

Abstract
Human actions are altering ecosystems worldwide. Among human‐released pollutants, ionizing radiation arises as a rare but potentially devastating threat to natural systems. The Chornobyl accident (1986) represents the largest release of radioactive material to the environment. Our aim was to examine how exposure to radiation from the Chornobyl accident influences dorsal skin coloration of Eastern tree frog (Hyla orientalis) males sampled across a wide gradient of radioactive contamination in northern Ukraine. We assessed the relationship between skin frog coloration (which can act as a protective mechanism against ionizing radiation), radiation conditions and oxidative stress levels. Skin coloration was darker in localities closest to areas with high radiation levels at the time of the accident, whereas current radiation levels seemed not to influence skin coloration in Chornobyl tree frogs. Tree frogs living within the Chornobyl Exclusion Zone had a remarkably darker dorsal skin coloration than frogs from outside the Zone. The maintenance of dark skin coloration was not linked to physiological costs in terms of frog body condition or oxidative status, and we did not detect short‐term changes in frog coloration. Dark coloration is known to protect against different sources of radiation by neutralizing free radicals and reducing DNA damage, and, particularly melanin pigmentation has been proposed as a buffering mechanism against ionizing radiation. Our results suggest that exposure to high levels of ionizing radiation, likely at the time of the accident, may have been selected for darker coloration in Chornobyl tree frogs. Further studies are needed to determine the underlying mechanisms and evolutionary consequences of the patterns found here.
Keywords: animal colouration, Chernobyl, environmental pollution, evolutionary physiology, melanism, oxidative stress, radioactivity
1. INTRODUCTION
Habitat destruction, climate change and pollution are key factors damaging natural systems nowadays (Rands et al., 2010). Pollutants, in particular, are widespread in nature and represent one of the more powerful forces of ecological and evolutionary change (Palumbi, 2001; Spurgeon et al., 2020). Strong selective factors can induce fast adaptive responses, and signs of rapid adaptation to some pollutants have been observed across many taxa (Nacci et al., 2016; Pitelka, 1988; Reid et al., 2016), although these responses are far from generalized (e.g. Brady et al., 2017; Loria et al., 2019). Among the large diversity of pollutants introduced into the environment, radioactive contamination caused by human activities generates broad societal and scientific concern (e.g. Beresford & Copplestone, 2011). The release of radionuclides from nuclear power plants, as occurred after the accidents in Chornobyl (Ukraine) in 1986, and Fukushima (Japan) in 2011, represents the most evident case of public concern about the impact that ionizing radiation can have on living organisms.
The accident at the Chornobyl nuclear power plant on the 26 April 1986 caused the largest release of radioactive material to the environment in human history (UNSCEAR, 1988). Hence, the Chornobyl area constitutes a key scenario for evaluating the eco‐evolutionary consequences of the exposure to ionizing radiation on wildlife. The exposure to extremely high levels of radiation after the Chornobyl accident caused rapid environmental damage (Beresford et al., 2016; Møller & Mousseau, 2006; Sazykina & Kryshev, 2006; Yablokov et al., 2009). Such acute exposure to radiation negatively affected the physiology, morphology and genomics of different species inhabiting the Chornobyl area (e.g. Kryshev et al., 2005; Møller & Mousseau, 2006, 2015, 2016; Yablokov et al., 2009). However, recent studies have reported the presence of large and diverse animal communities, a lack of negative effects of current levels of radiation in many taxa, and even signs of adaptation to radiation (e.g. Burraco, Bonzom, et al., 2021; Deryabina et al., 2015; Galván et al., 2014; Jernfors et al., 2018; Kovalchuk et al., 2004; Møller & Mousseau, 2016; Schlichting et al., 2019). Thus, more than three decades after the accident, there is still high scientific discrepancy about the long‐term effects of the accident on wildlife (Beresford et al., 2020).
Ionizing radiation is harmful because it can damage DNA and other biomolecules, causing cell malfunctions and increasing mortality risk (Alizadeh et al., 2015; Lehnert, 2007; Mothersill & Seymour, 2014). Different mechanisms, such as DNA damage repair pathways have been proposed as candidates that may facilitate life in radio‐contaminated environments. Organisms exposed to ionizing radiation can plastically upregulate the expression of these repair and protection mechanisms (e.g. Jernfors et al., 2018; Kesäniemi et al., 2019; Mustonen et al., 2018). However, these processes can be physiologically costly and unable to fully buffer the impact of radiation, which often results in deleterious mutations or lower individual performance. An alternative protection from ionizing radiation may come through changes in pigmentation. In animals, coloration plays a key role in several ecological functions such as sexual selection, defence from predators and health maintenance (Caro, 2005; Cuthill et al., 2017; Mackintosh, 2001). Previous studies have also revealed that melanin‐based coloration can mitigate the impact of different forms of radiation, including ionizing radiation (Cordero & Casadevall, 2020; Dadachova et al., 2007, 2008; Dadachova & Casadevall, 2008). However, whilst melanization has been showed to have a protective role against ionizing radiation in organisms with relatively simple level of organization, such as fungi, evidence suggests that costs of melanin‐based colorations may be often higher than benefits in wild vertebrates (Galván et al., 2011, 2014).
Amphibians occupy both the terrestrial and the aquatic environments during their life cycle and are, thus, exposed to a full range of radiation sources in contaminated environments. Furthermore, they often show low vagility and are highly philopatric to their natal ponds (Cayuela et al., 2020), which facilitates the estimation of their potential exposure to radiation and makes them ideal subjects for evaluating the effects of radioactive contamination on wild vertebrates. However, work on amphibians in radio‐contaminated environments is still scarce (Orizaola, 2022). Recent studies have reported, for example, that radiation exposure was linked to an increase in mutation rates and mitochondrial DNA damage in frogs from radio‐contaminated areas (Car et al., 2022; Gombeau et al., 2020), but a lack of effects of radiation exposure on physiological biomarkers in Chornobyl tree frogs (Burraco, Bonzom, et al., 2021). Here, we examined the differences in dorsal skin coloration of Eastern tree frogs (Hyla orientalis) living across a large gradient of radioactive contamination around Chornobyl, and whether the maintenance of skin coloration is physiologically costly in terms of oxidative stress.
Based on the putative protective role of melanin against radiation, we predicted that frogs living in (or near) areas with high radiation levels would present a darker skin coloration, which may suggest that radiation acted as a selective pressure on that trait. Since coloration is known to quickly change in some amphibians during the breeding season (Hettyey et al., 2009; Nilsson Sköld et al., 2013), we empirically examined the short‐term lability of coloration through a small laboratory‐based experiment. Finally, we explored the links between coloration, radiation and oxidative stress. Ionizing radiation is predicted to induce an oxidative stress state in cells (Einor et al., 2016; Galván et al., 2014), a process that may be buffered by the production of melanin or accentuated by the physiological costs of producing pigmentation. However, as eumelanin is the main pigment involved in the production of dark coloration in frogs, and a few studies support that its production seems not to be oxidatively costly (Frost‐Mason & Mason, 1996; Galván et al., 2011; Prota, 1992), we predicted that darker frogs would not experience significant oxidative stress.
2. MATERIAL AND METHODS
2.1. Field sampling
We examined skin coloration on the Eastern tree frog (Hyla orientalis), a cryptic species of the European tree frog (Hyla arborea) group that inhabits from the Caspian to the Baltic Sea. Females start to breed at 2–3 years of age (Özdemir et al., 2012), which means that 10–15 generations have passed since the Chornobyl accident. The species usually presents a light green coloration (https://amphibiaweb.org/species/7317).
During three consecutive breeding seasons (2017–2019), we collected reproductive adult males of H. orientalis in ponds located in a 2.200 km2 area around Chornobyl (Northern Ukraine, Figure 1; Supporting Information). We sampled frogs in twelve localities: eight within the Chornobyl Exclusion Zone and four outside Chornobyl in an area situated ca. 40 km to the East of the Exclusion Zone and currently exposed only to background radiation levels (Figure 1; Table S1). These two areas are separated by a system of flood plains, channels, and rivers (maximum crossing distance for a frog ca. 100 m) and moist soils (see USDA Foreign Agricultural Service https://ipad.fas.usda.gov/cropexplorer/imageview.aspx?regionid=umb; or Copernicus Soil Moisture Index: https://land.copernicus.eu/global/products/ssm). Both areas are located within the Polesia woodland ecological zone biome (Fileccia et al., 2014), and our previous studies have revealed no geographic barriers, and the existence of gene flow, between H. orientalis inhabiting localities within these two areas (Car et al., 2022). Field and laboratory work were designed to reduce differences in handling time, temperature or experimental procedures between localities and areas. All localities were medium‐small wetlands with reed beds, within a matrix of forest and meadows, and located within the same soil type and color: soddy‐podzolized sandy and clay‐sandy soils (Soil Map of Ukraine, https://esdac.jrc.ec.europa.eu/content/title‐russia‐soil‐map‐ukraine). Water pH values, which may be associated with changes in coloration (Ancans et al., 2001), did not differ between localities within and outside the Chornobyl Exclusion Zone (average ± SE: 6.9 ± 0.17 for localities within Chornobyl, 7.2 ± 0.24 for localities outside Chornobyl; χ 2 [1,12] = 1.66, p = 0.226). Temperature during the sampling days did not differ between localities situated within and outside the Chornobyl Exclusion Zone for both daily maximum and minimum values (average ± SE: 20.0 ± 1.0 and 9.3 °C ± 0.9, respectively, for localities within Chornobyl; 22.5 ± 1.6 and 12.2 ± 1.5 °C, respectively, for localities outside Chornobyl; χ 2(1, 12) = 1.77, p = 0.210 and χ 2(1, 12) = 2.60, p = 0.133 for maximum and minimum temperature, respectively).
FIGURE 1.
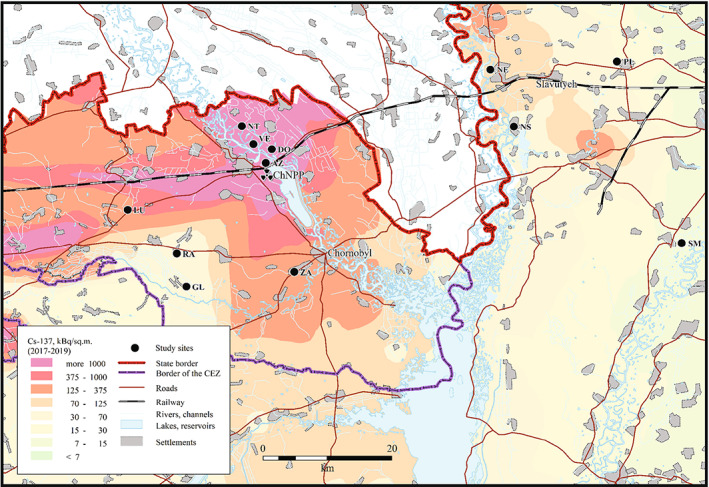
Map of the studied eastern tree frog (Hyla orientalis) locations (see also Table S1). The underlying 137Cs soil data (decay corrected to spring 2017–2019) is derived from the atlas of radioactive contamination of Ukraine (Intelligence Systems GEO, 2011).
In total, we examined 189 H. orientalis adult males, captured during active calling from 10 PM to 1 AM. Once captured, we placed frogs in plastic bags and transported them to the laboratory where we stored them individually in small plastic buckets with perforated lids, containing ca. 3 cm of water, and kept them in darkness during the rest of the night. On the next morning, we took a photograph of each individual for colorimetric evaluation (see below), we recorded morphological measurements (snout‐to‐vent length, body depth and width) using a calliper to the nearest 1 mm, and weighted each individual using a precision balance to the nearest 0.01 g. Morphometric measurements were used to estimate body condition index and individual dose rates (see below). All animals were collected, and experimental procedures were conducted, under permit of Ministry of Ecology and Natural Resources of Ukraine (No. 517, 21.04.2016).
2.2. Skin colour measurements
We quantified dorsal skin coloration of tree frogs following Troscianko and Stevens (2015). Individuals were placed on a black background and beside a ColorChecker passport (X‐Rite, Inc). To provide a uniform light spectrum, we used two LED lamps (Northlight Isac 5 W, 300 lm, 3000 K), placing one at each side of the frog. We took dorsal images of individuals placed in this setting using a Fuji XT‐1 digital camera with a Fujinon XF 35 mm F2 R WR lens, tripod‐mounted and provided with a Fujifilm RR‐100 remote release. We manually set lens aperture to f5.6, and ISO to 1250. All images were saved as. RAF, a file format containing fully uncompressed photography data. We linearized images with the help of a ColorChecker chart, and using the Multispectral Image Calibration and Analysis Toolbox for ImageJ software (Mica toolbox version 2.2.2) developed by Troscianko and Stevens (2015). Once completed the linearization process, we transformed images to greyscale and selected five squares of ca. 4 mm2 along the dorsal skin of frogs (two in the front part, two in the mid‐lateral part, and one in the back of the body) avoiding areas where light was reflected directly back (i.e. specular reflectance). We quantified dorsal coloration as luminance using the function Pattern Color and Luminance Measurements included in the Mica toolbox ImageJ plug‐in (Troscianko & Stevens, 2015). Luminance is an integrative trait that specifically describes the perceived lightness of an individual (see e.g. Carter et al., 2020). Luminance values were scaled to a range of 0–100% in reflectance, corresponding to the black and white luminance standards, respectively. For each individual, we calculated a single value for dorsal skin luminance, as the average luminance of the five selected areas.
2.3. Skin colour lability
Male frog coloration can change quickly in function of intrinsic and extrinsic factors such as the intensity of male calling activity or in response to some environmental conditions (e.g. Hettyey et al., 2009; Nilsson Sköld et al., 2013). To determine whether tree frog skin luminance can change over a short time period and how it may be affected by environmental background coloration, in May 2019, we collected 14 H. orientalis males from a single location within the Chornobyl Exclusion Zone (Azbuchyn, AZ, one of the localities with the highest variation in luminance; Figure 1) and kept them under contrasting experimental backgrounds. We transported the frogs to the laboratory and maintained them for 48 h into 1‐L plastic buckets with the walls covered with either a white (n = 7) or black (n = 7) plastic film (except the lid that was transparent and perforated to allow light and air to pass through) to simulate extreme background conditions. Buckets contained ca. 3 cm of water in order to maintain moisture. We photographed each individual at the start and at the end (48‐h later) of the experiment, and dorsal skin luminance was quantified as indicated above.
2.4. Oxidative stress
We determined the activity of three antioxidant enzymes (catalase, CAT; glutathione reductase, GR; and glutathione peroxidase, GPX) and lipid peroxidation (malondialdehyde concentration, MDA) in the liver of H. orientalis males inhabiting inside the Chornobyl Exclusion Zone. Antioxidant enzymes are present in different cellular compartments and have an essential role in transforming reactive oxygen substances produced during catabolism into more stable and non‐toxic molecules (Halliwell & Gutteridge, 2015). MDA is the main product formed during the peroxidation of membrane polyunsaturated fatty acids and is considered a marker of oxidative damage in lipids (Del Rio et al., 2005).
To explore the relationship between the frog redox status, skin coloration and radiation, we quantified oxidative stress parameters in individuals collected within the Chornobyl Exclusion Zone in 2018. Due to logistical limitations, we did not quantify oxidative levels in individuals inhabiting outside the Chornobyl Exclusion Zone. For the relationship between oxidative stress and coloration, high luminance values (i.e. brighter frogs, the most common coloration of the species) were considered as a reference. For the relationship between oxidative stress and radiation, individuals with low absorbed dose rates were considered as the reference ones, following the approach used in previous radioecological research in Chornobyl (e.g. Mappes et al., 2019; Webster et al., 2016). Once morphometric measurements were recorded, we euthanized frogs by pithing without decapitation (AVMA, 2020). We collected the liver of each individual and stored them at −80°C until assayed. Livers were immersed in a buffered solution (Burraco et al., 2017) to avoid proteolysis in a proportion tissue:solution of 1:4.5, and homogenized at 35,000 rpm. The homogenates were centrifuged at 20,817 g for 30 min at 4°C, and the supernatants were used to estimate levels of the oxidative stress markers. As the activity of each antioxidant enzyme is relative to the amount of protein contained in the sample, we first determined total protein content according to Bradford's procedure (Bradford, 1976). We quantified catalase activity according to Cohen and Somersonm (1969). This method uses potassium permanganate (KMnO4) as an oxidizing agent and coloured component of the reducing agent H2O2. Five minutes after adding KMnO4, we quantified the reduction in this compound at a wavelength of 490 nm. We quantified glutathione peroxidase and glutathione reductase following Paglia and Valentine (1967) and Cribb et al. (1989), respectively, measuring in both cases NADPH oxidation at a wavelength of 340 nm. Finally, we quantified malondialdehyde (MDA) by measuring the red colour formed after the reaction of MDA with thiobarbituric acids, at a wavelength of 535 nm and according to Buege and Aust (1978).
2.5. Exposure to radiation
To estimate current exposure to radiation, we quantified total dose rates absorbed by each frog (in μGy/h). Briefly, we estimated total activity of 90Sr and 137Cs by integrating radionuclide measurements with body mass of each individual, considering the relative mass of bones (10%) and muscles (69%). We combined radionuclide activity concentrations in frogs, soil, and water with dose coefficients (in μGy/h per Bq per unit of mass). Dose coefficients for H. orientalis were calculated for internal and external exposure using a theoretical habitat use scenario for the species during the breeding season (Burraco, Car, et al., 2021) with the help of EDEN v3 IRSN software (Beaugelin‐Seiller et al., 2006). For each frog, total individual dose rate was calculated by summing internal and external dose rates (see full details in Burraco, Car, et al., 2021). To estimate historical exposure to radiation in the studied localities, we calculated the distance from each place to the closest area with historical high radiation level, using as reference areas with 137Cs contamination levels above 3700 kBq/m2 in May 1986 (Cort et al., 2009).
2.6. Statistical analyses
All analyses were conducted in R version 3.6.1. We explored the relationship between historical and current radiation levels on dorsal skin luminance in tree frogs. We first measured the distance of sampling locations to the closest historical high radiation area (137Cs levels >3700 kBq/m2 in May 1986) using the maps of the Atlas of caesium deposition on Europe after the Chernobyl accident (Cort et al., 2009). As distance of sampling locations to the closest historical high radiation area and current individual dose rates were highly correlated (R 2 = 0.86), we conducted a linear regression between luminance and distance to the closest historical high radiation area and individual dose rate, including body condition as a covariate and locality as a random factor. We also conducted a linear regression between luminance and individual dose rate in individuals inhabiting the Chornobyl Exclusion Zone. Finally, to examine possible differences between sampling areas (i.e. Chornobyl Exclusion Zone and Outside Chornobyl) on the dorsal skin luminance of tree frogs, we ran a linear mixed model including luminance as the dependent variable, sampling area as the independent variable, body condition as a covariate, and sampling location as a random factor. We added 0.1 unit to each value of distance to the closest historical high radiation area and individual dose rate value, to avoid 0 values that prevent log transformations. We estimated body condition as the residuals of the regression between snout‐to‐vent‐length and body mass values (Green, 2001).
To check for the possible effect of time and experimental background conditions on short‐term changes in dorsal skin coloration, we ran a linear mixed model including luminance as a dependent variable, the interaction between time (0 or 48 h) and background (black or white) as independent variables, body condition as a covariate, and individual as a random factor accounting for within‐individual measurements. Finally, we conducted linear mixed regressions between dorsal skin luminance or total individual dose rate and each oxidative stress parameter (CAT, GR, GPX, and MDA, all log transformed), including body condition as a covariate, and sampling location as a random factor. Oxidative stress values were scaled to report comparable estimates in linear regressions. Data were plotted using the function ggplot included in the package ggplot2 (version 3.3.3).
3. RESULTS
Dorsal skin luminance varied substantially across sampling localities, ranging between 4.2 and 63.9 (4.2–45.6 and 22.4–63.9, within and outside the Chornobyl Exclusion Zone, respectively; Figure 2). In individuals inhabiting within and outside the Chornobyl Exclusion Zone, skin luminance was positively correlated with the distance to the closest area with historical high radiation level (χ 2 (189) = 7.77, p = 0.005; Figure 3a), but not to individual dose rate (χ 2 [189] = 2.24, p = 0.134). Radiation levels currently experienced by tree frogs living in the Chornobyl Exclusion Zone (i.e. total individual dose rates) were not associated with variation in dorsal skin luminance (χ 2 [1147] = 0.30, p = 0.586; Figure 3b). There was no effect of male body condition (proxy for health and fitness in amphibians) on dorsal skin luminance (p = 0.26). Overall, Eastern tree frogs (Hyla orientalis) living within the Chornobyl Exclusion Zone had 43.6% lower dorsal skin luminance, on average, than frogs living outside Chornobyl (χ 2 [189] = 27.18, p < 0.001; Figure 2a,b).
FIGURE 2.
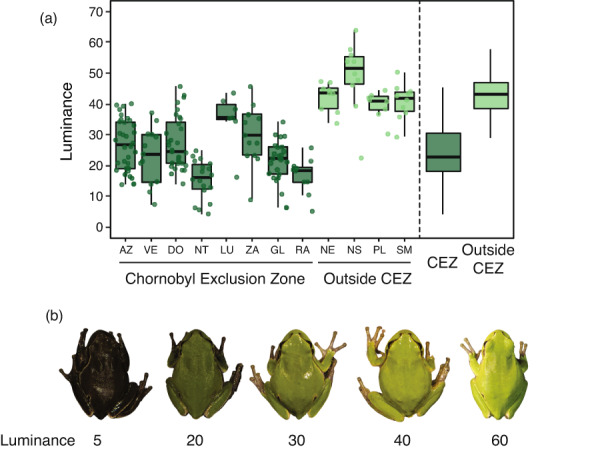
(a) Dorsal skin luminance in eastern tree frog (Hyla orientalis) males inhabiting across a gradient of radiation inside (CEZ) or outside the Chornobyl exclusion zone (outside CEZ). (b) Range of dorsal skin luminance in H. orientalis males (from left to right: Luminance values of 5, 20, 30, 40 and 60).
FIGURE 3.
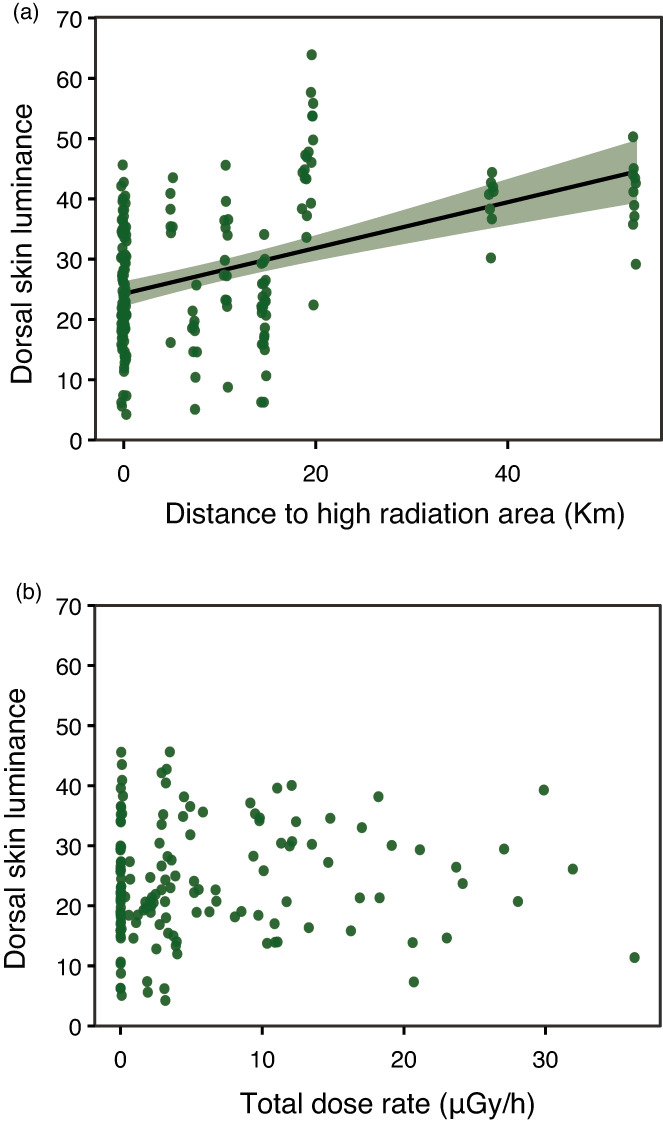
Correlation between dorsal skin luminance in eastern tree frog (Hyla orientalis) males and, (a) distance to the closest area with historical high radiation levels (137Cs levels >3700 kBq/m2 in May 1986), and (b) total individual dose rates of sampled individuals within the Chornobyl Exclusion Zone.
We did not find any correlation between skin luminance and oxidative stress markers (CAT: χ 2(1, 75) = 0.32, p = 0.574; GR: χ 2 (1, 77) = 1.06, p = 0.304; GPX: χ 2 (1, 78) = 0.76, p = 0.382; MDA: χ 2 (1, 81) = 0.59, p = 0.443), but we detected a negative correlation between individual dose rate and lipid peroxidation (i.e. MDA: χ 2 (1, 81) = 12.36, p < 0.001; see Supporting Information). All other correlations between individual dose rates and oxidative stress markers were nonsignificant (all p < 0.278). Frog dorsal skin luminance was stable during the 48 h that lasted the experiment on colour lability (χ 2 [1,14] = 0.51, p = 0.479) and was unaffected by background colour (χ 2 [1,14] = 0.14, p = 0.705). We also did not find a significant interaction between time and experimental background (χ 2 [1,14] = 2.96, p = 0.085).
4. DISCUSSION
Historical but not current radiation conditions seem to mediate differences in skin coloration in Chornobyl tree frogs. We detected a significant association between dark coloration and proximity to highly contaminated areas in May 1986 (i.e. areas with 137Cs levels >3700 kBq/m2; Cort et al., 2009). However, we did not detect a significant correlation between individual absorbed dose rates and pigmentation, suggesting that darker coloration is not induced by the current exposure to radiation. On average, tree frogs inhabiting the Chornobyl Exclusion Zone were remarkably darker than individuals from a closely located control area with background radiation levels. Additionally, we found no signs of oxidative costs linked to the maintenance of dark coloration in Chornobyl tree frogs, but lower lipid peroxidation in frogs with high dose rates. Finally, the observed differences in coloration do not seem to be due to short‐term changes in skin coloration, or caused by adjustments to background color. These results suggest that exposure to high levels of ionizing radiation may have selected for dark skin coloration in Chornobyl tree frogs.
Dark melanin‐based pigmentation is known to offer protection against different forms of radiation, including ionizing radiation (Cordero & Casadevall, 2020; Pacelli et al., 2017; Robertson et al., 2012). Exposure of fungi to high levels of ionizing radiation under laboratory conditions generates an increase in melanism and in the production of reactive oxygen species scavenging antioxidant enzymes, suggesting a key protective mechanism of melanin (Kothamasi et al., 2019). Likewise, exposure to ionizing radiation enhances the growth of some melanised fungi (Dadachova et al., 2007), which may even transform ionizing radiation into an energy source through the use of melanin pigments (Dadachova & Casadevall, 2008). Actually, melanised fungi can be found in areas with high natural (i.e. background) radiation such as those exposed to high cosmic radiation at the International Space Station (Dadachova & Casadevall, 2008), and they can even colonize highly radio‐contaminated structures such as the interior of the damaged Chornobyl reactor (Wember & Zhdanova, 2001). Laboratory studies have revealed the putative protective role of melanin against ionizing radiation in vertebrates, although the number of these studies is still very limited and they have been conducted under acute radiation conditions, and thus far from the ones experienced by wildlife in radio‐contaminated areas (e.g. Carotenuto et al., 2016; Kunwar et al., 2012).
Our study suggests that dark coloration in tree frogs is linked to historical but not to current radiation levels. This suggestion is based on a positive correlation between luminance and distance to the highest contaminated area at the moment of the accident, and on the lack of correlation between coloration and current dose rates absorbed by the examined frogs. Radiation levels have dropped several orders of magnitude since the accident and many short‐lived radionuclides known to induce significant biological damage have completely disappeared from the area (e.g. 131I; Intelligence Systems GEO, 2011), which can contribute to explain the role of historical exposure and the lack of correlation between luminance and current individual dose rate in our study. Variability in coloration, including dark or grey individuals, is often described in tree frogs of the Hyla genus (Wassef et al., 2019). In our study, dark individuals were also detected (although in very low proportions) in localities outside the Chornobyl area (see Figure 2a). It is, thus, plausible that selective processes acting on existing colour variability favoured individuals with darker coloration, linked to higher survival rates in dark frogs under extremely high radiation conditions shortly after the accident. Indeed, pollution can generate strong selective pressures inducing high rates of evolutionary change (Sanderson et al., 2022). In this line, a review by Geras'kin et al. (2008) showed that the severity of the effects of radiation on the physiology or fitness of different species and on the ecosystem structure after the Chornobyl accident was strongly dependent on the dose received in the early period after the accident. This agrees with the idea that historical exposure to radiation may be behind some of the effects detected in the area across taxa, a topic that deserves further research (Beresford et al., 2020; Hancock et al., 2020). If selection acted on frog coloration, low dispersal and high philopatry to the natal environment characteristic of many amphibians (including the study species; Angelone, 2009), may have favoured the prevalence of dark coloration in the Chornobyl tree frog metapopulation (Car et al., 2022). In addition, amphibians with dark skin coloration often show dark coloration in their internal organs (Franco‐Belussi et al., 2016, 2017), and stressful conditions such as oxygen and food deprivation are known to induce higher cellular pigmentation (Franco‐Belussi et al., 2017). Furthermore, although very few studies have examined the heritability of coloration in amphibians, work on different species has revealed that coloration has a genetic basis (Hoffman & Blouin, 2000; Stuckert et al., 2019), a knowledge that can foster the development of further studies investigating the evolutionary causes and consequences behind the observed divergence in coloration in Chornobyl frogs.
In vertebrates, dark coloration is mostly produced by two melanin‐based pigments, black eumelanin and yellow–red pheomelanin. Melanin production is often costly and can generate oxidative stress (in addition to increases in corticosterone levels and metabolic rate; Chang et al., 2021; Polo‐Cavia & Gomez‐Mestre, 2017), which is also one of the main negative effects of the exposure to ionizing radiation (Galván et al., 2014). However, we found no sign of oxidative stress linked to dark coloration in Chornobyl tree frogs, but lower lipid peroxidation in frogs with high dose rates. The latter may be a consequence of compensatory responses, earlier in life, of the oxidative stress machinery against radiation (similar to observed in amphibian larvae in response to early‐life detrimental conditions, e.g. Burraco et al., 2020). In amphibians, dark skin pigments are generated in melanophores, the skin chromatophores responsible for black, brown and darker green coloration (Duellman & Trueb, 1986). Although pheomelanin has been identified in the skin of some frogs (e.g. Wolnicka‐Glubisz et al., 2012), eumelanin seems to be the almost exclusive dark pigment in amphibians (Frost‐Mason & Mason, 1996; Prota, 1992). Eumelanin protects organisms against DNA damage (Galván et al., 2014), and its production may incur in lower costs than pheomelanin, since this pigment is produced without the need of cysteine and GSH (glutathione), a crucial intracellular antioxidant (García‐Borrón & Olivares Sánchez, 2011; Ito et al., 2011). Studies in wild vertebrates exposed to ionizing radiation have reported a strong demographic decline linked to radiation in bird species with pheomelanin‐based coloration, probably due to high consumption of GSH during pheomelanogenesis (e.g. Galván et al., 2011). By contrast, eumelanin levels in feathers were associated with lower oxidative stress and lower DNA damage in birds breeding in the Chornobyl area (Galván et al., 2014). Since oxidative damage is generally high in radio‐contaminated environments (Bonisoli‐Alquati et al., 2010; Einor et al., 2016), the production of eumelanin pigmentation may allow Chornobyl frogs to get protection without incurring in oxidative costs linked to the synthesis of pheomelanin.
Our study design aimed to minimize the effects of other factors potentially affecting coloration, such as environmental characteristics (e.g. habitat and soil type, water pH), capture time, or field and laboratory temperature, and although habitat changes have been substantial in the area, frog breeding habitats have remained much more stable (Santos, 2018). At the species/individual level, melanin‐based coloration can have pleiotropic effects on other traits (San‐Jose & Roulin, 2018). However, in our study, dorsal coloration did not correlate with body condition, a proxy for overall individual performance. Another putative confounding process may be a link between coloration and mating success (observed in some amphibian species, e.g. Rudh et al., 2011), although that seems not to be the case in tree frogs (Gomez et al., 2009). In our laboratory‐based experiment, skin coloration seemed to be stable over a short time period (48 h). In addition, skin coloration was not significantly affected by background coloration, and (even if not significant) those changes were much smaller than the observed differences in skin coloration between frogs inhabiting within and outside the Chornobyl Exclusion Zone. Despite we discarded the possible influence of several confounding factors on the observed coloration pattern, we acknowledge the limitations of our field approach regarding the lack of field‐based experiments and the small sample size used in the experiment testing for lability in coloration. Also, the absence (to the best of our knowledge) of tree frog samples collected in the Chernobyl area before or immediately after the accident, prevents us from comparing current and past skin coloration. Research addressing genome‐wide signatures of selection may also contribute to improving our understanding on the mechanism behind dark coloration in Chornobyl tree frogs.
5. CONCLUSIONS
Our results suggest that the protective role of melanin previously detected in Chornobyl in smaller living organisms such as fungi may extend to wild vertebrates exposed to ionizing radiation. Historical high radiation and lack of production costs of eumelanin‐based pigmentation may have facilitated the selection and maintenance of dark coloration in Chornobyl tree frogs. Further studies are needed to disentangle the causes and consequences of darker pigmentation in radio‐contaminated environments, which will help to develop a better understanding of the eco‐evolutionary effects of the long‐term exposure to ionizing radiation on wildlife.
FUNDING INFORMATION
This work was supported by the Swedish Radiation Protection Agency‐SSM (SSM2018‐2038), by Carl Tryggers Foundation (CT 16:344) and by the Spanish Society of Terrestrial Ecology (AEET). The Carl Tryggers Foundation, Marie Sklodowska‐Curie project METAGE‐797879, and Juan de la Cierva Incorporación (IJC2020‐044680) supported PB, and the Spanish Ministry of Science and Innovation (Ramón y Cajal program, RYC‐2016‐20,656) supported GO.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
Supporting information
Table S1
Table S2
Figure S1
ACKNOWLEDGMENTS
S. Gashchack, Y. Gulyaichenko, J.‐M. Bonzom and C. Car help us in the field or with radiation analyses, and the administrative personal of the Chornobyl Center for Nuclear Safety, Radioactive Waste and Radioecology (Ukraine) offered administrative assistance. C. Bradshaw and K. Stark helped us on initial stages of the project. F. Miranda helps us with oxidative stress analyses at the Ecophysiology laboratory (LEF) of the Doñana Biological Station. I. Gómez‐Mestre and N. Metcalfe gave us great insights into early versions of the manuscript. W. Jones advised us with English language.
Burraco, P. , & Orizaola, G. (2022). Ionizing radiation and melanism in Chornobyl tree frogs. Evolutionary Applications, 15, 1469–1479. 10.1111/eva.13476
DATA AVAILABILITY STATEMENT
Data associated with this study are made available in the figshare data repository https://doi.org/10.6084/m9.figshare.15124218.v1 Burraco and Orizaola (2022).
REFERENCES
- Alizadeh, E. , Orlando, T. M. , & Sanche, L. (2015). Biomolecular damage induced by ionizing radiation: The direct and indirect effects of low‐energy electrons on DNA. Annual Review of Physical Chemistry, 66, 379–398. [DOI] [PubMed] [Google Scholar]
- Ancans, J. , Tobin, D. J. , Hoogduijn, M. J. , Smit, N. P. , Wakamatsu, K. , & Thody, A. J. (2001). Melanosomal pH controls rate of melanogenesis, eumelanin/phaeomelanin ratio and melanosome maturation in melanocytes and melanoma cells. Experimental Cell Research, 268, 26–35. [DOI] [PubMed] [Google Scholar]
- Angelone, S. (2009). Dispersal success of European tree frogs thanks to habitat connectivity measures: A genetic evaluation. University of Zurich. [Google Scholar]
- AVMA . (2020). AVMA guidelines for the euthanasia of animals (2020th ed.). American Veterinary Medical Association. [Google Scholar]
- Beaugelin‐Seiller, K. , Jasserand, F. , Garnier‐Laplace, J. , & Gariel, J. C. (2006). Modeling radiological dose in non‐human species: Principles, computerization, and application. Health Physics, 90, 485–493. [DOI] [PubMed] [Google Scholar]
- Beresford, N. A. , & Copplestone, D. (2011). Effects of ionizing radiation on wildlife: What knowledge have we gained between the Chernobyl and Fukushima accidents? Integrative Environmental Assessment and Management, 7, 371–373. [DOI] [PubMed] [Google Scholar]
- Beresford, N. A. , Fesenko, S. , Konoplev, A. , Skuterud, L. , Smith, J. T. , & Voigt, G. (2016). Thirty years after the Chernobyl accident: What lessons have we learnt? Journal of Environmental Radioactivity, 157, 77–89. [DOI] [PubMed] [Google Scholar]
- Beresford, N. A. , Horemans, N. , Copplestone, D. , Raines, K. E. , Orizaola, G. , Wood, M. D. , Laanen, P. , Whitehead, H. C. , Burrows, J. E. , Tinsley, M. C. , Smith, J. T. , Bonzom, J.‐M. , Gagnaire, B. , Adam‐Guillermin, C. , Gashchak, S. , Jha, A. N. , de Menezes, A. , Willey, N. , & Spurgeon, D. (2020). Towards solving a scientific controversy ‐ the effects of ionising radiation on the environment. Journal of Environmental Radioactivity, 211, 106033. [DOI] [PubMed] [Google Scholar]
- Bonisoli‐Alquati, A. , Mousseau, T. A. , Møller, A. P. , Carioli, M. , & Saino, N. (2010). Increased oxidative stress in barn swallows from the Chernobyl region. Comparative, Biochemistry, and Physiology Part A, 155, 205–210. [DOI] [PubMed] [Google Scholar]
- Bradford, M. M. (1976). A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein‐dye binding. Analytical Biochemistry, 72, 248–254. [DOI] [PubMed] [Google Scholar]
- Brady, S. P. , Monosson, E. , Matson, C. W. , & Bickham, J. W. (2017). Evolutionary toxicology: Toward a unified understanding of life's response to toxic chemicals. Evolutionary Applications, 10, 745–751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buege, J. A. , & Aust, S. D. (1978). Microsomal lipid peroxidation. Methods in Enzymology, 52, 302–310. [DOI] [PubMed] [Google Scholar]
- Burraco, P. , Bonzom, J. M. , Car, C. , Beaugelin‐Seiller, K. , Gashchack, S. , & Orizaola, G. (2021). Lack of impact of radiation on blood physiology biomarkers of Chernobyl tree frogs. Frontiers in Zoology, 18, 33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burraco, P. , Car, C. , Bonzom, J. M. , & Orizaola, G. (2021). Assessment of exposure to ionizing radiation in Chernobyl tree frogs (Hyla orientalis). Scientific Reports, 11, 20509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burraco, P. , Díaz‐Paniagua, C. , & Gomez‐Mestre, I. (2017). Different effects of accelerated development and enhanced growth on oxidative stress and telomere shortening in amphibian larvae. Scientific Reports, 7, 7494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burraco, P. , & Orizaola, G. (2022). Dataset of: Ionizing radiation and melanism in Chornobyl tree frogs. 10.6084/m9.figshare.15124218.v1 [DOI] [PMC free article] [PubMed]
- Burraco, P. , Valdés, A. E. , & Orizaola, G. (2020). Metabolic costs of altered growth trajectories across life transitions in amphibians. Journal of Animal Ecology, 89, 855–866. [DOI] [PubMed] [Google Scholar]
- Car, C. , Gilles, A. , Armant, O. , Burraco, P. , Beaugelin‐Seiller, K. , Gashchak, S. , Camilleri, V. , Cavalie, I. , Laloi, P. , Adam‐Guillermin, C. , Orizaola, G. , & Bonzom, J.‐M. (2022). Unusual evolution of tree frog populations in the Chernobyl exclusion zone. Evolutionary Applications, 15, 203–219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caro, T. (2005). The adaptive significance of coloration in mammals. Bioscience, 55, 125–136. [Google Scholar]
- Carotenuto, R. , Tussellino, M. , Mettivier, G. , & Russo, P. (2016). Survival fraction and phenotype alterations of Xenopus laevis embryos at 3 Gy, 150 kV X‐ray irradiation. Biochemical and Biophysical Research Communications, 480, 580–585. [DOI] [PubMed] [Google Scholar]
- Carter, E. E. , Tregenza, T. , & Stevens, M. (2020). Ship noise inhibits colour change, camouflage, and anti‐predator behaviour in shore crabs. Current Biology, 30, R211–R212. [DOI] [PubMed] [Google Scholar]
- Cayuela, H. , Valenzuela‐Sánchez, A. , Teulier, L. , Martínez‐Solano, I. , Léna, J. P. , Merilä, J. , Muths, E. , Shine, R. , Quay, L. , Denoël, M. , Clobert, J. , & Schmidt, B. R. (2020). Determinants and consequences of dispersal in vertebrates with complex life cycles: A review of pond‐breeding amphibians. The Quarterly Review of Biology, 95, 1–36. [Google Scholar]
- Chang, L. , Wang, B. , Zhang, M. , Liu, J. , Zhao, T. , Zhu, W. , & Jiang, J. (2021). The effects of corticosterone and background colour on tadpole physiological plasticity. Comparative Biochemistry and Physiology Part D: Genomics and Proteomics, 39, 100872. [DOI] [PubMed] [Google Scholar]
- Cohen, G. , & Somersonm, N. L. (1969). Catalase‐aminotriazole method for measuring secretion of hydrogen peroxide by microorganisms. Journal of Bacteriology, 98, 543–546. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cordero, R. J. B. , & Casadevall, A. (2020). Melanin. Current Biology, 30, R142–R143. [DOI] [PubMed] [Google Scholar]
- Cort, E. V. , Dubois, G. , Fridman, Sh. D. , Germenchuk, M. G. , Izrael, Yu. A. , Janssens, A. , Jones, A. R. , Kelly, G. N. , Kvasnikova, E. V. , Matveenko, I. I. , Nazarov, I. M. , Pokumeiko, Yu. M. , Sitak, V. A. , Stukin, E. D. , Tabachny, L. Ya. , Tsaturov, Yu. S. , & Avdyushin, S. I. (2009). Atlas of caesium deposition on Europe after the chernobyl accident. Directorate‐general for research and innovation (European Commission). ISBN 92–828‐3140‐X.
- Cribb, A. E. , Leeder, J. S. , & Spielberg, S. P. (1989). Use of a microplate reader in an assay of glutathione reductase using 5, 5′‐dithiobis (2‐nitrobenzoic acid). Analytical Biochemistry, 183, 195–196. [DOI] [PubMed] [Google Scholar]
- Cuthill, I. C. , Allen, W. L. , Arbuckle, K. , Caspers, B. , Chaplin, G. , Hauber, M. E. , Hill, G. E. , Jablonski, N. G. , Jiggins, C. D. , Kelber, A. , Mappes, J. , Marshall, J. , Merrill, R. , Osorio, D. , Prum, R. , Roberts, N. W. , Roulin, A. , Rowland, H. M. , Sherratt, T. N. , … Caro, T. (2017). The biology of color. Science, 357, eaan0221. [DOI] [PubMed] [Google Scholar]
- Dadachova, E. , Bryan, R. A. , Howell, R. C. , Schweitzer, A. D. , Aisen, P. , Nosanchuk, J. D. , & Casadevall, A. (2008). The radioprotective properties of fungal melanin are a function of its chemical composition, stable radical presence and spatial arrangement. Pigment Cell & Melanoma Research, 21, 192–199. [DOI] [PubMed] [Google Scholar]
- Dadachova, E. , Bryan, R. A. , Huang, X. , Moadel, T. , Schweitzer, A. D. , Aisen, P. , Nosanchuk, J. D. , & Casadevall, A. (2007). Ionizing radiation changes the electronic properties of melanin and enhances the growth of melanized fungi. PLoS One, 2, e457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dadachova, E. , & Casadevall, A. (2008). Ionizing radiation: How fungi cope, adapt, and exploit with the help of melanin. Current Opinion in Microbiology, 11, 525–531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Del Rio, D. , Stewart, A. J. , & Pellegrini, N. (2005). A review of recent studies on malondialdehyde as toxic molecule and biological marker of oxidative stress. Nutrution, Metabolism and Cardiovascular Diseases, 15, 316–328. [DOI] [PubMed] [Google Scholar]
- Deryabina, T. G. , Kuchmel, S. V. , Nagorskaya, L. L. , Hinton, T. G. , Beasley, J. C. , Lerebours, A. , & Smith, J. T. (2015). Long‐term census data reveal abundant wildlife populations at Chernobyl. Current Biology, 25, 824–826. [DOI] [PubMed] [Google Scholar]
- Duellman, W. E. , & Trueb, L. (1986). Biology of amphibians. The John Hopkins University Press. [Google Scholar]
- Einor, D. , Bonisoli‐Alquati, A. , Costantini, D. , Mousseau, T. A. , & Møller, A. P. (2016). Ionizing radiation, antioxidant response and oxidative damage: A meta‐analysis. Science of the Total Environment, 548–549, 463–471. [DOI] [PubMed] [Google Scholar]
- Fileccia, T. , Guadagni, M. , Hovhera, V. , & Bernoux, M. (2014). Ukraine ‐ soil fertility to strengthen climate resilience: Preliminary assessment of the potential benefits of conservation agriculture. World Bank report number 191850. The World Bank. [Google Scholar]
- Franco‐Belussi, L. , Provete, D. B. , & Oliveira, C. (2017). Environmental correlates of internal coloration in frogs vary throughout space and lineages. Ecology and Evolution, 7, 9222–9233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Franco‐Belussi, L. , Sköld, H. N. , & de Oliveira, C. (2016). Internal pigment cells respond to external UV radiation in frogs. Journal of Experimental Biology, 219, 1378–1383. [DOI] [PubMed] [Google Scholar]
- Frost‐Mason, S. K. , & Mason, K. A. (1996). What insights into vertebrate pigmentation has the axolotl model system provided? The Integrative Journal of Developmental Biology, 40, 685–693. [PubMed] [Google Scholar]
- Galván, I. , Bonisoli‐Alquati, A. , Jenkinson, S. , Ghanem, G. , Wakamatsu, K. , Mousseau, T. A. , & Møller, A. P. (2014). Chronic exposure to low‐dose radiation at Chernobyl favours adaptation to oxidative stress in birds. Functional Ecology, 28, 1387–1403. [Google Scholar]
- Galván, I. , Mousseau, T. A. , & Møller, A. P. (2011). Bird population declines due to radiation exposure at Chernobyl are stronger in species with pheomelanin‐based coloration. Oecologia, 165, 827–835. [DOI] [PubMed] [Google Scholar]
- García‐Borrón, J. C. , & Olivares Sánchez, M. C. (2011). Biosynthesis of melanins. In Borovanský J. & Riley P. A. (Eds.), Melanins and melanosomes: Biosynthesis, biogenesis, physiological, and pathological functions (pp. 87–116). Wiley‐Blackwell. [Google Scholar]
- Geras'kin, S. A. , Fesenko, S. V. , & Alexakhin, R. M. (2008). Effects of non‐human species irradiation after the Chernobyl NPP accident. Environmental International, 34, 880–897. [DOI] [PubMed] [Google Scholar]
- Gombeau, K. , Bonzom, J.‐M. , Cavalié, I. , Camilleri, V. , Orjollet, D. , Dubourg, N. , Beaugelin‐Seiller, K. , Bourdineaud, J.‐P. , Lengagne, T. , Armant, O. , Ravanat, J.‐L. , & Adam‐Guillermin, C. (2020). Dose‐dependent genomic DNA hypermethylation and mitochondrial DNA damage in Japanese tree frogs sampled in the Fukushima Daiichi area. Journal of Environmental Radioactivity, 225, 106429. [DOI] [PubMed] [Google Scholar]
- Gomez, D. , Richardson, C. , Lengagne, T. , Plenet, S. , Joly, P. , Léna, J. P. , & Théry, M. (2009). The role of nocturnal vision in mate choice: Females prefer conspicuous males in the European tree frog (Hyla arborea). Proceedings of the Royal Society B, 276, 2351–2358. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Green, A. J. (2001). Mass/length residuals: Measures of body condition or generators of spurious results? Ecology, 82, 1473–1483. [Google Scholar]
- Halliwell, B. , & Gutteridge, J. M. (2015). Free radicals in biology and medicine. Oxford University Press. [Google Scholar]
- Hancock, S. , Vo, N. T. K. , Goncharova, R. I. , Seymour, C. B. , Byun, S. H. , & Mothersill, C. E. (2020). One‐decade‐spanning transgenerational effects of historic radiation dose in wild populations of bank voles exposed to radioactive contamination following the Chernobyl nuclear disaster. Environmental Research, 180, 108816. [DOI] [PubMed] [Google Scholar]
- Hettyey, A. , Crochet, P. A. , Merilä, J. , Herczeg, G. , & Laurila, A. (2009). Body temperature, size, nuptial colouration and mating success in male moor frogs (Rana arvalis). Amphibia‐Reptilia, 30, 37–43. [Google Scholar]
- Hoffman, E. A. , & Blouin, M. S. (2000). A review of colour and pattern polymorphisms in anurans. Biological Journal of the Linnean Society, 70, 633–665. [Google Scholar]
- Intelligence Systems GEO . (2011). Atlas of radioactive contamination of Ukraine. Ministry of Emergencies and Affairs of Population Protection from the Consequences of Chernobyl Catastrophe. http://radatlas.isgeo.com.ua/ [Google Scholar]
- Ito, S. , Wakamatsu, K. , d'Ischia, M. , Napolitano, A. , & Pezzella, A. (2011). Structure of melanins. In Borovanský J. & Riley P. A. (Eds.), Melanins and melanosomes: Biosynthesis, biogenesis, physiological, and pathological functions (pp. 167–185). Wiley‐Blackwell. [Google Scholar]
- Jernfors, T. , Kesäniemi, J. , Lavrinienko, A. , Mappes, T. , Milinevsky, G. , Møller, A. P. , Mousseau, T. A. , Tukalenko, E. , & Watts, P. C. (2018). Transcriptional upregulation of DNA damage response genes in bank voles (Myodes glareolus) inhabiting the Chernobyl exclusion zone. Frontiers in Environmental Science, 5, 95. [Google Scholar]
- Kesäniemi, J. , Jernfors, T. , Lavrinienko, A. , Kivisaari, K. , Kiljunen, M. , Mappes, T. , & Watts, P. (2019). Exposure to environmental radionuclides is associated with altered metabolic and immunity pathways in a wild rodent. Molecular Ecology, 28, 4620–4635. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kothamasi, D. , Wannijn, J. , Van Hees, M. , Nauts, R. , Van Gompel, A. , Vanhoudt, N. , & Vandenhove, H. (2019). Exposure to ionizing radiation affects the growth of ectomycorrhizal fungi and induces increased melanin production and increased capacities of reactive oxygen species scavenging enzymes. Journal of Environmental Radioactivity, 197, 16–22. [DOI] [PubMed] [Google Scholar]
- Kovalchuk, I. , Abramov, V. , Pogribny, I. , & Kovalchuk, O. (2004). Molecular aspects of plant adaptation to life in the Chernobyl zone. Plant Physiology, 135, 357–363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kryshev, I. I. , Sazykina, T. G. , & Beresford, N. A. (2005). Effects on wildlife. In Chernobyl: Catastrophe and consequences (pp. 267–287). Springer. [Google Scholar]
- Kunwar, A. , Adhikary, B. , Jayakumar, S. , Barik, A. , Chattopadhyay, S. , Raghukumar, S. , & Priyadarsini, K. I. (2012). Melanin, a promising radioprotector: Mechanisms of actions in a mice model. Toxicology and Applied Pharmacology, 264, 202–211. [DOI] [PubMed] [Google Scholar]
- Lehnert, S. (2007). Biomolecular action of ionizing radiation. Taylor & Francis. [Google Scholar]
- Loria, A. , Cristescu, M. E. , & Gonzalez, A. (2019). Mixed evidence for adaptation to environmental pollution. Evolutionary Applications, 12, 1259–1273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mackintosh, J. A. (2001). The antimicrobial properties of melanocytes, melanosomes and melanin and the evolution of black skin. Journal of Theoretical Biology, 211, 101–113. [DOI] [PubMed] [Google Scholar]
- Mappes, T. , Boratyński, Z. , Kivisaari, K. , Lavrinienko, A. , Milinevsky, G. , Mousseau, T. A. , Møller, A. P. , Tukalenko, E. , & Watts, P. C. (2019). Ecological mechanisms can modify radiation effects in a key forest mammal of Chernobyl. Ecosphere, 10, e02667. [Google Scholar]
- Møller, A. P. , & Mousseau, T. A. (2006). Biological consequences of Chernobyl: 20 years on. Trends in Ecology and Evolution, 21, 200–207. [DOI] [PubMed] [Google Scholar]
- Møller, A. P. , & Mousseau, T. A. (2015). Strong effects of ionizing radiation from Chernobyl on mutation rates. Scientific Reports, 5, 8363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Møller, A. P. , & Mousseau, T. A. (2016). Are organisms adapting to ionizing radiation at Chernobyl? Trends in Ecology and Evolution, 31, 281–289. [DOI] [PubMed] [Google Scholar]
- Mothersill, C. , & Seymour, C. (2014). Implications for human and environmental health of low doses of ionising radiation. Journal of Environmental Radioactivity, 133, 5–9. [DOI] [PubMed] [Google Scholar]
- Mustonen, V. , Kesäniemi, J. , Lavrinienko, A. , Tukalenko, E. , Mappes, T. , Watts, P. C. , & Jurvansuu, J. (2018). Fibroblasts from bank voles inhabiting Chernobyl have increased resistance against oxidative and DNA stresses. BMC Cell Biology, 19, 17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nacci, D. , Proestou, D. , Champlin, D. , Martinson, J. , & Waits, E. R. (2016). Genetic basis for rapidly evolved tolerance in the wild: Adaptation to toxic pollutants by an estuarine fish species. Molecular Ecology, 25, 5467–5482. [DOI] [PubMed] [Google Scholar]
- Nilsson Sköld, H. , Aspengren, A. , & Wallin, M. (2013). Rapid color change in fish and amphibians – Function, regulation, and emerging applications. Pigment Cell & Melanoma Research, 26, 29–38. [DOI] [PubMed] [Google Scholar]
- Orizaola, G. (2022). Amphibians in field radioecology: A review and perspective. In Wood M. D., Mothersill C. E., Tsakanova G., Cresswell T., & Woloschak G. E. (Eds.), Biomarkers of radiation in the environment ‐ robust tools for risk assessment NATO science for peace and security series a: Chemistry and biology (pp. 185–203). Springer. [Google Scholar]
- Özdemir, N. , Altunışık, A. , Ergül, T. , Gül, S. , Tosunoğlu, M. , Cadeddu, G. , & Giacoma, C. (2012). Variation in body size and age structure among three Turkish populations of the treefrog Hyla arborea . Amphibia‐Reptilia, 33, 25–35. [Google Scholar]
- Pacelli, C. , Bryan, R. A. , Onofri, S. , Selbmann, L. , Shuryak, I. , & Dadachova, E. (2017). Melanin is effective in protecting fast and slow growing fungi from various types of ionizing radiation. Environmental Microbiology, 19, 1612–1624. [DOI] [PubMed] [Google Scholar]
- Paglia, D. E. , & Valentine, W. N. (1967). Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. Journal of Laboratory and Clinical Medicine, 70, 158–169. [PubMed] [Google Scholar]
- Palumbi, S. R. (2001). Humans as the world's greatest evolutionary force. Science, 293, 1786–1790. [DOI] [PubMed] [Google Scholar]
- Pitelka, L. F. (1988). Evolutionary responses of plants to anthropogenic pollutants. Trends in Ecology and Evolution, 3, 233–236. [DOI] [PubMed] [Google Scholar]
- Polo‐Cavia, N. , & Gomez‐Mestre, I. (2017). Pigmentation plasticity enhances crypsis in larval newts: Associated metabolic cost and background choice behaviour. Scientific Reports, 7, 39739. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prota, G. (1992). Melanins and melanogenesis. Academic Press. [Google Scholar]
- Rands, M. R. W. , Adams, W. A. , Bennun, L. , Butchart, S. H. M. , Clements, A. , Coomes, D. , Entwistle, A. , Hodge, I. , Kapos, V. , Scharlemann, J. P. W. , Sutherland, W. J. , & Vira, B. (2010). Biodiversity conservation: Challenges beyond 2010. Science, 329, 1298–1303. [DOI] [PubMed] [Google Scholar]
- Reid, N. M. , Proestou, D. A. , Clark, B. W. , Warren, W. C. , Colbourne, J. K. , Shaw, J. R. , Karchner, S. I. , Hahn, M. E. , Nacci, D. , Oleksiak, M. F. , Crawford, D. L. , & Whitehead, A. (2016). The genomic landscape of rapid repeated evolutionary adaptation to toxic pollution in wild fish. Science, 354, 1305–1308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robertson, K. L. , Mostaghim, A. , Cuomo, C. A. , Soto, C. M. , Lebedev, N. , Bailey, R. F. , & Wang, Z. (2012). Adaptation of the black yeast Wangiella dermatitidis to ionizing radiation: Molecular and cellular mechanisms. PLoS One, 7, e48674. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rudh, A. , Rogell, B. , Håstad, O. , & Qvarnström, A. (2011). Rapid population divergence linked with co‐variation between coloration and sexual display in strawberry poison frogs. Evolution: International Journal of Organic Evolution, 65, 1271–1282. [DOI] [PubMed] [Google Scholar]
- Sanderson, S. , Beausoleil, M.‐O. , O'Dea, R. E. , Wood, Z. T. , Correa, C. , Frankel, V. , Gorné, L. D. , Haines, G. E. , Kinnison, M. T. , Oke, K. B. , Pelletier, F. , Pérez‐Jvostov, F. , Reyes‐Corral, W. D. , Ritchot, Y. , Sorbara, F. , Gotanda, K. M. , & Hendry, A. P. (2022). The pace of modern life, revisited. Molecular Ecology, 31, 1028–1043. [DOI] [PubMed] [Google Scholar]
- San‐Jose, L. M. , & Roulin, A. (2018). Toward understanding the repeated occurrence of associations between melanin‐based coloration and multiple phenotypes. The American Naturalist, 192, 111–130. [DOI] [PubMed] [Google Scholar]
- Santos, P. (2018). Landscape changes at Chernobyl. PhD, University of Porto. [Google Scholar]
- Sazykina, T. , & Kryshev, I. I. (2006). Radiation effects in wild terrestrial vertebrates — The EPIC collection. Journal of Environmental Radioactivity, 88, 11–48. [DOI] [PubMed] [Google Scholar]
- Schlichting, P. E. , Love, C. N. , Webster, S. C. , & Beasley, J. C. (2019). Efficiency and composition of vertebrate scavengers at the land‐water interface in the Chernobyl exclusion zone. Food Webs, 18, e00107. [Google Scholar]
- Spurgeon, D. , Lahive, E. , Robinson, A. , Short, A. , & Kille, P. (2020). Species sensitivity to toxic substances: Evolution, ecology and applications. Frontiers in Environmental Science, 8, 588380. [Google Scholar]
- Stuckert, A. M. , Moore, E. , Coyle, K. P. , Davison, I. , MacManes, M. D. , Roberts, R. , & Summers, K. (2019). Variation in pigmentation gene expression is associated with distinct aposematic color morphs in the poison frog Dendrobates auratus . BMC Evolutionary Biology, 19, 1–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Troscianko, J. , & Stevens, M. (2015). Image calibration and analysis toolbox–a free software suite for objectively measuring reflectance, colour and pattern. Methods in Ecology and Evolution, 6, 1320–1331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- UNSCEAR (1988). Sources, effects and risks of ionizing radiation. United Nations scientific committee on the effects of atomic radiation 1988 report to the general assembly, with annexes. United Nations. [Google Scholar]
- Wassef, J. , Savary, R. , Ghali, K. , Pasteur, B. , & Dufresnes, C. (2019). Biogeographic patterns of color and size polymorphisms in the European tree frog (Hyla arborea). Bulletin de la Societé Vaudoise Science des Science Naturelles, 98, 93–103. [Google Scholar]
- Webster, S. C. , Byrne, M. E. , Lance, S. L. , Love, C. N. , Hinton, T. G. , Shamovich, D. , & Beasley, J. C. (2016). Where the wild things are: Influence of radiation on the distribution of four mammalian species within the Chernobyl exclusion zone. Frontiers in Ecology and the Environment, 14, 185–190. [Google Scholar]
- Wember, V. V. , & Zhdanova, N. N. (2001). Peculiarities of linear growth of the melanin‐containing fungi Cladosporium sphaerospermum Penz. And Alternaria alternata (Fr.) Keissler. Mikrobiolohichnyĭ Zhurnal, 63, 3–12. [PubMed] [Google Scholar]
- Wolnicka‐Glubisz, A. , Pecio, A. , Podkowa, D. , Kolodziejczyk, L. M. , & Plonka, P. M. (2012). Pheomelanin in the skin of Hymenochirus boettgeri (amphibia: Anura: Pipidae). Experimental Dermatology, 21, 537–540. [DOI] [PubMed] [Google Scholar]
- Yablokov, A. V. , Nesterenko, V. B. , & Nesterenko, A. V. (2009). Consequences of the Chernobyl catastrophe for the environment. Annals of the New York Academy of Sciences, 1181, 221–286. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Table S1
Table S2
Figure S1
Data Availability Statement
Data associated with this study are made available in the figshare data repository https://doi.org/10.6084/m9.figshare.15124218.v1 Burraco and Orizaola (2022).