

Abstract
Cryptosporidium parvum induces moderate levels of apoptosis of cultured human intestinal epithelial cells, which are maximal at 24 h after infection. Apoptosis is further increased in C. parvum-infected cells by inhibition of NF-κB. C. parvum infection also attenuates epithelial apoptosis induced by strongly proapoptotic agents. The data suggest C. parvum has developed strategies to limit apoptosis in order to facilitate its growth and maturation in the early period after epithelial cell infection.
The protozoan parasite Cryptosporidium parvum causes diarrhea in both immunocompetent and immunosuppressed hosts. The primary site of C. parvum infection is the epithelium of the intestine, although epithelial cells in extraintestinal sites, including the stomach and biliary and respiratory tract, can also be infected (13, 27, 35). The infective stage of C. parvum is the oocyst, which usually contains four sporozoites. Following excystation, sporozoites attach to the intestinal epithelium, are enveloped by the apical membrane, and reside in an intracellular, extracytoplasmic parasitophorous vacuole (15, 21, 28). C. parvum undergoes a series of asexual reproductive stages in the first 48 h after infection in vivo and in cell culture (34). Infection of the intestinal epithelium with C. parvum can result in blunting of intestinal villi, crypt hyperplasia, and cytoskeletal remodeling, as well as decreased sodium absorption, increased prostaglandin production, and epithelial chemokine secretion (1, 2, 11–13, 24, 25, 33). C. parvum infection of intestinal epithelial cells in vitro also results in cell detachment and the apical release of the cytosolic enzyme lactate dehydrogenase (14, 25), but the exact nature of this cytopathic effect is poorly understood. Apoptosis is a regulated process of cell death that can be signaled from the external environment or from within the cell and, in contrast to intestinal epithelial cell necrosis, results in little disruption of intestinal epithelial barrier integrity (18). Apoptosis occurs in response to infection with several invasive and noninvasive microbial pathogens of the human gastrointestinal tract, including Salmonella spp., Shigella spp., enteropathogenic Escherichia coli, human immunodeficiency virus type 1, and Helicobacter pylori (6, 8, 17, 22, 23). Since C. parvum resides and undergoes critical phases of its life cycle within the intestinal epithelium, we investigated whether this pathogen has developed strategies to alter epithelial cell apoptosis that may enhance its survival within that environment.
Cells of the human ileocecal adenocarcinoma line HCT-8 (ATCC CCL 244) and colonic adenocarcinoma line Caco-2 (ATCC HTB 37) were grown in RPMI 1640 or Dulbecco's modified Eagle's medium, respectively, supplemented with 10% heat-inactivated fetal calf serum, 2 mM l-glutamine, 50 U of penicillin G per ml, and 50 μg of streptomycin per ml (23, 24). Calpain-1 inhibitor was from Calbiochem, La Jolla, Calif., and etoposide, 5-fluorouracil, and staurosporine were from Sigma Chemical Co., St. Louis, Mo. C. parvum was maintained and oocysts were isolated and used for infection as described before (24, 25). Recombinant adenovirus containing an IκBα-AA superrepressor (Ad5IκB-A32/36) was constructed as described before (9). Monolayers of HCT-8 or Caco-2 cells at 80 to 100% confluency in six-well Costar tissue culture plates were infected with oocysts at a ratio of three oocysts per one cell for 5 h, after which cells were washed and fresh supplemented medium was added. Cells were then incubated for an additional 12 to 48 h, after which nonadherent and adherent cells were removed, the latter by treatment with 0.25% trypsin–1 mM EDTA for 5 min. To detect epithelial cell apoptosis, cells were pooled, fixed with 4% formalin, stained with the DNA binding dye Hoechst 33258 (4 μg/ml), deposited on glass microscope slides by a cytocentrifuge, and analyzed by epifluorescence microscopy. Cells were defined as apoptotic based on compaction and segregation of chromatin into dense masses, segmentation of nuclei, and formation of apoptotic bodies. Nonapoptotic epithelial cells displayed intact regularly shaped nuclei and normal chromatin distribution. Five hundred cells were examined (100 in each of five separate fields), and the number of apoptotic cells was expressed as a percentage of the total number of cells examined. Data obtained by Hoechst staining were confirmed by assessing cleavage of keratin 18 and reorganization of intermediate filaments as a measure of apoptosis (4). Fragmented cytokeratin 18 was detected by using acetone-fixed cells with M30 cytoDeath (Roche Molecular Biochemicals, Indianapolis, Ind.) as the primary antibody and Cy2-labeled secondary antibody. To detect C. parvum-infected cells, fixed cells were stained with a rat anti-C. parvum serum (1:500) (24) and a 1:1,000 dilution of Cy3-labelled goat anti-rat immunoglobulin G (heavy plus light chains) (Amersham Corporation, Arlington Heights, Ill.), followed by staining with Hoechst 33258 (4 μg/ml). Statistical analysis was performed with a two-tailed Student t test.
Figure 1 demonstrates that the percentage of cells with apoptotic morphology significantly increased in C. parvum-infected HCT-8 cultures, compared to control uninfected cultures, over the first 24 h after infection. C. parvum infection also increased apoptosis of Caco-2 cells (11.7% ± 0.9% apoptotic cells in infected cultures versus 1.3% ± 0.3% in uninfected cultures at 24 h after infection [P < 0.0001, n = 3]). The percentage of intestinal epithelial cells undergoing apoptosis and the relatively slow kinetics of apoptosis resemble those recently reported for C. parvum-infected human biliary tract epithelial cells (5).
FIG. 1.

Time course of apoptosis in HCT-8 cells infected with C. parvum. Apoptotic cells were determined by staining with Hoechst 33258 dye. The percentage of apoptotic cells is shown for C. parvum-infected cells (●) and uninfected control cells (○). Data are means ± standard errors of four repeated experiments. Similar findings were obtained by assessing caspase cleavage of keratin 18 as a measure of apoptosis (data not shown).
To determine if the epithelial cells undergoing apoptosis were those directly infected with C. parvum, cells were double stained for C. parvum and with Hoechst 33258 dye to detect apoptosis or triple stained additionally with M30 antibody to detect caspase-cleaved cytokeratin 18 (Fig. 2). More than 93% of the cells undergoing apoptosis were concomitantly infected with C. parvum, although only a fraction (21%) of C. parvum-infected cells exhibited signs of apoptosis 24 h after infection (Table 1). Nonetheless within C. parvum-infected cultures, apoptosis was increased by 6.5-fold in infected compared with uninfected cells. This suggests that cellular infection is directly required for the induction of epithelial cell apoptosis and that the possible release of mediators from infected cells which might induce apoptosis in adjacent uninfected cells, as has been demonstrated in Salmonella-infected epithelial cell cultures (23), plays a minor, if any, role in the apoptotic response to C. parvum.
FIG. 2.
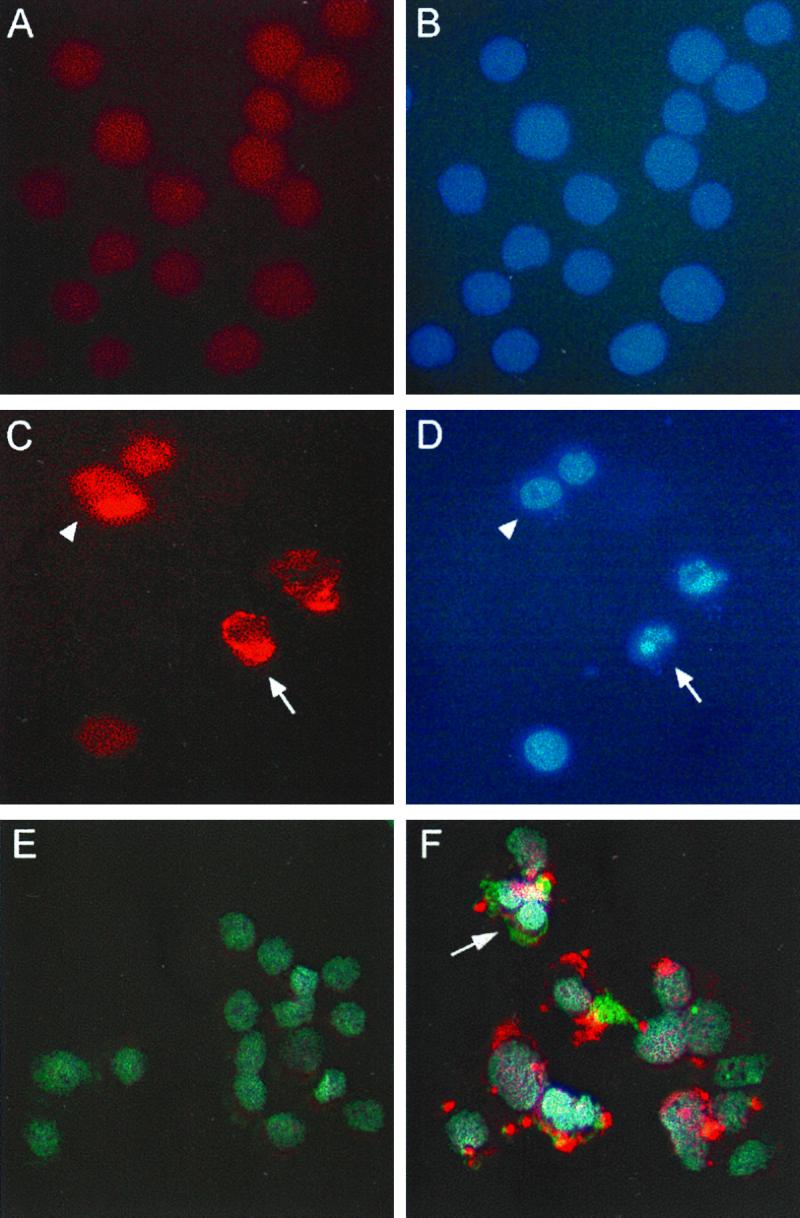
Colocalization of C. parvum infection and apoptosis in HCT-8 cells. Confluent monolayers of uninfected (A and B) and C. parvum-infected (C and D) HCT-8 cells were fixed 24 h after infection and stained for C. parvum and with Hoechst 33258 dye. Panels A and C are photomicrographs obtained with an Omega optical XF34 fluorescence filter to visualize C. parvum staining, while panels B and D are photomicrographs from the respective identical fields obtained with an Omega optical XF05 filter to visualize staining with Hoechst dye. A cell infected with C. parvum and undergoing apoptosis is indicated by the arrows, and a nonapoptotic cell infected with C. parvum is indicated by the solid arrowheads. Panels E and F display triple staining of uninfected (E) and C. parvum-infected (F) HCT-8 cultures. C. parvum was stained with Cy3-conjugated secondary antibody (red), nuclear morphology was stained with Hoechst 33258 dye (blue), and cleaved cytokeratin 18 was detected with a Cy2-conjugated secondary antibody (green), which was visualized with an Omega optical XF23 filter. Triple-staining images were layered by using the program Adobe Photoshop. The arrow depicts a C. parvum-infected cell undergoing apoptosis, as indicated by apoptotic nuclear morphology and the presence of cleaved cytokeratin 18. Overlapping areas of green and red appear as yellow. Original magnification, ×630.
TABLE 1.
Enumeration of apoptotic and nonapoptotic cells in C. parvum-infected HCT-8 culturesa
| Cells | % of cells
|
Apoptotic/total cell ratio | ||
|---|---|---|---|---|
| Apoptotic | Nonapoptotic | Total | ||
| C. parvum infected | 15 | 55 | 70 | 0.214 |
| Uninfected | 1 | 29 | 30 | 0.033 |
| Total | 16 | 84 | 100 | |
Apoptosis levels were assessed 24 h after infection by using Hoechst 33258 dye. The results are the means of two separate experiments.
The transcription factor NF-κB has been shown to prevent apoptosis of tumor necrosis factor alpha-stimulated cells in several experimental models (3, 26, 36, 37). Moreover, activation of NF-κB by infection of T cells with the parasite Theileria parva or endothelial cells with Rickettsia rickettsii protected infected cells from undergoing apoptosis (7, 16). Since C. parvum infection of intestinal epithelial cells is paralleled by the activation of NF-κB target genes (24, 25, 33), we investigated whether the relatively low proportion of infected cells undergoing apoptosis in response to C. parvum infection reflects a protective effect of NF-κB activation. C. parvum-infected HCT-8 monolayers were treated for 24 h with calpain-1 inhibitor (25 μM), a calcium proteinase inhibitor which prevents the activation of NF-κB by blocking IκB degradation (29). Note that there was a significant increase in the proportion of cells undergoing apoptosis in C. parvum-infected cultures treated with calpain-1 inhibitor, as follows. There was 1.7% ± 0.3% (mean ± standard error) apoptosis in control cells (n = 11 experiments) given no treatment versus 11.2% ± 1.1% apoptosis in infected cells (n = 11) given no treatment. There was 2.5% ± 1.2% apoptosis in control cells treated with calpain-1 inhibitor (n = 4) versus 26.8% ± 6.6% apoptosis in infected cells treated with calpain-1 inhibitor (n = 4). The difference between calpain-1 inhibitor-treated and untreated control cells was not significant, whereas the difference between calpain-1 inhibitor-treated infected cells versus untreated infected cells was significant (P < 0.05).
Since pharmacologic agents are not always completely specific, we used an additional approach in which cells were infected with a recombinant adenovirus expressing a mutant IκBα protein that has serine-to-alanine substitutions at positions 32 and 36 (Ad5IκB-A32/36) and acts as a superrepressor of NF-κB activation by preventing signal-induced IκBα phosphorylation (9). Subsequent to adenovirus infection, HCT-8 cells were infected with C. parvum, and apoptosis was assessed 24 h later. As shown in Table 2, apoptosis in response to C. parvum infection markedly increased in Ad5IκB-A32/36-infected cells, but not in cells infected with control adenovirus. Taken together, these findings suggest that C. parvum-induced apoptosis is limited by the concomitant activation of NF-κB. When NF-κB activation is blocked, the apoptosis-inducing capacity of C. parvum is increased, and a significantly greater fraction of infected cells undergoes apoptosis.
TABLE 2.
A superrepressor of NF-κB activation increases apoptosis in C. parvum-infected HCT-8 cells
| Treatment | % Apoptosis in HCT-8 cellsa
|
|
|---|---|---|
| Uninfected | C. parvum infected | |
| None | 1.5 ± 0.3 | 9.7 ± 1.9 |
| Ad5IκB-A32/36 | 6.7 ± 1.5 | 33.3 ± 1.2 |
| Ad5 vector control | 6.7 ± 0.3 | 10.7 ± 2.0 |
Data are means ± standard errors of three repeated experiments.
Pathogens such as Toxoplasma gondii and Chlamydia trachomatis, which alone did not significantly induce apoptosis of target cells, have been shown to decrease apoptosis of infected cells challenged with apoptosis-inducing agents (7, 10, 30). We investigated, therefore, whether C. parvum infection of intestinal epithelial cells could attenuate apoptosis in response to known inducers of apoptosis. For these experiments, HCT-8 monolayers were infected with C. parvum and then challenged for 24 h with staurosporine, an inhibitor of protein kinase C, or etoposide, a DNA topoisomerase II inhibitor, or were challenged for 48 h with 5-fluorouracil, a thymidine monophosphate synthesis inhibitor (19, 20, 31, 32). As shown in Table 3, C. parvum significantly attenuated apoptosis induced by these agents.
TABLE 3.
Apoptosis induction by chemical agents is attenuated in HCT-8 cells infected with C. parvum
| Treatment | % Apoptosis in HCT-8 cellsa
|
P (infected vs control) | |
|---|---|---|---|
| Control | Infected | ||
| None | 1.7 ± 0.3 (11) | 11.2 ± 1.1 (11) | <0.001 |
| Staurosporine | 41.1 ± 2.8 (11) | 22.0 ± 2.5 (11) | <0.005 |
| Etoposide | 35.5 ± 0.3 (4) | 18.0 ± 0.5 (4) | <0.001 |
| 5-Fluorouracil | 57.8 ± 3.9 (4) | 27.0 ± 3.5 (4) | <0.001 |
Data are means ± standard errors for n experiments (indicated in parentheses).
These data show that C. parvum induces a moderate degree of apoptosis in intestinal epithelial cells yet inhibits apoptosis of these cells in response to strong proapoptotic stimuli. The most likely explanation for these findings is that C. parvum prevents induction of high levels of epithelial cell apoptosis early after infection, when the parasite depends on the host cell for growth and development. Induction of moderate levels of epithelial cell apoptosis, rather than necrosis, early after infection may also limit the host inflammatory response, which could be detrimental to the survival of the parasite. On the other hand, deletion of infected epithelial cells by apoptosis may benefit the host, since it allows maintenance of epithelial barrier integrity.
Acknowledgments
This work was supported by National Institutes of Health grant DK35108.
We thank N. Varki for assistance with immunostaining analysis.
REFERENCES
- 1.Argenzio R A, Lecce J, Powell D W. Prostanoids inhibit intestinal NaCl absorption in experimental porcine cryptosporidiosis. Gastroenterology. 1993;104:440–447. doi: 10.1016/0016-5085(93)90412-6. [DOI] [PubMed] [Google Scholar]
- 2.Argenzio R A, Liacos J A, Levy M L, Meuten D J, Lecce J G, Powell D W. Villous atrophy, crypt hyperplasia, cellular infiltration, and impaired glucose-Na absorption in enteric cryptosporidiosis of pigs. Gastroenterology. 1990;98:1129–1140. doi: 10.1016/0016-5085(90)90325-u. [DOI] [PubMed] [Google Scholar]
- 3.Beg A A, Baltimore D. An essential role for NF-κB in preventing TNF-α-induced cell death. Science. 1996;274:782–784. doi: 10.1126/science.274.5288.782. [DOI] [PubMed] [Google Scholar]
- 4.Caulin C, Salvesen G S, Oshima R G. Caspase cleavage of keratin 18 and reorganization of intermediate filaments during epithelial cell apoptosis. J Cell Biol. 1997;138:1379–1394. doi: 10.1083/jcb.138.6.1379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Chen X-M, Gores G J, Paya C V, LaRusso N F. Cryptosporidium parvum induces apoptosis in biliary epithelia by a Fas/Fas ligand-dependent mechanism. Am J Physiol. 1999;277:G599–G608. doi: 10.1152/ajpgi.1999.277.3.G599. [DOI] [PubMed] [Google Scholar]
- 6.Clayton F, Reka S, Cronin W J, Torlakovic E, Sigal S H, Kotler D P. Rectal mucosal pathology varies with human immunodeficiency virus antigen content and disease stage. Gastroenterology. 1992;103:919–933. doi: 10.1016/0016-5085(92)90026-u. [DOI] [PubMed] [Google Scholar]
- 7.Clifton D R, Goss R A, Sahni S K, van Antwerp D, Baggs R B, Marder V J, Silverman D J, Sporn L A. NF-κB-dependent inhibition of apoptosis is essential for host cell survival during Rickettsia rickettsii infection. Proc Natl Acad Sci USA. 1998;95:4646–4651. doi: 10.1073/pnas.95.8.4646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Crane J K, Majumdar S, Pickhardt D F., III Host cell death due to enteropathogenic Escherichia coli has features of apoptosis. Infect Immun. 1999;67:2575–2584. doi: 10.1128/iai.67.5.2575-2584.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Elewaut D, DiDonato J A, Kim J M, Truong F, Eckmann L, Kagnoff M F. NF-κB is a central regulator of the intestinal epithelial cell innate immune response induced by infection with enteroinvasive bacteria. J Immunol. 1999;163:1457–1466. [PubMed] [Google Scholar]
- 10.Fan T, Lu H, Hu H, Shi L, McClarty G A, Nance D M, Greenberg A H, Zhong G. Inhibition of apoptosis in chlamydia-infected cells: blockade of mitochondrial cytochrome c release and caspase activation. J Exp Med. 1998;187:487–496. doi: 10.1084/jem.187.4.487. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Forney J R, DeWald D B, Yang S, Speer C A, Healey M C. A role for host phosphoinositide 3-kinase and cytoskeletal remodeling during Cryptosporidium parvum infection. Infect Immun. 1999;67:844–852. doi: 10.1128/iai.67.2.844-852.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Genta R M, Chappell C L, White A C J, Kimball K T, Goodgame R W. Duodenal morphology and intensity of infection in AIDS-related intestinal cryptosporidiosis. Gastroenterology. 1993;105:1769–1775. doi: 10.1016/0016-5085(93)91075-s. [DOI] [PubMed] [Google Scholar]
- 13.Godwin T A. Cryptosporidiosis in the acquired immunodeficiency syndrome: a study of 15 autopsy cases. Hum Pathol. 1991;22:1215–1224. doi: 10.1016/0046-8177(91)90103-v. [DOI] [PubMed] [Google Scholar]
- 14.Griffiths J K, Moore R, Dooley S, Keusch G T, Tzipori S. Cryptosporidium parvum infection of Caco-2 cell monolayers induces an apical monolayer defect, selectively increases transmonolayer permeability, and causes epithelial cell death. Infect Immun. 1994;62:4506–4514. doi: 10.1128/iai.62.10.4506-4514.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Hamer D H, Ward H, Tzipori S, Pereira M E A, Alroy J P, Keusch G T. Attachment of Cryptosporidium parvum sporozoites to MDCK cells in vitro. Infect Immun. 1994;62:2208–2213. doi: 10.1128/iai.62.6.2208-2213.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Heussler V T, Machado J J, Fernandez P C, Botteron C, Chen C G, Pearse M J, Dobbelaere D A. The intracellular parasite Theileria parva protects infected T cells from apoptosis. Proc Natl Acad Sci USA. 1999;96:7312–7317. doi: 10.1073/pnas.96.13.7312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Islam D, Veress B, Bardhan P K, Lindberg A A, Christensson B. In situ characterization of inflammatory responses in the rectal mucosae of patients with shigellosis. Infect Immun. 1997;65:739–749. doi: 10.1128/iai.65.2.739-749.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Iwanaga T, Han H, Adachi K, Fujita T. A novel mechanism for disposing of effete epithelial cells in the small intestine of guinea pigs. Gastroenterology. 1993;105:1089–1097. doi: 10.1016/0016-5085(93)90953-a. [DOI] [PubMed] [Google Scholar]
- 19.Jacobson M D, Weil M, Raff M C. Role of CED-3/ICE-family proteases in staurosporine-induced programmed cell death. J Cell Biol. 1996;133:1041–1051. doi: 10.1083/jcb.133.5.1041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Jiang S, Song M J, Shin E C, Lee M O, Kim S J, Park J H. Apoptosis in human hepatoma cell lines by chemotherapeutic drugs via Fas-dependent and Fas-independent pathways. Hepatology. 1999;29:101–110. doi: 10.1002/hep.510290102. [DOI] [PubMed] [Google Scholar]
- 21.Joe A, Verdon R, Tzipori S, Keusch G T, Ward H D. Attachment of Cryptosporidium parvum sporozoites to human intestinal epithelial cells. Infect Immun. 1998;66:3429–3432. doi: 10.1128/iai.66.7.3429-3432.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Jones N L, Day A S, Jennings H A, Sherman P M. Helicobacter pylori induces gastric epithelial cell apoptosis in association with increased Fas receptor expression. Infect Immun. 1999;67:4237–4242. doi: 10.1128/iai.67.8.4237-4242.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Kim J M, Eckmann L, Savidge T C, Lowe D C, Witthoft T, Kagnoff M F. Apoptosis of human intestinal epithelial cells after bacterial invasion. J Clin Investig. 1998;102:1815–1823. doi: 10.1172/JCI2466. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Laurent F, Eckmann L, Savidge T C, Morgan G, Theodos C, Naciri M, Kagnoff M F. Cryptosporidium parvum infection of human intestinal epithelial cells induces the polarized secretion of C-X-C chemokines. Infect Immun. 1997;65:5067–5073. doi: 10.1128/iai.65.12.5067-5073.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Laurent F, Kagnoff M F, Savidge T C, Naciri M, Eckmann L. Human intestinal epithelial cells respond to Cryptosporidium parvum infection with increased prostaglandin H synthase 2 expression and prostaglandin E2 and F2α production. Infect Immun. 1998;66:1787–1790. doi: 10.1128/iai.66.4.1787-1790.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Liu Z-G, Hsu H, Goeddel D V, Karin M. Dissection of TNF receptor 1 effector functions: JNK activation is not linked to apoptosis while NF-κB activation prevents cell death. Cell. 1996;87:565–576. doi: 10.1016/s0092-8674(00)81375-6. [DOI] [PubMed] [Google Scholar]
- 27.Lopez-Velez R, Tarazona R, Garcia C A, Gomez-Mampaso E, Guerrero A, Moreira V, Villanueva R. Intestinal and extraintestinal cryptosporidiosis in AIDS patients. Eur J Clin Microbiol Infect Dis. 1995;14:677–681. doi: 10.1007/BF01690873. [DOI] [PubMed] [Google Scholar]
- 28.Marcial M A, Madara J L. Cryptosporidium: cellular localization, structural analysis of absorptive cell-parasite membrane-membrane interactions in guineapigs, and suggestion of protozoan transport by M cells. Gastroenterology. 1986;90:583–594. doi: 10.1016/0016-5085(86)91112-1. [DOI] [PubMed] [Google Scholar]
- 29.Molinari M, Carafoli E. Calpain: a cytosolic proteinase active at the membranes. J Membr Biol. 1997;156:1–8. doi: 10.1007/s002329900181. [DOI] [PubMed] [Google Scholar]
- 30.Nash P B, Purner M B, Leon R P, Clarke P, Duke R C, Curiel T J. Toxoplasma gondii-infected cells are resistant to multiple inducers of apoptosis. J Immunol. 1998;160:1824–1830. [PubMed] [Google Scholar]
- 31.Qiao L, Koutsos M, Tsai L L, Kozoni V, Guzman J, Shiff S J, Rigas B. Staurosporine inhibits the proliferation, alters the cell cycle distribution and induces apoptosis in HT-29 human colon adenocarcinoma cells. Cancer Lett. 1996;107:83–89. doi: 10.1016/0304-3835(96)04346-7. [DOI] [PubMed] [Google Scholar]
- 32.Reyland M E, Anderson S M, Matassa A A, Barzen K A, Quissell D O. Protein kinase C δ is essential for etoposide-induced apoptosis in salivary gland acinar cells. J Biol Chem. 1999;274:19115–19123. doi: 10.1074/jbc.274.27.19115. [DOI] [PubMed] [Google Scholar]
- 33.Seydel K B, Zhang T, Champion G A, Fichtenbaum C, Swanson P E, Tzipori S, Griffiths J K, Stanley S L., Jr Cryptosporidium parvum infection of human intestinal xenografts in SCID mice induces production of human tumor necrosis factor alpha and interleukin-8. Infect Immun. 1998;66:2379–2382. doi: 10.1128/iai.66.5.2379-2382.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Upton S J, Tilley M, Brillhart D B. Comparative development of Cryptosporidium parvum (Apicomplexa) in 11 continuous host cell lines. FEMS Microbiol Lett. 1994;118:233–236. doi: 10.1111/j.1574-6968.1994.tb06833.x. [DOI] [PubMed] [Google Scholar]
- 35.Vakil N B, Schwartz S M, Buggy B P, Brummitt C F, Kherellah M, Letzer D M, Gilson I H, Jones P G. Biliary cryptosporidiosis in HIV-infected people after the waterborne outbreak of cryptosporidiosis in Milwaukee. N Engl J Med. 1996;334:19–23. doi: 10.1056/NEJM199601043340104. [DOI] [PubMed] [Google Scholar]
- 36.van Antwerp D, Martin S J, Kafri T, Green D R, Verma I M. Suppression of TNF-α-induced apoptosis by NF-κB. Science. 1996;274:787–789. doi: 10.1126/science.274.5288.787. [DOI] [PubMed] [Google Scholar]
- 37.Wang C Y, Mayo M W, Baldwin A S J. TNF- and cancer therapy-induced apoptosis: potentiation by inhibition of NF-κB. Science. 1996;274:784–787. doi: 10.1126/science.274.5288.784. [DOI] [PubMed] [Google Scholar]