

Abstract
Cancer is one of the leading causes of death worldwide, and the factors responsible for its progression need to be elucidated. Exosomes are structures with an average size of 100 nm that can transport proteins, lipids, and nucleic acids. This review focuses on the role of exosomes in cancer progression and therapy. We discuss how exosomes are able to modulate components of the tumor microenvironment and influence proliferation and migration rates of cancer cells. We also highlight that, depending on their cargo, exosomes can suppress or promote tumor cell progression and can enhance or reduce cancer cell response to radio- and chemo-therapies. In addition, we describe how exosomes can trigger chronic inflammation and lead to immune evasion and tumor progression by focusing on their ability to transfer non-coding RNAs between cells and modulate other molecular signaling pathways such as PTEN and PI3K/Akt in cancer. Subsequently, we discuss the use of exosomes as carriers of anti-tumor agents and genetic tools to control cancer progression. We then discuss the role of tumor-derived exosomes in carcinogenesis. Finally, we devote a section to the study of exosomes as diagnostic and prognostic tools in clinical courses that is important for the treatment of cancer patients. This review provides a comprehensive understanding of the role of exosomes in cancer therapy, focusing on their therapeutic value in cancer progression and remodeling of the tumor microenvironment.
Graphical Abstract
Keywords: Exosome, Cancer, Immunotherapy, Non-coding RNA, Biomarker, Prognosis
Introduction
Cancer remains one of the diseases that threaten the lives of many people around the world. It is the second leading cause of death worldwide after cardiovascular diseases. Cancer cells possess unique features such as high proliferation rate, self-renewal ability, cancer stem cell (CSC) characteristics, metastasis, and the ability to switch between different molecular pathways to develop drug resistance [1–4]. Based on these properties, novel therapeutics, including nucleic acid drugs and anti-cancer agents, have been developed to target cancer cells and suppress their progression [5–9]. In addition, novel methods such as the use of nanoparticles have been employed for targeted delivery of therapeutics to cancer cells [10].
Recently, attention has focused on the role of a new type of structure, called extracellular vesicles (EVs) in cancer [11, 12]. EVs originate from the cell membrane and are considered micro- or nanovesicles. These structures can be secreted by all prokaryotic and eukaryotic cells in an evolutionarily conserved manner [13, 14]. Initially, EVs were thought to be waste products of cells or entities formed by cellular damage [15]. However, further studies on EVs have shown that they have vital biological functions and are important cellular components [16, 17]. There are several types of EVs that are categorized based on their size, origin, and localization [18–20]. The best known EVs include exosomes, microparticles, shedding vesicles, apoptotic bodies, tolerosomes, proteasomes, and prominosomes [21, 22]. There are two different mechanisms for the formation of EVs. In the first mechanism, EVs arise directly from cell membrane budding [15]. In the second mechanism, EVs arise during exocytosis of multivesicular bodies as part of the endocytosis system [23]. EVs are involved in biological functions in cells and play an important role in pathological conditions. They can transfer various molecules between cells and are a means of communication [24]. Therefore, special attention should be paid to their role in diseases, especially cancer [25–30].
The present review focuses on the role of exosomes in cancer. This comprehensive review first provides an overview of the discovery of exosomes, their composition, and the pathway of their biogenesis, which are of important for understanding these structures. Then, we focus specifically on the role of exosomes in cancer by introducing a section on exosomes in tumor microenvironment (TME) remodeling and how they influence various cancer hallmarks, including proliferation, migration, and therapy response. Next, we discuss exosomal non-coding RNAs (ncRNAs) and how they can affect cancer cell progression. We then turn our attention to exosomes and the key molecular signaling pathways that regulate cancer progression. Finally, we provide insight into tumor-derived exosomes and the clinical applications of exosomes relevant to the treatment of cancer patients.
Exosome structure, isolation and dosing
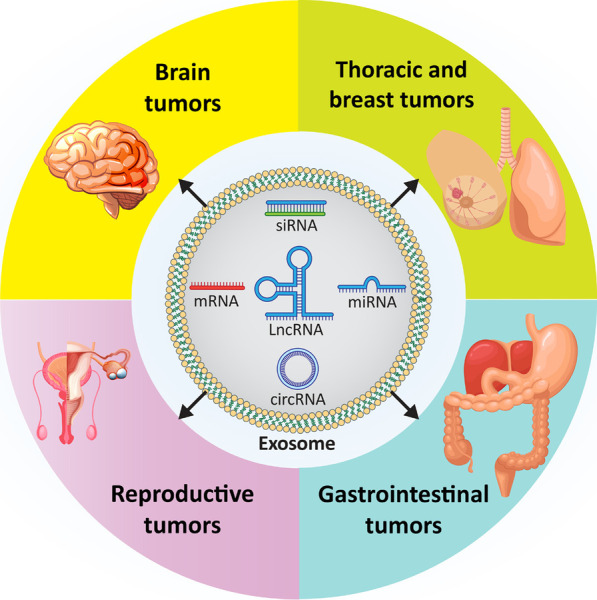
Exosomes are double-membraned vesicles (30–150 nm in size; average particle size: 100 nm) secreted by different types of cells. Their specific functions depend on their origin. For example, exosomes originating from tumor cells provide cell-to-cell communication and are mainly involved in migration and invasion [31]. In the phospholipid membrane of exosomes originating from the parent cell, there are a variety of proteins and lipids [32, 33]. Among the lipid molecules, phosphatidylcholine, phosphatidylethanolamines, phosphatidylinositol, phosphatidylserine, and sphingomyelin are present in the exosome membrane. The composition and levels of these lipid molecules mainly influence the properties of exosomes. For example, the high stability of exosomes in body fluids and at different pH values is due to the high levels of sphingomyelin and phosphatidylinositol in their membrane. Therefore, these lipid molecules protect exosomes from degradation by proteolytic or lipolytic enzymes [34]. The phospholipid membrane of exosomes has lipid rafts containing proteins such as tyrosine kinase Src and glycosylphosphatidylinositol-containing proteins [35]. The presence of proteins in exosomes is a bit complex. Exosomes are thought to contain both general and specific proteins. The general or nonspecific proteins are present in all cell types, including CD63, tetraspanins, CD81, and CD9, whereas specific proteins include MHC II found in exosomes from dendritic cells and B lymphocytes, HER2 in exosomes from breast cancer, and EGFR in exosomes from gliomas [36]. It is worth noting that nonspecific proteins are critical for exosome function. Tetraspanins, for example, are nonspecific proteins that can interact with integrin or MHC molecules and form complexes. In addition to proteins, exosomes may also contain ncRNAs including microRNAs (miRNAs), long noncoding RNAs (lncRNAs) and circular RNAs (circRNAs) [37–39].
Since exosomes are present in various body fluids, they can be considered as novel biomarkers for the detection and diagnosis of various diseases. Therefore, it is important to develop methods for their isolation. A total of six strategies have been developed for the isolation of exosomes, including ultracentrifugation, ultrafiltration, size exclusion chromatography, precipitation, immunoaffinity-based capture, and microfluidics. Each method has its own advantages and problems that should be addressed [40–45]. Ultracentrifugation is capable of detecting exosomes based on their density, size, and shape, and its advantages include affordability, large sample capacity, and ability to isolate high concentrations of exosomes. The disadvantages of ultracentrifugation are the time-consuming process, the risk of exosome damage to exosomes from high-speed centrifugation, and the need for complex equipment [46–48]. Ultrafiltration isolates exosomes based on size differences from other particles. This strategy is fast and portable but has drawbacks such as low purity, shear stress, exosome loss, and clogging [49–51]. Size exclusion chromatography also uses size differences and has the advantage that it can accurately separate exosomes and isolate the intact exosomes without damaging them. Its disadvantage is the time-consuming process, which needs further advancement and development [52–54]. The precipitation method is based on changing the solubility of exosomes, and its advantages include the ease of performance, applicability to large sample volumes, and little damage to exosomes. Its disadvantages include the time-consuming process and the possibility of precipitating other particles such as polymeric materials and proteins [55, 56]. Immunoaffinity-based capture is based on the interaction between antibodies and antigens. Advantages of this method include high purity and the possibility of subtyping, whereas problems include high cost, low yield, risk of antigen blockade, and loss of exosome functionality [57]. The final technique for exosome isolation is the microfluidic strategy, which has the advantage of being inexpensive, time-saving, and requiring only a small amount of sample, but it has low sensitivity [58–62]. Further information on exosome isolation techniques has been reviewed elsewhere [63, 64].
The dosing of exosome has been the subject of debate and investigation in recent years. Three different methods have been used to determine exosome dosage, including cell equivalents, protein concentration, and/or specific quantitative analytical measurements using tools, with each with its own advantages and disadvantages. However, there is still a need to develop a standardized method for exosome dosing and currently available technologies suffer from accurate and precise assessment of exosomes at the level of individual vesicles. To improve the accuracy in exosome dosing, it is proposed to use multiple methods. For example, although the protein method that assesses total protein levels is fast and inexpensive, it may also assess proteins that are not exosome-related and may not indicate bioactive ingredients. TRPS, NTA, ELISA, cell equivalents, and flow cytometry are other methods for exosome dosing. A review by Willis and colleagues provides more details on techniques related to exosome dosing [65]. With regard to the use of exosomes in clinical trials, good manufacturing practices (GMPs) are important. Indeed, exosomes used in clinical trials should comply with GMPs. GMPs for exosomes consider three major factors, including upstream cell cultivation, downstream purification process, and exosome quality control [66].
Biogenesis route of exosomes
Exosomes are formed by the endocytic pathway after passing through several steps [67]. In the first step, invaginations of the cytoplasmic membrane generate an early secretory endosome. Then, biogenesis of multivesicular bodies (MVBs) occurs by inward sprouting, generating intraluminal vesicles (ILVs) surrounded by endosomes. Acidification is then required for maturation of the late endosomes. In the final step, the ILVs fuse with the cell membrane and the exosomes are released [68]. MVBs have a size of 250–100 nm and therefore multiple ILVs with a particle size of 30–150 nm can be formed within the MVBs [69]. A number of proteins are involved in the formation of ILVs and MVBs, and in cargo selection [70]. The best known proteins for exosome biogenesis are the endosomal sorting complexes required for transport (ESCRT), which consist of four members, including ESCRT-0, -I, -II, and -III, that play a special role in membrane formation and cargo sorting [71]. Association of ubiquitylated cargoes with lipid microdomains is performed by ESCRT-0 and ESCRT-I. Then, ESCRT-II and -III are involved in invagination and formation of MVBs and ILVs. ALIX (Apoptosis-linked gene 2-interacting protein X, encoded by PDCD6IP), VTA1 (Vesicle Trafficking 1), VPS4 (Vacuolar protein sorting-associated protein 4), and TSG101 (Tumor susceptibility gene 101 protein) are other proteins that help the ESCRT machinery in exosome biogenesis.
Of note, there is another pathway for exosome biogenesis that is independent of ESCRT. In this ESCRT-independent pathway, heat shock protein-60 (HSP60), HSP70, and HSP90 act as chaperones, and CD63, CD81, CD82, CD37, and CD9 act as tetraspanins, which play important roles in membrane formation and cargo binding to lipid microdomains. Therefore, the mechanisms of exosome biogenesis are divided into two categories: ESCRT-dependent and -independent mechanisms [13, 72–75]. Among the tetraspanins, CD63 and CD81 are the most abundant proteins in the membranes of ILVs and are considered to be markers for exosomes [75, 76].
The preferred mechanism of exosome biogenesis (either ESCRT-dependent or -independent pathway) is determined by cargo and the specific cell type [74]. Two major proteins play notable roles in the transfer and fusion of MVBs: the Ras-associated binding (Rab) family of GTPases and the soluble NSF attachment protein receptor (SNARE) [72, 77, 78]. It is worth noting that some of the MVBs are not fused to cell membranes and are transferred to lysosomes for degradation [72, 77]. The ILVs secreted from MVBs are known as exosomes. There are some limitations to exosome biogenesis and related mechanisms that may be considered in future studies. The underlying mechanism of differentiation of MVBs destined for degradation or fusion with the cell membrane is not known and needs to be studied in detail. Another limitation is the mechanism by which the endocytic system regulates the percentage of MVBs destined for fusion with the cell membrane. In addition, how the sorting of materials from ILVs to MVBs is regulated is still unknown [79]. Figure 1 shows a schematic representation of the biogenesis of exosomes.
Fig. 1.

The biogenesis of exosomes. Exosomes contain various types of cargoes such as siRNA, circRNA, lncRNA, mRNA, miRNA, lipids, and proteins, and are therefore involved in various biological mechanisms in cells. They have a particle size of 30–150 nm and various types of proteins shown in the figure may be involved in the biogenesis of exosomes. Targeting these proteins may regulate exosome biogenesis and provide new insights for the development of therapeutics
Exosomes and the tumor microenvironment
Most of the tumor mass is occupied by the TME, which comprises the stroma of the tumor [80]. Low oxygen levels, high lactate levels, extracellular acidosis, and poor nutrient content are prominent features of the TME [81, 82]. A variety of cells, including mesenchymal stem cells, fibroblasts, endothelial cells, and immune cells, are present in the TME and can secrete cytokines and growth factors [83]. Cancer-associated fibroblasts are one of the most abundant cells in the TME, creating conditions for tumor growth and progression [84, 85]. The interactions that occur in the TME and the activation/inhibition of signaling networks may determine tumor progression. Therefore, much attention has been devoted to understanding the interactions and developing targeted therapies for the TME [86–88]. This section summarizes the role of exosomes in influencing TME components.
Macrophages are abundant in the TME and have two distinct phenotypes, including M1- and M2-polarized macrophages [89]. Changing the polarization of macrophages toward the M2 phenotype leads to tumor progression and an event that mediates therapy resistance [90, 91]. One of the molecular signaling pathways shown to play an oncogenic role is the signal transducer and activator of transcription 3 (STAT3) pathway [92–96]. A recent experiment attempted to establish a link between STAT3, exosomes, and macrophage polarization in gliomas. The hypoxic state leads to the secretion of exosomes from glioma cells, which subsequently promote cancer progression by inducing M2 polarization of macrophages by triggering autophagy. Exosomes contain high levels of interleukin-6 (IL-6) and miRNA-155-3p. Activation of STAT3 occurs through IL-6, which in turn enhances the expression of miRNA-155-3p to induce autophagy. Due to a positive feedback loop, induced autophagy enhances STAT3 phosphorylation and thus tumorigenesis. Exosome-induced autophagy leads to M2 polarization of macrophages and paves the way for enhanced glioma progression [97]. Similar to glioma, the presence of hypoxia leads to the secretion of exosomes in the TME of colorectal carcinoma. These exosomes contain high levels of miRNA-210-3p, which inhibit apoptosis and promote the transition from G1 to S cycle by downregulating the expression of CELF2. Clinical investigation has also revealed that exosomes containing miRNA-210-3p have high levels in colorectal cancer patients and are correlated with an unfavorable prognosis [98]. Therefore, the signaling networks affected by exosomes may determine tumor progression by influencing TME [99, 100].
Now, the question arises: how can macrophages promote cancer cell migration and invasion? Polarized M2 macrophages are capable of secreting exosomes that promote hepatocellular carcinoma cell metastasis. M2 macrophages-derived exosomes transfer CD11b/CD18 to hepatocellular carcinoma cells. Subsequently, matrix metalloproteinase-9 (MMP-9) is activated, which significantly promotes cancer migration and metastasis [101]. Considering this important role of macrophages in cancer progression, exosomes targeting the TME have been developed. Galectin-9 siRNA was loaded into exosomes and then oxaliplatin was embedded as an antitumor agent and trigger of immunogenic cell death. Exosome-delivered galectin-9 siRNA suppressed M2 polarization of macrophages and oxaliplatin inhibited pancreatic cancer progression [102]. This study demonstrates how exosomes can reprogram the TME in favor of anticancer activity.
Because of the potential of exosomes to affect the TME, efforts have been made to develop exosomes that target the TME and regulate cancer progression. In a recent experiment, exosomes were loaded with manganese carbonyl to mediate their delivery to the TME. This resulted in increased formation of reactive oxygen species (ROS) and was able to reduce tumor proliferation by up to 90% during low-dose radiotherapy [103]. In addition, exosomes that are responsive to inflammatory TME were developed and, because of their ability to cross the blood–brain barrier (BBB), effectively transport doxorubicin into the TME and suppress glioma progression [104]. Therefore, exosomes may be considered promising candidates for targeting the TME and influencing cancer progression. When exosomes are present in the TME, a number of agents such as cytokines can alter their surface. For example, a recent experiment has shown that the surface of exosomes is modified by the CCL2 cytokine via binding to glycosaminoglycan side chains of proteoglycans, altering their cellular uptake and tropism toward certain cells and tissues [105]. Thus, if exosomes are to be manipulated, their interaction with components of the TME and the modification of their cellular uptake should be emphasized. Overall, exosomes exhibit interactions with the TME [106] and further experimentation is needed in basic research, in the development of exosomes for targeting the TME, and also in the introduction of these concepts into clinical courses (Fig. 2).
Fig. 2.

Exosomes in the regulation of the TME. Proliferation and metastasis of tumor cells are strongly modulated by the TME. Exosomes can influence various cellular interactions in the TME and affect tumor progression. In addition, exosomes can transport both anti-tumor agents (oxaliplatin) and siRNA into the TME and modulate tumor growth
Exosomes and tumor angiogenesis
Angiogenesis and vasculogenesis are considered the two most important mechanisms for the formation of new vessels [107–110]. However, there are major differences between angiogenesis and vasculogenesis. Vasculogenesis is involved in the formation of a whole vessel during embryonic development and is responsible for the development of the cardiovascular system. Thus, the endoderm releases vascular endothelial growth factor (VEGF), which induces VEGF receptor 2 (VEGFR2) on mesodermal cells in a paracrine manner [111]. Subsequently, mesodermal cells are converted into angioblasts or endothelial progenitor cells in the periphery of blood islands, whose fusion leads to the formation of primitive capillary networks [112]. Whereas vasculogenesis refers to the formation of new vessels, angiogenesis is the process of vessel formation from preexisting vessels [113]. The process of angiogenesis is inactive in adults and can be observed in physiological processes such as placental angiogenesis and embryo implantation [114–116]. Both angiogenesis and vasculogenesis are critical to the process of wound healing and facilitate this process [117]. Recently, attention has focused on the role of angiogenesis in cancer. Tumor cells should induce angiogenesis to ensure their survival, grow, and spread to different parts of the body. It has been reported that cancer cells cannot grow beyond a size of 1–2 mm if angiogenesis does not occur. Therefore, a promising strategy in cancer treatment could be the inhibition of angiogenesis. The best known factor responsible for the induction of angiogenesis is VEGF, a cytokine involved in cancer progression [118]. The activity of VEGF in neovascularization is related to its binding to receptors such as VEGFR1 and VEGFR2. In addition, VEGF has an affinity for binding to cofactors such as neuropilin-1 (NRP-1) and NRP-2. VEGFR2 expression is mainly observed in endothelial cells, whereas VEGFR1 is found on macrophages, cancer cells, and fibroblasts. The use of monoclonal antibodies is of interest for inhibition of VEGF or VEGFR and suppression of angiogenesis [119].
Since induction of angiogenesis promotes cancer progression, tumor cells secrete exosomes to trigger this mechanism. In this case, multiple molecular signaling pathways are involved that ultimately induce angiogenesis. Oral squamous cell carcinoma (OSCC) cells are able to secrete exosomes containing miRNA-210-3p. Upregulation of miRNA-210-3p occurs in OSCC cells and acts as a tumor-promoting factor by increasing microvessel density (MD) and tumor grade. Mechanistically, exosomal miRNA-210-3p reduces ephrin A3 expression to stimulate the PI3K/Akt axis, trigger angiogenesis, and promote OSCC progression [120]. Indeed, exosomes function as tools of cell–cell communication and can influence the conditions that promote cancer progression. Nasopharyngeal carcinoma (NPC) cells have a high migratory capacity that has been linked to their ability to trigger angiogenesis. Exosomal miRNA-23a binds to the 3’-UTR of TSGA10 and reduces its expression, leading to angiogenesis and increased metastasis of NPC cells [121]. The question now arises: since exosomes are able to regulate angiogenesis, can we isolate exosomes that suppress angiogenesis and thereby impair cancer progression? The answer is affirmative, and such a strategy has already been used in the treatment of lung cancer. It has been reported that exosomes derived from Plasmodium-infected mice inhibit angiogenesis. To test this hypothesis, an animal model of Plasmodium infection was developed in an experiment and then exosomes were isolated for the treatment of lung cancer. These exosomes contained high levels of miRNA-16, -322, -497, and -17, and when injected into a mouse model of lung cancer, there was a significant reduction in the expression of VEGFR2, resulting in inhibition of angiogenesis and reduced tumor progression [122]. This experiment clearly indicates that more studies should be conducted on exosomes and their role in affecting angiogenesis. By developing isolation methods, such exosomes can be obtained and their potential for cancer treatment can be revealed.
In addition to miRNAs, exosomes may also contain lncRNAs involved in the regulation of angiogenesis. In this case, the induction or inhibition of angiogenesis depends on the role of lncRNA as a tumor-suppressor or tumor-promoter. The lncRNA GAS5 is thought to suppress lung cancer progression. Exosomes containing high levels of GAS5 stimulate apoptosis in lung cancer and impair its growth. To this end, exosomal lncRNA GAS5 reduces miRNA-29-3p expression to increase PTEN expression. Subsequently, activated PTEN signaling suppresses PI3K/Akt phosphorylation to inhibit angiogenesis [123]. The role of exosomal ncRNAs in cancer progression will be discussed mechanistically in the next sections. However, it is clear that one way to modulate cancer progression is to influence angiogenesis through exosomes.
Angiopoietin-2 (ANGPT2) is thought to mediate resistance to antiangiogenic therapy by destroying vascular stability and promoting angiogenesis [124]. Suppression of the ANGPT2/Tie2 axis is a promising target [125, 126] because studies have shown the role of this factor in angiogenesis of cancer angiogenesis and in inflammation [127, 128]. Hepatocellular carcinoma (HCC) cells are capable of secreting ANGPT2-containing exosomes. These exosomes are introduced into HUVECs by endocytosis, and increased expression of ANGPT2 induces angiogenesis that promotes cancer progression [129]. As more experiments are performed, the novel signaling networks involved in angiogenesis are revealed. Hypoxia is a common feature of the TME. Recent experiments have shown that hypoxia can induce the secretion of exosomes from tumor cells, which increases their stemness and proliferation rate [130, 131]. A similar phenomenon occurs in colorectal cancer, where hypoxia leads to the secretion of exosomes, which in turn promote both growth and migration of tumor cells. Inhibition of exosome secretion by silencing RAB27a impairs proliferation and growth of colorectal tumors. Under hypoxic conditions, hypoxia-inducible factor-1α (HIF-1α) induces the secretion of exosomes containing Wnt4a. Subsequently, the β-catenin signaling pathway is activated and the nucleus is translocated, leading to angiogenesis and colorectal cancer progression [132].
In the previous sections, we have shown that exosomes affect the TME. The interaction of exosomes with the components of the TME may influence angiogenesis and thus cancer progression. Several experiments have shown that macrophages can induce angiogenesis. Recruitment of macrophages can induce angiogenesis to enhance nerve regeneration [133]. In addition, reduction or depletion of macrophages suppresses angiogenesis [134]. Tumor-derived exosomes (TEX) are capable of carrying CD39/CD73 and adenosine, which are enzymatically active. The TEX leads to polarization of macrophages into the M2 phenotype via A2BR. Subsequently, M2 macrophages secrete angiogenic factors (ANGPT2, IL-8, MMP9, PF4, and TIMP-1) that induce angiogenesis and promote cancer progression [135]. Overall, the studies are consistent with the fact that angiogenic factors are strongly regulated by exosomes. Depending on the cargo of exosomes, they can act as tumor suppressive or tumor promoting factors to influence angiogenesis in cancer cells. Exosomes can affect various molecular signaling pathways such as MAPK, YAP, VEGF, and miRNAs in modulating angiogenesis in cancer cells (Table 1) [136–141]. Figure 3 illustrates the role of exosomes in regulating angiogenesis in cancer cells.
Table 1.
Exosomes and their association with angiogenesis in different cancers
| Cancer type | In vitro/In vivo | Cell line/animal model | Signaling network | Remarks | Refs |
|---|---|---|---|---|---|
| Thyroid cancer |
In vitro In vivo |
Nthy-ori-3–1 cells |
miRNA-21-5p/TGFBI miRNA-21-5p/COL4A1 |
miRNA-21-5p in exosomes is upregulated under hypoxic conditions Angiogenesis is induced TGFBI and COL4A1 are inhibited by miRNA-21-5p to promote angiogenesis and cancer progression |
[217] |
| Esophageal squamous cell carcinoma |
In vitro In vivo |
ECA109, KYSE410 and HET-1A cell lines Nude mice |
– | Angiogenesis is promoted by the increased levels of exosomes under hypoxic conditions | [218] |
| Head and neck squamous cell carcinoma |
In vitro In vivo |
PCI-13 (HPV−) and UMSCC47 (HPV+) cell lines Mouse model |
– |
Functional reprogramming and phenotypic modulation are observed in endothelial cells Vascular structure formation is increased Proliferation and invasion are promoted Angiogenesis is induced by exosomes carrying angiogenic proteins |
[219] |
| Gastric cancer |
In vitro In vivo |
SGC7901 cells Xenograft model |
miRNA-155/FOXO3a | FOXO3a is inhibited by miRNA-155 in exosomes to induce angiogenesis in gastric to drive cancer progression | [220] |
| Gastric cancer | In vitro | HUVECs | YB-1/VEGF |
Exosomes derived from gastric cancer cells have high levels of YB-1 Apoptosis is inhibited, and metastasis and angiogenesis are enhanced Protein and mRNA levels of VEGF are increased |
[221] |
| Gastric cancer | In vitro | SGC7901 and MGC803 cells | miRNA-6785-5p/INHBA | INHBA expression is reduced by exosomes containing miRNA-6785-5p to impair migration and angiogenesis of cancer cells | [222] |
| Gastric tumor | In vitro | SGC7901 cells | miRNA-135b/FOXO1 | FOXO1 expression is decreased by the overexpression of miRNA-135b in exosomes to induce angiogenesis and exert tumor-promoting effects | [223] |
| Endometrial cancer | In vitro | SPEC2 and ISK cells | LGALS3BP/PI3K/Akt/VEGFA |
Cancer progression is enhanced by exosomes containing LGALS3BP Associated with unfavorable prognosis VEGFA expression is induced by triggering the PI3K/Akt axis Angiogenesis is promoted |
[224] |
| Breast cancer | In vitro | MDA-MB-231, MCF-7 and T47D cells | miRNA-100/mTOR/HIF-1α/VEGF |
miRNA-100 is delivered by exosomes in breast cancer therapy Angiogenesis is suppressed to impair cancer progression VEGF expression is downregulated in a time-dependent manner The mTOR/HIF-1α axis is suppressed |
[225] |
| Breast cancer | In vitro | 4T1 cells | miRNA-16/VEGF |
VEGF is downregulated by exosomes containing miRNA-16 Angiogenesis is suppressed Cancer progression is impaired |
[226] |
| Ovarian cancer | In vitro | A2780 and HO-8910 cells | PKR1/STAT3 |
Ovarian cancer migration is promoted by exosomes containing PKR1 through induction of angiogenesis Phosphorylation level of STAT3 is increased by PKR1 |
[227] |
| Ovarian cancer | In vitro | SKOV3 cells | miRNA-130a |
miRNA-130a is delivered by exosomes Angiogenesis is increased to promote cancer progression and trigger drug resistance |
[228] |
| Small-cell lung cancer |
In vitro In vivo |
H446 cells Xenograft model |
Profilin 2 |
Migration and tube formation capacity of endothelial cells are enhanced Smad2/3 is stimulated by profilin 2 in H446 cells Cancer development and metastasis are enhanced by exosomes |
[229] |
| Colon cancer |
In vitro In vivo |
HCT-15 cells Nude mice |
GDF15/Smad/periostin |
Angiogenesis is enhanced by exosomes derived from cancer cells Smad is inhibited by GDF15 to enhance periostin expression to promote angiogenesis |
[229] |
| Colorectal cancer |
In vitro In vivo |
LoVo and HT29 cells | miRNA-135b-5p | Angiogenesis is triggered, and proliferation and migration are enhanced | [230] |
| Renal cancer | In vitro | 789-0 cells | hepaCAM/VEGF | VEGF is downregulated and angiogenesis is inhibited by exosomes enriched with hepaCAM | [231] |
| Renal cancer |
In vitro In vivo |
786-0 cells Nude mice |
miRNA-27a/SFRP1/VEGF | SFRP1 is downregulated and VEGF expression is increased by miRNA-27a delivered by exosomes to trigger angiogenesis and promote cancer progression | [232] |
Fig. 3.

Exosomes in the modulation of angiogenesis in cancer cells. The molecular signaling pathways that regulate angiogenesis, including Akt, PTEN, β-catenin, TSGA10, and ANGPT2, are regulated by exosomes. Induction of angiogenesis promotes tumor progression and therapeutic targeting of exosomes may impair cancer growth
Exosomes and tumor growth
Proliferation of cancer cells increases abnormally and is one of the factors by which cancer cells differ from normal cells. Increased cell cycle progression, inhibition of apoptosis, and glycolysis are considered to be major main factors in cancer growth [142–146]. The aim of this section is to evaluate the role of exosomes in modulating proliferation of cancer cells. Rapidly dividing cancer cells require high levels of energy to sustain their proliferation. Oxidative phosphorylation is a slow process and cannot provide tumor cells with energy to meet their needs. Therefore, a switch in metabolism from oxidative phosphorylation to glycolysis is initiated. Therefore, suppression of glycolysis can be considered a promising strategy in cancer therapy [147–150]. Exosomes have been shown to increase the growth rate of lung cancer cells via the induction of glycolysis. To this end, exosomes were isolated from irradiated lung cancer cells. They exhibited high levels of ALDOA and ALDH3A1, which stimulate glycolysis to increase lung cancer proliferation [151]. With regard to the close link between drug resistance and glycolysis, studies have attempted to elucidate a link between exosomes, glycolysis, and chemoresistance. Macrophages are capable of secreting exosomes that play a tumor-promoting role. A recent experiment has shown that macrophage-derived exosomes enhance glycolysis, mediating lung cancer cell resistance to cisplatin chemotherapy. Molecular pathway study shows that these exosomes have high levels of miRNA-3679-5p and decrease the expression of NEDD4L to promote the stability of c-Myc, leading to lung cancer growth, induction of glycolysis, and mediation of cisplatin resistance [151]. Therefore, exosomes can induce glycolysis in favor of cancer growth and induce drug resistance [152].
STAT3 signaling is an oncogenic pathway that inhibits apoptosis and cell cycle arrest and promotes growth and metastasis [153]. STAT3 induces EMT and promotes cancer invasion. Overexpression of STAT3 is associated with poor prognosis and triggers chemoresistance [154–158]. Exosomes alter the polarization of macrophages and transform them into cancer-associated macrophages. The exosomes are enriched in gp130 and induce STAT3 signaling via IL-6 upregulation [159]. STAT3-containing exosomes are able to promote ovarian cancer progression by inducing an imbalance between T cells and tumor-associated macrophages in favor of immunosuppression [160]. Cyclin D1, MMP-2, and MMP-9 are upregulated by STAT3-containing exosomes and promote proliferation and invasion of breast cancer cells [161]. Similarly, hypoxic conditions in the TME enhance the ability of colon cancer cells to self-proliferate by upregulating STAT3 expression [162]. Overall, several molecular signaling pathways are affected by exosomes, and understanding their interaction may pave the way for the development of novel therapeutics [163–165].
Apoptosis is an important signaling pathway regulated by exosomes in tumors. Inhibition of apoptosis may pave the way for tumor progression and resistance to therapy [166]. A recent experiment has shown that cancer-associated fibroblasts secrete exosomes containing miRNA-92a-3p, which act as a tumor-promoting factor and induce the Wnt/β-catenin axis, leading to inhibition of mitochondrial apoptosis and inducing resistance of colorectal cancer cells to 5-fluorouracil [167]. The ROS can induce apoptosis in cancer cells. It has been reported that modulation of the levels of ROS may be important for the response of cancer cells to therapy [168–171]. In pancreatic cancer, exosomes containing miRNA-155 reduce the expression of DCK, an enzyme involved in the metabolism of gemcitabine. This is followed by an increase in superoxide dismutase and catalase, leading to a reduction in ROS and subsequent growth of cancer cells and mediating their resistance to chemotherapy [172, 173]. Interestingly, not only can apoptosis in cancer cells reduce their proliferation, but apoptosis in immune cells can also affect cancer progression. A recent experiment has shown that pancreatic cancer-derived exosomes are taken up by lymphocytes to induce p38 MAPK signaling and mediate apoptosis triggered by endoplasmic reticulum stress apoptosis to stimulate immunosuppression and pave the way for cancer progression [174]. On the other hand, exosomes derived from colorectal cancer cells stimulate extracellular signal-regulated kinase (ERK) to suppress apoptosis and promote growth [175]. Apoptosis as a mechanism of programmed cell death is closely related to autophagy. In general, autophagy is involved in cell homeostasis by degrading aged and toxic organelles and macromolecules. However, the activation of autophagy in cancer cells is controversial and requires further clarification because it plays both tumor-promoting and tumor-suppressive roles [176–179]. A recent experiment has shown that sirtuin 2 (SIRT2) increases the mRNA stability of transcription factor EB (TFEB) and induces the release of exosomes to trigger autophagy and decrease apoptosis in non-small cell lung cancer cells [180]. Therefore, special attention should be paid to autophagy in cancer progression when studying apoptosis regulation by exosomes. Overall, studies support the fact that exosomes can either increase or decrease cancer cell proliferation [181–185].
Exosomes and tumor metastasis
Cancer cell invasion threatens the lives of many cancer patients around the world by enabling the spread of tumor cells to various organs and tissues of the body and mediating their malignancy [186–188]. Therefore, the factors involved in cancer metastasis should be highlighted to direct future experiments to target them [189–192]. Exosomes have been shown to be critical regulators of cancer metastasis. RelA and RelB are able to decrease the levels of MCAM and CD146 adhesion molecules in the release of EVs, leading to breast cancer metastasis. Silencing of RelA and RelB decreases the organotropic ability of exosomes in vivo and significantly reduces their ability to promote breast cancer migration and invasion [193]. It appears that exosomes containing Eph receptor A2 (EphA2) are able to transfer metastatic potential to pancreatic cancer cells and promote their invasion [194]. In contrast, there are exosomes capable of suppressing the cancer cell metastasis. For example, migration and invasion of non-small cell lung cancer cells were significantly decreased by miRNA-let7e-containing exosomes. LSD1 is upregulated in lung cancer and reduces E-cadherin levels to promote migration. Exosomes containing miRNA-let7e are able to increase CDH1 expression via LSD1 down-regulation to impair lung cancer metastasis [195]. Therefore, exosomes are important modulators of cancer migration and invasion.
The molecular mechanisms responsible for cancer migration and invasion are influenced by exosomes. The epithelial-to-mesenchymal transition (EMT) is among the best known mechanisms involved in cancer migration and invasion [196]. The decrease in E-cadherin, and the increase in N-cadherin and vimentin mediate EMT-induced metastasis in cancer cells [197, 198]. There are a number of factors known as EMT-inducing transcription factors (EMT-TFs), including ZEB1/2, TGF-β, Snail, Slug, and Twist, which can stimulate EMT in cancer cells and promote tumor invasion [199, 200]. A recent experiment has shown that exosomes containing the integrin alpha 2 subunit (ITAG2) are able to induce EMT and enhance prostate cancer cell metastasis [201]. On the other hand, exosomes containing miRNA-204 exhibit anti-tumor activity and reduce lung tumor cell invasion and migration by inhibiting EMT. To this end, exosomal miRNA-204 reduces the expression of KLF7 to inhibit the Akt/HIF-1α axis, resulting in a reduction of lung cancer migration and invasion by inhibiting EMT [202]. The ascites of ovarian cancer secretes exosomes containing miRNA-6780b-5p, which increase cancer migration and invasion in patients. It appears that exosomes containing miRNA-6780b-5p induce EMT to promote ovarian cancer metastasis [203]. The ability of exosomes to inhibit or induce EMT depends on their cargo. For example, miRNA-381-3p plays a tumor-suppressive role and exosomes containing this miRNA suppress EMT-mediated metastasis of breast cancer cells [204]. Therefore, there is increasing evidence for the role of exosomes in regulating metastasis by targeting the EMT mechanism [205, 206].
In addition to EMT, matrix metalloproteinases (MMPs) are also involved in increasing cancer metastasis [207–211], and recent experiments have confirmed this. MMP-2 induces EMT to increase squamous cell carcinoma metastasis, and it may act as an independent factor in patient prognosis [212]. A clinical experiment demonstrated overexpression of MMP-7 in bladder cancer, which is associated with unfavorable prognosis and shortened overall survival of patients [213]. Moreover, MMP-3 is involved in the induction of angiogenesis, which promotes cancer progression [214]. Therefore, suppression of MMP activity may be of interest to inhibit cancer metastasis. Overexpression of trefoil factor 3 (TFF3) leads to upregulation of MMP-2 and MMP-9, enhancing prostate cancer cell invasion. Mesenchymal stroma cell-derived exosomes containing miRNA-143 exhibit anti-tumor activity and inhibit TFF3 to downregulate MMP-2 and MMP-9, leading to suppression of metastasis [215]. In contrast, exosomes derived from renal cancer cells are able to increase the expression of MMP-9 to promote invasion [216]. Although some studies have focused on the interaction between exosomes and MMP, there is still a long way to go to uncover the signaling networks involved (Fig. 4).
Fig. 4.

Exosomes in the regulation of cancer cell growth and invasion. Glycolysis responsible for tumor growth is regulated by exosomes. CAFs are able to secrete exosomes to modulate tumor progression. EMT, metastasis, ROS and apoptosis are other signaling pathways affected by exosomes in tumor cells
Exosomes and cancer resistance
Drug resistance
In the field of cancer therapy, a variety of antitumor agents have been developed, including cisplatin, 5-fluorouracil (5-FU), sorafenib, and oxaliplatin [233]. However, long-term use of these chemotherapeutic agents leads to drug resistance and an unfavorable prognosis for cancer patients. A specific mechanism is responsible for chemoresistance. Among others, drug efflux, upregulation of anti-apoptotic factors, DNA damage repair, epigenetic changes, and the TME may influence drug resistance [234–239]. The current section focuses on the potential role of exosomes in drug resistance of cancer cells.
A growing body of evidence suggests that exosomes are capable of influencing the response of cancer cells to chemotherapy [240]. The ability of exosomes to transport cargoes has made them promising agents in cancer chemotherapy. As nanostructures, exosomes can mediate the co-delivery of a miRNA-21 inhibitor and 5-FU in colon tumor chemotherapy. The 5-FU and miRNA-21 inhibitor were loaded into exosomes via electroporation. Systematic administration of exosomes containing the miRNA-21 inhibitor and 5-FU suppressed tumor growth in mice. Exosomes administration enhances cellular uptake and reduces miRNA-21 expression in favor of colon cancer suppression. Moreover, miRNA-21 inhibitor and exosomes loaded with 5-FU induce cell cycle arrest and apoptosis. These anti-tumor activities are mediated via the upregulation of PTEN and hMSH2 as tumor suppressor factors in colon cancer [241]. The process of exosome secretion, cargo transport, and involvement in drug resistance are complex and should be elucidated. The epithelial ovarian cancer cells are able to recruit macrophages and stimulate their tumor-associated phenotype. Hypoxia in the TME leads to the secretion of exosomes from macrophages containing high levels of miRNA-223 as a tumor-promoting factor. The process of mediating drug resistance is that miRNA-223 delivered by exosomes reduces PTEN expression to induce PI3K/Akt signaling. To establish a link between hypoxia and exosome secretion, patients with ovarian cancer were studied. It was found that overexpression of HIF-1α, a hypoxia marker, occurs in ovarian cancer patients and is associated with upregulation of miRNA-223. Therefore, complicated molecular pathways and mechanisms are involved in the secretion of exosomes and the triggering of chemoresistance [242]. Another experiment demonstrates the potential role of macrophage-derived exosomes in triggering drug resistance in pancreatic cancer. An interesting point is that exosomes may be involved in the inactivation of chemotherapeutic agents in triggering drug resistance. Macrophage-derived exosomes contain miRNA-365 as a tumor-promoting factor and are able to induce gemcitabine resistance in pancreatic cancer. To this end, exosome-derived miRNA-365 stimulates the cytidine deaminase enzyme to inactivate gemcitabine, leading to chemoresistance in pancreatic cancer [243].
In addition to inactivating chemotherapeutic agents, exosomes can direct cancer cells toward cell death. It has been reported that exosomes can be obtained from CSCs in pancreatic cancer. These exosomes contain miRNA-210, which can induce gemcitabine resistance via inducing mTOR signaling. Moreover, these exosomes suppress gemcitabine-mediated apoptosis and cell cycle arrest [244]. Consequently, various signaling networks are affected by exosomes in triggering chemoresistance. In addition, the accumulation of chemotherapeutic agents in tumor cells is impaired. Exosomes are able to induce efflux of cisplatin from ovarian cancer cells under hypoxic conditions, demonstrating that they can prevent internalization of chemotherapeutic agents. Furthermore, STAT3 plays an important role in this case. Overexpression of STAT3 in hypoxic condition is crucial for exosome release and triggering cisplatin resistance in ovarian cancer. Suppression of STAT3 signaling alters the levels of Rab7 and Rab27a proteins, preventing the secretion of exosomes [245].
Tumor cells exhibiting features of drug resistance are able to secrete exosomes that accelerates chemoresistance. Such a strategy has been studied in lung cancer. Exosomes derived from cisplatin-resistant lung cancer cells have high levels of miRNA-100-5p, which decrease the expression of mTOR, leading to cisplatin resistance [246]. In addition, exosomes may act as a means of communication between normal and cancer cells in inducing drug resistance. Endothelial cells are able to secrete exosomes with a particle size of 40–100 nm, which trigger EMT-mediated metastasis in nasopharyngeal carcinomas and mediate their resistance to chemotherapy [247]. Exosomes can be used to suppress chemoresistance. In one experiment, exosomes were used to deliver anti-miRNA-214 to gastric cancer cells to induce apoptosis and decrease proliferation and invasion, leading to drug sensitivity [248]. Overall, the studies are consistent with the fact that exosomes can affect the growth and invasion of cancer cells to influence their response to chemotherapy. They contain various cargoes and can modulate molecular signaling pathways in favor of chemoresistance or chemosensitivity. Such exosomes and associated signaling networks should be elucidated to prevent chemoresistance in cancer cells [249–259]. Table 2 provides an overview of exosomes and their association with drug resistance in cancer. Figure 5 shows a schematic representation of exosomes in regulating drug action.
Table 2.
Exosomes and their function in mediating drug resistance/sensitivity in cancer
| Cancer type | Chemotherapeutic agent | Signaling network | Remark | Refs |
|---|---|---|---|---|
| Breast cancer | Adriamycin | – | Drug resistance is induced by the transfer of P-gp and UCH-L1 proteins through exosomes into the extracellular microenvironment | [260] |
| Breast cancer | Anthracycline and taxane agents | – | Chemoresistance is observed in breast cancer patients who had high levels of GSTP1-containing exosomes | [261] |
| Breast cancer | Adriamycin |
MDR1 P-glycoprotein |
Drug resistance is induced by exosomes by enhancing the expression of MDR1 and P-glycoprotein Chemoresistance is inhibited by suppression of exosome formation and secretion by psoralen |
[262] |
| Breast cancer | Gemcitabine |
Autophagy EMT/HIF-α |
Autophagy is inhibited by exosomes containing siMTA1 EMT is suppressed Tumor growth in vitro and in vivo is retarded |
[263] |
| Liver cancer | Sorafenib | – |
Selectivity of exosomes against cancer cells is increased by modifying the surface of exosomes Drug resistance is suppressed by synergistic cancer chemotherapy with sgIQ 1.1 plasmid-loaded exosomes |
[263] |
| Leukemia | Etoposide |
Bax Bcl-2 PARP Caspase-3 |
Drug resistance is induced by exosomes derived from bone mesenchymal stem cells by increasing the expression of Bcl-2 and decreasing the expression of Bax, caspase-3, and PARP | [264] |
| Leukemia | Imatinib |
Bax Bcl-2 Caspase-3 Caspase-9 |
Apoptosis is prevented by exosomes derived from mesenchymal stromal cells, and leukemia cell survival is increased The expression of Bax, caspase-3 and caspase-9 is downregulated, and the expression of Bcl-2 is increased |
[265] |
| Leukemia | Imatinib | miRNA-328/ABCG2 | Drug sensitivity is increased by decreasing ABCG2 expression through miRNA-328 in exosomes | [266] |
| Glioblastoma | Temozolomide | PD-L1/AMPK/ULK1/autophagy | Autophagy is induced by the exosomes containing PD-L1 through stimulation of the AMPK/ULK1 axis, which mediates drug resistance | [267] |
| Glioblastoma | Temozolomide | STAT3/miRNA-21/PDCD4 |
STAT3 is downregulated by a combination of temozolomide and pacritinib miRNA-21 expression is reduced to upregulate the PDCD4 tumor suppressor M2 polarization of macrophages is inhibited Glioblastoma tumorigenesis is prevented |
[268] |
| Non-small cell lung cancer | Cisplatin | miRNA-146a-5p | Low levels of miRNA-146a-5p are observed in cisplatin-resistant A549 cells and can be used to predict cancer recurrence | [269] |
| Oral cancer | Cisplatin | miRNA-155/FOXO3a |
FOXO3a expression is enhanced by exosomes containing the miRNA-155-inhibitor Mesenchymal-to-epithelial transition is triggered to suppress cancer cell migration and invasion |
[270] |
| Hepatocellular carcinoma | Oxaliplatin |
miRNA-214/P-gp miRNA-214/SF3B3 |
P-gp and SF3B3 expression is decreased by exosomal miRNA-214 Drug sensitivity is increased |
[271] |
| Hepatocellular carcinoma | Cisplatin | miRNA-199a-3p | Drug resistance is suppressed by apoptosis induction through the increased expression of miRNA-199a-3p delivered by exosomes | [272] |
| Prostate cancer | Docetaxel | CD44v8-10 mRNA | Drug resistance is mediated by the presence of CD44v8-10-containing exosomes in the serum of prostate cancer patients | [273] |
Fig. 5.

The role of exosomes in modulating the response to drug therapy. Most experiments focused on exosomal miRNAs and their downstream targets such as PTEN and JAK2. PTEN suppresses cancer progression, while JAK2 promotes cancer malignancy. Depending on the function of each molecular mechanism, the role of exosomes in cancer progression or inhibition varies
Radio-resistance
Radiotherapy is another cancer treatment option that uses radiation to inhibit cancer progression and induce cell death [274]. However, due to specific conditions in the TME such as hypoxia, cancer cells could develop resistance to radiotherapy, and the factors involved in this phenomenon should be elucidated [275, 276].
Most experiments have focused on the relationship between exosomes and drug resistance. However, there are also a few studies examining the role of exosomes in radioresistance. For example, a recent experiment has shown that cancer-associated fibroblasts are able to secrete exosomes to promote stemness of colorectal tumors and trigger their clonogenicity and radioresistance. Mechanistically, these exosomes induce transforming growth factor-beta (TGF-β) to mediate radioresistance. When this signaling pathway is suppressed using antibodies, colorectal tumor progression is impaired and sensitivity to radiotherapy is increased [277]. In contrast, there are exosomes capable of suppressing radioresistance. Exosomes containing miRNA-34c suppress proliferation, invasion, and EMT in nasopharyngeal carcinomas. In addition, miRNA-34c-loaded exosomes induce apoptosis and mediate radiosensitivity. Molecular pathway study shows that miRNA-34c-loaded exosomes suppress the β-catenin signaling pathway, thereby increasing the sensitivity of nasopharyngeal carcinoma cells to radiotherapy [278]. In the previous section, it was shown that chemotherapy of cancer cells induces the secretion of exosomes. Moreover, chemoresistant cancer cells are capable of secreting exosomes, which favors their progression and promotes drug resistance [279, 280]. A recent experiment has shown that exosomes can be obtained from irradiated gastric cancer cells [281]. However, further studies are needed to determine whether exosomes are involved in the development of radioresistance.
Immune evasion and inflammation
Although few experiments have investigated the role of exosomes in immune resistance and evasion, these studies show that exosomes are promising candidates in this case because of their modulatory effect on immune cells. The T-regulatory cells (Treg cells) are well known because of their immunosuppressive effects. In breast cancer, the CD73 + Treg cells are able to facilitate immune evasion by producing adenosine. The exosomes containing the lncRNA SNHG16 increase the expression level of CD73 on Treg cells. To this end, exosomal SNHG16 decreases the expression of miRNA-16-5p via sponging to induce the TGF-β/SMAD5 axis, resulting in overexpression of CD73 on Treg cells. Therefore, the ability of exosomes to transmit SNHG16 may mediate overexpression of CD73 on Treg cells and lead to immunosuppression in breast cancer [282]. Programmed death-1 (PD-1) is a molecular pathway that causes T cell exhaustion and prevents their proliferation. Moreover, PD-1 induces apoptosis in cytotoxic T cells and inhibits their anti-tumor activity to mediate immune evasion. Binding of PD-L1 to PD-1 triggers this pathway [283]. A recent experiment has shown that exosomes derived from cancer-associated fibroblasts contain high levels of miRNA-92 as a tumor-promoting factor. Exosomal miRNA-92 mediates the interaction between LATS2 and YAP1 in breast cancer cells. Subsequently, YAP1 translocates to the nucleus and binds to the PD-L1 promoter to enhance its expression, leading to the apoptosis of T cells and a decrease in proliferation of these cytotoxic cells [284]. Exosomes can not only evade immune defences but also influence immune cells to promote cancer progression. Indeed, interactions between exosomes and immune cells can create optimal conditions for increased cancer growth and invasion. NF-κB signaling is related to the inflammatory process and may promote cancer progression. NF-κB expression in cancer is regulated by other molecular signaling pathways, of which miRNAs are the best known [285]. On the other hand, there is growing evidence that chronic inflammation and pro-inflammatory cytokines promote cancer progression [286–288]. A recent experiment has shown that exosomes derived from breast cancer cells have high levels of miRNA-183-5p and are able to decrease the expression of PPP2CA. Decreased expression of PPP2CA paves the way for triggering NF-κB signaling and mediating chronic inflammation. In addition, this signaling network increases the levels of pro-inflammatory cytokines such as IL-1β, IL-6 and TNF-α. Therefore, the transmission of miRNA-183-5p by exosomes and its effect on inflammation may promote the proliferation and invasion of breast cancer cells [289].
TGF-β mediates immune evasion of breast cancer cells. To this end, TGF-β increases the levels of PD-L1 in exosomes and stimulates the dysfunction of cytotoxic CD8 T cells [290]. Another experiment shows that exosomes derived from multiple myeloma suppress apoptosis and increase the growth rate of Treg cells, triggering immune dysfunction [291]. In addition, exosomes are able to promote the progression of gastric cancer by suppressing the maturation of dendritic cells [292]. Exosomes containing indoleamine 2,3-dioxygenase (IDO) may induce T-cell dysfunction via triggering the IL-6/TNF-α axis [293]. Future experiments may focus on targeting exosomes in preventing immune evasion and suppressing inflammation to impair cancer progression [294, 295].
Exosomal miRNAs
miRNAs are endogenous, short noncoding RNAs with a length of 19–24 nucleotides that can regulate gene expression at the posttranscriptional level by binding to the 3’-untranslated region (3’-UTR) of target mRNAs [296, 297]. Recent experiments have shed light on the role of miRNAs in cancer. For example, hypoxic conditions enhance lung cancer progression by decreasing the expression of miRNA-495 and miRNA-5688 and subsequently increasing IL-11 levels [298]. Moreover, decreased expression of miRNA-100 and miRNA-125b paves the way for overexpression of IGF2 and subsequent cancer stem cell features in hepatocellular carcinoma [299]. Further studies have shown that miRNAs can be considered as reliable biomarkers for cancer diagnosis [300, 301]. Because exosomes are capable of transmitting miRNAs, we dedicated this section to the study of exosomal miRNAs in the regulation of cancer progression.
In the previous sections, the role of exosomes in cancer progression has been clearly demonstrated as they affect the TME and the therapeutic response of cancer cells. It has been discussed that exosomes may contain various genes that influence cancer progression. The current section focuses specifically on exosomal miRNAs and how they may modulate cancer progression. In a recent experiment, exosomes were isolated exosomes by centrifugation from colorectal cancer cells infected with Fusobacterium nucleatum and transferred to uninfected cancer cells. The exosomes were found to contain high levels of miRNA-1264, miRNA-92b-3p, and miRNA-27a-3p, which are able to enhance metastasis and tumor stage of colorectal cancer [302]. There is increasing evidence that hypoxic conditions in the TME significantly promote carcinogenesis in gastric cancer [303, 304]. It appears that hypoxia induces the secretion of exosomes from gastric cancer cells. These exosomes promote both growth and migration of gastric cancer. These exosomes contain miRNA-301a-3p, which acts as a tumor-promoting factor and increases the stability of HIF-1α and inhibits its degradation by targeting PDH3 and hydroxylating HIF-1α subunits. Moreover, there is a positive feedback loop between HIF-1α and miRNA-301a-3p in enhancing the proliferation and invasion of gastric cancer cells. Clinical investigation shows that exosomal miRNA-301a-3p is upregulated in gastric cancer patients and mediates peritoneal metastasis [305].
The PI3K/Akt axis is a trigger of cancer progression and its induction promotes cancer cell proliferation and invasion [306–308]. In addition, the PI3K/Akt axis enhances the aggressive behavior of cancer and is associated with drug resistance. PTEN is the negative regulator of the PI3K/Akt axis and increasing its expression is a promising strategy to interrupt cancer progression [309–311]. A recent experiment shows that exosomes containing miRNA-22-3p have a tumor suppressive effect and prevent colorectal cancer progression by downregulating PI3K/Akt [312]. In addition to increased proliferation, exosomes may facilitate the transfer of apolipoprotein E between cells to induce the PI3K/Akt axis, which mediates cytoskeletal remodeling and promotes gastric cancer cell migration and invasion [313]. Therefore, the PI3K/Akt axis is strongly regulated by exosomes [314]. The question now arises: is there a link between exosomes and PTEN as upstream mediators of the PI3K/Akt axis? The answer is positive, and this potential has been confirmed in several experiments. In non-small cell lung cancer, exosomes containing miRNA-126 are able to enhance PTEN expression in suppressing the PI3K/Akt axis and impair metastasis. The in vivo experiment showed that miRNA-126 reduced lung cancer metastasis by modulating the PTEN/PI3K/Akt axis [315]. On the other hand, hypoxia leads to the secretion of exosomes from colorectal cancer cells. The exosomes contain high levels of miRNA-410-3p, which decrease PTEN expression to induce the PI3K/Akt axis and promote cancer cell invasion. Moreover, exosomal miRNA-410-3p is associated with unfavorable prognosis of colorectal cancer patients [316]. The growth rate of esophageal cancer cells is significantly increased by the transfer of miRNA-93-5p through exosomes and the downregulation of PTEN [317].
Another molecular signaling pathway involved in cancer progression is the Wnt/β-catenin axis. Nuclear translocation of β-catenin promotes cancer growth and invasion [318–320]. Activation of Wnt signaling is associated with poor prognosis. In addition, the Wnt/β-catenin axis can mediate features of drug resistance in cancer cells [321–325]. Exosomes are able to induce Wnt5b signaling, thereby increasing the progression of lung cancer cells [326]. The exosomes containing miRNA-320a act as tumor-promoting factors and reduce the expression of SOX4. As a result, Wnt/β-catenin activation occurs, which significantly promotes the growth and metastasis of lung cancer cells [327]. Another experiment shows that exosomes from breast cancer cells contain high levels of miRNA-146a, which reduces the expression of TXNIP to induce the Wnt/β-catenin axis, leading to activation of cancer-associated fibroblasts in the TME and promoting breast cancer progression [328]. Therefore, regulation of Wnt signaling by exosomes modulates cancer progression [329].
One of the increasing challenges in breast cancer is bone metastasis, which is associated with pain, decreased overall patient survival, and an unfavorable prognosis. Therefore, efforts have been made to uncover the role of exosomal miRNAs in bone metastasis of breast cancer cells in order to target them in future experiments. A clinical study has shown that serum exosomes containing miRNA-21 promote bone metastasis in breast cancer patients [330]. Although these studies demonstrate the tumor-promoting role of exosomal miRNAs, there are also experiments showing that exosomal miRNAs can suppress cancer progression. In pancreatic cancer cells, exosomal miRNA-34a can effectively enter the cell membrane and decrease the expression of Bcl-2 to induce apoptosis and reduce growth and viability. The in vivo experiment on nude mouse xenografts also demonstrated the role of exosomal miRNA-34a in retarding tumor growth [331]. In addition to apoptotic factors, other signaling networks responsible for cancer progression may also be influenced by exosomal miRNAs. It is suggested that exosomal miRNA-210 is a tumor-promoting factor in lung cancer. Secretion of exosomal miRNA-210 by cancer-associated fibroblasts significantly promotes lung cancer migration and invasion. Molecular pathway study reveals that exosomal miRNA-210 induces the PI3K/Akt axis via PTEN down-regulation to induce EMT and enhance lung cancer cell metastasis [332]. UbiA prenyltransferase domain-containing protein 1 (UBIAD1) is downregulated by exosomal miRNA-4644 via binding to its 3’-UTR to enhance bladder cancer cell invasion [333]. Thus, exosomal miRNAs influence a variety of molecular pathways in regulating cancer progression [334–337].
Exosomal long noncoding RNAs
Recently, lncRNAs have attracted considerable attention because of their potential role in modulating of various molecular signaling pathways in cancer therapy [338–341]. Briefly, lncRNAs are RNA molecules longer than 200 nucleotides and their function differs depending on their localization in the nucleus or cytoplasm [342, 343]. There are five types of lncRNAs and they are able to affect proteins and genes under physiological and pathological conditions [93, 344–347]. The lncRNA DILA1 functions as a tumor-promoting factor and increases the stability of cyclin D1 to promote breast cancer progression and mediate resistance to tamoxifen [348]. Upregulation of lncRNA ENO1-IT1 by the gut microbiota mediates glycolysis and increases the proliferation rate of colorectal cancer cells [349]. Similar to miRNAs, lncRNAs may function as diagnostic and prognostic tools in cancer [350].
Similar to miRNAs, lncRNAs can also be transferred between cells via exosomes. Depending on the function of lncRNAs, they can reduce or promote cancer progression. The lncRNA H19 is considered a tumor-promoting factor because its upregulation induces drug resistance and promote both proliferation and invasion of cancer cells [351]. Exosomes transfer lncRNA H19 to non-small cell lung cancer cells to inhibit apoptosis and induce resistance to gefitinib chemotherapy [352]. Cancer cell migration is also regulated by exosomal lncRNAs. The lncRNA linc-ROR can be transferred into the TME to promote distant metastasis through EMT induction [353]. The exosomal lncRNAs are able to modulate the expression level of miRNAs to target other molecular pathways. The exosomal lncRNA CASC15 is overexpressed in osteosarcomas and increases growth and metastasis. Silencing of CASC15 impairs progression of osteosarcoma cells. Further studies show that exosomal CASC15 decreased the expression of miRNA-338-3p by sponging and increases the expression of RAB14 in osteosarcomas [354]. Delivery of lncRNAs through exosomes is a challenge for the treatment of some kinds of tumor types, particularly brain tumors. The blood–brain barrier (BBB) is an obstacle that prevents antitumor drugs from entering the brain and limits our ability to target brain tumors [355]. However, exosomes are able to disrupt BBB when transporting lncRNAs into the brain. A recent experiment has shown that exosomes are capable of crossing the BBB and transport the lncRNA GS1-6000G8.5 into the brain and mediate metastasis of breast cancer cells to the brain [356].
Due to the interaction between lncRNAs and miRNAs, downregulation of tumor-promoting lncRNAs may pave the way for upregulation of miRNAs with anti-tumor activity. It has been reported that downregulation of the exosomal lncRNA SBF2-AS1 in polarized M2 macrophages leads to the expression of miRNA-122-5p, a tumor suppressor factor. Subsequently, upregulated miRNA-122-5p suppresses pancreatic cancer progression via inhibition of XIAP [357]. As for the ability of exosomal lncRNAs to regulate apoptosis pathways, they can modulate the therapeutic response of cancer cells. The exosomal lncRNA UCA1 shows overexpression in breast cancer cells (MCF-7) and suppresses apoptosis via downregulation of caspase-3 to mediate tamoxifen resistance [358]. In addition to chemotherapy, exosomal lncRNAs regulate the response of cancer cells to radiotherapy. Because of the tumor-promoting role of the lncRNA HOTAIR, its transfer to laryngeal cancer cells via exosomes induces the expression of E2F2 via downregulation of miRNA-454-3p. This accelerates the progression of laryngeal cancer and reduces their sensitivity to radiotherapy [359]. Therefore, the identification of exosomal lncRNAs may increase our understanding of the factors involved in cancer progression and develop novel therapeutics in the near future [360–363].
Exosomal circular RNAs
CircRNAs are another subset of noncoding RNAs that have a covalently closed loop structure and exhibit vital functions under physiological and pathological conditions [364–367]. Aberrant expression of circRNAs is observed in various cancers. The hsa-circRNA-000166 increases the progression of colorectal cancer by downregulating miRNA-326 and subsequently overexpressing LASP1 [368]. Downregulation of miRNA-665 by circ-100876 occurs in gastric cancer, which triggers EMT via upregulation of YAP1 [369]. Experiments have shown that circRNAs affect cancer growth and metastasis mainly by regulating the expression of miRNAs [369, 370].
The circRNA IARS (circ-IARS) is thought to promote cancer metastasis. This circRNA is located in exosomes and enters HUVECs to increase cancer metastasis. Exosomal circ-IARS decreases overall survival and increases metastasis and TNM stage. Mechanistically, exosomal circ-IARS decreases the levels of miRNA-122 and ZO-1, whereas it increases the levels of RhoA and RhoA-GTP and increases the permeability of endothelial monolayers. Moreover, exosomal circ-IARS enhances F-actin expression and focal adhesion to promote invasion and metastasis [371]. The Wnt signaling pathway is related to cancer proliferation and metastasis. In the context of Wnt pathway, β-catenin translocates to the nucleus to promote cancer progression [372]. The exosomal circ-ABCC1 is overexpressed in colorectal cancer and promotes stemness and invasion. Mechanistically, circ-ABCC1 induces β-catenin to enhance colorectal cancer progression [373]. Similar to lncRNAs, exosomal circRNAs can regulate the response of cancer cells to chemotherapy. Exosomal circ-0002130 shows overexpression in lung cancer and mediates osimertinib resistance. To this end, exosomal circ-0002130 reduces miRNA-498 expression via sponging to enhance GLUT1, HK2, and LDHA expression, leading to lung cancer progression and drug resistance [374]. Another experiment shows how exosomal circRNAs can regulate drug sensitivity via affecting HK2. A recent experiment has shown that exosomal circ-0008928 can increase lung cancer progression and glycolysis. Indeed, exosomal circ-0008928 increases the proliferation rate of lung cancer cells via inducing glycolysis and then, decreases their sensitivity to cisplatin. Molecular pathway study shows that exosomal circ-0008928 enhances HK2 expression in triggering glycolysis and mediating drug resistance in lung cancer [375].
Exosomal circRNAs can also be considered as diagnostic and prognostic tools. For example, the expression levels of circ_0047921, circ_0056285, and circ_0007761 can be used to diagnose non-small cell lung cancer in Chinese. In addition, circ-0056285 is positively associated with the clinical stage and may increase lymph node metastasis [376]. The potential of exosomes as diagnostic and prognostic tools will be specifically discussed in the next sections. However, exosomal circRNAs can be used independently in this case [377–380]. All in all, exosomal ncRNAs regulate proliferation, invasion, immune response and drug sensitivity of cancer cells and can be considered as diagnostic and prognostic factors in cancer (Fig. 6, Table 3) [381–389].
Fig. 6.

The exosomal ncRNAs in modulating cancer progression. A variety of signaling networks are influenced by exosomal ncRNAs. Metastasis, growth, apoptosis and response to therapy are strongly modulated by exosomal ncRNAs. Further experiments are needed to identify other exosomal circRNAs, as studies have focused more on exosomal miRNAs and lncRNAs
Table 3.
The exosomal ncRNAs in cancer cells
| Exosomal ncRNA | Signaling network | Cancer type | Remarks | Refs |
|---|---|---|---|---|
| miRNA-34a | – | Breast cancer | Proliferation of cancer cells is suppressed | [390] |
| miRNA-145 |
MMP-9 TP53 |
Breast cancer |
Apoptosis is induced Metastasis is inhibited |
[391] |
| miRNA-21-5p | ZNF367 | Breast cancer | Cancer cell invasion is suppressed by downregulation of ZNF367 | [392] |
| miRNA-5100 | CXC12/CXCR4/EMT | Breast cancer |
CXC12/CXCR4 axis is suppressed by miRNA-5100, which acts as a tumor suppressor EMT is inhibited, and cancer cell invasion and migration are decreased |
[393] |
| miRNA-3613-3p | SOCS2 | Breast cancer |
Cancer cell proliferation and metastasis are enhanced SOCS2 is downregulated |
[394] |
| miRNA-423-5p | – | Breast cancer | The sensitivity of breast cancer cells to cisplatin is reduced | [395] |
| miRNA-19b-3p | PTEN/EMT | Esophageal cancer |
miRNA-19b-3p is upregulated EMT is induced by exosomal miRNA-19b-3p by downregulating PTEN Apoptosis is inhibited Growth and metastasis of cancer cells are enhanced |
[396] |
| miRNA-124 | EZH2 | Pancreatic cancer |
Exosomal miRNA-124 is downregulated Apoptosis is induced, EMT is inhibited and cancer cell migration is decreased by miRNA-124 EZH2 is downregulated by miRNA-124 |
[397] |
|
miRNA-21-5p miRNA-155-5p |
BRG1 | Colon cancer |
miRNA-21-5p and miRNA-155-5p are transferred from exosomes BRG1 expression is reduced M2 polarization of cancer cells is induced Cancer metastasis is enhanced |
[398] |
| miRNA-34c-3p | Integrin α2β1 | Non-small cell lung cancer | Metastasis and invasion of A549 cells are promoted by increased expression of integrins | [399] |
| miRNA-7 | YAP | Lung cancer | YAP expression is inhibited and cancer cell sensitivity to gefitinib is increased | [400] |
| miRNA-126a | – | Lung cancer |
Secretion of exosomes by lung cancer cells is induced by exposure to doxorubicin Cancer cell migration and invasion are increased by exosomal miRNA-126a |
[401] |
| miRNA-122 | – | Hepatocellular carcinoma | Sensitivity of cancer cells to chemotherapy is increased by exosomal miRNA-122 | [402] |
| miRNA-302b |
ERK1/2 MMP-9 TGFβRII |
Lung cancer |
Cancer cell growth and invasion are inhibited ERK1/2, MMP-9, and TGFβRII are downregulated |
[403] |
| miRNA-21 | PDCD4 | Lung cancer |
Lung cancer proliferation is increased Anti-tumor immunity is suppressed by the proliferation of myeloid-derived suppressor cells PDCD4 is downregulated |
[404] |
| miRNA-375 | ENAH | Esophageal cancer | Cancer progression is suppressed by decreasing the expression of ENAH | [405] |
|
miRNA-146b miRNA-222 |
– | Papillary thyroid cancer | Proliferation of cancer cells is increased | [406] |
| miRNA-200b | KLF6 | Ovarian cancer |
KLF6 is downregulated by miRNA-200b M2 polarization of macrophages is induced |
[407] |
| miRNA-92b-3p | SOX4 | Ovarian cancer |
Cancer progression is suppressed by inhibiting angiogenesis SOX4 is downregulated |
[408] |
| miRNA-224-5p | – | Renal cancer | Invasion and growth of cancer cells are suppressed | [407] |
| miRNA-1228 | MMP-14 | Gastric cancer | Cancer progression is suppressed by downregulation of MMP-14 | [408] |
| lncRNA ZFAS1 | Gastric cancer | – |
Association with lymph node metastasis and TNM stage is observed EMT is induced Apoptosis is inhibited |
[409] |
| lncRNA KCNQ1OT1 | Colorectal cancer | miRNA-30a-5p/USP22/PD-L1 |
Immune evasion is induced CD8 + T cell response is suppressed Expression of miRNA-30a-5p is decreased by acting as ceRNA USPP22 expression is upregulated to prevent PD-L1 ubiquitination PD-L1 expression is enhanced |
[410] |
| lncRNA HOTAIR | Breast cancer | ErB2 |
A positive association is observed between HOTAIR and ErB2 HOTAIR expression is increased by ErB2 in a MAPK-dependent manner |
[411] |
| LINC01133 | Bladder cancer | Wnt |
Low levels of LINC01133 in exosomes from bladder cancer cells are observed Wnt signaling is suppressed to impair cancer cell growth and metastasis |
[412] |
| Circ-ABCC1 | Colorectal cancer | Wnt/β-catenin | Cancer cell progression is enhanced by circ-ABCC1 via induction of β-catenin signaling | [376] |
| Circ-0002130 | Non-small cell lung cancer | miRNA-498/HK2-GLUT1-LDHA |
Cancer cell proliferation and invasion are increased in vitro and in vivo Osimertinib resistance is observed miRNA-498 is downregulated via sponging Expression of HK2, GLUT1, and LDHA is increased |
[374] |
| Circ-0008928 | Non-small cell lung cancer | miRNA-488/HK2 |
Glycolysis, proliferation and cisplatin resistance of cancer cells are induced Expression of miRNA-488 is decreased to induce HK2 expression |
(375) |
Exosomes as carrier systems
Anti-tumor agents
The previous sections have obviously shown that exosomes can affect cancer progression in several ways and are able to modulate the TME. These effects are based on exosome cargo. In this section, we discuss how exosomes can be used to deliver anti-tumor agents in cancer therapy. Remarkably, exosomes can deliver both synthetic and natural agents. In a recent experiment, exosomes with triptolide were used in the treatment of ovarian cancer. The exosomes showed high encapsulation efficiency and were able to slow tumor growth in vivo. Triptolide-loaded exosomes induce apoptosis in ovarian cancer cells and suppress their proliferation and viability [413]. Paclitaxel (PTX) is an anticancer agent that arrests the cell cycle by disrupting microtubule polymerization. Some cancer cells have developed resistance to PTX chemotherapy. Various techniques including nanoscale delivery systems have been developed to suppress chemoresistance. In one study, exosomes were used as delivery vehicles for PTX in lung cancer therapy. Exosomes were derived from macrophages and then modified with aminoethylanisamide-polyethylene glycol (AA-PEG) to selectively target sigma receptors that are upregulated on the surface of lung cancer cells. These exosomes are preferentially internalized into lung cancer cells and release PTX to suppress lung cancer cell progression [414]. One of the advantages of exosomes is their biocompatibility. In addition, they can deliver drugs as well as act as and imaging agents, which is referred to as theranostics. In a recent experiment, exosomes were isolated from cancer cells (e.g., HeLa cells) and then loaded with doxorubicin. In addition, silver nanoclusters were loaded into doxorubicin-coated exosomes. These exosomes enable imaging while delivering doxorubicin to suppress cancer progression, while exhibiting high biocompatibility and safety profile [415]. In the same study, exosomes were also used to deliver geldanamycin as an HSP90 inhibitor to affect the growth rate of cancer cells [416]. Drug-loaded exosomes can also regulate the TME in favor of cancer therapy. It has been reported that exosomes derived from M1-polarized macrophages can be loaded with PTX. PTX-loaded exosomes induced a pro-inflammatory environment and enhanced inflammation, which promoted the upregulation of caspase-3 expression, triggered apoptosis, and the subsequent enhancement of the anti-tumor activity of PTX [417]. Overall, these studies suggest that exosomes are promising candidates for drug delivery. Further experiments should be performed to elucidate their role in drug delivery, their encapsulation efficiency, and how their surface can be modified to increase their selectivity toward cancer cells [418–421].
Genetic tools
Small interfering RNAs (siRNAs) are double-stranded RNA molecules of up to 25 nucleotides in length. They are produced from mRNA and lncRNAs via the function of the RNase III enzyme Dicer [422]. The actual function of siRNA is achieved it is incorporated into the RNA-induced silencing complex (RISC) to direct the RNAi machinery to target mRNA for degradation after complementary sequences are found. Recently, siRNA has paved the way to treat various diseases in preclinical and clinical research, such as viral infections, neurological disorders, ocular diseases, autoimmune diseases, and cancer [421, 423]. Although siRNA has shown great capacity in suppressing gene expression and subsequently treating disease, naked siRNA appears to require modification in alleviating disease. Degradation of siRNA by RNase enzymes, tumor barriers, and off-targeting are drawbacks of siRNA that can be solved using delivery systems [424, 425]. Another genetic tool used in cancer therapy is the CRISPR/Cas system. The CRISPR/Cas9 system is the best known type of CRISPR system that has recently been used in the treatment of diseases [426]. The CRISPR/Cas9 system was discovered in prokaryotes and its main function is adaptive immunity [427]. The CRISPR/Cas9 system consists of three main components, including Cas9, sgRNA, and tracrRNA. The specificity, efficiency, and accuracy of the CRISPR/Cas9 system are provided by sgRNA. Cas9 acts as a scissor and is responsible for the destruction of double-stranded DNA [428]. Various experiments have been conducted on the use of CRISPR/Cas9 system in cancer therapy. CRISPR/Cas9 is able to target fusion oncogenes or transcription factors to suppress cancer progression and reduce growth and mortality [429]. Downregulation of ZEB1 and ZEB2 by the CRISPR/Cas9 system significantly reduces lung cancer cell migration and invasion [430]. The present section addresses the role of exosomes in the delivery of siRNA and the CRISPR/Cas system in cancer therapy.
siRNA
In one experiment, exosomes were isolated from embryonic kidney cells (HEK-293 cells) by ultracentrifugation and loaded with siRNA. The resulting exosomes had a diameter of 107 nm and an encapsulation efficiency of 10–20%. The exosomes efficiently transported siRNA to PANC-1 cells, demonstrating their potential as a transport system [431]. Induction of apoptosis is an ideal strategy to suppress cancer proliferation. To this end, Bcl-2-siRNA was loaded into exosomes and its anti-tumor activity against digestive system tumors was evaluated. Bcl-2-siRNA-loaded exosomes penetrated the cell membrane and delivered siRNA, resulting in apoptosis induction and reduced tumor growth. The anti-tumor activity of these exosomes was confirmed in both in vitro and in vivo experiments [432].
Hepatocyte growth factor (HGF) was first identified in mouse liver and is considered a cytokine with physiological functions in cell proliferation, survival, and migration [433, 434]. Recent experiments have revealed the tumor-promoting role of HGF in cancer. Overexpression of HGF enhances the growth and invasion of cervical cancer cells via affecting c-Met [435]. By inducing the c-Met/PI3K/Akt axis, HGF induces EMT and mediates drug resistance in pancreatic cancer [436]. HGF-siRNA-loaded exosomes may serve as nanostructures for cargo transport in gastric cancer treatment. These exosomes effectively transport siRNA to gastric cancer cells, leading to a significant reduction in their growth and migration and inhibition of angiogenesis [437]. Polo-like kinase 1 (PLK1) is another tumor-promoting factor in cancer. Overexpression of PLK1 inhibits autophagic cell death in prostate cancer [438]. In addition, Silencing of PLK1 suppresses breast cancer cell migration and invasion and promotes their sensitivity to drugs [439]. PLK1-siRNA was introduced into exosomes by electroporation, and exposure of bladder cancer cells to these exosomes resulted in a significant decrease in PLK1 mRNA levels and subsequent cancer eradication [440].
Efforts are underway to engineer exosomes to increase their selectivity toward cancer cells. One of the promising methods is to modify exosomes with ligands. The exosomes carrying DARPin G3 on their surface can bind to HER2/Neu on breast cancer cells. These targeted exosomes deliver TPD52-siRNA and reduce the expression of HER2/Neu by up to 70% [441]. Future experiments may therefore focus on the development of engineered and surface-modified exosomes for cancer therapy. Another ligand that can be used for surface modification of exosomes is the tLyp-1 peptide with the amino acid sequence CGNKRTR. The tLyp-1 peptide is able to bind to receptors such as neuropilin-1 (NRP1) and neuropilin-2 (NRP2), which are overexpressed on the surface of lung cancer cells [442–444]. Recently, tLyp-1 exosomes were used for siRNA delivery. These nanostructures were 100 nm in diameter and, thanks due to their selectivity toward lung cancer cells, they efficiently delivered siRNA and significantly reduced cancer stemness [445].
The factors involved in cancer metabolism and growth can be targeted by siRNA to increase drug sensitivity. Recently, CPT1A-siRNA was loaded into exosomes and surface modification with iRGD was performed to promote their selectivity toward colon cancer cells. These exosomes increased the sensitivity of colon cancer cells to oxaliplatin by downregulating CPT1A. Simultaneous administration of these exosomes and oxaliplatin induced apoptosis in colon cancer cells and suppressed their proliferation [446]. In another experiment, exosomes were modified with an epidermal growth factor receptor (EGFR) aptamer and then loaded with survivin-siRNA. Since survivin is involved in cancer progression and functions as an anti-apoptotic factor, its downregulation by exosomes sensitizes lung cancer cells to apoptosis. Exosomes provide endosomal escape of siRNA, which is important for increasing its efficiency in anticancer activity [447]. Overall, it is evident that siRNA is an efficient tool in cancer therapy and exosomes enhance its potential to suppress cancer and promote chemosensitivity [448]. Short hairpin RNA (shRNA) is another genetic tool that has a similar function to siRNA and can be used to regulate gene expression in cancer therapy. However, there is no study evaluating the potential of exosomes in shRNA, and further experiments could focus on this aspect.
CRISPR/Cas9 system
To date, only two studies have focused on CRISPR/Cas9 transfer through exosomes. In one experiment, cancer-derived exosomes were used to transfer the CRISPR/Cas9 system in the treatment of ovarian cancer. It seems that cancer-derived exosomes have high selectivity toward ovarian cancer cells due to cell tropism. CRISPR/Cas9-loaded exosomes significantly stimulated apoptosis by downregulating PARP-1. In addition, CRISPR/Cas9-loaded exosomes are able to increase the cytotoxicity of cisplatin (CP) against ovarian cancer cells [449]. Limitations of the CRISPR/Cas9 system include its difficulty in specifically targeting all cancer cells and its low efficacy in vivo. Therefore, it seems crucial to use exosomes for the delivery of the CRISPR/Cas9 system in cancer therapy. Exosomes can induce necroptosis in cancer cells through the CRISPR/Cas9 system (Fig. 7) [450]. However, few studies have investigated this potential of exosomes, and the development of engineered and surface-modified exosomes is encouraged.
Fig. 7.

The use of exosomes in the administration of genetic tools. Downregulation of tumor-promoting molecular signaling pathways such as survivin, Bcl-2, PLK1, HGF, and TPD52 by exosomes loaded with genetic tools leads to induction of apoptosis, impairment of tumor progression, and suppression of cancer metastasis
Tumor-derived exosomes
Brain tumors
The exosomes can be derived from glioblastoma cells. These exosomes mediate immune evasion of cancer cells via induction of PD-L1 expression and transfer of STAT3. In addition, exosomes induce M2 polarization of macrophages, which promotes glioblastoma progression [451]. When glioblastoma cells are exposed to hypoxia, the secretion of exosomes is triggered. These exosomes contain miRNA-1246, which is a tumor-promoting factor to promote cancer progression by upregulating STAT3 expression, suppressing NF-κB signaling, and mediating M2 polarization of macrophages [452]. Glioblastoma-derived exosomes contain high levels of VEGF-C, which inhibits Hippo signaling and enhances tafazzin (TAZ) expression, leading to angiogenesis [453]. Proteomics can be used to identify proteins embedded in exosomes and use them as biomarkers [454]. It appears that glioblastoma-derived exosomes have immunomodulatory effects. As mentioned previously, these exosomes are able to mediate M2 polarization of macrophages, which is attributed to the induction of NF-κB signaling. Then, macrophages secrete factors responsible for cancer progression. Moreover, these exosomes suppress the activity of cytotoxic CD4 + T cells to evade the immune response in glioblastomas [455]. Furthermore, exosomes from glioblastomas promote cancer stemness by transferring Notch1 protein [456].
A recent experiment has shown that exosomes derived from glioma stem cells contain high levels of Linc01060, which acts as a tumor-promoting factor and promotes cancer progression. The exosomal Linc01060 increases the stability of myeloid zinc finger 1 (MZF1) as a transcription factor and induces its nuclear translocation. Then, MZF1 induces HIF-1α via upregulation of c-Myc to enhance glioma progression [457]. Glioma-derived exosomes contain high levels of miRNA-10a and miRNA-21, which regulate PTEN and RORA, leading to activation of myeloid-derived suppressor cells and impairing immune function [458]. miRNA-1246 and mIRNA-10b-5p are other miRNAs found in glioma-derived exosomes that promote cancer cell metastasis [459]. Furthermore, glioma cells secrete exosomes containing the lncRNA CCAT2 to stimulate angiogenesis and inhibit apoptosis, setting the stage for tumor progression [460]. To trigger angiogenesis, glioma-derived exosomes may deliver miRNA-21, which upregulates VEGF expression [461]. Overall, these studies are consistent with the fact that glioma-derived exosomes modulate proliferation and migration by transporting various cargoes [462–464].
Thoracic and breast tumors
Breast cancer-derived exosomes are capable of suppressing immune function to enhance tumor progression. Injection of exosomes into naïve mice leads to accumulation of myeloid-derived suppressor cells in the lungs and the liver. Breast cancer-derived exosomes prevent T cell proliferation and suppress natural killer cell cytotoxicity to mediate immune evasion [465]. In enhancing cancer progression, breast cancer-derived exosomes transfer miRNA-155 to induce cachexia via downregulation of PARP-1 expression. Further studies revealed that these exosomes can also induce EMT-mediated metastasis in breast cancer by triggering catabolism and release of metabolites in adipocytes and muscle cells [466]. Metastatic breast cancer cells secrete exosomes containing miRNA-21 and miRNA-200c, which can be detected in patients and are used as diagnostic and prognostic factors [467]. The protein content of exosomes can be analyzed to distinguish breast cancer subtypes [468]. The presence of CD44 in breast cancer-derived exosomes leads to doxorubicin resistance [469]. Moreover, activation of fibroblasts by exosomes containing survivin can promote both growth and metastasis of breast cancer cells [470]. By inducing M2 polarization of macrophages, breast cancer-derived exosomes enhance lymph node metastasis of breast cancer cells [471]. Therefore, breast cancer-derived exosomes may modulate the progression of these tumor cells [472].
Exosomes derived from lung cancer cells, on the other hand, may act as triggers of EMT via upregulation of vimentin [473]. Exosomes derived from gemcitabine-resistant cancer cells may promote the progression of non-small cell lung cancer cells, mediate their drug resistance, and enhance their malignant phenotype through the transmission of miRNA-222-3p [474]. Exosomes derived from lung cancer cells are able to induce the Wnt3a/β-catenin axis to promote growth and survival [475]. Irradiation stimulates the release of exosomes from non-small cell lung cancer and induces Akt, STAT3, and ERK signaling pathways that mediate resistance to kinase inhibitors [476]. In additions, some of the proteins, such as MUC1, are enriched in exosomes to determine their localization and biological function [477].
Gastrointestinal tumors
Exosomes derived from gastric cancer enhance peritoneal metastasis and disrupt the mesothelial barrier [478]. These exosomes induce NF-κB signaling in macrophages to mediate secretion of pro-inflammatory factors and promote gastric cancer progression [479]. Induction of NF-κB signaling by gastric cancer-derived exosomes maintains inflammatory conditions in the TME that promote gastric cancer progression [480]. Gastric cancer-derived exosomes are able to stimulate PI3K/Akt and MERK/ERK signaling pathways. Moreover, inhibition of BMP prevents the potential of exosomes to transform pericytes into cancer-associated fibroblasts [481]. Exposure of gastric cancer cells to various antitumor agents may affect their ability to secrete exosomes. Pyrotinib, for example, induces the release of exosomes from gastric cancer cells. The secreted exosomes enhance migration, and the use of apatinib as a VEGFR inhibitor, suppresses this condition [482].
Hepatocellular carcinoma is another gastrointestinal tumor. Exosomes derived from hepatocellular carcinoma cells can induce ERK signaling to mediate EMT via upregulation of ZEB1/2, leading to cancer metastasis [483]. In addition to metastasis, exosomes derived from hepatocellular carcinomas also mediate cancer recurrence and can be used for early diagnosis of this malignancy [484]. By triggering chaperone-mediated autophagy, hepatocellular carcinoma-derived exosomes induce drug resistance and inhibit apoptosis [485]. Exosomes are also known to transfer Linc-ROR to liver cancer cells to increase their growth rate and inhibit their apoptosis [485].
Pancreatic cancer cells may also secrete exosomes. A recent experiment has shown that Dickkopf1 (DKK1)-dependent endocytosis is involved in the biogenesis of exosomes. Pancreatic cancer cell-derived exosomes have high levels of CKAP4 and are associated with poor prognosis in patients [486]. Moreover, pancreatic cancer-derived exosomes mediate M2 polarization of macrophages to suppress immune function against cancer cells [487]. To demonstrate the potential of exosomes in cancer migration, an experiment isolated serum exosomes from pancreatic cancer patients and showed that they can induce EMT and promote metastasis [488]. To enhance invasion and migration ability, pancreatic cancer cell-derived exosomes recruit cancer-associated fibroblasts and transfer Lin28B to reduce let-7 expression, leading to upregulation of HMGA2 and subsequent overexpression of PDGFB [489]. All in all, exosomes derived from pancreatic cancer cells regulate cancer progression, and their isolation and targeting may be important for cancer therapy [489–494].
Reproductive tumors
Most experiments on reproductive tumor-derived exosomes have focused on ovarian cancer. Proteomic and lipidomic analysis of exosomes can be used in the early diagnosis of ovarian cancer [495]. Ovarian cancer cell-derived exosomes may be involved in the development of malignant TME by promoting fibroblast migration [496]. They can be considered as potential therapeutic targets, as their modulation can suppress growth and invasion of ovarian cancer cells [497]. Moreover, ovarian cancer cell-derived exosomes can transport miRNAs into the TME and induce M2 polarization of macrophages that promote cancer progression [498]. Exosomal miRNA-940 stimulates ovarian cancer progression by inducing polarization of macrophages to the M2 phenotype [499]. Ovarian cancer-cell derived exosomes can induce angiogenesis and migration via upregulation of VEGF [500]. The same phenomenon occurs in cervical cancer. It has been reported that cervical cancer cell-derived exosomes can promote angiogenesis via Hedgehog-GLI signaling and enhancement of VEGF-A, VEGFR2, and angiopoietin-2 expression [501]. Loading dendritic cells with exosomes derived from HeLa cells stimulates anti-tumor immunity by increasing T-cell cytotoxicity [502]. Overall, these experiments highlight the role of ovarian and cervical cancer cell-derived exosomes in modulating migration, growth, TME, anti-tumor immunity, and angiogenesis. An experiment was conducted to investigate the role of prostate cancer-derived exosomes in immunomodulation. These exosomes impair dendritic cell function and suppress CD8 + T cell activity. The exosomes mediate the expression of CD73 on dendritic cells, which subsequently upregulate the expression of CD39, resulting in ATP-dependent inhibition of TNF-α and IL-12 production. In addition, exosomes have been found to contain prostaglandin E2, which enhances CD73 expression (Fig. 8) [503].
Fig. 8.

Tumor-derived exosomes and their role in cancer progression
Clinical application and role of exosomes as biomarkers
According to the role of exosomes in influencing cancer progression, a significant attempt has been made to reveal the role of exosomes in clinical background [504–508]. The genes contained in exosomes can be considered as biomarkers for the diagnosis of lung cancer. A recent experiment has shown that exosomes derived from non-small cell lung cancer cells have Hippo, Rap1, and Wnt as important signaling networks and can be considered as prognostic tools [509]. Another study shows that exosomes derived from non-small cell lung cancer contain high levels of PRPS2 and can mediate cisplatin resistance. In addition, PRPS2-containing exosomes are capable of inducing M2 polarization of macrophages and are associated with an unfavorable prognosis [510]. Exosomes containing high levels of miRNA-3362, miRNA-146a, and miRNA-1290 are observed in breast cancer patients and mediate lymph node metastasis and clinical stage [511]. In colorectal cancer patients, the abundance of QSOX1 in plasma exosomes decreases and can be considered as a diagnostic factor [511]. These studies demonstrate how exosomes can be analyzed to determine the prognosis of cancer patients (Table 4).
Table 4.
Clinical trials on the use of exosomes in cancer patients
| Status | Remarks | Reference |
|---|---|---|
| Completed | Use of exosomes as reliable biomarkers for the diagnosis of men with prostate cancer | NCT02702856 |
| Unknown | Combination of computed tomography and exosomes for diagnosis of early stage lung cancer | NCT03542253 |
| Active, not recruiting | Use of exosomes present in blood plasma to diagnose lung cancer in patients | NCT04529915 |
| Unknown | Use of circulating exosomes for diagnosis of advanced gastric cancer | NCT01779583 |
| Recruiting | New diagnostic method for colorectal cancer using exosomes | NCT04394572 |
| Recruiting | Presence of exosomes in tumor-draining vein and their molecular profiling | NCT04939324 |
| Recruiting | Use of plant exosome for delivery of curcumin in the treatment of colon cancer | NCT01294072 |
Conclusion and future perspectives
Thanks to the attempts made in recent years to uncover the factors involved in cancer progression, it is now clear that each factor has a unique fingerprint in cancer pathogenesis. If we know exactly how these factors interact in cancer, we can develop novel and effective therapeutics. Exosomes are minute structures that are involved in the regulation of biological processes through their cargo, which can be proteins, lipids, or nucleic acids. Genetic tools and anti-tumor agents can also be loaded into exosomes. Therefore, they provide intercellular communication and their involvement in cancer progression or inhibition is inevitable. Depending on the cargo, the effect of exosomes on the target cell may be different. In addition, normal cells such as macrophages and mesenchymal stem cells are capable of secreting exosomes to affect cancer cell progression. Therefore, exosomes can mediate both normal cell-cancer cell and cancer cell-cancer cell interactions.
Growth and invasion are the two most important aspects of tumor cells. When exosomes contain tumor-promoting substances, they can promote cell cycle progression and glycolysis and inhibit apoptosis. The role of autophagy is a bit confusing. It has been mentioned that induction of autophagy by exosomes can prevent apoptosis in cancer cells. Therefore, further experiments should be performed to reveal the interaction between apoptosis and autophagy in cancer cells affected by exosomes. Similar to proliferation, the cargo of exosomes determines the function of these structures in increasing or decreasing cancer migration and invasion. EMT and MMPs are strongly influenced by exosomes in regulating cancer progression. However, most studies have focused on the EMT mechanism, and it is proposed to uncover the signaling networks affected by exosomes in targeting MMPs and modulating cancer metastasis, because of the important role of MMPs in this case. Cancer cell proliferation and invasion rates determine response to therapy. If cancer cells have a high capacity to migrate and grow, they may develop resistance to therapy. Therefore, by targeting exosomes, proliferation and invasion of cancer cells can be modulated and their response to therapy can be predicted. The aggressive behavior of cancer cells depends mainly on interactions in the TME. The best known interaction in the TME is macrophage polarization mediated by exosomes. Exosomes can induce M2 polarization of macrophages, promoting cancer cell progression.
Since response to therapy is a major concern for physicians treating cancer patients, we have provided a section specifically addressing the role of exosomes in this case. The sensitivity of cancer cells to chemotherapy-mediated apoptosis can be reduced by exosomes. Because of the potential of exosomes to transfer various genes, they can influence the progression of cancer cells and determine their response to therapy. In addition to drug resistance, exosomes may also be involved in triggering radioresistance. In addition, exosomes can induce immune cell exhaustion, decrease T cell cytotoxicity, and mediate immune evasion. By triggering chronic inflammation, exosomes promote cancer progression. When new therapeutics are to be introduced into clinics for the treatment of cancer patients, they can focus on these aspects.
For internalization into cells, exosomes can follow different pathways. Exosomes are able to bind to receptors on the surface of cells, and can be internalized by binding to integrins, tetraspanins and intercellular adhesion molecules. Clathrin- and caveolin-mediated endocytosis, lipid raft uptake, macropinocytosis, phagocytosis and fusion with the plasma membrane [512]. Therefore, if exosomes are to be used for cargo transport in cancer therapy, the method of their internalization should be elucidated and subsequent functionalization should be performed to improve their intracellular accumulation.
Exosomes may contain miRNAs, lncRNAs, circRNAs, and other genes such as PTEN, PI3K/Akt, and STAT3 that affect cancer progression. Indeed, cancer progression is strongly influenced by exosome cargo. Since exosomes are capable of delivering various drugs, they have the potential to be used as delivery systems for anti-tumor agents and genetic tools in cancer therapy. The various anti-cancer agents, including plant-derived natural compounds such as triptolides and synthetic agents such as cisplatin, doxorubicin and paclitaxel, can be transferred by exosomes in cancer therapy. The siRNA and CRISPR/Cas9 are genetic tools embedded in exosomes for cancer therapy. Delivery of therapeutics using exosomes can potentially lead to increased intracellular accumulation and improved therapeutic efficacy. In addition, the surface of exosomes can be modified with ligands to increase their selectivity toward cancer cells. Since exosomes affect various aspects of cancer cells, they can be isolated from the serum of patients and are considered reliable biomarkers for the diagnosis and prognosis of cancer patients. As shown in Table 4, exosomes have been used as biomarkers in various experiments in cancer patients. In addition to diagnosis, exosomes have also been used to increase the accuracy of other methods of detecting cancer patients, such as CT. Of note, a clinical trial is currently underway to deliver curcumin as an anti-cancer agent to treat colon cancer. The results of this clinical trial are of great importance, as they may provide novel insights into the role of exosomes as drug delivery systems in the clinical course and their safety. Furthermore, a number of clinical trials on molecular profiling of exosomes are currently ongoing, which could be useful in the field of precision medicine in the near future.
Acknowledgements
We would like to thank Academic Proofreading (www.academicproofreading.uk) for their excellent English language editing services.
Abbreviations
- CSC
Cancer stem cell
- EVs
Extracellular vesicles
- TME
Tumor microenvironment
- ncRNAs
Non-coding RNAs
- miRNA
MicroRNA
- lncRNA
Long non-coding RNA
- circRNA
Circular RNA
- MVBs
Multi-vesicular bodies
- ILVs
Intraluminal vesicles
- ESCRT
Endosomal sorting complex required for transport
- VTA1
Vesicle trafficking 1
- VPS4
Vacuolar protein sorting-associated protein 4
- TSG101
Tumor susceptibility gene 101 protein
- HSP60
Heat shock protein 60
- Rab
Ras-associated binding
- SNARE
Soluble NSF-attachment protein receptor
- STAT3
Signal transducer and activator of transcription 3
- IL-6
Interleukin-6
- MMP-9
Matrix metalloproteinase-9
- ROS
Reactive oxygen species
- BBB
Blood–brain barrier
- ERK
Extracellular-signal-regulated kinase
- SIRT2
Sirtuin 2
- TFEB
Transcription factor EB
- EphA2
Eph receptor A2
- EMT
Epithelial-to-mesenchymal transition
- EMT-TFs
EMT-inducing transcription factors
- MMPs
Matrix metalloproteinases
- TFF3
Trefoil factor 3
- VEGF
Vascular endothelial growth factor
- VEGFR2
VEGF receptor 2
- NRP-1
Neuropilin-1
- OSCC
Oral squamous cell carcinoma
- NPS
Nasopharyngeal carcinoma
- ANGPT2
Angiopoietin-2
- HCC
Hepatocellular carcinoma
- HIF-1α
Hypoxia inducible factor-1α
- TEX
Tumor-derived exosomes
- 5-FU
5-Fluorouracil
- CP
Cisplatin
- TGF-β
Transforming growth factor-beta
- Treg cells
T regulatory cells
- PD-1
Programmed death-1
- IDO
Indoleamine 2:3-dioxygenase
- 3/-UTR
3/-Untranslated region
- PTEN
Phosphatase and tensin homolog
- PTX
Paclitaxel
- AA-PEG
Aminoethylanisamide-polyethylene glycol
- RISC
RNA-induced silencing complex
- HGF
Hepatocyte growth factor
- PLK1
Polo-like kinase 1
- EGFR
Epidermal growth factor receptor
- shRNA
Short hairpin RNA
- TA2
Tafazzin
- MZF1
Myeloid zinc finger 1
- DKK1
Dickkopf1
Author contributions
MA and APK conceptualized the topic. MDAP, ME, SM, AZ (Amirhossein Zabolian), MJN, SS, MAK, MH, KH and MA participated in literature searches, writing and preparing the first draft. AZ (Ali Zarrabi) performed software works, depicting figures and participated in the draft writing. GS MA, AZ (Ali Zarrabi), APK, SCT, MP, and AA supervised and critically edited the draft for submission. All authors read and approved the final manuscript.
Funding
Open Access funding enabled and organized by Projekt DEAL. The work was supported by a grant from the Singapore Ministry of Education Tier 2 (MOE-T2EP30120-0016) to A.P.K. A.P.K. is also supported by the National Medical Research Council of Singapore and the Singapore Ministry of Education under its Research Centers of Excellence initiative to Cancer Science Institute of Singapore, National University of Singapore. S.C.T. is supported by the Research University Grant of Universiti Kebangsaan Malaysia (No. GUP-2020-076) and the Fundamental Research Grant Scheme of the Ministry of Higher Education, Malaysia (No. FRGS/1/2019/SKK08/UKM/02/9).
Availability of data and materials
Not applicable.
Declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interest.
Footnotes
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Mahshid Deldar Abad Paskeh, Maliheh Entezari, and Sepideh Mirzaei contributed equally to this paper and are considered co-first authors.
Contributor Information
Shing Cheng Tan, Email: sctan@ukm.edu.my, Email: shingchengtan@gmail.com.
Marios Papadakis, Email: marios_papadakis@yahoo.gr.
Milad Ashrafizadeh, Email: milad.ashrafizadeh@sabanciuniv.edu.
References
- 1.Cai W, Xiong Chen Z, Rane G, Satendra Singh S, Choo Z, Wang C, et al. Wanted DEAD/H or alive: helicases winding up in cancers. J Natl Cancer Inst. 2017;109(6):djw278. doi: 10.1093/jnci/djw278. [DOI] [PubMed] [Google Scholar]
- 2.Manu KA, Shanmugam MK, Ramachandran L, Li F, Siveen KS, Chinnathambi A, et al. Isorhamnetin augments the anti-tumor effect of capecitabine through the negative regulation of NF-κB signaling cascade in gastric cancer. Cancer Lett. 2015;363(1):28–36. doi: 10.1016/j.canlet.2015.03.033. [DOI] [PubMed] [Google Scholar]
- 3.Wang C, Kar S, Lai X, Cai W, Arfuso F, Sethi G, et al. Triple negative breast cancer in Asia: an insider's view. Cancer Treat Rev. 2018;62:29–38. doi: 10.1016/j.ctrv.2017.10.014. [DOI] [PubMed] [Google Scholar]
- 4.Li F, Shanmugam MK, Chen L, Chatterjee S, Basha J, Kumar AP, et al. Garcinol, a polyisoprenylated benzophenone modulates multiple proinflammatory signaling cascades leading to the suppression of growth and survival of head and neck carcinoma. Cancer Prev Res (Phila) 2013;6(8):843–854. doi: 10.1158/1940-6207.CAPR-13-0070. [DOI] [PubMed] [Google Scholar]
- 5.Ang HL, Yuan Y, Lai X, Tan TZ, Wang L, Huang BB, et al. Putting the BRK on breast cancer: From molecular target to therapeutics. Theranostics. 2021;11(3):1115–1128. doi: 10.7150/thno.49716. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Sethi G, Shanmugam MK, Warrier S, Merarchi M, Arfuso F, Kumar AP, et al. Pro-apoptotic and anti-cancer properties of diosgenin: a comprehensive and critical review. Nutrients. 2018;10(5):645. doi: 10.3390/nu10050645. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Patel SM, Nagulapalli Venkata KC, Bhattacharyya P, Sethi G, Bishayee A. Potential of neem (Azadirachta indica L.) for prevention and treatment of oncologic diseases. Semin Cancer Biol. 2016;40–41:100–15. doi: 10.1016/j.semcancer.2016.03.002. [DOI] [PubMed] [Google Scholar]
- 8.Chopra P, Sethi G, Dastidar SG, Ray A. Polo-like kinase inhibitors: an emerging opportunity for cancer therapeutics. Expert Opin Investig Drugs. 2010;19(1):27–43. doi: 10.1517/13543780903483191. [DOI] [PubMed] [Google Scholar]
- 9.Warrier S, Patil M, Bhansali S, Varier L, Sethi G. Designing precision medicine panels for drug refractory cancers targeting cancer stemness traits. Biochim Biophys Acta. 2021;1875(1):188475. doi: 10.1016/j.bbcan.2020.188475. [DOI] [PubMed] [Google Scholar]
- 10.Kashyap D, Tuli HS, Yerer MB, Sharma A, Sak K, Srivastava S, et al. Natural product-based nanoformulations for cancer therapy: opportunities and challenges. Semin Cancer Biol. 2021;69:5–23. doi: 10.1016/j.semcancer.2019.08.014. [DOI] [PubMed] [Google Scholar]
- 11.Ailuno G, Baldassari S, Lai F, Florio T, Caviglioli GJC. Exosomes and extracellular vesicles as emerging theranostic platforms in cancer research cells. Semin Cancer Biol. 2020;9(12):2569. doi: 10.3390/cells9122569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Jayasinghe MK, Tan M, Peng B, Yang Y, Sethi G, Pirisinu M, et al. New approaches in extracellular vesicle engineering for improving the efficacy of anti-cancer therapies. Semin Cancer Biol. 2021 doi: 10.1016/j.semcancer.2021.02.010. [DOI] [PubMed] [Google Scholar]
- 13.Colombo M, Raposo G, Théry C. Biogenesis, secretion, and intercellular interactions of exosomes and other extracellular vesicles. Annu Rev Cell Dev Biol. 2014;30:255–289. doi: 10.1146/annurev-cellbio-101512-122326. [DOI] [PubMed] [Google Scholar]
- 14.Weng J, Xiang X, Ding L, Wong AL, Zeng Q, Sethi G, et al. Extracellular vesicles, the cornerstone of next-generation cancer diagnosis? Semin Cancer Biol. 2021 doi: 10.1016/j.semcancer.2021.05.011. [DOI] [PubMed] [Google Scholar]
- 15.Cocucci E, Racchetti G, Meldolesi J. Shedding microvesicles: artefacts no more. Trends Cell Biol. 2009;19(2):43–51. doi: 10.1016/j.tcb.2008.11.003. [DOI] [PubMed] [Google Scholar]
- 16.Hristov M, Erl W, Linder S, Weber PC. Apoptotic bodies from endothelial cells enhance the number and initiate the differentiation of human endothelial progenitor cells in vitro. Blood. 2004;104(9):2761–2766. doi: 10.1182/blood-2003-10-3614. [DOI] [PubMed] [Google Scholar]
- 17.Raposo G, Stoorvogel W. Extracellular vesicles: exosomes, microvesicles, and friends. J Cell Biol. 2013;200(4):373–383. doi: 10.1083/jcb.201211138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Wu P, Zhang B, Ocansey DKW, Xu W, Qian H. Extracellular vesicles: a bright star of nanomedicine. Biomaterials. 2021;269:120467. doi: 10.1016/j.biomaterials.2020.120467. [DOI] [PubMed] [Google Scholar]
- 19.Möller A, Lobb RJ. The evolving translational potential of small extracellular vesicles in cancer. Nat Rev Cancer. 2020;20(12):697–709. doi: 10.1038/s41568-020-00299-w. [DOI] [PubMed] [Google Scholar]
- 20.Mathew M, Zade M, Mezghani N, Patel R, Wang Y, Momen-Heravi F. Extracellular vesicles as biomarkers in cancer immunotherapy. Cancers. 2020;12(10):2825. doi: 10.3390/cancers12102825. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Vlassov AV, Magdaleno S, Setterquist R, Conrad R. Exosomes: current knowledge of their composition, biological functions, and diagnostic and therapeutic potentials. Biochem Biophys Acta. 2012;1820(7):940–948. doi: 10.1016/j.bbagen.2012.03.017. [DOI] [PubMed] [Google Scholar]
- 22.Wee I, Syn N, Sethi G, Goh BC, Wang L. Role of tumor-derived exosomes in cancer metastasis. Biochim Biophys Acta. 2019;1871(1):12–19. doi: 10.1016/j.bbcan.2018.10.004. [DOI] [PubMed] [Google Scholar]
- 23.Denzer K, Kleijmeer MJ, Heijnen HF, Stoorvogel W, Geuze HJ. Exosome: from internal vesicle of the multivesicular body to intercellular signaling device. J Cell Sci. 2000;113(Pt 19):3365–3374. doi: 10.1242/jcs.113.19.3365. [DOI] [PubMed] [Google Scholar]
- 24.Ashrafizaveh S, Ashrafizadeh M, Zarrabi A, Husmandi K, Zabolian A, Shahinozzaman M, et al. Long non-coding RNA in the doxorubicin resistance of cancer cells. Cancer Lett. 2021 doi: 10.1016/j.canlet.2021.03.018. [DOI] [PubMed] [Google Scholar]
- 25.Hu T, Wolfram J, Srivastava S. Extracellular vesicles in cancer detection: hopes and hypes. Trends Cancer. 2021;7(2):122–133. doi: 10.1016/j.trecan.2020.09.003. [DOI] [PubMed] [Google Scholar]
- 26.Shoucair I, Weber Mello F, Jabalee J, Maleki S, Garnis C. The role of cancer-associated fibroblasts and extracellular vesicles in tumorigenesis. Int J Mol Sci. 2020;21(18):6837. doi: 10.3390/ijms21186837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Schubert A, Boutros M. Extracellular vesicles and oncogenic signaling. Mol Oncol. 2021;15(1):3–26. doi: 10.1002/1878-0261.12855. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Palazzolo S, Memeo L, Hadla M, Duzagac F, Steffan A, Perin T, et al. Cancer extracellular vesicles: next-generation diagnostic and drug delivery nanotools. Cancers. 2020;12(11):3165. doi: 10.3390/cancers12113165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Sun H, Burrola S, Wu J, Ding WQ. Extracellular vesicles in the development of cancer therapeutics. Int J Mol Sci. 2020;21(17):6097. doi: 10.3390/ijms21176097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Huyan T, Li H, Peng H, Chen J, Yang R, Zhang W, et al. Extracellular vesicles - advanced nanocarriers in cancer therapy: progress and achievements. Int J Nanomed. 2020;15:6485–6502. doi: 10.2147/IJN.S238099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Deb A, Gupta S, Mazumder PJ. Exosomes: a new horizon in modern medicine. Life Sci. 2020;264:118623. doi: 10.1016/j.lfs.2020.118623. [DOI] [PubMed] [Google Scholar]
- 32.Théry C, Zitvogel L, Amigorena SJ. Exosomes: composition, biogenesis and function. Nat Rev Immunol. 2002;2(8):569–79. doi: 10.1038/nri855. [DOI] [PubMed] [Google Scholar]
- 33.Syn N, Wang L, Sethi G, Thiery JP, Goh BC. Exosome-mediated metastasis: from epithelial-mesenchymal transition to escape from immunosurveillance. Trends Pharmacol Sci. 2016;37(7):606–617. doi: 10.1016/j.tips.2016.04.006. [DOI] [PubMed] [Google Scholar]
- 34.Laulagnier K, Motta C, Hamdi S, Roy S, Fauvelle F, Pageaux J-F, et al. Mast cell-and dendritic cell-derived exosomes display a specific lipid composition and an unusual membrane organization. Biochem J. 2004;380(1):161–171. doi: 10.1042/BJ20031594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Staubach S, Razawi H, Hanisch FGJ. Proteomics of MUC1-containing lipid rafts from plasma membranes and exosomes of human breast carcinoma cells MCF-7. Proteomics. 2009;9(10):2820–35. doi: 10.1002/pmic.200800793. [DOI] [PubMed] [Google Scholar]
- 36.Tamkovich S, Tutanov O, Laktionov P. Exosomes: generation, structure, transport, biological activity, and diagnostic application. Biochem Moscow Suppl Ser A. 2016;10(3):163–73. [Google Scholar]
- 37.Lu GD, Cheng P, Liu T, Wang Z. BMSC-derived exosomal miR-29a promotes angiogenesis and osteogenesis. Front Cell Dev Biol. 2020;8:608521. doi: 10.3389/fcell.2020.608521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Jia J, Guo S, Zhang D, Tian X, Xie X. Exosomal-lncRNA DLEU1 Accelerates the Proliferation, Migration, and Invasion of Endometrial Carcinoma Cells by Regulating microRNA-E2F3. Onco Targets Ther. 2020;13:8651–8663. doi: 10.2147/OTT.S262661. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Tian C, Liu J, Di X, Cong S, Zhao M, Wang K. Exosomal hsa_circRNA_104484 and hsa_circRNA_104670 may serve as potential novel biomarkers and therapeutic targets for sepsis. Sci Rep. 2021;11(1):14141. doi: 10.1038/s41598-021-93246-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Romano E, Netti PA, Torino EJP. Exosomes in gliomas: biogenesis, isolation, and preliminary applications in nanomedicine. Pharmaceuticals. 2020;13(10):319. doi: 10.3390/ph13100319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Burkova EE, Sedykh SE, Nevinsky GA. Human placenta exosomes: Biogenesis, isolation, composition, and prospects for use in diagnostics. Int J Mol Sci. 2021;22(4):2158. doi: 10.3390/ijms22042158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Whiteside TL, Diergaarde B, Hong C-S. Tumor-derived exosomes (TEX) and their role in immuno-oncology. Int J Mol Sci. 2021;22(12):6234. doi: 10.3390/ijms22126234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Jafari D, Shajari S, Jafari R, Mardi N, Gomari H, Ganji F, et al. Designer exosomes: a new platform for biotechnology therapeutics. BioDrugs. 2020 doi: 10.1007/s40259-020-00434-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Jiang Z, Liu G, Li J. Recent progress on the isolation and detection methods of exosomes. Chem Asian J. 2020;15(23):3973–3982. doi: 10.1002/asia.202000873. [DOI] [PubMed] [Google Scholar]
- 45.Alzhrani GN, Alanazi ST, Alsharif SY, Albalawi AM, Alsharif AA, Abdel-Maksoud MS, et al. Exosomes: isolation, characterization, and biomedical applications. Cell Biol Int. 2021 doi: 10.1002/cbin.11620. [DOI] [PubMed] [Google Scholar]
- 46.An M, Wu J, Zhu J, Lubman DM. Comparison of an optimized ultracentrifugation method versus size-exclusion chromatography for isolation of exosomes from human serum. J Proteome Res. 2018;17(10):3599–3605. doi: 10.1021/acs.jproteome.8b00479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Greening DW, Xu R, Ji H, Tauro BJ, Simpson RJ. A protocol for exosome isolation and characterization: evaluation of ultracentrifugation, density-gradient separation, and immunoaffinity capture methods. Methods Mol Biol (Clifton, NJ) 2015;1295:179–209. doi: 10.1007/978-1-4939-2550-6_15. [DOI] [PubMed] [Google Scholar]
- 48.Momen-Heravi F. Isolation of extracellular vesicles by ultracentrifugation. Methods Mol Biol (Clifton, NJ) 2017;1660:25–32. doi: 10.1007/978-1-4939-7253-1_3. [DOI] [PubMed] [Google Scholar]
- 49.Barreiro K, Huber TB, Holthofer H. Isolating urinary extracellular vesicles as biomarkers for diabetic disease. Methods Mol Biol (Clifton, NJ) 2020;2067:175–188. doi: 10.1007/978-1-4939-9841-8_13. [DOI] [PubMed] [Google Scholar]
- 50.Lobb RJ, Becker M, Wen SW, Wong CS, Wiegmans AP, Leimgruber A, et al. Optimized exosome isolation protocol for cell culture supernatant and human plasma. J Extracell Vesicles. 2015;4:27031. doi: 10.3402/jev.v4.27031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Shu S, Yang Y, Allen CL, Hurley E, Tung KH, Minderman H, et al. Purity and yield of melanoma exosomes are dependent on isolation method. J Extracell Vesicles. 2020;9(1):1692401. doi: 10.1080/20013078.2019.1692401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Feng Y, Huang W, Wani M, Yu X, Ashraf M. Ischemic preconditioning potentiates the protective effect of stem cells through secretion of exosomes by targeting Mecp2 via miR-22. PLoS One. 2014;9(2):e88685. doi: 10.1371/journal.pone.0088685. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Takov K, Yellon DM, Davidson SM. Comparison of small extracellular vesicles isolated from plasma by ultracentrifugation or size-exclusion chromatography: yield, purity and functional potential. J Extracell Vesicles. 2019;8(1):1560809. doi: 10.1080/20013078.2018.1560809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Buschmann D, Kirchner B, Hermann S, Märte M, Wurmser C, Brandes F, et al. Evaluation of serum extracellular vesicle isolation methods for profiling miRNAs by next-generation sequencing. J Extracell Vesicles. 2018;7(1):1481321. doi: 10.1080/20013078.2018.1481321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Rider MA, Hurwitz SN, Meckes DG., Jr ExtraPEG: a polyethylene glycol-based method for enrichment of extracellular vesicles. Sci Rep. 2016;6:23978. doi: 10.1038/srep23978. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Alvarez ML, Khosroheidari M, Kanchi Ravi R, DiStefano JK. Comparison of protein, microRNA, and mRNA yields using different methods of urinary exosome isolation for the discovery of kidney disease biomarkers. Kidney Int. 2012;82(9):1024–1032. doi: 10.1038/ki.2012.256. [DOI] [PubMed] [Google Scholar]
- 57.Zarovni N, Corrado A, Guazzi P, Zocco D, Lari E, Radano G, et al. Integrated isolation and quantitative analysis of exosome shuttled proteins and nucleic acids using immunocapture approaches. Methods (San Diego, Calif) 2015;87:46–58. doi: 10.1016/j.ymeth.2015.05.028. [DOI] [PubMed] [Google Scholar]
- 58.Helwa I, Cai J, Drewry MD, Zimmerman A, Dinkins MB, Khaled ML, et al. A comparative study of serum exosome isolation using differential ultracentrifugation and three commercial reagents. PLoS One. 2017;12(1):e0170628. doi: 10.1371/journal.pone.0170628. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Salafi T, Zeming KK, Zhang Y. Advancements in microfluidics for nanoparticle separation. Lab Chip. 2016;17(1):11–33. doi: 10.1039/c6lc01045h. [DOI] [PubMed] [Google Scholar]
- 60.Lin S, Yu Z, Chen D, Wang Z, Miao J, Li Q, et al. Progress in microfluidics-based exosome separation and detection technologies for diagnostic applications. Small. 2020;16(9):e1903916. doi: 10.1002/smll.201903916. [DOI] [PubMed] [Google Scholar]
- 61.He M, Crow J, Roth M, Zeng Y, Godwin AK. Integrated immunoisolation and protein analysis of circulating exosomes using microfluidic technology. Lab Chip. 2014;14(19):3773–3780. doi: 10.1039/c4lc00662c. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Contreras-Naranjo JC, Wu HJ, Ugaz VM. Microfluidics for exosome isolation and analysis: enabling liquid biopsy for personalized medicine. Lab Chip. 2017;17(21):3558–3577. doi: 10.1039/c7lc00592j. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Zhu L, Sun H-T, Wang S, Huang S-L, Zheng Y, Wang C-Q, et al. Isolation and characterization of exosomes for cancer research. J Hematol Oncol. 2020;13(1):1–24. doi: 10.1186/s13045-020-00987-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Kurian TK, Banik S, Gopal D, Chakrabarti S, Mazumder N. Elucidating methods for isolation and quantification of exosomes: a review. Mol Biotechnol. 2021;63:249–266. doi: 10.1007/s12033-021-00300-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Willis GR, Kourembanas S, Mitsialis SA. Toward exosome-based therapeutics: isolation, heterogeneity, and fit-for-purpose potency. Front Cardiovasc Med. 2017;4:63. doi: 10.3389/fcvm.2017.00063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Chen Y-S, Lin E-Y, Chiou T-W, Harn H-J. Exosomes in clinical trial and their production in compliance with good manufacturing practice. Tzu-Chi Med J. 2019;32(2):113–120. doi: 10.4103/tcmj.tcmj_182_19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Dai J, Su Y, Zhong S, Cong L, Liu B, Yang J, et al. Exosomes: key players in cancer and potential therapeutic strategy. Sig Transduct Target Ther. 2020;5(1):1–10. doi: 10.1038/s41392-020-00261-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Ibrahim A, Marbán E. Exosomes: fundamental biology and roles in cardiovascular physiology. Annu Rev Physiol. 2016;78:67–83. doi: 10.1146/annurev-physiol-021115-104929. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Rastogi S, Sharma V, Bharti PS, Rani K, Modi GP, Nikolajeff F, et al. The evolving landscape of exosomes in neurodegenerative diseases: exosomes characteristics and a promising role in early diagnosis. Int J Mol Sci. 2021;22(1):440. doi: 10.3390/ijms22010440. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Heijnen HF, Schiel AE, Fijnheer R, Geuze HJ, Sixma JJ. Activated platelets release two types of membrane vesicles: microvesicles by surface shedding and exosomes derived from exocytosis of multivesicular bodies and alpha-granules. Blood. 1999;94(11):3791–3799. [PubMed] [Google Scholar]
- 71.Radulovic M, Stenmark H. ESCRTs in membrane sealing. Biochem Soc Trans. 2018;46(4):773–778. doi: 10.1042/BST20170435. [DOI] [PubMed] [Google Scholar]
- 72.Stuffers S, Sem Wegner C, Stenmark H, Brech A. Multivesicular endosome biogenesis in the absence of ESCRTs. Traffic (Copenhagen, Denmark) 2009;10(7):925–937. doi: 10.1111/j.1600-0854.2009.00920.x. [DOI] [PubMed] [Google Scholar]
- 73.Henne WM, Buchkovich NJ, Emr SD. The ESCRT pathway. Dev Cell. 2011;21(1):77–91. doi: 10.1016/j.devcel.2011.05.015. [DOI] [PubMed] [Google Scholar]
- 74.Mulcahy LA, Pink RC, Carter DR. Routes and mechanisms of extracellular vesicle uptake. J Extracell Vesicles. 2014;3:24641. doi: 10.3402/jev.v3.24641. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Lötvall J, Hill AF, Hochberg F, Buzás EI, Di Vizio D, Gardiner C, et al. Minimal experimental requirements for definition of extracellular vesicles and their functions: a position statement from the international society for extracellular vesicles. J Extracell Vesicles. 2014;3:26913. doi: 10.3402/jev.v3.26913. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Kowal J, Arras G, Colombo M, Jouve M, Morath JP, Primdal-Bengtson B, et al. Proteomic comparison defines novel markers to characterize heterogeneous populations of extracellular vesicle subtypes. Proc Natl Acad Sci USA. 2016;113(8):E968–E977. doi: 10.1073/pnas.1521230113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Klumperman J, Raposo G. The complex ultrastructure of the endolysosomal system. Cold Spring Harb Perspect Biol. 2014;6(10):a016857. doi: 10.1101/cshperspect.a016857. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Anand S, Samuel M, Kumar S, Mathivanan S. Ticket to a bubble ride: Cargo sorting into exosomes and extracellular vesicles. Biochim Biophys Acta. 2019;1867(12):140203. doi: 10.1016/j.bbapap.2019.02.005. [DOI] [PubMed] [Google Scholar]
- 79.Alenquer M, Amorim MJ. Exosome biogenesis, regulation, and function in viral infection. Viruses. 2015;7(9):5066–83. doi: 10.3390/v7092862. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Camuzard O, Santucci-Darmanin S, Carle GF, Pierrefite-Carle V. Autophagy in the crosstalk between tumor and microenvironment. Cancer Lett. 2020;490:143–153. doi: 10.1016/j.canlet.2020.06.015. [DOI] [PubMed] [Google Scholar]
- 81.Ma Z, Wang LZ, Cheng JT, Lam WST, Ma X, Xiang X, et al. Targeting hypoxia-inducible factor-1-mediated metastasis for cancer therapy. Antioxid Redox Signal. 2021;34(18):1484–1497. doi: 10.1089/ars.2019.7935. [DOI] [PubMed] [Google Scholar]
- 82.Shanmugam MK, Warrier S, Kumar AP, Sethi G, Arfuso F. Potential role of natural compounds as anti-angiogenic agents in cancer. Curr Vasc Pharmacol. 2017;15(6):503–519. doi: 10.2174/1570161115666170713094319. [DOI] [PubMed] [Google Scholar]
- 83.Ye J, Wu D, Wu P, Chen Z, Huang JJ. The cancer stem cell niche: cross talk between cancer stem cells and their microenvironment. Tumor Biol. 2014;35(5):3945–3951. doi: 10.1007/s13277-013-1561-x. [DOI] [PubMed] [Google Scholar]
- 84.Kalluri R, Zeisberg MJ. Fibroblasts in cancer. Nat Rev Cancer. 2006;6(5):392–401. doi: 10.1038/nrc1877. [DOI] [PubMed] [Google Scholar]
- 85.Qian L-W, Mizumoto K, Maehara N, Ohuchida K, Inadome N, Saimura M, et al. Co-cultivation of pancreatic cancer cells with orthotopic tumor-derived fibroblasts: fibroblasts stimulate tumor cell invasion via HGF secretion whereas cancer cells exert a minor regulative effect on fibroblasts HGF production. Cancer Lett. 2003;190(1):105–12. doi: 10.1016/s0304-3835(02)00517-7. [DOI] [PubMed] [Google Scholar]
- 86.Jin M-Z, Jin W-L. The updated landscape of tumor microenvironment and drug repurposing. Signal Transduct Target Ther. 2020;5(1):166. doi: 10.1038/s41392-020-00280-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Datta A, Deng S, Gopal V, Yap KC, Halim CE, Lye ML, et al. Cytoskeletal dynamics in epithelial-mesenchymal transition: insights into therapeutic targets for cancer metastasis. Cancers. 2021;13(8):1882. doi: 10.3390/cancers13081882. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Cheng JT, Wang L, Wang H, Tang FR, Cai WQ, Sethi G, et al. Insights into biological role of LncRNAs in epithelial-mesenchymal transition. Cells. 2019;8(10):1178. doi: 10.3390/cells8101178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Moraes LA, Kar S, Foo SL, Gu T, Toh YQ, Ampomah PB, et al. Annexin-A1 enhances breast cancer growth and migration by promoting alternative macrophage polarization in the tumour microenvironment. Sci Rep. 2017;7(1):17925. doi: 10.1038/s41598-017-17622-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Yang Y, Guo Z, Chen W, Wang X, Cao M, Han X, et al. M2 macrophage-derived exosomes promote angiogenesis and growth of pancreatic ductal adenocarcinoma by targeting E2F2. Mol Ther J Am Soc Gene Ther. 2021;29(3):1226–1238. doi: 10.1016/j.ymthe.2020.11.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Dong F, Ruan S, Wang J, Xia Y, Le K, Xiao X, et al. M2 macrophage-induced lncRNA PCAT6 facilitates tumorigenesis and angiogenesis of triple-negative breast cancer through modulation of VEGFR2. Cell Death Dis. 2020;11(9):728. doi: 10.1038/s41419-020-02926-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Jung YY, Um J-Y, Nasif O, Alharbi SA, Sethi G, Ahn KS. Blockage of the JAK/STAT3 signaling pathway in multiple myeloma by leelamine. Phytomedicine. 2021;87:153574. doi: 10.1016/j.phymed.2021.153574. [DOI] [PubMed] [Google Scholar]
- 93.Mohan CD, Rangappa S, Nayak SC, Sethi G, Rangappa KS. Paradoxical functions of long noncoding RNAs in modulating STAT3 signaling pathway in hepatocellular carcinoma. Biochim Biophys Acta (BBA) Rev Cancer. 2021;1876:188574. doi: 10.1016/j.bbcan.2021.188574. [DOI] [PubMed] [Google Scholar]
- 94.Garg M, Shanmugam MK, Bhardwaj V, Goel A, Gupta R, Sharma A, et al. The pleiotropic role of transcription factor STAT3 in oncogenesis and its targeting through natural products for cancer prevention and therapy. Med Res Rev. 2021;41(3):1291–336. doi: 10.1002/med.21761. [DOI] [PubMed] [Google Scholar]
- 95.Jung YY, Ha IJ, Um J-Y, Sethi G, Ahn KS. Fangchinoline diminishes STAT3 activation by stimulating oxidative stress and targeting SHP-1 protein in multiple myeloma model. J Adv Res. 2021 doi: 10.1016/j.jare.2021.03.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Lee JH, Mohan CD, Deivasigamani A, Jung YY, Rangappa S, Basappa S, et al. Brusatol suppresses STAT3-driven metastasis by downregulating epithelial-mesenchymal transition in hepatocellular carcinoma. J Adv Res. 2020;26:83–94. doi: 10.1016/j.jare.2020.07.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Xu J, Zhang J, Zhang Z, Gao Z, Qi Y, Qiu W, et al. Hypoxic glioma-derived exosomes promote M2-like macrophage polarization by enhancing autophagy induction. Cell Death Dis. 2021;12(4):373. doi: 10.1038/s41419-021-03664-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Ge L, Zhou F, Nie J, Wang X, Zhao Q. Hypoxic colorectal cancer-secreted exosomes deliver miR-210–3p to normoxic tumor cells to elicit a protumoral effect. Exp Biol Med (Maywood, NJ) 2021 doi: 10.1177/15353702211011576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Yin H, Qiu X, Shan Y, You B, Xie L, Zhang P, et al. HIF-1α downregulation of miR-433-3p in adipocyte-derived exosomes contributes to NPC progression via targeting SCD1. Cancer Sci. 2021;112(4):1457–1470. doi: 10.1111/cas.14829. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Li Y, Zhang X, Zheng Q, Zhang Y, Ma Y, Zhu C, et al. YAP1 inhibition in HUVECs Is associated with released exosomes and increased hepatocarcinoma invasion and metastasis. Mol Ther Nucleic Acids. 2020;21:86–97. doi: 10.1016/j.omtn.2020.05.021. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 101.Wu J, Gao W, Tang Q, Yu Y, You W, Wu Z, et al. M2 macrophage-derived exosomes facilitate HCC metastasis by transferring α(M) β(2) integrin to tumor cells. Hepatology (Baltimore, MD) 2021;73(4):1365–1380. doi: 10.1002/hep.31432. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 102.Zhou W, Zhou Y, Chen X, Ning T, Chen H, Guo Q, et al. Pancreatic cancer-targeting exosomes for enhancing immunotherapy and reprogramming tumor microenvironment. Biomaterials. 2021;268:120546. doi: 10.1016/j.biomaterials.2020.120546. [DOI] [PubMed] [Google Scholar]
- 103.Zhu D, Liu Z, Li Y, Huang Q, Xia L, Li K. Delivery of manganese carbonyl to the tumor microenvironment using tumor-derived exosomes for cancer gas therapy and low dose radiotherapy. Biomaterials. 2021;274:120894. doi: 10.1016/j.biomaterials.2021.120894. [DOI] [PubMed] [Google Scholar]
- 104.Wang J, Tang W, Yang M, Yin Y, Li H, Hu F, et al. Inflammatory tumor microenvironment responsive neutrophil exosomes-based drug delivery system for targeted glioma therapy. Biomaterials. 2021;273:120784. doi: 10.1016/j.biomaterials.2021.120784. [DOI] [PubMed] [Google Scholar]
- 105.Lima LG, Ham S, Shin H, Chai EPZ, Lek ESH, Lobb RJ, et al. Tumor microenvironmental cytokines bound to cancer exosomes determine uptake by cytokine receptor-expressing cells and biodistribution. Nat Commun. 2021;12(1):3543. doi: 10.1038/s41467-021-23946-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Ferguson Bennit HR, Gonda A, Kabagwira J, Oppegard L, Chi D, Licero Campbell J, et al. Natural killer cell phenotype and functionality affected by exposure to extracellular survivin and lymphoma-derived exosomes. Int J Mol Sci. 2021;22(3):1255. doi: 10.3390/ijms22031255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Markiewski MM, Daugherity E, Reese B, Karbowniczek M. The role of complement in angiogenesis. Antibodies. 2020;9(4):67. doi: 10.3390/antib9040067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Lee JH, Kim C, Um JY, Sethi G, Ahn KS. Casticin-induced inhibition of cell growth and survival are mediated through the dual modulation of Akt/mTOR signaling cascade. Cancers. 2019;11(2):254. doi: 10.3390/cancers11020254. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Padmavathi G, Banik K, Monisha J, Bordoloi D, Shabnam B, Arfuso F, et al. Novel tumor necrosis factor-α induced protein eight (TNFAIP8/TIPE) family: functions and downstream targets involved in cancer progression. Cancer Lett. 2018;432:260–271. doi: 10.1016/j.canlet.2018.06.017. [DOI] [PubMed] [Google Scholar]
- 110.Siveen KS, Ahn KS, Ong TH, Shanmugam MK, Li F, Yap WN, et al. Y-tocotrienol inhibits angiogenesis-dependent growth of human hepatocellular carcinoma through abrogation of AKT/mTOR pathway in an orthotopic mouse model. Oncotarget. 2014;5(7):1897–1911. doi: 10.18632/oncotarget.1876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Flamme I, Breier G, Risau W. Vascular endothelial growth factor (VEGF) and VEGF receptor 2 (flk-1) are expressed during vasculogenesis and vascular differentiation in the quail embryo. Dev Biol. 1995;169(2):699–712. doi: 10.1006/dbio.1995.1180. [DOI] [PubMed] [Google Scholar]
- 112.Risau W, Sariola H, Zerwes HG, Sasse J, Ekblom P, Kemler R, et al. Vasculogenesis and angiogenesis in embryonic-stem-cell-derived embryoid bodies. Development (Cambridge, England) 1988;102(3):471–478. doi: 10.1242/dev.102.3.471. [DOI] [PubMed] [Google Scholar]
- 113.Folkman J. Tumor angiogenesis: therapeutic implications. N Engl J Med. 1971;285(21):1182–1186. doi: 10.1056/NEJM197111182852108. [DOI] [PubMed] [Google Scholar]
- 114.Eelen G, Treps L, Li X, Carmeliet P. Basic and therapeutic aspects of angiogenesis updated. Circ Res. 2020;127(2):310–329. doi: 10.1161/CIRCRESAHA.120.316851. [DOI] [PubMed] [Google Scholar]
- 115.Anbalagan D, Yap G, Yuan Y, Pandey VK, Lau WH, Arora S, et al. Annexin-A1 regulates microRNA-26b* and microRNA-562 to directly target NF-κB and angiogenesis in breast cancer cells. PLoS One. 2014;9(12):e114507. doi: 10.1371/journal.pone.0114507. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Puar YR, Shanmugam MK, Fan L, Arfuso F, Sethi G, Tergaonkar V. Evidence for the involvement of the master transcription factor NF-κB in cancer initiation and progression. Biomedicines. 2018;6(3):82. doi: 10.3390/biomedicines6030082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Eming SA, Martin P, Tomic-Canic M. Wound repair and regeneration: mechanisms, signaling, and translation. Sci Transl Medi. 2014;6(265):265sr6. doi: 10.1126/scitranslmed.3009337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Singh N, Badrun D, Ghatage PJ. State of the art and up-and-coming angiogenesis inhibitors for ovarian cancer. Expert Opin Pharmacother. 2020;21(13):1579–90. doi: 10.1080/14656566.2020.1775813. [DOI] [PubMed] [Google Scholar]
- 119.Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011;144(5):646–74. doi: 10.1016/j.cell.2011.02.013. [DOI] [PubMed] [Google Scholar]
- 120.Wang H, Wang L, Zhou X, Luo X, Liu K, Jiang E, et al. OSCC exosomes regulate miR-210-3p targeting EFNA3 to promote oral cancer angiogenesis through the PI3K/AKT pathway. Biomed Res Int. 2020;2020:2125656. doi: 10.1155/2020/2125656. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Bao L, You B, Shi S, Shan Y, Zhang Q, Yue H, et al. Metastasis-associated miR-23a from nasopharyngeal carcinoma-derived exosomes mediates angiogenesis by repressing a novel target gene TSGA10. Oncogene. 2018;37(21):2873–2889. doi: 10.1038/s41388-018-0183-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Yang Y, Liu Q, Lu J, Adah D, Yu S, Zhao S, et al. Exosomes from Plasmodium-infected hosts inhibit tumor angiogenesis in a murine Lewis lung cancer model. Oncogenesis. 2017;6(6):e351. doi: 10.1038/oncsis.2017.52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Cheng Y, Dai X, Yang T, Zhang N, Liu Z, Jiang Y. Low long noncoding RNA growth arrest-specific transcript 5 expression in the exosomes of lung cancer cells promotes tumor angiogenesis. J Oncol. 2019;2019:2476175. doi: 10.1155/2019/2476175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Fukumura D, Kloepper J, Amoozgar Z, Duda DG, Jain RK. Enhancing cancer immunotherapy using antiangiogenics: opportunities and challenges. Nat Rev Clin Oncol. 2018;15(5):325–340. doi: 10.1038/nrclinonc.2018.29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Jayson GC, Kerbel R, Ellis LM, Harris AL. Antiangiogenic therapy in oncology: current status and future directions. Lancet (London, England) 2016;388(10043):518–529. doi: 10.1016/S0140-6736(15)01088-0. [DOI] [PubMed] [Google Scholar]
- 126.Saharinen P, Eklund L, Alitalo K. Therapeutic targeting of the angiopoietin-TIE pathway. Nat Rev Drug Discov. 2017;16(9):635–661. doi: 10.1038/nrd.2016.278. [DOI] [PubMed] [Google Scholar]
- 127.Akwii RG, Sajib MS, Zahra FT, Mikelis CM. Role of angiopoietin-2 in vascular physiology and pathophysiology. Cells. 2019;8(5):471. doi: 10.3390/cells8050471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Scholz A, Plate KH, Reiss Y. Angiopoietin-2: a multifaceted cytokine that functions in both angiogenesis and inflammation. Ann N Y Acad Sci. 2015;1347:45–51. doi: 10.1111/nyas.12726. [DOI] [PubMed] [Google Scholar]
- 129.Xie JY, Wei JX, Lv LH, Han QF, Yang WB, Li GL, et al. Angiopoietin-2 induces angiogenesis via exosomes in human hepatocellular carcinoma. Cell Commun Signal. 2020;18(1):46. doi: 10.1186/s12964-020-00535-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Kling MJ, Chaturvedi NK, Kesherwani V, Coulter DW, McGuire TR, Sharp JG, et al. Exosomes secreted under hypoxia enhance stemness in Ewing's sarcoma through miR-210 delivery. Oncotarget. 2020;11(40):3633–3645. doi: 10.18632/oncotarget.27702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Sun H, Meng Q, Shi C, Yang H, Li X, Wu S, et al. Hypoxia-inducible exosomes facilitate liver-tropic pre-metastatic niche in colorectal cancer. Hepatology (Baltimore, Md) 2021 doi: 10.1002/hep.32009. [DOI] [PubMed] [Google Scholar]
- 132.Huang Z, Feng Y. Exosomes derived from hypoxic colorectal cancer cells promote angiogenesis through Wnt4-induced β-catenin signaling in endothelial cells. Oncol Res. 2017;25(5):651–661. doi: 10.3727/096504016X14752792816791. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Pan D, Acevedo-Cintrón JA, Sayanagi J, Snyder-Warwick AK, Mackinnon SE, Wood MD. The CCL2/CCR2 axis is critical to recruiting macrophages into acellular nerve allograft bridging a nerve gap to promote angiogenesis and regeneration. Exp Neurol. 2020;331:113363. doi: 10.1016/j.expneurol.2020.113363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Tay H, Du Cheyne C, Demeyere K, De Craene J, De Bels L, Meyer E, et al. Depletion of embryonic macrophages leads to a reduction in angiogenesis in the ex ovo chick chorioallantoic membrane assay. Cells. 2020;10(1):5. doi: 10.3390/cells10010005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Ludwig N, Yerneni SS, Azambuja JH, Gillespie DG, Menshikova EV, Jackson EK, et al. Tumor-derived exosomes promote angiogenesis via adenosine A(2B) receptor signaling. Angiogenesis. 2020;23(4):599–610. doi: 10.1007/s10456-020-09728-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Mao L, Li X, Gong S, Yuan H, Jiang Y, Huang W, et al. Serum exosomes contain ECRG4 mRNA that suppresses tumor growth via inhibition of genes involved in inflammation, cell proliferation, and angiogenesis. Cancer Gene Ther. 2018;25(9–10):248–259. doi: 10.1038/s41417-018-0032-3. [DOI] [PubMed] [Google Scholar]
- 137.Yi H, Ye J, Yang XM, Zhang LW, Zhang ZG, Chen YP. High-grade ovarian cancer secreting effective exosomes in tumor angiogenesis. Int J Clin Exp Pathol. 2015;8(5):5062–5070. [PMC free article] [PubMed] [Google Scholar]
- 138.Horie K, Kawakami K, Fujita Y, Sugaya M, Kameyama K, Mizutani K, et al. Exosomes expressing carbonic anhydrase 9 promote angiogenesis. Biochem Biophys Res Commun. 2017;492(3):356–361. doi: 10.1016/j.bbrc.2017.08.107. [DOI] [PubMed] [Google Scholar]
- 139.Alcayaga-Miranda F, González PL, Lopez-Verrilli A, Varas-Godoy M, Aguila-Díaz C, Contreras L, et al. Prostate tumor-induced angiogenesis is blocked by exosomes derived from menstrual stem cells through the inhibition of reactive oxygen species. Oncotarget. 2016;7(28):44462–44477. doi: 10.18632/oncotarget.9852. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Umezu T, Imanishi S, Azuma K, Kobayashi C, Yoshizawa S, Ohyashiki K, et al. Replenishing exosomes from older bone marrow stromal cells with miR-340 inhibits myeloma-related angiogenesis. Blood Adv. 2017;1(13):812–823. doi: 10.1182/bloodadvances.2016003251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Wang Y, Cen A, Yang Y, Ye H, Li J, Liu S, et al. miR-181a, delivered by hypoxic PTC-secreted exosomes, inhibits DACT2 by downregulating MLL3, leading to YAP-VEGF-mediated angiogenesis. Mol Ther Nucleic Acids. 2021;24:610–621. doi: 10.1016/j.omtn.2021.02.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Dai X, Wang L, Deivasigamni A, Looi CY, Karthikeyan C, Trivedi P, et al. A novel benzimidazole derivative, MBIC inhibits tumor growth and promotes apoptosis via activation of ROS-dependent JNK signaling pathway in hepatocellular carcinoma. Oncotarget. 2017;8(8):12831–12842. doi: 10.18632/oncotarget.14606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Ong PS, Wang LZ, Dai X, Tseng SH, Loo SJ, Sethi G. Judicious toggling of mTOR activity to combat insulin resistance and cancer: current evidence and perspectives. Front Pharmacol. 2016;7:395. doi: 10.3389/fphar.2016.00395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Mohan CD, Bharathkumar H, Bulusu KC, Pandey V, Rangappa S, Fuchs JE, et al. Development of a novel azaspirane that targets the Janus kinase-signal transducer and activator of transcription (STAT) pathway in hepatocellular carcinoma in vitro and in vivo. J Biol Chem. 2014;289(49):34296–34307. doi: 10.1074/jbc.M114.601104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Manu KA, Shanmugam MK, Li F, Chen L, Siveen KS, Ahn KS, et al. Simvastatin sensitizes human gastric cancer xenograft in nude mice to capecitabine by suppressing nuclear factor-kappa B-regulated gene products. J Mol Med (Berl) 2014;92(3):267–276. doi: 10.1007/s00109-013-1095-0. [DOI] [PubMed] [Google Scholar]
- 146.Tan SC. Low penetrance genetic polymorphisms as potential biomarkers for colorectal cancer predisposition. J Gene Med. 2018;20(4):e3010. doi: 10.1002/jgm.3010. [DOI] [PubMed] [Google Scholar]
- 147.Jeong H, Kim S, Hong BJ, Lee CJ, Kim YE, Bok S, et al. Tumor-associated macrophages enhance tumor hypoxia and aerobic glycolysis. Can Res. 2019;79(4):795–806. doi: 10.1158/0008-5472.CAN-18-2545. [DOI] [PubMed] [Google Scholar]
- 148.Nie H, Ju H, Fan J, Shi X, Cheng Y, Cang X, et al. O-GlcNAcylation of PGK1 coordinates glycolysis and TCA cycle to promote tumor growth. Nat Commun. 2020;11(1):36. doi: 10.1038/s41467-019-13601-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Deng F, Zhou R, Lin C, Yang S, Wang H, Li W, et al. Tumor-secreted dickkopf2 accelerates aerobic glycolysis and promotes angiogenesis in colorectal cancer. Theranostics. 2019;9(4):1001–1014. doi: 10.7150/thno.30056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Liang Y, Wang H, Chen B, Mao Q, Xia W, Zhang T, et al. circDCUN1D4 suppresses tumor metastasis and glycolysis in lung adenocarcinoma by stabilizing TXNIP expression. Mol Ther Nucleic Acids. 2021;23:355–368. doi: 10.1016/j.omtn.2020.11.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Wang H, Wang L, Pan H, Wang Y, Shi M, Yu H, et al. Exosomes derived from macrophages enhance aerobic glycolysis and chemoresistance in lung cancer by stabilizing c-Myc via the inhibition of NEDD4L. Front Cell Dev Biol. 2020;8:620603. doi: 10.3389/fcell.2020.620603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Wang B, Wang X, Hou D, Huang Q, Zhan W, Chen C, et al. Exosomes derived from acute myeloid leukemia cells promote chemoresistance by enhancing glycolysis-mediated vascular remodeling. J Cell Physiol. 2019;234(7):10602–10614. doi: 10.1002/jcp.27735. [DOI] [PubMed] [Google Scholar]
- 153.Lee JH, Kim C, Baek SH, Ko JH, Lee SG, Yang WM, et al. Capsazepine inhibits JAK/STAT3 signaling, tumor growth, and cell survival in prostate cancer. Oncotarget. 2017;8(11):17700–17711. doi: 10.18632/oncotarget.10775. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Liu Y, Liao S, Bennett S, Tang H, Song D, Wood D, et al. STAT3 and its targeting inhibitors in osteosarcoma. Cell Prolif. 2021;54(2):e12974. doi: 10.1111/cpr.12974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Wang H, Tao Z, Feng M, Li X, Deng Z, Zhao G, et al. Dual PLK1 and STAT3 inhibition promotes glioblastoma cells apoptosis through MYC. Biochem Biophys Res Commun. 2020;533(3):368–375. doi: 10.1016/j.bbrc.2020.09.008. [DOI] [PubMed] [Google Scholar]
- 156.Xu Y, Zhu Y, Yue Y, Pu S, Wu J, Lv Y, et al. Tamoxifen attenuates reactive astrocyte-induced brain metastasis and drug resistance through the IL-6/STAT3 signaling pathway. Acta Biochim Biophys Sin. 2020;52(12):1299–1305. doi: 10.1093/abbs/gmaa127. [DOI] [PubMed] [Google Scholar]
- 157.Cong Y, Cui Y, Zhu S, Cao J, Zou H, Martin TA, et al. Tim-3 promotes cell aggressiveness and paclitaxel resistance through NF-κB/STAT3 signalling pathway in breast cancer cells. Chin J Cancer Res. 2020;32(5):564–79. doi: 10.21147/j.issn.1000-9604.2020.05.02. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Jiang Y, Chen P, Hu K, Dai G, Li J, Zheng D, et al. Inflammatory microenvironment of fibrotic liver promotes hepatocellular carcinoma growth, metastasis and sorafenib resistance through STAT3 activation. J Cell Mol Med. 2021;25(3):1568–1582. doi: 10.1111/jcmm.16256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Ham S, Lima LG, Chai EPZ, Muller A, Lobb RJ, Krumeich S, et al. Breast cancer-derived exosomes alter macrophage polarization via gp130/STAT3 signaling. Front Immunol. 2018;9:871. doi: 10.3389/fimmu.2018.00871. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Zhou J, Li X, Wu X, Zhang T, Zhu Q, Wang X, et al. Exosomes released from tumor-associated macrophages transfer miRNAs that induce a Treg/Th17 cell imbalance in epithelial ovarian cancer. Cancer Immunol Res. 2018;6(12):1578–1592. doi: 10.1158/2326-6066.CIR-17-0479. [DOI] [PubMed] [Google Scholar]
- 161.Yu X, Zhang Q, Zhang X, Han Q, Li H, Mao Y, et al. Exosomes from macrophages exposed to apoptotic breast cancer cells promote breast cancer proliferation and metastasis. J Cancer. 2019;10(13):2892–2906. doi: 10.7150/jca.31241. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Ren R, Sun H, Ma C, Liu J, Wang H. Colon cancer cells secrete exosomes to promote self-proliferation by shortening mitosis duration and activation of STAT3 in a hypoxic environment. Cell Biosci. 2019;9:62. doi: 10.1186/s13578-019-0325-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.He C, Hua W, Liu J, Fan L, Wang H, Sun G. Exosomes derived from endoplasmic reticulum-stressed liver cancer cells enhance the expression of cytokines in macrophages via the STAT3 signaling pathway. Oncol Lett. 2020;20(1):589–600. doi: 10.3892/ol.2020.11609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Xing H, Liang C, Xu X, Sun H, Ma X, Jiang Z. Mesenchymal stroma/stem-like cells of GARP knockdown inhibits cell proliferation and invasion of mouse colon cancer cells (MC38) through exosomes. J Cell Mol Med. 2020;24(23):13984–13990. doi: 10.1111/jcmm.16008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Dorayappan KDP, Gardner ML, Hisey CL, Zingarelli RA, Smith BQ, Lightfoot MDS, et al. A microfluidic chip enables isolation of exosomes and establishment of their protein profiles and associated signaling pathways in ovarian cancer. Can Res. 2019;79(13):3503–3513. doi: 10.1158/0008-5472.CAN-18-3538. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Tan SC, Ankathil R. Genetic susceptibility to cervical cancer: role of common polymorphisms in apoptosis-related genes. Tumour Biol J Int Soc Oncodev Biol Med. 2015;36(9):6633–6644. doi: 10.1007/s13277-015-3868-2. [DOI] [PubMed] [Google Scholar]
- 167.Hu JL, Wang W, Lan XL, Zeng ZC, Liang YS, Yan YR, et al. CAFs secreted exosomes promote metastasis and chemotherapy resistance by enhancing cell stemness and epithelial-mesenchymal transition in colorectal cancer. Mol Cancer. 2019;18(1):91. doi: 10.1186/s12943-019-1019-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Kim C, Lee SG, Yang WM, Arfuso F, Um JY, Kumar AP, et al. Formononetin-induced oxidative stress abrogates the activation of STAT3/5 signaling axis and suppresses the tumor growth in multiple myeloma preclinical model. Cancer Lett. 2018;431:123–141. doi: 10.1016/j.canlet.2018.05.038. [DOI] [PubMed] [Google Scholar]
- 169.Kirtonia A, Sethi G, Garg M. The multifaceted role of reactive oxygen species in tumorigenesis. Cell Mol Life Sci CMLS. 2020;77(22):4459–4483. doi: 10.1007/s00018-020-03536-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Lee JH, Kim C, Lee SG, Sethi G, Ahn KS. Ophiopogonin D, a steroidal glycoside abrogates STAT3 signaling cascade and exhibits anti-cancer activity by causing GSH/GSSG imbalance in lung carcinoma. Cancers. 2018;10(11):427. doi: 10.3390/cancers10110427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Ko JH, Lee JH, Jung SH, Lee SG, Chinnathambi A, Alharbi SA, et al. 2,5-Dihydroxyacetophenone induces apoptosis of multiple myeloma cells by regulating the MAPK activation pathway. Molecules (Basel, Switzerland) 2017;22(7):1157. doi: 10.3390/molecules22071157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Patel GK, Khan MA, Bhardwaj A, Srivastava SK, Zubair H, Patton MC, et al. Exosomes confer chemoresistance to pancreatic cancer cells by promoting ROS detoxification and miR-155-mediated suppression of key gemcitabine-metabolising enzyme. DCK Br J Cancer. 2017;116(5):609–619. doi: 10.1038/bjc.2017.18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.Subramaniam A, Loo SY, Rajendran P, Manu KA, Perumal E, Li F, et al. An anthraquinone derivative, emodin sensitizes hepatocellular carcinoma cells to TRAIL induced apoptosis through the induction of death receptors and downregulation of cell survival proteins. Apoptosis Int J Progr Cell Death. 2013;18(10):1175–1187. doi: 10.1007/s10495-013-0851-5. [DOI] [PubMed] [Google Scholar]
- 174.Shen T, Huang Z, Shi C, Pu X, Xu X, Wu Z, et al. Pancreatic cancer-derived exosomes induce apoptosis of T lymphocytes through the p38 MAPK-mediated endoplasmic reticulum stress. FASEB J Off Publ Federation Am Soc Exp Biol. 2020;34(6):8442–8458. doi: 10.1096/fj.201902186R. [DOI] [PubMed] [Google Scholar]
- 175.Wang B, Wang Y, Yan Z, Sun Y, Su C. Colorectal cancer cell-derived exosomes promote proliferation and decrease apoptosis by activating the ERK pathway. Int J Clin Exp Pathol. 2019;12(7):2485–2495. [PMC free article] [PubMed] [Google Scholar]
- 176.Deng S, Shanmugam MK, Kumar AP, Yap CT, Sethi G, Bishayee A. Targeting autophagy using natural compounds for cancer prevention and therapy. Cancer. 2019;125(8):1228–1246. doi: 10.1002/cncr.31978. [DOI] [PubMed] [Google Scholar]
- 177.Patra S, Mishra SR, Behera BP, Mahapatra KK, Panigrahi DP, Bhol CS, et al. Autophagy-modulating phytochemicals in cancer therapeutics: Current evidences and future perspectives. Semin Cancer Biol. 2020 doi: 10.1016/j.semcancer.2020.05.008. [DOI] [PubMed] [Google Scholar]
- 178.Hwang ST, Kim C, Lee JH, Chinnathambi A, Alharbi SA, Shair OHM, et al. Cycloastragenol can negate constitutive STAT3 activation and promote paclitaxel-induced apoptosis in human gastric cancer cells. Phytomed Int J Phytother Phytopharmacol. 2019;59:152907. doi: 10.1016/j.phymed.2019.152907. [DOI] [PubMed] [Google Scholar]
- 179.Praharaj PP, Naik PP, Panigrahi DP, Bhol CS, Mahapatra KK, Patra S, et al. Intricate role of mitochondrial lipid in mitophagy and mitochondrial apoptosis: its implication in cancer therapeutics. Cell Mol Life Sci CMLS. 2019;76(9):1641–1652. doi: 10.1007/s00018-018-2990-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Wang L, Xu P, Xie X, Hu F, Jiang L, Hu R, et al. Down regulation of SIRT2 reduced ASS induced NSCLC apoptosis through the release of autophagy components via exosomes. Front Cell Dev Biol. 2020;8:601953. doi: 10.3389/fcell.2020.601953. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181.Zhang X, Shi H, Yuan X, Jiang P, Qian H, Xu W. Tumor-derived exosomes induce N2 polarization of neutrophils to promote gastric cancer cell migration. Mol Cancer. 2018;17(1):146. doi: 10.1186/s12943-018-0898-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 182.Yang P, Cao X, Cai H, Feng P, Chen X, Zhu Y, et al. The exosomes derived from CAR-T cell efficiently target mesothelin and reduce triple-negative breast cancer growth. Cell Immunol. 2021;360:104262. doi: 10.1016/j.cellimm.2020.104262. [DOI] [PubMed] [Google Scholar]
- 183.Wang S, Su X, Xu M, Xiao X, Li X, Li H, et al. Exosomes secreted by mesenchymal stromal/stem cell-derived adipocytes promote breast cancer cell growth via activation of Hippo signaling pathway. Stem Cell Res Ther. 2019;10(1):117. doi: 10.1186/s13287-019-1220-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 184.Lu Z, Zuo B, Jing R, Gao X, Rao Q, Liu Z, et al. Dendritic cell-derived exosomes elicit tumor regression in autochthonous hepatocellular carcinoma mouse models. J Hepatol. 2017;67(4):739–748. doi: 10.1016/j.jhep.2017.05.019. [DOI] [PubMed] [Google Scholar]
- 185.Li H, Chi X, Li R, Ouyang J, Chen Y. HIV-1-infected cell-derived exosomes promote the growth and progression of cervical cancer. Int J Biol Sci. 2019;15(11):2438–2447. doi: 10.7150/ijbs.38146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 186.Shanmugam MK, Ahn KS, Hsu A, Woo CC, Yuan Y, Tan KHB, et al. Thymoquinone inhibits bone metastasis of breast cancer cells through abrogation of the CXCR4 signaling axis. Front Pharmacol. 2018;9:1294. doi: 10.3389/fphar.2018.01294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 187.Chua AW, Hay HS, Rajendran P, Shanmugam MK, Li F, Bist P, et al. Butein downregulates chemokine receptor CXCR4 expression and function through suppression of NF-κB activation in breast and pancreatic tumor cells. Biochem Pharmacol. 2010;80(10):1553–1562. doi: 10.1016/j.bcp.2010.07.045. [DOI] [PubMed] [Google Scholar]
- 188.Shanmugam MK, Ong TH, Kumar AP, Lun CK, Ho PC, Wong PT, et al. Ursolic acid inhibits the initiation, progression of prostate cancer and prolongs the survival of TRAMP mice by modulating pro-inflammatory pathways. PLoS One. 2012;7(3):e32476. doi: 10.1371/journal.pone.0032476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189.Feng W, Dean DC, Hornicek FJ, Shi H, Duan Z. Exosomes promote pre-metastatic niche formation in ovarian cancer. Mol Cancer. 2019;18(1):124. doi: 10.1186/s12943-019-1049-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 190.Yoshida K, Tsuda M, Matsumoto R, Semba S, Wang L, Sugino H, et al. Exosomes containing ErbB2/CRK induce vascular growth in premetastatic niches and promote metastasis of bladder cancer. Cancer Sci. 2019;110(7):2119–2132. doi: 10.1111/cas.14080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 191.Wang H, Wei H, Wang J, Li L, Chen A, Li Z. MicroRNA-181d-5p-containing exosomes derived from CAFs promote EMT by regulating CDX2/HOXA5 in breast cancer. Mol Ther Nucleic Acids. 2020;19:654–667. doi: 10.1016/j.omtn.2019.11.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192.Maziveyi M, Dong S, Baranwal S, Mehrnezhad A, Rathinam R, Huckaba TM, et al. Exosomes from nischarin-expressing cells reduce breast cancer cell motility and tumor growth. Can Res. 2019;79(9):2152–2166. doi: 10.1158/0008-5472.CAN-18-0842. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 193.Ghoroghi S, Mary B, Larnicol A, Asokan N, Klein A, Osmani N, et al. Ral GTPases promote breast cancer metastasis by controlling biogenesis and organ targeting of exosomes. eLife. 2021 doi: 10.7554/eLife.61539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194.Wei Q, Wei L, Zhang J, Li Z, Feng H, Ren L. EphA2-enriched exosomes promote cell migration and are a potential diagnostic serum marker in pancreatic cancer. Mol Med Rep. 2020;22(4):2941–2947. doi: 10.3892/mmr.2020.11384. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 195.Xu S, Zheng L, Kang L, Xu H, Gao L. microRNA-let-7e in serum-derived exosomes inhibits the metastasis of non-small-cell lung cancer in a SUV39H2/LSD1/CDH1-dependent manner. Cancer Gene Ther. 2021;28(3–4):250–264. doi: 10.1038/s41417-020-00216-1. [DOI] [PubMed] [Google Scholar]
- 196.Hwang ST, Yang MH, Kumar AP, Sethi G, Ahn KS. Corilagin represses epithelial to mesenchymal transition process through modulating Wnt/β-catenin signaling cascade. Biomolecules. 2020;10(10):1406. doi: 10.3390/biom10101406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 197.Yang MH, Lee JH, Ko JH, Jung SH, Sethi G, Ahn KS. Brassinin represses invasive potential of lung carcinoma cells through deactivation of PI3K/Akt/mTOR signaling cascade. Molecules (Basel, Switzerland) 2019;24(8):1584. doi: 10.3390/molecules24081584. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 198.Ko JH, Nam D, Um JY, Jung SH, Sethi G, Ahn KS. Bergamottin suppresses metastasis of lung cancer cells through abrogation of diverse oncogenic signaling cascades and epithelial-to-mesenchymal transition. Molecules (Basel, Switzerland) 2018;23(7):1601. doi: 10.3390/molecules23071601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 199.Dai X, Ahn KS, Wang LZ, Kim C, Deivasigamni A, Arfuso F, et al. Ascochlorin enhances the sensitivity of doxorubicin leading to the reversal of epithelial-to-mesenchymal transition in hepatocellular carcinoma. Mol Cancer Ther. 2016;15(12):2966–2976. doi: 10.1158/1535-7163.MCT-16-0391. [DOI] [PubMed] [Google Scholar]
- 200.Ong MS, Deng S, Halim CE, Cai W, Tan TZ, Huang RY, et al. Cytoskeletal proteins in cancer and intracellular stress: a therapeutic perspective. Cancers. 2020;12(1):238. doi: 10.3390/cancers12010238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 201.Gaballa R, Ali HEA, Mahmoud MO, Rhim JS, Ali HI, Salem HF, et al. Exosomes-mediated transfer of Itga2 promotes migration and invasion of prostate cancer cells by inducing epithelial-mesenchymal transition. Cancers. 2020;12(8):2300. doi: 10.3390/cancers12082300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 202.Liu XN, Zhang CB, Lin H, Tang XY, Zhou R, Wen HL, et al. microRNA-204 shuttled by mesenchymal stem cell-derived exosomes inhibits the migration and invasion of non-small-cell lung cancer cells via the KLF7/AKT/HIF-1α axis. Neoplasma. 2021;68(4):719–731. doi: 10.4149/neo_2021_201208N1328. [DOI] [PubMed] [Google Scholar]
- 203.Cai J, Gong L, Li G, Guo J, Yi X, Wang Z. Exosomes in ovarian cancer ascites promote epithelial-mesenchymal transition of ovarian cancer cells by delivery of miR-6780b-5p. Cell Death Dis. 2021;12(2):210. doi: 10.1038/s41419-021-03490-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 204.Shojaei S, Hashemi SM, Ghanbarian H, Sharifi K, Salehi M, Mohammadi-Yeganeh S. Delivery of miR-381-3p mimic by mesenchymal stem cell-derived exosomes inhibits triple negative breast cancer aggressiveness; an in vitro study. Stem Cell Rev Rep. 2021;17(3):1027–1038. doi: 10.1007/s12015-020-10089-4. [DOI] [PubMed] [Google Scholar]
- 205.Scavo MP, Rizzi F, Depalo N, Fanizza E, Ingrosso C, Curri ML, et al. A possible role of FZD10 delivering exosomes derived from colon cancers cell lines in inducing activation of epithelial-mesenchymal transition in normal colon epithelial cell line. Int J Mol Sci. 2020;21(18):6705. doi: 10.3390/ijms21186705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 206.Zhou J, Wang H, Sun Q, Liu X, Wu Z, Wang X, et al. miR-224-5p-enriched exosomes promote tumorigenesis by directly targeting androgen receptor in non-small cell lung cancer. Mol Ther Nucleic Acids. 2021;23:1217–1228. doi: 10.1016/j.omtn.2021.01.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 207.Monisha J, Roy NK, Padmavathi G, Banik K, Bordoloi D, Khwairakpam AD, et al. NGAL is downregulated in oral squamous cell carcinoma and leads to increased survival, proliferation, migration and chemoresistance. Cancers. 2018;10(7):228. doi: 10.3390/cancers10070228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 208.Ko JH, Um JY, Lee SG, Yang WM, Sethi G, Ahn KS. Conditioned media from adipocytes promote proliferation, migration, and invasion in melanoma and colorectal cancer cells. J Cell Physiol. 2019;234(10):18249–18261. doi: 10.1002/jcp.28456. [DOI] [PubMed] [Google Scholar]
- 209.Kothapalli R, Sivaraman Siveen K, Tan TZ, Thiery JP, Kumar AP, Sethi G, et al. Functional characterization of selective exosite-binding inhibitors of matrix metalloproteinase-13 (MMP-13) - experimental validation in human breast and colon cancer. Biosci Biotechnol Biochem. 2016;80(11):2122–2131. doi: 10.1080/09168451.2016.1200456. [DOI] [PubMed] [Google Scholar]
- 210.Lee H, Baek SH, Lee JH, Kim C, Ko JH, Lee SG, et al. Isorhynchophylline, a potent plant alkaloid, induces apoptotic and anti-metastatic effects in human hepatocellular carcinoma cells through the modulation of diverse cell signaling cascades. Int J Mol Sci. 2017;18(5):1095. doi: 10.3390/ijms18051095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 211.Jung YY, Lee JH, Nam D, Narula AS, Namjoshi OA, Blough BE, et al. Anti-myeloma effects of icariin are mediated through the attenuation of JAK/STAT3-dependent signaling cascade. Front Pharmacol. 2018;9:531. doi: 10.3389/fphar.2018.00531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 212.Sáenz-de-Santa-María I, Celada L, San José Martínez A, Cubiella T, Chiara MD. Blockage of squamous cancer cell collective invasion by FAK inhibition is released by CAFs and MMP-2. Cancers. 2020;12(12):3708. doi: 10.3390/cancers12123708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 213.Szarvas T, Hoffmann MJ, Olah C, Szekely E, Kiss A, Hess J, et al. MMP-7 serum and tissue levels are associated with poor survival in platinum-treated bladder cancer patients. Diagnostics (Basel, Switzerland) 2020;11(1):48. doi: 10.3390/diagnostics11010048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 214.Frieling JS, Li T, Tauro M, Lynch CC. Prostate cancer-derived MMP-3 controls intrinsic cell growth and extrinsic angiogenesis. Neoplasia (New York, NY) 2020;22(10):511–521. doi: 10.1016/j.neo.2020.08.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 215.Che Y, Shi X, Shi Y, Jiang X, Ai Q, Shi Y, et al. Exosomes derived from miR-143-overexpressing MSCs inhibit cell migration and invasion in human prostate cancer by downregulating TFF3. Mol Ther Nucleic Acids. 2019;18:232–244. doi: 10.1016/j.omtn.2019.08.010. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 216.Chen G, Zhang Y, Wu X. 786–0 renal cancer cell line-derived exosomes promote 786–0 cell migration and invasion in vitro. Oncol Lett. 2014;7(5):1576–1580. doi: 10.3892/ol.2014.1962. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 217.Wu F, Li F, Lin X, Xu F, Cui RR, Zhong JY, et al. Exosomes increased angiogenesis in papillary thyroid cancer microenvironment. Endocr Relat Cancer. 2019;26(5):525–538. doi: 10.1530/ERC-19-0008. [DOI] [PubMed] [Google Scholar]
- 218.Mao Y, Wang Y, Dong L, Zhang Y, Zhang Y, Wang C, et al. Hypoxic exosomes facilitate angiogenesis and metastasis in esophageal squamous cell carcinoma through altering the phenotype and transcriptome of endothelial cells. J Exp Clin Cancer Res CR. 2019;38(1):389. doi: 10.1186/s13046-019-1384-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 219.Ludwig N, Yerneni SS, Razzo BM, Whiteside TL. Exosomes from HNSCC promote angiogenesis through reprogramming of endothelial cells. Mol Cancer Res MCR. 2018;16(11):1798–1808. doi: 10.1158/1541-7786.MCR-18-0358. [DOI] [PubMed] [Google Scholar]
- 220.Zhou Z, Zhang H, Deng T, Ning T, Liu R, Liu D, et al. Exosomes carrying MicroRNA-155 target forkhead box O3 of endothelial cells and promote angiogenesis in gastric cancer. Mol Ther Oncolytics. 2019;15:223–233. doi: 10.1016/j.omto.2019.10.006. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 221.Xue X, Huang J, Yu K, Chen X, He Y, Qi D, et al. YB-1 transferred by gastric cancer exosomes promotes angiogenesis via enhancing the expression of angiogenic factors in vascular endothelial cells. BMC Cancer. 2020;20(1):996. doi: 10.1186/s12885-020-07509-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 222.Chen Z, Xie Y, Chen W, Li T, Chen X, Liu B. microRNA-6785-5p-loaded human umbilical cord mesenchymal stem cells-derived exosomes suppress angiogenesis and metastasis in gastric cancer via INHBA. Life Sci. 2021;284:119222. doi: 10.1016/j.lfs.2021.119222. [DOI] [PubMed] [Google Scholar]
- 223.Bai M, Li J, Yang H, Zhang H, Zhou Z, Deng T, et al. miR-135b delivered by gastric tumor exosomes inhibits FOXO1 expression in endothelial cells and promotes angiogenesis. Mol Ther J Am Soc Gene Ther. 2019;27(10):1772–1783. doi: 10.1016/j.ymthe.2019.06.018. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 224.Song Y, Wang M, Tong H, Tan Y, Hu X, Wang K, et al. Plasma exosomes from endometrial cancer patients contain LGALS3BP to promote endometrial cancer progression. Oncogene. 2021;40(3):633–646. doi: 10.1038/s41388-020-01555-x. [DOI] [PubMed] [Google Scholar]
- 225.Pakravan K, Babashah S, Sadeghizadeh M, Mowla SJ, Mossahebi-Mohammadi M, Ataei F, et al. MicroRNA-100 shuttled by mesenchymal stem cell-derived exosomes suppresses in vitro angiogenesis through modulating the mTOR/HIF-1α/VEGF signaling axis in breast cancer cells. Cell Oncol (Dordr) 2017;40(5):457–470. doi: 10.1007/s13402-017-0335-7. [DOI] [PubMed] [Google Scholar]
- 226.Lee JK, Park SR, Jung BK, Jeon YK, Lee YS, Kim MK, et al. Exosomes derived from mesenchymal stem cells suppress angiogenesis by down-regulating VEGF expression in breast cancer cells. PLoS One. 2013;8(12):e84256. doi: 10.1371/journal.pone.0084256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 227.Zhang X, Sheng Y, Li B, Wang Q, Liu X, Han J. Ovarian cancer derived PKR1 positive exosomes promote angiogenesis by promoting migration and tube formation in vitro. Cell Biochem Funct. 2021;39(2):308–316. doi: 10.1002/cbf.3583. [DOI] [PubMed] [Google Scholar]
- 228.Li Z, Yan-Qing W, Xiao Y, Shi-Yi L, Meng-Qin Y, Shu X, et al. Exosomes secreted by chemoresistant ovarian cancer cells promote angiogenesis. J Ovarian Res. 2021;14(1):7. doi: 10.1186/s13048-020-00758-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 229.Zheng X, Ma N, Wang X, Hu J, Ma X, Wang J, et al. Exosomes derived from 5-fluorouracil-resistant colon cancer cells are enriched in GDF15 and can promote angiogenesis. J Cancer. 2020;11(24):7116–7126. doi: 10.7150/jca.49224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 230.Yin H, Yu S, Xie Y, Dai X, Dong M, Sheng C, et al. Cancer-associated fibroblasts-derived exosomes upregulate microRNA-135b-5p to promote colorectal cancer cell growth and angiogenesis by inhibiting thioredoxin-interacting protein. Cell Signal. 2021;84:110029. doi: 10.1016/j.cellsig.2021.110029. [DOI] [PubMed] [Google Scholar]
- 231.Zhang L, Wu X, Luo C, Chen X, Yang L, Tao J, et al. The 786–0 renal cancer cell-derived exosomes promote angiogenesis by downregulating the expression of hepatocyte cell adhesion molecule. Mol Med Rep. 2013;8(1):272–276. doi: 10.3892/mmr.2013.1458. [DOI] [PubMed] [Google Scholar]
- 232.Hou Y, Fan L, Li H. Oncogenic miR-27a delivered by exosomes binds to SFRP1 and promotes angiogenesis in renal clear cell carcinoma. Mol Ther Nucleic Acids. 2021;24:92–103. doi: 10.1016/j.omtn.2020.11.019. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 233.Song H, Liu D, Dong S, Zeng L, Wu Z, Zhao P, et al. Epitranscriptomics and epiproteomics in cancer drug resistance: therapeutic implications. Signal Transduct Target Ther. 2020;5(1):193. doi: 10.1038/s41392-020-00300-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 234.Asano T. Drug resistance in cancer therapy and the role of epigenetics. J Nippon Med Scl. 2020;87(5):244–51. doi: 10.1272/jnms.JNMS.2020_87-508. [DOI] [PubMed] [Google Scholar]
- 235.Haider T, Pandey V, Banjare N, Gupta PN, Soni V. Drug resistance in cancer: mechanisms and tackling strategies. Pharmacol Rep. 2020;72(5):1125–1151. doi: 10.1007/s43440-020-00138-7. [DOI] [PubMed] [Google Scholar]
- 236.Holohan C, Van Schaeybroeck S, Longley DB, Johnston PG. Cancer drug resistance: an evolving paradigm. Nat Rev Cancer. 2013;13(10):714–726. doi: 10.1038/nrc3599. [DOI] [PubMed] [Google Scholar]
- 237.Kansara S, Pandey V, Lobie PE, Sethi G, Garg M, Pandey AK. Mechanistic involvement of long non-coding RNAs in oncotherapeutics resistance in triple-negative breast cancer. Cells. 2020;9(6):1511. doi: 10.3390/cells9061511. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 238.Li F, Sethi G. Targeting transcription factor NF-kappaB to overcome chemoresistance and radioresistance in cancer therapy. Biochem Biophys Acta. 2010;1805(2):167–180. doi: 10.1016/j.bbcan.2010.01.002. [DOI] [PubMed] [Google Scholar]
- 239.Aggarwal V, Tuli HS, Thakral F, Singhal P, Aggarwal D, Srivastava S, et al. Molecular mechanisms of action of hesperidin in cancer: recent trends and advancements. Exp Biol Med (Maywood) 2020;245(5):486–497. doi: 10.1177/1535370220903671. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 240.Hon KW, Ab-Mutalib NS, et al. Extracellular vesicle-derived circular RNAs confers chemoresistance in colorectal cancer. Sci Rep. 2019;9(1):16497. doi: 10.1038/s41598-019-53063-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 241.Liang G, Zhu Y, Ali DJ, Tian T, Xu H, Si K, et al. Engineered exosomes for targeted co-delivery of miR-21 inhibitor and chemotherapeutics to reverse drug resistance in colon cancer. J Nanobiotechnol. 2020;18(1):10. doi: 10.1186/s12951-019-0563-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 242.Zhu X, Shen H, Yin X, Yang M, Wei H, Chen Q, et al. Macrophages derived exosomes deliver miR-223 to epithelial ovarian cancer cells to elicit a chemoresistant phenotype. J Exp Clin Cancer Res. 2019;38(1):81. doi: 10.1186/s13046-019-1095-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 243.Binenbaum Y, Fridman E, Yaari Z, Milman N, Schroeder A, Ben David G, et al. Transfer of miRNA in macrophage-derived exosomes induces drug resistance in pancreatic adenocarcinoma. Can Res. 2018;78(18):5287–5299. doi: 10.1158/0008-5472.CAN-18-0124. [DOI] [PubMed] [Google Scholar]
- 244.Yang Z, Zhao N, Cui J, Wu H, Xiong J, Peng T. Exosomes derived from cancer stem cells of gemcitabine-resistant pancreatic cancer cells enhance drug resistance by delivering miR-210. Cell Oncol (Dordr) 2020;43(1):123–136. doi: 10.1007/s13402-019-00476-6. [DOI] [PubMed] [Google Scholar]
- 245.Dorayappan KDP, Wanner R, Wallbillich JJ, Saini U, Zingarelli R, Suarez AA, et al. Hypoxia-induced exosomes contribute to a more aggressive and chemoresistant ovarian cancer phenotype: a novel mechanism linking STAT3/Rab proteins. Oncogene. 2018;37(28):3806–3821. doi: 10.1038/s41388-018-0189-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 246.Qin X, Yu S, Zhou L, Shi M, Hu Y, Xu X, et al. Cisplatin-resistant lung cancer cell-derived exosomes increase cisplatin resistance of recipient cells in exosomal miR-100-5p-dependent manner. Int J Nanomed. 2017;12:3721–3733. doi: 10.2147/IJN.S131516. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 247.Huang L, Hu C, Chao H, Zhang Y, Li Y, Hou J, et al. Drug-resistant endothelial cells facilitate progression, EMT and chemoresistance in nasopharyngeal carcinoma via exosomes. Cell Signal. 2019;63:109385. doi: 10.1016/j.cellsig.2019.109385. [DOI] [PubMed] [Google Scholar]
- 248.Wang X, Zhang H, Bai M, Ning T, Ge S, Deng T, et al. Exosomes serve as nanoparticles to deliver anti-miR-214 to reverse chemoresistance to cisplatin in gastric cancer. Mol Ther J Am Soc Gene Ther. 2018;26(3):774–783. doi: 10.1016/j.ymthe.2018.01.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 249.Zhang X, Yang Y, Yang Y, Chen H, Tu H, Li J. Exosomes from bone marrow microenvironment-derived mesenchymal stem cells affect CML cells growth and promote drug resistance to tyrosine kinase inhibitors. Stem Cells Int. 2020;2020:8890201. doi: 10.1155/2020/8890201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 250.Sadegh-Nejadi S, Afrisham R, Emamgholipour S, Izadi P, Eivazi N, Tahbazlahafi B, et al. Influence of plasma circulating exosomes obtained from obese women on tumorigenesis and tamoxifen resistance in MCF-7 cells. IUBMB Life. 2020;72(9):1930–1940. doi: 10.1002/iub.2305. [DOI] [PubMed] [Google Scholar]
- 251.Liu J, Zhu S, Tang W, Huang Q, Mei Y, Yang H. Exosomes from tamoxifen-resistant breast cancer cells transmit drug resistance partly by delivering miR-9-5p. Cancer Cell Int. 2021;21(1):55. doi: 10.1186/s12935-020-01659-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 252.Deng X, Ruan H, Zhang X, Xu X, Zhu Y, Peng H, et al. Long noncoding RNA CCAL transferred from fibroblasts by exosomes promotes chemoresistance of colorectal cancer cells. Int J Cancer. 2020;146(6):1700–1716. doi: 10.1002/ijc.32608. [DOI] [PubMed] [Google Scholar]
- 253.Qu Z, Wu J, Wu J, Luo D, Jiang C, Ding Y. Exosomes derived from HCC cells induce sorafenib resistance in hepatocellular carcinoma both in vivo and in vitro. J Exp Clin Cancer Res. 2016;35(1):159. doi: 10.1186/s13046-016-0430-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 254.Liu X, Jiang T, Li X, Zhao C, Li J, Zhou F, et al. Exosomes transmit T790M mutation-induced resistance in EGFR-mutant NSCLC by activating PI3K/AKT signalling pathway. J Cell Mol Med. 2020;24(2):1529–1540. doi: 10.1111/jcmm.14838. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 255.Shi S, Huang X, Ma X, Zhu X, Zhang Q. Research of the mechanism on miRNA193 in exosomes promotes cisplatin resistance in esophageal cancer cells. PLoS One. 2020;15(5):e0225290. doi: 10.1371/journal.pone.0225290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 256.Kunou S, Shimada K, Takai M, Sakamoto A, Aoki T, Hikita T, et al. Exosomes secreted from cancer-associated fibroblasts elicit anti-pyrimidine drug resistance through modulation of its transporter in malignant lymphoma. Oncogene. 2021;40(23):3989–4003. doi: 10.1038/s41388-021-01829-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 257.Zhang S, Zhang Y, Qu J, Che X, Fan Y, Hou K, et al. Exosomes promote cetuximab resistance via the PTEN/Akt pathway in colon cancer cells. Braz J Med Biol Res. 2017;51(1):e6472. doi: 10.1590/1414-431X20176472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 258.Pan Y, Lin Y, Mi C. Cisplatin-resistant osteosarcoma cell-derived exosomes confer cisplatin resistance to recipient cells in an exosomal circ_103801-dependent manner. Cell Biol Int. 2021;45(4):858–868. doi: 10.1002/cbin.11532. [DOI] [PubMed] [Google Scholar]
- 259.Chang WH, Nguyen TT, Hsu CH, Bryant KL, Kim HJ, Ying H, et al. KRAS-dependent cancer cells promote survival by producing exosomes enriched in Survivin. Cancer Lett. 2021;517:66–77. doi: 10.1016/j.canlet.2021.05.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 260.Ning K, Wang T, Sun X, Zhang P, Chen Y, Jin J, et al. UCH-L1-containing exosomes mediate chemotherapeutic resistance transfer in breast cancer. J Surg Oncol. 2017;115(8):932–940. doi: 10.1002/jso.24614. [DOI] [PubMed] [Google Scholar]
- 261.Yang SJ, Wang DD, Li J, Xu HZ, Shen HY, Chen X, et al. Predictive role of GSTP1-containing exosomes in chemotherapy-resistant breast cancer. Gene. 2017;623:5–14. doi: 10.1016/j.gene.2017.04.031. [DOI] [PubMed] [Google Scholar]
- 262.Wang X, Xu C, Hua Y, Sun L, Cheng K, Jia Z, et al. Exosomes play an important role in the process of psoralen reverse multidrug resistance of breast cancer. J Exp Clin Cancer Res CR. 2016;35(1):186. doi: 10.1186/s13046-016-0468-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 263.He C, Jaffar Ali D, Li Y, Zhu Y, Sun B, Xiao Z. Engineering of HN3 increases the tumor targeting specificity of exosomes and upgrade the anti-tumor effect of sorafenib on HuH-7 cells. PeerJ. 2020;8:e9524. doi: 10.7717/peerj.9524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 264.Wang J, Li D, Zhuang Y, Fu J, Li X, Shi Q, et al. Exosomes derived from bone marrow stromal cells decrease the sensitivity of leukemic cells to etoposide. Oncol Lett. 2017;14(3):3082–3088. doi: 10.3892/ol.2017.6509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 265.Liu Y, Song B, Wei Y, Chen F, Chi Y, Fan H, et al. Exosomes from mesenchymal stromal cells enhance imatinib-induced apoptosis in human leukemia cells via activation of caspase signaling pathway. Cytotherapy. 2018;20(2):181–188. doi: 10.1016/j.jcyt.2017.11.006. [DOI] [PubMed] [Google Scholar]
- 266.Dong Y, Lin Y, Gao X, Zhao Y, Wan Z, Wang H, et al. Targeted blocking of miR328 lysosomal degradation with alkalized exosomes sensitizes the chronic leukemia cells to imatinib. Appl Microbiol Biotechnol. 2019;103(23–24):9569–9582. doi: 10.1007/s00253-019-10127-3. [DOI] [PubMed] [Google Scholar]
- 267.Zheng Y, Liu L, Wang Y, Xiao S, Mai R, Zhu Z, et al. Glioblastoma stem cell (GSC)-derived PD-L1-containing exosomes activates AMPK/ULK1 pathway mediated autophagy to increase temozolomide-resistance in glioblastoma. Cell Biosci. 2021;11(1):63. doi: 10.1186/s13578-021-00575-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 268.Chuang HY, Su YK, Liu HW, Chen CH, Chiu SC, Cho DY, et al. Preclinical evidence of STAT3 inhibitor pacritinib overcoming temozolomide resistance via downregulating miR-21-enriched exosomes from M2 glioblastoma-associated macrophages. J Clin Med. 2019;8(7):959. doi: 10.3390/jcm8070959. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 269.Yuwen DL, Sheng BB, Liu J, Wenyu W, Shu YQ. MiR-146a-5p level in serum exosomes predicts therapeutic effect of cisplatin in non-small cell lung cancer. Eur Rev Med Pharmacol Sci. 2017;21(11):2650–2658. [PubMed] [Google Scholar]
- 270.Sayyed AA, Gondaliya P, Mali M, Pawar A, Bhat P, Khairnar A, et al. MiR-155 inhibitor-laden exosomes reverse resistance to Cisplatin in a 3D tumor spheroid and xenograft model of oral cancer. Mol Pharm. 2021;18(8):3010–3025. doi: 10.1021/acs.molpharmaceut.1c00213. [DOI] [PubMed] [Google Scholar]
- 271.Semaan L, Zeng Q, Lu Y, Zhang Y, Zreik MM, Chamseddine MB, et al. MicroRNA-214 enriched exosomes from human cerebral endothelial cells (hCEC) sensitize hepatocellular carcinoma to anti-cancer drugs. Oncotarget. 2021;12(3):185–198. doi: 10.18632/oncotarget.27879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 272.Zhang K, Shao CX, Zhu JD, Lv XL, Tu CY, Jiang C, et al. Exosomes function as nanoparticles to transfer miR-199a-3p to reverse chemoresistance to cisplatin in hepatocellular carcinoma. Bioscience reports. 2020;40(7):BSR20194026. doi: 10.1042/BSR20194026. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 273.Kato T, Mizutani K, Kawakami K, Fujita Y, Ehara H, Ito M. CD44v8-10 mRNA contained in serum exosomes as a diagnostic marker for docetaxel resistance in prostate cancer patients. Heliyon. 2020;6(7):e04138. doi: 10.1016/j.heliyon.2020.e04138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 274.Kabakov AE, Yakimova AO. Hypoxia-induced cancer cell responses driving radioresistance of hypoxic tumors: approaches to targeting and radiosensitizing. Cancers. 2021;13(5):1102. doi: 10.3390/cancers13051102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 275.Graham K, Unger E. Overcoming tumor hypoxia as a barrier to radiotherapy, chemotherapy and immunotherapy in cancer treatment. Int J Nanomed. 2018;13:6049–6058. doi: 10.2147/IJN.S140462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 276.Forster JC, Marcu LG, Bezak E. Approaches to combat hypoxia in cancer therapy and the potential for in silico models in their evaluation. Phys Med PM Int J Devot Appl Phys Med Biol Off J Ital Assoc Biomed Phys (AIFB) 2019;64:145–156. doi: 10.1016/j.ejmp.2019.07.006. [DOI] [PubMed] [Google Scholar]
- 277.Liu L, Zhang Z, Zhou L, Hu L, Yin C, Qing D, et al. Cancer associated fibroblasts-derived exosomes contribute to radioresistance through promoting colorectal cancer stem cells phenotype. Exp Cell Res. 2020;391(2):111956. doi: 10.1016/j.yexcr.2020.111956. [DOI] [PubMed] [Google Scholar]
- 278.Wan FZ, Chen KH, Sun YC, Chen XC, Liang RB, Chen L, et al. Exosomes overexpressing miR-34c inhibit malignant behavior and reverse the radioresistance of nasopharyngeal carcinoma. J Transl Med. 2020;18(1):12. doi: 10.1186/s12967-019-02203-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 279.Tang Z, He J, Zou J, Yu S, Sun X, Qin L. Cisplatin-resistant HepG2 cell-derived exosomes transfer cisplatin resistance to cisplatin-sensitive cells in HCC. PeerJ. 2021;9:e11200. doi: 10.7717/peerj.11200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 280.Min QH, Wang XZ, Zhang J, Chen QG, Li SQ, Liu XQ, et al. Exosomes derived from imatinib-resistant chronic myeloid leukemia cells mediate a horizontal transfer of drug-resistant trait by delivering miR-365. Exp Cell Res. 2018;362(2):386–393. doi: 10.1016/j.yexcr.2017.12.001. [DOI] [PubMed] [Google Scholar]
- 281.Li G, Lin H, Tian R, Zhao P, Huang Y, Pang X, et al. VEGFR-2 inhibitor apatinib hinders endothelial cells progression triggered by irradiated gastric cancer cells-derived exosomes. J Cancer. 2018;9(21):4049–4057. doi: 10.7150/jca.25370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 282.Ni C, Fang QQ, Chen WZ, Jiang JX, Jiang Z, Ye J, et al. Breast cancer-derived exosomes transmit lncRNA SNHG16 to induce CD73+γδ1 Treg cells. Signal Transduct Target Ther. 2020;5(1):41. doi: 10.1038/s41392-020-0129-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 283.Ashrafizadeh M, Zarrabi A, Hushmandi K, Zarrin V, Moghadam ER, Zabolian A, et al. PD-1/PD-L1 axis regulation in cancer therapy: the role of long non-coding RNAs and microRNAs. Life Sci. 2020;256:117899. doi: 10.1016/j.lfs.2020.117899. [DOI] [PubMed] [Google Scholar]
- 284.Dou D, Ren X, Han M, Xu X, Ge X, Gu Y, et al. Cancer-associated fibroblasts-derived exosomes suppress immune cell function in breast cancer via the miR-92/PD-L1 pathway. Front Immunol. 2020;11:2026. doi: 10.3389/fimmu.2020.02026. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 285.Mirzaei S, Zarrabi A, Hashemi F, Zabolian A, Saleki H, Ranjbar A, et al. Regulation of nuclear factor-kappaB (NF-κB) signaling pathway by non-coding RNAs in cancer: inhibiting or promoting carcinogenesis? Cancer Lett. 2021;509:63–80. doi: 10.1016/j.canlet.2021.03.025. [DOI] [PubMed] [Google Scholar]
- 286.Neurath MF. IL-36 in chronic inflammation and cancer. Cytokine Growth Factor Rev. 2020;55:70–79. doi: 10.1016/j.cytogfr.2020.06.006. [DOI] [PubMed] [Google Scholar]
- 287.Hirano T. IL-6 in inflammation, autoimmunity and cancer. Int Immunol. 2021;33(3):127–148. doi: 10.1093/intimm/dxaa078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 288.Fishbein A, Hammock BD, Serhan CN, Panigrahy D. Carcinogenesis: failure of resolution of inflammation? Pharmacol Ther. 2021;218:107670. doi: 10.1016/j.pharmthera.2020.107670. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 289.Guo J, Duan Z, Zhang C, Wang W, He H, Liu Y, et al. Mouse 4T1 breast cancer cell-derived exosomes induce proinflammatory cytokine production in macrophages via miR-183. J Immunol (Baltimore, Md) 2020;205(10):2916–25. doi: 10.4049/jimmunol.1901104. [DOI] [PubMed] [Google Scholar]
- 290.Chatterjee S, Chatterjee A, Jana S, Dey S, Roy H, Das MK, et al. Transforming growth factor beta orchestrates PD-L1 enrichment in tumor-derived exosomes and mediates CD8 T-cell dysfunction regulating early phosphorylation of TCR signalome in breast cancer. Carcinogenesis. 2021;42(1):38–47. doi: 10.1093/carcin/bgaa092. [DOI] [PubMed] [Google Scholar]
- 291.Shao Q, Deng L, Liu H, Liu Z, Chen J, Jiang F, et al. Involvement of MM cell-derived exosomes in T lymphocytes immune responses. Oncol Lett. 2020;20(4):31. doi: 10.3892/ol.2020.11892. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 292.Hinata M, Kunita A, Abe H, Morishita Y, Sakuma K, Yamashita H, et al. Exosomes of epstein-barr virus-associated gastric carcinoma suppress dendritic cell maturation. Microorganisms. 2020;8(11):1776. doi: 10.3390/microorganisms8111776. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 293.Burassakarn A, Srisathaporn S, Pientong C, Wongjampa W, Vatanasapt P, Patarapadungkit N, et al. Exosomes-carrying Epstein-Barr virus-encoded small RNA-1 induces indoleamine 2, 3-dioxygenase expression in tumor-infiltrating macrophages of oral squamous-cell carcinomas and suppresses T-cell activity by activating RIG-I/IL-6/TNF-α pathway. Oral Oncol. 2021;117:105279. doi: 10.1016/j.oraloncology.2021.105279. [DOI] [PubMed] [Google Scholar]
- 294.Pakravan N, Abbasi A, Hassan ZM. Immunotherapy using oxygenated water and tumor-derived exosomes potentiates antitumor immune response and attenuates malignancy tendency in mice model of breast cancer. Oxid Med Cell Longev. 2021;2021:5529484. doi: 10.1155/2021/5529484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 295.Othman N, Jamal R, Abu N. Cancer-derived exosomes as effectors of key inflammation-related players. Front Immunol. 2019;10:2103. doi: 10.3389/fimmu.2019.02103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 296.Rishabh K, Khadilkar S, Kumar A, Kalra I, Kumar AP, Kunnumakkara AB. MicroRNAs as modulators of oral tumorigenesis-a focused review. Int J Mol Sci. 2021;22(5):2561. doi: 10.3390/ijms22052561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 297.Tan SC, Lim PY, Fang J, Mokhtar MFM, Hanif EAM, Jamal R. Association between MIR499A rs3746444 polymorphism and breast cancer susceptibility: a meta-analysis. Sci Rep. 2020;10(1):3508. doi: 10.1038/s41598-020-60442-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 298.Zhao M, Chang J, Liu R, Liu Y, Qi J, Wang Y, et al. miR-495 and miR-5688 are down-regulated in non-small cell lung cancer under hypoxia to maintain interleukin-11 expression. Cancer Commun (London, England) 2020;40(9):435–452. doi: 10.1002/cac2.12076. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 299.Seol HS, Akiyama Y, Lee SE, Shimada S, Jang SJ. Loss of miR-100 and miR-125b results in cancer stem cell properties through IGF2 upregulation in hepatocellular carcinoma. Sci Rep. 2020;10(1):21412. doi: 10.1038/s41598-020-77960-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 300.Jin L, Zhang Z. Serum miR-3180-3p and miR-124-3p may function as noninvasive biomarkers of cisplatin resistance in gastric cancer. Clin Lab. 2020 doi: 10.7754/Clin.Lab.2020.200302. [DOI] [PubMed] [Google Scholar]
- 301.Guo Y, Cui X, Zhang Y, Ma X, Ren A, Huang H. Diagnostic and prognostic value of serum miR-296-5p and miR-28-3p in human gastric cancer. Cancer Biother Radiopharm. 2020 doi: 10.1089/cbr.2020.4144. [DOI] [PubMed] [Google Scholar]
- 302.Guo S, Chen J, Chen F, Zeng Q, Liu WL, Zhang G. Exosomes derived from Fusobacterium nucleatum-infected colorectal cancer cells facilitate tumour metastasis by selectively carrying miR-1246/92b-3p/27a-3p and CXCL16. Gut. 2020;70(8):1507–1519. doi: 10.1136/gutjnl-2020-321187. [DOI] [PubMed] [Google Scholar]
- 303.Wang XH, Jiang ZH, Yang HM, Zhang Y, Xu LH. Hypoxia-induced FOXO4/LDHA axis modulates gastric cancer cell glycolysis and progression. Clin Transl Med. 2021;11(1):e279. doi: 10.1002/ctm2.279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 304.Zhou X, Ma W, Li X, Xu J. Glaucocalyxin a prevents hypoxia-induced epithelial-mesenchymal transition in human gastric cancer cells through the PI3K/Akt signaling pathway. J Recept Signal Transduct Res. 2020;42(2):1–8. doi: 10.1080/10799893.2020.1853160. [DOI] [PubMed] [Google Scholar]
- 305.Xia X, Wang S, Ni B, Xing S, Cao H, Zhang Z, et al. Hypoxic gastric cancer-derived exosomes promote progression and metastasis via MiR-301a-3p/PHD3/HIF-1α positive feedback loop. Oncogene. 2020;39(39):6231–6244. doi: 10.1038/s41388-020-01425-6. [DOI] [PubMed] [Google Scholar]
- 306.Mohan CD, Srinivasa V, Rangappa S, Mervin L, Mohan S, Paricharak S, et al. Trisubstituted-imidazoles induce apoptosis in human breast cancer cells by targeting the oncogenic PI3K/Akt/mTOR signaling pathway. PLoS One. 2016;11(4):e0153155. doi: 10.1371/journal.pone.0153155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 307.Kim C, Lee JH, Ko JH, Chinnathambi A, Alharbi SA, Shair OHM, et al. Formononetin regulates multiple oncogenic signaling cascades and enhances sensitivity to bortezomib in a multiple myeloma mouse model. Biomolecules. 2019;9(7):262. doi: 10.3390/biom9070262. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 308.Lee JH, Chinnathambi A, Alharbi SA, Shair OHM, Sethi G, Ahn KS. Farnesol abrogates epithelial to mesenchymal transition process through regulating Akt/mTOR pathway. Pharmacol Res. 2019;150:104504. doi: 10.1016/j.phrs.2019.104504. [DOI] [PubMed] [Google Scholar]
- 309.Abadi AJ, Zarrabi A, Gholami MH, Mirzaei S, Hashemi F, Zabolian A, et al. Small in size, but large in action: microRNAs as potential modulators of PTEN in breast and lung cancers. Biomolecules. 2021;11(2):304. doi: 10.3390/biom11020304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 310.Ashrafizadeh M, Najafi M, Ang HL, Moghadam ER, Mahabady MK, Zabolian A, et al. PTEN, a barrier for proliferation and metastasis of gastric cancer cells: from molecular pathways to targeting and regulation. Biomedicines. 2020;8(8):264. doi: 10.3390/biomedicines8080264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 311.Ashrafizadeh M, Zarrabi A, Samarghandian S, Najafi M. PTEN: What we know of the function and regulation of this onco-suppressor factor in bladder cancer? Eur J Pharmacol. 2020;881:173226. doi: 10.1016/j.ejphar.2020.173226. [DOI] [PubMed] [Google Scholar]
- 312.Wang Y, Lin C. Exosomes miR-22-3p derived from mesenchymal stem cells suppress colorectal cancer cell proliferation and invasion by regulating RAP2B and PI3K/AKT pathway. J Oncol. 2021;2021:3874478. doi: 10.1155/2021/3874478. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 313.Zheng P, Luo Q, Wang W, Li J, Wang T, Wang P, et al. Tumor-associated macrophages-derived exosomes promote the migration of gastric cancer cells by transfer of functional Apolipoprotein E. Cell Death Dis. 2018;9(4):434. doi: 10.1038/s41419-018-0465-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 314.Dai G, Yao X, Zhang Y, Gu J, Geng Y, Xue F, et al. Colorectal cancer cell-derived exosomes containing miR-10b regulate fibroblast cells via the PI3K/Akt pathway. Bull Cancer. 2018;105(4):336–349. doi: 10.1016/j.bulcan.2017.12.009. [DOI] [PubMed] [Google Scholar]
- 315.Nie H, Xie X, Zhang D, Zhou Y, Li B, Li F, et al. Use of lung-specific exosomes for miRNA-126 delivery in non-small cell lung cancer. Nanoscale. 2020;12(2):877–887. doi: 10.1039/c9nr09011h. [DOI] [PubMed] [Google Scholar]
- 316.Hu X, Mu Y, Liu J, Mu X, Gao F, Chen L, et al. Exosomes derived from hypoxic colorectal cancer cells transfer miR-410-3p to regulate tumor progression. J Cancer. 2020;11(16):4724–4735. doi: 10.7150/jca.33232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 317.Liu MX, Liao J, Xie M, Gao ZK, Wang XH, Zhang Y, et al. miR-93-5p transferred by exosomes promotes the proliferation of esophageal cancer cells via intercellular communication by targeting PTEN. Biomed Environ Sci BES. 2018;31(3):171–185. doi: 10.3967/bes2018.023. [DOI] [PubMed] [Google Scholar]
- 318.Hiremath IS, Goel A, Warrier S, Kumar AP, Sethi G, Garg M. The multidimensional role of the Wnt/β-catenin signaling pathway in human malignancies. J Cell Physiol. 2021 doi: 10.1002/jcp.30561. [DOI] [PubMed] [Google Scholar]
- 319.Ong MS, Cai W, Yuan Y, Leong HC, Tan TZ, Mohammad A, et al. 'Lnc'-ing Wnt in female reproductive cancers: therapeutic potential of long non-coding RNAs in Wnt signalling. Br J Pharmacol. 2017;174(24):4684–4700. doi: 10.1111/bph.13958. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 320.Bhuvanalakshmi G, Gamit N, Patil M, Arfuso F, Sethi G, Dharmarajan A, et al. Stemness, pluripotentiality, and wnt antagonism: sFRP4, a wnt antagonist mediates pluripotency and stemness in glioblastoma. Cancers. 2018;11(1):25. doi: 10.3390/cancers11010025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 321.Wong CC, Xu J, Bian X, Wu JL, Kang W, Qian Y, et al. In colorectal cancer cells with mutant KRAS, SLC25A22-mediated glutaminolysis reduces DNA demethylation to increase WNT signaling, stemness, and drug resistance. Gastroenterology. 2020;159(6):2163–80.e6. doi: 10.1053/j.gastro.2020.08.016. [DOI] [PubMed] [Google Scholar]
- 322.Menck K, Heinrichs S, Baden C, Bleckmann A. The WNT/ROR pathway in cancer: from signaling to therapeutic intervention. Cells. 2021;10(1):142. doi: 10.3390/cells10010142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 323.Wei Y, Wei L, Li J, Ma Z, Zhang Q, Han Z, et al. SLCO4A1-AS1 promotes cell growth and induces resistance in lung adenocarcinoma by modulating miR-4701-5p/NFE2L1 axis to activate WNT pathway. Cancer Med. 2020;9(19):7205–7217. doi: 10.1002/cam4.3270. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 324.Castagnoli L, Tagliabue E, Pupa SM. Inhibition of the wnt signalling pathway: an avenue to control breast cancer aggressiveness. Int J Mol Sci. 2020;21(23):9069. doi: 10.3390/ijms21239069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 325.Jiang T, Gao W, Lin S, Chen H, Du B, Liu Q, et al. FNDC1 promotes the invasiveness of gastric cancer via Wnt/β-catenin signaling pathway and correlates with peritoneal metastasis and prognosis. Front Oncol. 2020;10:590492. doi: 10.3389/fonc.2020.590492. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 326.Harada T, Yamamoto H, Kishida S, Kishida M, Awada C, Takao T, et al. Wnt5b-associated exosomes promote cancer cell migration and proliferation. Cancer Sci. 2017;108(1):42–52. doi: 10.1111/cas.13109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 327.Xie H, Wang J. MicroRNA-320a-containing exosomes from human umbilical cord mesenchymal stem cells curtail proliferation and metastasis in lung cancer by binding to SOX4. J Recept Signal Transduct Res. 2021 doi: 10.1080/10799893.2021.1918166. [DOI] [PubMed] [Google Scholar]
- 328.Yang SS, Ma S, Dou H, Liu F, Zhang SY, Jiang C, et al. Breast cancer-derived exosomes regulate cell invasion and metastasis in breast cancer via miR-146a to activate cancer associated fibroblasts in tumor microenvironment. Exp Cell Res. 2020;391(2):111983. doi: 10.1016/j.yexcr.2020.111983. [DOI] [PubMed] [Google Scholar]
- 329.Alharbi M, Lai A, Guanzon D, Palma C, Zuñiga F, Perrin L, et al. Ovarian cancer-derived exosomes promote tumour metastasis in vivo: an effect modulated by the invasiveness capacity of their originating cells. Clin Sci (London, England) 2019;133(13):1401–19. doi: 10.1042/CS20190082. [DOI] [PubMed] [Google Scholar]
- 330.Yuan X, Qian N, Ling S, Li Y, Sun W, Li J, et al. Breast cancer exosomes contribute to pre-metastatic niche formation and promote bone metastasis of tumor cells. Theranostics. 2021;11(3):1429–1445. doi: 10.7150/thno.45351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 331.Zuo L, Tao H, Xu H, Li C, Qiao G, Guo M, et al. Exosomes-coated miR-34a displays potent antitumor activity in pancreatic cancer both in vitro and in vivo. Drug Des Dev Ther. 2020;14:3495–3507. doi: 10.2147/DDDT.S265423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 332.Yang F, Yan Y, Yang Y, Hong X, Wang M, Yang Z, et al. MiR-210 in exosomes derived from CAFs promotes non-small cell lung cancer migration and invasion through PTEN/PI3K/AKT pathway. Cell Signal. 2020;73:109675. doi: 10.1016/j.cellsig.2020.109675. [DOI] [PubMed] [Google Scholar]
- 333.Yan L, Li Q, Sun K, Jiang F. MiR-4644 is upregulated in plasma exosomes of bladder cancer patients and promotes bladder cancer progression by targeting UBIAD1. Am J Transl Res. 2020;12(10):6277–6289. [PMC free article] [PubMed] [Google Scholar]
- 334.Chen QL, Xie CF, Feng KL, Cui DY, Sun SL, Zhang JC, et al. microRNAs carried by exosomes promote epithelial-mesenchymal transition and metastasis of liver cancer cells. Am J Transl Res. 2020;12(10):6811–6826. [PMC free article] [PubMed] [Google Scholar]
- 335.Moradi-Chaleshtori M, Bandehpour M, Soudi S, Mohammadi-Yeganeh S, Hashemi SM. In vitro and in vivo evaluation of anti-tumoral effect of M1 phenotype induction in macrophages by miR-130 and miR-33 containing exosomes. Cancer Immunol Immunother CII. 2021;70(5):1323–1339. doi: 10.1007/s00262-020-02762-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 336.Yu L, Sui B, Fan W, Lei L, Zhou L, Yang L, et al. Exosomes derived from osteogenic tumor activate osteoclast differentiation and concurrently inhibit osteogenesis by transferring COL1A1-targeting miRNA-92a-1-5p. J Extracell Vesicles. 2021;10(3):e12056. doi: 10.1002/jev2.12056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 337.Fang K, Chen X, Qiu F, Xu J, Xiong H, Zhang Z. Serum-derived exosomes-mediated circular RNA ARHGAP10 modulates the progression of non-small-cell lung cancer through the miR-638/FAM83F axis. Cancer Biother Radiopharm. 2020;37:96–110. doi: 10.1089/cbr.2019.3534. [DOI] [PubMed] [Google Scholar]
- 338.Sheng Y, Han C, Yang Y, Wang J, Gu Y, Li W, et al. Correlation between LncRNA-LINC00659 and clinical prognosis in gastric cancer and study on its biological mechanism. J Cell Mol Med. 2020;24(24):14467–14480. doi: 10.1111/jcmm.16069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 339.Fan Y, Sheng W, Meng Y, Cao Y, Li R. LncRNA PTENP1 inhibits cervical cancer progression by suppressing miR-106b. Artif Cells Nanomed Biotechnol. 2020;48(1):393–407. doi: 10.1080/21691401.2019.1709852. [DOI] [PubMed] [Google Scholar]
- 340.Cui Y, Pu R, Ye J, Huang H, Liao D, Yang Y, et al. LncRNA FAM230B promotes gastric cancer growth and metastasis by regulating the miR-27a-5p/TOP2A axis. Dig Dis Sci. 2021;66(8):2637–2650. doi: 10.1007/s10620-020-06581-z. [DOI] [PubMed] [Google Scholar]
- 341.Xu L, Huan L, Guo T, Wu Y, Liu Y, Wang Q, et al. LncRNA SNHG11 facilitates tumor metastasis by interacting with and stabilizing HIF-1α. Oncogene. 2020;39(46):7005–7018. doi: 10.1038/s41388-020-01512-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 342.Ma Z, Wang YY, Xin HW, Wang L, Arfuso F, Dharmarajan A, et al. The expanding roles of long non-coding RNAs in the regulation of cancer stem cells. Int J Biochem Cell Biol. 2019;108:17–20. doi: 10.1016/j.biocel.2019.01.003. [DOI] [PubMed] [Google Scholar]
- 343.Najafi S, Tan SC, Raee P, Rahmati Y, Asemani Y, Lee EHC, et al. Gene regulation by antisense transcription: a focus on neurological and cancer diseases. Biomed Pharmacother. 2022;145:112265. doi: 10.1016/j.biopha.2021.112265. [DOI] [PubMed] [Google Scholar]
- 344.Thakur KK, Kumar A, Banik K, Verma E, Khatoon E, Harsha C, et al. Long noncoding RNAs in triple-negative breast cancer: a new frontier in the regulation of tumorigenesis. J Cell Physiol. 2021 doi: 10.1002/jcp.30463. [DOI] [PubMed] [Google Scholar]
- 345.Pandya G, Kirtonia A, Sethi G, Pandey AK, Garg M. The implication of long non-coding RNAs in the diagnosis, pathogenesis and drug resistance of pancreatic ductal adenocarcinoma and their possible therapeutic potential. Biochim Biophys Acta. 2020;1874(2):188423. doi: 10.1016/j.bbcan.2020.188423. [DOI] [PubMed] [Google Scholar]
- 346.Chen X, Tang FR, Arfuso F, Cai WQ, Ma Z, Yang J, et al. The emerging role of long non-coding RNAs in the metastasis of hepatocellular carcinoma. Biomolecules. 2019;10(1):66. doi: 10.3390/biom10010066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 347.Mishra S, Verma SS, Rai V, Awasthee N, Chava S, Hui KM, et al. Long non-coding RNAs are emerging targets of phytochemicals for cancer and other chronic diseases. Cell Mol Life Sci CMLS. 2019;76(10):1947–1966. doi: 10.1007/s00018-019-03053-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 348.Shi Q, Li Y, Li S, Jin L, Lai H, Wu Y, et al. LncRNA DILA1 inhibits Cyclin D1 degradation and contributes to tamoxifen resistance in breast cancer. Nat Commun. 2020;11(1):5513. doi: 10.1038/s41467-020-19349-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 349.Hong J, Guo F, Lu SY, Shen C, Ma D, Zhang X, et al. F nucleatum targets lncRNA ENO1-IT1 to promote glycolysis and oncogenesis in colorectal cancer. Gut. 2020 doi: 10.1136/gutjnl-2020-322780. [DOI] [PubMed] [Google Scholar]
- 350.Chang KC, Diermeier SD, Yu AT, Brine LD, Russo S, Bhatia S, et al. MaTAR25 lncRNA regulates the Tensin1 gene to impact breast cancer progression. Nat Commun. 2020;11(1):6438. doi: 10.1038/s41467-020-20207-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 351.Shermane Lim YW, Xiang X, Garg M, Le MTN, Li-Ann Wong A, Wang L, et al. The double-edged sword of H19 lncRNA: insights into cancer therapy. Cancer Lett. 2021;500:253–262. doi: 10.1016/j.canlet.2020.11.006. [DOI] [PubMed] [Google Scholar]
- 352.Lei Y, Guo W, Chen B, Chen L, Gong J, Li W. Tumor-released lncRNA H19 promotes gefitinib resistance via packaging into exosomes in non-small cell lung cancer. Oncol Rep. 2018;40(6):3438–3446. doi: 10.3892/or.2018.6762. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 353.Hardin H, Helein H, Meyer K, Robertson S, Zhang R, Zhong W, et al. Thyroid cancer stem-like cell exosomes: regulation of EMT via transfer of lncRNAs. Lab Invest J Tech Methods Pathol. 2018;98(9):1133–42. doi: 10.1038/s41374-018-0065-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 354.Zhang H, Wang J, Ren T, Huang Y, Yu Y, Chen C, et al. LncRNA CASC15 is upregulated in osteosarcoma plasma exosomes and CASC15 knockdown inhibits osteosarcoma progression by regulating miR-338-3p/RAB14 axis. Onco Targets Ther. 2020;13:12055–12066. doi: 10.2147/OTT.S282053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 355.Ashrafizadeh M, Zarabi A, Hushmandi K, Moghadam ER, Hashemi F, Daneshi S, et al. C-Myc signaling pathway in treatment and prevention of brain tumors. Curr Cancer Drug Targets. 2021;21(1):2–20. doi: 10.2174/1568009620666201016121005. [DOI] [PubMed] [Google Scholar]
- 356.Lu Y, Chen L, Li L, Cao Y. Exosomes derived from brain metastatic breast cancer cells destroy the blood-brain barrier by carrying lncRNA GS1-600G8.5. BioMed Res Int. 2020;2020:7461727. doi: 10.1155/2020/7461727. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 357.Yin Z, Zhou Y, Ma T, Chen S, Shi N, Zou Y, et al. Down-regulated lncRNA SBF2-AS1 in M2 macrophage-derived exosomes elevates miR-122-5p to restrict XIAP, thereby limiting pancreatic cancer development. J Cell Mol Med. 2020;24(9):5028–5038. doi: 10.1111/jcmm.15125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 358.Xu CG, Yang MF, Ren YQ, Wu CH, Wang LQ. Exosomes mediated transfer of lncRNA UCA1 results in increased tamoxifen resistance in breast cancer cells. Eur Rev Med Pharmacol Sci. 2016;20(20):4362–4368. [PubMed] [Google Scholar]
- 359.Cui X, Xiao D, Cui Y, Wang X. Exosomes-derived long non-coding RNA HOTAIR reduces laryngeal cancer radiosensitivity by regulating microRNA-454-3p/E2F2 Axis. Onco Targets Ther. 2019;12:10827–10839. doi: 10.2147/OTT.S224881. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 360.Yang YN, Zhang R, Du JW, Yuan HH, Li YJ, Wei XL, et al. Predictive role of UCA1-containing exosomes in cetuximab-resistant colorectal cancer. Cancer Cell Int. 2018;18:164. doi: 10.1186/s12935-018-0660-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 361.Barbagallo C, Brex D, Caponnetto A, Cirnigliaro M, Scalia M, Magnano A, et al. LncRNA UCA1, upregulated in CRC biopsies and downregulated in serum exosomes, controls mRNA expression by RNA-RNA interactions. Mol Thera Nucleic Acids. 2018;12:229–241. doi: 10.1016/j.omtn.2018.05.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 362.Wu Q, Wu X, Ying X, Zhu Q, Wang X, Jiang L, et al. Suppression of endothelial cell migration by tumor associated macrophage-derived exosomes is reversed by epithelial ovarian cancer exosomal lncRNA. Cancer Cell Int. 2017;17:62. doi: 10.1186/s12935-017-0430-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 363.Xian J, Zeng Y, Chen S, Lu L, Liu L, Chen J, et al. Discovery of a novel linc01125 isoform in serum exosomes as a promising biomarker for NSCLC diagnosis and survival assessment. Carcinogenesis. 2021;42(6):831–841. doi: 10.1093/carcin/bgab034. [DOI] [PubMed] [Google Scholar]
- 364.Holdt LM, Kohlmaier A, Teupser D. Molecular roles and function of circular RNAs in eukaryotic cells. Cell Mol Life Sci CMLS. 2018;75(6):1071–1098. doi: 10.1007/s00018-017-2688-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 365.Chen LL. The biogenesis and emerging roles of circular RNAs. Nat Rev Mol Cell Biol. 2016;17(4):205–211. doi: 10.1038/nrm.2015.32. [DOI] [PubMed] [Google Scholar]
- 366.Haque S, Harries LW. Circular RNAs (circRNAs) in health and disease. Genes. 2017;8(12):353. doi: 10.3390/genes8120353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 367.Meng S, Zhou H, Feng Z, Xu Z, Tang Y, Li P, et al. CircRNA: functions and properties of a novel potential biomarker for cancer. Mol Cancer. 2017;16(1):94. doi: 10.1186/s12943-017-0663-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 368.Hao Q, Zhang Z. hsa_circRNA_000166 facilitated cell growth and limited apoptosis through targeting miR-326/LASP1 axis in colorectal cancer. Gastroenterol Res Pract. 2020;2020:8834359. doi: 10.1155/2020/8834359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 369.Zhou ZF, Wei Z, Yao JC, Liu SY, Wang F, Wang Z, et al. CircRNA_102179 promotes the proliferation, migration and invasion in non-small cell lung cancer cells by regulating miR-330-5p/HMGB3 axis. Pathol Res Pract. 2020;216(11):153144. doi: 10.1016/j.prp.2020.153144. [DOI] [PubMed] [Google Scholar]
- 370.Zhang X, Yang H, Jia Y, Xu Z, Zhang L, Sun M, et al. circRNA_0005529 facilitates growth and metastasis of gastric cancer via regulating miR-527/Sp1 axis. BMC Mol Cell Biol. 2021;22(1):6. doi: 10.1186/s12860-020-00340-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 371.Li J, Li Z, Jiang P, Peng M, Zhang X, Chen K, et al. Circular RNA IARS (circ-IARS) secreted by pancreatic cancer cells and located within exosomes regulates endothelial monolayer permeability to promote tumor metastasis. J Exp Clin Cancer Res CR. 2018;37(1):177. doi: 10.1186/s13046-018-0822-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 372.Ashrafizadeh M, Ahmadi Z, Farkhondeh T, Samarghandian S. Resveratrol targeting the Wnt signaling pathway: a focus on therapeutic activities. J Cell Physiol. 2020;235(5):4135–45. doi: 10.1002/jcp.29327. [DOI] [PubMed] [Google Scholar]
- 373.Zhao H, Chen S, Fu Q. Exosomes from CD133(+) cells carrying circ-ABCC1 mediate cell stemness and metastasis in colorectal cancer. J Cell Biochem. 2020;121(5–6):3286–3297. doi: 10.1002/jcb.29600. [DOI] [PubMed] [Google Scholar]
- 374.Ma J, Qi G, Li L. A novel serum exosomes-based biomarker hsa_circ_0002130 facilitates osimertinib-resistance in non-small cell lung cancer by sponging miR-498. Onco Targets Ther. 2020;13:5293–5307. doi: 10.2147/OTT.S243214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 375.Shi Q, Ji T, Ma Z, Tan Q, Liang J. Serum exosomes-based biomarker circ_0008928 regulates cisplatin sensitivity, tumor progression, and glycolysis metabolism by miR-488/HK2 axis in cisplatin-resistant nonsmall cell lung carcinoma. Cancer Biother Radiopharm. 2021 doi: 10.1089/cbr.2020.4490. [DOI] [PubMed] [Google Scholar]
- 376.Xian J, Su W, Liu L, Rao B, Lin M, Feng Y, et al. Identification of three circular RNA cargoes in serum exosomes as diagnostic biomarkers of non-small-cell lung cancer in the chinese population. J Mol Diagn JMD. 2020;22(8):1096–1108. doi: 10.1016/j.jmoldx.2020.05.011. [DOI] [PubMed] [Google Scholar]
- 377.Chen F, Huang C, Wu Q, Jiang L, Chen S, Chen L. Circular RNAs expression profiles in plasma exosomes from early-stage lung adenocarcinoma and the potential biomarkers. J Cell Biochem. 2020;121(3):2525–2533. doi: 10.1002/jcb.29475. [DOI] [PubMed] [Google Scholar]
- 378.Wang J, Zhang Q, Zhou S, Xu H, Wang D, Feng J, et al. Circular RNA expression in exosomes derived from breast cancer cells and patients. Epigenomics. 2019;11(4):411–421. doi: 10.2217/epi-2018-0111. [DOI] [PubMed] [Google Scholar]
- 379.Dou Y, Cha DJ, Franklin JL, Higginbotham JN, Jeppesen DK, Weaver AM, et al. Circular RNAs are down-regulated in KRAS mutant colon cancer cells and can be transferred to exosomes. Sci Rep. 2016;6:37982. doi: 10.1038/srep37982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 380.Guo S, Hu C, Zhai X, Sun D. Circular RNA 0006602 in plasma exosomes: a new potential diagnostic biomarker for hepatocellular carcinoma. Am J Transl Res. 2021;13(6):6001–6015. [PMC free article] [PubMed] [Google Scholar]
- 381.Ouyang Y, Tang Y, Fu L, Peng S, Wu W, Tan D, et al. Exosomes secreted by chronic hepatitis B patients with PNALT and liver inflammation grade ≥ A2 promoted the progression of liver cancer by transferring miR-25-3p to inhibit the co-expression of TCF21 and HHIP. Cell Prolif. 2020;53(7):e12833. doi: 10.1111/cpr.12833. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 382.Li L, Li C, Wang S, Wang Z, Jiang J, Wang W, et al. Exosomes derived from hypoxic oral squamous cell carcinoma cells deliver miR-21 to normoxic cells to elicit a prometastatic phenotype. Can Res. 2016;76(7):1770–1780. doi: 10.1158/0008-5472.CAN-15-1625. [DOI] [PubMed] [Google Scholar]
- 383.Park EJ, Jung HJ, Choi HJ, Jang HJ, Park HJ, Nejsum LN, et al. Exosomes co-expressing AQP5-targeting miRNAs and IL-4 receptor-binding peptide inhibit the migration of human breast cancer cells. FASEB J Off Publ Fed Am Soc Exp Biol. 2020;34(2):3379–3398. doi: 10.1096/fj.201902434R. [DOI] [PubMed] [Google Scholar]
- 384.Yang Y, Li J, Geng Y. Exosomes derived from chronic lymphocytic leukaemia cells transfer miR-146a to induce the transition of mesenchymal stromal cells into cancer-associated fibroblasts. J Biochem. 2020;168(5):491–498. doi: 10.1093/jb/mvaa064. [DOI] [PubMed] [Google Scholar]
- 385.Wang C, Wang Y, Chang X, Ba X, Hu N, Liu Q, et al. Melanoma-derived exosomes endow fibroblasts with an invasive potential via miR-21 target signaling pathway. Cancer Manag Res. 2020;12:12965–12974. doi: 10.2147/CMAR.S273718. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 386.Wang Y, Wang P, Zhao L, Chen X, Lin Z, Zhang L, et al. miR-224-5p carried by human umbilical cord mesenchymal stem cells-derived exosomes regulates autophagy in breast cancer cells via HOXA5. Front Cell Dev Biol. 2021;9:679185. doi: 10.3389/fcell.2021.679185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 387.Zhou Y, Zhu Y, Dong X, Cao G, Li Y, Fan Y, et al. Exosomes derived from pancreatic cancer cells induce osteoclast differentiation through the miR125a-5p/TNFRSF1B pathway. Onco Targets Ther. 2021;14:2727–2739. doi: 10.2147/OTT.S282319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 388.Rezaei R, Baghaei K, Hashemi SM, Zali MR, Ghanbarian H, Amani D. Tumor-derived exosomes enriched by miRNA-124 promote anti-tumor immune response in CT-26 tumor-bearing mice. Front Med. 2021;8:619939. doi: 10.3389/fmed.2021.619939. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 389.Zhang N, Li L, Luo J, Tan J, Hu W, Li Z, et al. Inhibiting microRNA-424 in bone marrow mesenchymal stem cells-derived exosomes suppresses tumor growth in colorectal cancer by upregulating TGFBR3. Arch Biochem Biophys. 2021;709:108965. doi: 10.1016/j.abb.2021.108965. [DOI] [PubMed] [Google Scholar]
- 390.Vakhshiteh F, Rahmani S, Ostad SN, Madjd Z, Dinarvand R, Atyabi F. Exosomes derived from miR-34a-overexpressing mesenchymal stem cells inhibit in vitro tumor growth: a new approach for drug delivery. Life Sci. 2021;266:118871. doi: 10.1016/j.lfs.2020.118871. [DOI] [PubMed] [Google Scholar]
- 391.Sheykhhasan M, Kalhor N, Sheikholeslami A, Dolati M, Amini E, Fazaeli H. Exosomes of mesenchymal stem cells as a proper vehicle for transfecting miR-145 into the breast cancer cell line and its effect on metastasis. Biomed Res Int. 2021;2021:5516078. doi: 10.1155/2021/5516078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 392.Du L, Tao X, Shen X. Human umbilical cord mesenchymal stem cell-derived exosomes inhibit migration and invasion of breast cancer cells via miR-21-5p/ZNF367 pathway. Breast Cancer (Tokyo, Japan) 2021;28(4):829–837. doi: 10.1007/s12282-021-01218-z. [DOI] [PubMed] [Google Scholar]
- 393.Yue S, Ye X, Zhou T, Gan D, Qian H, Fang W, et al. PGRN(-/-) TAMs-derived exosomes inhibit breast cancer cell invasion and migration and its mechanism exploration. Life Sci. 2021;264:118687. doi: 10.1016/j.lfs.2020.118687. [DOI] [PubMed] [Google Scholar]
- 394.Liu Y, Yang Y, Du J, Lin D, Li F. MiR-3613-3p from carcinoma-associated fibroblasts exosomes promoted breast cancer cell proliferation and metastasis by regulating SOCS2 expression. IUBMB Life. 2020;72(8):1705–1714. doi: 10.1002/iub.2292. [DOI] [PubMed] [Google Scholar]
- 395.Wang B, Zhang Y, Ye M, Wu J, Ma L, Chen H. Cisplatin-resistant MDA-MB-231 cell-derived exosomes increase the resistance of recipient cells in an exosomal miR-423-5p-dependent manner. Curr Drug Metab. 2019;20(10):804–814. doi: 10.2174/1389200220666190819151946. [DOI] [PubMed] [Google Scholar]
- 396.Zeng Q, Zhu Z, Song L, He Z. Transferred by exosomes-derived MiR-19b-3p targets PTEN to regulate esophageal cancer cell apoptosis, migration and invasion. 2020. Biosci Rep. 10.1042/BSR20201858. [DOI] [PMC free article] [PubMed]
- 397.Xu Y, Liu N, Wei Y, Zhou D, Lin R, Wang X, et al. Anticancer effects of miR-124 delivered by BM-MSC derived exosomes on cell proliferation, epithelial mesenchymal transition, and chemotherapy sensitivity of pancreatic cancer cells. Aging. 2020;12(19):19660–19676. doi: 10.18632/aging.103997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 398.Lan J, Sun L, Xu F, Liu L, Hu F, Song D, et al. M2 macrophage-derived exosomes promote cell migration and invasion in colon cancer. Can Res. 2019;79(1):146–158. doi: 10.1158/0008-5472.CAN-18-0014. [DOI] [PubMed] [Google Scholar]
- 399.Huang W, Yan Y, Liu Y, Lin M, Ma J, Zhang W, et al. Exosomes with low miR-34c-3p expression promote invasion and migration of non-small cell lung cancer by upregulating integrin α2β1. Signal Transduct Target Ther. 2020;5(1):39. doi: 10.1038/s41392-020-0133-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 400.Chen R, Qian Z, Xu X, Zhang C, Niu Y, Wang Z, et al. Exosomes-transmitted miR-7 reverses gefitinib resistance by targeting YAP in non-small-cell lung cancer. Pharmacol Res. 2021;165:105442. doi: 10.1016/j.phrs.2021.105442. [DOI] [PubMed] [Google Scholar]
- 401.Deng Z, Rong Y, Teng Y, Zhuang X, Samykutty A, Mu J, et al. Exosomes miR-126a released from MDSC induced by DOX treatment promotes lung metastasis. Oncogene. 2017;36(5):639–651. doi: 10.1038/onc.2016.229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 402.Lou G, Song X, Yang F, Wu S, Wang J, Chen Z, et al. Exosomes derived from miR-122-modified adipose tissue-derived MSCs increase chemosensitivity of hepatocellular carcinoma. J Hematol Oncol. 2015;8:122. doi: 10.1186/s13045-015-0220-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 403.Li J, Yu J, Zhang H, Wang B, Guo H, Bai J, et al. Exosomes-derived MiR-302b suppresses lung cancer cell proliferation and migration via TGFβRII inhibition. Cell Physiol Biochem Int J Exp Cell Physiol Biochem Pharmacol. 2016;38(5):1715–1726. doi: 10.1159/000443111. [DOI] [PubMed] [Google Scholar]
- 404.Zhang X, Li F, Tang Y, Ren Q, Xiao B, Wan Y, et al. miR-21a in exosomes from Lewis lung carcinoma cells accelerates tumor growth through targeting PDCD4 to enhance expansion of myeloid-derived suppressor cells. Oncogene. 2020;39(40):6354–6369. doi: 10.1038/s41388-020-01406-9. [DOI] [PubMed] [Google Scholar]
- 405.He Z, Li W, Zheng T, Liu D, Zhao S. Human umbilical cord mesenchymal stem cells-derived exosomes deliver microRNA-375 to downregulate ENAH and thus retard esophageal squamous cell carcinoma progression. J Exp Clin Cancer Res CR. 2020;39(1):140. doi: 10.1186/s13046-020-01631-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 406.Lee JC, Zhao JT, Gundara J, Serpell J, Bach LA, Sidhu S. Papillary thyroid cancer-derived exosomes contain miRNA-146b and miRNA-222. J Surg Res. 2015;196(1):39–48. doi: 10.1016/j.jss.2015.02.027. [DOI] [PubMed] [Google Scholar]
- 407.Liu Y, Fu W, Cao X, Li S, Xiong T, Zhang X, et al. Delivery of miR-224-5p by exosomes from cancer-associated fibroblasts potentiates progression of clear cell renal cell carcinoma. Comput Math Methods Med. 2021;2021:5517747. doi: 10.1155/2021/5517747. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 408.Chang L, Gao H, Wang L, Wang N, Zhang S, Zhou X, et al. Exosomes derived from miR-1228 overexpressing bone marrow-mesenchymal stem cells promote growth of gastric cancer cells. Aging. 2021;13(8):11808–11821. doi: 10.18632/aging.202878. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 409.Pan L, Liang W, Fu M, Huang ZH, Li X, Zhang W, et al. Exosomes-mediated transfer of long noncoding RNA ZFAS1 promotes gastric cancer progression. J Cancer Res Clin Oncol. 2017;143(6):991–1004. doi: 10.1007/s00432-017-2361-2. [DOI] [PubMed] [Google Scholar]
- 410.Xian D, Niu L, Zeng J, Wang L. LncRNA KCNQ1OT1 secreted by tumor cell-derived exosomes mediates immune escape in colorectal cancer by regulating PD-L1 ubiquitination via MiR-30a-5p/USP22. Front Cell Dev Biol. 2021;9:653808. doi: 10.3389/fcell.2021.653808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 411.Wang YL, Liu LC, Hung Y, Chen CJ, Lin YZ, Wu WR, et al. Long non-coding RNA HOTAIR in circulatory exosomes is correlated with ErbB2/HER2 positivity in breast cancer. Breast (Edinburgh, Scotland) 2019;46:64–69. doi: 10.1016/j.breast.2019.05.003. [DOI] [PubMed] [Google Scholar]
- 412.Yang H, Qu H, Huang H, Mu Z, Mao M, Xie Q, et al. Exosomes-mediated transfer of long noncoding RNA LINC01133 represses bladder cancer progression via regulating the Wnt signaling pathway. Cell Biol Int. 2021;45(7):1510–1522. doi: 10.1002/cbin.11590. [DOI] [PubMed] [Google Scholar]
- 413.Liu H, Shen M, Zhao D, Ru D, Duan Y, Ding C, et al. The effect of triptolide-loaded exosomes on the proliferation and apoptosis of human ovarian cancer SKOV3 cells. Biomed Res Int. 2019;2019:2595801. doi: 10.1155/2019/2595801. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 414.Kim MS, Haney MJ, Zhao Y, Yuan D, Deygen I, Klyachko NL, et al. Engineering macrophage-derived exosomes for targeted paclitaxel delivery to pulmonary metastases: in vitro and in vivo evaluations. Nanomed Nanotechnol Biol Med. 2018;14(1):195–204. doi: 10.1016/j.nano.2017.09.011. [DOI] [PubMed] [Google Scholar]
- 415.Qambrani A, Rehman FU, Tanziela T, Shaikh S, Semcheddine F, Du T, et al. Biocompatible exosomes nanodrug cargo for cancer cell bioimaging and drug delivery. Biomed Mater (Bristol, England) 2021;16(2):025026. doi: 10.1088/1748-605X/abaaa2. [DOI] [PubMed] [Google Scholar]
- 416.Dhayapulay A, Kanapathipillai M. Exosomes based geldanamycin delivery to cancer cells with increased therapeutic efficacy. J Biomed Nanotechnol. 2019;15(11):2202–2208. doi: 10.1166/jbn.2019.2844. [DOI] [PubMed] [Google Scholar]
- 417.Wang P, Wang H, Huang Q, Peng C, Yao L, Chen H, et al. Exosomes from M1-polarized macrophages enhance paclitaxel antitumor activity by activating macrophages-mediated inflammation. Theranostics. 2019;9(6):1714–1727. doi: 10.7150/thno.30716. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 418.Thakur A, Sidu RK, Zou H, Alam MK, Yang M, Lee Y. Inhibition of glioma cells' proliferation by doxorubicin-loaded exosomes via microfluidics. Int J Nanomed. 2020;15:8331–8343. doi: 10.2147/IJN.S263956. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 419.Zhang X, Liu L, Tang M, Li H, Guo X, Yang X. The effects of umbilical cord-derived macrophage exosomes loaded with cisplatin on the growth and drug resistance of ovarian cancer cells. Drug Dev Ind Pharm. 2020;46(7):1150–1162. doi: 10.1080/03639045.2020.1776320. [DOI] [PubMed] [Google Scholar]
- 420.Li Y, Gao Y, Gong C, Wang Z, Xia Q, Gu F, et al. A33 antibody-functionalized exosomes for targeted delivery of doxorubicin against colorectal cancer. Nanomed Nanotechnol Biol Med. 2018;14(7):1973–1985. doi: 10.1016/j.nano.2018.05.020. [DOI] [PubMed] [Google Scholar]
- 421.Zhou Y, Zhu F, Liu Y, Zheng M, Wang Y, Zhang D, et al. Blood-brain barrier-penetrating siRNA nanomedicine for Alzheimer's disease therapy. Sci Adv. 2020 doi: 10.1126/sciadv.abc7031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 422.Lu ZR, Laney VE, Hall R, Ayat N. Environment-responsive lipid/siRNA nanoparticles for cancer therapy. Adv Healthc Mater. 2021;10(5):2001294. doi: 10.1002/adhm.202001294. [DOI] [PubMed] [Google Scholar]
- 423.Zhang MM, Bahal R, Rasmussen TP, Manautou JE, Zhong XB. The growth of siRNA-based therapeutics: updated clinical studies. Biochem Pharmacol. 2021;189:114432. doi: 10.1016/j.bcp.2021.114432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 424.Mirzaei S, Gholami MH, Hashemi F, Zabolian A, Hushmandi K, Rahmanian V, et al. Employing siRNA tool and its delivery platforms in suppressing cisplatin resistance: approaching to a new era of cancer chemotherapy. Life Sci. 2021;277:119430. doi: 10.1016/j.lfs.2021.119430. [DOI] [PubMed] [Google Scholar]
- 425.Ashrafizade M, Delfi M, Hashemi F, Zabolian A, Saleki H, Bagherian M, et al. Biomedical application of chitosan-based nanoscale delivery systems: Potential usefulness in siRNA delivery for cancer therapy. Carbohydr Polym. 2021;260:117809. doi: 10.1016/j.carbpol.2021.117809. [DOI] [PubMed] [Google Scholar]
- 426.Najafi S, Tan SC, Aghamiri S, Raee P, Ebrahimi Z, Jahromi ZK, et al. Therapeutic potentials of CRISPR-Cas genome editing technology in human viral infections. Biomed Pharmacother. 2022;148:112743. doi: 10.1016/j.biopha.2022.112743. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 427.Khalaf K, Janowicz K, Dyszkiewicz-Konwińska M, Hutchings G, Dompe C, Moncrieff L, et al. CRISPR/Cas9 in cancer immunotherapy: animal models and human clinical trials. Genes. 2020;11(8):921. doi: 10.3390/genes11080921. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 428.Hazafa A, Mumtaz M, Farooq MF, Bilal S, Chaudhry SN, Firdous M, et al. CRISPR/Cas9: a powerful genome editing technique for the treatment of cancer cells with present challenges and future directions. Life Sci. 2020;263:118525. doi: 10.1016/j.lfs.2020.118525. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 429.Martinez-Lage M, Torres-Ruiz R, Puig-Serra P, Moreno-Gaona P, Martin MC, Moya FJ, et al. In vivo CRISPR/Cas9 targeting of fusion oncogenes for selective elimination of cancer cells. Nat Commun. 2020;11(1):5060. doi: 10.1038/s41467-020-18875-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 430.Mohammadinejad R, Sassan H, Pardakhty A, Hashemabadi M, Ashrafizadeh M, Dehshahri A, et al. ZEB1 and ZEB2 gene editing mediated by CRISPR/Cas9 in A549 cell line. Bratislava Med J. 2020;121(1):31–6. doi: 10.4149/BLL_2020_005. [DOI] [PubMed] [Google Scholar]
- 431.Faruqu FN, Xu L, Al-Jamal KT. Preparation of exosomes for siRNA delivery to cancer cells. J Vis Exp. 2018 doi: 10.3791/58814. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 432.Tao H, Xu H, Zuo L, Li C, Qiao G, Guo M, et al. Exosomes-coated bcl-2 siRNA inhibits the growth of digestive system tumors both in vitro and in vivo. Int J Biol Macromol. 2020;161:470–480. doi: 10.1016/j.ijbiomac.2020.06.052. [DOI] [PubMed] [Google Scholar]
- 433.Koh SA, Kim MK, Lee KH, Kim SW, Kim J-R. RhoGDI2 is associated with HGF-mediated tumor invasion through VEGF in stomach cancer. Clin Exp Metastasis. 2014;31(7):805–15. doi: 10.1007/s10585-014-9671-4. [DOI] [PubMed] [Google Scholar]
- 434.Yang X, Zhang XF, Lu X, Jia HL, Liang L, Dong QZ, et al. MicroRNA-26a suppresses angiogenesis in human hepatocellular carcinoma by targeting hepatocyte growth factor-cMet pathway. Hepatology. 2014;59(5):1874–85. doi: 10.1002/hep.26941. [DOI] [PubMed] [Google Scholar]
- 435.Zhai Y, Wu W, Xi X, Yu R. Adipose-derived stem cells promote proliferation and invasion in cervical cancer by targeting the HGF/c-MET pathway. Cancer Manag Research. 2020;12:11823–11832. doi: 10.2147/CMAR.S277130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 436.Xu J, Liu S, Yang X, Cao S, Zhou Y. Paracrine HGF promotes EMT and mediates the effects of PSC on chemoresistance by activating c-Met/PI3K/Akt signaling in pancreatic cancer in vitro. Life Sci. 2020;263:118523. doi: 10.1016/j.lfs.2020.118523. [DOI] [PubMed] [Google Scholar]
- 437.Zhang H, Wang Y, Bai M, Wang J, Zhu K, Liu R, et al. Exosomes serve as nanoparticles to suppress tumor growth and angiogenesis in gastric cancer by delivering hepatocyte growth factor siRNA. Cancer Sci. 2018;109(3):629–641. doi: 10.1111/cas.13488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 438.Zhang Z, Cheng L, Zhang Q, Kong Y, He D, Li K, et al. Co-targeting Plk1 and DNMT3a in advanced prostate cancer. Adv Sci (Weinheim, Baden-Wurttemberg, Germany) 2021;8(13):e2101458. doi: 10.1002/advs.202101458. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 439.Montaudon E, Nikitorowicz-Buniak J, Sourd L, Morisset L, El Botty R, Huguet L, et al. PLK1 inhibition exhibits strong anti-tumoral activity in CCND1-driven breast cancer metastases with acquired palbociclib resistance. Nat Commun. 2020;11(1):4053. doi: 10.1038/s41467-020-17697-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 440.Greco KA, Franzen CA, Foreman KE, Flanigan RC, Kuo PC, Gupta GN. PLK-1 silencing in bladder cancer by siRNA delivered With exosomes. Urology. 2016;91(241):e1–7. doi: 10.1016/j.urology.2016.01.028. [DOI] [PubMed] [Google Scholar]
- 441.Limoni SK, Moghadam MF, Moazzeni SM, Gomari H, Salimi F. Engineered exosomes for targeted transfer of siRNA to HER2 positive breast cancer cells. Appl Biochem Biotechnol. 2019;187(1):352–364. doi: 10.1007/s12010-018-2813-4. [DOI] [PubMed] [Google Scholar]
- 442.Teesalu T, Sugahara KN, Kotamraju VR, Ruoslahti E. C-end rule peptides mediate neuropilin-1-dependent cell, vascular, and tissue penetration. Proc Natl Acad Sci USA. 2009;106(38):16157–16162. doi: 10.1073/pnas.0908201106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 443.Kawakami T, Tokunaga T, Hatanaka H, Kijima H, Yamazaki H, Abe Y, et al. Neuropilin 1 and neuropilin 2 co-expression is significantly correlated with increased vascularity and poor prognosis in nonsmall cell lung carcinoma. Cancer. 2002;95(10):2196–2201. doi: 10.1002/cncr.10936. [DOI] [PubMed] [Google Scholar]
- 444.Roth L, Agemy L, Kotamraju VR, Braun G, Teesalu T, Sugahara KN, et al. Transtumoral targeting enabled by a novel neuropilin-binding peptide. Oncogene. 2012;31(33):3754–3763. doi: 10.1038/onc.2011.537. [DOI] [PubMed] [Google Scholar]
- 445.Bai J, Duan J, Liu R, Du Y, Luo Q, Cui Y, et al. Engineered targeting tLyp-1 exosomes as gene therapy vectors for efficient delivery of siRNA into lung cancer cells. Asian J Pharm Sci. 2020;15(4):461–471. doi: 10.1016/j.ajps.2019.04.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 446.Lin D, Zhang H, Liu R, Deng T, Ning T, Bai M, et al. iRGD-modified exosomes effectively deliver CPT1A siRNA to colon cancer cells, reversing oxaliplatin resistance by regulating fatty acid oxidation. Mol Oncol. 2021 doi: 10.1002/1878-0261.13052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 447.Li Z, Yang L, Wang H, Binzel DW, Williams TM, Guo P. Non-small-cell lung cancer regression by sirna delivered through exosomes that display EGFR RNA aptamer. Nucleic Acid Ther. 2021 doi: 10.1089/nat.2021.0002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 448.Li H, Yang C, Shi Y, Zhao L. Exosomes derived from siRNA against GRP78 modified bone-marrow-derived mesenchymal stem cells suppress Sorafenib resistance in hepatocellular carcinoma. J Nanobiotechnol. 2018;16(1):103. doi: 10.1186/s12951-018-0429-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 449.Kim SM, Yang Y, Oh SJ, Hong Y, Seo M, Jang M. Cancer-derived exosomes as a delivery platform of CRISPR/Cas9 confer cancer cell tropism-dependent targeting. J Controll Release Off J Controll Release Soc. 2017;266:8–16. doi: 10.1016/j.jconrel.2017.09.013. [DOI] [PubMed] [Google Scholar]
- 450.Gulei D, Berindan-Neagoe I. Activation of necroptosis by engineered self tumor-derived exosomes loaded with CRISPR/Cas9. Mol Ther Nucleic Acids. 2019;17:448–451. doi: 10.1016/j.omtn.2019.05.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 451.Gabrusiewicz K, Li X, Wei J, Hashimoto Y, Marisetty AL, Ott M, et al. Glioblastoma stem cell-derived exosomes induce M2 macrophages and PD-L1 expression on human monocytes. Oncoimmunology. 2018;7(4):e1412909. doi: 10.1080/2162402X.2017.1412909. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 452.Qian M, Wang S, Guo X, Wang J, Zhang Z, Qiu W, et al. Hypoxic glioma-derived exosomes deliver microRNA-1246 to induce M2 macrophage polarization by targeting TERF2IP via the STAT3 and NF-κB pathways. Oncogene. 2020;39(2):428–442. doi: 10.1038/s41388-019-0996-y. [DOI] [PubMed] [Google Scholar]
- 453.Wang Z, Yuan Y, Ji X, Xiao X, Li Z, Yi X, et al. The Hippo-TAZ axis mediates vascular endothelial growth factor C in glioblastoma-derived exosomes to promote angiogenesis. Cancer Lett. 2021;513:1–13. doi: 10.1016/j.canlet.2021.05.002. [DOI] [PubMed] [Google Scholar]
- 454.Naryzhny S, Volnitskiy A, Kopylov A, Zorina E, Kamyshinsky R, Bairamukov V, et al. Proteome of glioblastoma-derived exosomes as a source of biomarkers. Biomedicines. 2020;8(7):216. doi: 10.3390/biomedicines8070216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 455.Azambuja JH, Ludwig N, Yerneni S, Rao A, Braganhol E, Whiteside TL. Molecular profiles and immunomodulatory activities of glioblastoma-derived exosomes. Neuro-Oncol Adv. 2020;2(1):vdaa 56. doi: 10.1093/noajnl/vdaa056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 456.Sun Z, Wang L, Zhou Y, Dong L, Ma W, Lv L, et al. Glioblastoma stem cell-derived exosomes enhance stemness and tumorigenicity of glioma cells by transferring Notch1 protein. Cell Mol Neurobiol. 2020;40(5):767–784. doi: 10.1007/s10571-019-00771-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 457.Li J, Liao T, Liu H, Yuan H, Ouyang T, Wang J, et al. Hypoxic glioma stem cell-derived exosomes containing linc01060 promote progression of glioma by regulating the MZF1/c-Myc/HIF1α axis. Can Res. 2021;81(1):114–128. doi: 10.1158/0008-5472.CAN-20-2270. [DOI] [PubMed] [Google Scholar]
- 458.Guo X, Qiu W, Liu Q, Qian M, Wang S, Zhang Z, et al. Immunosuppressive effects of hypoxia-induced glioma exosomes through myeloid-derived suppressor cells via the miR-10a/Rora and miR-21/Pten pathways. Oncogene. 2018;37(31):4239–4259. doi: 10.1038/s41388-018-0261-9. [DOI] [PubMed] [Google Scholar]
- 459.Qian M, Chen Z, Guo X, Wang S, Zhang Z, Qiu W, et al. Exosomes derived from hypoxic glioma deliver miR-1246 and miR-10b-5p to normoxic glioma cells to promote migration and invasion. Lab Investig J Tech Methods Pathol. 2021;101(5):612–24. doi: 10.1038/s41374-020-00522-0. [DOI] [PubMed] [Google Scholar]
- 460.Lang HL, Hu GW, Zhang B, Kuang W, Chen Y, Wu L, et al. Glioma cells enhance angiogenesis and inhibit endothelial cell apoptosis through the release of exosomes that contain long non-coding RNA CCAT2. Oncol Rep. 2017;38(2):785–798. doi: 10.3892/or.2017.5742. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 461.Sun X, Ma X, Wang J, Zhao Y, Wang Y, Bihl JC, et al. Glioma stem cells-derived exosomes promote the angiogenic ability of endothelial cells through miR-21/VEGF signal. Oncotarget. 2017;8(22):36137–36148. doi: 10.18632/oncotarget.16661. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 462.Guo X, Qiu W, Wang J, Liu Q, Qian M, Wang S, et al. Glioma exosomes mediate the expansion and function of myeloid-derived suppressor cells through microRNA-29a/Hbp1 and microRNA-92a/Prkar1a pathways. Int J Cancer. 2019;144(12):3111–3126. doi: 10.1002/ijc.32052. [DOI] [PubMed] [Google Scholar]
- 463.Domenis R, Cesselli D, Toffoletto B, Bourkoula E, Caponnetto F, Manini I, et al. Systemic T cells immunosuppression of glioma stem cell-derived exosomes is mediated by monocytic myeloid-derived suppressor cells. PLoS One. 2017;12(1):e0169932. doi: 10.1371/journal.pone.0169932. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 464.Figueroa J, Phillips LM, Shahar T, Hossain A, Gumin J, Kim H, et al. Exosomes from glioma-associated mesenchymal stem cells increase the tumorigenicity of glioma stem-like cells via transfer of miR-1587. Can Res. 2017;77(21):5808–5819. doi: 10.1158/0008-5472.CAN-16-2524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 465.Wen SW, Sceneay J, Lima LG, Wong CS, Becker M, Krumeich S, et al. The biodistribution and immune suppressive effects of breast cancer-derived exosomes. Can Res. 2016;76(23):6816–6827. doi: 10.1158/0008-5472.CAN-16-0868. [DOI] [PubMed] [Google Scholar]
- 466.Wu Q, Sun S, Li Z, Yang Q, Li B, Zhu S, et al. Breast cancer-released exosomes trigger cancer-associated cachexia to promote tumor progression. Adipocyte. 2019;8(1):31–45. doi: 10.1080/21623945.2018.1551688. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 467.Inubushi S, Kawaguchi H, Mizumoto S, Kunihisa T, Baba M, Kitayama Y, et al. Oncogenic miRNAs identified in tear exosomes from metastatic breast cancer patients. Anticancer Res. 2020;40(6):3091–3096. doi: 10.21873/anticanres.14290. [DOI] [PubMed] [Google Scholar]
- 468.Wen SW, Lima LG, Lobb RJ, Norris EL, Hastie ML, Krumeich S, et al. Breast cancer-derived exosomes reflect the cell-of-origin phenotype. Proteomics. 2019;19(8):e1800180. doi: 10.1002/pmic.201800180. [DOI] [PubMed] [Google Scholar]
- 469.Wang X, Cheng K, Zhang G, Jia Z, Yu Y, Guo J, et al. Enrichment of CD44 in exosomes from breast cancer cells treated with doxorubicin promotes chemoresistance. Front Oncol. 2020;10:960. doi: 10.3389/fonc.2020.00960. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 470.Li K, Liu T, Chen J, Ni H, Li W. Survivin in breast cancer-derived exosomes activates fibroblasts by up-regulating SOD1, whose feedback promotes cancer proliferation and metastasis. J Biol Chem. 2020;295(40):13737–13752. doi: 10.1074/jbc.RA120.013805. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 471.Piao YJ, Kim HS, Hwang EH, Woo J, Zhang M, Moon WK. Breast cancer cell-derived exosomes and macrophage polarization are associated with lymph node metastasis. Oncotarget. 2018;9(7):7398–7410. doi: 10.18632/oncotarget.23238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 472.Sen K, Sheppe AEF, Singh I, Hui WW, Edelmann MJ, Rinaldi C. Exosomes released by breast cancer cells under mild hyperthermic stress possess immunogenic potential and modulate polarization in vitro in macrophages. Int J Hyperth Off J Eur Soc Hyperth Oncol North Am Hyperth Gr. 2020;37(1):696–710. doi: 10.1080/02656736.2020.1778800. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 473.Rahman MA, Barger JF, Lovat F, Gao M, Otterson GA, Nana-Sinkam P. Lung cancer exosomes as drivers of epithelial mesenchymal transition. Oncotarget. 2016;7(34):54852–54866. doi: 10.18632/oncotarget.10243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 474.Wei F, Ma C, Zhou T, Dong X, Luo Q, Geng L, et al. Exosomes derived from gemcitabine-resistant cells transfer malignant phenotypic traits via delivery of miRNA-222-3p. Mol Cancer. 2017;16(1):132. doi: 10.1186/s12943-017-0694-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 475.Xu H, Jiao X, Wu Y, Li S, Cao L, Dong L. Exosomes derived from PM2.5-treated lung cancer cells promote the growth of lung cancer via the Wnt3a/β-catenin pathway. Oncol Rep. 2019;41(2):1180–8. doi: 10.3892/or.2018.6862. [DOI] [PubMed] [Google Scholar]
- 476.Wu H, Zeng C, Ye Y, Liu J, Mu Z, Xie Y, et al. Exosomes from irradiated nonsmall cell lung cancer cells reduced sensitivity of recipient cells to anaplastic lymphoma kinase inhibitors. Mol Pharm. 2018;15(5):1892–1900. doi: 10.1021/acs.molpharmaceut.8b00059. [DOI] [PubMed] [Google Scholar]
- 477.Pan D, Chen J, Feng C, Wu W, Wang Y, Tong J, et al. Preferential localization of MUC1 glycoprotein in exosomes secreted by non-small cell lung carcinoma cells. Int J Mol Sci. 2019;20(2):323. doi: 10.3390/ijms20020323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 478.Deng G, Qu J, Zhang Y, Che X, Cheng Y, Fan Y, et al. Gastric cancer-derived exosomes promote peritoneal metastasis by destroying the mesothelial barrier. FEBS Lett. 2017;591(14):2167–2179. doi: 10.1002/1873-3468.12722. [DOI] [PubMed] [Google Scholar]
- 479.Wu L, Zhang X, Zhang B, Shi H, Yuan X, Sun Y, et al. Exosomes derived from gastric cancer cells activate NF-κB pathway in macrophages to promote cancer progression. Tumour Biol J Int Soc Oncodev Biol Med. 2016;37(9):12169–12180. doi: 10.1007/s13277-016-5071-5. [DOI] [PubMed] [Google Scholar]
- 480.Shen Y, Xue C, Li X, Ba L, Gu J, Sun Z, et al. Effects of gastric cancer cell-derived exosomes on the immune regulation of mesenchymal stem cells by the NF-kB signaling pathway. Stem Cells Dev. 2019;28(7):464–476. doi: 10.1089/scd.2018.0125. [DOI] [PubMed] [Google Scholar]
- 481.Ning X, Zhang H, Wang C, Song X. Exosomes released by gastric cancer cells induce transition of pericytes into cancer-associated fibroblasts. Med Sci Monit Int Med J Exp Clin Res. 2018;24:2350–2359. doi: 10.12659/MSM.906641. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 482.Gao Z, Song C, Li G, Lin H, Lian X, Zhang N, et al. Pyrotinib treatment on HER2-positive gastric cancer cells promotes the released exosomes to enhance endothelial cell progression, which can be counteracted by apatinib. Onco Targets Ther. 2019;12:2777–2787. doi: 10.2147/OTT.S194768. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 483.Chen L, Guo P, He Y, Chen Z, Chen L, Luo Y, et al. HCC-derived exosomes elicit HCC progression and recurrence by epithelial-mesenchymal transition through MAPK/ERK signalling pathway. Cell Death Dis. 2018;9(5):513. doi: 10.1038/s41419-018-0534-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 484.Ge Y, Mu W, Ba Q, Li J, Jiang Y, Xia Q, et al. Hepatocellular carcinoma-derived exosomes in organotropic metastasis, recurrence and early diagnosis application. Cancer Lett. 2020;477:41–48. doi: 10.1016/j.canlet.2020.02.003. [DOI] [PubMed] [Google Scholar]
- 485.He X, Yu J, Xiong L, Liu Y, Fan L, Li Y, et al. Exosomes derived from liver cancer cells reprogram biological behaviors of LO2 cells by transferring Linc-ROR. Gene. 2019;719:144044. doi: 10.1016/j.gene.2019.144044. [DOI] [PubMed] [Google Scholar]
- 486.Kimura H, Yamamoto H, Harada T, Fumoto K, Osugi Y, Sada R, et al. CKAP4, a DKK1 receptor, is a biomarker in exosomes derived from pancreatic cancer and a molecular target for therapy. Clin Cancer Res Off J Am Assoc Cancer Res. 2019;25(6):1936–1947. doi: 10.1158/1078-0432.CCR-18-2124. [DOI] [PubMed] [Google Scholar]
- 487.Linton SS, Abraham T, Liao J, Clawson GA, Butler PJ, Fox T, et al. Tumor-promoting effects of pancreatic cancer cell exosomes on THP-1-derived macrophages. PLoS One. 2018;13(11):e0206759. doi: 10.1371/journal.pone.0206759. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 488.Tang P, Tao L, Yuan C, Zhang L, Xiu D. Serum derived exosomes from pancreatic cancer patients promoted metastasis: an iTRAQ-based proteomic analysis. Onco Targets Ther. 2019;12:9329–9339. doi: 10.2147/OTT.S229494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 489.Zhang YF, Zhou YZ, Zhang B, Huang SF, Li PP, He XM, et al. Pancreatic cancer-derived exosomes promoted pancreatic stellate cells recruitment by pancreatic cancer. J Cancer. 2019;10(18):4397–4407. doi: 10.7150/jca.27590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 490.Stefanius K, Servage K, de Souza SM, Gray HF, Toombs JE, Chimalapati S, et al. Human pancreatic cancer cell exosomes, but not human normal cell exosomes, act as an initiator in cell transformation. eLife. 2019 doi: 10.7554/eLife.40226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 491.Masamune A, Yoshida N, Hamada S, Takikawa T, Nabeshima T, Shimosegawa T. Exosomes derived from pancreatic cancer cells induce activation and profibrogenic activities in pancreatic stellate cells. Biochem Biophys Res Commun. 2018;495(1):71–77. doi: 10.1016/j.bbrc.2017.10.141. [DOI] [PubMed] [Google Scholar]
- 492.Zhang Y, Huang S, Li P, Chen Q, Li Y, Zhou Y, et al. Pancreatic cancer-derived exosomes suppress the production of GIP and GLP-1 from STC-1 cells in vitro by down-regulating the PCSK1/3. Cancer Lett. 2018;431:190–200. doi: 10.1016/j.canlet.2018.05.027. [DOI] [PubMed] [Google Scholar]
- 493.Yu Z, Zhao S, Ren L, Wang L, Chen Z, Hoffman RM, et al. Pancreatic cancer-derived exosomes promote tumor metastasis and liver pre-metastatic niche formation. Oncotarget. 2017;8(38):63461–63483. doi: 10.18632/oncotarget.18831. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 494.Jiao YJ, Jin DD, Jiang F, Liu JX, Qu LS, Ni WK, et al. Characterization and proteomic profiling of pancreatic cancer-derived serum exosomes. J Cell Biochem. 2019;120(1):988–999. doi: 10.1002/jcb.27465. [DOI] [PubMed] [Google Scholar]
- 495.Cheng L, Zhang K, Qing Y, Li D, Cui M, Jin P, et al. Proteomic and lipidomic analysis of exosomes derived from ovarian cancer cells and ovarian surface epithelial cells. J Ovarian Res. 2020;13(1):9. doi: 10.1186/s13048-020-0609-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 496.Lee AH, Ghosh D, Quach N, Schroeder D, Dawson MR. Ovarian cancer exosomes trigger differential biophysical response in tumor-derived fibroblasts. Sci Rep. 2020;10(1):8686. doi: 10.1038/s41598-020-65628-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 497.Yang C, Kim HS, Song G, Lim W. The potential role of exosomes derived from ovarian cancer cells for diagnostic and therapeutic approaches. J Cell Physiol. 2019;234(12):21493–21503. doi: 10.1002/jcp.28905. [DOI] [PubMed] [Google Scholar]
- 498.Chen X, Zhou J, Li X, Wang X, Lin Y, Wang X. Exosomes derived from hypoxic epithelial ovarian cancer cells deliver microRNAs to macrophages and elicit a tumor-promoted phenotype. Cancer Lett. 2018;435:80–91. doi: 10.1016/j.canlet.2018.08.001. [DOI] [PubMed] [Google Scholar]
- 499.Chen X, Ying X, Wang X, Wu X, Zhu Q, Wang X. Exosomes derived from hypoxic epithelial ovarian cancer deliver microRNA-940 to induce macrophage M2 polarization. Oncol Rep. 2017;38(1):522–528. doi: 10.3892/or.2017.5697. [DOI] [PubMed] [Google Scholar]
- 500.Ghorbanian M, Babashah S, Ataei F. The effects of ovarian cancer cell-derived exosomes on vascular endothelial growth factor expression in endothelial cells. EXCLI J. 2019;18:899–907. doi: 10.17179/excli2019-1800. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 501.Bhat A, Yadav J, Thakur K, Aggarwal N, Tripathi T, Chhokar A, et al. Exosomes from cervical cancer cells facilitate pro-angiogenic endothelial reconditioning through transfer of Hedgehog-GLI signaling components. Cancer Cell Int. 2021;21(1):319. doi: 10.1186/s12935-021-02026-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 502.Ren G, Wang Y, Yuan S, Wang B. Dendritic cells loaded with HeLa-derived exosomes simulate an antitumor immune response. Oncol Lett. 2018;15(5):6636–6640. doi: 10.3892/ol.2018.8126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 503.Salimu J, Webber J, Gurney M, Al-Taei S, Clayton A, Tabi Z. Dominant immunosuppression of dendritic cell function by prostate-cancer-derived exosomes. J Extracell Vesicles. 2017;6(1):1368823. doi: 10.1080/20013078.2017.1368823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 504.Esfandyari S, Elkafas H, Chugh RM, Park HS, Navarro A, Al-Hendy A. Exosomes as biomarkers for female reproductive diseases diagnosis and therapy. Int J Mol Sci. 2021;22(4):2165. doi: 10.3390/ijms22042165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 505.Bestard-Escalas J, Reigada R, Reyes J, de la Torre P, Liebisch G, Barceló-Coblijn G. Fatty acid unsaturation degree of plasma exosomes in colorectal cancer patients: a promising biomarker. Int J Mol Sci. 2021;22(10):5060. doi: 10.3390/ijms22105060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 506.He R, Wang Z, Shi W, Yu L, Xia H, Huang Z, et al. Exosomes in hepatocellular carcinoma microenvironment and their potential clinical application value. Biomed Pharmacother. 2021;138:111529. doi: 10.1016/j.biopha.2021.111529. [DOI] [PubMed] [Google Scholar]
- 507.Damanti CC, Gaffo E, Lovisa F, Garbin A, Di Battista P, Gallingani I, et al. MiR-26a-5p as a reference to normalize microRNA qRT-PCR levels in plasma exosomes of pediatric hematological malignancies. Cells. 2021;10(1):101. doi: 10.3390/cells10010101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 508.Fu C, Zhang Q, Wang A, Yang S, Jiang Y, Bai L, et al. EWI-2 controls nucleocytoplasmic shuttling of EGFR signaling molecules and miRNA sorting in exosomes to inhibit prostate cancer cell metastasis. Mol Oncol. 2021;15(5):1543–1565. doi: 10.1002/1878-0261.12930. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 509.Cao B, Wang P, Gu L, Liu J. Use of four genes in exosomes as biomarkers for the identification of lung adenocarcinoma and lung squamous cell carcinoma. Oncol Lett. 2021;21(4):249. doi: 10.3892/ol.2021.12510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 510.Liu G, Luo Y, Hou P. PRPS2 enhances resistance to cisplatin via facilitating exosomes-mediated macrophage M2 polarization in non-small cell lung cancer. Immunol Invest. 2021 doi: 10.1080/08820139.2021.1952217. [DOI] [PubMed] [Google Scholar]
- 511.Ganig N, Baenke F, Thepkaysone ML, Lin K, Rao VS, Wong FC, et al. Proteomic analyses of fibroblast- and serum-derived exosomes identify QSOX1 as a marker for non-invasive detection of colorectal cancer. Cancers. 2021;13(6):1351. doi: 10.3390/cancers13061351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 512.Sinha D, Roy S, Saha P, Chatterjee N, Bishayee AJC. Trends in research on exosomes in cancer progression and anticancer therapy. Cancers. 2021;13(2):326. doi: 10.3390/cancers13020326. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Not applicable.