

Abstract
We sought to identify resting-state characteristics related to attention deficit/hyperactivity disorder, both as a categorical diagnosis and as a trait feature, using large-scale samples which were processed according to a standardized pipeline. In categorical analyses, we considered 1301 subjects with diagnosed ADHD, contrasted against 1301 unaffected controls (total N = 2602; 1710 males (65.72%); mean age = 10.86 years, sd = 2.05). Cases and controls were 1:1 nearest neighbor matched on in-scanner motion and key demographic variables and drawn from multiple large cohorts. Associations between ADHD-traits and resting-state connectivity were also assessed in a large multi-cohort sample (N = 10,113). ADHD diagnosis was associated with less anticorrelation between the default mode and salience/ventral attention (B = 0.009, t = 3.45, p-FDR = 0.004, d = 0.14, 95% CI = 0.004, 0.014), somatomotor (B = 0.008, t = 3.49, p-FDR = 0.004, d = 0.14, 95% CI = 0.004, 0.013), and dorsal attention networks (B = 0.01, t = 4.28, p-FDR < 0.001, d = 0.17, 95% CI = 0.006, 0.015). These results were robust to sensitivity analyses considering comorbid internalizing problems, externalizing problems and psychostimulant medication. Similar findings were observed when examining ADHD traits, with the largest effect size observed for connectivity between the default mode network and the dorsal attention network (B = 0.0006, t = 5.57, p-FDR < 0.001, partial-r = 0.06, 95% CI = 0.0004, 0.0008). We report significant ADHD-related differences in interactions between the default mode network and task-positive networks, in line with default mode interference models of ADHD. Effect sizes (Cohen’s d and partial-r, estimated from the mega-analytic models) were small, indicating subtle group differences. The overlap between the affected brain networks in the clinical and general population samples supports the notion of brain phenotypes operating along an ADHD continuum.
Subject terms: Attention, Diagnostic markers
Introduction
Attention deficit/hyperactivity disorder (ADHD) is a neurodevelopmental disorder characterized by age-inappropriate inattention, hyperactivity and impulsivity [1]. Its pathophysiology is unknown, although widely considered to involve alterations in the interplay of large-scale brain networks [2–4]. Such networks can be defined according to synchronized fluctuations in low frequency blood-oxygen-level-dependent signal examined in the absence of an external cognitive task [2, 5, 6]. The most prominent models in the literature implicate ADHD-related abnormalities in interactions between the default mode network and task-positive networks [2, 3, 7, 8]. The default mode network consists of interconnected midline regions including anterior/ventromedial prefrontal cortex, the posterior cingulate and precuneus, and inferolateral parietal, middle and medial temporal regions [5, 9]. This network shows attenuated activity during externally-oriented attention, is activated during rest, attentional lapses and spontaneous mind‐wandering, is putatively involved in self-referential and internally-oriented cognitions, and shows antagonistic relationships with task-positive networks implicated in externally-directed attention and cognition [5, 9–11]. Default mode network interference models of ADHD propose that greater positive connectivity and/or less negative connectivity between the default mode and task-positive networks interferes with the performance of the latter during tasks requiring executive control and effortful attention [2, 3, 7]. These network alterations are suggested to underlie the increased instances of attention lapses, behavioral/cognitive instability and mind-wandering observed in ADHD [2, 3, 7, 8].
This model arose initially from a series of studies that reported increased connectivity/decreased anti-correlation between the default mode network and task positive networks in individuals with ADHD, relative to unaffected controls [8, 12–16]. However, these findings were not always replicated [17–19]. Furthermore, a wide range of significant associations involving regions and networks outside of the default mode network have been reported, again with inconsistency (see Cortese and colleagues [20] for a systematic review). These discrepant findings are likely due in part to the typically small sample sizes, which can inflate type-I and type-II errors [20, 21].
To detect more consistent themes, several meta-analyses have attempted quantitative summaries of published findings, most of which have reported connectivity between candidate regions (‘seeds’) and the remaining brain (‘voxels’) [20, 22, 23]. However, these meta-analyses have also reached different conclusions. For example, one meta-analysis reported hyperconnectivity between the fronto-parietal network and the default mode network and between the fronto-parietal network and other task-positive networks in individuals with ADHD relative to unaffected controls [22], while a second reported abnormal connectivity within the default mode network [23]. A third meta-analysis reported no significant convergence in implicated brain regions between studies [20]. The discrepant findings likely reflect the different search strategies, different meta-analytic approaches, and the reliance on published seed-to-voxel resting-state data, which are not suited to retrospective meta-analyses due to the highly heterogenous region-of-interest definitions in the different papers [20]. Moreover, as these meta-analyses relied on the published literature, they were susceptible to publication bias and highly limited in their ability to assess potentially important subject-level confounds such as in-scanner motion, medication status and comorbid emotional and behavioral problems.
Here, we aimed to overcome the limitations of prior meta-analyses of published data by examining resting-state connectivity in patients with ADHD and unaffected controls through a mega-analysis of multiple large datasets which were processed using the same standardized preprocessing pipeline [24–27]. To explore diagnostic signals, we used data from the ADHD-200 [28], Healthy Brain Network (HBN) [29], Adolescent Brain Cognitive Development (ABCD) [30], and Neurobehavioral Clinical Research (NCR) [31] cohorts. All of these cohorts provide sufficient data to make a diagnosis of the presence of ADHD. The large size of our sample allows this investigation to closely match cases with unaffected controls on features that have all been shown to act as potential confounds in prior research, including age, gender, in-scanner motion and minutes of useable data, medication and comorbid emotional and behavioral problems [25, 32–35].
Furthermore, since research findings suggest that ADHD may exist at the extreme end of a continuum of ADHD traits in the general population [27, 36, 37], we also examined associations between resting-state connectivity and ADHD-traits assessed using the Child Behavior Checklist DSM-Oriented ADHD (CBCL-ADHD) subscale (N = 10,113) [38]. This analysis included data from the large National Consortium on Alcohol and Neurodevelopment in Adolescence (NCANDA) [39] and Human Connectome Project-Development (HCP-D) [40] cohorts. Based on previous theoretical models, we hypothesized that both the diagnosis of ADHD and ADHD-traits would be associated with alterations in default mode connectivity [2, 3, 7]. However, given the inconsistencies in the findings from the existing ADHD resting-state literature and meta-analyses, we also tested for potential ADHD-related alterations in network metrics outside of those involving the default mode network [20].
Materials and methods
Samples
We included data from the ADHD-200, HBN, ABCD, NCR, NCANDA and HCP-D datasets [28–31, 40–47]. Details on ADHD diagnostic methods for each study are given in the Supplement, along with narrative summaries of each study’s recruitment methods, protocols, assent/consent procedures and image acquisition parameters. All studies had IRB/ethical approval and acquired informed assent and/or consent using IRB approved procedures (see Supplement).
The ADHD-200, HBN, ABCD and NCR datasets included youths with ADHD diagnoses as well as unaffected controls. Each of the sites from the NCANDA dataset had only a small number of individuals with ADHD diagnoses, and diagnostic information was not available for HCP-D. We therefore included these datasets in the analyses examining associations with the CBCL-ADHD scale, but not in the case-control comparisons.
In the case-control comparison, 1:1 nearest neighbor matching without replacement was conducted using the MatchIt::matchit function in R [48]. Groups were matched on age, sex, in-scanner motion and site. Details for this matched sample are given in Table 1. Inclusion/exclusion criteria are given in the Supplement. In brief, subjects were aged ≥6 years and ≤19 with an IQ > 70. For the categorical analysis, both ADHD and non-ADHD subjects were excluded if they met criteria for possible psychosis or bipolar disorder. Further, to be eligible as an unaffected control, subjects had to have minimal ADHD symptoms and not be on psychostimulant medication. Other medications were allowed for both groups, including anti-depressants (ADHD group N = 29; matched controls = 17), anti-psychotics (ADHD group N = 12; matched controls = 2) and anti-convulsant medications (ADHD group N = 4; matched controls = 4). Emotional and behavioral problems were assessed using the CBCL and are reported for both groups in Table 1. In the analysis examining associations between the ADHD-CBCL scale and resting-state connectivity, we used N = 10,113 subjects from the HBN, ABCD, NCR, NCANDA and HCP-D datasets who provided useable resting-state data and the required covariate data.
Table 1.
Characteristics of N = 1301 subjects with ADHD and N = 1301 matched unaffected controls included in the case-control analysis, as well as N = 10,113 subjects included in the analyses of ADHD-traits.
| Non-ADHD | ADHD | CBCL sample | |||||
|---|---|---|---|---|---|---|---|
| Variable | Mean (sd) or count (%) | Mean (sd) or count (%) | Stat | p | sig | es | Mean (sd) or count (%) |
| Age, years | 10.86 (2.05) | 10.86 (2.05) | t = 0.00 | ~1 | d = 0.00 | 10.74 (1.89) | |
| Subtype | — | — | — | — | — | — | — |
| Combined Type | — | 370 (28.44%) | — | — | — | — | 392 (4.07%) |
| Hyperactive/Impulsive Type | — | 70 (5.38%) | — | — | — | — | 90 (0.93%) |
| Inattentive Type | — | 519 (39.89%) | — | — | — | — | 528 (5.49%) |
| ADHD-NOS | — | 103 (7.92%) | — | — | — | — | 116 (1.2%) |
| past-ADHD/ADHD in partial remission | — | 239 (18.37%) | — | — | — | — | 314 (3.26%) |
| Gender (M) | 855 (65.72%) | 855 (65.72%) | χ2 < 0.01 | ~1 | OR = 1.00 | 5045 (49.88%) | |
| Minutes of data | 12.73 (4.22) | 12.64 (4.29) | t = −0.54 | 0.59 | d = −0.02 | 15.37(4.64) | |
| Mean-RMS | 0.17 (0.05) | 0.17 (0.05) | t = −0.06 | 0.95 | d = −0.002 | 0.17 (0.05) | |
| CBCL-ADHD | 51.42 (2.9) | 61.03 (7.4) | t = 39.53 | <0.001 | *** | d = 1.71 | 53.18 (5.64) |
| CBCL-Internalizing | 47.84 (10.47) | 55.28 (10.66) | t = 16.39 | <0.001 | *** | d = 0.70 | 48.54 (10.72) |
| CBCL-CD | 51.99 (4.09) | 56.33 (7.77) | t = 16.19 | <0.001 | *** | d = 0.70 | 52.81 (5.36) |
| CBCL-ODD | 52.49 (4.23) | 57.57 (7.73) | t = 18.87 | <0.001 | *** | d = 0.81 | 53.38 (5.42) |
| Race | — | — | χ2 = 9.25 | 0.06 | V = 0.03 | ||
| Asian | 32 (2.89%) | 17 (1.57%) | — | — | — | — | 272 (2.74%) |
| Black/African American | 119 (10.75%) | 142 (13.12%) | — | — | — | — | 1248 (12.51%) |
| Mixed | 142 (12.83%) | 158 (14.6%) | — | — | — | — | 1213 (12.15%) |
| Other | 61 (5.51%) | 65 (6.01%) | — | — | — | — | 529 (5.3%) |
| White | 753 (68.02%) | 700 (64.7%) | — | — | — | — | 6716 (67.29%) |
| Ethnicity | — | — | χ2 = 1.57 | 0.21 | OR = 0.87 | ||
| Hispanic | 205 (18.65%) | 218 (20.9%) | — | — | — | 1955 (19.66%) | |
| Non-Hispanic/Latino | 894 (81.35%) | 825 (79.1%) | — | — | — | 7985 (80.33%) |
Note. For the case-control comparison there were N = 434 missing CBCL-ADHD, N = 434 missing CBCL-internalizing, N = 434 missing CBCL-CD, N = 434 missing CBCL-ODD, N = 413 missing race. N = 460 missing ethnicity. For the CBCL models, there were N = 494 missing diagnoses, N = 133 missing race, N = 173 missing ethnicity.
ADHD attention deficit/hyperactivity disorder, ADHD-NOS attention deficit/hyperactivity disorder not otherwise specified, CBCL child behavior checklist, CBCL-ADHD child behavior checklist attention deficit/hyperactivity disorder t-score, CBCL-Internalizing child behavior checklist internalizing t-score, CBCL-CD child behavior checklist conduct disorder t-score, CBCL-ODD oppositional defiant disorder t-score, RMS root‐mean‐square.
*p < 0.05, **p < 0.01, ***p < 0.001.
Resting-state connectivity
Details on MRI acquisition are provided in the Supplement. Preprocessing was performed using a well validated and standardized 36-parameter plus despiking pipeline [24], implemented in fMRIPrep and xcpEngine [24, 49]. Runs with mean-RMS ≤ 0.3 were concatenated using fslmerge to create a single time-series per subject [50, 51]. This mean-RMS threshold is stricter than the ones adopted in some previous studies of ADHD [52–55], but similar to thresholds used to remove runs with gross motion in recent developmental fMRI studies adopting the same 36-parameter plus despiking pipeline [24, 25]. We examined resting-state connectivity within and between resting-state networks using the Schaefer 400 ROI parcellation, which has been mapped to the 7 networks described by Yeo [6, 56]. These networks include the default mode network, task-positive networks including the dorsal attention network, frontoparietal network and salience/ventral attention network, primary motor and visual networks and a limbic network. Given that the limbic network consists of very ventral portions of orbitofrontal cortex and temporal lobe, which often suffer from signal loss and poor coverage, we excluded this network from our analyses. Time courses from all voxels within a 5-mm spherical radius around each of the ROIs were averaged and then correlated with each other, thereby creating an ROI-to-ROI connectivity matrix for each subject. These matrices were Fisher-z transformed. We then calculated within and between network connectivity for each of the networks. Within network connectivity for each network was defined as the average Fisher-z score for connections involving two regions from that network. Between network connectivity was calculated for each pairwise combination of networks (e.g., default mode network to dorsal attention network) by averaging all the Fisher-z scores of inter-network connections involving the relevant networks.
Modeling approach
The mega-analysis was performed using the lmerTest (version 3.1-3) software package [57] for R (version 4.0.2; http://www.r-project.org). Our mega-analytic approach was adapted from recent consortia-based mega-analyses including the Enhancing Neuro Imaging Genetics Through Meta-Analysis (ENIGMA) and REST-meta-MDD studies, and has been associated with increased statistical power relative to traditional meta-analytic methods while also allowing for the consideration of subject-level covariates [26, 27, 58, 59]. For the case-control comparison, the model included ADHD group (ADHD = 1 and non-ADHD = 0) as the predictor variable of interest. Although subjects were nearest-neighbor matched on age, sex, motion and site, we took a double-adjustment approach to control for potential residual associations between covariates and the network metrics by retaining covariates in the models [60]. Nested random intercepts were included for family ID and site and cohort [59]. This model was applied to each of the connectivity metrics, with multiple comparison correction performed using the Benjamini-Hochberg method [61]. A similar modeling approach examined CBCL-ADHD t-scores as the predictor variable of interest. Effect sizes were estimated based on the t-values from the linear mixed effects models. See Supplement for model syntax.
Sensitivity analyses
Comorbid psychiatric problems were most consistently assessed across studies using the parent-rated CBCL, available for all but the ADHD-200 cohorts [38]. We therefore used the internalizing broadband subscale from the CBCL to control for internalizing problems including anxiety and depression. This involved using scores on this scale as an additional variable in the matching algorithm for the case-control analysis, and as an additional covariate in the statistical models. To rule out the confounding influence of non-ADHD externalizing problems, in a sensitivity analysis we matched groups based on the CBCL subscales for conduct disorder and oppositional defiant disorder and included scores on these scales as extra covariates. As in-scanner motion has significant effects on estimates of resting-state connectivity [24], we repeated the primary analyses using a strict motion threshold of mean-RMS ≤ 0.15 which retained only subjects with no or only very minimal in-scanner motion (half the original threshold of mean-RMS ≤ 0.3). We re-ran case-control analyses while excluding subjects with past-ADHD (ADHD in full remission). Finally, we performed analyses excluding ADHD subjects taking psychostimulants, or for whom medication status was unclear. There are presently unresolved “known issues” with the ADHD diagnoses included in ABCD release 3.0 [62]. To maximize the sample size, and following other recent work [62], we initially applied a broad definition of ADHD to subjects in the ABCD dataset. Nonetheless, to confirm that effect sizes were not underestimated due to the potential inclusion of subjects with subclinical ADHD presentations, we also performed sensitivity analyses applying stricter ADHD diagnostic criteria for subjects in the ABCD study. The KSADS-COMP algorithm required that symptoms cause interference in at least one setting (e.g, home, social or school functioning), which is in line with current DSM-5 criteria. However, it did not assess the presence of symptoms in two or more contexts, as required by DSM-5 [1]. We therefore used item-level data from the KSADS-COMP to ensure that ADHD symptoms caused difficulties for the subject in at least two contexts (See Supplement). Note that since the items were intended to assess interference associated with symptoms rather than their mere presence, this is arguably a stricter requirement than current DSM-V criteria [1]. Sensitivity analyses were performed on network metrics that were significant in the primary models and examined whether models retained statistical significance (uncorrected p < 0.05) and similar effect sizes.
Results
Details on the matched sample of N = 1301 ADHD cases and N = 1301 unaffected controls are given in Table 1. Following correction for multiple comparisons, mixed effects mega-analytic models indicated significant group differences in resting-state connectivity between the default mode network and the dorsal attention network (B = 0.01, t = 4.28, p-FDR < 0.001, d = 0.17, 95% CI = 0.006, 0.015), between the default mode network and the salience/ventral attention network (B = 0.009, t = 3.45, p-FDR = 0.004, d = 0.14, 95% CI = 0.004, 0.014), and between the default mode network and somatomotor network (B = 0.008, t = 3.49, p-FDR = 0.004, d = 0.14, 95% CI = 0.004, 0.013). These networks showed a stronger anti-correlation in unaffected controls than in ADHD subjects. A relationship with increased positive connectivity between the default mode network and the other major task positive attention network, the frontoparietal network, was observed at p < 0.05 uncorrected threshold but did not survive FDR correction. Mean connectivity values are presented in Supplementary Table 1. See Table 2 and Fig. 1 & 2. Results are summarized for all models, including non-significant ones, in Supplementary Table 2.
Table 2.
Significant associations (p < 0.05) between resting-state connectivity and ADHD diagnosis (N = 2602) and between resting-state connectivity and ADHD-traits (N = 10,113).
| Network 1 | Network 2 | Β | SE | t | 95% CI | FDR p | ES |
|---|---|---|---|---|---|---|---|
| Significant case-control differences (N = 2602) | |||||||
| Default mode | Dorsal attention | 0.01 | 0.002 | 4.28 | 0.006, 0.015 | <0.001*** | d = 0.17 |
| Default mode | Salience/ventral attention | 0.009 | 0.003 | 3.45 | 0.004, 0.014 | 0.004** | d = 0.14 |
| Default mode | Somatomotor | 0.008 | 0.002 | 3.49 | 0.004, 0.013 | 0.004** | d = 0.14 |
| Significant associations with scores on the CBCL-ADHD scale (N = 10,113) | |||||||
| Default mode | Dorsal attention | 0.0006 | 0.0001 | 5.57 | 0.0004, 0.0008 | <0.001*** | r = 0.06 |
| Default mode | Frontoparietal | 0.0004 | 0.00009 | 4.31 | 0.0002, 0.0005 | <0.001*** | r = 0.04 |
| Dorsal attention | Dorsal attention | −0.0005 | 0.0001 | −4.02 | −0.0007, −0.0002 | <0.001*** | r =−0.04 |
| Default mode | Salience/ventral attention | 0.0005 | 0.0001 | 4.06 | 0.0002, 0.0007 | <0.001*** | r = 0.04 |
| Salience/ventral attention | Somatomotor | 0.0004 | 0.0001 | 3.56 | 0.0002, 0.0006 | 0.002** | r = 0.04 |
| Default mode | Somatomotor | 0.0003 | 0.0001 | 2.58 | 0.00007, 0.0005 | 0.035* | r = 0.03 |
CBCL-ADHD child behavior checklist ADHD t-score; FDR false discovery rate.
*p < 0.05, **p < 0.01, ***p < 0.001.
Fig. 1. Forest plots for brain network metrics that differed significantly (corrected p < 0.05) between N = 1301 ADHD cases and N = 1301 non-ADHD controls.

The top line shows the effect size (Cohen’s d) and corresponding 95% confidence intervals for the main mega-analytic model. The subsequent rows show the results of models performed within each cohort (NCR N = 122; HBN = 575; ADHD200-Peking N = 199; ADHD200-NYU N = 182; ABCD N = 1524). Panel (A) shows the results for connectivity between the default mode network and the dorsal attention network. Panel (B) shows the results for connectivity between the default mode network and the salience/ventral attention network. Panel (C) shows the results for connectivity between the default mode network and somatomotor network.
Fig. 2. Associations between ADHD diagnosis and connectivity within and between brain networks.
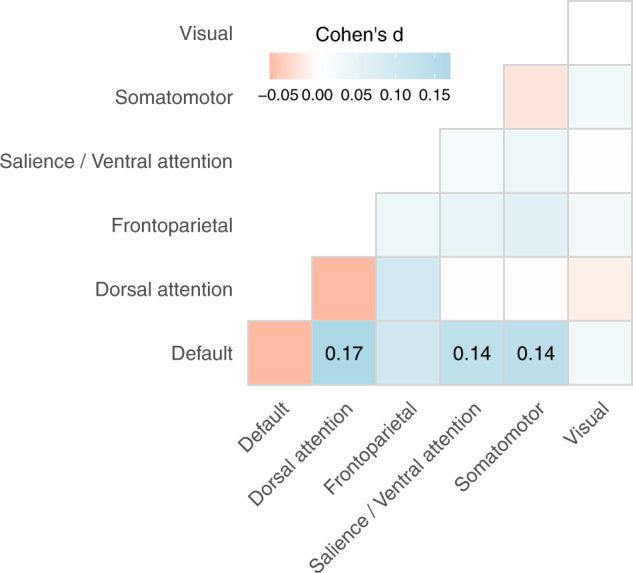
Red indicates a negative association. Blue indicates a positive association. Darker shades represent stronger associations. Effect sizes from the linear mixed models are presented for network connections significant at p < 0.05 after correction for multiple comparisons.
Effects of ADHD traits on functional connectivity
We next considered ADHD as a trait, incorporating data from additional cohorts, to give a sample size of 10,113 (Table 1). ADHD traits were associated with decreased anti-correlation between the default mode network and the dorsal attention network (B = 0.0006, t = 5.57, p-FDR < 0.001, partial r = 0.06, 95% CI = 0.0004, 0.0008), between the default mode network and the salience/ventral attention network (B = 0.0005, t = 4.06, p-FDR < 0.001, partial r = 0.04, 95% CI = 0.0002, 0.0007) and more positive connectivity between the default mode network and the frontoparietal network (B = 0.0004, t = 4.31, p-FDR = <0.001, partial r = 0.04, 95% CI = 0.0002, 0.0005). Additional significant associations are listed in Table 2. See Fig. 3 and Supplementary Table 3 and Supplementary Figs. 1 and 2.
Fig. 3. Forest plots for significant associations (corrected p < 0.05) between ADHD-traits and resting-state connectivity (N = 10,113).

The top line shows the effect size (partial r) and corresponding 95% confidence intervals for the main mega-analytic model. The subsequent rows show the results of models performed within each cohort (NCR N = 100; NCANDA N = 504; HCP-D N = 494; HBN N = 692; ABCD N = 8323). Panel (A) shows the results for connectivity between the default mode network and the dorsal attention network. Panel (B) Shows the results for connectivity between the default mode network and the frontoparietal network. Panel (C) Shows the results for connectivity within the dorsal attention network. Panel (D) Shows the results for connectivity between the default mode network and the salience/ventral attention network. Panel (E) Shows the results for connectivity between the salience/ventral attention network and the somatomotor network. Panel (F) Shows the results for connectivity between the default mode network and the somatomotor network.
Sensitivity analyses
We used the internalizing broadband subscale from the CBCL to control for internalizing problems including anxiety and depression, which have been reported to be elevated in individuals with ADHD and associated with alterations in resting-state connectivity [58, 63]. The case-control differences remained significant. To rule out the confounding influence of commonly comorbid non-ADHD externalizing problems, we matched groups based on the CBCL subscales for conduct disorder and oppositional defiant disorder and included scores on these scales as extra covariates in our mega-analytic models, with case-control differences remaining significant.
As in-scanner motion has significant effects on estimates of resting-state connectivity [24], we repeated the primary analyses using a strict motion threshold of mean-RMS ≤ 0.15 (half the original threshold) which retained only subjects with no or only very minimal in-scanner motion, with the results remaining significant. The case-control findings were robust to including only psychostimulant-free subjects, which is important in light of work showing a modulatory effect of psychostimulants on default mode connectivity [64], and to excluding subjects with past-ADHD. The findings held when considering a stricter definition of ADHD in the ABCD cohort. Associations with CBCL scores were also typically robust in these sensitivity analyses. See Supplementary Tables 4 and 5 for details.
Discussion
In this study, we applied meta-analytic methods to examine resting-state connectivity metrics associated with ADHD diagnosis and ADHD-traits. Using both approaches, we found that ADHD was associated with relatively more positive connectivity/decreased negative connectivity between the default mode network and task-positive networks. Significant associations had consistently small effect sizes (ranging from d = 0.14 to d = 0.17 for case-control comparisons, and partial-r = 0.03 to partial-r = 0.06 for relationships with CBCL-ADHD). These findings were robust to sensitivity analyses which considered in-scanner motion, medication exposure and comorbid internalizing and externalizing problems.
We report that ADHD was associated with stronger positive connectivity/weaker anticorrelations between the default mode and task-positive networks. The findings of more positive/decreased negative connectivity between the default mode network and dorsal and ventral attention networks is consistent with numerous previous reports on individual cohorts [8, 12–16]. The dorsal attention network is comprised of bilateral intraparietal sulcus and the frontal eye fields and enables top-down control over spatial attention [6, 65, 66]. The salience/ventral attention network, on the other hand, consists of portions of bilateral inferior frontal gyrus/anterior insula, temporo-parietal junction and posterior medial frontal cortex, and is commonly engaged across tasks requiring externally-focused attention [67]. Network segregation between the default mode and task-positive networks is argued to act as a buffer against the disruption of attentional processes by internally generated cognition [68], and has been associated with better performance on attention and executive function tasks [8, 69, 70], while positive connectivity or co-activation of these networks has been associated with attention lapses and mind-wandering [10, 69, 71, 72]. Attention deficits are one of the key domains of impairment in ADHD, and previous work using dorsal attention and salience/ventral attention network-dependent attention paradigms has reported decreased default mode network deactivation in individuals with ADHD relative to controls, and impaired performance [35, 65]. Our results are consistent with dominant models of network abnormalities associated with the disorder, which propose an important role for decreased default mode segregation in the cognitive and behavioral impairments which characterize ADHD [2, 3, 7, 73].
The network findings were highly similar whether performing a case-control comparison or examining ADHD as a continuous trait. This is analogous to the findings of the ENIGMA-ADHD Working Group, which reported that ADHD diagnosis and traits were associated with similar patterns of smaller cortical surface area [27], and provides further support for associations between ADHD traits and brain phenotypes operating along a continuum and independently of clinical diagnosis [27, 37]. These results are also compatible with a large genetic literature, with numerous twin studies and studies of common risk variants showing that the same genetic risks contribute to both diagnosable ADHD and dimensional ADHD traits in the population [36, 74].
We advance the literature by applying mega-analysis to ‘raw’ data, rather than integrating published data through meta-analyses, as the latter is biased by reported signals that attained thresholds for declaring significance within each study, and cannot account for important subject-level confounders. By using ‘raw’ data from each cohort, we could use a standardized preprocessing pipeline and uniform network parcellation. Further our use of ‘raw’ data allowed us to focus on diagnostic signals that hold when cases and controls were matched on critical confounders. For instance, cases and controls were 1:1 matched on in-scanner motion, and slightly larger effect sizes were observed when applying an exceptionally strict motion threshold. This is important considering the known effects of in-scanner motion on estimates of resting-state connectivity and the commonly observed correlations between ADHD diagnosis and in-scanner motion [75]. Relatedly, as in previous work, ADHD was associated with elevated internalizing and externalizing problems [63]. However, our results were robust after further matching subjects with available CBCL data on these symptom domains, with modest increases in effect sizes. Finally, using a subgroup of unmedicated subjects we were able to rule out the confounding influence of psychostimulant medication on case-control differences.
The effect sizes were small but in a similar range to those reported previously by the ENIGMA-ADHD mega-analyses of alterations in structural brain phenotypes [26, 27, 59], as well as recent mega-analyses and “big data” brain-wise association studies of multiple psychiatric disorders and related behavioral traits [58, 76–79]. Our findings support the notion that although subtle, between-group differences in default mode connectivity are detectable using large neuroimaging samples, atypical connectivity is neither necessary nor sufficient for the development of ADHD [20]. Our mega-analytic effect sizes are smaller than those observed in previous, smaller studies of ADHD [12, 19, 20]. This is expected, as findings surviving significance in underpowered studies, even if they are true positives, are likely to be associated with inflated effect sizes [21, 30, 76]. However, we only considered network-level metrics in the present work, as they were deemed best suited to testing default mode interference models of the disorder. Future studies may report larger effect sizes for alternative resting-state metrics relevant to ADHD, such as those assessed at the network-edge or voxel-level. Furthermore, we combined data from multiple datasets with differing imaging protocols, recruitment procedures and diagnostic tools. Studies of more homogenous samples may be associated with larger effect sizes, with the trade-off that the results may be less generalizable. Moreover, recent work has suggested the utility of aggregating disorder-related brain phenotypes into single cumulative value estimates, which are analogous to polygenic risk scores for common genetic variant risk [80, 81]. Relatedly, normative modeling methods aggregate the overall number and size of deviations from developmentally typical templates of brain phenotypes regardless of sign, network or spatial location, with initial results based on structural brain phenotypes pointing towards ADHD being best characterized as a disorder of broadly atypical brain development, rather than one of any particular set of brain regions or networks [82]. In addition, several important studies have shown promise in using multivariate predictive modelling methods, including machine learning algorithms, to aggregate distributed patterns from across the entire brain associated with the cognitive, clinical or demographic phenotype of interest, and to test how well such multivariate patterns predict these phenotypes in unseen data [76, 83–86]. Considering the small effect sizes at the level of individual brain phenotypes, these methods based on cumulative risk scores and/or multivariate prediction methods may offer better predictive power for imaging-based classification of ADHD [76, 80, 86]. Finally, longitudinal work using structural MRI has suggested ADHD to be a disorder of abnormal neurodevelopmental trajectories [37, 87]. Future work is needed in large samples to examine whether detected effect sizes are larger for associations with dynamic properties of functional brain maturation compared with static estimates of resting-state connectivity [37, 53].
Further limitations must also be kept in mind. First, our cross-sectional case-control design cannot speak to the direction of any potential causal relationships between ADHD and default mode connectivity, and work combining genomics, heritability and longitudinal methods is required to delineate any causal relationships. Second, given the large sample size we were only able to process the data using a single pipeline. Future work should consider the effects of preprocessing pipelines by adopting a multiverse approach [88]. Third, while applying motion thresholds and controlling for mean-RMS was essential for reducing motion-related noise and potential type-I errors resulting from group differences in in-scanner motion, since ADHD diagnosis and in-scanner motion have been reported to be correlated and to share common genetic influences, these steps may have reduced our statistical power to detect true between-group differences [89]. Fourth, while we controlled carefully for psychiatric comorbidities such as internalizing and externalizing problems, which were well characterized across most included datasets using the available CBCL scales, common neurodevelopmental comorbidities such as autism spectrum disorder have also been associated with hyperconnectivity between the default mode and task-positive networks. It is therefore possible that the hyperconnectivity reported here may reflect a feature of neurodevelopmental problems defined more broadly [14, 77].
In summary, we conducted the largest assessment to date of resting-state connectivity alterations associated with ADHD diagnosis and ADHD traits in the general population. The results provided converging evidence for an association between ADHD and connectivity between the default mode network and task positive networks, which were in line with existing models of the disorder and robust in numerous sensitivity analyses. Nonetheless, the small effect sizes suggest the resting-state abnormalities may at best form only a small part of the overall pathophysiology of ADHD, and future work is required to confirm the causal associations between brain functioning and ADHD symptoms often assumed in current models in the literature.
Supplementary information
Acknowledgements
Data used in the preparation of this article were obtained from the Adolescent Brain Cognitive Development (ABCD) Study (https://abcdstudy.org), held in the NIMH Data Archive (NDA). This is a multisite, longitudinal study designed to recruit more than 10,000 children aged 9 to 10 years and follow them over 10 years into early adulthood. The ABCD Study is supported by the National Institutes of Health and additional federal partners under award numbers U01DA041048, U01DA050989, U01DA051016, U01DA041022, U01DA051018, U01DA051037, U01DA050987, U01DA041174, U01DA041106, U01DA041117, U01DA041028, U01DA041134, U01DA050988, U01DA051039, U01DA041156, U01DA041025, U01DA041120, U01DA051038, U01DA041148, U01DA041093, U01DA041089, U24DA041123, and U24DA041147. A full list of supporters is available at https://abcdstudy.org/ federal-partners.html. A listing of participating sites and a complete listing of the study investigators can be found at https://abcdstudy.org/consortium_ members/. ABCD consortium investigators designed and implemented the study and/or provided data but did not necessarily participate in analysis or writing of this report. The ABCD data repository grows and changes over time. The ABCD data used in this report came from DOI: 10.15154/1519007. The DOIs can be found at 10.15154/1519007 and from the fast-track data release (raw neuroimaging data). The raw data are available at https://nda.nih.gov/edit_collection.html?id=2573. Instructions on how to create an NDA study are available at https://nda.nih.gov/training/modules/study.html). Data and/or research tools used in the preparation of this manuscript were obtained from the National Institute of Mental Health (NIMH) Data Archive (NDA). NDA is a collaborative informatics system created by the National Institutes of Health to provide a national resource to support and accelerate research in mental health. Dataset identifier (s): #2573, #2846 and #3165. This manuscript reflects the views of the authors and may not reflect the opinions or views of the NIH or of the Submitters submitting original data to NDA (10.15154/1527788). The Human Connectome Project-Development was supported by the NIMH under Award Number U01MH109589. This manuscript included data from a limited access dataset obtained from the Child Mind Institute Biobank, the HBN study (http://www.healthybrainnetwork.org). Data collection as for the ADHD200-NYU cohort was funded by the NIMH (R01MH083246), Autism Speaks, The Stavros Niarchos Foundation, The Leon Levy Foundation, and an endowment provided by Phyllis Green and Randolph Cōwen. Data collection for the ADHD200-Peking cohort was supported by The Commonwealth Sciences Foundation, Ministry of Health, China (200802073), The National Foundation, Ministry of Science and Technology, China (2007BAI17B03), The National Natural Sciences Foundation, China (30970802), The Funds for International Cooperation of the National Natural Science Foundation of China (81020108022), The National Natural Science Foundation of China (8100059), Open Research Fund of the State Key Laboratory of Cognitive Neuroscience and Learning. Data collection and sharing for this project was provided by the National Consortium on Alcohol and Neurodevelopment in Adolescence (NCANDA), which is funded by the U.S. National Institute on Alcohol Abuse and Alcoholism with co-funding from the National Institute on Drug Abuse, the National Institute of Mental Health, the National Institute of Child Health and Human Development, and the National Institute of Health Office of the Director (AA021695 (MPI: SA Brown, SF Tapert), AA021697 (MPI: A Pfefferbaum, KM Pohl), AA021692 (PI: SF Tapert), AA021681 (PI: MD DeBellis), AA021690 (PI: DB Clark), AA021691 (PI: B Nagel), AA021696 (MPI: IM Colrain, FC Baker), AA021697-04S1 (PI: KM Pohl)). NCANDA data are disseminated by the Center for Health Sciences, SRI International. We used data from NCANDA_PUBLIC_4Y_REDCAP_V01 (10.7303/syn22216455), NCANDA_PUBLIC_BASE_RESTINGSTATE_V01 (10.7303/syn11605291) and NCANDA_PUBLIC_BASE_STRUCTURAL_V01 (10.7303/syn11541569). This study reflects the views of the authors and may not reflect the opinions or views of other individuals or institutions including the NIH, the ABCD, NCANDA, HCP or ADHD200 consortium investigators, the Child Mind Institute, SRI International, or other funding agencies. Image processing was conducted using the high-performance computing capabilities of the NIH Biowulf cluster. The authors thank the NIMH Data Science and Sharing Team for help with accessing and processing the ABCD-BIDS dataset.
Author contributions
Study concept and design: LJN. Acquisition, analysis, or interpretation of data: all authors. Drafting of the manuscript: LJN, PS. Critical revision of the manuscript for important intellectual content: All authors. Statistical analysis: LJN, GS. Administrative, technical, or material support: LJN, GS, PS. Study supervision: PS. Obtained funding: PS.
Funding
Funded by the intramural research program of the National Institute of Mental Health and the National Human Genome Research Institute (ZIAHG200378 to PS). This funding supported data collection for the NCR cohort (ClinicalTrials.gov identifier: NCT01721720).
Competing interests
The authors declare no competing interests.
Footnotes
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
These authors contributed equally: Luke J. Norman, Gustavo Sudre.
Supplementary information
The online version contains supplementary material available at 10.1038/s41386-022-01408-z.
References
- 1.Association AP. Diagnostic and statistical manual of mental disorders (DSM-5®). American Psychiatric Pub; 2013. [DOI] [PubMed]
- 2.Bozhilova NS, Michelini G, Kuntsi J, Asherson P. Mind wandering perspective on attention-deficit/hyperactivity disorder. Neurosci Biobehav Rev. 2018;92:464–76. doi: 10.1016/j.neubiorev.2018.07.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Sonuga-Barke EJS, Castellanos FX. Spontaneous attentional fluctuations in impaired states and pathological conditions: a neurobiological hypothesis. Neurosci Biobehav Rev. 2007;31:977–86. doi: 10.1016/j.neubiorev.2007.02.005. [DOI] [PubMed] [Google Scholar]
- 4.Castellanos FX, Proal E. Large-scale brain systems in ADHD: beyond the prefrontal–striatal model. Trends Cogn Sci. 2012;16:17–26. doi: 10.1016/j.tics.2011.11.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Raichle ME. The brain’s default mode network. Annu Rev Neurosci. 2015;38:433–47. doi: 10.1146/annurev-neuro-071013-014030. [DOI] [PubMed] [Google Scholar]
- 6.Thomas Yeo BT, Krienen FM, Sepulcre J, Sabuncu MR, Lashkari D, Hollinshead M, et al. The organization of the human cerebral cortex estimated by intrinsic functional connectivity. J Neurophysiol. 2011;106:1125–65. doi: 10.1152/jn.00338.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Castellanos FX, Kelly C, Milham MP. The restless brain: attention-deficit hyperactivity disorder, resting—state functional connectivity, and intrasubject variability. Can J Psychiatry. 2009;54:665–72. doi: 10.1177/070674370905401003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Mills BD, Miranda-Dominguez O, Mills KL, Earl E, Cordova M, Painter J, et al. ADHD and attentional control: Impaired segregation of task positive and task negative brain networks. Network. Neuroscience. 2018;2:200–17. doi: 10.1162/netn_a_00034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Andrews-Hanna JR, Reidler JS, Sepulcre J, Poulin R, Buckner RL. Functional-anatomic fractionation of the brain’s default network. Neuron. 2010;65:550–62. doi: 10.1016/j.neuron.2010.02.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Weissman DH, Roberts KC, Visscher KM, Woldorff MG. The neural bases of momentary lapses in attention. Nat Neurosci. 2006;9:971–8. doi: 10.1038/nn1727. [DOI] [PubMed] [Google Scholar]
- 11.Christoff K, Gordon AM, Smallwood J, Smith R, Schooler JW. Experience sampling during fMRI reveals default network and executive system contributions to mind wandering. Proc Natl Acad Sci USA. 2009;106:8719–24. doi: 10.1073/pnas.0900234106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Sudre G, Szekely E, Sharp W, Kasparek S, Shaw P. Multimodal mapping of the brain’s functional connectivity and the adult outcome of attention deficit hyperactivity disorder. Proc Natl Acad Sci. 2017;114:11787–92. doi: 10.1073/pnas.1705229114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Sripada C, Kessler D, Fang Y, Welsh RC, Prem Kumar K, Angstadt M. Disrupted network architecture of the resting brain in attention‐deficit/hyperactivity disorder. Hum Brain Mapp. 2014;35:4693–705. doi: 10.1002/hbm.22504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Kernbach JM, Satterthwaite TD, Bassett DS, Smallwood J, Margulies D, Krall S, et al. Shared endo-phenotypes of default mode dysfunction in attention deficit/hyperactivity disorder and autism spectrum disorder. Transl Psychiatry. 2018;8:1–11. doi: 10.1038/s41398-018-0179-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Sun L, Cao Q, Long X, Sui M, Cao X, Zhu C, et al. Abnormal functional connectivity between the anterior cingulate and the default mode network in drug-naïve boys with attention deficit hyperactivity disorder. Psychiatry Res: Neuroimaging. 2012;201:120–7. doi: 10.1016/j.pscychresns.2011.07.001. [DOI] [PubMed] [Google Scholar]
- 16.Lin H-Y, Kessler D, Tseng W-YI, Gau SS-F. Increased functional segregation related to the salience network in unaffected siblings of youths with attention-deficit/hyperactivity disorder. J Am Acad Child Adolesc Psychiatry. 2021;60:152–65. doi: 10.1016/j.jaac.2019.11.012. [DOI] [PubMed] [Google Scholar]
- 17.Mostert JC, Shumskaya E, Mennes M, Onnink AMH, Hoogman M, Kan CC, et al. Characterising resting-state functional connectivity in a large sample of adults with ADHD. Prog Neuro-Psychopharmacol Biol Psychiatry. 2016;67:82–91. doi: 10.1016/j.pnpbp.2016.01.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Jung M, Tu Y, Park J, Jorgenson K, Lang C, Song W, et al. Surface-based shared and distinct resting functional connectivity in attention-deficit hyperactivity disorder and autism spectrum disorder. Br J Psychiatry. 2019;214:339–44. doi: 10.1192/bjp.2018.248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Sidlauskaite J, Sonuga-Barke E, Roeyers H, Wiersema JR. Altered intrinsic organisation of brain networks implicated in attentional processes in adult attention-deficit/hyperactivity disorder: a resting-state study of attention, default mode and salience network connectivity. Eur Arch Psychiatry Clin Neurosci. 2016;266:349–57. doi: 10.1007/s00406-015-0630-0. [DOI] [PubMed] [Google Scholar]
- 20.Cortese S, Aoki YY, Itahashi T, Castellanos FX, Eickhoff SB. Systematic review and meta-analysis: resting state functional magnetic resonance imaging studies of attention-deficit/hyperactivity disorder. J Am Acad Child Adolescent Psych. 2021;60:61–75. [DOI] [PubMed]
- 21.Button KS, Ioannidis JP, Mokrysz C, Nosek BA, Flint J, Robinson ES, et al. Power failure: why small sample size undermines the reliability of neuroscience. Nat Rev Neurosci. 2013;14:365–76. doi: 10.1038/nrn3475. [DOI] [PubMed] [Google Scholar]
- 22.Gao Y, Shuai D, Bu X, Hu X, Tang S, Zhang L, et al. Impairments of large-scale functional networks in attention-deficit/hyperactivity disorder: a meta-analysis of resting-state functional connectivity. Psychological Med. 2019;49:2475–85. doi: 10.1017/S003329171900237X. [DOI] [PubMed] [Google Scholar]
- 23.Sutcubasi B, Metin B, Kurban MK, Metin ZE, Beser B, Sonuga-Barke E. Resting-state network dysconnectivity in ADHD: A system-neuroscience-based meta-analysis. The. World J Biol Psychiatry. 2020;21:662–72. doi: 10.1080/15622975.2020.1775889. [DOI] [PubMed] [Google Scholar]
- 24.Ciric R, Rosen AF, Erus G, Cieslak M, Adebimpe A, Cook PA, et al. Mitigating head motion artifact in functional connectivity MRI. Nat Protoc. 2018;13:2801–26. doi: 10.1038/s41596-018-0065-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Ciric R, Wolf DH, Power JD, Roalf DR, Baum GL, Ruparel K, et al. Benchmarking of participant-level confound regression strategies for the control of motion artifact in studies of functional connectivity. Neuroimage. 2017;154:174–87. doi: 10.1016/j.neuroimage.2017.03.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Hoogman M, Bralten J, Hibar DP, Mennes M, Zwiers MP, Schweren L, et al. Subcortical brain volume differences of participants with ADHD across the lifespan: an ENIGMA collaboration. Lancet Psychiatry. 2017;4:310. doi: 10.1016/S2215-0366(17)30049-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Hoogman M, Muetzel R, Guimaraes JP, Shumskaya E, Mennes M, Zwiers MP, et al. Brain imaging of the cortex in ADHD: a coordinated analysis of large-scale clinical and population-based samples. Am J Psychiatry. 2019;176:531–42. doi: 10.1176/appi.ajp.2019.18091033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Milham MP, Fair D, Mennes M, Mostofsky SH. The ADHD-200 consortium: a model to advance the translational potential of neuroimaging in clinical neuroscience. Frontiers in Systems. Neuroscience. 2012;6:62. doi: 10.3389/fnsys.2012.00062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Alexander LM, Escalera J, Ai L, Andreotti C, Febre K, Mangone A, et al. An open resource for transdiagnostic research in pediatric mental health and learning disorders. Sci Data. 2017;4:1–26. doi: 10.1038/sdata.2017.181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Casey BJ, Cannonier T, Conley MI, Cohen AO, Barch DM, Heitzeg MM, et al. The adolescent brain cognitive development (ABCD) study: imaging acquisition across 21 sites. Developmental Cogn Neurosci. 2018;32:43–54. doi: 10.1016/j.dcn.2018.03.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Sudre G, Sharp W, Kundzicz P, Bouyssi-Kobar M, Norman L, Choudhury S, et al. Predicting the course of ADHD symptoms through the integration of childhood genomic, neural, and cognitive features. Mol Psychiatry. 2021;26:4046–54. doi: 10.1038/s41380-020-00941-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Power JD, Barnes KA, Snyder AZ, Schlaggar BL, Petersen SE. Spurious but systematic correlations in functional connectivity MRI networks arise from subject motion. Neuroimage. 2012;59:2142–54. doi: 10.1016/j.neuroimage.2011.10.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Satterthwaite TD, Wolf DH, Roalf DR, Ruparel K, Erus G, Vandekar S, et al. Linked sex differences in cognition and functional connectivity in youth. Cereb Cortex. 2015;25:2383–94. doi: 10.1093/cercor/bhu036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Chabernaud C, Mennes M, Kelly C, Nooner K, Di Martino A, Castellanos FX, et al. Dimensional brain-behavior relationships in children with attention-deficit/hyperactivity disorder. Biol Psychiatry. 2012;71:434–42. doi: 10.1016/j.biopsych.2011.08.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Liddle EB, Hollis C, Batty MJ, Groom MJ, Totman JJ, Liotti M, et al. Task‐related default mode network modulation and inhibitory control in ADHD: Effects of motivation and methylphenidate. J Child Psychol Psychiatry. 2011;52:761–71. doi: 10.1111/j.1469-7610.2010.02333.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Thapar A. Discoveries on the genetics of ADHD in the 21st century: new findings and their implications. Am J Psychiatry. 2018;175:943–50. doi: 10.1176/appi.ajp.2018.18040383. [DOI] [PubMed] [Google Scholar]
- 37.Shaw P, Gilliam M, Liverpool M, Weddle C, Malek M, Sharp W, et al. Cortical development in typically developing children with symptoms of hyperactivity and impulsivity: support for a dimensional view of attention deficit hyperactivity disorder. Am J Psychiatry. 2011;168:143–51. doi: 10.1176/appi.ajp.2010.10030385. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.LA, ATR. Manual for the ASEBA School-Age Forms & Profiles. Burlington, VT: University of Vermont. Research Center for Children, Youth, & Families. 2001.
- 39.Brown SA, Brumback T, Tomlinson K, Cummins K, Thompson WK, Nagel BJ, et al. The National Consortium on Alcohol and NeuroDevelopment in Adolescence (NCANDA): a multisite study of adolescent development and substance use. J Stud Alcohol Drugs. 2015;76:895–908. doi: 10.15288/jsad.2015.76.895. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Somerville LH, Bookheimer SY, Buckner RL, Burgess GC, Curtiss SW, Dapretto M, et al. The Lifespan Human Connectome Project in Development: A large-scale study of brain connectivity development in 5–21 year olds. Neuroimage. 2018;183:456–68. doi: 10.1016/j.neuroimage.2018.08.050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Pfefferbaum A, Rohlfing T, Pohl KM, Lane B, Chu W, Kwon D, et al. Adolescent development of cortical and white matter structure in the NCANDA sample: role of sex, ethnicity, puberty, and alcohol drinking. Cereb Cortex. 2016;26:4101–21. doi: 10.1093/cercor/bhv205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Müller-Oehring EM, Kwon D, Nagel BJ, Sullivan EV, Chu W, Rohlfing T, et al. Influences of age, sex, and moderate alcohol drinking on the intrinsic functional architecture of adolescent brains. Cereb Cortex. 2018;28:1049–63. doi: 10.1093/cercor/bhx014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Zhao Q, Sullivan EV, Honnorat N, Adeli E, Podhajsky S, De Bellis MD, et al. Association of heavy drinking with deviant fiber tract development in frontal brain systems in adolescents. JAMA Psychiatry. 2021;78:407–15. doi: 10.1001/jamapsychiatry.2020.4064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Pohl KM, Sullivan EV, Pfefferbaum A. The NCANDA_PUBLIC_BASE_RESTINGSTATE_V01 Data Release of the National Consortium on Alcohol and NeuroDevelopment in Adolescence (NCANDA). Sage Bionetworks Synapse. 2017;2017.
- 45.Pohl KM, Sullivan EV, Pfefferbaum A. The NCANDA_PUBLIC_4Y_REDCAP_V01 Data Release of the National Consortium on Alcohol and NeuroDevelopment in Adolescence (NCANDA). Sage Bionetworks Synapse. 2021;2021.
- 46.Pohl KM, Sullivan EV, Pfefferbaum A. The NCANDA_PUBLIC_BASE_STRUCTURAL_V01 Data Release of the National Consortium on Alcohol and NeuroDevelopment in Adolescence (NCANDA). Sage Bionetworks Synapse. 2017;2017.
- 47.Sudre G, Bouyssi-Kobar M, Norman L, Sharp W, Choudhury S, Shaw P. Estimating the heritability of developmental change in neural connectivity, and its association with changing symptoms of attention-deficit/hyperactivity disorder. Biol Psychiatry. 2021;89:443–50. doi: 10.1016/j.biopsych.2020.06.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Ho DE, Imai K, King G, Stuart EA. MatchIt: Nonparametric Preprocessing for Parametric Causal Inference. J Stat Softw. 2011;42:1–28.
- 49.Esteban O, Markiewicz CJ, Blair RW, Moodie CA, Isik AI, Erramuzpe A, et al. fMRIPrep: a robust preprocessing pipeline for functional MRI. Nat Methods. 2019;16:111–6. doi: 10.1038/s41592-018-0235-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Jenkinson M, Beckmann CF, Behrens TE, Woolrich MW, Smith SM. FSL. Neuroimage. 2012;62:782–90. doi: 10.1016/j.neuroimage.2011.09.015. [DOI] [PubMed] [Google Scholar]
- 51.Cho JW, Korchmaros A, Vogelstein JT, Milham MP, Xu T. Impact of concatenating fMRI data on reliability for functional connectomics. Neuroimage. 2021;226:117549. doi: 10.1016/j.neuroimage.2020.117549. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Pruim RH, Beckmann CF, Oldehinkel M, Oosterlaan J, Heslenfeld D, Hartman CA, et al. An integrated analysis of neural network correlates of categorical and dimensional models of attention-deficit/hyperactivity disorder. Biol Psychiatry: Cogn Neurosci Neuroimaging. 2019;4:472–83. doi: 10.1016/j.bpsc.2018.11.014. [DOI] [PubMed] [Google Scholar]
- 53.Norman LJ, Sudre G, Bouyssi-Kobar M, Sharp W, Shaw P. A longitudinal study of resting-state connectivity and response to psychostimulant treatment in ADHD. Am J Psychiatry. 2021;178:744–51. [DOI] [PMC free article] [PubMed]
- 54.Norman LJ, Sudre G, Bouyssi-Kobar M, Sharp W, Shaw P. An examination of the relationships between attention/deficit hyperactivity disorder symptoms and functional connectivity over time. Neuropsychopharmacology. 2022;47:704–10. [DOI] [PMC free article] [PubMed]
- 55.Von Rhein D, Oldehinkel M, Beckmann CF, Oosterlaan J, Heslenfeld D, Hartman CA, et al. Aberrant local striatal functional connectivity in attention‐deficit/hyperactivity disorder. J Child Psychol Psychiatry. 2016;57:697–705. doi: 10.1111/jcpp.12529. [DOI] [PubMed] [Google Scholar]
- 56.Schaefer A, Kong R, Gordon EM, Laumann TO, Zuo X-N, Holmes AJ, et al. Local-global parcellation of the human cerebral cortex from intrinsic functional connectivity MRI. Cereb Cortex. 2018;28:3095–114. doi: 10.1093/cercor/bhx179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Kuznetsova A, Brockhoff PB, Christensen RH. lmerTest package: tests in linear mixed effects models. J Stat Softw. 2017;82:1–26. doi: 10.18637/jss.v082.i13. [DOI] [Google Scholar]
- 58.Yan C-G, Chen X, Li L, Castellanos FX, Bai T-J, Bo Q-J, et al. Reduced default mode network functional connectivity in patients with recurrent major depressive disorder. Proc Natl Acad Sci. 2019;116:9078–83. doi: 10.1073/pnas.1900390116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Boedhoe PS, Heymans MW, Schmaal L, Abe Y, Alonso P, Ameis SH, et al. An empirical comparison of meta-and mega-analysis with data from the ENIGMA obsessive-compulsive disorder working group. Front Neuroinformatics. 2019;12:102. doi: 10.3389/fninf.2018.00102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Miller GA, Chapman JP. Misunderstanding analysis of covariance. J Abnorm Psychol. 2001;110:40. doi: 10.1037/0021-843X.110.1.40. [DOI] [PubMed] [Google Scholar]
- 61.Benjamini Y, Hochberg Y. Controlling the false discovery rate: a practical and powerful approach to multiple testing. J R Stat Soc: Ser B (Methodol) 1995;57:289–300. [Google Scholar]
- 62.Cordova M, Antovich DM, Ryabinin P, Neighbor C, Mooney MA, Dieckmann N, et al. ADHD: Restricted Phenotypes Prevalence, Comorbidity, and Polygenic Risk Sensitivity in ABCD Baseline Cohort. 2021;2021. [DOI] [PMC free article] [PubMed]
- 63.D’Agati E, Curatolo P, Mazzone L. Comorbidity between ADHD and anxiety disorders across the lifespan. Int J Psychiatry Clin Pract. 2019;23:238–44. doi: 10.1080/13651501.2019.1628277. [DOI] [PubMed] [Google Scholar]
- 64.Picon FA, Sato JR, Anés M, Vedolin LM, Mazzola AA, Valentini BB, et al. Methylphenidate alters functional connectivity of default mode network in drug-naive male adults with ADHD. J Atten Disord. 2020;24:447–55. doi: 10.1177/1087054718816822. [DOI] [PubMed] [Google Scholar]
- 65.Christakou A, Murphy CM, Chantiluke K, Cubillo AI, Smith AB, Giampietro V, et al. Disorder-specific functional abnormalities during sustained attention in youth with attention deficit hyperactivity disorder (ADHD) and with autism. Mol Psychiatry. 2013;18:236–44. doi: 10.1038/mp.2011.185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Corbetta M, Shulman GL. Control of goal-directed and stimulus-driven attention in the brain. Nat Rev Neurosci. 2002;3:201–15. doi: 10.1038/nrn755. [DOI] [PubMed] [Google Scholar]
- 67.Uddin LQ, Kelly AC, Biswal BB, Margulies DS, Shehzad Z, Shaw D, et al. Network homogeneity reveals decreased integrity of default-mode network in ADHD. J Neurosci Methods. 2008;169:249–54. doi: 10.1016/j.jneumeth.2007.11.031. [DOI] [PubMed] [Google Scholar]
- 68.Baum GL, Ciric R, Roalf DR, Betzel RF, Moore TM, Shinohara RT, et al. Modular segregation of structural brain networks supports the development of executive function in youth. Curr Biol. 2017;27:1561–72. e8. doi: 10.1016/j.cub.2017.04.051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Kelly AC, Uddin LQ, Biswal BB, Castellanos FX, Milham MP. Competition between functional brain networks mediates behavioral variability. Neuroimage. 2008;39:527–37. doi: 10.1016/j.neuroimage.2007.08.008. [DOI] [PubMed] [Google Scholar]
- 70.Duffy KA, Rosch KS, Nebel MB, Seymour KE, Lindquist MA, Pekar JJ, et al. Increased integration between default mode and task-relevant networks in children with ADHD is associated with impaired response control. Dev Cogn Neurosci. 2021;50:100980. [DOI] [PMC free article] [PubMed]
- 71.Godwin CA, Hunter MA, Bezdek MA, Lieberman G, Elkin-Frankston S, Romero VL, et al. Functional connectivity within and between intrinsic brain networks correlates with trait mind wandering. Neuropsychologia. 2017;103:140–53. doi: 10.1016/j.neuropsychologia.2017.07.006. [DOI] [PubMed] [Google Scholar]
- 72.Sadaghiani S, Poline J-B, Kleinschmidt A, D’Esposito M. Ongoing dynamics in large-scale functional connectivity predict perception. Proc Natl Acad Sci. 2015;112:8463–8. doi: 10.1073/pnas.1420687112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Shappell HM, Duffy KA, Rosch KS, Pekar JJ, Mostofsky SH, Lindquist MA, et al. Children with attention-deficit/hyperactivity disorder spend more time in hyperconnected network states and less time in segregated network states as revealed by dynamic connectivity analysis. NeuroImage. 2021;229:117753. doi: 10.1016/j.neuroimage.2021.117753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Demontis D, Walters RK, Martin J, Mattheisen M, Als TD, Agerbo E, et al. Discovery of the first genome-wide significant risk loci for attention deficit/hyperactivity disorder. Nat Genet. 2019;51:63–75. doi: 10.1038/s41588-018-0269-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Pardoe HR, Hiess RK, Kuzniecky R. Motion and morphometry in clinical and nonclinical populations. Neuroimage. 2016;135:177–85. doi: 10.1016/j.neuroimage.2016.05.005. [DOI] [PubMed] [Google Scholar]
- 76.Marek S, Tervo-Clemmens B, Calabro FJ, Montez DF, Kay BP, Hatoum AS, et al. Reproducible brain-wide association studies require thousands of individuals. Nature. 2022;603:654–60. doi: 10.1038/s41586-022-04492-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Karcher NR, Michelini G, Kotov R, Barch DM. Associations between resting-state functional connectivity and a hierarchical dimensional structure of psychopathology in middle childhood. Biol Psychiatry: Cogn Neurosci Neuroimaging. 2021;6:508–17. doi: 10.1016/j.bpsc.2020.09.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Norman LJ, Taylor SF, Liu Y, Radua J, Chye Y, De Wit SJ, et al. Error processing and inhibitory control in obsessive-compulsive disorder: a meta-analysis using statistical parametric maps. Biol Psychiatry. 2019;85:713–25. doi: 10.1016/j.biopsych.2018.11.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Bernanke J, Luna A, Chang L, Bruno E, Dworkin J, Posner J. Structural brain measures among children with and without ADHD in the Adolescent Brain and Cognitive Development Study cohort: a cross-sectional US population-based study. Lancet Psychiatry. 2022;9:222–31. doi: 10.1016/S2215-0366(21)00505-8. [DOI] [PubMed] [Google Scholar]
- 80.Mooney MA, Hermosillo RJ, Feczko E, Miranda-Dominguez O, Moore LA, Perrone A, et al. Cumulative Effects of Resting-state Connectivity Across All Brain Networks Significantly Correlate with ADHD Symptoms. MedRxiv. 2021;2021. [DOI] [PMC free article] [PubMed]
- 81.Shen X, Finn ES, Scheinost D, Rosenberg MD, Chun MM, Papademetris X, et al. Using connectome-based predictive modeling to predict individual behavior from brain connectivity. Nat Protoc. 2017;12:506–18. doi: 10.1038/nprot.2016.178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Wolfers T, Beckmann CF, Hoogman M, Buitelaar JK, Franke B, Marquand AF. Individual differences v. the average patient: mapping the heterogeneity in ADHD using normative models. Psychological Med. 2020;50:314–23. doi: 10.1017/S0033291719000084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Scheinost D, Noble S, Horien C, Greene AS, Lake EM, Salehi M, et al. Ten simple rules for predictive modeling of individual differences in neuroimaging. NeuroImage. 2019;193:35–45. doi: 10.1016/j.neuroimage.2019.02.057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Sripada C, Rutherford S, Angstadt M, Thompson WK, Luciana M, Weigard A, et al. Prediction of neurocognition in youth from resting state fMRI. Mol Psychiatry. 2020;25:3413–21. doi: 10.1038/s41380-019-0481-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Sripada C, Angstadt M, Rutherford S, Kessler D, Kim Y, Yee M, et al. Basic units of inter-individual variation in resting state connectomes. Sci Rep. 2019;9:1–12. doi: 10.1038/s41598-018-38406-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Woo C-W, Chang LJ, Lindquist MA, Wager TD. Building better biomarkers: brain models in translational neuroimaging. Nat Neurosci. 2017;20:365–77. doi: 10.1038/nn.4478. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Shaw P, Eckstrand K, Sharp W, Blumenthal J, Lerch JP, Greenstein D, et al. Attention-deficit/hyperactivity disorder is characterized by a delay in cortical maturation. Proc Natl Acad Sci. 2007;104:19649–54. doi: 10.1073/pnas.0707741104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Dafflon J, Da Costa PF, Váša F, Monti RP, Bzdok D, Hellyer PJ, et al. Neuroimaging: into the Multiverse. BioRxiv. 2020;2020.
- 89.Couvy-Duchesne B, Ebejer JL, Gillespie NA, Duffy DL, Hickie IB, Thompson PM, et al. Head motion and inattention/hyperactivity share common genetic influences: implications for fMRI studies of ADHD. PloS One. 2016;11:e0146271. doi: 10.1371/journal.pone.0146271. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.