

Abstract
Prostate cancer is one of the leading causes of death for men worldwide. The development of resistance, toxicity, and side effects of conventional therapies have made prostate cancer treatment become more intensive and aggressive. Many phytochemicals isolated from plants have shown to be tumor cytotoxic. In vitro laboratory studies have revealed that natural compounds can affect cancer cell proliferation by modulating many crucial cellular signaling pathways frequently dysregulated in prostate cancer. A multitude of natural compounds have been found to induce cell cycle arrest, promote apoptosis, inhibit cancer cell growth, and suppress angiogenesis. In addition, combinatorial use of natural compounds with hormone and/or chemotherapeutic drugs seems to be a promising strategy to enhance the therapeutic effect in a less toxic manner, as suggested by pre-clinical studies. In this context, we systematically reviewed the currently available literature of naturally occurring compounds isolated from vegetables, fruits, teas, and herbs, with their relevant mechanisms of action in prostate cancer. As there is increasing data on how phytochemicals interfere with diverse molecular pathways in prostate cancer, this review discusses and emphasizes the implicated molecular pathways of cell proliferation, cell cycle control, apoptosis, and autophagy as important processes that control tumor angiogenesis, invasion, and metastasis. In conclusion, the elucidation of the natural compounds’ chemical structure-based anti-cancer mechanisms will facilitate drug development and the optimization of drug combinations. Phytochemicals, as anti-cancer agents in the treatment of prostate cancer, can have significant health benefits for humans.
Keywords: natural products, chemotherapy, prostate cancer, mechanisms studies
1. Introduction
Prostate cancer (PCa) is the second most commonly diagnosed cancer and the fifth leading cause of cancer death among men worldwide [1]. The American Joint Committee on Cancer (AJCC) developed the TNM (tumor, node, metastasis) system to describe the five stages of prostate cancer: Stage 0, and Stages I to IV. Hormone-naïve advanced prostate cancer is very sensitive to hormones; thus, androgen deprivation therapy (ADT) has been used as the conventional therapy for advanced PCa [2]. Hormone naïve advanced prostate cancer will eventually progress to castration-resistant prostate cancer (CRPC) within 2–3 years after the initial ADT treatment [3]. With an improved understanding that the growth of most CRPC tumors is still dependent on androgen and androgen receptors (AR), several second-generation antiandrogens, such as abiraterone acetate (preventing androgen biosynthesis) and enzalutamide (preventing AR translocation to the nucleus), were developed. However, resistance to these antiandrogens has also appeared clinically [4]. After the failure of antiandrogens, patients will typically be treated with chemotherapy, such as docetaxel/prednisone treatment. Although some patients are initially responsive to chemotherapy, resistance usually occurs in 1–2 years [5]. Traditional herb medicine has used plants to treat various diseases for many years. A number of natural products have been isolated from plants to test their tumor cytotoxic efficacy in the last few decades. The phytochemicals’ natural and raw forms are important for the effectiveness of the agents. Laboratory studies have revealed that natural compounds can affect cellular proliferation by modulating many crucial cellular signaling pathways. Given that the magnitude of decreasing PCa mortality, in company with increasing PCa incidence due to the adoption of screening-based early detection, has been attenuated globally in recent years [1], and both hormone therapy and chemotherapy have potential resistance and severe side effects, there is an intensive need to develop less toxic, practical approaches to improve these conventional treatments. Increasing evidence from laboratory studies has suggested that natural compounds are potential to enhance the therapeutic effect of hormone or chemotherapy in prostate cancer in a non-/less-toxic manner through selectively targeting diverse dysfunctional molecular pathways in cancer cells.
As there is increasing data on how phytochemicals interfere with diverse molecular pathways in PCa, this review discusses and emphasizes the implicated molecular pathways of cell proliferation, cell cycle control, apoptosis, and autophagy as important processes that control tumor angiogenesis, invasion, and metastasis. We reviewed the latest progress on phytochemicals in prostate cancer studies, being particularly interested in information on their mechanism of action. The phytochemicals and their major molecular targets are summarized in Table 1, illustrating the effectiveness of phytochemicals on the major signaling pathways involved in prostate cancer. Some detailed information for each of the phytochemicals is described below.
Table 1.
Bioactive natural products against prostate cancer.
| Natural Compound | Constituent | Plant Source | Chemical Structure | Mechanism | Molecular Pathway (Reference) | In Vitro | In Vivo |
|---|---|---|---|---|---|---|---|
| 1. Polyphenol | |||||||
| 1. 1 Polyphenol Flavonols | |||||||
| 1.1.1 Quercetin | Flavonol | Apple and onion |
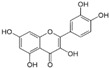
|
Induction of apoptosis. | PI3K/Akt and MAPK/ERK [6]; p21, FA, Hsp90 [7]. | LNCaP, DU 145, PC-3, Prostate cancer stem cells. | |
| 1.1.2 Apigenin | Flavone | Anthemis sp. |
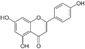
|
Induction of apoptosis and cell cycly arrest. | IGF-IR; PI3k/Akt [8]; HDACs [9]; ROS [10]; VEGF [11]. | PC-3 and 22Rv1. | 22Rv1 and PC-3 xenografts. |
| 1.1.3 Baicalin | Flavone | Genus Scutellaria |
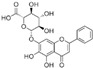
|
Inhibition of proliferation and induction of apoptosis. | Caveolin-1/AKT/mTOR [12]; AR target genes [13]; Ezrin [14]. | DU 145 and PC-3. | PC-3 xenografts. |
| 1.1.4 Cyanidin-3-glucoside | Flavonoid anthocyanin | Berries, red cabbages, teas, and coloured grains. |
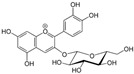
|
Inhibition of EMT. | Snail/E-cadherin [15]. | PC-3. | |
| 1.1.5 Daidzein | Isoflavone | Soybeans |
|
Induction of cell cycle arrest; CpG island demethylation. | CDK [16]; GSTP1 and EPHB2 [17]; ERβ [18]. | DU 145, LNCaP and PC-3. | |
| 1.1.6 Delphinidin | Flavonoid anthocyanin | Viola sp. |
|
Inhibition of cell growth; Induction of apoptosis. | NF-κB [19]; β-catenin [20]; DR5 [21]; HDAC [22]. | PC-3 and LNCaP. | PC-3 xenografts. |
| 1.1.7 Epigallocatechin-3-gallate (EGCG) | Catechin | Green tea |
|
Suppression of the proliferation, invasion and migration, and vasculogenic mimicry; Induction of apoptosis and autophagy. | ERK1/2 [23], VEGF, uPA, angiopoietin 1/2, MMP-2, and MMP-9 [24], PSA [25], Caspase 9 [26]; Twist/VE-Cadherin/AKT [27]; AR [28]; CHOP and p-eIF2α [29]. | LNCaP, PC-3, 22Rv1. | PC-3 xenografts. |
| 1.1.8 Fisetin | Flavonol | Acacia greggii |
|
Inhibition of adhesion, migration, and metastasis; Induction of autophagy. | NF-κB [30]; mTOR and PI3K/Akt [31]; AR [32]; NF-κB [33]; Nudc [34]. | DU 145, LNCaP, PC-3. | CWR22 Rupsilon1 cells xenograft. |
| 1.1.9 Formononetin (FN) | Omethylated isoflavone | Trifolium pretense. |
|
Induction of apoptosis. | ERK1/2, MAPK-Bax [35]; IGF-1/IGF-1R [36]; p38/Akt [37]; RASD1 [38]. | LNCaP, PC-3, and DU 145. | |
| 1.1.10 Genistein | Flavanone | Glycine max |
|
Inhibition of the proliferation; Suppression of cancer stem cells; Activation of epigenetic modification; Induction of apoptosis. |
IGF-1/IGF-1R [39]; miR-1260b [40]; miR-34a and HOTAIR [41]. | LNCaP, PC-3, and DU 145. | TRAMP mice. |
| 1.1.11 Glycyrrhiza | |||||||
| 1.1.11.1 Isoangustone A | Flavonoid | Glycyrrhiza uralensis |

|
Induction of apoptosis; G1 phase arrest. | Caspase [42]; CDK2/4, cyclin A [43]. | DU 145. | |
| 1.1.11.2 Licoricidin | Flavonoid | Glycyrrhiza uralensis |
|
Inhibition of metastasis and invasion. | MMP, TIMP-1, VEGF [44]. | DU 145. | |
| 1.1.12 Licochalcone | Flavonoid | Licorice root |
|
Induction of G2/M phase arrest and apoptosis. | Cyclin B1 and cdc2 [45]. | LNCaP, PC-3. | |
| 1.1.13 Luteolin | Flavone | Terminalia chebula |
|
Suppression of angiogenesis; Induction of cell cycle arrest. | IGF-1 [46]; VEGF-2R, AR [47]. | LNCaP and PC-3. | PC-3 xenografts. |
| 1.2 Polyphenol lignans | |||||||
| 1.2.1 Arctigenin | lignan | Arctium lappa |
|
Inhibition of the proliferation; Induction of apoptosis and autophagy. | AR [48], FFA [49], PI3K/Akt/mTOR [50]. | LAPC-4 and LNCaP. | LAPC-4 xenograft. |
| 1.2.2 Honokiol | Lignin | Magnolia officinalis |
|
Induction of G0-G1 phase arrest and apoptosis; Inhibition of angiogenesis. | Bax, Bak, Bad, Bcl-xL, and Mcl-1 [51,52]. | LNCaP, PC-3, and C4-2. | PC-3 xenografts. |
| 1.2.3 Magnolol | Hydroxylated biphenyl lignan | Magnolia officinalis |
|
Induction of apoptosis and inhibition of the adhesion, invasion, and migration. | EGFR [53]. | PC-3. | |
| 1.2.4 Obovatol | Biphenyl ether lignan | Magnolia obovate |
|
Induction of apoptosis. | NF-κB [54]. | LNCaP and PC-3. | |
| 1.2.5 Silibinin | Flavolignan | Silybum marianum |
|
Induction of G1 phase arrest; Induction of apoptosis; Inhibition of EMT; Restriction of tumor microvessel density. | p21 and p27 [55]; NF-κB [56]; ZEB1 and SLUG [57]. | DU 145. | PC-3 xenografts and TRAMP mice [58]. |
| 1.3 Polyphenol Stilbenoids | |||||||
| 1.3.1 Resveratrol | Stilbenoid | Grape, raspberry, mulberry. |
|
Induction of apoptosis; Cellular antioxidant defense. | FKHRL1 [59]; VEGF, MMPs [60]; CXCR4 [61]; ROS, RNS, HO-1 [62]; AR [63]. | TRAMP mouse [61]. | |
| 1.4 Other Polyphenols | |||||||
| 1.4.1 Curcumin | Polyphenols | Curcuma longa |
|
Induction of apoptosis; Induction of autophagy. | EGF-R [64]; AIF [65]; Wnt/β-catenin [66]; Nrf-2 [67]; MMPs [68]. | DU 145, LNCaP and PC-3. | LNCaP xenograft [69]; DU 145 xenografts [68]. |
| 1.4.2 Ellagitannins | Polyphenols | Pomegranate fruit |
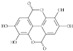
|
Induction of apoptosis and S phase cell cycle arrest; Inhibition of metastasis and angiogenesis. | SIRT1, p21, AIF [70]; Eicosanoid [71]; cyclin B1 and D1 [72]. MMP-2 [73]; p53-MDM2 [74]. | LNCaP, 22RV1 and PC-3. | murine studies [75]. |
| 1.4.3 Gallic acid | Polyphenols | Grape seed |
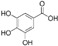
|
Induction of G2/M cell cycle arrest; Induction of apoptosis; Inhibition of invasion and migration. | cdc25A/C-cdc2 [76]; PARP [77]; p38, JNK, PKC, and PI3K/AKT [78]; Cdk, cyclin B1, and E [79]. | DU 145; PC-3. | TRAMP mice [79]; DU 145 and 22Rv1 xenografts [80]. |
| 1.4.4 Gossypol | Polyphenolic aldehyde | Cotton seed |
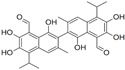
|
Induction of G0/G1 phase arrest; Induction of apoptosis; Induction of autophagy; Inhibition of angiogenesis; Reduction of the microvessel density. | TGF-β1 and Akt [81]; Bcl-xL [82]; Beclin1 [83]. AP-1, NF-κB [84]. | MAT-LyLu; PC-3. | PC-3 xenografts [85]. |
| 2. Terpenoids | |||||||
| 2.1 Artemisinin | Sesquiterpene | Artemisia annua |
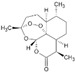
|
Induction of G1 cell cycle arrest; Inhibition of viability. | CDK4 and Sp1 [86], caspases 8/9 [87]. | LNCaP, DU 145 and PC-3. | |
| 2.2 Betulinic Acid (BA) | Triterpene | Betula papyrifera |
|
Induction of apoptosis; Inhibition of angiogenesis. | Bax/Bcl-2 [88], DUBs [89], AR and cyclin D [90]. | DU 145, LNCaP, and PC-3. | TRAMP mice [90]. |
| 2.3 Germacrone | Sesquiterpene | Geranium macrorrhizum. |
|
Induction of apoptosis and autophagy. | Akt/mTOR [91]. | PC-3 and 22RV1. | |
| 2.4 Ginsenosides | Steroid glycosides and triterpene saponins | Genus Panax |
|
Inhibition of proliferation; Inhibition of migration and metastasis; Induction of apoptosis. | p38 [92]; MAP [93]. | PC-3M, LNCaP and PC-3. | PC-3 xenograft [94]. |
| 2.5 Glycyrrhizin | Saponins | Fabaceae |
|
Induction of apoptosis and autophagy. | Apoptosis and autophagy [95]. | PC-3. | |
| 2.6 Lycopene | Carotenoid | Tomatoes |
|
Reduction in cholesterol synthesis; Induction of apoptosis. | CDK7, EGFR, IGF-1R, and BCL2 [96]; PPARγ-LXRα-ABCA1 [97]; CDK7,EGFR, IGF-1R, BCL2 [96]; IGF-I, IGF-IR, and IGFBP-3 [98]. | LNCaP, PC-3 and DU 145. | PC-3 xenograft [98]; TRAMP mice [99]. |
| 2.7 Oridonin | Isoprenoid | Rabdosia rubescens |
|
Induction of apoptosis and G0/G1 cell cycle arrest; Induction of apoptosis, autophagy, and G2/M phase arrest. | p53 and Bax [100]; p21 [101]. | LNCaP and PC-3. | |
| 2.8 Thymoquinone (TQ) | Monoterpene | Nigella sativa |
|
Induction of apoptosis; Inhibition of angiogenesis. | AR and E2F-1 [102]; ROS [103]; AKT, VEGF [104]. | LNCaP, PC-3, C4-B, and DU 145. | |
| 2.9 Ursolic acid (UA) | Pentacyclic triterpenoid | Cornus Officinalis |
|
Induction of apoptosis. | Akt and MMP9 [105]; JNK [106]; CXCR4 [107]. NF-κB and STAT3 [108]; | PC-3, LNCaP, LNCaP-AI, and DU 145. | TRAMP mice [108]. |
| 2.10 β-Elemonic acid | Triterpene | Ganoderma tsugae, lucidum, and Boswellia. |
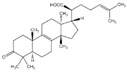
|
Induction of apoptosis. | JAK2/STAT3/MCL-1 and NF-κB [109]. | 22RV1. | 22RV1xenograft [109]. |
| 3. Taxanes | |||||||
| 3.1 Cabazitaxel | Taxane | European yew tree |
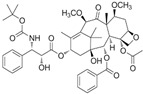
|
Inhibition of proliferation. | P-gp [110]. | C4-2. | |
| 3.2 Docetaxel | Taxane | European yew tree |
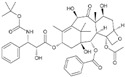
|
Induction of apoptosis. | p38/p53/p21 [111]. | LNCaP, PC3 and DU 145. | |
| 3.3 Paclitaxel | Taxane | Pacific yew tree |
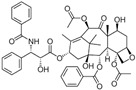
|
Induction of apoptosis. | Bcl-2 [112]; AR [113]. | C4-2. | 22Rv1 xenografts [113]. |
| 4. Alkaloids | |||||||
| 4.1 Anibamine, | Pyridine quaternary alkaloid | Aniba sp. |
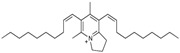
|
Inhibition of proliferation. | CCR5 [114]. | LNCaP. | |
| 4.2 Berberine | Isoquinoline Alkaloid | Genus Berberis |
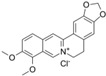
|
Induction of G1 phase arrest; enhancement of the radiosensitivity. | p53-p21 [115], AR [116], MAPK/caspase-3 and ROS [117]. | RM-1. | LNCaP xenografts [116]. |
| 4.3 Capsaicin | Alkaloid | Red pepper |
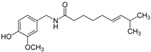
|
Induction of apoptosis. | JNK and ERK [118]. p53, p21, and Bax, AR [119]. | LNCaP, PC-3, and DU 145. | |
| 4.4 Neferine | Bisbenzylisoquinoline alkaloid | Nelumbo nucifera |
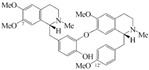
|
Inhibition of proliferation and migration of prostate cancer stem cells. | p38 mapk/jnk [120]. | PC3. | |
| 4.5 Piperine | Alkaloid | Black pepper |
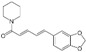
|
Induction of apoptosis, autophagy, and G0/G1 phase arrest. | NF-kB, STAT-3 [121]. | LNCaP, PC-3, and DU 145. | Animal xeno-transplanted model [122]; CRPC xenograft model [123]. |
| 4.6 Sanguinarine | Alkaloid | Sanguinaria Canadensis |
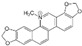
|
Inhibition of ubiquitin-proteasome system. | Stat3 [124], survivin [125], RGS17 [126]. | DU 145, C4-2B, and LNCaP. | DU 145 xenografts [125]. |
| 5. Other | |||||||
| 5.1 Gambogic acid | Xanthone | Garcinia hanburyi |
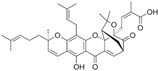
|
Inhibition of angiogenesis. | PI3K/Akt and NF-κB [127]; VEGF-2R [128]. | PC-3. | PC-3 xenograft model [128]. |
| 5.2 Glucoraphanin → Thiocyanates + Isothiocyanates + Indoles | |||||||
| 5.2.1 Sulforaphane (SFN) | Isothiocyanates (ITCs) | Cruciferous vegetables |
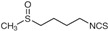
|
Induction of apoptosis; Induction of G2/M cell cycle arrest; Inhibition of FA metabolism. | ERK1/2 [129]; Hsp90 [130]; AR [131]; ROS [132]; FA [133]; HDACs [134]. | LNCaP, DU 145 and PC-3. | TRAMP mice [134]. |
| 5.2.2 Phenethyl-Isothiocyan (PEITC) | Isothiocyanates (ITCs) | Cruciferous vegetables |
|
Induction of G2/M cell cycle arrest; Induction of apoptosis; Inhibition of angiogenesis. | α- and β-tubulin [135]; Bax [136]; Akt [137]; PECAM1-CD31 [138]. | C4-2B, DU 145, PC-3, and LNCaP. | LNCaP xenograft model [138]; TRAMP mice [139]. |
| 5.2.3 Indole-3-Carbinol | Indoles | Cruciferous vegetables |
|
Induction of apoptosis and cell cycle arrest; Modulation of epigenetic alterations of cancer stem cells. | Bax [140]; NF-ĸB, Nrf2 [140]. | LNCaP, and PC-3. | |
2. Polyphenols
2.1. Polyphenol Flavonol
2.1.1. Quercetin
Quercetin is a bioactive flavonol pigment present in onions and apples. Quercetin augmented TRAIL-mediated PCa cell apoptosis and interfered with many oncogenes and tumor suppressor genes, showing cancer-protecting effects [141,142]. Quercetin provoked PCa cell apoptosis by inhibition of FA synthase and downregulation of Hsp90 [7]. Further, quercetin also could inhibit PCa stem cells by PI3K/Akt and MAPK/ERK signaling [6]. Moreover, quercetin exacerbated TRAIL-induced cytotoxicity by a reduction in survivin and Akt phosphorylation and caspase activation [143]. PCa cell xenograft mouse models demonstrated quercetin interacted with the VEGF-R2-regulated autophagic pathway, depicting its anti-angiogenetic effects [144].
2.1.2. Apigenin
Apigenin is a flavone extracted from Anthemis spp. Apigenin suppressed cell proliferation by inhibiting the PI3k/Akt pathway and induced apoptosis by inactivating the IGF-IGF-IR signaling in PC-3 cells [8]. Apigenin also inhibited class I HDACs as a mediator of epigenetic events in PC-3 and 22Rv1 cells [9]. Apigenin-activated p53 pathways by ROS generation induced apoptosis in 22Rv1 cells [10]. Apigenin has shown an anti-angiogenetic potential by reduced vascular endothelial growth factor (VEGF) production in PC-3, LNCaP, and C4-2B cells, attenuating cancer progression and metastasis [11]. Additionally, apigenin inhibited PC-3 and 22Rv1 xenografts in athymic nude mice [145]. Apigenin also suppressed cancer progression in TRAMP mice [146].
2.1.3. Baicalin
Baicalin is a flavone extracted from several species in the genus Scutellaria, such as Scutellaria baicalensis and Scutellaria lateriflora. Baicalein induced apoptosis and inhibited metastasis through inhibition of the caveolin-1/AKT/mTOR pathway in DU 145 and PC-3 cells [12]. Baicalein effectively suppressed the growth of AR-positive PCa cells, by inhibiting the AR N/C dimerization and AR-coactivators interaction in LNCaP cells and CWR22Rv1 cells [13]. Baicalein inhibited PC-3 cell proliferative activity by downregulating Ezrin [14].
2.1.4. Cyanidin-3-Glucoside
Cyanidin-3-glucoside (C3G) is a major flavonoid anthocyanin in plant-based foods, such as leafy vegetables, berries, red cabbages, teas, and colored grains. It is a well-known natural anthocyanin and possesses anti-oxidant and anti-inflammatory properties. Jongsomchai et al. have demonstrated that C3G delays progressive cancer cell behaviors by inhibiting EMT through mediating Snail/E-cadherin expression [15].
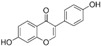
2.1.5. Daidzein
Daidzein is an isoflavone extracted from soybeans. Daidzein elicited cell cycle arrest by modulations of the CDK-related genes and a reduction in EGF and IGF in LNCaP, PC-3, and DU 145 cells [16]. Daidzein also caused epigenetic modifications of tumor suppressor genes, such as CpG island demethylation, thus showing a therapeutic role [17]. Novel daidzein molecules exhibited anti-prostate cancer activity through nuclear receptor ERβ modulation [18].
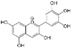
2.1.6. Delphinidin
Delphinidin is an anthocyanidin extracted from Viola spp. Delphinidin inhibited NF-κB signaling and the activation of the subsequent caspase, leading to cell growth inhibition and apoptosis induction in a dose-dependent manner in LNCaP, C4-2, 22Rv1, and PC-3 cells [19]. Delphinidin inhibited cell growth through modulation of β-catenin signaling [20]. Delphinidin sensitized prostate cancer cells to TRAIL-induced apoptosis by inducing DR5 [21]. Delphinidin also induced p53 acetylation-mediated apoptosis by suppressing HDAC activity in LNCaP cells [22]. In addition, delphinidin administration markedly suppressed PC-3 xenografts growth in athymic nude mice [147].
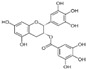
2.1.7. Epigallocatechin-3-Gallate
Epigallocatechin-3-gallate (EGCG) is the most abundant catechin derived from green tea. EGCG reduced the PSA levels, resulting in the suppression of their proliferation in LNCaP cells [25]. EGCG activated extracellular signal-regulated kinase (ERK1/2) and mitogen-activated protein kinase kinase (MEK) signaling, preventing the proliferation of PC-3 cells [23]. EGCG inhibited the invasion and migration of 22Rv1 cells via regulation of protein expression in VEGF, uPA, angiopoietin 1 and 2, MMP-2, and MMP-9 [24]. Combining EGCG and cisplatin promoted the expression of caspase 9, a pro-apoptotic splice isoform in PC-3 cells [26]. EGCG suppressed vasculogenic mimicry through inhibiting the Twist/VE-Cadherin/AKT pathway in PC-3 cells [27]. EGCG suppressed prostate cancer cell growth by modulating acetylation of androgen receptor [28]. EGCG antagonized bortezomib cytotoxicity by an autophagic mechanism in PC-3 cells [29].
Tea, especially green tea, has shown potential in preventing prostate cancer based on pre-clinical and preliminary clinical studies (reviewed by Henning et al. [148]). Our studies have also demonstrated that green tea and quercetin could significantly enhance the therapeutic effect of docetaxel in vitro and in mouse models at no increased risk of side effects [149,150]. However, the role of tea polyphenols in PCa still needs more long-term and intensive studies.
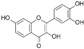
2.1.8. Fisetin
Fisetin is a flavonol derived from Acacia greggii. Fisetin inhibited adhesion, migration, and metastasis by downregulating MMP-2 and MMP-9 and interfering with NF-κB signaling in PC-3 cells [30]. Fisetin sensitized the DU 145, LNCaP, and PC-3 cells to TRAIL-caused death by activating the receptor-mediated mitochondrial apoptotic pathways [33]. Fisetin also induced autophagic cell death by inhibiting mTOR and PI3K/Akt signaling [31]. Fisetin significantly inhibited proliferation, migration, and invasion by disrupting the microtubule dynamics in prostate cancer cells [34]. Moreover, Fisetin attenuated tumor growth and reduced serum PSA levels by competing with the AR ligand and decreasing AR stability in a CWR22 Rupsilon1 cell xenograft model in athymic nude mice [32].
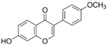
2.1.9. Formononetin
Formononetin (FN) is an O-methylated isoflavone isolated in Trifolium pretense. Formononetin-induced apoptosis was associated with the ERK/MAPK/Bax signaling in LNCaP and PC-3 cells [35]. Another study showed that formononetin provoked apoptosis by inhibiting IGF-1/IGF-1R signaling in PC-3 cells [36], modulating the Bax/Bcl-2 ratio, and altering the p38/Akt signaling [37]. Formononetin also triggers the mitochondrial apoptotic pathway, following the upregulation of RASD1 in DU 145 cells [38].
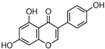
2.1.10. Genistein
Genistein is a flavanone extracted from Glycine max. Genistein suppressed cell growth by inhibiting the IGF-1/IGF-1R signaling in PC-3 cells [39]. The genistein–gold nanoparticle conjugates have shown anti-proliferative activities on three malignant prostate carcinoma cell lines by MTT testing in vitro [151]. Genistein was reported to be essential for inhibiting PCa relapse and metastasis by targeting cancer stem cells (CSC) [152]. Genistein activated epigenetic modification of tumor suppression genes by reversing DNA hypermethylation, inhibiting cancer progression in DU 145 and PC-3 cells [153]. Genistein induced apoptosis by miR-1260b downregulating its target genes, sRRP1 and Smad4 [40]. Furthermore, genistein inhibited cell growth by regulating miR-34a and HOTAIR levels in PC-3 and DU 145 cells [41]. A low percentage of poorly differentiated tumors was observed in TRAMP mice fed genistein [154].
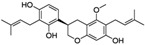
2.1.11. Glycyrrhiza Compounds
The hexane/ethanol extract of Glycyrrhiza uralensis (HEGU) consists of the two active compounds, isoangustone A and licoricidin. Isoangustone A, an active flavonoid, induced apoptosis through increasing cleaved caspase-3, caspase-7, and caspase-9 in DU 145 cells [42]. In addition, isoangustone A diminished DNA synthesis by decreasing the CDK2 activity and caused G1 phase arrest by reducing the CDK2, CDK4, cyclin A, and cyclin D1 expression in DU 145 cells [43]. Licoricidin, another active flavonoid, inhibited the metastatic and invasive capacity of malignant PCa cells by suppressing the secretion of the matrix metalloproteinases (MMP-2, MMP-9), TIMP-1, urokinase-type plasminogen activator (uPA), and VEGF [44].
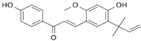
2.1.12. Licochalcone
Licochalcone is an estrogenic flavonoid extracted from licorice root. Licochalcone caused G2/M phase arrest of PC-3 cells by suppressing cyclin B1 and cdc2 [45]. Licochalcone also induced autophagic cell death in LNCaP cells [155].
2.1.13. Luteolin
Luteolin is a flavone extracted from Terminalia chebula. Luteolin suppressed angiogenesis and invasion by reducing AR, PSA, and VEGF-2R in LNCaP and PC-3 cells [47]. Luteolin acted as a ligand for the nuclear type II estradiol binding site, leading to epigenetic alterations in genes involved in the cell cycle in PC-3 cells [156]. Luteolin suppressed PC-3 cell xenografts by inhibiting IGF-1 and the subsequent activation of IGF-1R, AKT, EGFR, and MAPK/ERK signaling [46].
2.2. Polyphenol Lignans
2.2.1. Arctigenin
Arctigenin is a phenylpropanoid dibenzyl butyrolactone found in the seeds of Arctium lappa. Our previous study showed that Arctigenin combined with quercetin synergistically enhanced the antiproliferative effect in prostate cancer cells [48]. Recently, we found that arctigenin inhibited prostate tumor growth in high-fat diet-fed mice through dual actions on adipose tissue and tumor [49]. Sun et al. have demonstrated that arctigenin may induce apoptosis and autophagy through the PI3K/Akt/mTOR pathway in PC-3M Cells [50].
2.2.2. Honokiol
Honokiol is a lignan found in Magnolia officinalis. Honokiol caused G0-G1 phase arrest by triggering apoptotic DNA fragmentation in LNCaP, PC-3, and C4-2 cells [51]. Likewise, honokiol exhibited growth inhibitory effects by the pro-apoptotic and anti-angiogenic mechanism on PCa xenografts [52].
2.2.3. Magnolol
Magnolol is a hydroxylated biphenyl lignan extracted from the root and stem bark of Magnolia officinalis. Magnolol induced apoptotic cell death through epidermal growth factor receptor (EGFR)-mediated pathways and inhibited the adhesion, invasion, and migration in PC-3 cells [53].
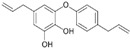
2.2.4. Obovatol
Obovatol, a biphenyl ether lignan found from Magnolia obovate. Obovatol engaged LNCaP and PC-3 cells to apoptotic cell death by inhibiting NF-κB activity, thereby enhancing the therapeutics’ inhibitory effect on PCa cell growth [54].
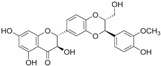
2.2.5. Silibinin
Silibinin or silybin is a flavonolignan derived from the fruits of Silybum marianum. Silibinin induced G1 phase arrest by reducing the expression of p21 and p27 in DU 145 cells [55]. Silibinin induced apoptosis by inhibiting active Stat3 and restraining Wnt/LRp6 signaling, while it sensitized DU 145 cells to TNFα-induced apoptosis by inactivating NF-κB [56]. Silibinin inhibited the epithelial to mesenchymal transition (EMT) of PCa cells by interfering with the NF-κB signaling and subsequent reduction in ZEB1 and SLUG transcription factors [57]. Silibinin also prevented PCa cells-induced osteoclastogenesis in a high bone metastatic prostate model [157]. Silibinin’s inhibitory effects on PC-3 cells xenografts were attributed to increased expression of IGFBP-3, Cip1/p21, and Kip1/p27, and a reduced expression of Bcl-2 and VEGF [158]. Silibinin confined tumor microvessel density by a reduction in VEGF, VEGFR-2, MMPs, and vimentin, thus blocking PCa growth and progression in TRAMP mice [58].
2.3. Polyphenol Stilbenes
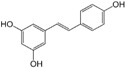
Resveratrol
Resveratrol, a natural stilbenoid polyphenol, is present in several fruits, such as tomatoes, grapes, blueberries, mulberries, and raspberries. Resveratrol increased the pro-apoptotic potential of TNF-related apoptosis-inducing ligand (TRAIL) by activating FKHRL1 [59]. Resveratrol reduced the metastatic potential of PCa cells by decreasing the expression of vascular endothelial growth factor (VEGF), VEGF receptor 2 (VEGFR2), and matrix metalloproteinases (MMPs) [60]. Resveratrol downregulated AR expression and altered the chemokine receptor type 4 (CXCR4) signaling in TRAMP mouse models [61]. Resveratrol enhanced cellular antioxidant defense ability by reducing ROS, reactive nitrogen species (RNS), and inducing the antioxidant enzyme heme-oxygenase-1 (HO-1) [62]. Resveratrol inhibited hypoxia-inducible factor (HIF)-1α and β-catenin-mediated AR signaling in CRPC [63]. These results merit further studies of resveratrol for its potential efficacy against PCa.
2.4. Other Polyphenols
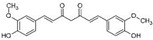
2.4.1. Curcumin
Curcumin is a polyphenolic compound extracted from the rhizome of the plant Curcuma longa. Curcumin was first described to induce apoptosis of PCa cells by interfering with the EGF-R signaling [64]. Curcumin induced apoptosis through apoptosis-inducing factor (AIF) and caspase-independent pathway in PC-3 cells [65]. Curcumin blocked the occupancy at sites of AR function by interacting with cAMP response element binding protein (CBP) and co-activator protein p300 in LNCaP and PC-3 cells, thus decreasing tumor growth and delaying the onset of hormone-resistant disease [159]. Curcumin affected Wnt/β-catenin signaling, resulting in autophagy in early-stage PCa [66]. Curcumin was also an ideal candidate for treating PCa by activating the Nrf-2 signaling [67]. Additionally, derivates of curcumin enhanced expressions of Nrf-2 and phase II detoxifying genes by epigenetic regulation in TRAMP C1 PCa cells [160]. Curcumin reduced the PCa growth and metastasis rate in LNCaP xenograft models [69]. Curcumin suppressed the invasion and metastasis of DU 145 xenografts by a reduction in metalloproteinase expression [68].
2.4.2. Ellagitannin
Ellagitannins, belonging to polyphenols, are enriched in the pomegranate fruit’s seeds and juice. Punicalagin, a kind of ellagitannin, is broken down to ellagic acid (EA), and then further metabolized to urolithin A at intestinal pH by gut microbiota. EA modulated AIF expression, leading to ROS generation and caspase-mediated apoptosis in LNCaP cells [70]. EA decreased the eicosanoid biosynthesis levels in LNCaP cells, depicting the anti-angiogenetic effects [71]. EA could induce apoptosis and cell cycle arrest in the S phase by decreasing cyclin B1 and D1 expression in a caspase-dependent pathway in DU 145 and PC-3 cells [72]. Moreover, EA interfered with protease activity and reduced the MMP-2 secretion, confining the invasive potential of PC-3 and rat PCa cells [73]. The presence of urolithin A led to the inhibition of MDM2-mediated p53 polyubiquitination in 22RV1 and PC-3 cells, indicating that urolithin A inhibited PCa via the p53-MDM2 signaling [74]. In addition, the metabolites ellagic acid, luteolin, and punicic acid have shown the capacity to inhibit PCa cell metastasis and angiogenesis in murine studies [75].
2.4.3. Gallic Acid
Gallic acid (GA) is a polyphenolic constituent extracted from grape seeds. GA caused DNA damage through increasing cdc25A/C-cdc2 phosphorylation, resulting in growth inhibition and G2/M cell cycle arrest in DU 145 cells [76]. GA triggered mitochondrial-mediated mechanisms, including ROS generation, the cleavage of caspase-3, caspase-9, and poly (ADP-ribose) polymerase (PARP), inducing PCa cells apoptosis [77]. GA blocked the p38, JNK, PKC, and PI3K/AKT pathways, inhibiting the invasion and migration of PC-3 cells [78]. The oral feeding with water supplemented with GA decreased the expression of Cdk2, Cdk4, Cdk6, cyclin B1, and E proteins, inhibiting PCa progression to advanced-stage adenocarcinoma in TRAMP mice [79]. GA reduced the microvessel density of DU 145 and 22Rv1 xenografts, suppressing tumor growth in nude mice [80].
2.4.4. Gossypol
Gossypol is a polyphenolic aldehyde extracted from cottonseed. Gossypol modulated the expression of cyclin D1, Cdk4, and phospho-Rb through TGF-β1 and Akt signaling, leading to G0/G1 phase arrest in MAT-LyLu cells [81]. Gossypol inhibited the heterodimerization of Bcl-xL/Bcl-2 with pro-apoptosis molecules, resulting in apoptotic processes [82]. In addition, gossypol induced autophagy by releasing the BH3-only pro-autophagic protein Beclin1, which, in turn, triggered the autophagic cascade in androgen-independent PCa cells [83]. Gossypol inhibited metastatic behaviors and angiogenesis by repressing NF-κB and AP-1 activity in PC-3 cells [84]. Gossypol suppressed the phosphorylation of focal adhesion kinase, extracellular signal-related kinase, and key intracellular proangiogenic kinases by blocking VEGF receptor 2 kinase activation, causing the subsequent suppression of angiogenesis in PC-3 xenografts [85].
3. Terpenoids
3.1. Artemisinin
Artemisinin, a potent anti-malarial compound, is a sesquiterpene lactone isolated from Artemisia annua. Artemisinin transcriptionally downregulated the CDK4 expression by disrupting Sp1 interactions with the CDK4 promoter, triggering G1 cell cycle arrest in LNCaP cells [86]. Dihydroartemisinin (DHA), a derivative of artemisinin, has been shown to reduce cell viability by activating caspases 8/9 in DU 145, LNCaP, and PC-3 cells [87].
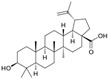
3.2. Betulinic Acid
Betulinic acid (BA) is a pentacyclic triterpene extracted from the bark of Betula papyrifera. BA shifted the Bax/Bcl-2 ratio and cleaved PARP by inhibition of NF-κB, thus inducing PC-3 cell apoptosis [88]. BA decreased oncoproteins expression by inhibiting multiple deubiquitinases (DUBs), which resulted in poly-ubiquitinated protein accumulation, thus increasing apoptosis in DU 145, LNCaP, and PC-3 cells [89]. Furthermore, BA inhibited angiogenesis and tumor growth by decreasing the expression of AR and cyclin D in TRAMP mice [90].
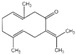
3.3. Germacrone
Germacrone is a sesquiterpene that has been isolated from Geranium macrorrhizum. Yu et al. have shown that Germacrone induces apoptosis and protective autophagy in human prostate cancer cells [91].
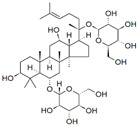
3.4. Ginsenosides
Ginsenosides are a class of natural steroid glycosides and triterpene saponins extracted exclusively from the genus Panax. Ginsenoside Rg3, one of the bioactive components extracted from ginseng root, downregulated aquaporin 1 (AQP1) expression by interfering with the p38 MAPK signaling, inhibiting migration and metastasis of PC-3M cells [92]. Rh2 ginsenoside, another bioactive glycoside, modulated MAP kinase expression, causing cell detachment and showing an anti-proliferative effect in LNCaP and PC-3 cells [93]. Oral administration of Rh2 significantly increased the apoptosis rate, eventually reducing the tumor growth in a PC-3 xenograft model [94].
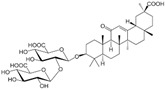
3.5. Glycyrrhizin
Glycyrrhizin, also known as liquorice, is extracted from the root of a flowering plant of the bean family, Fabaceae. Gioti et al. have demonstrated liquorice’s anti-proliferative properties by both apoptosis and autophagy mechanisms in PC-3 cells [95].
3.6. Lycopene
Lycopene is the most abundant antioxidant carotenoid that colors tomatoes. It was reported that lycopene inhibited PCa cell proliferation via modulation of CDK7, epidermal growth factor receptor (EGFR), insulin-like growth factor-1 receptor (IGF-1R), and BCL2 [96]. Lycopene reduced cholesterol synthesis by activating the PPARγ–LXRα–ABCA1 axis in LNCaP and DU 145 cells [97]. Increasing concentrations of lycopene induced the release of mitochondrial cytochrome c, and reduced mitochondrial potential in LNCaP cells [161]. A high concentration of lycopene induced apoptosis through alterations in IGF-I, IGF-IR, and IGFBP-3 expression in PC-3 cells and xenograft models [98]. Finally, the therapeutic activity of lycopene was also observed in TRAMP mice fed lycopene [99].
3.7. Oridonin
Oridonin is an isoprenoid extracted from Rabdosia rubescens. Oridonin elicited apoptosis and G0/G1 cell cycle arrest by upregulating p53 and Bax and downregulating Bcl-2 in LNCaP cells [100]. Oridonin triggered apoptosis, autophagy, and G2/M phase arrest by the up-regulation of p21 in LNCaP and PC-3 cells [101].
3.8. Thymoquinone
Thymoquinone (TQ) is a monoterpene extracted from Nigella sativa. TQ inhibited cell growth by decreasing AR, E2F-1, and E2F-1 in LNCaP, PC-3, C4-B, and DU 145 cells [102]. TQ induced apoptosis through increasing ROS generation and decreasing GSH levels in C4-B and PC-3 cells [103]. Furthermore, TQ was demonstrated to prevent tumor angiogenesis by suppressing AKT in PC-3 cells [104].
3.9. Ursolic Acid
Ursolic acid (UA) is a pentacyclic triterpenoid compound isolated from Cornus officinalis. UA evoked apoptosis and confined invasion by inhibiting Akt and MMP9 in PC-3 cells [105]. UA also induced apoptosis through JNK activation, resulting in phosphorylation and degradation of Bcl-2 and activation of caspase 9 in LNCaP, LNCaP-AI, and DU 145 cells [106]. UA restricted metastasis by repressing CXCR4 expression in PCa cells [107]. In addition, UA induced apoptosis by downregulating the NF-κB and STAT3 target genes involved in proliferation, survival, and angiogenesis in TRAMP mice [108].
3.10. β-Elemonic Acid
β-Elemonic acid (β-EA), a known triterpene isolated from Ganoderma tsugae, Ganoderma lucidum, and Boswellia, exhibits anti-inflammatory effects. Bao et al. have shown that β-EA inhibited the activation of JAK2/STAT3/MCL-1 and the NF-κB pathway in PCa cells. Furthermore, tumor growth in a murine xenograft model was retarded by the administration of β-EA [109].
4. Taxanes
Taxane is a class of diterpenes, initially isolated from the genus Taxus (yews). Therapies aimed at decreasing androgen signaling have been the principal treatment for advanced prostate cancer for most of a century [162]. The breakthrough in cytotoxic chemotherapy for prostate cancer came with the taxanes. Taxanes function through microtubule interaction and polymerization, stabilizing microtubule stabilization and mitotic arrest.
4.1. Cabazitaxel
Cabazitaxel is a semi-synthetic derivative of a natural taxoid extracted from yew needles. Cabazitaxel is a third-generation taxane drug developed after resistance was seen with the other taxanes. Cabazitaxel effectively inhibited the proliferation of CRPC cells with acquired docetaxel resistance [163,164]. The distinct activity of cabazitaxel to overcome resistance to prior taxanes or hormonal therapies in prostate cancer was observed in pre-clinical and clinical data [110]. In conclusion, the success of taxanes in PCa treatment suggests a need for a further understanding of the taxane mechanisms of action and a better development of rational taxane-based combination therapies.
4.2. Docetaxel (DTX)
Docetaxel is a taxane derivative isolated from the European yew tree’s renewable and more readily available leaves. Docetaxel is a semi-synthetic analog of paclitaxel as a cytotoxic chemotherapeutic agent. Gan et al. have shown that silencing the p38/p53/p21 signaling could be crucial to sensitizing LNCaP cells to docetaxel treatment [111].
4.3. Paclitaxel (PTX)
Paclitaxel is a chemotherapy medication in the taxane family of drugs, first isolated from the Pacific yew, Taxus brevifolia. Paclitaxel phosphorylated the B-cell lym phoma-2 (Bcl-2) protein at serine residues, which inhibited the mitochondrial release of cytochrome c and subsequently blocked the caspase cascade, thereby inducing apoptosis [112]. Paclitaxel induced nuclear accumulation of FOXO1, thus inhibiting AR nuclear translocation from reducing the expression of AR target genes in PCa cells [113].
5. Alkaloids
5.1. Anibamine
Anibamine, a novel pyridine quaternary alkaloid recently isolated from Aniba spp., is a natural antagonist of C–C chemokine receptor type 5 (CCR5). Haney et al. found anibamine and their analogs, with micromolar range affinity to CCR5, exerted anti-proliferative activity against several prostate cancer cell lines [114].
5.2. Berberine
Berberine is an isoquinoline alkaloid isolated from the genus Berberis. Low concentrations of berberine triggered G1 arrest by activating the p53-p21 cascade in RM-1 cells [115]. Berberine inhibited tumor growth by reducing AR expression in nude mice bearing LNCaP xenografts [116]. Berberine inhibited the expression of HIF-1 alpha and VEGF by interfering with the MAPK/caspase-3 and ROS pathways, thereby enhancing the radiosensitivity of human PCa cells [117].
5.3. Capsaicin
Capsaicin is a vanilloid extracted from red pepper. Capsaicin induced apoptosis through ROS generation and activation of caspase 3 in PC-3 cells [165]. Further studies revealed that capsaicin induced apoptosis by ceramide accumulation and activation of JNK and ERK in PC-3 cells [118]. Likewise, the various concentrations of capsaicin stimulated apoptosis by increasing p53, p21, and Bax, decreasing prostate-specific antigen (PSA) and AR, and inhibiting proteasome activity in LNCaP and DU 145 cells [119].
5.4. Neferine
Neferine is the major bisbenzylisoquinoline alkaloid isolated from the seed embryo of a traditional medicinal plant, Nelumbo nucifera (Lotus). Erdogan et al. have shown that neferine inhibits the proliferation and migration of human prostate cancer stem cells through p38 MAPK/JNK activation [120].
5.5. Piperine
Piperine is a major bioactive alkaloid present in black pepper. Piperine induced apoptosis, promoted autophagy, triggered cell cycle arrest at G0/G1, and inhibited proliferation in PCa cells and animal xeno-transplanted models [122]. Piperine also inhibited the transcription factor NF-kB expression and downregulate phosphorylated STAT-3 in LNCaP, PC-3, and DU 145 cells [121]. Additionally, piperine remarkably enhanced the anti-cancer effects of docetaxel in a xenograft model of CRPC [123].
5.6. Sanguinarine
Sanguinarine is a benzophenanthridine alkaloid isolated from Sanguinaria canadensis. Sanguinarine restricted PCa cell growth, migration, and invasion by inactivating Stat3 in DU 145, C4-2B, and LNCaP cells [124]. The administration of sanguinarine confined tumor weight and volume by suppressing survivin via the ubiquitin–proteasome system in DU 145 cell xenografts [125]. Using a high-throughput screen, Bodle et al. discovered two natural compounds, sanguinarine and celastrol, to be cytotoxic against prostate cancer cell lines by inhibiting interaction with regulator of G protein signaling 17 (RGS17) [126].
6. Other
6.1. Gambogic Acid
Gambogic acid (GA) is a xanthone found in Garcinia hanburyi. GA suppressed TNF-α-caused invasion of PC-3 cells by inactivating PI3K/Akt and NF-κB pathways [127]. Injection of GA suppressed angiogenesis and tumor growth by inhibiting vascular endothelial growth factor receptor 2 (VEGF-2R) and its downstream protein kinases, such as c-Src and AKT, in a xenograft model [128].
6.2. Glucoraphanin
Glucoraphanin (GRA) is the most abundant glucosinolate (GSLs) and is extracted from cruciferous vegetables such as brussels sprouts, broccoli, and cauliflower, contributing to these vegetables’ organoleptic properties. GSLs are enzymatically hydrolyzed to breakdown derivates, including thiocyanates, isothiocyanates, and indoles, exhibiting biological effects (Figure 1), either by bacteria with microbial thioglucosidase activity in the gut or by the plant enzyme myrosinase [166].
Figure 1.

Breakdown of glucosinolates.
6.2.1. Sulforaphane
Sulforaphane (SFN) was first isolated from Brassica oleracea and is a natural isothiocyanate (ITC). SFN could reduce the progression and incidence of PCa by metabolic regulation in PCa cells [167]. SFN interfered with heat shock protein 90 (Hsp90), a critical androgen receptor (AR) chaperone, in prostate cancer [130]. SFN directly attenuated the androgen receptor (AR) pathways by inhibiting HDAC6 activity in the LNCaP cells [131]. SFN metabolites, sulforaphane-cysteine (SFN-Cys), and sulforaphane-N-acetyl-cysteine (SFN-NAC) disrupted the microtubules and induced apoptosis in DU 145 and PC-3 cells by phosphorylation of extracellular signal-regulated protein kinases 1 and 2 (ERK1/2) [129]. SFN could cause G2/M cell cycle arrest and apoptotic cell death via signaling disruption within tumor microenvironments [168]. SFN has been reported to strongly inhibit the nuclear translocation of p65 and nuclear factor-kappa B (NF-ĸb) activity, subsequently regulating gene expression of Bcl-XL, cyclin D1, and VEGF in PCa cells [169]. SFN also restricts prostate cancer cell migration because of the modulations of the Notch pathway [170]. SFN induced apoptosis by reactive oxygen species (ROS) generation, triggering intrinsic and extrinsic caspase cascades in LNCaP, PC-3, and DU 145 cells [132]. SFN significantly reduced the level of free fatty acid (FA) and relevant metabolism proteins in PCa cells and the prostatic adenocarcinoma of TRAMP mice, suggesting that the SFN inhibits PCa progression by reducing FA metabolism [133]. In addition, SFN inhibited HDAC3, an epigenetic regulator, in PC-3 cells and decreased HDAC3 expression in TRAMP mice [134]. These results explain the mechanism of how the cruciferous vegetable broccoli may reduce the risk of PCa progression.
6.2.2. Phenethyl-Isothiocyanate
Phenethyl-isothiocyanate (PEITC) is another extensively studied ITC found in cruciferous vegetables. PEITC induced G2/M cell cycle arrest by inhibiting the expression of α- and β-tubulin and generation of ROS in C4-2B, DU 145, PC-3, and LNCaP cells [135]. PEITC activated Bax and ROS production to trigger apoptotic mechanisms in LNCaP and PC-3 cells [136]. It was noteworthy that PEITC interfered with the Notch pathway and inactivated Akt with subsequent suppression of VEGF, restraining the migration of PC-3 and LNCaP cells [137]. PEITC caused apoptosis and cell cycle arrest in the G2/M phase in a dose-dependent manner in DU 145 cells [171]. Finally, PEITC slowed tumor growth by suppressing angiogenesis in an LNCaP xenograft model [138]. Studies in TRAMP mice also revealed that the overexpression of E-cadherin-induced prostate carcinogenesis was inhibited by PEITC [139].
6.2.3. Indole-3-Carbinol
Indole-3-carbinol is a main breakdown product of GRA by the action of myrosinase enzymes. Indole-3-carbinol caused cell cycle arrest by downregulating cyclin-dependent kinase2 (CDK2), CDK4, and CDK6, and upregulating p15, p21, and p27 [140]. Indole-3-carbinol induces apoptosis by activating cas-3 and cas-9, the subsequent release of mitochondrial cytochrome C, and upregulation of the pro-apoptotic protein Bax [140]. Indole-3-carbinol potentiated the effects of tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) by inactivation of NF-ĸB, estrogen receptor (ER), androgen receptor (AR), and nuclear factor-E2-related factor 2 (Nrf2) signaling [140]. Additionally, indole-3-carbinol could modulate epigenetic alterations of cancer stem cells, including aberrant microRNA expression, CpG methylation, and histone modification [168].
7. Conclusions
This review represents the latest findings on phytochemicals from vegetables, fruit, tea, and herbs that have shown multi-modal anti-cancer effects on PCa cells, including cell cycle arrest, triggering apoptotic cell death, and suppression of invasion/metastasis, and their underlying molecular mechanisms (Figure 2). The multi-targeting capacity of these phytochemicals seems to be a favorable feature against cancer cells that typically have lots of dysregulated signals that may crosstalk with each other. Moreover, many phytochemicals have been selective in targeting cancer cells without toxicity to normal cells, making them more promising as cancer preventive/therapeutic agents. In addition, increasing evidence from laboratory studies has shown the potential of phytochemicals to enhance the therapeutic effect of drugs in a non-/less-toxic manner in treating different cancers, including prostate cancer.
Figure 2.

Mechanism of action of bioactive natural products in prostate cancer cells.
However, several limitations with these phytochemicals need to be addressed on the road to a successful clinical application. First, the low bioavailability is one of the major limitations of most phytochemicals. In vitro effective concentrations are barely achievable in vivo through oral consumption at safe doses, limiting their clinical success. Second, the long-term toxicity of these phytochemicals will still need to be tested, particularly due to the relatively high concentrations consumed during disease treatment. In addition, complete profiling of their molecular targets may help address the concern of possible side effects of their multi-targeting activities.
Relative to the extensive pre-clinical studies on phytochemicals, information from clinical trials is limited. All phases of clinical trial studies are needed to advance this field. In addition, efforts should continue to identify novel phytochemicals of improved bioavailability and anti-cancer potency. Despite all the challenges, natural products remain a promise to be an alternative or complementary approach to cancer treatment. In conclusion, elucidation of the chemical structure-based precise mechanisms of action will facilitate the identification of novel natural compounds with significant anti-cancer properties, for both drug development and optimization of drug combinations, which are essential to make the phytochemical an anti-cancer agent in a non-/less-toxic manner in the treatment of prostate cancer, thereby providing significant health benefits for humans.
Abbreviations
ATP-Binding Cassette transporter 1 (ABCA1); Androgen Deprivation Therapy (ADT); Apoptosis Inducing Factor (AIF); American Joint Committee on Cancer (AJCC); Aquaporin 1 (AQP1); Androgen Receptor (AR); Betulinic acid (BA); B-cell lymphoma 2 (Bcl-2); B cell lymphoma extra-large (Bcl-xL); Benign Prostatic Hyperplasia (BPH); cAMP response element-binding protein (CBP); C–C chemokine receptor type 5 (CCR5); Cell Division Cycle 6 (CDC6); Cyclin-Dependent Kinases (CDKs); Cyclooxygenase-2 (COX-2); cAMP response element-binding protein (CREB); Castrate-Resistant Prostate Cancer (CRPC); Cancer Stem Cells (CSC); Chemokine receptor type 4 (CXCR4); Diallyl Disulfide (DADS); Diallyl Aulfide (DAS); Dihydroartemisinin (DHA); Deubiquitinases (DUBs); Ellagic acid (EA); Epicatechin (EC); Epicatechin-3-gallate (ECG); Epigallocatechin-3-gallate (EGCG); Epidermal Growth Factor Receptor (EGFR); Epithelial-to-Mesenchymal Transition (EMT); Estrogen Receptor (ER); Extracellular signal-regulated protein kinases 1 and 2 (ERK1/2); Fatty Acid (FA); Fas-associated death domain protein-like interleukin-1-betaconverting enzyme inhibitory protein (FLIP); Gallic acid (GA); Gambogic acid (GA); Gut Microbiota (GM); Glucoraphanin (GRA); Glucosinolates (GSLs); Histone Deacetylase (HDAC); Hexane/ethanol Extract of Glycyrrhiza Uralensis (HEGU); Hypoxia-Inducible Factor (HIF); Heme-Oxygenase-1 (HO-1); Heat Shock Protein 90 (Hsp90); Human Umbilical Vein Endothelial Cells (HUVEC); Inhibitor-of Apoptosis Protein (IAP); Inhibitor of DNA binding 1 (Id1); Insulin-like Growth Factor (IGF); Insulin-like Growth Factor-1 Receptor (IGF-1R); Insulin like Growth Insulin-like Growth Factors Binding Protein-3 (IGFBP-3); Isothiocyanates (ITCs); Liver X Receptor alpha (LXRα); Mitogen-activated protein kinase (MAPK); MAPK kinase 1/2 (MEK1/2); Matrix Metalloproteinases (MMPs); Methanethiosulfinate (MMTSI); Methanethiolsulfonate (MMTSO); Metastasis associated protein 1 (MTA1); Nuclear Factor-kappa B (NF-ĸB); Nuclear factor-E2-related factor 2 (Nrf2); Poly (ADP-ribose) polymerase (PARP); Prostate cancer (PCa); Prostate-Derived Epithelial Factor (PDEF); Platelet/Endothelial Cell Adhesion Molecule (PECAM1-CD31); Phenethyl-isothiocyanate (PEITC); Protein Kinase C (PKC); Peroxisome Proliferator-Activated Receptor gamma (PPARγ); Prostate-Specific Antigen (PSA); Retinoblastoma (Rb); Regulator of G Protein Signaling 17 (RGS17); Reactive Nitrogen Species (RNS); Reactive Oxygen Species (ROS); S-Alk(en)yl-L-Cysteine Sulfoxides (SACSOs); Sulforaphane (SFN); Sulforaphane-Cysteine (SFN-Cys) and Sulforaphane-N-Acetyl-Cysteine (SFN-NAC); Silent Information Regulator 1 (SIRT1); S-Methyl-L-Cysteine Sulfoxide (SMCSO); TNM (Tumor, Node, Metastasis); TNF-related apoptosis-inducing ligand (TRAIL); Ursolic acid (UA); urokinase-type Plasminogen Activator (uPA); Vascular Endothelial Growth Factor (VEGF); Vascular Endothelial Growth Factor Receptor 2 (VEGF-2R); X chromosome-linked IAP (XIAP).
Author Contributions
Investigation: Q.H.; visualization: Q.H.; conceptualization: P.W.; funding acquisition: Q.H., Y.W., J.V.V. and P.W.; writing—original draft preparation: Q.H.; writing—review and editing: P.W. All authors have read and agreed to the published version of the manuscript.
Conflicts of Interest
The authors declare no potential conflict of interest.
Funding Statement
This work was supported by NIH/NIMHD Accelerating Excellence in Translational Science Pilot Grants G0814C01 (Q. Hao); NIH/NCI 1U54CA14393, NIH/NIMHD U54MD007598, Department of Defense Breast Cancer Research Program Grant BC043180, and NIH/NCATS CTSI UL1TR000124 (J.V. Vadgama). NIH/NCI 1R03CA208221, NIH/NIGMS 1SC1GM121202, and NIH/NIMHD 5S21MD000103 (P. Wang).
Footnotes
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Wang L., Lu B., He M., Wang Y., Wang Z., Du L. Prostate Cancer Incidence and Mortality: Global Status and Temporal Trends in 89 Countries From 2000 to 2019. Front. Public Health. 2022;10:811044. doi: 10.3389/fpubh.2022.811044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Crawford E.D., Heidenreich A., Lawrentschuk N., Tombal B., Pompeo A.C.L., Mendoza-Valdes A., Miller K., Debruyne F.M.J., Klotz L. Androgen-targeted therapy in men with prostate cancer: Evolving practice and future considerations. Prostate Cancer Prostatic Dis. 2019;22:24–38. doi: 10.1038/s41391-018-0079-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Dong L., Zieren R.C., Xue W., de Reijke T.M., Pienta K.J. Metastatic prostate cancer remains incurable, why? Asian J. Urol. 2019;6:26–41. doi: 10.1016/j.ajur.2018.11.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Pal S.K., Patel J., He M., Foulk B., Kraft K., Smirnov D.A., Twardowski P., Kortylewski M., Bhargava V., Jones J.O. Identification of mechanisms of resistance to treatment with abiraterone acetate or enzalutamide in patients with castration-resistant prostate cancer (CRPC) Cancer. 2018;124:1216–1224. doi: 10.1002/cncr.31161. [DOI] [PubMed] [Google Scholar]
- 5.Gjyrezi A., Xie F., Voznesensky O., Khanna P., Calagua C., Bai Y., Kung J., Wu J., Corey E., Montgomery B., et al. Taxane resistance in prostate cancer is mediated by decreased drug-target engagement. J. Clin. Investig. 2020;130:3287–3298. doi: 10.1172/JCI132184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Erdogan S., Turkekul K., Dibirdik I., Doganlar O., Doganlar Z.B., Bilir A., Oktem G. Midkine downregulation increases the efficacy of quercetin on prostate cancer stem cell survival and migration through PI3K/AKT and MAPK/ERK pathway. Biomed. Pharm. 2018;107:793–805. doi: 10.1016/j.biopha.2018.08.061. [DOI] [PubMed] [Google Scholar]
- 7.Aalinkeel R., Bindukumar B., Reynolds J.L., Sykes D.E., Mahajan S.D., Chadha K.C., Schwartz S.A. The dietary bioflavonoid, quercetin, selectively induces apoptosis of prostate cancer cells by down-regulating the expression of heat shock protein 90. Prostate. 2008;68:1773–1789. doi: 10.1002/pros.20845. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Shukla S., Gupta S. Apigenin suppresses insulin-like growth factor I receptor signaling in human prostate cancer: An in vitro and in vivo study. Mol. Carcinog. 2009;48:243–252. doi: 10.1002/mc.20475. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Pandey M., Kaur P., Shukla S., Abbas A., Fu P., Gupta S. Plant flavone apigenin inhibits HDAC and remodels chromatin to induce growth arrest and apoptosis in human prostate cancer cells: In vitro and in vivo study. Mol. Carcinog. 2012;51:952–962. doi: 10.1002/mc.20866. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Shukla S., Gupta S. Apigenin-induced prostate cancer cell death is initiated by reactive oxygen species and p53 activation. Free Radic. Biol. Med. 2008;44:1833–1845. doi: 10.1016/j.freeradbiomed.2008.02.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Mirzoeva S., Franzen C.A., Pelling J.C. Apigenin inhibits TGF-beta-induced VEGF expression in human prostate carcinoma cells via a Smad2/3- and Src-dependent mechanism. Mol. Carcinog. 2014;53:598–609. doi: 10.1002/mc.22005. [DOI] [PubMed] [Google Scholar]
- 12.Guo Z., Hu X., Xing Z., Xing R., Lv R., Cheng X., Su J., Zhou Z., Xu Z., Nilsson S., et al. Baicalein inhibits prostate cancer cell growth and metastasis via the caveolin-1/AKT/mTOR pathway. Mol. Cell Biochem. 2015;406:111–119. doi: 10.1007/s11010-015-2429-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Xu D., Chen Q., Liu Y., Wen X. Baicalein suppresses the androgen receptor (AR)-mediated prostate cancer progression via inhibiting the AR N-C dimerization and AR-coactivators interaction. Oncotarget. 2017;8:105561–105573. doi: 10.18632/oncotarget.22319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Ma S.C., Chen R., Yang T.N., Zhang F., Zhao D. Baicalein inhibits the proliferative activity of human prostate cancer cell line PC3 by downregulating Ezrin. J. Biol. Regul. Homeost. Agents. 2020;34:885–892. doi: 10.23812/20-145-A-44. [DOI] [PubMed] [Google Scholar]
- 15.Jongsomchai K., Leardkamolkarn V., Mahatheeranont S. A rice bran phytochemical, cyanidin 3-glucoside, inhibits the progression of PC3 prostate cancer cell. Anat. Cell Biol. 2020;53:481–492. doi: 10.5115/acb.20.085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Rabiau N., Kossai M., Braud M., Chalabi N., Satih S., Bignon Y.J., Bernard-Gallon D.J. Genistein and daidzein act on a panel of genes implicated in cell cycle and angiogenesis by polymerase chain reaction arrays in human prostate cancer cell lines. Cancer Epidemiol. 2010;34:200–206. doi: 10.1016/j.canep.2009.12.018. [DOI] [PubMed] [Google Scholar]
- 17.Vardi A., Bosviel R., Rabiau N., Adjakly M., Satih S., Dechelotte P., Boiteux J.P., Fontana L., Bignon Y.J., Guy L., et al. Soy phytoestrogens modify DNA methylation of GSTP1, RASSF1A, EPH2 and BRCA1 promoter in prostate cancer cells. In Vivo. 2010;24:393–400. [PubMed] [Google Scholar]
- 18.Ranjithkumar R., Saravanan K., Balaji B., Hima S., Sreeja S., Timane S.R., Ram Pravin Kumar M., Kabilan S., Ramanathan M. Novel daidzein molecules exhibited anti-prostate cancer activity through nuclear receptor ERbeta modulation, in vitro and in vivo studies. J. Chemother. 2021;33:582–594. doi: 10.1080/1120009X.2021.1924935. [DOI] [PubMed] [Google Scholar]
- 19.Bin Hafeez B., Asim M., Siddiqui I.A., Adhami V.M., Murtaza I., Mukhtar H. Delphinidin, a dietary anthocyanidin in pigmented fruits and vegetables: A new weapon to blunt prostate cancer growth. Cell Cycle. 2008;7:3320–3326. doi: 10.4161/cc.7.21.6969. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Lee W., Yun J.M. Suppression of beta-catenin Signaling Pathway in Human Prostate Cancer PC3 Cells by Delphinidin. J. Cancer Prev. 2016;21:110–114. doi: 10.15430/JCP.2016.21.2.110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Ko H., Jeong M.H., Jeon H., Sung G.J., So Y., Kim I., Son J., Lee S.W., Yoon H.G., Choi K.C. Delphinidin sensitizes prostate cancer cells to TRAIL-induced apoptosis, by inducing DR5 and causing caspase-mediated HDAC3 cleavage. Oncotarget. 2015;6:9970–9984. doi: 10.18632/oncotarget.3667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Jeong M.H., Ko H., Jeon H., Sung G.J., Park S.Y., Jun W.J., Lee Y.H., Lee J., Lee S.W., Yoon H.G., et al. Delphinidin induces apoptosis via cleaved HDAC3-mediated p53 acetylation and oligomerization in prostate cancer cells. Oncotarget. 2016;7:56767–56780. doi: 10.18632/oncotarget.10790. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Albrecht D.S., Clubbs E.A., Ferruzzi M., Bomser J.A. Epigallocatechin-3-gallate (EGCG) inhibits PC-3 prostate cancer cell proliferation via MEK-independent ERK1/2 activation. Chem. Biol. Interact. 2008;171:89–95. doi: 10.1016/j.cbi.2007.09.001. [DOI] [PubMed] [Google Scholar]
- 24.Siddiqui I.A., Asim M., Hafeez B.B., Adhami V.M., Tarapore R.S., Mukhtar H. Green tea polyphenol EGCG blunts androgen receptor function in prostate cancer. FASEB J. 2011;25:1198–1207. doi: 10.1096/fj.10-167924. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Chuu C.P., Chen R.Y., Kokontis J.M., Hiipakka R.A., Liao S. Suppression of androgen receptor signaling and prostate specific antigen expression by (-)-epigallocatechin-3-gallate in different progression stages of LNCaP prostate cancer cells. Cancer Lett. 2009;275:86–92. doi: 10.1016/j.canlet.2008.10.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Hagen R.M., Chedea V.S., Mintoff C.P., Bowler E., Morse H.R., Ladomery M.R. Epigallocatechin-3-gallate promotes apoptosis and expression of the caspase 9a splice variant in PC3 prostate cancer cells. Int. J. Oncol. 2013;43:194–200. doi: 10.3892/ijo.2013.1920. [DOI] [PubMed] [Google Scholar]
- 27.Yeo C., Han D.S., Lee H.J., Lee E.O. Epigallocatechin-3-Gallate Suppresses Vasculogenic Mimicry through Inhibiting the Twist/VE-Cadherin/AKT Pathway in Human Prostate Cancer PC-3 Cells. Int. J. Mol. Sci. 2020;21:439. doi: 10.3390/ijms21020439. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Lee Y.H., Kwak J., Choi H.K., Choi K.C., Kim S., Lee J., Jun W., Park H.J., Yoon H.G. EGCG suppresses prostate cancer cell growth modulating acetylation of androgen receptor by anti-histone acetyltransferase activity. Int. J. Mol. Med. 2012;30:69–74. doi: 10.3892/ijmm.2012.966. [DOI] [PubMed] [Google Scholar]
- 29.Modernelli A., Naponelli V., Giovanna Troglio M., Bonacini M., Ramazzina I., Bettuzzi S., Rizzi F. EGCG antagonizes Bortezomib cytotoxicity in prostate cancer cells by an autophagic mechanism. Sci. Rep. 2015;5:15270. doi: 10.1038/srep15270. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Chien C.S., Shen K.H., Huang J.S., Ko S.C., Shih Y.W. Antimetastatic potential of fisetin involves inactivation of the PI3K/Akt and JNK signaling pathways with downregulation of MMP-2/9 expressions in prostate cancer PC-3 cells. Mol. Cell Biochem. 2010;333:169–180. doi: 10.1007/s11010-009-0217-z. [DOI] [PubMed] [Google Scholar]
- 31.Adhami V.M., Syed D.N., Khan N., Mukhtar H. Dietary flavonoid fisetin: A novel dual inhibitor of PI3K/Akt and mTOR for prostate cancer management. Biochem. Pharm. 2012;84:1277–1281. doi: 10.1016/j.bcp.2012.07.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Khan N., Asim M., Afaq F., Abu Zaid M., Mukhtar H. A novel dietary flavonoid fisetin inhibits androgen receptor signaling and tumor growth in athymic nude mice. Cancer Res. 2008;68:8555–8563. doi: 10.1158/0008-5472.CAN-08-0240. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Szliszka E., Helewski K.J., Mizgala E., Krol W. The dietary flavonol fisetin enhances the apoptosis-inducing potential of TRAIL in prostate cancer cells. Int. J. Oncol. 2011;39:771–779. doi: 10.3892/ijo.2011.1116. [DOI] [PubMed] [Google Scholar]
- 34.Mukhtar E., Adhami V.M., Sechi M., Mukhtar H. Dietary flavonoid fisetin binds to beta-tubulin and disrupts microtubule dynamics in prostate cancer cells. Cancer Lett. 2015;367:173–183. doi: 10.1016/j.canlet.2015.07.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Ye Y., Hou R., Chen J., Mo L., Zhang J., Huang Y., Mo Z. Formononetin-induced apoptosis of human prostate cancer cells through ERK1/2 mitogen-activated protein kinase inactivation. Horm. Metab. Res. 2012;44:263–267. doi: 10.1055/s-0032-1301922. [DOI] [PubMed] [Google Scholar]
- 36.Huang W.J., Bi L.Y., Li Z.Z., Zhang X., Ye Y. Formononetin induces the mitochondrial apoptosis pathway in prostate cancer cells via downregulation of the IGF-1/IGF-1R signaling pathway. Pharm. Biol. 2013;52:466–470. doi: 10.3109/13880209.2013.842600. [DOI] [PubMed] [Google Scholar]
- 37.Zhang X., Bi L., Ye Y., Chen J. Formononetin induces apoptosis in PC-3 prostate cancer cells through enhancing the Bax/Bcl-2 ratios and regulating the p38/Akt pathway. Nutr. Cancer. 2014;66:656–661. doi: 10.1080/01635581.2014.894098. [DOI] [PubMed] [Google Scholar]
- 38.Liu X.J., Li Y.Q., Chen Q.Y., Xiao S.J., Zeng S.E. Up-regulating of RASD1 and apoptosis of DU-145 human prostate cancer cells induced by formononetin in vitro. Asian Pac. J. Cancer Prev. 2014;15:2835–2839. doi: 10.7314/APJCP.2014.15.6.2835. [DOI] [PubMed] [Google Scholar]
- 39.Lee J., Ju J., Park S., Hong S.J., Yoon S. Inhibition of IGF-1 signaling by genistein: Modulation of E-cadherin expression and downregulation of beta-catenin signaling in hormone refractory PC-3 prostate cancer cells. Nutr. Cancer. 2012;64:153–162. doi: 10.1080/01635581.2012.630161. [DOI] [PubMed] [Google Scholar]
- 40.Hirata H., Hinoda Y., Shahryari V., Deng G., Tanaka Y., Tabatabai Z.L., Dahiya R. Genistein downregulates onco-miR-1260b and upregulates sFRP1 and Smad4 via demethylation and histone modification in prostate cancer cells. Br. J. Cancer. 2014;110:1645–1654. doi: 10.1038/bjc.2014.48. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Chiyomaru T., Yamamura S., Fukuhara S., Yoshino H., Kinoshita T., Majid S., Saini S., Chang I., Tanaka Y., Enokida H., et al. Genistein inhibits prostate cancer cell growth by targeting miR-34a and oncogenic HOTAIR. PLoS ONE. 2013;8:e70372. doi: 10.1371/journal.pone.0070372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Seon M.R., Lim S.S., Choi H.J., Park S.Y., Cho H.J., Kim J.K., Kim J., Kwon D.Y., Park J.H. Isoangustone A present in hexane/ethanol extract of Glycyrrhiza uralensis induces apoptosis in DU145 human prostate cancer cells via the activation of DR4 and intrinsic apoptosis pathway. Mol. Nutr. Food Res. 2010;54:1329–1339. doi: 10.1002/mnfr.200900260. [DOI] [PubMed] [Google Scholar]
- 43.Seon M.R., Park S.Y., Kwon S.J., Lim S.S., Choi H.J., Park H., Lim D.Y., Kim J.S., Lee C.H., Kim J., et al. Hexane/ethanol extract of Glycyrrhiza uralensis and its active compound isoangustone A induce G1 cycle arrest in DU145 human prostate and 4T1 murine mammary cancer cells. J. Nutr. Biochem. 2012;23:85–92. doi: 10.1016/j.jnutbio.2010.11.010. [DOI] [PubMed] [Google Scholar]
- 44.Park S.Y., Lim S.S., Kim J.K., Kang I.J., Kim J.S., Lee C., Kim J., Park J.H. Hexane-ethanol extract of Glycyrrhiza uralensis containing licoricidin inhibits the metastatic capacity of DU145 human prostate cancer cells. Br. J. Nutr. 2010;104:1272–1282. doi: 10.1017/S0007114510002114. [DOI] [PubMed] [Google Scholar]
- 45.Fu Y., Hsieh T.C., Guo J., Kunicki J., Lee M.Y., Darzynkiewicz Z., Wu J.M. Licochalcone-A, a novel flavonoid isolated from licorice root (Glycyrrhiza glabra), causes G2 and late-G1 arrests in androgen-independent PC-3 prostate cancer cells. Biochem. Biophys. Res. Commun. 2004;322:263–270. doi: 10.1016/j.bbrc.2004.07.094. [DOI] [PubMed] [Google Scholar]
- 46.Fang J., Zhou Q., Shi X.L., Jiang B.H. Luteolin inhibits insulin-like growth factor 1 receptor signaling in prostate cancer cells. Carcinogenesis. 2007;28:713–723. doi: 10.1093/carcin/bgl189. [DOI] [PubMed] [Google Scholar]
- 47.Pratheeshkumar P., Son Y.O., Budhraja A., Wang X., Ding S., Wang L., Hitron A., Lee J.C., Kim D., Divya S.P., et al. Luteolin inhibits human prostate tumor growth by suppressing vascular endothelial growth factor receptor 2-mediated angiogenesis. PLoS ONE. 2012;7:e52279. doi: 10.1371/journal.pone.0052279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Wang P., Phan T., Gordon D., Chung S., Henning S.M., Vadgama J.V. Arctigenin in combination with quercetin synergistically enhances the antiproliferative effect in prostate cancer cells. Mol. Nutr. Food Res. 2015;59:250–261. doi: 10.1002/mnfr.201400558. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Hao Q., Diaz T., Verduzco A.D.R., Magyar C.E., Zhong J., Elshimali Y., Rettig M.B., Henning S.M., Vadgama J.V., Wang P. Arctigenin inhibits prostate tumor growth in high-fat diet fed mice through dual actions on adipose tissue and tumor. Sci. Rep. 2020;10:1403. doi: 10.1038/s41598-020-58354-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Sun B.L., Cai E.B., Zhao Y., Wang Y., Yang L.M., Wang J.Y. Arctigenin Triggers Apoptosis and Autophagy via PI3K/Akt/mTOR Inhibition in PC-3M Cells. Chem. Pharm. Bull. 2021;69:472–480. doi: 10.1248/cpb.c21-00021. [DOI] [PubMed] [Google Scholar]
- 51.Hahm E.R., Singh S.V. Honokiol causes G0-G1 phase cell cycle arrest in human prostate cancer cells in association with suppression of retinoblastoma protein level/phosphorylation and inhibition of E2F1 transcriptional activity. Mol. Cancer Ther. 2007;6:2686–2695. doi: 10.1158/1535-7163.MCT-07-0217. [DOI] [PubMed] [Google Scholar]
- 52.Hahm E.R., Arlotti J.A., Marynowski S.W., Singh S.V. Honokiol, a constituent of oriental medicinal herb magnolia officinalis, inhibits growth of PC-3 xenografts in vivo in association with apoptosis induction. Clin. Cancer Res. 2008;14:1248–1257. doi: 10.1158/1078-0432.CCR-07-1926. [DOI] [PubMed] [Google Scholar]
- 53.Lee D.H., Szczepanski M.J., Lee Y.J. Magnolol induces apoptosis via inhibiting the EGFR/PI3K/Akt signaling pathway in human prostate cancer cells. J. Cell Biochem. 2009;106:1113–1122. doi: 10.1002/jcb.22098. [DOI] [PubMed] [Google Scholar]
- 54.Lee S.Y., Yuk D.Y., Song H.S., Yoon D.Y., Jung J.K., Moon D.C., Lee B.S., Hong J.T. Growth inhibitory effects of obovatol through induction of apoptotic cell death in prostate and colon cancer by blocking of NF-kappaB. Eur. J. Pharm. 2008;582:17–25. doi: 10.1016/j.ejphar.2007.12.027. [DOI] [PubMed] [Google Scholar]
- 55.Roy S., Kaur M., Agarwal C., Tecklenburg M., Sclafani R.A., Agarwal R. p21 and p27 induction by silibinin is essential for its cell cycle arrest effect in prostate carcinoma cells. Mol. Cancer Ther. 2007;6:2696–2707. doi: 10.1158/1535-7163.MCT-07-0104. [DOI] [PubMed] [Google Scholar]
- 56.Agarwal C., Tyagi A., Kaur M., Agarwal R. Silibinin inhibits constitutive activation of Stat3, and causes caspase activation and apoptotic death of human prostate carcinoma DU145 cells. Carcinogenesis. 2007;28:1463–1470. doi: 10.1093/carcin/bgm042. [DOI] [PubMed] [Google Scholar]
- 57.Wu K., Zeng J., Li L., Fan J., Zhang D., Xue Y., Zhu G., Yang L., Wang X., He D. Silibinin reverses epithelial-to-mesenchymal transition in metastatic prostate cancer cells by targeting transcription factors. Oncol. Rep. 2010;23:1545–1552. [PubMed] [Google Scholar]
- 58.Singh R.P., Raina K., Sharma G., Agarwal R. Silibinin inhibits established prostate tumor growth, progression, invasion, and metastasis and suppresses tumor angiogenesis and epithelial-mesenchymal transition in transgenic adenocarcinoma of the mouse prostate model mice. Clin. Cancer Res. 2008;14:7773–7780. doi: 10.1158/1078-0432.CCR-08-1309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Kumar A., Dhar S., Rimando A.M., Lage J.M., Lewin J.R., Zhang X., Levenson A.S. Epigenetic potential of resveratrol and analogs in preclinical models of prostate cancer. Ann. N. Y. Acad. Sci. 2015;1348:1–9. doi: 10.1111/nyas.12817. [DOI] [PubMed] [Google Scholar]
- 60.Ganapathy S., Chen Q., Singh K.P., Shankar S., Srivastava R.K. Resveratrol enhances antitumor activity of TRAIL in prostate cancer xenografts through activation of FOXO transcription factor. PLoS ONE. 2010;5:e15627. doi: 10.1371/journal.pone.0015627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Jang Y.G., Go R.E., Hwang K.A., Choi K.C. Resveratrol inhibits DHT-induced progression of prostate cancer cell line through interfering with the AR and CXCR4 pathway. J. Steroid Biochem. Mol. Biol. 2019;192:105406. doi: 10.1016/j.jsbmb.2019.105406. [DOI] [PubMed] [Google Scholar]
- 62.Xue D., Zhou C., Shi Y., Lu H., Xu R., He X. Nuclear transcription factor Nrf2 suppresses prostate cancer cells growth and migration through upregulating ferroportin. Oncotarget. 2016;7:78804–78812. doi: 10.18632/oncotarget.12860. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Mitani T., Harada N., Tanimori S., Nakano Y., Inui H., Yamaji R. Resveratrol inhibits hypoxia-inducible factor-1alpha-mediated androgen receptor signaling and represses tumor progression in castration-resistant prostate cancer. J. Nutr. Sci. Vitaminol. 2014;60:276–282. doi: 10.3177/jnsv.60.276. [DOI] [PubMed] [Google Scholar]
- 64.Dorai T., Gehani N., Katz A. Therapeutic potential of curcumin in human prostate cancer. II. Curcumin inhibits tyrosine kinase activity of epidermal growth factor receptor and depletes the protein. Mol. Urol. 2000;4:1–6. [PubMed] [Google Scholar]
- 65.Hilchie A.L., Furlong S.J., Sutton K., Richardson A., Robichaud M.R., Giacomantonio C.A., Ridgway N.D., Hoskin D.W. Curcumin-induced apoptosis in PC3 prostate carcinoma cells is caspase-independent and involves cellular ceramide accumulation and damage to mitochondria. Nutr. Cancer. 2010;62:379–389. doi: 10.1080/01635580903441238. [DOI] [PubMed] [Google Scholar]
- 66.Teiten M.H., Gaascht F., Cronauer M., Henry E., Dicato M., Diederich M. Anti-proliferative potential of curcumin in androgen-dependent prostate cancer cells occurs through modulation of the Wingless signaling pathway. Int. J. Oncol. 2011;38:603–611. doi: 10.3892/ijo.2011.905. [DOI] [PubMed] [Google Scholar]
- 67.Khurana N., Sikka S.C. Targeting Crosstalk between Nrf-2, NF-kappaB and Androgen Receptor Signaling in Prostate Cancer. Cancers. 2018;10:352. doi: 10.3390/cancers10100352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Hong J.H., Ahn K.S., Bae E., Jeon S.S., Choi H.Y. The effects of curcumin on the invasiveness of prostate cancer in vitro and in vivo. Prostate Cancer Prostatic Dis. 2006;9:147–152. doi: 10.1038/sj.pcan.4500856. [DOI] [PubMed] [Google Scholar]
- 69.Killian P.H., Kronski E., Michalik K.M., Barbieri O., Astigiano S., Sommerhoff C.P., Pfeffer U., Nerlich A.G., Bachmeier B.E. Curcumin inhibits prostate cancer metastasis in vivo by targeting the inflammatory cytokines CXCL1 and -2. Carcinogenesis. 2012;33:2507–2519. doi: 10.1093/carcin/bgs312. [DOI] [PubMed] [Google Scholar]
- 70.Vanella L., Di Giacomo C., Acquaviva R., Barbagallo I., Cardile V., Kim D.H., Abraham N.G., Sorrenti V. Apoptotic markers in a prostate cancer cell line: Effect of ellagic acid. Oncol. Rep. 2013;30:2804–2810. doi: 10.3892/or.2013.2757. [DOI] [PubMed] [Google Scholar]
- 71.Vanella L., Di Giacomo C., Acquaviva R., Barbagallo I., Li Volti G., Cardile V., Abraham N.G., Sorrenti V. Effects of ellagic Acid on angiogenic factors in prostate cancer cells. Cancers. 2013;5:726–738. doi: 10.3390/cancers5020726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Naiki-Ito A., Chewonarin T., Tang M., Pitchakarn P., Kuno T., Ogawa K., Asamoto M., Shirai T., Takahashi S. Ellagic acid, a component of pomegranate fruit juice, suppresses androgen-dependent prostate carcinogenesis via induction of apoptosis. Prostate. 2015;75:151–160. doi: 10.1002/pros.22900. [DOI] [PubMed] [Google Scholar]
- 73.Pitchakarn P., Chewonarin T., Ogawa K., Suzuki S., Asamoto M., Takahashi S., Shirai T., Limtrakul P. Ellagic acid inhibits migration and invasion by prostate cancer cell lines. Asian Pac. J. Cancer Prev. 2013;14:2859–2863. doi: 10.7314/APJCP.2013.14.5.2859. [DOI] [PubMed] [Google Scholar]
- 74.Mohammed Saleem Y.I., Albassam H., Selim M. Urolithin A induces prostate cancer cell death in p53-dependent and in p53-independent manner. Eur. J. Nutr. 2020;59:1607–1618. doi: 10.1007/s00394-019-02016-2. [DOI] [PubMed] [Google Scholar]
- 75.Wang D., Ozen C., Abu-Reidah I.M., Chigurupati S., Patra J.K., Horbanczuk J.O., Jozwik A., Tzvetkov N.T., Uhrin P., Atanasov A.G. Vasculoprotective Effects of Pomegranate (Punica granatum L.) Front. Pharm. 2018;9:544. doi: 10.3389/fphar.2018.00544. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Agarwal C., Tyagi A., Agarwal R. Gallic acid causes inactivating phosphorylation of cdc25A/cdc25C-cdc2 via ATM-Chk2 activation, leading to cell cycle arrest, and induces apoptosis in human prostate carcinoma DU145 cells. Mol. Cancer Ther. 2006;5:3294–3302. doi: 10.1158/1535-7163.MCT-06-0483. [DOI] [PubMed] [Google Scholar]
- 77.Chen H.M., Wu Y.C., Chia Y.C., Chang F.R., Hsu H.K., Hsieh Y.C., Chen C.C., Yuan S.S. Gallic acid, a major component of Toona sinensis leaf extracts, contains a ROS-mediated anti-cancer activity in human prostate cancer cells. Cancer Lett. 2009;286:161–171. doi: 10.1016/j.canlet.2009.05.040. [DOI] [PubMed] [Google Scholar]
- 78.Liu K.C., Huang A.C., Wu P.P., Lin H.Y., Chueh F.S., Yang J.S., Lu C.C., Chiang J.H., Meng M., Chung J.G. Gallic acid suppresses the migration and invasion of PC-3 human prostate cancer cells via inhibition of matrix metalloproteinase-2 and -9 signaling pathways. Oncol. Rep. 2011;26:177–184. doi: 10.3892/or.2011.1264. [DOI] [PubMed] [Google Scholar]
- 79.Raina K., Rajamanickam S., Deep G., Singh M., Agarwal R., Agarwal C. Chemopreventive effects of oral gallic acid feeding on tumor growth and progression in TRAMP mice. Mol. Cancer Ther. 2008;7:1258–1267. doi: 10.1158/1535-7163.MCT-07-2220. [DOI] [PubMed] [Google Scholar]
- 80.Kaur M., Velmurugan B., Rajamanickam S., Agarwal R., Agarwal C. Gallic acid, an active constituent of grape seed extract, exhibits anti-proliferative, pro-apoptotic and anti-tumorigenic effects against prostate carcinoma xenograft growth in nude mice. Pharm. Res. 2009;26:2133–2140. doi: 10.1007/s11095-009-9926-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Jiang J., Ye W., Lin Y.C. Gossypol inhibits the growth of MAT-LyLu prostate cancer cells by modulation of TGFbeta/Akt signaling. Int. J. Mol. Med. 2009;24:69–75. [PubMed] [Google Scholar]
- 82.Zhang M., Liu H., Tian Z., Griffith B.N., Ji M., Li Q.Q. Gossypol induces apoptosis in human PC-3 prostate cancer cells by modulating caspase-dependent and caspase-independent cell death pathways. Life Sci. 2007;80:767–774. doi: 10.1016/j.lfs.2006.11.004. [DOI] [PubMed] [Google Scholar]
- 83.Lian J., Karnak D., Xu L. The Bcl-2-Beclin 1 interaction in (-)-gossypol-induced autophagy versus apoptosis in prostate cancer cells. Autophagy. 2010;6:1201–1203. doi: 10.4161/auto.6.8.13549. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Jiang J., Slivova V., Jedinak A., Sliva D. Gossypol inhibits growth, invasiveness, and angiogenesis in human prostate cancer cells by modulating NF-kappaB/AP-1 dependent- and independent-signaling. Clin. Exp. Metastasis. 2012;29:165–178. doi: 10.1007/s10585-011-9439-z. [DOI] [PubMed] [Google Scholar]
- 85.Pang X., Wu Y., Wu Y., Lu B., Chen J., Wang J., Yi Z., Qu W., Liu M. (-)-Gossypol suppresses the growth of human prostate cancer xenografts via modulating VEGF signaling-mediated angiogenesis. Mol. Cancer Ther. 2011;10:795–805. doi: 10.1158/1535-7163.MCT-10-0936. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Willoughby J.A., Sr., Sundar S.N., Cheung M., Tin A.S., Modiano J., Firestone G.L. Artemisinin blocks prostate cancer growth and cell cycle progression by disrupting Sp1 interactions with the cyclin-dependent kinase-4 (CDK4) promoter and inhibiting CDK4 gene expression. J. Biol. Chem. 2009;284:2203–2213. doi: 10.1074/jbc.M804491200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.He Q., Shi J., Shen X.L., An J., Sun H., Wang L., Hu Y.J., Sun Q., Fu L.C., Sheikh M.S., et al. Dihydroartemisinin upregulates death receptor 5 expression and cooperates with TRAIL to induce apoptosis in human prostate cancer cells. Cancer Biol. Ther. 2010;9:819–824. doi: 10.4161/cbt.9.10.11552. [DOI] [PubMed] [Google Scholar]
- 88.Rabi T., Shukla S., Gupta S. Betulinic acid suppresses constitutive and TNFalpha-induced NF-kappaB activation and induces apoptosis in human prostate carcinoma PC-3 cells. Mol. Carcinog. 2008;47:964–973. doi: 10.1002/mc.20447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Reiner T., Parrondo R., de Las Pozas A., Palenzuela D., Perez-Stable C. Betulinic acid selectively increases protein degradation and enhances prostate cancer-specific apoptosis: Possible role for inhibition of deubiquitinase activity. PLoS ONE. 2013;8:e56234. doi: 10.1371/journal.pone.0056234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.de Las Pozas A., Reiner T., De Cesare V., Trost M., Perez-Stable C. Inhibiting Multiple Deubiquitinases to Reduce Androgen Receptor Expression in Prostate Cancer Cells. Sci. Rep. 2018;8:13146. doi: 10.1038/s41598-018-31567-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Yu Z., Xu J., Shao M., Zou J. Germacrone Induces Apoptosis as Well as Protective Autophagy in Human Prostate Cancer Cells. Cancer Manag. Res. 2020;12:4009–4016. doi: 10.2147/CMAR.S250522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Pan X.Y., Guo H., Han J., Hao F., An Y., Xu Y., Xiaokaiti Y., Pan Y., Li X.J. Ginsenoside Rg3 attenuates cell migration via inhibition of aquaporin 1 expression in PC-3M prostate cancer cells. Eur. J. Pharm. 2012;683:27–34. doi: 10.1016/j.ejphar.2012.02.040. [DOI] [PubMed] [Google Scholar]
- 93.Kim H.S., Lee E.H., Ko S.R., Choi K.J., Park J.H., Im D.S. Effects of ginsenosides Rg3 and Rh2 on the proliferation of prostate cancer cells. Arch. Pharm. Res. 2004;27:429–435. doi: 10.1007/BF02980085. [DOI] [PubMed] [Google Scholar]
- 94.Musende A.G., Eberding A., Wood C., Adomat H., Fazli L., Hurtado-Coll A., Jia W., Bally M.B., Guns E.T. Pre-clinical evaluation of Rh2 in PC-3 human xenograft model for prostate cancer in vivo: Formulation, pharmacokinetics, biodistribution and efficacy. Cancer Chemother. Pharm. 2009;64:1085–1095. doi: 10.1007/s00280-009-0965-1. [DOI] [PubMed] [Google Scholar]
- 95.Gioti K., Papachristodoulou A., Benaki D., Beloukas A., Vontzalidou A., Aligiannis N., Skaltsounis A.L., Mikros E., Tenta R. Glycyrrhiza glabra-Enhanced Extract and Adriamycin Antiproliferative Effect on PC-3 Prostate Cancer Cells. Nutr. Cancer. 2020;72:320–332. doi: 10.1080/01635581.2019.1632357. [DOI] [PubMed] [Google Scholar]
- 96.Rafi M.M., Kanakasabai S., Reyes M.D., Bright J.J. Lycopene modulates growth and survival associated genes in prostate cancer. J. Nutr. Biochem. 2013;24:1724–1734. doi: 10.1016/j.jnutbio.2013.03.001. [DOI] [PubMed] [Google Scholar]
- 97.Yang C.M., Lu I.H., Chen H.Y., Hu M.L. Lycopene inhibits the proliferation of androgen-dependent human prostate tumor cells through activation of PPARgamma-LXRalpha-ABCA1 pathway. J. Nutr. Biochem. 2012;23:8–17. doi: 10.1016/j.jnutbio.2010.10.006. [DOI] [PubMed] [Google Scholar]
- 98.Yang C.M., Yen Y.T., Huang C.S., Hu M.L. Growth inhibitory efficacy of lycopene and beta-carotene against androgen-independent prostate tumor cells xenografted in nude mice. Mol. Nutr. Food Res. 2011;55:606–612. doi: 10.1002/mnfr.201000308. [DOI] [PubMed] [Google Scholar]
- 99.Konijeti R., Henning S., Moro A., Sheikh A., Elashoff D., Shapiro A., Ku M., Said J.W., Heber D., Cohen P., et al. Chemoprevention of prostate cancer with lycopene in the TRAMP model. Prostate. 2010;70:1547–1554. doi: 10.1002/pros.21190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Chen S., Gao J., Halicka H.D., Huang X., Traganos F., Darzynkiewicz Z. The cytostatic and cytotoxic effects of oridonin (Rubescenin), a diterpenoid from Rabdosia rubescens, on tumor cells of different lineage. Int. J. Oncol. 2005;26:579–588. doi: 10.3892/ijo.26.3.579. [DOI] [PubMed] [Google Scholar]
- 101.Li X., Li X., Wang J., Ye Z., Li J.C. Oridonin up-regulates expression of P21 and induces autophagy and apoptosis in human prostate cancer cells. Int. J. Biol. Sci. 2012;8:901–912. doi: 10.7150/ijbs.4554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Kaseb A.O., Chinnakannu K., Chen D., Sivanandam A., Tejwani S., Menon M., Dou Q.P., Reddy G.P. Androgen receptor and E2F-1 targeted thymoquinone therapy for hormone-refractory prostate cancer. Cancer Res. 2007;67:7782–7788. doi: 10.1158/0008-5472.CAN-07-1483. [DOI] [PubMed] [Google Scholar]
- 103.Koka P.S., Mondal D., Schultz M., Abdel-Mageed A.B., Agrawal K.C. Studies on molecular mechanisms of growth inhibitory effects of thymoquinone against prostate cancer cells: Role of reactive oxygen species. Exp. Biol. Med. 2010;235:751–760. doi: 10.1258/ebm.2010.009369. [DOI] [PubMed] [Google Scholar]
- 104.Yi T., Cho S.G., Yi Z., Pang X., Rodriguez M., Wang Y., Sethi G., Aggarwal B.B., Liu M. Thymoquinone inhibits tumor angiogenesis and tumor growth through suppressing AKT and extracellular signal-regulated kinase signaling pathways. Mol. Cancer Ther. 2008;7:1789–1796. doi: 10.1158/1535-7163.MCT-08-0124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Zhang Y., Kong C., Zeng Y., Wang L., Li Z., Wang H., Xu C., Sun Y. Ursolic acid induces PC-3 cell apoptosis via activation of JNK and inhibition of Akt pathways in vitro. Mol. Carcinog. 2010;49:374–385. doi: 10.1002/mc.20610. [DOI] [PubMed] [Google Scholar]
- 106.Zhang Y.X., Kong C.Z., Wang L.H., Li J.Y., Liu X.K., Xu B., Xu C.L., Sun Y.H. Ursolic acid overcomes Bcl-2-mediated resistance to apoptosis in prostate cancer cells involving activation of JNK-induced Bcl-2 phosphorylation and degradation. J. Cell Biochem. 2010;109:764–773. doi: 10.1002/jcb.22455. [DOI] [PubMed] [Google Scholar]
- 107.Shanmugam M.K., Manu K.A., Ong T.H., Ramachandran L., Surana R., Bist P., Lim L.H., Kumar A.P., Hui K.M., Sethi G. Inhibition of CXCR4/CXCL12 signaling axis by ursolic acid leads to suppression of metastasis in transgenic adenocarcinoma of mouse prostate model. Int. J. Cancer. 2011;129:1552–1563. doi: 10.1002/ijc.26120. [DOI] [PubMed] [Google Scholar]
- 108.Shanmugam M.K., Ong T.H., Kumar A.P., Lun C.K., Ho P.C., Wong P.T., Hui K.M., Sethi G. Ursolic acid inhibits the initiation, progression of prostate cancer and prolongs the survival of TRAMP mice by modulating pro-inflammatory pathways. PLoS ONE. 2012;7:e32476. doi: 10.1371/journal.pone.0032476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Bao X., Zhu J., Ren C., Zhao A., Zhang M., Zhu Z., Lu X., Zhang Y., Li X., Sima X., et al. beta-elemonic acid inhibits growth and triggers apoptosis in human castration-resistant prostate cancer cells through the suppression of JAK2/STAT3/MCL-1 and NF-kB signal pathways. Chem. Biol. Interact. 2021;342:109477. doi: 10.1016/j.cbi.2021.109477. [DOI] [PubMed] [Google Scholar]
- 110.Mellado B., Jimenez N., Marin-Aguilera M., Reig O. Diving Into Cabazitaxel’s Mode of Action: More Than a Taxane for the Treatment of Castration-Resistant Prostate Cancer Patients. Clin. Genitourin Cancer. 2016;14:265–270. doi: 10.1016/j.clgc.2015.12.030. [DOI] [PubMed] [Google Scholar]
- 111.Gan L., Wang J., Xu H., Yang X. Resistance to docetaxel-induced apoptosis in prostate cancer cells by p38/p53/p21 signaling. Prostate. 2011;71:1158–1166. doi: 10.1002/pros.21331. [DOI] [PubMed] [Google Scholar]
- 112.Haldar S., Chintapalli J., Croce C.M. Taxol induces bcl-2 phosphorylation and death of prostate cancer cells. Cancer Res. 1996;56:1253–1255. [PubMed] [Google Scholar]
- 113.Gan L., Chen S., Wang Y., Watahiki A., Bohrer L., Sun Z., Wang Y., Huang H. Inhibition of the androgen receptor as a novel mechanism of taxol chemotherapy in prostate cancer. Cancer Res. 2009;69:8386–8394. doi: 10.1158/0008-5472.CAN-09-1504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Haney K.M., Zhang F., Arnatt C.K., Yuan Y., Li G., Ware J.L., Gewirtz D.A., Zhang Y. The natural product CCR5 antagonist anibamine and its analogs as anti-prostate cancer agents. Bioorganic Med. Chem. Lett. 2011;21:5159–5163. doi: 10.1016/j.bmcl.2011.07.058. [DOI] [PubMed] [Google Scholar]
- 115.Wang Y., Liu Q., Liu Z., Li B., Sun Z., Zhou H., Zhang X., Gong Y., Shao C. Berberine, a genotoxic alkaloid, induces ATM-Chk1 mediated G2 arrest in prostate cancer cells. Mutagenesis Res. 2012;734:20–29. doi: 10.1016/j.mrfmmm.2012.04.005. [DOI] [PubMed] [Google Scholar]
- 116.Li J., Cao B., Liu X., Fu X., Xiong Z., Chen L., Sartor O., Dong Y., Zhang H. Berberine suppresses androgen receptor signaling in prostate cancer. Mol. Cancer Ther. 2011;10:1346–1356. doi: 10.1158/1535-7163.MCT-10-0985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Zhang Q., Zhang C., Yang X., Yang B., Wang J., Kang Y., Wang Z., Li D., Huang G., Ma Z., et al. Berberine inhibits the expression of hypoxia induction factor-1alpha and increases the radiosensitivity of prostate cancer. Diagn. Pathol. 2014;9:98. doi: 10.1186/1746-1596-9-98. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Sanchez A.M., Malagarie-Cazenave S., Olea N., Vara D., Chiloeches A., Diaz-Laviada I. Apoptosis induced by capsaicin in prostate PC-3 cells involves ceramide accumulation, neutral sphingomyelinase, and JNK activation. Apoptosis. 2007;12:2013–2024. doi: 10.1007/s10495-007-0119-z. [DOI] [PubMed] [Google Scholar]
- 119.Mori A., Lehmann S., O’Kelly J., Kumagai T., Desmond J.C., Pervan M., McBride W.H., Kizaki M., Koeffler H.P. Capsaicin, a component of red peppers, inhibits the growth of androgen-independent, p53 mutant prostate cancer cells. Cancer Res. 2006;66:3222–3229. doi: 10.1158/0008-5472.CAN-05-0087. [DOI] [PubMed] [Google Scholar]
- 120.Erdogan S., Turkekul K. Neferine inhibits proliferation and migration of human prostate cancer stem cells through p38 MAPK/JNK activation. J. Food Biochem. 2020;44:e13253. doi: 10.1111/jfbc.13253. [DOI] [PubMed] [Google Scholar]
- 121.Samykutty A., Shetty A.V., Dakshinamoorthy G., Bartik M.M., Johnson G.L., Webb B., Zheng G., Chen A., Kalyanasundaram R., Munirathinam G. Piperine, a Bioactive Component of Pepper Spice Exerts Therapeutic Effects on Androgen Dependent and Androgen Independent Prostate Cancer Cells. PLoS ONE. 2013;8:e65889. doi: 10.1371/journal.pone.0065889. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Ouyang D.Y., Zeng L.H., Pan H., Xu L.L.H., Wang Y., Liu K.P., He X.H. Piperine inhibits the proliferation of human prostate cancer cells via induction of cell cycle arrest and autophagy. Food Chem. Toxicol. 2013;60:424–430. doi: 10.1016/j.fct.2013.08.007. [DOI] [PubMed] [Google Scholar]
- 123.Makhov P., Golovine K., Canter D., Kutikov A., Simhan J., Corlew M.M., Uzzo R.G., Kolenko V.M. Co-administration of piperine and docetaxel results in improved anti-tumor efficacy via inhibition of CYP3A4 activity. Prostate. 2012;72:661–667. doi: 10.1002/pros.21469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Sun M., Liu C., Nadiminty N., Lou W., Zhu Y., Yang J., Evans C.P., Zhou Q., Gao A.C. Inhibition of Stat3 activation by sanguinarine suppresses prostate cancer cell growth and invasion. Prostate. 2012;72:82–89. doi: 10.1002/pros.21409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Sun M., Lou W., Chun J.Y., Cho D.S., Nadiminty N., Evans C.P., Chen J., Yue J., Zhou Q., Gao A.C. Sanguinarine suppresses prostate tumor growth and inhibits survivin expression. Genes Cancer. 2010;1:283–292. doi: 10.1177/1947601910368849. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Bodle C.R., Mackie D.I., Hayes M.P., Schamp J.H., Miller M.R., Henry M.D., Doorn J.A., Houtman J.C.D., James M.A., Roman D.L. Natural Products Discovered in a High-Throughput Screen Identified as Inhibitors of RGS17 and as Cytostatic and Cytotoxic Agents for Lung and Prostate Cancer Cell Lines. J. Nat. Prod. 2017;80:1992–2000. doi: 10.1021/acs.jnatprod.7b00112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Lu L., Tang D., Wang L., Huang L.Q., Jiang G.S., Xiao X.Y., Zeng F.Q. Gambogic acid inhibits TNF-alpha-induced invasion of human prostate cancer PC3 cells in vitro through PI3K/Akt and NF-kappaB signaling pathways. Acta Pharmacol. Sin. 2012;33:531–541. doi: 10.1038/aps.2011.180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Yi T., Yi Z., Cho S.G., Luo J., Pandey M.K., Aggarwal B.B., Liu M. Gambogic acid inhibits angiogenesis and prostate tumor growth by suppressing vascular endothelial growth factor receptor 2 signaling. Cancer Res. 2008;68:1843–1850. doi: 10.1158/0008-5472.CAN-07-5944. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Zhou Y., Yang G., Tian H., Hu Y., Wu S., Geng Y., Lin K., Wu W. Sulforaphane metabolites cause apoptosis via microtubule disruption in cancer. Endocr. -Relat. Cancer. 2018;25:255–268. doi: 10.1530/ERC-17-0483. [DOI] [PubMed] [Google Scholar]
- 130.Ferreira P.M.P., Rodrigues L., de Alencar Carnib L.P., de Lima Sousa P.V., Nolasco Lugo L.M., Nunes N.M.F., do Nascimento Silva J., da Silva Araujo L., de Macedo Goncalves Frota K. Cruciferous Vegetables as Antioxidative, Chemopreventive and Antineoplasic Functional Foods: Preclinical and Clinical Evidences of Sulforaphane Against Prostate Cancers. Curr. Pharm. Des. 2018;24:4779–4793. doi: 10.2174/1381612825666190116124233. [DOI] [PubMed] [Google Scholar]
- 131.Gibbs A., Schwartzman J., Deng V., Alumkal J. Sulforaphane destabilizes the androgen receptor in prostate cancer cells by inactivating histone deacetylase 6. Proc. Natl. Acad. Sci. USA. 2009;106:16663–16668. doi: 10.1073/pnas.0908908106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Clarke J.D., Hsu A., Yu Z., Dashwood R.H., Ho E. Differential effects of sulforaphane on histone deacetylases, cell cycle arrest and apoptosis in normal prostate cells versus hyperplastic and cancerous prostate cells. Mol. Nutr. Food Res. 2011;55:999–1009. doi: 10.1002/mnfr.201000547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Singh K.B., Kim S.H., Hahm E.R., Pore S.K., Jacobs B.L., Singh S.V. Prostate cancer chemoprevention by sulforaphane in a preclinical mouse model is associated with inhibition of fatty acid metabolism. Carcinogenesis. 2018;39:826–837. doi: 10.1093/carcin/bgy051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Beaver L.M., Lhr C.V., Clarke J.D., Glasser S.T., Watson G.W., Wong C.P., Zhang Z., Williams D.E., Dashwood R.H., Shannon J., et al. Broccoli Sprouts Delay Prostate Cancer Formation and Decrease Prostate Cancer Severity with a Concurrent Decrease in HDAC3 Protein Expression in Transgenic Adenocarcinoma of the Mouse Prostate (TRAMP) Mice. Curr. Dev. Nutr. 2018;2:nzy002. doi: 10.1093/cdn/nzy002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Yin P., Kawamura T., He M., Vanaja D.K., Young C.Y. Phenethyl isothiocyanate induces cell cycle arrest and reduction of alpha- and beta-tubulin isotypes in human prostate cancer cells. Cell Biol. Int. 2009;33:57–64. doi: 10.1016/j.cellbi.2008.09.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Sakao K., Desineni S., Hahm E.R., Singh S.V. Phenethyl isothiocyanate suppresses inhibitor of apoptosis family protein expression in prostate cancer cells in culture and in vivo. Prostate. 2012;72:1104–1116. doi: 10.1002/pros.22457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Kim S.H., Sehrawat A., Sakao K., Hahm E.R., Singh S.V. Notch activation by phenethyl isothiocyanate attenuates its inhibitory effect on prostate cancer cell migration. PLoS ONE. 2011;6:e26615. doi: 10.1371/journal.pone.0026615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Hudson T.S., Perkins S.N., Hursting S.D., Young H.A., Kim Y.S., Wang T.C., Wang T.T. Inhibition of androgen-responsive LNCaP prostate cancer cell tumor xenograft growth by dietary phenethyl isothiocyanate correlates with decreased angiogenesis and inhibition of cell attachment. Int. J. Oncol. 2012;40:1113–1121. doi: 10.3892/ijo.2012.1335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Powolny A.A., Bommareddy A., Hahm E.R., Normolle D.P., Beumer J.H., Nelson J.B., Singh S.V. Chemopreventative potential of the cruciferous vegetable constituent phenethyl isothiocyanate in a mouse model of prostate cancer. J. Natl. Cancer Inst. 2011;103:571–584. doi: 10.1093/jnci/djr029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Aggarwal B.B., Ichikawa H. Molecular targets and anticancer potential of indole-3-carbinol and its derivatives. Cell Cycle. 2005;4:1201–1215. doi: 10.4161/cc.4.9.1993. [DOI] [PubMed] [Google Scholar]
- 141.Chahar M.K., Sharma N., Dobhal M.P., Joshi Y.C. Flavonoids: A versatile source of anticancer drugs. Pharm. Rev. 2011;5:1–12. doi: 10.4103/0973-7847.79093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Rauf A., Imran M., Khan I.A., Ur-Rehman M., Gilani S.A., Mehmood Z., Mubarak M.S. Anticancer potential of quercetin: A comprehensive review. Phytother. Res. 2018;32:2109–2130. doi: 10.1002/ptr.6155. [DOI] [PubMed] [Google Scholar]
- 143.Kim Y.H., Lee D.H., Jeong J.H., Guo Z.S., Lee Y.J. Quercetin augments TRAIL-induced apoptotic death: Involvement of the ERK signal transduction pathway. Biochem. Pharm. 2008;75:1946–1958. doi: 10.1016/j.bcp.2008.02.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Pratheeshkumar P., Budhraja A., Son Y.O., Wang X., Zhang Z., Ding S., Wang L., Hitron A., Lee J.C., Xu M., et al. Quercetin inhibits angiogenesis mediated human prostate tumor growth by targeting VEGFR- 2 regulated AKT/mTOR/P70S6K signaling pathways. PLoS ONE. 2012;7:e47516. doi: 10.1371/journal.pone.0047516. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Shukla S., Gupta S. Molecular targets for apigenin-induced cell cycle arrest and apoptosis in prostate cancer cell xenograft. Mol. Cancer Ther. 2006;5:843–852. doi: 10.1158/1535-7163.MCT-05-0370. [DOI] [PubMed] [Google Scholar]
- 146.Shukla S., Bhaskaran N., Babcook M.A., Fu P., Maclennan G.T., Gupta S. Apigenin inhibits prostate cancer progression in TRAMP mice via targeting PI3K/Akt/FoxO pathway. Carcinogenesis. 2014;35:452–460. doi: 10.1093/carcin/bgt316. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Hafeez B.B., Siddiqui I.A., Asim M., Malik A., Afaq F., Adhami V.M., Saleem M., Din M., Mukhtar H. A dietary anthocyanidin delphinidin induces apoptosis of human prostate cancer PC3 cells in vitro and in vivo: Involvement of nuclear factor-kappaB signaling. Cancer Res. 2008;68:8564–8572. doi: 10.1158/0008-5472.CAN-08-2232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Henning S.M., Wang P., Heber D. Chemopreventive effects of tea in prostate cancer: Green tea versus black tea. Mol. Nutr. Food Res. 2011;55:905–920. doi: 10.1002/mnfr.201000648. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Wang P., Henning S.M., Heber D., Vadgama J.V. Sensitization to docetaxel in prostate cancer cells by green tea and quercetin. J. Nutr. Biochem. 2015;26:408–415. doi: 10.1016/j.jnutbio.2014.11.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Wang P., Henning S.M., Magyar C.E., Elshimali Y., Heber D., Vadgama J.V. Green tea and quercetin sensitize PC-3 xenograft prostate tumors to docetaxel chemotherapy. J. Exp. Clin. Cancer Res. 2016;35:73. doi: 10.1186/s13046-016-0351-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Vodnik V.V., Mojic M., Stamenovic U., Otonicar M., Ajdzanovic V., Maksimovic-Ivanic D., Mijatovic S., Markovic M.M., Barudzija T., Filipovic B., et al. Development of genistein-loaded gold nanoparticles and their antitumor potential against prostate cancer cell lines. Mater. Sci. Eng. C Mater. Biol. Appl. 2021;124:112078. doi: 10.1016/j.msec.2021.112078. [DOI] [PubMed] [Google Scholar]
- 152.Zhang L., Li L., Jiao M., Wu D., Wu K., Li X., Zhu G., Yang L., Wang X., Hsieh J.T., et al. Genistein inhibits the stemness properties of prostate cancer cells through targeting Hedgehog-Gli1 pathway. Cancer Lett. 2012;323:48–57. doi: 10.1016/j.canlet.2012.03.037. [DOI] [PubMed] [Google Scholar]
- 153.Adjakly M., Bosviel R., Rabiau N., Boiteux J.P., Bignon Y.J., Guy L., Bernard-Gallon D. DNA methylation and soy phytoestrogens: Quantitative study in DU-145 and PC-3 human prostate cancer cell lines. Epigenomics. 2011;3:795–803. doi: 10.2217/epi.11.103. [DOI] [PubMed] [Google Scholar]
- 154.Mentor-Marcel R., Lamartiniere C.A., Eltoum I.E., Greenberg N.M., Elgavish A. Genistein in the diet reduces the incidence of poorly differentiated prostatic adenocarcinoma in transgenic mice (TRAMP) Cancer Res. 2001;61:6777–6782. [PubMed] [Google Scholar]
- 155.Yo Y.T., Shieh G.S., Hsu K.F., Wu C.L., Shiau A.L. Licorice and licochalcone-A induce autophagy in LNCaP prostate cancer cells by suppression of Bcl-2 expression and the mTOR pathway. J. Agric. Food Chem. 2009;57:8266–8273. doi: 10.1021/jf901054c. [DOI] [PubMed] [Google Scholar]
- 156.Shoulars K., Rodriguez M.A., Thompson T., Markaverich B.M. Regulation of cell cycle and RNA transcription genes identified by microarray analysis of PC-3 human prostate cancer cells treated with luteolin. J. Steroid Biochem. Mol. Biol. 2010;118:41–50. doi: 10.1016/j.jsbmb.2009.09.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Kavitha C.V., Deep G., Gangar S.C., Jain A.K., Agarwal C., Agarwal R. Silibinin inhibits prostate cancer cells- and RANKL-induced osteoclastogenesis by targeting NFATc1, NF-kappaB, and AP-1 activation in RAW264.7 cells. Mol. Carcinog. 2014;53:169–180. doi: 10.1002/mc.21959. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Singh R.P., Deep G., Blouin M.J., Pollak M.N., Agarwal R. Silibinin suppresses in vivo growth of human prostate carcinoma PC-3 tumor xenograft. Carcinogenesis. 2007;28:2567–2574. doi: 10.1093/carcin/bgm218. [DOI] [PubMed] [Google Scholar]
- 159.Shah S., Prasad S., Knudsen K.E. Targeting pioneering factor and hormone receptor cooperative pathways to suppress tumor progression. Cancer Res. 2012;72:1248–1259. doi: 10.1158/0008-5472.CAN-11-0943. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Li W., Su Z.Y., Guo Y., Zhang C., Wu R., Gao L., Zheng X., Du Z.Y., Zhang K., Kong A.N. Curcumin Derivative Epigenetically Reactivates Nrf2 Antioxidative Stress Signaling in Mouse Prostate Cancer TRAMP C1 Cells. Chem. Res. Toxicol. 2018;31:88–96. doi: 10.1021/acs.chemrestox.7b00248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Applegate C.C., Rowles J.L., 3rd, Erdman J.W., Jr. Can Lycopene Impact the Androgen Axis in Prostate Cancer?: A Systematic Review of Cell Culture and Animal Studies. Nutrients. 2019;11:633. doi: 10.3390/nu11030633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Hurwitz M. Chemotherapy in Prostate Cancer. Curr. Oncol. Rep. 2015;17:44. doi: 10.1007/s11912-015-0468-7. [DOI] [PubMed] [Google Scholar]
- 163.De Leeuw R., Berman-Booty L.D., Schiewer M.J., Ciment S.J., Den R.B., Dicker A.P., Kelly W.K., Trabulsi E.J., Lallas C.D., Gomella L.G., et al. Novel actions of next-generation taxanes benefit advanced stages of prostate cancer. Clin. Cancer Res. 2015;21:795–807. doi: 10.1158/1078-0432.CCR-14-1358. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Vrignaud P., Semiond D., Lejeune P., Bouchard H., Calvet L., Combeau C., Riou J.F., Commercon A., Lavelle F., Bissery M.C. Preclinical antitumor activity of cabazitaxel, a semisynthetic taxane active in taxane-resistant tumors. Clin. Cancer Res. 2013;19:2973–2983. doi: 10.1158/1078-0432.CCR-12-3146. [DOI] [PubMed] [Google Scholar]
- 165.Sanchez A.M., Sanchez M.G., Malagarie-Cazenave S., Olea N., Diaz-Laviada I. Induction of apoptosis in prostate tumor PC-3 cells and inhibition of xenograft prostate tumor growth by the vanilloid capsaicin. Apoptosis. 2006;11:89–99. doi: 10.1007/s10495-005-3275-z. [DOI] [PubMed] [Google Scholar]
- 166.Maina S., Misinzo G., Bakari G., Kim H.Y. Human, Animal and Plant Health Benefits of Glucosinolates and Strategies for Enhanced Bioactivity: A Systematic Review. Molecules. 2020;25:3682. doi: 10.3390/molecules25163682. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Livingstone T.L., Beasy G., Mills R.D., Plumb J., Needs P.W., Mithen R., Traka M.H. Plant Bioactives and the Prevention of Prostate Cancer: Evidence from Human Studies. Nutrients. 2019;11:2245. doi: 10.3390/nu11092245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Atwell L.L., Beaver L.M., Shannon J., Williams D.E., Dashwood R.H., Ho E. Epigenetic Regulation by Sulforaphane: Opportunities for Breast and Prostate Cancer Chemoprevention. Curr. Pharm. Rep. 2015;1:102–111. doi: 10.1007/s40495-014-0002-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Saw C.L., Cintron M., Wu T.Y., Guo Y., Huang Y., Jeong W.S., Kong A.N. Pharmacodynamics of dietary phytochemical indoles I3C and DIM: Induction of Nrf2-mediated phase II drug metabolizing and antioxidant genes and synergism with isothiocyanates. Biopharm. Drug Dispos. 2011;32:289–300. doi: 10.1002/bdd.759. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Hahm E.R., Chandra-Kuntal K., Desai D., Amin S., Singh S.V. Notch activation is dispensable for D, L-sulforaphane-mediated inhibition of human prostate cancer cell migration. PLoS ONE. 2012;7:e44957. doi: 10.1371/journal.pone.0044957. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Tang N.Y., Huang Y.T., Yu C.S., Ko Y.C., Wu S.H., Ji B.C., Yang J.S., Yang J.L., Hsia T.C., Chen Y.Y., et al. Phenethyl isothiocyanate (PEITC) promotes G2/M phase arrest via p53 expression and induces apoptosis through caspase- and mitochondria-dependent signaling pathways in human prostate cancer DU 145 cells. Anticancer Res. 2011;31:1691–1702. [PubMed] [Google Scholar]