

Abstract
Blood trimethylamine-N-oxide (TMAO) has been associated with cardiovascular disease. Black raspberry (Rubus occidentalis, BR) has been regarded to be beneficial for cardiovascular health. This study aimed to investigate how BR extract affects serum lipid profile, gut microbial composition, metabolites in rats fed TMAO with a high-fat diet. Dietary TMAO increased serum LDL cholesterol, while BR extract decreased its level. α-Diversity of gut microbiota was not changed; however, in the rats fed TMAO, Macellibacteroides and Mucispirillum were enriched, while Ruminococcaceae was reduced. The BR supplementation could restore Macellibacteroides, Clostridium, and Ruminococcaceae. The BR supplementation increased cecal hippuric acid and serum farnesoid X receptor-antagonistic bile acids, including ursodeoxycholic acid (UDCA), tauro-α-muricholic acid, and tauro-UDCA. The BR supplementation tended to upregulate Cyp7a1 and Abcg5 expressions while downregulating Srebf2 and Hmgcr expressions. BR extract affects the gut bacterial community and microbial metabolites, lowering serum LDL cholesterol in rats with elevated serum TMAO.
Supplementary Information
The online version contains supplementary material available at 10.1007/s10068-022-01079-y.
Keywords: Black raspberry, Trimethylamine-N-oxide, Gut microbiota, FXR-antagonistic bile acid, Hippuric acid
Introduction
Cardiovascular disease (CVD) is one of the major causes of mortality worldwide. About a decade ago, it emerged that trimethylamine-N-oxide (TMAO) might be a putative promoter of CVD (Wang et al., 2011; Tang et al., 2013). Since then, the link between TMAO and CVD risk has revealed that cholesterol uptake, protein conformation, platelet hyperreactivity, and thrombosis potential could be induced by TMAO, leading to increase in CVD risk (Seldin et al., 2016; Zhu et al., 2016). Since TMAO can be produced from trimethylamine (TMA), a microbial metabolite generated from dietary quaternary amines, attention has also been paid to TMA precursors in developing CVD (Koeth et al., 2013; Tang et al., 2013; Zhu et al., 2016). The intakes of TMA-containing compounds, such as choline, lecithin, L-carnitine, and betaine, have been shown to increase blood TMAO level (Koeth et al., 2013; Tang et al., 2013; Wang et al., 2011).
Blood TMAO level can be increased by consuming foods rich in TMAO or TMA-containing compounds. Several studies have been reported to investigate the dietary effects of food containing TMAO or TMA-containing compounds on CVD risk factors (Wang et al., 2019; Yin et al., 2020). Some seafood and saltwater fish are commonly rich in TMAO (Summers et al., 2017). However, there has been no clinical evidence that CVD risk could be elevated by consuming TMAO-rich foods, although consuming those foods can result in a higher level of TMAO (Svensson et al., 1994). The point of view that TMAO itself may be harmful has been highly debated so far. Therefore, modulation of the gut microbiome could be a potential strategy to decrease blood TMAO level in a higher consumption of a diet rich in TMA-containing compounds. It has been investigated which dietary components can reduce CVD risk in models elevated with serum TMAO level via modulating gut microbial composition (He et al., 2019; Hu et al., 2015).
Black raspberry (Rubus occidentalis, BR) contains high amounts of polyphenols and anthocyanins (Jung et al., 2015; Tulio Jr. et al., 2008), of which cyanidin-3-glucoside, cyanidin-3-rutinoside (C3R), and ellagitannins were reported to have strong anti-oxidative and anti-inflammatory activities (Jung et al., 2014; Kula and Krauze-Baranowska, 2016). Therefore, several in vitro and in vivo studies also suggested that mechanisms of action of BR for cardio-protective effect may be explained at the endothelial level (Jung et al., 2015; Krga and Milenkovic, 2019). However, flavonoid glycosides with a di-/tri-saccharide and ellagitannins in BR were reported to be barely absorbed in the small intestine (de Ferrars et al., 2014). Thus, it may be explained that the majority of the cardio-protective effect of BR resulted from their influences on the gut environment.
Gut microbiota can alter host physiology via various kinds of microbial metabolites such as secondary bile acids (BA) and short-chain fatty acids (SCFA). These metabolites act as endocrine fashions for crosstalk between gut microbiota and host. Gut microbiota contributes to diversification and concentrations of BA and SCFA in the host, thereby influencing host metabolism since microbial metabolites bind distinctively to corresponding receptors (Wahlström et al., 2016; Winston and Theriot, 2020).
Based on this point of view, this study aimed to investigate the effects of BR extract on changes in serum lipid profile, gut microbial composition, and metabolite profile (BA and SCFA) in rats fed TMAO water with a high-fat diet. The hepatic mRNA expressions of the genes involved in cholesterol and BA metabolisms were also investigated.
Materials and methods
Standards
α-Muricholic acid (α-MCA), β-MCA, ω-MCA, tauro-α-MCA (Tα-MCA), tauro-β-MCA(Tβ-MCA), and glycine-β-MCA (Gβ-MCA) were purchased from Cayman Chemical (Ann Arbor, MI, USA). All the other standards were purchased from Sigma-Aldrich (St. Louis, MO, USA).
BR extract preparation and its characteristics
BR fruits harvested in Gochang (Korea) were purchased and extracted to obtain the powder form of BR extract (Lim et al., 2020). Briefly, BR fruits crushed by hand were added into 80% (v/v) aqueous ethanol solution and extracted for 1 h. The extract was filtered using Whatman No. 2 filter paper (Whatman International Ltd., Maidstone, England) and the filtrate was concentrated using a vacuum rotary evaporator (A-10005, Eyela Co., Tokyo, Japan). The concentrated BR extract was freeze-dried and powdered to prepare the experimental diets. Total phenolic content and anthocyanin composition were determined using spectroscopy and high-performance liquid chromatography (Lim et al., 2020). The total polyphenol content in the BR extract was 251.0 ± 40.3 μmol gallic acid equivalent/g (Lim et al., 2020). The contents of C3R, cyanidin-3-xylosylrutinoside, and cyanidin-3-glucoside in the BR extract were 3.49 ± 0.13 μmol/g, 1.39 ± 0.08 μmol C3R equivalent/g, and 1.03 ± 0.02 μmol/g, respectively (Lim et al., 2020).
Experimental design
All protocols used in this study were conducted in accordance with institutional policies for animal health and well-being and approved by the Institutional Animal Care and Use Committee of Seoul National University (Approval No.: SNU-190417-12-1). Fifty Sprague–Dawley rats were randomly divided into 5 groups after 1-week acclimation and two were housed per cage: CON, fed AIN-93G diet (16% calories from fat); HF, fed high-fat diet (45% calories from fat); HFB, fed high-fat diet with 0.6% (w/w) BR extract; HFT, fed high-fat diet and 0.15% (w/w) TMAO water; and HFTB, fed high-fat diet with 0.6% BR extract and 0.15% TMAO water. The concentrations of BR extract and TMAO water were decided based on our previous study (Lim et al., 2020) and preliminary experiment results for the determination of TMAO water concentration (Figure S1 in the Supplementary Material), respectively. All the groups were fed ad libitum experimental diet and water and raised for 8 weeks under controlled temperature (23 ± 3 °C), humidity (50 ± 10%), and a 12/12 h light–dark cycle. Water was replaced every two days.
Blood and tissue collection
All the rats were fasted for 6 h and were euthanized by asphyxiation with CO2. Blood was centrifuged at 3000×g at 4 °C for 20 min after coagulation. Liver tissue and cecal content were isolated and immediately stored at − 80 °C until analysis.
Quantification of serum TMAO
The serum sample was mixed with 80% (v/v) ice-cold methanol solution, vortexed for 1 min, and centrifuged at 12,000×g for 5 min at 4 °C. The supernatant was filtered using a 0.22 μm syringe filter (Pall Co., Port Washington, NY, USA) and the filtrate was used for future analysis. The analyte was separated on an Acquity ultra-performance liquid chromatography (UPLC) (Waters Co., Milford, MA, USA) equipped with Acquity UPLC BEH amide column (2.1 mm × 100 mm, 1.7 μm, Waters Co.) heated at 50 °C. The mobile phase consisted of two eluents: 0.5 mM ammonium formate (pH 8.1) in water (A) and acetonitrile (B). The gradient program was: 0–2.5 min, 95–5% B; 2.5–5 min, 5–95% B; and 5–6 min, 95% B. Flow rate was 0.6 mL/min. The ion transition (m/z 76.07 → 59.07 for TMAO) was used for quantitation. The sample was analyzed using SYNAPT G2-Si mass spectrometer (Waters Co.) in positive ion electrospray mode. Capillary and sampling cone voltages were set at 0.5 kV and 15 V, respectively. Flow rates of desolvation gas and cone gas were 650 L/h and 250 L/h, respectively. Desolvation temperature was 150 °C. Data acquisition and quantitation were carried out using MassLynx software 4.1 (Waters Co.).
Serum lipid profiling
Serum total glycerides (TG), total cholesterols (TC), and HDL cholesterol (HDL-C) were determined with commercial kits (Asan Pharmaceutical Co., Ltd., Seoul, Korea) according to the manufacturer’s instructions based on enzymatic colorimetric methods. Absorbance was measured by a spectrophotometer (Spectramax190, Molecular Devices). Serum LDL cholesterol (LDL-C) level was calculated by the following formula (Friedewald et al., 1972): LDL-C (mg/dL) = TC (mg/dL) − HDL-C (mg/dL) − TG (mg/dL)/5.
Total bacterial gDNA extraction and 16S rRNA gene sequencing analysis
Total bacterial genomic DNA (gDNA) was extracted from cecal content using QIAamp Fast DNA Stool Mini Kit (Qiagen, Hilden, Germany) according to the manufacturer’s instruction with some modifications. Briefly, about 100 mg of cecal content was added into 0.7 mL InhibitEX buffer containing sterilized 0.1 mm glass bead (Scientific Industries, Inc., Bohemia, NY, USA) and homogenized using TissueLyser (Qiagen) at 30 Hz for 5 min and another 5 min. The homogenated soup was then heated at 95 °C for 5 min. After loading and washing steps, the gDNA was incubated with 50 μL ATE buffer for 5 min and eluted by centrifugation (16,000×g, 1 min).
The V3 and V4 hypervariable regions of the 16S rRNA gene were amplified with TaKaRa Ex Taq DNA Polymerase (Takara Bio, Otsu, Shiga, Japan) and universal primers (341F/805R) with overhang adapter attached, followed by AMPure XP bead (Beckman Coulter, Brea, CA, USA) cleanup. The amplicon was quantified by Qubit 3.0 System (Thermo Fisher Scientific, Hampton, NH, USA). Miseq libraries were prepared using Illumina Nextera XT Index Kit (Illumina Inc.) according to the manufacturer’s instruction. Paired-end sequencing (2 × 300 bp) was performed on Illumina Miseq Platform (Illumina Inc., San Diego, CA, USA).
Bioinformatics analysis
Quantitative Insights Into Microbial Ecology 2 (QIIME2, v 2020.2) software (Bolyen et al., 2019) was used to analyze the raw data obtained from 16S rRNA gene sequencing. DADA2 plugin (Callahan et al., 2016) in QIIME2 was used to merge and filter the sequence data. Forward and reverse sequences were trimmed (–p-trim-left-f 17 and –p-trim-left-r 21, respectively) and the parameters for truncation was set at 280 bp and 220 bp (–p-trunc-len-f 280 and –p-trunc-len-r 220, respectively) to get rid of reads having low quality. For α- and β-diversity analyses, the feature table was rarefied not to exclude any individuals by subsampling randomly (15,883 reads). Taxonomic analysis was conducted by pre-trained classifier based on Greengenes 13_8 99% operational taxonomic units (OTUs).
BA profile and hippuric acid level analyses
To analyze the BA profile (in cecum and serum) and hippuric acid (in cecum), UPLC SYNAPT G2-Si Q-TOF mass spectrometer (Waters Co., Milford, MA, USA) was used. Each of the cecal content (about 40 mg) and serum samples (100 μL) was immersed in 80% (v/v) ice-cold methanol (800 μL and 400 μL, respectively), vortexed for 5 min and 1 min, respectively, and then centrifuged at 12,000×g for 5 min at 4 °C. The supernatant was filtered using a 0.22 μm syringe filter (Pall Co., Port Washington, NY, USA) and the filtrate was concentrated only for the cecal samples by centrifugation at 15,000×g for 25 min at 4 °C in a Vivaspin centrifugal concentrator (Vivaspin 500, MWCO 3000, VS0192; Sartorius Stedim Lab, Stonehouse, UK).
Sixteen BAs and hippuric acid (Table S1 in the Supplementary Material) were separated using an Acquity UPLC system (Waters Co.) equipped with an HSS T3 column (2.1 mm × 100 mm, 1.7 μm, Waters Co.) heated at 50 °C. Mobile phase consisted of two eluents: water with 0.1% (v/v) formic acid (A) and acetonitrile with 0.1% (v/v) formic acid (B). Flow rate was set at 0.4 mL/min and gradient was as follows: 0–2.5 min, 15% B; 9 min, 95% B; 9-13 min, 95% B; 13.1 min, 95% B; and 13.1–15 min 15% B. BAs were detected by SYNAPT G2-Si Q-TOF mass spectrometer (Waters Co.) in negative ionization mode. Mass parameters were set as follows: Tof-MRM mode; capillary voltage, 2.5 kV; sampling cone voltage, 25 V; desolvation gas, 600 L/h; cone gas, 50 L/h; and desolvation temperature, 400 °C. The ion transitions used for quantitation are shown in Table S1 in the Supplementary Material. Data quantitation was performed using MassLynx software 4.1 (Waters Co.).
Cecal SCFA composition analysis
SCFA concentrations in the cecum were determined by gas chromatography (GC) equipped with a Nukol fused silica capillary column (30 m × 0.25 mm, 0.25 μm) and a flame ionization detector. Cecal content (about 100 mg) was homogenized with 1 mL distilled water. Sulfuric acid (20 μL) was then added into the homogenate and remained for 5 min at room temperature. The mixture was centrifuged at 13,000×g for 5 min and 500 μL of the supernatant was mixed with the same volume of diethyl ether. The mixture was vortexed for 1 min and centrifuged for 13,000×g for 5 min. The organic phase was collected and transferred into a vial for GC analysis. GC analysis was performed as follows: Oven temperature was held at 170 °C and the injector and detector temperatures were 225 °C. The injected volume was 2 μL and the split ratio was 100:1.
RNA extraction and real-time qPCR analysis
RNA extraction and cDNA synthesis were carried out according to the method in our previous paper (Lim et al., 2020). qPCR was performed using Applied Biosystems StepOne Real-Time PCR system (Applied Biosystems, Foster City, CA, USA) with SYBR Green PCR Master Mix (Applied Biosystems) with 2 min denaturation at 95 °C followed by 40 cycles of 15 s at 95 °C and 60 s at 58 °C. All primer sequences used in this study are listed in Table S2 in the Supplementary Material. All the gene expressions were normalized to glyceraldehyde-3-phosphate dehydrogenase (Gapdh) expression.
Statistical analysis
All statistical analyses were conducted using SPSS program (version 26.0, SPSS, Chicago, IL, USA). One-way analysis of variance (ANOVA) with Duncan’s multiple range test (for analyses of SCFA composition, hippuric acid concentrations, and mRNA expression of genes; at p < 0.05) or Kruskal–Wallis test with Dunn’s test (for analyses of α-diversity, bacterial composition at phylum and genus levels, and BA profile) was used to compare statistical significance between the groups. Permutational multivariate analysis of variance (PERMANOVA) test was used to analyze significant differences in β-diversity analysis using QIIME2 command line (qiime diversity beta-group-significance) (Bolyen et al., 2019).
Results and discussion
Body weights and daily intakes of food and water
Daily food intake was the highest in the CON among the groups and was not affected by dietary BR extract and TMAO (Table S3 in the Supplementary Material). However, there was no significant difference in daily energy intake and weight gain among the groups, although the high-fat diet-fed groups showed significantly lower food intake. Daily water intake was not significantly different among the groups regardless of the TMAO supplementation.
Serum TMAO and lipids
The effect of the black raspberry extract on the circulating level of TMAO was evaluated. The groups fed TMAO water showed a significantly higher level of TMAO than those fed plain water as expected. However, the BR supplementation did not affect the circulating level of TMAO (Figure S2 in the Supplementary Material). Serum levels of TG and HDL-C were not significantly different among the groups (Fig. 1). However, serum levels of TC (p = 0.123) and LDL-C (p < 0.05) were the highest in the HFT, indicating that dietary TMAO may contribute to an increase in serum cholesterol level in the rats fed a high-fat diet. On the other hand, the dietary BR extract reduced serum levels of TC (p = 0.123) and LDL-C (p < 0.05).
Fig. 1.
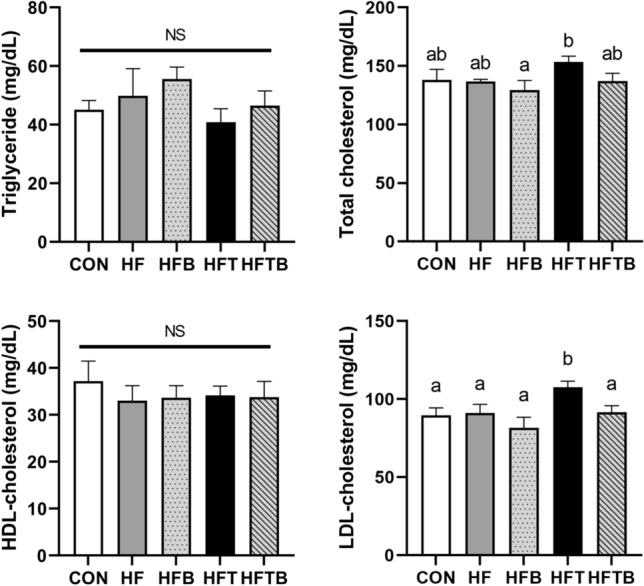
Serum lipid profile of female rats fed a high-fat diet with trimethylamine-N-oxide (TMAO) and/or black raspberry (BR) extract. All data represent the means and standard errors of means. Different small letters above bars indicate significant differences among the groups (p < 0.05; one-way ANOVA and Duncan’s multiple range test). CON (AIN-93G diet), HF (45% high-fat diet), HFB (HF + 0.6% (w/w) BR extract), HFT (HF + 0.15% (w/w) TMAO water), and HFTB (HFT + 0.6% (w/w) BR extract). NS not significant
To determine whether CVD risk was elevated in our model, we first measured the serum levels of lipids and TMAO. In this study, levels of TG and HDL-C were not changed by TMAO with a high-fat diet, while those of TC (p = 0.123) and LDL-C (p < 0.05) increased. The role of dietary TMAO in lipid abnormality is controversial. Recently, it was reported that 0.02% (w/w) TMAO might promote atherosclerosis in ApoE−/− mice under a low-fat diet (under 10% calories from fat) rather than a high-fat diet (Yu et al., 2020). Hence, the 45% high-fat diet with 0.15% TMAO feeding in this study may tend to increase serum TC level with no significance. Elevation of serum levels of LDL-C in the rats fed TMAO with a high-fat diet might be due to an imbalance in cholesterol transport by TMAO (Chen et al., 2016; Ren et al., 2016). Interestingly, the BR supplementation did not lower serum TMAO level, suggesting that BR extract may lower serum cholesterol level by regulating cholesterol metabolism, not by reducing TMAO level.
α-Diversity and β-diversity of gut microbiota
After denoise and merge, the mean depth of sequences was 29,717 reads per sample and the number of features was 1203. α-Rarefaction (observed OTUs) curves at different sampling depths are shown in Fig. 2A. The curves reached the plateau phase by 21,109 reads, indicating that the sampling depths were sufficient to investigate following diversity analysis. α-Diversity (Shannon, Pieolu’s evenness, and Faith’s phylogenetic diversity (PD)) was analyzed to explore the richness and evenness of the gut microbiome in each rat. Although the median values of the Shannon index and Pielou’s evenness were the lowest in the HFT, there were no significant differences in the α-diversity of gut microbiota based on the three indices (Fig. 2B). Principal coordinate analyses (PCoA) were carried out to evaluate gut microbiome variation among the groups. As shown in the PCoA plots based on the weighted and unweighted UniFrac (Fig. 2C) distances, only PCoA of the unweighted UniFrac showed significant clustering of samples.
Fig. 2.

A α-Rarefaction curves based on the number of observed operational taxonomic units (OTUs). B Box plots for α-diversity (Shannon index, Pielou_E, and Faith_PD) of the bacterial communities in the rat cecum. C Principal coordinate analysis plots for β-diversity based on the distances of weighted and unweighted UniFrac. Pieloue_E, Pielou’s evenness; and Faith_PD, Faith’s phylogenetic diversity. Statistically significant differences in α-diversity between the groups were assessed by Kruskal–Wallis test with Dunn’s test. PERMANOVA test was used to measure dissimilarity of gut community between the groups based on the distance matrices of weighted and unweighted UniFrac (number of permutations: 999). CON (AIN-93G diet), HF (45% high-fat diet), HFB (HF + 0.6% (w/w) BR extract), HFT (HF + 0.15% (w/w) TMAO water), and HFTB (HFT + 0.6% (w/w) BR extract)
Gut microbial composition
Taxonomic analysis was performed to compare relative bacterial abundance in rat cecum among the groups at the phylum and genus levels. There was no significant difference among the 5 dominant phyla (data not shown). At genus level, Macellibacteroides was only detected in the rats fed TMAO with a high-fat diet (Fig. 3). The proportion of genera Mucispirilum and Paraeggerthella (Coriobacteraceae) increased in the rats fed TMAO with a high-fat diet (HFT and HFTB) compared to the HF (p < 0.01) and CON (p < 0.01), respectively. Dietary TMAO with a high-fat diet enriched the genera rc4-4 (Peptococcaceae) and Clostridium (Erysipelotrichaceae) compared to the CON, which was restored by the supplementation with BR extract. Relative abundance of unclassified Ruminococcaceae showed a significant decrease in the HFT compared to the CON (p < 0.05), HF (p < 0.05), and HFTB (p < 0.01). Unclassified S24-7 (p < 0.05) and RF32 (p < 0.01) in the HFB were reduced in their relative abundance compared to those in the HF.
Fig. 3.

Relative abundances of 8 operational taxonomic units at the genus level of gut microbiota in the cecal content of female rats fed a high-fat diet with trimethylamine-N-oxide (TMAO) and/or black raspberry (BR) extract. Statistically significant differences between the groups were assessed by Kruskal–Wallis test with Dunn’s test (*p < 0.05 and **p < 0.01). g_, genus; f_, family; and o_, order. Uncl., unclassified. g_Clostridium, Erysipelotrichaceae Clostridium. CON (AIN-93G diet), HF (45% high-fat diet), HFB (HF + 0.6% (w/w) BR extract), HFT (HF + 0.15% (w/w) TMAO water), and HFTB (HFT + 0.6% (w/w) BR extract)
We found that both dietary TMAO and BR extract could modulate the growths of specific gut bacteria in rats fed a high-fat diet. Yardeni et al. (2019) reported that the redox status in the gut plays a crucial role in the relative abundance of S24-7, Lachnospiraceae, Ruminococcaceae, and Clostridiales. Especially, aged mice, which are abundant in high reactive oxygen species (ROS), showed a significant increase in the relative abundance of S24-7 and a significant decrease in Ruminococcaceae (Yardeni et al., 2019). Since BR extract has a strong anti-oxidative activity, it could scavenge ROS in the gut environment, leading to modulation of the growths of Clostridium, S24-7, and Ruminococcaceae. Moreover, it was reported that Coriobacteriaceae, including Parraeggerthella and Adlercreutzia, can cleave polyphenol nucleus and improve the absorption of phenolic metabolites, enhancing the bioavailability of polyphenols (Espín et al., 2017). Therefore, it suggests that elevated levels of Clostridiaceae and Coriobacteriaceae by dietary TMAO might degrade polyphenols in BR extract so that their bioavailability can be improved. It indicates that the absorbed polyphenols can more circulate in the blood, directly and positively affecting cardiovascular health.
Bile acid profile
Since individual BAs showed differential activities for the receptors (Song et al., 2015), concentrations of 16 individual BAs in the rat cecum and serum were identified. In the cecum, dietary TMAO did not show any significant differences in the levels of individual BAs, while the high-fat diet seemed to exert changes in the levels of DCA (Fig. 4A). Among conjugated bile acids, the levels of Tα-MCA (p < 0.05) and TUDCA (p < 0.01) were significantly lower in the HFT than in the CON (Fig. 4B). In serum BA profile, dietary TMAO and a high-fat diet affect the concentrations of various kinds of BAs, including β-MCA, ω-MCA, and UDCA (Fig. 4C), and Tα-MCA and TUDCA (Fig. 4D). The HF and HFT showed a significant reduction in cecal levels of β-MCA, ω-MCA, and UDCA compared to the CON. Interestingly, meanwhile, the levels of UDCA (p = 0.067), Tα-MCA (p < 0.05), and TUDCA (p < 0.05), which were considered as Farnesoid X receptor (FXR)-antagonistic BAs, were higher in the HFTB than in the HFT.
Fig. 4.

Unconjugated and conjugated bile acids in the cecal content (A, B) and serum (C, D) of female rats fed a high-fat diet with trimethylamine-N-oxide (TMAO) and/or black raspberry (BR) extract. All data represent the means and standard errors of means. Statistically significant differences between the groups were assessed by Kruskal–Wallis test with Dunn’s test (*p < 0.05 and **p < 0.01). CON, AIN-93G diet; HF, 45% high-fat diet; HFB, HF + 0.6% (w/w) BR extract; HFT, HF + 0.15% (w/w) TMAO water; and HFTB, HFT + 0.6% (w/w) BR extract. MCA, muricholic acid; CA, cholic acid; CDCA, chenodeoxycholic acid; DCA, deoxycholic acid, LCA, lithocholic acid, UDCA, ursodeoxycholic acid; G, glycine-conjugated; and T, taurine-conjugated
Metabolites produced by gut microbiota, such as secondary BAs and SCFAs, can affect the pathogenesis of CVD (Ding et al., 2018; Kriaa et al., 2019; Poll et al., 2020; Wahlström et al., 2016). BAs can act as ligands for BA receptors and regulate expressions of genes involved in their synthesis, transport, conjugation, and excretion (Ding et al., 2018). Primary BAs are synthesized in hepatocytes and transformed into secondary BAs by gut microbiota. Therefore, abnormal changes in physiological status may commonly perturbate the gut environment, resulting in alteration of gut microbial composition, subsequently bile acid dysmetabolism (Winston and Theriot, 2020). In addition, BA composition is highly associated with cholesterol and bile acid metabolisms that may play critical roles in CVD development (Winston and Theriot, 2020). In this study, dietary TMAO did not affect BA composition in rat cecum. However, in the serum of the BR-fed groups, FXR-antagonistic BAs such as UDCA, Tα-MCA, and TUDCA increased. Likewise, Ding et al. (2018) reported 0.3% TMAO administration for 8 weeks induced aortic lesion development by altering BA profile in male ApoE−/− mice. Since various receptors that can bind BAs as hormones, including FXR, pregnane X receptor, vitamin D receptor, and G-protein-coupled bile acid receptor (TGR5), were differentially activated depending on the type of BAs (Parks et al., 1999), changes in the composition of BAs by the BR extract can be a crucial factor regulating cholesterol metabolism.
Short-chain fatty acid profile and hippuric acid level in the cecum
SCFAs as well as BAs are regarded as other major signaling molecules produced by gut bacteria. Among six SCFAs (acetate, propionate, butyrate, isobutyrate, valerate, and isovalerate), dietary TMAO with a high-fat diet increased the levels of acetate, propionate, isobutyrate, and isovalerate (Fig. 5A). However, the supplementation of BR extract did not show any change in the levels of SCFAs. Therefore, BR extract may lower LDL-C due to the changes in the composition of secondary microbial metabolites, mainly BAs, but not SCFAs.
Fig. 5.

Short-chain fatty acid profile (A) and hippuric acid concentration (B) in the cecal content of female rats fed a high-fat diet with trimethylamine-N-oxide (TMAO) and/or black raspberry (BR) extract. All data represent the means and standard errors of means. Different small letters above bars indicate significant differences among the groups (p < 0.05 by one-way ANOVA and Duncan’s multiple test). CON (AIN-93G diet), HF (45% high-fat diet), HFB (HF + 0.6% (w/w) BR extract), HFT (HF + 0.15% (w/w) TMAO water), and HFTB (HFT + 0.6% (w/w) BR extract)
We examined if the BR supplementation changed the concentrations of small phenolic metabolites in the gut environment. Among various candidates for phenolic metabolites, only hippuric acid significantly increased in the rats fed BR extract (Fig. 5B). Hippuric acid, one of the microbial metabolites converted from phenolic compounds, can be a metabolomic marker for microbial diversity due to the correlation between its increasing trend and diversity (Pallister et al., 2017). Furthermore, hippuric acid was also reported to be negatively correlated with the prevalence of metabolic syndrome (Pallister et al., 2017). The increased hippuric acid in cecum after consuming BR extract might be one of the contributing factors in lowering CVD risk.
Expression of the genes involved in cholesterol and bile acid metabolisms
To investigate the potential mechanism underlying the protective effect of BR extract in rats with elevated levels of serum TMAO and cholesterol, the mRNA expressions of the genes responsible for cholesterol and BA metabolisms in the liver tissue were explored.
The mRNA expressions of Cyp7a1 and Cyp27a1 were higher in the HFTB than in the other groups without significant differences (Fig. 6). Polyphenols have been reported to modulate hepatic Cyp7a1 expression and enhance reverse cholesterol transport (RCT) (Xu et al., 2010). On the other hand, Liang et al. (2020) reported that TMAO could inhibit RCT and cholesterol removal from peripheral macrophages and down-regulate hepatic expressions of Cyp7a1 and Cyp27a1. In this study, it seemed that the BR supplementation tended to upregulate hepatic Cyp7a1 expression due to the activity of increased levels of FXR-antagonistic BAs in serum. UDCA, TUDCA, and conjugated MCAs are considered as FXR antagonists that can suppress FXR signaling and promote hepatic Cyp7a1 expression (Zhang et al., 2018). FXR-fibroblast growth factor 15/19 (FGF15/19) signaling pathway is a well-known system to regulate cholesterol metabolism in a BA-dependent manner. Once ileal FXR is activated, FGF15/19 circulates and binds to fibroblast growth factor receptor 4, which rapidly inhibits mRNA expression of Cyp7a1 (Song et al., 2009).
Fig. 6.

Hepatic mRNA expressions of the genes responsible for cholesterol and bile acid metabolisms in female rats fed a high-fat diet with trimethylamine-N-oxide (TMAO) and/or black raspberry (BR) extract. Relative gene expressions were normalized to Gapdh expression. All data represent the means and standard errors of means. Different small letters above bars indicate significant differences among the groups (p < 0.05 by one-way ANOVA and Duncan’s multiple range test). CON (AIN-93G diet), HF (45% high-fat diet), HFB (HF + 0.6% (w/w) BR extract), HFT (HF + 0.15% (w/w) TMAO water), and HFTB (HFT + 0.6% (w/w) BR extract)
Among transcription factors that can activate transcription of the genes involved in the regulation of cholesterol and BAs including FXR and sterol regulatory element-binding protein 2 (SREBP-2) (Nr1h4 and Srebf2, respectively), the administration of TMAO tended to upregulate the mRNA expressions of Nr1h4 and Srebf2, while the supplementation of BR extract tended to down-regulate them, although not significant. The mRNA expression of hmgcr showed a similar tendency to the result of Srebf2. Among the genes involved in cholesterol and BA transporters, the mRNA expressions of ATP-binding cassette transporter 5 (ABCG5) and bile salt export pump (BSEP) (Abcg5 and Abcb11, respectively) were measured. Among them, the mRNA expression of Abcg5, a transporter facilitating the excretion of cholesterol into bile (Molusky et al., 2018), significantly increased in the HFTB. The upregulated expression of ABCG5 can contribute to avoiding hypercholesterolemia and atherosclerosis (Seeram, 2008). Similar to our result, polyphenol-rich extract from black chokeberry was reported to modulate mRNA expressions of Srebf2 and Abcg5 in Caco-2 cells (Kim et al., 2013). FMO3 is an enzyme converting TMAO from TMA in liver (Liang et al., 2020). In this study, TMAO itself did not change Fmo3 expression. Hepatic Fmo3 expression was not affected by the BR supplementation as well.
In conclusion, the dietary administration of BR extract affected lipid profile, gut microbial composition, microbial metabolites, and hepatic gene expressions in the rats fed TMAO with a high-fat diet. The possible mechanism is that BR extract consumption can affect gut microbial composition and microbial meatoblites shifted by TMAO in a positive way, leading to lowering serum LDL-C. To be specific, the BR supplementation could restore gut microbial composition, such as Macellibacteroides, Clostridium, and Ruminococcaceae and increase cecal hippuric acid and serum FXR-antagonistic BAs. Collectively, BR extract may regulate cholesterol efflux and clearance by modifying the gut microbiome and subsequently affecting microbial metabolites and cholesterol metabolism.
Supplementary Information
Below is the link to the electronic supplementary material.
Acknowledgements
This research was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (NRF-2017R1D1A1B03028407).
Declarations
Conflict of interest
The authors declare no conflict of interest.
Footnotes
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Taehwan Lim, Email: imtae86@snu.ac.kr.
Kiuk Lee, Email: leku@snu.ac.kr.
Ryun Hee Kim, Email: ryunheekim@snu.ac.kr.
Kwang Hyun Cha, Email: chakh79@kist.re.kr.
Song Yi Koo, Email: ninesong2@kist.re.kr.
Eun Chae Moon, Email: luna5833@snu.ac.kr.
Keum Taek Hwang, Email: keum@snu.ac.kr.
References
- Bolyen E, Rideout JR, Dillon MR, Bokulich NA, Abnet CC, Al-Ghalith GA, Alexander H, Alm EJ, Arumugam M, Asnicar F. Reproducible, interactive, scalable and extensible microbiome data science using QIIME2. Nature Biotechnology. 2019;37:852–857. doi: 10.1038/s41587-019-0209-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Callahan BJ, McMurdie PJ, Rosen MJ, Han AW, Johnson AJA, Holmes SP. DADA2 is an open-source software package that denoises and removes sequencing errors from Illumina amplicon. Nature Methods. 2016;13:581–583. doi: 10.1038/nmeth.3869. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen ML, Yi L, Zhang Y, Zhou X, Ran L, Yang J, Zhu JD, Zhang QY, Mi MT. Resveratrol attenuates trimethylamine-N-oxide (TMAO)-induced atherosclerosis by regulating TMAO synthesis and bile acid metabolism via remodeling of the gut microbiota. mBio. 2016;7:e02210–e2215. doi: 10.1128/mBio.02210-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Ferrars RM, Czank C, Zhang Q, Botting NP, Kroon PA, Cassidy A, Kay CD. The pharmacokinetics of anthocyanins and their metabolites in humans. British Journal of Pharmacology. 2014;171:3268–3282. doi: 10.1111/bph.12676. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ding L, Chang M, Guo Y, Zhang L, Xue C, Yanagita T, Zhang T, Wang Y. Trimethylamine-N-oxide (TMAO)-induced atherosclerosis is associated with bile acid metabolism. Lipids in Health and Disease. 2018;17:286. doi: 10.1186/s12944-018-0939-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Espín JC, González-Sarrías A, Tomás-Barberán FA. The gut microbiota: A key factor in the therapeutic effects of (poly)phenols. Biochemical Pharmacology. 2017;139:82–93. doi: 10.1016/j.bcp.2017.04.033. [DOI] [PubMed] [Google Scholar]
- Friedewald WT, Levy RI, Fredrickson DS. Estimation of the concentration of low-density lipoprotein cholesterol in plasma, without use of the preparative ultracentrifuge. Clinical Chemistry. 1972;18:499–502. doi: 10.1093/clinchem/18.6.499. [DOI] [PubMed] [Google Scholar]
- He Z, Lei L, Kwek E, Zhao Y, Liu J, Hao W, Zhu H, Liang N, Ma KY, Ho HM, He W, Chen Z. Ginger attenuates trimethylamine-N-oxide (TMAO)-exacerbated disturbance in cholesterol metabolism and vascular inflammation. Journal of Functional Foods. 2019;52:25–33. doi: 10.1016/j.jff.2018.10.022. [DOI] [Google Scholar]
- Hu Y, Zhao Y, Yuan L, Yang X. Protective effects of tartary buckwheat flavonoids on high TMAO diet-induced vascular dysfunction and liver injury in mice. Food and Function. 2015;6:3359–3372. doi: 10.1039/C5FO00581G. [DOI] [PubMed] [Google Scholar]
- Jung H, Kwak HK, Hwang KT. Antioxidant and antiinflammatory activities of cyanidin-3-glucoside and cyanidin-3-rutinoside in hydrogen peroxide and lipopolysaccharide-treated RAW264.7 cells. Food Science and Biotechnology. 2014;23:2053–2062. doi: 10.1007/s10068-014-0279-x. [DOI] [Google Scholar]
- Jung H, Lee HJ, Cho H, Hwang KT. Anti-inflammatory activities of Rubus fruit anthocyanins in inflamed human intestinal epithelial cells. Journal of Food Biochemistry. 2015;39:300–309. doi: 10.1111/jfbc.12130. [DOI] [Google Scholar]
- Kim B, Park Y, Wegner CJ, Bolling BW, Lee J. Polyphenol-rich black chokeberry (Aronica melanocarpa) extract regulates the expression of genes critical for intestinal cholesterol flux in Caco-2 cells. The Journal of Nutritional Biochemistry. 2013;24:1564–1570. doi: 10.1016/j.jnutbio.2013.01.005. [DOI] [PubMed] [Google Scholar]
- Koeth RA, Wang Z, Levison BS, Buffa JA, Org E, Sheehy BT, Britt EB, Fu X, Wu Y, Li L, Smith JD, DiDonato JA, Chen J, Li H, Wu GD, Lewis JD, Warrier M, Brown JM, Krauss RM, Tang WHW, Bushman FD, Lusis AJ, Hazen SL. Intestinal microbiota metabolism of L-carnitine, a nutrient in red meat, promotes atherosclerosis. Nature Medicine. 2013;19:576–585. doi: 10.1038/nm.3145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krga I, Milenkovic D. Anthocyanins: From sources and bioavailability to cardiovascular health benefits and molecular mechanisms of action. Journal of Agricultural and Food Chemistry. 2019;67:1771–1783. doi: 10.1021/acs.jafc.8b06737. [DOI] [PubMed] [Google Scholar]
- Kriaa A, Bourgin M, Potiron A, Mkaouar H, Jablaoui A, Gérard P, Maguin E, Rhimi M. Microbial impact on cholesterol and bile acid metabolism: Current status and future prospects. Journal of Lipid Research. 2019;60:323–332. doi: 10.1194/jlr.R088989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kula M, Krauze-Baranowska M. Rubus occidentalis: The black raspberry—Its potential in the prevention of cancer. Nutrition and Cancer. 2016;68:18–28. doi: 10.1080/01635581.2016.1115095. [DOI] [PubMed] [Google Scholar]
- Liang Xi, Zhang Zhe, Lv Youyou, Tong Linjun, Liu Tongjie, Yi Huaxi, Zhou Xiaohong, Yu Zhuang, Tian Xiaoying, Cui Qingyu, Yang Jiebing, Gong Pimin, Zhang Lanwei. Reduction of intestinal trimethylamine by probiotics ameliorated lipid metabolic disorders associated with atherosclerosis. Nutrition. 2020;1:79–80. doi: 10.1016/j.nut.2020.110941. [DOI] [PubMed] [Google Scholar]
- Lim T, Ryu J, Lee K, Park SY, Hwang KT. Protective effects of black raspberry (Rubus occidentalis) extract on hypercholesterolemia and hepatic inflammation in rats fed high-fat and high-choline diets. Nutrients. 2020;12:2448. doi: 10.3390/nu12082448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Molusky MM, Hsieh J, Lee SX, Ramakrishnan R, Tascau L, Haeusler RA, Accili D, Tall AR. Metformin and AMP Kinase activation increase expression of the sterol transporters ABCG5/8 (ATP-binding cassette transporter G5/G8) with potential antiatherogenic consequences. Atherosclerosis, Thrombosis, and Vascular Biology. 2018;38:1493–1503. doi: 10.1161/ATVBAHA.118.311212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pallister T, Jackson MA, Martin TC, Zierer J, Jennings A, Mohney RP, MacGregor A, Steves CJ, Cassidy A, Spector TD, Menni C. Hippurate as a metabolomic marker of gut microbiome diversity: Modulation by diet and relationship to metabolic syndrome. Scientific Reports. 2017;7:13670. doi: 10.1038/s41598-017-13722-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parks DJ, Blanchard SG, Bledsoe RK, Chandra G, Consler TG, Kliewer SA, Stimmel JB, Wilson TM, Zavacki AM, Moore DD, Lehmann JM. Bile acids: Natural ligands for an orphan nuclear receptor. Science. 1999;284:1365–1368. doi: 10.1126/science.284.5418.1365. [DOI] [PubMed] [Google Scholar]
- Poll BG, Cheema MU, Pluznick JL. Gut microbial metabolites and blood pressure regulation: Focus on SCFAs and TMAO. Physiology. 2020;35:275–284. doi: 10.1152/physiol.00004.2020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ren D, Liu Y, Zhao Y, Yang X. Hepatotoxicity and endothelial dysfunction induced by high choline diet and the protective effects of phloretin in mice. Food and Chemical Toxicology. 2016;94:203–212. doi: 10.1016/j.fct.2016.06.004. [DOI] [PubMed] [Google Scholar]
- Seldin MM, Meng Y, Qi H, Zhu W, Wang Z, Hazen SL, Lusis AJ, Shih DM. Trimethylamine N-oxide promotes vascular inflammation through signaling of mitogen-activated protein kinase and nuclear factor-κB. Journal of the American Heart Association. 2016;5:e002767. doi: 10.1161/JAHA.115.002767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Song Kwang-Hoon, Li Tiangang, Owsley Erika, Strom Stephen, Chiang John Y. L. Bile acids activate fibroblast growth factor 19 signaling in human hepatocytes to inhibit cholesterol 7α-hydroxylase gene expression. Hepatology. 2009;49(1):297–305. doi: 10.1002/hep.22627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Summers G., Wibisono R. D., Hedderley D. I., Fletcher G. C. Trimethylamine oxide content and spoilage potential of New Zealand commercial fish species. New Zealand Journal of Marine and Freshwater Research. 2017;51(3):393–405. doi: 10.1080/00288330.2016.1250785. [DOI] [Google Scholar]
- Tang WH, Wang Z, Levison BS, Koeth RA, Britt EB, Fu X, Wu Y, Hazen SL. Intestinal microbial metabolism of phosphatidylcholine and cardiovascular risk. New England Journal of Medicine. 2013;368:1575–1584. doi: 10.1056/NEJMoa1109400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tulio AZ, Jr, Reese RN, Wyzgoski FJ, Rinaldi PL, Fu R, Scheerens JC, Miller AR. Cyanidin-3-rutinoside and cyanidin-3-xylosylrutinoside as primary phenolic antioxidants in black raspberry. Journal of Agricultural and Food Chemistry. 2008;56:1880–1888. doi: 10.1021/jf072313k. [DOI] [PubMed] [Google Scholar]
- Wahlström A, Sayin SI, Marschall H, Bäckhed F. Intestinal crosstalk between bile acids and microbiota and its impact on host metabolism. Cell Metabolism. 2016;24:41–50. doi: 10.1016/j.cmet.2016.05.005. [DOI] [PubMed] [Google Scholar]
- Wang Z, Klipfell E, Bennett BJ, Koeth R, Levison BS, DuGar B, Feldstein AE, Britt EB, Fu X, Chung YM. Gut flora metabolism of phosphatidylcholine promotes cardiovascular disease. Nature. 2011;472:57–63. doi: 10.1038/nature09922. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang Z, Bergeron N, Levison BS, Li XS, Chiu S, Jia X, Koeth RA, Li L, Wu Y, Tang WH, Krauss RM, Hazen SL. Impact of chronic dietary red meat, white meat, or non-meat protein on trimethylamine N-oxide metabolism and renal excretion in healthy men and women. European Heart Journal. 2019;40:583–594. doi: 10.1093/eurheartj/ehy799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Winston JA, Theriot CM. Diversification of host bile acids by members of the gut microbiota. Gut Microbes. 2020;11:158–171. doi: 10.1080/19490976.2019.1674124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu ZJ, Fan JG, Ding XD, Qiao L, Wang GL. Characterization of high-fat, diet-induced, non-alcoholic steatohepatitis with fibrosis in rats. Digestive Diseases and Sciences. 2010;55:931–940. doi: 10.1007/s10620-009-0815-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yardeni T, Tanes CE, Bittinger K, Mattei LM, Schaefer PM, Singh LN, Wu GD, Murdock DG, Wallace DC. Host mitochondria influence gut microbiome diversity: A role for ROS. Science Signaling. 2019;12:eaaw3159. doi: 10.1126/scisignal.aaw3159. [DOI] [PubMed] [Google Scholar]
- Yin X, Gibbons H, Rundle M, Frost G, McNulty BA, Nugent AP, Walton J, Flynn A, Brennan L. The relationship between fish intake and urinary trimethylamine-N-oxide. Molecular Nutrition and Food Research. 2020;64:1900799. doi: 10.1002/mnfr.201900799. [DOI] [PubMed] [Google Scholar]
- Yu ZL, Ding L, Xue CH, Zhang TT, Wang YM. Dietary trimethylamine N-oxide exacerbated atherosclerosis under a low-fat rather than high-fat diet. Journal of Agricultural and Food Chemistry. 2020;68:6789–6791. doi: 10.1021/acs.jafc.0c03190. [DOI] [PubMed] [Google Scholar]
- Zhu W, Gregory JC, Org E, Buffa JA, Gupta N, Wang Z, Li L, Fu X, Wu Y, Mehrabian M. Gut microbial metabolite TMAO enhances platelet hyperreactivity and thrombosis risk. Cell. 2016;24:111–124. doi: 10.1016/j.cell.2016.02.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.