants that seroconverted to DENV-2 after receiving one dose of TV003 led to the development of a DENV-2 challenge model in which the original rDEN2D30 was used as a challenge virus 6 months post-vaccination, since it induced viremias 100-fold higher than the vaccine strain (rDEN2/4D30). The results showed that all the participants developed protective DENV-2 responses and induced specific neutralizing antibodies against DENV-2 [21].
These results led to the licensing of the TV003 vaccine by the Butantan Institute in Brazil under the name Butantan-DV, which was manufactured as a lyophilized tetravalent DENV vaccine and subjected to a two-step, double-blind, randomized, placebo-controlled, phase II clinical trial in 155 DENV-naïve and 145 DENV-exposed healthy individuals, aged between 18 and 59 years [48]. The vaccine was safe and well tolerated and induced robust balanced neutralizing antibody responses with seroconversion frequencies above 78% for the four DENV serotypes, in both DENV-naïve and DENV-exposed participants. Significant T-CD8 responses were observed in DENV-naïve and DENV-exposed participants 91 days after receiving one dose of the Butantan-DV vaccine, suggesting that this vaccine elicits broader protective immune responses in adults, in comparison to tetravalent vaccines based on the expression of structural DENV proteins. A randomized, multicenter, double-blind, placebo-controlled phase III clinical trial for the Butantan-DV vaccine is currently being conducted in Brazil with 16,944 participants divided into three age groups (18–59 years, 7–17 years, and 2–6 years); results for this trial are still to be published.
TAK-003 (DENVax)
The clinical evaluation of DENVax started with a randomized, double-blind, dose-escalation phase I clinical trial conducted by Takeda in Rionegro, Antioquia, Colombia, aimed to evaluate the safety and the immunogenicity against the four DENV serotypes of a two-dose scheme of DENVax administered intradermally (ID) or intramuscularly (IM) to DENV-naïve adults between 18 and 45 years of age [49]. The vaccine was safe and well tolerated among the participants in the study, inducing transient local reactogenicity and mild systemic adverse events. Vaccination induced the production of neutralizing antibodies against the four DENV serotypes, yet antibody titers against DENV-3 and DENV-4 were lower among the participants [50].
Due to the lower antibody responses against DENV-4, DENVax (now named TDV) was reformulated to increase the amount of the DENVax4 component, and different formulations and dosing schedules were tested in a randomized, multicenter, phase 1b study carried out in the USA with 140 DENV-naïve individuals between 18 and 45 years of age [51]. The seroconversion rates observed among the participants in the study were 84–100% (DENV-1), 96–100% (DENV-2), DENV-3: 83–100% (DENV-3), and 33–77% (DENV-4).
The evaluation of the safety and immunogenicity of a two-dose scheme (0, 90 days) of the TDV vaccine in DENV-exposed individuals during a randomized, double-blind, placebo-controlled phase II clinical trial carried out in Puerto Rico, Colombia, Singapore, and Thailand using a “high-dose” vaccine with higher titers of the DENVax3 and DENVax4 components, revealed that the vaccine induced lower neutralizing antibody responses against the DENVax4 component regardless of prior DENV exposure [52].
A large-scale phase III clinical trial to evaluate the efficacy of DENVax in a cohort of 20,071 healthy children, between 4 ando 16 years in dengue-endemic countries from Latin America and Asia, is currently being conducted. Primary results at 12 months post vaccination showed efficacy variations according to DENV serotype: 97.7% for DENV-2, 73.7% for DENV-1, and 62.6% against DENV-3, while the efficacy results for DENV-4 were inconclusive. Interestingly, the overall vaccine efficacy was similar between participants who were seronegative at baseline (74.9%) and those who were seropositive at baseline (82.2%), and were independent of age range. Overall vaccine efficacy against dengue leading to hospitalization was shown to be 95.4% amongst seronegative individuals and 94.4% for seropositive individuals [14].
Data from the same trial at 18 months post-vaccination revealed an overall vaccine efficacy of 76.1% in seropositive individuals and 66.2% in seronegative individuals, with an overall efficacy against different DENV serotypes ranging from 95.1% against DENV-2 to 48.9% against DENV-3. The overall efficacy against dengue requiring hospitalization was 90.4%, and 85.9% against DHF. Yet, when stratified by age group, the vaccine efficacy against requiring hospitalization was significantly lower in those previously seronegative children aged between 4 and 5 years (59.1%), as the efficacy to prevent hospitalization in seropositive children the same age was 51.6% [53].
The cumulative efficacy data of the DENVax vaccine 3 years post-vaccination were recently published, showing an overall vaccine efficacy of 62% against VCD, significantly lower than the one observed at 18 months post-vaccination. A similar effect was observed with the overall efficacy against dengue requiring hospitalization, which reduced from 90.4 to 83.6%. In baseline seropositives, vaccine efficacy against VCD ranged between 52.3% for DENV-3 and 83.4 % for DENV-2, yet, in baseline seronegatives the vaccine was only efficacious against DENV-1 and DENV-2 (43.5% and 91.9%, respectively), while no efficacy was observed for DENV-3. The efficacy against dengue requiring hospitalization remained high amongst seropositive individuals but not in seronegative individuals, when the vaccine was only efficacious against DENV-1 and DENV-2. Interestingly, the vaccine efficacy did not show any clear patterns that associated the efficacy observed with the age of the participants [54].
These significant variations in vaccine efficacy, assessed 18 months apart, should be further analyzed during long-term phase IV surveillance trials to rule out that the protective efficacy observed is due to cross-protection and might decline over time, as has been observed in pediatric populations during natural infections [55].
Challenges in DENV Vaccine Development
Immunopathological events are a common feature of DENV infections, with several underlying mechanisms such as an overreactive proinflammatory immune response (cytokine storm) characterized by an elevation of the plasmatic concentrations of several proinflammatory cytokines such as granulocyte-macrophage colony-stimulating growth factor (GM-CSF), macrophage inflammatory protein 1 beta (MIP-1β), interferon gamma (IFN-γ), and Intereukin 10 IL-10 [56], which eventually leads to vascular leak, hemorrhages [56, 57], and some other thrombotic events [58, 59]. Molecular mimicry involving E and NS1 viral proteins has also been documented to activate cross-reactive antibodies against platelets and endothelium leading to severe dengue [60–62]. Moreover, naturally infected individuals with one DENV serotype, showing low levels of anti-DENV antibodies (<1:80) suffer complications during secondary infections with a heterologous DENV serotype [63]. Specific antibodies produced during infections constitute a significant part of immune response to neutralize invaders; however, for pathogens such as DENV, under certain conditions antibodies generated in an initial encounter may enhance further infections [64, 65]. Interestingly, patients who develop high titers of anti-DENV antibodies (>1:320) exhibit protection against further symptomatic DENV infections.
The infection enhancement process known as antibody-dependent enhancement (ADE) in dengue is well documented [66], but the details of this event in dengue remain elusive. Virus entry is known to be facilitated by Fcγ (Fig. 3) and mediates T-cell activation and release of TNF-α and other cytokines that cause endothelial dysfunction. Hence, effective DENV vaccines must induce a strong and highly neutralizing response against all four DENV serotypes to avoid vaccine-induced immunopathologic events.
Fig. 3.
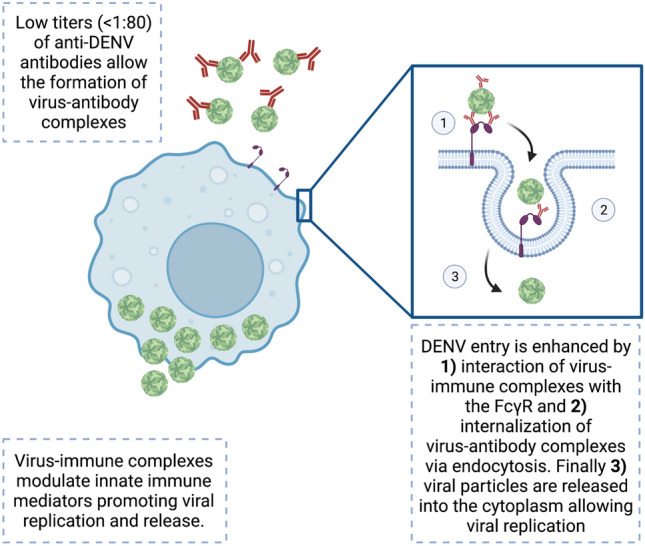
Schematic representation of antibody-dependent enhancement (ADE) in dengue virus (DENV) infection. Low levels of anti-DENV antibodies (< 1:80) against one DENV serotype promote the formation of virus-immune complexes during secondary infections with a heterologous DENV serotype. These virus-immune complexes are internalized into monocytes, macrophages and dendritic cells via the Fcγ receptor, promoting viral release into the cell cytoplasm. Virus-immune complexes modulate innate immune pathways promoting viral replication and release
Evidence shows that this process might be potentiated mostly by anti-prM antibodies. prM is present in immature virus particles. Antibodies like anti-prM not only facilitate Fc uptake, but these antibodies in high concentrations also poorly neutralize virus [67]. Different IgG subclasses (IgG1–4) exist and exhibit dissimilar properties regarding half-lives, levels in serum, complement activation, and binding to Fc receptors. Therefore, not only the targeted viral protein, but also the IgG subclass could help to better understand how neutralization/protection can be favored rather than ADE/immunopathogenesis. ADE occurrence in vaccinees must be examined regarding not only the IgG subclasses, but also the glycosylation stage [68, 69] to separate protective from immunopathogenic events. This is because glycosylation might alter affinity to Fc receptors as has been documented [70].
It seems all the live attenuated vaccines against dengue show capacity to induce antibodies against prM and fusion loop epitope resulting in serotype cross-reactivity and increasing the risk of ADE [71]. A new and important concern is how individuals will react to dengue vaccines, which arises in regions affected by the Zika virus (ZIKV). ADE has also been reported in individuals infected by DENV-2 who were previously infected by ZIKV [72]. Evidence in animal models show this event for both DENV and ZIKV when using live attenuated vaccines [73].
An inefficient innate immune response followed by production of subneutralizing antibodies and hyper-reactive T-cell response might contribute to immunopathologic events [67] in both dengue-infected individuals and vaccinees, but these events must be carefully analyzed in both vaccinees and naturally infected individuals to assess biological significance. Maps of epitopes targeted by highly neutralizing antibodies have been developed in macaques [74]; however, similar studies are needed in humans and these data must be considered for future vaccine development.
Safety Issues with Dengue Vaccines
To develop vaccines against dengue it is important for these to recognize the four existing serotypes; the three more advanced vaccines have considered this for their design (Table 3). An important consideration in natural infection is that a primary infection with one DENV serotype would establish a long-term memory against that specific serotype, but might result in a short-term, subneutralizing, and enhancing response for the other serotypes [75–77]. The enhancing phenomenon in subsequent dengue infections with a different DENV serotype could result due to previous heterotypic exposure, and avoiding these events is a constant concern in the field of dengue vaccines [78, 79].
Table 3.
Dengue vaccine characteristics
| Molecular design | Efficacy | Main side effects | Immunogenicity | |
|---|---|---|---|---|
| Dengvaxia® |
DENV-1 (prM-E) YFV-17D genome DENV-2 (prM-E) YFV-17D genome DENV-3 (prM-E) YFV-17D genome DENV-4 (prM-E) YFV-17D genome |
45–65% 44.6% under 9 yo 65.6% over 9 yo |
Injection site pain, headache, malaise, low-grade fever, among others | CD8+ reponse mostly to NS3 and neutralizing antibodies mostly against DENV-4 |
| [16, 17, 85] | [35, 38, 39] | [30, 31] | [34, 81] | |
| TV003/TV005 |
Complete DENV-1 Δ30 (3´UTR) DENV-2 (prM-E) in DENV-4 Δ30 (3´UTR) Complete DENV-3 Δ30 (3´UTR) Complete DENV-4 Δ30 (3´UTR) |
Low grade rash |
Reported as close to sterilizing Strong neutralizing antibodies in rhesus macaques |
|
| [21, 22, 85] | Data not available | [47, 48] | [21, 46, 81] | |
| TAK003 (DENVax) |
DENV-1 (prM-E) in DENV-2 genome* DENV-2 complete genome* DENV-3 (prM-E) in DENV-2 genome* DENV-4 (prM-E) in DENV-2 genome* *Referring to genetic construction |
74.9–76.1% in seropositive (previously exposed) 66.2–82.2% in seronegative (naïve) |
Transient local reactogenicity and mild systemic adverse events | Neutralizing antibodies against all four serotypes |
| [27, 85] | [14, 53] | [49, 50] | [50, 52] |
Hence, from the beginning, the need for tetravalent, equally effective immunization for all the four DENV serotypes was recognized as the most important challenge for design and development. Other concerns emerged later, such as the stimulation of high titers of neutralizing antibodies over those that sensitized individuals and enhance infection by DENV [71].
Dengvaxia®, one of the most advanced developments in vaccines against dengue, raised serious safety concerns when data showed an increased risk of hospitalization in naïve vaccinees when they were exposed to natural infection [80]. Takeda vaccine TAK-003 is still in phase III trials and has been shown to be safe and to induce protection against dengue-related hospitalization with efficacies around 75–80% independent of serostatus of vaccinated individuals [9]. Numerous phase II studies are still being carried out for this live-attenuated vaccine over the globe and in different age-group populations [31, 43, 81]. However, a phase III trial is still ongoing and the final conclusions on efficacy, side or rare adverse effects for this vaccine have not been drawn.
Regarding TV003/TV005, another live-attenuated vaccine developed in the National Institute of Allergy and Infectious Disease, data show these do not allow immunodominant expression of any of the four serotypes, resulting in a desirable immune response. Moreover, initial reports showed that a single dose is protective and nearly sterilizing immunity is achieved in a second dose 6–12 months after the first immunization [10, 82, 83]. This vaccine has been licensed by different manufacturers and the Instituto Butantan started a phase III trial to evaluate its efficacy [47].
Data of clinical trials for other promising developments using different vaccine platforms are still necessary, but much work is directed towards improving the existing vaccines. Developers aim to assure vaccines raise a broad and long-lasting highly neutralizing antibody response against all four serotypes [84]. Nevertheless, cellular response must also be evaluated and be efficient in vaccinees along with the antibody responses to assure the desirable protection. Most of all, vaccines should not pose any risk for developing severe disease in either naïve or previously exposed to DENV individuals. Thus, the scientific community is still working towards the safe and universal dengue vaccine the world needs.
Challenges and Opportunities
Reviewing clinical data on current dengue vaccines, clearly these pose a singular challenge for developers since protection must be achieved equally for all four serotypes without causing any potentially immunopathogenic event in further DENV encounters. Despite all efforts, dengue epidemics have intensified in number and frequency, and are affecting new geographical areas, thus it is critical to greatly improve control measures in dengue areas, and to develop a universal and highly effective vaccine to counteract dengue as one of those important measures. As safety remains a concern, the race to get an effective and safe dengue vaccine continues to be an imperative necessity.
Acknowledgements
The authors thank Dr. Brad Blitvich for suggestions in the manuscript, as well as reviewers and the editor for their revisions and comments, and also Instituto Politécnico Nacional for all its support.
Declarations
Funding
No funding was received for the preparation of this review.
Conflict of interest
Jesús M. Torres-Flores, Arturo Reyes-Sandoval, and Ma. Isabel Salazar have no conflicts of interest to declare.
Ethics approval
Not applicable.
Consent to participate/publish
Not applicable.
Availability of data and material
Not applicable.
Code availability
Not applicable.
Authors contributions
JMTF and MIS designed the content and wrote the first draft, ARS reviewed the manuscript and made suggestions, JMTF produced the figures and revised the manuscript, MIS revised and submitted the manuscript.
References
- 1.Bhatt S, Gething PW, Brady OJ, Messina JP, Farlow AW, Moyes CL, Drake JM, et al. The global distribution and burden of dengue. Nature. 2013;496(7446):504–507. doi: 10.1038/nature12060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Zeng Z, Zhan J, Chen L, Chen H, Cheng S. Global, regional, and national dengue burden from 1990 to 2017: a systematic analysis based on the global burden of disease study 2017. E Clin Med. 2021;32:100712. doi: 10.1016/j.eclinm.2020.100712. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Colón-González FJ, Fezzi C, Lake IR, Hunter PR. The effects of weather and climate change on dengue. PLoS Negl Trop Dis. 2013;7(11):e2503. doi: 10.1371/journal.pntd.0002503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Deming R, Manrique-Saide P, Medina Barreiro A, Cardeña EU, Che-Mendoza A, Jones B, Liebman K, et al. Spatial variation of insecticide resistance in the dengue vector Aedes aegypti presents unique vector control challenges. Parasit Vectors. 2016;9:67. doi: 10.1186/s13071-016-1346-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Kraemer MUG, Reiner RC, Jr, Brady OJ, Messina JP, Gilbert M, Pigott DM, et al. Past and future spread of the arbovirus vectors Aedes aegypti and Aedes albopictus. Nat Microbiol. 2019;4(5):854–863. doi: 10.1038/s41564-019-0376-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Tripathi NK, Shrivastava A. Recent developments in recombinant protein-based dengue vaccines. Front Immunol. 2018;9:1919. doi: 10.3389/fimmu.2018.01919. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.da Silveira L, Tura B, Santos M. Systematic review of dengue vaccine efficacy. BMC Infect Dis. 2019;19(1):750. doi: 10.1186/s12879-019-4369-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Narayan R, Tripathi S. Intrinsic ADE: the dark side of antibody dependent enhancement during dengue infection. Front Cell Infect Microbiol. 2020;10:580096. doi: 10.3389/fcimb.2020.580096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Lin L, Koren MA, Paolino KM, Eckels KH, De La Barrera R, Friberg H, et al. Immunogenicity of a live-attenuated dengue vaccine using a heterologous prime-boost strategy in a phase 1 randomized clinical trial. J Infect Dis. 2021;223(10):1707–1716. doi: 10.1093/infdis/jiaa603. [DOI] [PubMed] [Google Scholar]
- 10.Fernandez S, Thomas SJ, De La Barrera R, Im-Erbsin R, Jarman RG, Baras B, et al. An adjuvanted, tetravalent dengue virus purified inactivated vaccine candidate induces long-lasting and protective antibody responses against dengue challenge in rhesus macaques. Am J Trop Med Hyg. 2015;92(4):698–708. doi: 10.4269/ajtmh.14-0268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Danko JR, Kochel T, Teneza-Mora N, Luke TC, Raviprakash K, Sun P, et al. Safety and immunogenicity of a tetravalent dengue DNA vaccine administered with a cationic lipid-based adjuvant in a phase 1 clinical trial. Am J Trop Med Hyg. 2018;98(3):849–856. doi: 10.4269/ajtmh.17-0416. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Manoff SB, Sausser M, Falk Russell A, Martin J, Radley D, Hyatt D, et al. Immunogenicity and safety of an investigational tetravalent recombinant subunit vaccine for dengue: results of a phase I randomized clinical trial in flavivirus-naïve adults. Hum Vaccin Immunother. 2019;15(9):2195–2204. doi: 10.1080/21645515.2018.1546523. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Endy TP, Wang D, Polhemus ME, Jarman RG, Jasper LE, Gromowski G, et al. A phase 1, open-label assessment of a dengue virus-1 live virus human challenge strain. J Infect Dis. 2021;223(2):258–267. doi: 10.1093/infdis/jiaa351. [DOI] [PubMed] [Google Scholar]
- 14.Biswal S, Reynales H, Saez-Llorens X, Lopez P, Borja-Tabora C, Kosalaraksa P, et al. Efficacy of a tetravalent dengue vaccine in healthy children and adolescents. N Engl J Med. 2019;381:2009–2019. doi: 10.1056/NEJMoa1903869. [DOI] [PubMed] [Google Scholar]
- 15.Whitehead SS. Development of TV003/TV005, a single dose, highly immunogenic live attenuated dengue vaccine; what makes this vaccine different from the Sanofi-Pasteur CYD™ vaccine? Expert Rev Vaccines. 2016;15(4):509–517. doi: 10.1586/14760584.2016.1115727. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Gruenberg A, Woo WS, Biedrzycka A, Wright PJ. Partial nucleotide sequence and deduced amino acid sequence of the structural proteins of dengue virus type 2, new Guinea C and PUO-218 strains. J Gen Virol. 1988;69:1391–1398. doi: 10.1099/0022-1317-69-6-1391. [DOI] [PubMed] [Google Scholar]
- 17.Guirakhoo F, Weltzin R, Chambers TJ, Zhang ZX, Soike K, Ratterree M, et al. Recombinant chimeric yellow fever-dengue type 2 virus is immunogenic and protective in nonhuman primates. J Virol. 2000;74:5477–5485. doi: 10.1128/JVI.74.12.5477-5485.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Guirakhoo F, Arroyo J, Pugachev KV, Miller C, Zhang ZX, Weltzin R, et al. Construction, safety, and immunogenicity in nonhuman primates of a chimeric yellow fever-dengue virus tetravalent vaccine. J Virol. 2001;75:7290–7304. doi: 10.1128/JVI.75.16.7290-7304.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Guirakhoo F, Pugachev K, Zhang Z, Myers G, Levenbook I, Draper K, et al. Safety and efficacy of chimeric yellow Fever-dengue virus tetravalent vaccine formulations in nonhuman primates. J Virol. 2004;78(9):4761–4775. doi: 10.1128/jvi.78.9.4761-4775.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Men R, Bray M, Clark D, Chanock RM, Lai CJ. Dengue type 4 virus mutants containing deletions in the 3’ noncoding region of the RNA genome: analysis of growth restriction in cell culture and altered viremia pattern and immunogenicity in rhesus monkeys. J Virol. 1996;70:3930–3937. doi: 10.1128/JVI.70.6.3930-3937.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Whitehead SS, Falgout B, Hanley KA, Blaney JE, Markoff L, Murphy BR. A live, attenuated dengue virus type 1 vaccine candidate with a 30-nucleotide deletion in the 3′ untranslated region is highly attenuated and immunogenic in monkeys. J Virol. 2003;77:1653–1657. doi: 10.1128/JVI.77.2.1653-1657.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Blaney JE, Sathe NS, Goddard L, Hanson CT, Romero TA, Hanley KA, et al. Dengue virus type 3 vaccine candidates generated by introduction of deletions in the 3′ untranslated region (3′-UTR) or by exchange of the DENV-3 3′-UTR with that of DENV-4. Vaccine. 2008;26:817–828. doi: 10.1016/j.vaccine.2007.11.082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Blaney JE, Whitehead SS, Hanley KA, Murphy BR, Hanson CT, Firestone CY. Genetically modified, live attenuated dengue virus type 3 vaccine candidates. Am J Trop Med Hyg. 2004;71:811–821. doi: 10.4269/ajtmh.2004.71.811. [DOI] [PubMed] [Google Scholar]
- 24.Yoksan S, Bhamarapravati N, Halstead S. Dengue virus vaccine development: study on biological markers of cloned dengue 1–4 viruses serially passaged in primary kidney cells. In: St. George TD, Kay BH, Blok J, editors. Arbovirus research in Australia, Proceedings of the fourth symposium, Brisbane: CSIRO/QIMR. 1986. p. 35–8.
- 25.Kinney RM, Butrapet S, Chang GJ, Tsuchiya KR, Roehrig JT, Bhamarapravati N, et al. Construction of infectious cDNA clones for dengue 2 virus: strain 16681 and its attenuated vaccine derivative, strain PDK-53. Virology. 1997;230:300–308. doi: 10.1006/viro.1997.8500. [DOI] [PubMed] [Google Scholar]
- 26.Butrapet S, Huang CY, Pierro DJ, Bhamarapravati N, Gubler DJ, Kinney RM. Attenuation markers of a candidate dengue type 2 vaccine virus, strain 16681 (PDK-53), are defined by mutations in the 5 noncoding region and nonstructural proteins 1 and 3. J Virol. 2000;74:3011–3019. doi: 10.1128/jvi.74.7.3011-3019.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Huang CY, Butrapet S, Tsuchiya KR, Bhamarapravati N, Gubler DJ, Kinney RM. Dengue 2 PDK-53 virus as a chimeric carrier for tetravalent dengue vaccine development. J Virol. 2003;77:11436–11447. doi: 10.1128/jvi.77.21.11436-11447.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Osorio JE, Brewoo J, Silengo SJ, Arguello J, Moldovan IR, Tary-Lehmann M, et al. Efficacy of a tetravalent chimeric dengue vaccine (DENVax) in cynomolgus macaques. Am J Trop Med Hyg. 2011;84(6):978–987. doi: 10.4269/ajtmh.2011.10-0592. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Thomas SJ, Yoon IK. A review of Dengvaxia®: development to deployment. Hum Vaccin Immunother. 2019;15(10):2295–2314. doi: 10.1080/21645515.2019.1658503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Tricou V, Low JG, Oh HM, Leo YS, Kalimuddin S, Wijaya L, et al. Safety and immunogenicity of a single dose of a tetravalent dengue vaccine with two different serotype-2 potencies in adults in Singapore: a phase 2, double-blind, randomised, controlled trial. Vaccine. 2020;38(6):1513–1519. doi: 10.1016/j.vaccine.2019.11.061. [DOI] [PubMed] [Google Scholar]
- 31.Tricou V, Sáez-Llorens X, Yu D, Rivera L, Jimeno J, Villarreal AC, et al. Safety and immunogenicity of a tetravalent dengue vaccine in children aged 2–17 years: a randomised, placebo-controlled, phase 2 trial. Lancet. 2020;395(10234):1434–1443. doi: 10.1016/S0140-6736(20)30556-0. [DOI] [PubMed] [Google Scholar]
- 32.van Der Most RG, Murali-Krishna K, Ahmed R, Strauss JH. Chimeric yellow fever/dengue virus as a candidate dengue vaccine: quantitation of the dengue virus-specific CD8 T-cell response. J Virol. 2000;74(17):8094–8101. doi: 10.1128/jvi.74.17.8094-8101.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Torresi J, Heron LG, Qiao M, Marjason J, Chambonneau L, Bouckenooghe A, et al. Lot-to-lot consistency of a tetravalent dengue vaccine in healthy adults in Australia: a randomised study. Vaccine. 2015;33(39):5127–5134. doi: 10.1016/j.vaccine.2015.08.008. [DOI] [PubMed] [Google Scholar]
- 34.Henein S, Swanstrom J, Byers AM, Moser JM, Shaik SF, Bonaparte M, et al. Dissecting antibodies induced by a chimeric yellow fever dengue, live-attenuated, tetravalent dengue vaccine (CYD-TDV) in naive and dengue-exposed individuals. J Infect Dis. 2017;215(3):351–358. doi: 10.1093/infdis/jiw576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Hadinegoro SR, Arredondo-Garcia JL, Capeding MRC, Deseda T, Chotpitayasunondh R, Dietze HI, et al. Efficacy and long-term safety of a dengue vaccine in regions of endemic disease. N Engl J Med. 2015;373:1195–1206. doi: 10.1056/NEJMoa1506223. [DOI] [PubMed] [Google Scholar]
- 36.Sridhar S, Luedtke A, Langevin E, Zhu M, Bonaparte M, Machabert T, et al. Effect of dengue serostatus on dengue vaccine safety and efficacy. N Engl J Med. 2018;379:327–340. doi: 10.1056/NEJMoa1800820. [DOI] [PubMed] [Google Scholar]
- 37.Sabchareon A, Wallace D, Sirivichayakul C, Limkittikul K, Chanthavanich P, Suvannadabba S, et al. Protective efficacy of the recombinant, live-attenuated, CYD tetravalent dengue vaccine in Thai schoolchildren: a randomised, controlled phase 2b trial. Lancet. 2012;380:1559–1567. doi: 10.1016/S0140-6736(12)61428-7. [DOI] [PubMed] [Google Scholar]
- 38.Villar L, Dayan GH, Arredondo-García JL, Rivera DM, Cunha R, Deseda C, et al. Efficacy of a tetravalent dengue vaccine in children in latin America. N Engl J Med. 2015;372:113–123. doi: 10.1056/NEJMoa1411037. [DOI] [PubMed] [Google Scholar]
- 39.Hadinegoro SR, Arredondo-García JL, Capeding MR, Deseda C, Chotpitayasunondh T, Dietze R, Saville M. Efficacy and long-term safety of a dengue vaccine in regions of endemic disease. N Engl J Med. 2015;373(13):1195–1206. doi: 10.1056/NEJMoa1506223. [DOI] [PubMed] [Google Scholar]
- 40.Halstead SB, Katzelnick LC, Russell PK, Markoff L, Aguiar M, Dans LR, et al. Ethics of a partially effective dengue vaccine: lessons from the Philippines. Vaccine. 2020;38:5572–5576. doi: 10.1016/j.vaccine.2020.06.079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Durbin AP, Kirkpatrick BD, Pierce KK, Schmidt AC, Whitehead SS. Development and clinical evaluation of multiple investigational monovalent DENV vaccines to identify components for inclusion in a live attenuated tetravalent DENV vaccine. Vaccine. 2011;29:7242–7250. doi: 10.1016/j.vaccine.2011.07.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Durbin AP, Schmidt A, Elwood D, Wanionek KA, Lovchik J, Thumar B, et al. Heterotypic dengue infection with live attenuated monotypic dengue virus vaccines: implications for vaccination of populations in areas where dengue is endemic. J Infect Dis. 2011;203:327–334. doi: 10.1093/infdis/jiq059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Durbin AP, Whitehead SS, Shaffer D, Elwood D, Wanionek K, Blaney JE, Jr, et al. A single dose of the DENV-1 candidate vaccine rDEN1D30 is strongly immunogenic and induces resistance to a second dose in a randomized trial. PLoS Negl Trop Dis. 2011;5:e1267. doi: 10.1371/journal.pntd.0001267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Lindow JC, Durbin AP, Whitehead SS, Pierce KK, Carmolli MP, Kirkpatrick BD. Vaccination of volunteers with low-dose, live-attenuated, dengue viruses leads to serotype-specific immunologic and virologic profiles. Vaccine. 2013;31:3347–3352. doi: 10.1016/j.vaccine.2013.05.075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Troyer JM, Hanley KA, Whitehead SS, Strickman D, Karron RA, Durbin AP, Murphy BR. A live attenuated recombinant dengue-4 virus vaccine candidate with restricted capacity for dissemination in mosquitoes and lack of transmission from vaccinees to mosquitoes. Am J Trop Med Hyg. 2001;65:414–419. doi: 10.4269/ajtmh.2001.65.414. [DOI] [PubMed] [Google Scholar]
- 46.Kirkpatrick BD, Durbin AP, Pierce KK, Carmolli MP, Tibery CM, Grier PL, et al. Robust and balanced immune responses to all 4 dengue virus serotypes following administration of a single dose of a live attenuated tetravalent dengue vaccine to healthy, flavivirus- naive adults. J Infect Dis. 2015;212:702–710. doi: 10.1093/infdis/jiv082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Durbin AP. Historical discourse on the development of the live attenuated tetravalent dengue vaccine candidate TV003/TV005. Curr Opin Virol. 2020;43:79–87. doi: 10.1016/j.coviro.2020.09.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Kallas EG, Precioso AR, Palacios R, Thome B, Braga PE, Vanni T, et al. Safety and immunogenicity of the tetravalent, live-attenuated dengue vaccine Butantan-DV in adults in Brazil: a two-step, double-blind, randomised placebo-controlled phase 2 trial. Lancet Infect Dis. 2020;20(7):839–850. doi: 10.1016/S1473-3099(20)30023-2. [DOI] [PubMed] [Google Scholar]
- 49.Jackson LA, Rupp R, Papadimitriou A, Wallace D, Raanan M, Moss KJ. A phase 1 study of safety and immunogenicity following intradermal administration of a tetravalent dengue vaccine candidate. Vaccine. 2018;36:3976–3983. doi: 10.1016/j.vaccine.2018.05.028. [DOI] [PubMed] [Google Scholar]
- 50.Osorio JE, Wallace D, Stinchcomb DT. A recombinant, chimeric tetravalent dengue vaccine candidate based on a dengue virus serotype 2 backbone. Expert Rev Vaccines. 2016;15:497–508. doi: 10.1586/14760584.2016.1128328. [DOI] [PubMed] [Google Scholar]
- 51.Rupp R, Luckasen GJ, Kirstein JL, Osorio JE, Santangelo JD, Raanan M, et al. Safety and immunogenicity of different doses and schedules of a live attenuated tetravalent dengue vaccine (TDV) in healthy adults: a Phase 1b randomized study. Vaccine. 2015;33(46):6351–6359. doi: 10.1016/j.vaccine.2015.09.008. [DOI] [PubMed] [Google Scholar]
- 52.Sáez-Llorens X, Tricou V, Yu D, Rivera L, Tuboi S, Garbes P, Borkowski A, Wallace D. Safety and immunogenicity of one versus two doses of Takeda's tetravalent dengue vaccine in children in Asia and Latin America: interim results from a phase 2, randomised, placebo-controlled study. Lancet Infect Dis. 2017;17(6):615–625. doi: 10.1016/S1473-3099(17)30166-4. [DOI] [PubMed] [Google Scholar]
- 53.Biswal S, Borja-Tabora C, Martinez Vargas L, Velásquez H, Theresa Alera M, Sierra V, TIDES study group et al. Efficacy of a tetravalent dengue vaccine in healthy children aged 4–16 years: a randomised, placebo-controlled, phase 3 trial. Lancet. 2020;395(10234):1423–1433. doi: 10.1016/S0140-6736(20)30414-1. [DOI] [PubMed] [Google Scholar]
- 54.Rivera L, Biswal S, Sáez-Llorens X, Reynales H, López-Medina E, Borja-Tabora C, TIDES study group et al. Three years efficacy and safety of Takeda's dengue vaccine candidate (TAK-003) Clin Infect Dis. 2021 doi: 10.1093/cid/ciab864. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Anderson KB, Gibbons RV, Cummings DA, Nisalak A, Green S, Libraty DH, et al. A shorter time interval between first and second dengue infections is associated with protection from clinical illness in a school-based cohort in Thailand. J Infect Dis. 2013;209:360–368. doi: 10.1093/infdis/jit436. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Patro A, Mohanty S, Prusty BK, Singh DK, Gaikwad S, Saswat T, et al. Cytokine signature associated with disease severity in dengue. Viruses. 2019;11(1):34. doi: 10.3390/v11010034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Srikiatkhachorn A, Mathew A, Rothman AL. Immune-mediated cytokine storm and its role in severe dengue. Semin Immunopathol. 2017;39(5):563–574. doi: 10.1007/s00281-017-0625-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Guo L, Rondina MT. The era of thromboinflammation: platelets are dynamic sensors and effector cells during infectious diseases. Front Immunol. 2019;10:2204. doi: 10.3389/fimmu.2019.02204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Hassan J, Borhany M, Abid M, Zaidi U, Fatima N, Shamsi T. Coagulation abnormalities in dengue and dengue haemorrhagic fever patients. Transfus Med. 2020;30(1):46–50. doi: 10.1111/tme.12658. [DOI] [PubMed] [Google Scholar]
- 60.Imad HA, Phumratanaprapin W, Phonrat B, Chotivanich K, Charunwatthana P, Muangnoicharoen S, et al. Cytokine expression in dengue fever and dengue hemorrhagic fever patients with bleeding and severe hepatitis. Am J Trop Med Hyg. 2020;102(5):943–950. doi: 10.4269/ajtmh.19-0487. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Lei HY, Yeh TM, Liu HS, Lin YS, Chen SH, Liu CC. Immunopathogenesis of dengue virus infection. J Biomed Sci. 2001;8(5):377–388. doi: 10.1007/BF02255946. [DOI] [PubMed] [Google Scholar]
- 62.Chuang YC, Lin YS, Li HS, Yeh TM. Molecular mimicry between dengue virus and coagulation factors induces antibodies to inhibit thrombin activity and enhance fibrinolysis. J Virol. 2014;88(23):13759–13768. doi: 10.1128/JVI.02166-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Katzelnick LC, Gresh L, Halloran ME, Mercado JC, Kuan G, Gordon A, et al. Antibody-dependent enhancement of severe dengue disease in humans. Science. 2017;358(6365):929–932. doi: 10.1126/science.aan6836. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Taylor A, Fo SS, Bruzzone R, Dinh LV, King NJ, Mahalinga S. Fc receptors in antibody-dependent enhancement of viral infections. Immunol Rev. 2015;268(1):340–364. doi: 10.1111/imr.12367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Priyamvada L, Cho A, Onlamoon N, Zheng NY, Huang M, Kovalenkov Y, et al. B cell responses during secondary dengue virus infection are dominated by highly cross-reactive, memory-derived plasmablasts. J Virol. 2016;90(12):5574–5585. doi: 10.1128/JVI.03203-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Halstead SB. Dengue antibody-dependent enhancement: knowns and unknowns. Microbiol Spectr. 2014 doi: 10.1128/microbiolspec.AID-0022-2014. [DOI] [PubMed] [Google Scholar]
- 67.Screaton G, Mongkolsapaya J, Yacoub S, Roberts C. New insights into the immunopathology and control of dengue virus infection. Nat Rev Immunol. 2015;15:745–759. doi: 10.1038/nri3916. [DOI] [PubMed] [Google Scholar]
- 68.Vidarsson G, Dekkers G, Rispens T. IgG subclasses and allotypes: from structure to effector functions. Front Immunol. 2014;5:520. doi: 10.3389/fimmu.2014.00520. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.de Taeye SW, Rispens T, Vidarsson G. The ligands for human IgG and their effector functions. Antibodies (Basel, Switzerland). 2019;8(2):30. doi: 10.3390/antib8020030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Jennewein MF, Alter G. The immunoregulatory roles of antibody glycosylation. Trends Immunol. 2017;38(5):358–372. doi: 10.1016/j.it.2017.02.004. [DOI] [PubMed] [Google Scholar]
- 71.Shukla R, Ramasamy V, Shanmugam RK, Ahuja R, Khanna N. Antibody-dependent enhancement: a challenge for developing a safe dengue vaccine. Front Cell Infect Microbiol. 2020;10:572681. doi: 10.3389/fcimb.2020.57268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Kawiecki AB, Christofferson RC. Zika virus-induced antibody response enhances dengue virus serotype 2 replication in vitro. J Infect Dis. 2016;214(9):1357–1360. doi: 10.1093/infdis/jiw377. [DOI] [PubMed] [Google Scholar]
- 73.Shukla R, Beesetti H, Brown JA, Ahuja R, Ramasamy V, Shanmugam RK, et al. Dengue and Zika virus infections are enhanced by live attenuated dengue vaccine but not by recombinant DSV4 vaccine candidate in mouse models. EBioMedicine. 2020;60:102991. doi: 10.1016/j.ebiom.2020.102991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Li L, Meng W, Horton M, DiStefano DR, Thoryk EA, Pfaff JM, et al. Potent neutralizing antibodies elicited by dengue vaccine in rhesus macaque target diverse epitopes. PLoS Pathog. 2019;15(6):e1007716. doi: 10.1371/journal.ppat.1007716. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.de Alwis R, Williams KL, Schmid MA, Lai CY, Patel B, Smith SA, et al. Dengue viruses are enhanced by distinct populations of serotype cross-reactive antibodies in human immune sera. PLoS Pathog. 2014;10(10):e1004386. doi: 10.1371/journal.ppat.1004386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Wahala WM, Silva AM. The human antibody response to dengue virus infection. Viruses. 2011;3(12):2374–2395. doi: 10.3390/v3122374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Izmirly AM, Alturki SO, Alturki SO, Connors J, Haddad EK. Challenges in dengue vaccines development: pre-existing infections and cross-reactivity. Front Immunol. 2020;11:1055. doi: 10.3389/fimmu.2020.01055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Murphy BR, Whitehead SS. Immune response to dengue virus and prospects for a vaccine. Annu Rev Immunol. 2011;29:587–619. doi: 10.1146/annurev-immunol-031210-101315. [DOI] [PubMed] [Google Scholar]
- 79.Ly MHP, Moi ML, Vu TBH, Tun MMN, Saunders T, Nguyen CN, et al. Dengue virus infection-enhancement activity in neutralizing antibodies of healthy adults before dengue season as determined by using FcγR-expressing cells. BMC Infect Dis. 2018;18:31. doi: 10.1186/s12879-017-2894-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Salmon DA, Lambert PH, Nohynek HM, Gee J, Parashar UD, Tate JE, et al. Novel vaccine safety issues and areas that would benefit from further research. BMJ Glob Health. 2021;6(Suppl 2):e003814. doi: 10.1136/bmjgh-2020-003814. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Turner M, Papadimitriou A, Winkle P, Segall N, Levin M, Doust M, et al. Immunogenicity and safety of lyophilized and liquid dengue tetravalent vaccine candidate formulations in healthy adults: a randomized, phase 2 clinical trial. Hum Vaccin Immunother. 2020;16(10):2456–2464. doi: 10.1080/21645515.2020.1727697. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Magnani DM, Silveira C, Ricciardi MJ, Gonzalez-Nieto L, Pedreño-Lopez N, Bailey VK, et al. Potent plasmablast-derived antibodies elicited by the national institutes of health dengue vaccine. J Virol. 2017;91(22):e00867–e917. doi: 10.1128/JVI.00867-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Nivarthi UK, Swanstrom J, Delacruz MJ, et al. A tetravalent live attenuated dengue virus vaccine stimulates balanced immunity to multiple serotypes in humans. Nat Commun. 2021;12:1102. doi: 10.1038/s41467-021-21384-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Galula JU, Salem GM, Chang GJ, Chao DY. Does structurally-mature dengue virion matter in vaccine preparation in post-Dengvaxia®era? Hum Vaccin Immunother. 2019;15(10):2328–2336. doi: 10.1080/21645515.2019.1643676. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Wilken L, Rimmelzwaan GF. Adaptive immunity to dengue virus: slippery slope or solid ground for rational vaccine design? Pathogens. 2020;9(6):470. doi: 10.3390/pathogens9060470.PMID:32549226;PMCID:PMC7350362. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Capeding MR, Tran NH, Hadinegoro SR, Ismail HI, Chotpitayasunondh T, Chua MN, CYD14 Study Group et al. Clinical efficacy and safety of a novel tetravalent dengue vaccine in healthy children in Asia: a phase 3, randomised, observer-masked, placebo-controlled trial. Lancet. 2014;384(9951):1358–1365. doi: 10.1016/S0140-6736(14)61060-6. [DOI] [PubMed] [Google Scholar]