

Abstract
Mammals almost always have seven cervical vertebrae. The strong evolutionary constraint on changes in this number has been broken in sloths and manatees. We have proposed that the extremely low activity and metabolic rates of these species relax the stabilizing selection against changes in the cervical count. Our hypothesis is that strong stabilizing selection in other mammals is largely indirect and due to associated pleiotropic effects, including juvenile cancers. Additional direct selection can occur due to biomechanical problems (thoracic outlet syndrome). Low metabolic and activity rates are thought to diminish these direct and indirect effects. To test this hypothesis within the primates, we have compared the number of cervical vertebrae of three lorisid species with particularly low activity and metabolic rates with those of more active primate species, including with their phylogenetically closest active relatives, the galagids (bushbabies). In support of our hypothesis, we found that 37.6% of the lorisid specimens had an abnormal cervical count, which is a higher percentage than in the other nine primate families, in which the incidence varied from zero to 2.2%. We conclude that our data support the importance of internal selection in constraining evolvability and of a relaxed stabilizing selection for increasing evolvability. Additionally, we discuss that there is no support for a role of the muscularized diaphragm in the evolutionary constraint.
Keywords: cervical ribs, evolutionary constraint, homeotic transformations, mammals, pleiotropic constraint, primates, rudimentary first ribs, stabilizing selection, transitional vertebrae
The number of neck vertebrae in mammals is virtually always seven. Sloths and manatees are famous exceptions to this rule. This is hypothesized to be due to relaxed stabilizing selection against changes, made possible by their extremely low metabolic and activity rates. Here we present further support for this hypothesis, as we find that lorisid primates, also known for their extreme slowness, provide another example of relaxation of this strong evolutionary constraint.
1. INTRODUCTION
The mammalian vertebral column is highly variable in morphology, reflecting adaptations to a wide range of lifestyles, from burrowing in moles to flying in bats (Flower, 1885; Galliari et al., 2010; Granatosky et al., 2014; Jenkins, 1970; Schultz, 1961; Slijper, 1946; Williams & Russo, 2015). However, the number of cervical vertebrae is surprisingly constant at seven (Bateson, 1894; Galis, 1999; Starck, 1979). To retain this number seems, however, not adaptive. Even giraffes with their long necks have seven cervical vertebrae. A giraffe's neck is quite stiff, it requires substantial force to lift it and it is too short to reach the ground unless the front legs are spread wide apart (Figure 1). This awkward position renders giraffes vulnerable to predators while drinking (Périquet et al., 2010; Valeix et al., 2009). Long‐necked birds and reptiles, on the other hand, always have a larger number of cervical vertebrae, allowing for longer and more flexible necks (Figure 1). The constancy in the mammalian neck region contrasts with variability between mammalian species in other vertebral regions: Bottlenose whales have nine thoracic vertebrae while elephants have 19–21. Anteaters have two or three lumbar vertebrae and Indri lemurs eight or nine (Narita & Kuratani, 2005; Varela‐Lasheras et al., 2011).
Figure 1.
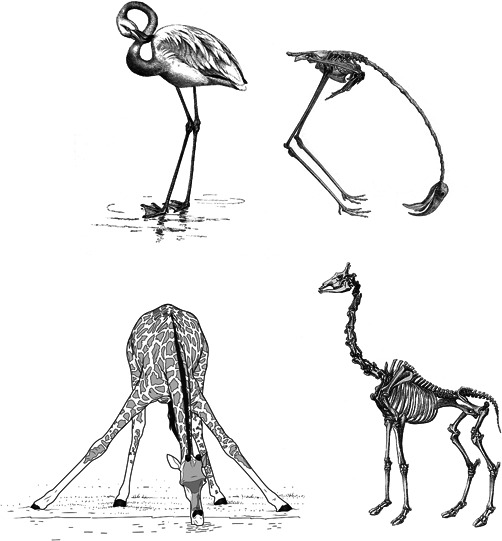
The long neck of the giraffe has only seven vertebrae, which makes it rather stiff, such that lifting it costs considerable force. Despite the length of each cervical vertebra, the neck is not long enough to reach the ground, unless the front legs are spread wide apart, which is an awkward position that makes the giraffe vulnerable to predators. A large number of vertebrae make a long and flexible neck in flamingoes. (a) and (c) Reproduced from Owen (1866). (b) from Evans (1900). (d) Drawing by Erik‐Jan Bosch (Naturalis).
The remarkable conservation of the number of cervical vertebrae is probably caused by strong stabilizing selection. Most hypotheses propose that this selection is indirect and internal, as it is caused by deleterious pleiotropic effects that are associated with the development of an abnormal number of cervical vertebrae (Buchholtz et al., 2012; Galis, 1999; Galis et al., 2006; Hirasawa & Kuratani, 2013; Kuratani, 2004; Narita & Kuratani, 2005). Our hypothesis, based on studies of human and other mammalian individuals with abnormal numbers of cervical vertebrae, is that juvenile cancers and a wide variety of congenital abnormalities are associated with these cervical number changes, leading to an increased mortality before the reproductive age (Furtado et al., 2011; Galis & Metz, 2003; Galis et al., 2006; Galis, 1999; Varela‐Lasheras et al., 2011; Schut et al., 2020a,b; Ten Broek et al., 2012). The association with many different pleiotropic effects is thought to be due to the early determination of the number of neck vertebrae during the phylotypic stage. The strong global interactivity in this stage implies that any perturbation affects many processes and leads to a multitude of changes in different parts of the body (Galis & Metz, 2007; Galis et al., 2018; see also Galis et al., 2002). As a result, the evolvability of the phylotypic stage is severely limited, as is the evolvability of traits determined during this stage.
Kuratani (2004), Hirasawa and Kuratani (2013), and Buchholtz et al. (2012) proposed that the mammalian innovation of a muscularized diaphragm is causally involved in the evolutionary constraint of the number of cervical vertebrae. Associations should then be found between abnormal numbers of cervical vertebrae and anomalies of the muscular diaphragm in mammals.
Direct stabilizing selection against changes in the number of cervical vertebrae can also arise from biomechanical problems. The determination of the neck‐thorax boundary forms part of the early head‐to‐tail patterning of the embryo in which Hox genes play a crucial mediating role (Deschamps & Duboule, 2017). Changes of this boundary necessarily involve homeotic transformations of the seventh and, or eighth vertebra: the seventh normally ribless cervical vertebra changes into a thoracic vertebra with a rib, or the eighth normally thoracic vertebra with a rib changes into a ribless cervical vertebra. Initial mutations for homeotic transformations tend to be incomplete and asymmetric, resulting in so‐called transitional vertebrae (Fischel, 1906; Galis et al., 2014; Jeannotte et al., 1993; Schultz, 1961; Streissler‐Graz, 1913; Varela‐Lasheras et al., 2011). Transitional cervicothoracic vertebrae have rudimentary ribs on the seventh and/or eighth vertebra, which are called cervical ribs (Figures 2 and 3) and rudimentary first ribs, respectively. The rudimentary ribs, or the ligamentous bands attached to them, may cause compression of nerves and/or blood vessels which run into the arm. This can provoke severe degeneration (wasting) of the arm known as the Thoracic Outlet Syndrome (TOS; Adson & Coffey, 1927; Illig et al., 2021; Makhoul & Machleder, 1992; Roos, 1976; Sargent, 1921; Thompson, 1908; Weber, 1913). TOS is particularly disadvantageous when forelimbs are intensively used (Friedman et al., 1959; Rayan, 1988; Sargent, 1921).
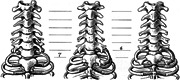
Figure 2.
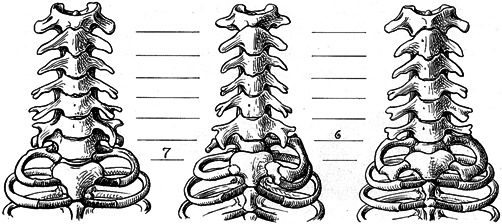
Cervical and upper thoracic regions in three slow lorises, showing the range of variation at the cervicothoracic boundary, drawn by Adolph H. Schultz (1961). On the left, a normal cervical pattern with seven cervical vertebrae. In the middle, an asymmetric pattern with on the left a full cervical rib connected to the seventh vertebra, and on the right a transitional cervicothoracic vertebra with a rudimentary cervical rib. Homeotic transformations of vertebrae often show strong left‐right asymmetry. On the right, two full cervical ribs on the seventh vertebrae. Complete cervical ribs are more rare than rudimentary ribs and were not encountered in this study. Reproduced with permission from Karger, A.G.
Figure 3.
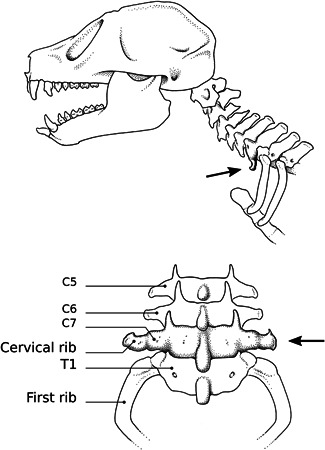
Slow loris skeleton with rudimentary cervical ribs attached to the seventh cervical vertebra (C7). Above: lateral view, with cervical rib indicated by arrow. Below: dorsal view. Drawing by Esmée Winkel (Naturalis).
Stabilizing selection against cervical number variants is probably not as strong in all mammal species. Sloths and manatees are well‐known exceptions among mammals with an aberrant number of cervical vertebrae, with three‐toed sloths (Bradypus tridactylus) usually having eight or nine cervical vertebrae and two‐toed sloths (Choloepus) and manatees (Trichechus) only five or six (Bateson, 1894; Böhmer et al., 2018; Varela‐Lasheras et al., 2011). TOS is not expected to occur in these species, as they move infrequently and when they do, they move slowly (Galis, 1999; Varela‐Lasheras et al., 2011). Furthermore, adult sloths and manatees frequently have skeletal abnormalities (e.g., ossification defects and fused vertebrae, Varela‐Lasheras et al., 2011) which are commonly associated with cervical ribs in deceased human fetuses and infants, but these skeletal abnormalities apparently pose no major problem here. Vestibular system anomalies are also commonly found in sloths (Billet et al., 2013). Furthermore, the extremely low metabolic rates found in sloths and manatees (Irvine, 1983; McNab, 1978, 1988; Pauli et al., 2016) are expected to lower the incidence of pediatric cancers, a pleiotropic effect of cervical ribs found in humans (~120‐fold increase, Galis & Metz, 2003; Galis, 1999; Merks et al., 2005; Schumacher et al., 1992). Low metabolic rates are associated with low levels of DNA damage; hence cancer incidences are expected to be lower in such organisms (Behrend et al., 2003; Cooke et al., 2003; Jackson & Loeb, 2001; Perez‐Campo et al., 1998; Shigenawa & Ames, 1993; Toyokuni et al., 1995). Cancer rates in manatees and sloths have indeed been found to be low (Galis & Metz, 2003; Tollis et al., 2020). Thus, in sloths and manatees, their low activity and metabolic rates presumably relax the stabilizing selection against changes in the number of cervical vertebrae and allow the removal of pleiotropic constraints (Varela‐Lasheras et al., 2011).
In other extremely slow mammals, selection may be relaxed as well relative to their faster sister taxa. To test this, we analyzed the number of cervical vertebrae in three slow primate species, i.e. the red slender loris (Loris tardigradus), the greater slow lori (Nycticebus coucang) and the potto (Perodictitis potto; McNab, 1988; Mueller, 1983, 1985; Napier & Napier, 1967; Rasmussen & Izard, 1988; Shapiro, 2007; Whittow et al., 1977). Locomotion of lorises and pottos has been characterized as slow quadrupedalism, without leaping or running, and branches are gripped with three limbs at any time (Jouffroy, 1989; Napier & Napier, 1967; Shapiro, 2007; Walker, 1969). The greater slow lori has basal and field metabolic rates that are amongst the lowest found in mammals, while the slender lori and potto are slightly less extreme (Genoud, 2002; McNab, 1988; Mueller, 1983, 1985; Rasmussen & Izard, 1988; Whittow et al., 1977). We compared the cervical vertebral patterns of the lorisids with those of various Old World and New World haplorrhine taxa, using phylogenetic comparative analysis (Garamszegi, 2014). Additionally, we compared their pattern frequencies with their phylogenetically closest active relatives, the galagids (bushbabies). Galagids are vertical clingers and spectacular leapers, or active arboreal quadrupeds (Fleagle, 2013; Napier & Napier, 1967; Shapiro, 2007). In agreement with their more active lifestyles, galagids have higher metabolic rates (McNab, 1988; Mueller, 1983, 1985; Rasmussen & Izard, 1988). Additionally, we compared the lorisid cervical patterns with those of lemurids and indriids within the Lemuriformes/Strepsirrhini. Our hypothesis predicts that the frequency of abnormal numbers of cervical vertebrae will be highest in the lorises and pottos in each comparison.
2. METHODS
2.1. Data‐collection
Specimens. We analysed 1090 skeletons of wild‐born primates belonging to 60 species of 10 families (Table 1). These skeletons are held in collections of 10 European and American natural history museums (Naturalis Biodiversity Center, Leiden (Naturalis); The Natural History Museum, London (NHMUK); the Royal Museum for Central Africa, Tervuren (RMCA); the Royal Belgian Institute of Natural Sciences, Brussels (RBINS); the Natural History Museum of Denmark, Copenhagen (ZMUC); Naturhistorisches Museum Wien, Vienna (NHMW); the Swedish Museum of Natural History, Stockholm (NRM); Museum fur Naturkunde, Berlin (MfN); and the National Museum for Natural History, Paris (MNHN), Natural History Museum Oslo, American Museum of Natural History, New York, Field Museum of Natural History, Chicago (FMNH). Five families belonged to the Strepsirrhini (Lorisidae, Galagidae, Daubentoniidae, Lemuridae, Indriidae) and five to the Haplorrhini, of which two Platyrrhini (Cebidae, Atelidae) and three Catarrhini (Cercopithecidae, Hylobatidae, Hominidae).
Table 1.
Normal and abnormal numbers of cervical vertebrae in primates
| Number of individuals evaluated for cervical count | No. of Individuals with seven cervical vertebrae | No. of Individuals with cervical rib or rudimentary first rib | % with cervical rib or rudimentary first rib | ||
|---|---|---|---|---|---|
| Lorisidae | 85 | 50 | 32 | 39.02 | |
| Loris tardigradus | 5 | 3 | 2 | 40.00 | |
| Nycticebus coucang | 40 | 20 | 19 | 48.72 | |
| Perodicticus potto | 40 | 27 | 11 | 28.95 | |
| Galagidae | 90 | 88 | 0 | 0.00 | |
| Galago moholi | 10 | 10 | 0 | 0.00 | |
| Galago senegalensis | 33 | 32 | 0 | 0.00 | |
| Otolemur crassicaudatus | 47 | 46 | 0 | 0.00 | |
| Indriidae | 56 | 54 | 1 | 1.82 | |
| Indri indri | 20 | 20 | 0 | 0.00 | |
| Propithecus coquereli | 14 | 14 | 0 | 0.00 | |
| Propithecus deckenii | 5 | 5 | 0 | 0.00 | |
| Propithecus verreauxi | 17 | 15 | 1 | 6.25 | |
| Lemuridae | 95 | 93 | 1 | 1.06 | |
| Lemur catta | 39 | 39 | 0 | 0.00 | |
| Eulemur mongoz | 31 | 31 | 0 | 0.00 | |
| Eulemur albifrons | 16 | 14 | 1 | 6.67 | |
| Eulemur collaris | 3 | 3 | 0 | 0.00 | |
| Eulemur fulvus | 5 | 5 | 0 | 0.00 | |
| Eulemur rufus | 1 | 1 | 0 | 0.00 | |
| Daubentoniidae | 16 | 16 | 0 | 0.00 | |
| Daubentonia madagascariensis | 16 | 16 | 0 | 0.00 | |
| Atelidae | 98 | 92 | 2 | 2.13 | |
| Ateles paniscus | 23 | 22 | 0 | 0.00 | |
| Ateles chamek | 1 | 1 | 0 | 0.00 | |
| Ateles belzebuth | 15 | 14 | 1 | 6.67 | |
| Ateles marginatus | 1 | 1 | 0 | 0.00 | |
| Ateles geoffroyi | 8 | 7 | 1 | 12.50 | |
| Ateles fuscipes | 1 | 1 | 0 | 0.00 | |
| Ateles sp. | 5 | 4 | 0 | 0.00 | |
| Alouatta seniculus | 37 | 36 | 0 | 0.00 | |
| Alouatta belzebul | 2 | 2 | 0 | 0.00 | |
| Alouatta macconnelli | 4 | 3 | 0 | 0.00 | |
| Alouatta guariba | 1 | 1 | 0 | 0.00 | |
| Cebidae | 37 | 36 | 0 | 0.00 | |
| Saimiri sciureus | 35 | 34 | 0 | 0.00 | |
| Saimiri sp. | 2 | 2 | 0 | 0.00 | |
| Cercopithecidae | 359 | 347 | 0 | 0.00 | |
| Chlorocebus aethiops | 20 | 19 | 0 | 0.00 | |
| Chlorocebus sabaeus | 10 | 10 | 0 | 0.00 | |
| Chlorocebus tantalus | 1 | 1 | 0 | 0.00 | |
| Chlorocebus pygerythrus | 8 | 8 | 0 | 0.00 | |
| Chlorocebus cynosuros | 12 | 12 | 0 | 0.00 | |
| Cercopithecus albulogaris | 31 | 29 | 0 | 0.00 | |
| Cercopithecus mitis | 43 | 40 | 0 | 0.00 | |
| Cercopithecus nictitans | 26 | 25 | 0 | 0.00 | |
| Cercopithecus lhoesti | 20 | 19 | 0 | 0.00 | |
| Cercopithecus solatus | 6 | 6 | 0 | 0.00 | |
| Cercopithecus preussi | 5 | 5 | 0 | 0.00 | |
| Cercopithecus ascanius | 28 | 28 | 0 | 0.00 | |
| Erythrocebus patas | 47 | 46 | 0 | 0.00 | |
| Theropithecus gelada | 20 | 20 | 0 | 0.00 | |
| Papio hamadryas | 39 | 39 | 0 | 0.00 | |
| Papio anubis | 42 | 39 | 0 | 0.00 | |
| Hylobatidae | 68 | 68 | 0 | 0.00 | |
| Symphalangus syndactylus | 21 | 21 | 0 | 0.00 | |
| Hylobates klossii | 3 | 3 | 0 | 0.00 | |
| Hylobates lar | 9 | 9 | 0 | 0.00 | |
| Hylobates moloch | 16 | 16 | 0 | 0.00 | |
| Hylobates agilis | 7 | 7 | 0 | 0.00 | |
| Hylobates pileatus | 1 | 1 | 0 | 0.00 | |
| Hylobates muelleri | 4 | 4 | 0 | 0.00 | |
| Hylobates albibarbis | 1 | 1 | 0 | 0.00 | |
| Hylobates sp. | 6 | 6 | 0 | 0.00 | |
| Hominidae | 186 | 179 | 4 | 2.19 | |
| Pongo pygmaeus | 31 | 30 | 0 | 0.00 | |
| Gorilla gorilla | 15 | 15 | 0 | 0.00 | |
| Gorilla beringei | 37 | 37 | 0 | 0.00 | |
| Pan troglodytes | 72 | 67 | 3 | 4.29 | |
| Pan paniscus | 31 | 179 | 1 | 0.56 | |
2.1.1. Cervical vertebrae and transitional cervicothoracic vertebrae
We determined the number of cervical vertebrae and transitional cervicothoracic vertebrae (vertebrae with both cervical and thoracic characteristics, that is, a seventh vertebra with a rudimentary rib or one full rib instead of two, or an eighth vertebra with rudimentary ribs or without ribs on one side). The identification of transitional cervicothoracic vertebrae was based on the presence of cervical or rudimentary first ribs. In the case of a fusion of rudimentary cervical ribs with the transverse process (apophysomegaly), the vertebra was counted as a transitional cervicothoracic vertebra when the transverse process was at least 15% longer than that of the first thoracic vertebra, or when traces of the articulation were still visible.
2.1.2. Explanatory variables
Per specimen where we determined the vertebral pattern, we recorded the species, lifestyle (“fast” vs. “slow”), individual age class and sex, and whether the animal was kept in a zoo later in life or not. This last indicator variable can accommodate effects of relaxed selection in captive environments on the probability of finding an abnormal pattern.
2.1.3. Phylogeny
We used the consensus phylogeny of primates provided by the 10k Trees Project (Arnold & Nunn, 2010) to represent our data per species graphically and to calculate correlations between species effects in the phylogenetic mixed model analyses described below.
2.2. Statistical analysis
The data consist per primate of an indicator variable whether that individual had a normal pattern with seven cervical vertebrae or not, with cervical and rudimentary first ribs on the seventh and eight vertebrae (abnormal pattern), and of the values of the explanatory variables. Our analysis of the incidence of abnormal patterns treated most explanatory variables as fixed effects and the species effects as random. Random species effects can consist of independent random environmental and sampling effects and so‐called phylogenetic effects (Garamszegi, 2014) which are heritable and assume covariances between species which decrease with phylogenetic distance. Correlated phylogenetic effects are often believed to “correct” for the nonindependence of data on different species due to shared evolutionary history (Revell, 2010; Rohlf, 2006). We note, however, that other explanatory variables shared between species or correlated environmental effects due to correlated sampling are other sources of nonindependence of data values.
The precise pattern of the dependence of phylogenetic species covariances on phylogenetic distance is determined by how selection and drift are assumed to operate on the heritable part of species effects (Hansen & Martins, 1996). With drift and directional selection, correlations between species effects scale with the proportion of evolutionary time on the phylogeny between the root and the most recent common ancestor of each pair of species. With drift and strong stabilizing selection, covariances are expected to become minimal and difficult to distinguish from independence. With weaker stabilizing selection, correlations decrease exponentially with distance, and the decrease is characterized by the parameter α of the OU (Ornstein‐Uhlenbeck) process describing the change in heritable species effects over time due to stabilizing selection (Hansen & Martins, 1996).
We hypothesize strong directional selection against the incidence of cervical ribs in primate species which are mobile and much less so in slow movers. This would imply that drift would have a larger impact on species covariances among slow movers. However, the number of species in that group is too small to estimate a separate covariance pattern for them. Additionally, binary traits are zero‐one responses. In generalized mixed linear models, these are modeled as proportions constrained between zero and one which depend on an underlying continuous liability. Proportions increase in a nonlinear manner as a function of the liability, usually via an inverse logit function. It is for the random species effects contributing to these underlying liabilities that we can model non‐zero species covariances. For very low and very high probabilities of cervical ribs, the effect of changing the liability on the predicted incidence is weak such that selection on the liability must weaken in species with low incidences of cervical ribs. This pattern of selection is akin to stabilizing selection which decreases when approaching an optimum. However, here the selection optimum is on the boundary of trait space and therefore at minus infinity for the liabilities. It is therefore unclear if predictions of models of stabilizing selection apply fully. Stronger selection in fast moving species would imply that their heritable species liability effects are closer to independent (Cooper et al., 2016). Summarizing, there might be non‐zero covariances that depend on phylogenetic distance but it is unclear which random species effects model applies best. We, therefore, modeled different correlation patterns.
The molecular phylogeny of primates was trimmed to the species with data and explanatory variables. Eight species with data could not be attributed to a species in the phylogeny were removed from the mixed model analysis. We used function pglmm() from the phyr package (Ives & Helmus, 2011) for R (R Core team, 2021) to fit the binary phylogenetic generalized mixed models. First, we investigated which pattern of correlations in species effects fitted the data best. Different models including all fixed effects (sex, age class, life style, kept in zoo) but with different species effect specifications were fitted using REML. They had either (1) joint phylogenetic and independent species effects, (2) only independent species effects or (3) phylogenetic species effects only. For (1) and (3) the phylogenetic effect correlations were either calculated assuming Brownian motion (function vcv. phylo ()) or for OU processes with different strengths of stabilizing selection (α = .01, .02, .04, .06, and .10) using the corMartins() function from the ape library (Paradis & Schliep, 2019) to calculate species correlation matrices. For the smallest value of α, correlations between species effects were only slightly smaller than for Brownian motion. We then used a numerical optimizer to find the value of α which minimized the deviance of the phylogenetic mixed model, this for a model including all fixed effects and starting from the α among the values above which had the largest likelihood. We compared model deviances among these models fitted and carried out likelihood ratio tests (Self & Liang, 1987) on the random effects in the model with minimal deviance. Subsequently, we refitted the best model using ML and used likelihood ratio tests to test for the significance of each fixed effect.
Our data do not need to satisfy all assumptions of the mixed models fitted and the tail probabilities of the likelihood ratio tests for the fixed effects above might be slightly conservative or anticonservative (Pinheiro & Bates, 2000). The change to a slow life style also occurred only once across the primates. Therefore, two additional specific pairwise comparisons between families were made based on generalized linear models: (1) a comparison of proportions of normal patterns between slow lorisids and galagids; (3) a comparison of lorisids with indriids and lemurids combined. To account for heterogeneity in incidence probabilities between species within these families, we fitted generalized linear models to the data with quasibinomial distributions, allowing variance estimates which differ from the expectation based on the binomial distribution. All explanatory variables are categorical and our individual data could therefore show overdispersion or underdispersion (Hilbe, 2013). We found that underdispersion might occur and report nevertheless difference tests from models for binomial distributions, as these tests were more conservative, i.e., they had larger tail probabilities. The R script with our analysis is available as supplementary material.
3. RESULTS
Overall, we found that only 40 out of 1063 primate individuals (3.8%) did not have seven cervical vertebrae (1063 of 1089 skeletons could be scored; Figure 4 and Table 1). Lorisids were the only family with a high number of abnormal cervical counts, all due to transitional cervicothoracic vertebrae (39.0% of 82 lorisids). In the other families, the incidences ranged from 0% to 2.2%. The change in the number of cervical vertebrae resulted most often from rudimentary cervical ribs (homeotic transformation of the seventh vertebra) and, more exceptionally (in two pottos), from rudimentary first ribs (homeotic transformation of the eighth vertebra). In the statistical modeling, we analyzed the proportions of individuals with a normal cervical vertebral pattern (absence of transitional vertebrae).
Figure 4.
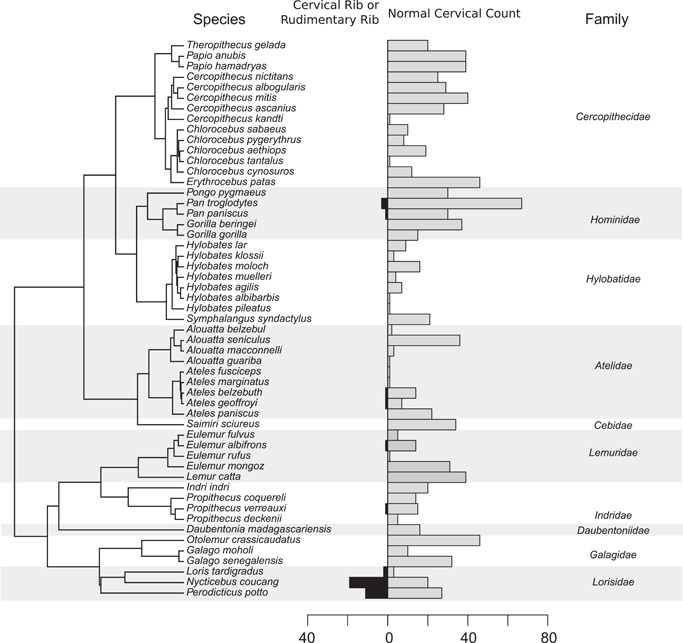
Molecular phylogeny of primate species involved in our study. Bars to the right of each species indicate the number of individuals with an abnormal and a normal cervical vertebral pattern. Primate families are indicated.
Our phylogenetic comparative analysis using mixed models found that a model without independent species effects and with a phylogenetic random species effect calculated from an OU model with α = .098 fitted the data best. However, for this random effect specification, correlations are only above 0.01 for closely related species and accordingly a likelihood ratio comparison determined that a model with only independent species effects was not significantly worse (χ 2(1) = 2.84, p = .09). Likelihood ratio tests did indicate that including a random species effect contributed significantly to explaining the data pattern (independent effects χ 2(0 | 1) = 2.74, p = .049, OU correlated χ 2(0 | 1) = 6.046, p = .007). We tested the significance of fixed effects in the best OU model. The effect of life style was significant in the model with all explanatory variables. Slow primates have a reduced probability of a normal cervical vertebral pattern (estimate liability effect −4.45 (SE 0.59), χ 2(1) = 64.33, p < .0001). Age, sex and whether the individual was kept in a zoo did not have significant effects. We, therefore, did not add these in the generalized linear models below.
When we compared family effects between lorisids and galagids using generalized linear models, the incidence of normal cervical vertebral patterns differed between both (χ 2(1) = 54.75, p < .0001), but confidence intervals of the effects were so wide that no meaningful interval for the difference resulted. This was due to the absence of abnormal patterns in the galagids, all individuals having seven cervical vertebrae. A Fisher exact test not depending on parameter estimates and generalized linear model assumptions, also found that the incidence of abnormal patterning differed between both (p < .0001). For the comparison between Lorisidae and Indriidae and Lemuridae combined, we found that the incidence of seven cervical vertebrae was significantly lower in Lorisidae (χ 2(1) = 62.11, p < .0001), parameter estimate effect size difference −3.85 (95% confidence interval −5.69 to −2.61).
4. DISCUSSION
In an earlier study, we compared sloths and manatees with sister taxa with a generally normal cervical pattern (Varela‐Lasheras et al., 2011). We found a strong association between abnormal cervical numbers and congenital abnormalities in all investigated taxa. This supported the hypothesis that the extremely low activity and metabolic rates of sloths and manatees reduce the strength of stabilizing selection against changes in the number of cervical vertebrae, thus relaxing a strong evolutionary constraint. In this study we found further support for this hypothesis. Relative to sister taxa and primates overall, an increased incidence of transitional cervicothoracic vertebrae was found in lorises and pottos, primates with particularly low activity and metabolic rates. Therefore, the strength of stabilizing selection is most likely reduced because of a mitigation of at least some of the pleiotropic effects that tend to be associated with changes in the cervical count in other mammals, that is, pediatric cancers, thoracic outlet syndrome and congenital abnormalities.
A substantial number of pottos and lorises had an abnormal number of cervical vertebrae, usually caused by cervical ribs and, more rarely, by rudimentary first ribs (homeotic transformations of the seventh and eighth vertebrae, respectively). In all other primate families where we collected data, we found a low incidence of abnormal numbers of cervical vertebrae, virtually always due to cervical ribs. Previously, we found an incidence of cervical ribs in dugongs of 45% (Varela‐Lasheras et al., 2011), which is comparable to the incidence in Lorisidae. Dugongs also have low activity and metabolic rates, although somewhat less extreme than manatees (Lanyon et al., 2006; Zeh et al., 2018). In sloths and manatees, however, the deviation from the canonical number of seven is larger than in lorisids and in dugongs. The finding that the largest incidences of deviations from the canonical cervical number occurs in the species with the lowest known metabolic and activity rates of mammals (Gallivan & Best, 1980; Hammond & Diamond, 1997; Koteja, 1991; McNab, 1988), further supports our hypothesis. A reduced incidence and severity of deleterious pleiotropic effects potentially allows directional selection for more or fewer cervical vertebrae to dominate. This may well have been the case for three‐toed sloths, known to have a remarkable ability to rotate their long necks with their many neck vertebrae (Bell, 1834; Mendel, 1985), whereas the short neck of manatees may have been selected to improve streamlining (cf. dolphins with short necks due to fusion of the seven cervical vertebrae). A reduced strength of stabilizing selection also allows a larger trait variability. In lorisids, we found no clear indication of directional selection on the number of cervical vertebrae, only an increased incidence of transitional vertebrae.
Deleterious pleiotropic effects and the resulting strong selection against changes in the number of cervical vertebrae have been most thoroughly investigated in humans (Furtado et al., 2011; Galis et al., 2006; Schut et al., 2020a,b; Ten Broek et al., 2012). However, studies on a wide variety of mammals, including afrotherians and xenarthrans (Varela‐Lasheras et al., 2011), thoroughbred horses (May‐Davis, 2017), extinct woolly mammoths and woolly rhinos (Reumer et al., 2014; van der Geer & Galis, 2017) and domesticated dogs (Brocal et al., 2018) further support the association of deleterious pleiotropic effects with abnormal cervical numbers. One of the pleiotropic effects which can occur in humans, an increased probability of juvenile cancer in humans, has an estimated probability of occurrence of 12% in individuals with a cervical rib (a 120‐fold increase in comparison to the general population, Galis, 1999; see also Merks et al., 2005; Schumacher et al., 1992). The low metabolic rates of lorisids are expected to drastically lower cancer rates and, hence, weaken indirect selection against changes in the cervical number (Galis & Metz, 2003; Galis, 1999; Varela‐Lasheras et al., 2011). Indeed, despite the remarkably high incidence of cervical ribs in lorisids, among 233 neonatal and juvenile deaths of lorisids in zoos, none was found to have a tumor (Simpson et al., 2018). Similarly, in manatees and sloths, cancer rates appeared to be exceptionally low (Galis & Metz, 2003; Tollis et al., 2020). The moderate use of the forelimbs of lorisids makes it unlikely that they will have TOS problems caused by pressure on the nerves and blood vessels. In humans, TOS is in particular a problem among athletes with well‐developed muscles which decrease the available space in the thoracic outlet (Illig et al., 2021; Rayan, 1988; Roos, 1976, 1996). In thoroughbred racing horses, rudimentary first ribs and cervical ribs (described as flared or bicipital first ribs) are associated with anatomical changes in the thoracic outlet, with disturbances of forelimb locomotion and with pain, in agreement with TOS in humans (May‐Davis, 2017). The moderate use of the forelimbs of lorisids makes it unlikely that they will have TOS problems. Finally, congenital abnormalities are further pleiotropic effects associated with abnormal cervical numbers in humans, sloths, manatees, dugongs, hyraxes, and horses (Galis et al., 2006; May‐Davis, 2017; Schut et al., 2020a,b; Ten Broek et al., 2012; Varela‐Lasheras et al., 2011). We only had access to skeletons for this analysis and can therefore only conjecture that selection against some potential congenital anomalies might be weaker (Billet et al., 2013; Varela‐Lasheras et al., 2011).
Kuratani (2004), Hirasawa and Kuratani (2013), and Buchholtz et al. (2012) have proposed that the evolutionary novelty of the muscularization of the diaphragm in mammals is involved in the fixation of the number of cervical vertebrae. They hypothesized that during early development, the migration of precursor muscles cells from the cervical somites to the membranous scaffold of the diaphragm constrains the development of an abnormal number of cervical vertebrae. However, empirical data do not support this hypothesis, which requires an association between anomalous patterns in cervical vertebrae and in the muscularized diaphragm. No diaphragm abnormalities have been reported in many mouse transgenic mutants with a changed number of cervical vertebrae (e.g. Horan et al., 1994; Rancourt et al., 1995; Takihara et al., 1997), nor in mice and rats with cervical ribs induced by teratological treatments (Abdulrazzaq et al., 1997; Leung & Ballantyne, 1999; Narotsky et al., 1998). A changed number of cervical vertebrae in one mouse Hox mutant was found together with an abnormality of the diaphragm (Hoxa5‐/‐) (Hoxa5‐/‐), however, it concerned improper innervation of the diaphragm and not an abnormality of the muscularized diaphragm itself (Boucherat et al., 2013). Furthermore, mice mutants without diaphragm muscle cells are also not associated with changes in the number of cervical vertebrae (e.g., six1‐/‐ mutants; Laclef et al., 2003). In Splotch mutants, the occasional cervical ribs are due to a lack of early activity of Pax3 before the formation of somites, whereas the failure of migration of diaphragm muscle cells is due to the lack of later activity of Pax3 with wider effects. (Schubert et al., 2001). Furthermore, among 68 children with diaphragmatic hernia none had an abnormal number of cervical vertebrae, or other vertebral abnormalities (Chiba et al., 1991). There is, thus, a general lack of empirical support that the migration of muscle precursor cells through the lateral plate mesoderm to the membranous diaphragm interferes with the mesodermal cervical vertebral development. In agreement with this observation, almost all lorisid individuals with an abnormal number of cervical vertebrae survived until adulthood and are, therefore unlikely to have had serious diaphragm abnormalities.
This also fits with the observation that the evolutionary constraint on the number of cervical vertebrae is probably not restricted to mammals, but holds more broadly for all limbed amniotes, as long as they don't have long necks (Galis, 2023). In reptiles, the cervical count is quite constant in families with nine or fewer neck vertebrae, such as pterosaurs, crocodiles, turtles, geckos, and many other lizards (Bennett, 2014; Hofstetter & Gasc, 1969). In contrast, the number can be highly variable in reptiles with many cervical vertebrae (e.g., many plesiosaurs and dinosaurs) and in birds, which have numbers that vary between 12 in pigeons and 23–25 in swans (Woolfenden, 1961). A larger number of neck vertebrae implies a later determination of the cervicothoracic boundary, after the vulnerable and highly interactive phylotypic stage, when development becomes more modular. The increased modularity should lead to a weakening of the constraint, as fewer side‐effects are expected to be associated with changes, similar to a weaker evolutionary constraint on changes at the thoracolumbar boundary (Galis et al., 2006). Other examples of late‐determined structures are carpal and tarsal elements, phalanges, teeth, trunk and caudal vertebrae in amniotes, and nipples in mammals, which are all highly evolvable (Galis, 2023; Galis & Metz, 2007).
We conclude that lorisids provide another example where a strong evolutionary constraint in mammals has been relaxed. The data support our hypothesis that stabilizing selection against changes in the number of cervical vertebrae is weaker due to low metabolic rates and limited activity. More generally, the data support the importance of internal selection in constraining evolvability and of a relaxed stabilizing selection for increasing evolvability.
CONFLICTS OF INTEREST
The authors declare no conflicts of interest.
ACKNOWLEDGEMENTS
We thank the following curators and collection managers for their assistance with the primate material in their care: Wim Wendelen and Emmanuel Gilissen (RMCA), Georges Lenglet (IRSNB‐KBIN), Mogens Andersen, Daniel Klingberg Johansson and Trine Sørensen (ZMUC), Christine Lefèvre, Joséphine Lesur, Jacques Cuisin (MNHN), Nora Lange and Manja Voss (NHMB), Roberto Portela‐Miguez (NHM), Alex Bibl (NMW), Daniela Kalthoff, Peter Mortensen and Peter Nilsson (NRM), Eileen Westwig, Ross MacPhee and Neil Duncan (AMNH), Lawrence Heaney and the late Bill Stanley (FMNH), and Pepijn Kamminga (Naturalis). We thank Erik‐Jan Bosch and Esmée Winkel (Naturalis) for the beautiful drawings. This study was in part supported by Synthesys travel grants to F.G. and A.v.d.G. to visit the Royal Museum for Central Africa Tervuren, The Natural History Museum Berlin, the Zoological Museum Copenhagen, the National Museum for Natural History in Paris, the Natural History Museum of Stockholm, The Natural History Museum London (Grants BE‐TAF‐1649, BE‐TAF‐6522, DE‐TAF‐2114, DE‐TAF‐5865, DK‐TAF‐2183, DK‐TAF‐4886, FR‐TAF‐4104, SE‐TAF‐3009, GB‐TAF‐2762, respectively).
Galis, F. , Van Dooren, T. J. M. , & Geer, A. A. E. (2022). Breaking the constraint on the number of cervical vertebrae in mammals: On homeotic transformations in lorises and pottos. Evolution & Development, 24, 196–210. 10.1111/ede.12424
DATA AVAILABILITY STATEMENT
The data and the R script used for analysis are accessible via the Zenodo.org platform at https://doi.org/10.5281/zenodo.72210911, under licence CC BY‐NC‐SA 4.0.
REFERENCES
- Abdulrazzaq, Y. M. , Bastaki, S. M. A. , & Padmanabhan, R. (1997). Teratogenic effects of vigabatrin in TO mouse fetuses. Teratology, 55, 165–176. [DOI] [PubMed] [Google Scholar]
- Adson, A. W. , & Coffey, J. R. (1927). Cervical rib: A method of anterior approach for relief of symptoms by division of the scalenus anticus. Annals of Surgery, 85(6), 839–857. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arnold, C. , & Nunn, C. L. (2010). Phylogenetic targeting of research effort in evolutionary biology. The American Naturalist, 176, 601–612. [DOI] [PubMed] [Google Scholar]
- Bateson, W. (1894). Materials for the study of variation. London: MacMillan. [Google Scholar]
- Behrend, L. , Henderson, G. , & Zwacka, R. M. (2003). Reactive oxygen species in oncogenic transformation. Biochemical Society Transactions, 31(6), 1441–1444. [DOI] [PubMed] [Google Scholar]
- Bell, T. (1834). XII. Observations on the neck of the three‐toed sloth, Bradypus tridactylus, Linn. The Transactions of the Zoological Society of London, 1, 113–116. [Google Scholar]
- Bennett, S. C. (2014). A new specimen of the pterosaur Scaphogranthus crassirostris, with comments on constraint of cervical vertebrae number in pterosaurs. Neues Jahrbuch für Geologie und Paläontologischen Abhandlungen, 27, 327–348. [Google Scholar]
- Billet, G. , Germain, D. , Ruf, I. , de Muizon, C. , & Hautier, L. (2013). The inner ear of megatherium and the evolution of the vestibular system in sloths. Journal of Anatomy, 223, 557–567. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blair Irvine, A. (1983). Manatee metabolism and its influence on distribution in Florida. Biological Conservation, 25(4), 315–334. [Google Scholar]
- Böhmer, C. , Amson, E. , Arnold, P. , van Heteren, A. H. , & Nyakatura, J. A. (2018). Homeotic transformations reflect departure from the mammalian ‘rule of seven’ cervical vertebrae in sloths: Inferences on the Hox code and morphological modularity of the mammalian neck. BMC Evolutionary Biology, 18, 84. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boucherat, O. , Montaron, S. , Bérubé‐Simard, F.‐A. , Aubin, J. , Philippidou, P. , Wellik, D. M. , Dasen, J. S. , & Jeannotte, L. (2013). Partial functional redundancy between Hoxa5 and Hoxb5 paralog genes during lung morphogenesis. American Journal of Physiology‐Lung Cellular and Molecular Physiology, 304(12), L817–L830. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brocal, J. , De Decker, S. , José‐López, R. , Manzanilla, E. G. , Penderis, J. , Stalin, C. , Bertram, S. , Schoenebeck, J. J. , Rusbridge, C. , Fitzpatrick, N. , & Gutierrez‐Quintana, R. (2018). C7 vertebra homeotic transformation in domestic dogs – Are Pug dogs breaking mammalian evolutionary constraints? Journal of Anatomy, 233(2), 255–265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buchholtz, E. A. , Bailin, H. G. , Laves, S. A. , Yang, J. T. , Chan, M.‐Y. , & Drozd, L. E. (2012). Fixed cervical count and the origin of the mammalian diaphragm. Evolution & Development, 14, 399–411. [DOI] [PubMed] [Google Scholar]
- Chiba, T. , Kamiyama, T. , & Ohi, R. (1991). Two cases of bizarre rib anomalies associated with intestinal malformation. Nihon geka hokan. Archiv fur japanische Chirurgie, 60(1), 75–79. [PubMed] [Google Scholar]
- Cooke, M. S. , Evans, M. D. , Dizdaroglu, M. , & Lunec, J. (2003). Oxidative DNA damage: Mechanisms, mutation, and disease. The FASEB Journal, 17, 1195–1214. [DOI] [PubMed] [Google Scholar]
- Cooper, N. , Thomas, G. H. , & FitzJohn, R. G. (2016). Shedding light on the ‘dark side’ of phylogenetic comparative methods. Methods in Ecology and Evolution, 7, 693–699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deschamps, J. , & Duboule, D. (2017). Embryonic timing, axial stem cells, chromatin dynamics, and the Hox clock. Genes & Development, 31, 1406–1416. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Evans, T. H. (1900). Birds. The Cambridge natural history (3). Macmillan. [Google Scholar]
- Fischel, A. (1906). Untersuchungen über die Wirbelsäule und den Brustkorb des Menschen. Beiträge und Referate zur Anatomie und Entwickelungsgeschichte, 31(3), 462–588. [Google Scholar]
- Fleagle, J. G. (2013). Primate adaptation & evolution (3rd edition.). Elsevier. [Google Scholar]
- Flower, W. H. (1885). An introduction to the osteology of the mammalia (3rd edition.). Macmillan. [Google Scholar]
- Friedman, H. H. , Argyros, T. G. , & Steinbrocker, O. (1959). Neurovascular syndromes of the shoulder girdle and upper extremity, the compression disorders and the shoulder‐hand syndrome. Postgraduate Medical Journal, 35, 397–425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Furtado, L. V. , Thaker, H. M. , Erickson, L. K. , Shirts, B. H. , & Opitz, J. M. (2011). Cervical ribs are more prevalent in stillborn fetuses than in live‐born infants and are strongly associated with fetal aneuploidy. Pediatric and Developmental Pathology, 14, 431–437. 10.2350/11-01-0974-OA.1 [DOI] [PubMed] [Google Scholar]
- Galis, F. (2023). Evolvability of body plans: On phylotypic Stages, developmental Modularity, and an ancient metazoan constraint. In Wagner G. P., Hansen T. F., Pelabon C., & Pavlicev M. (Eds.), Evolvability. MIT Press. [Google Scholar]
- Galis, F. (1999). Why do almost all mammals have seven cervical vertebrae? Developmental constraints, hox genes, and cancer. Journal of Experimental Zoology, 285, 19–26. 10.1002/(SICI)1097-010X(19990415)285:1<19::AID-JEZ3>3.0.CO;2-Z [DOI] [PubMed] [Google Scholar]
- Galis, F. , Carrier, D. R. , van Alphen, J. , van der Mije, S. D. , Van Dooren, T. J. M. , Metz, J. A. J. , & ten Broek, C. M. A. (2014). Fast running restricts evolutionary change of the vertebral column in mammals. Proceedings of the National Academy of Sciences, 111(31), 11401–11406. 10.1073/pnas.1401392111 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Galis, F. , Van Dooren, T. J. M. , & Metz, J. A. J. (2002). Conservation of the segmented germband stage: robustness or pleiotropy? Trends in Genetics, 18, 504–509. [DOI] [PubMed] [Google Scholar]
- Galis, F. , Dooren, T. J. M. V. , Feuth, J. D. , Metz, J. A. J. , Witkam, A. , Ruinard, S. , Steigenga, M. J. , & Wunaendts, L. C. D. (2006). Extreme selection in humans against homeotic transformations of cervical vertebrae. Evolution, 60, 2643–2654. 10.1111/j.0014-3820.2006.tb01896.x [DOI] [PubMed] [Google Scholar]
- Galis, F. , & Metz, J. A. J. (2003). Anti‐cancer selection as a source of developmental and evolutionary constraints. BioEssays, 25, 1035–1039. 10.1002/bies.10366 [DOI] [PubMed] [Google Scholar]
- Galis, F. , & Metz, J. A. J. (2007). Evolutionary novelties: The making and breaking of pleiotropic constraints. Integrative and Comparative Biology, 47, 409–419. [DOI] [PubMed] [Google Scholar]
- Galis, F. , Metz, J. A. J. , & van Alphen, J. J. M. (2018). Development and evolutionary constraints in animals. Annual Review of Ecology Evolution and Systematics, 49, 499–522. [Google Scholar]
- Galliari, F. C. , Carlini, A. A. , & Sánchez‐Villagra, M. R. (2010). Evolution of the axial skeleton in armadillos (mammalia, Dasypodidae). Mammalian Biology, 75, 326–333. [Google Scholar]
- Gallivan, G. J. , & Best, R. C. (1980). Metabolism and respiration of the Amazonian manatee (Trichechus inunguis). Physiological Zoology, 53, 245–253. [Google Scholar]
- Garamszegi L. Z. (Ed.). (2014). Modern phylogenetic comparative methods and their application in Evolutionary Biology ‐ Concepts and Practice. Springer. [Google Scholar]
- van der Geer, A. A. E. , & Galis, F. (2017). High incidence of cervical ribs indicates vulnerable condition in late Pleistocene woolly rhinoceroses. PeerJ, 5, e3684. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Genoud, M. (2002). Comparative studies of basal rate of metabolism in primates. Evolutionary Anthropology: Issues, News, and Reviews, 11, 108–111. [Google Scholar]
- Granatosky, M. C. , Lemelin, P. , Chester, S. G. B. , Pampush, J. D. , & Schmitt, D. (2014). Functional and evolutionary aspects of axial stability in Euarchontans and other mammals. Journal of Morphology, 275, 313–327. [DOI] [PubMed] [Google Scholar]
- Hammond, K. A. , & Diamond, J. (1997). Maximal sustained energy budgets in humans and animals. Nature, 386, 457–462. [DOI] [PubMed] [Google Scholar]
- Hansen, T. F. , & Martins, E. P. (1996). Translating between microevolutionary process and macroevolutionary patterns: the correlation structure of interspecific data. Evolution, 50, 1404–1417. [DOI] [PubMed] [Google Scholar]
- Hilbe, J. M. (2013). Can binary logistic models be overdispersed? Available at http://highstat.com/Books/BGS/GLMGLMM/pdfs/HILBE-Can_binary_logistic_models_be_overdispersed2Jul2013.pdf
- Hirasawa, T. , & Kuratani, S. (2013). A new scenario of the evolutionary derivation of the mammalian diaphragm from shoulder muscles. Journal of Anatomy, 222, 504–517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hofstetter, R. , & Gasc, J.‐P. (1969). Vertebrae and ribs of modern reptiles. In Gans C. (Ed.), Biology of reptiles (pp. 201–310). Academic Press. [Google Scholar]
- Horan, G. S. , Wu, K. , Wolgemuth, D. J. , & Behringer, R. R. (1994). Homeotic transformation of cervical vertebrae in Hoxa‐4 mutant mice. Proceedings of the National Academy of Sciences, 91, 12644–12648. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Illig, K. A. , Thompson, R. W. , Freischlag, J. A. , Donaghue, D. M. , Jordan, S. E. & Edgelow, P. I. , (Eds.). (2021). Thoracic outlet Syndrome. Springer Verlag. [Google Scholar]
- Ives, A. R. , & Helmus, M. R. (2011). Generalized linear mixed models for phylogenetic analyses of community structure. Ecological Monographs, 81, 511–525. [Google Scholar]
- Jackson, A. L. , & Loeb, L. A. (2001). The contribution of endogenous sources of DNA damage to the multiple mutations in cancer. Mutation Research/Fundamental and Molecular Mechanisms of Mutagenesis, 477, 7–21. [DOI] [PubMed] [Google Scholar]
- Jeannotte, L. , Lemieux, M. , Charron, J. , Poirier, F. , & Robertson, E. J. (1993). Specification of axial identity in the mouse: role of the Hoxa‐5 (Hox1. 3) gene. Genes & Development, 7, 2085–2096. 10.1101/gad.7.11.2085 [DOI] [PubMed] [Google Scholar]
- Jenkins, F. A. (1970). Anatomy and function of expanded ribs in certain edentates and primates. Journal of Mammalogy, 51, 288–301. [PubMed] [Google Scholar]
- Jouffroy, F. (1989). Quantitative and experimental approaches to primate locomotion: a review of recent advances. In Eds Seth P. & Seth S., Perspectives in Primate Biology (Vol 2, pp. 47–108). Today and Tomorrow's Printers & Publishers. [Google Scholar]
- Koteja, P. (1991). On the relation between basal and field metabolic rates in birds and mammals. Functional Ecology, 5(1), 56–64. [Google Scholar]
- Kuratani, S. (2004). Evolutionary Morphology: Bauplan and Embryonic Development of Vertebrates. University of Tokyo Press. [Google Scholar]
- Laclef, C. , Hamard, G. , Demignon, J. , Souil, E. , Houbron, C. , & Maire, P. (2003). Altered myogenesis in Six1‐deficient mice. Development, 130, 2239–2252. [DOI] [PubMed] [Google Scholar]
- Lanyon, J. M. , Newgrain, K. , & Sahir Syah Alli, T. (2006). Estimation of water turnover rate in captive dugongs (Dugong dugon) . Aquatic Mammals, 32(1), 103–108. [Google Scholar]
- Leung, H. W. , & Ballantyne, B. (1999). Developmental toxicity evaluation of rats dosed orally or cutaneously with octoxynol‐9. Journal of Applied Toxicology, 19, 267–273. [DOI] [PubMed] [Google Scholar]
- Makhoul, R. G. , & Machleder, H. I. (1992). Developmental anomalies at the thoracic outlet an analysis of 200 consecutive cases. Journal of Vascular Surgery, 16, 534–545. [PubMed] [Google Scholar]
- May‐Davis, S. (2017). Congenital malformations of the first sternal rib. Journal of Equine Veterinary Science, 49, 92–100. [Google Scholar]
- McNab, B. K. (1988). Complications inherent in scaling the basal rate of metabolism in mammals. The Quarterly Review of Biology, 63, 25–54. [DOI] [PubMed] [Google Scholar]
- McNab, B. K. (1978). Energetics of arboreal folivores: physiological problems and ecological consequences of feeding on an ubiquitous food supply. In Montgomery G. G. (Ed.), The ecology of arboreal folivores (pp. 153–162). Washington Smithsonian Institution Press. [Google Scholar]
- Mendel, F. C. (1985). Use of hands and feet of three‐toed sloths (Bradypus variegatus) during climbing and terrestrial locomotion. Journal of Mammalogy, 66, 359–366. [Google Scholar]
- Merks, J. H. M. , Smets, A. M. , van Rijn, R. R. , Kobes, J. , Caron, H. N. , Maas, M. , & Hennekam, R. C. M. (2005). Prevalence of rib anomalies in normal Caucasian children and childhood cancer patients. European Journal of Medical Genetics, 48, 113–129. [DOI] [PubMed] [Google Scholar]
- Mueller, E. F. (1983). Wärme und energiehaushalt bei Halbaffen (prosimiae). Bonner Zoologische Beiträge, 34, 29–71. [Google Scholar]
- Mueller, E. F. (1985). Basal metabolic rates in primates – the possible role of phylogenetic and ecological factors. Comparative Biochemistry and Physiology, 81A, 717–711. [DOI] [PubMed] [Google Scholar]
- Napier, J. R. , & Napier, P. H. (1967). A handbook of living primates. London and New York Academic Press. [Google Scholar]
- Narita, Y. , & Kuratani, S. (2005). Evolution of the vertebral formulae in mammals: a perspective on developmental constraints. Journal of Experimental Zoology Part B: Molecular and Developmental Evolution, 304(2), 91–106. [DOI] [PubMed] [Google Scholar]
- Narotsky, M. G. , Schmid, J. E. , Andrews, J. E. , & Kavlock, R. J. (1998). Effects of boric acid on axial skeletal development in rats. Biological Trace Element Research, 66, 373–394. [DOI] [PubMed] [Google Scholar]
- Owen, R. (1866). On the anatomy of vertebrates. Green, and Co Longmans. [Google Scholar]
- Paradis, E. , & Schliep, K. (2019). “Ape 5.0: an environment for modern phylogenetics and evolutionary analyses in R.” Bioinformatics, 35, 526–528. [DOI] [PubMed] [Google Scholar]
- Pauli, J. N. , Peery, M. Z. , fountain, E. D. , & Karasov, W. H. (2016). Arboreal folivores limit their energetic output, all the way to slothfulness. The American Naturalist, 188, 196–204. [DOI] [PubMed] [Google Scholar]
- Perez‐Campo, R. , López‐Torres, M. , Cadenas, S. , Rojas, C. , & Barja, G. (1998). The rate of free radical production as a determinant of the rate of aging: evidence from the comparative approach. Journal of Comparative Physiology B: Biochemical, Systemic, and Environmental Physiology, 168, 149–158. [DOI] [PubMed] [Google Scholar]
- Périquet, S. , Valeix, M. , Loveridge, A. J. , Madzikanda, H. , Macdonald, D. W. , & Fritz, H. (2010). Individual vigilance of African herbivores while drinking: the role of immediate predation risk and context. Animal Behaviour, 79, 665–671. [Google Scholar]
- Pinheiro, J. C. , & Bates, D. M. (2000). Mixed‐effects models in S and Splus. Springer. [Google Scholar]
- R Core Team . 2021. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/.
- Rancourt, D. E. , Tsuzuki, T. , & Capecchi, M. R. (1995). Genetic interaction between hoxb‐5 and hoxb‐6 is revealed by nonallelic noncomplementation. Genes & Development, 9, 108–122. [DOI] [PubMed] [Google Scholar]
- Rasmussen, D. T. , & Izard, M. K. (1988). Scaling of growth and life history traits relative to body size, brain size and metabolic rate in lorises and galagos (Lorisidae, Primates). American Journal of Physical Anthropology, 75, 357–367. [DOI] [PubMed] [Google Scholar]
- Rayan, G. M. (1988). Lower trunk brachial plexus compression neuropathy due to cervical rib in young athletes. The American Journal of Sports Medicine, 16, 77–79. [DOI] [PubMed] [Google Scholar]
- Reumer, J. W. F. , Ten Broek, C. M. A. , & Galis, F. (2014). Extraordinary incidence of cervical ribs indicates vulnerable condition in Late Pleistocene mammoths. PeerJ, 2, e318. 10.7717/peerj.318 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Revell, L. J. (2010). Phylogenetic signal and linear regression on species data. Methods in Ecology and Evolution, 1, 319–329. [Google Scholar]
- Rohlf, F. J. (2006). A comment on phylogenetic correction. Evolution, 60, 1509–1515. [DOI] [PubMed] [Google Scholar]
- Roos, D. B. (1976). Congenital anomalies associated with thoracic outlet syndrome. The American Journal of Surgery, 132, 771–778. [DOI] [PubMed] [Google Scholar]
- Roos, D. B. (1996). Historical perspectives and anatomic considerations. Seminars in Thoracic and Cardiovascular Surgery, 8, 183–189. [PubMed] [Google Scholar]
- Sargent, P. (1921). Lesions of the brachial plexus associated with rudimentary ribs. Brain, 44(2), 95–124. [Google Scholar]
- Schubert, F. R. , Tremblay, P. , Mansouri, A. , Faisst, A. M. , Kammandel, B. , Lumsden, A. , Gruss, P. , & Dietrich, S. (2001). Early mesodermal phenotypes insplotch suggest a role forPax3 in the formation of epithelial somites. Developmental Dynamics, 222, 506–521. [DOI] [PubMed] [Google Scholar]
- Schultz, A. 1961. Vertebral column and thorax. Basel: S. Karger.
- Schumacher, R. , Mai, A. , & Gutjahr, P. (1992). Association of rib anomalies and malignancy in childhood. European Journal of Pediatrics, 151, 432–434. [DOI] [PubMed] [Google Scholar]
- Schut, P. C. , Brosens, E. , Eggink, A. J. , Van Dooren, T. J. M. , Galis, F. , ten Broek, C. M. A. , Baijens, I. M. M. , Dremmen, M. H. G. , Tibboel, D. , Schol, M. P. , & Cohen‐Overbeek, T. E. (2020a). Exploring copy number variants in deceased fetuses and neonates with abnormal vertebral patterns and cervical ribs. Birth Defects Research, 112, 1513–1525. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schut, P. C. , Eggink, A. J. , Cohen‐Overbeek, T. E. , Van Dooren, T. J. M. , de Borst, G. J. , & Galis, F. (2020b). Miscarriage is associated with cervical ribs in thoracic outlet syndrome. Early Human Development, 144, 105027. [DOI] [PubMed] [Google Scholar]
- Self, S. G. , & Liang, K.‐Y. (1987). Asymptotic properties of maximum likelihood estimators and likelihood ratio tests under nonstandard conditions. Journal of the American Statistical Association, 82(398), 605–610. 10.2307/2289471 [DOI] [Google Scholar]
- Shapiro, L. J. (2007). Morphological and functional differentiation in the lumbar spine of lorisids and galagids. American Journal of Primatology, 69, 86–102. [DOI] [PubMed] [Google Scholar]
- Shigenawa, M. K. , & Ames, B. N. (1993). Oxidants and mitogenesis as causes of mutation and cancer: the influence of diet. In Bronzetti G. (Ed.), Antimutagenesis and anticarcinogenesis mechanisms III (pp. 419–436). Plenum press. [DOI] [PubMed] [Google Scholar]
- Simpson, G. M. , Fuller, G. , Lukas, K. E. , Kuhar, C. W. , Fitch‐Snyder, H. , Taylor, J. , & Dennis, P. M. (2018). Sources of morbidity in lorises and pottos in North American zoos: A retrospective review, 1980‐2010. Zoo Biology, 37, 245–257. [DOI] [PubMed] [Google Scholar]
- Slijper, E. J. (1946). Comparative Biologic‐Anatomical investigations on the vertebral column and spinal musculature of mammals. Verhandelingen der Koninklijke Nederlandsche Akademie van Wetenschappen, Afd. Natuurkunde. Tweede Sectie, Deel, 42(nr 5), 128. [Google Scholar]
- Starck, D. (1979). Vergleichende Anatomie der Wirbeltiere. Springer Verlag. [Google Scholar]
- Streissler‐Graz, E. (1913). Die Halsrippen. Ergebnisse der Chirurgie und Orthopädie, 5280–5355. [Google Scholar]
- Takihara, Y. , Tomotsune, D. , Shirai, M. , Katoh‐Fukui, Y. , Nishii, K. , Motaleb, M. A. , Nomura, M. , Tsuchiya, R. , Fujita, Y. , Shibata, Y. , Higashinakagawa, T. , & Shimada, K. (1997). Targeted disruption of the mouse homologue of the Drosophila polyhomeotic gene leads to altered anteroposterior patterning and neural crest defects. Development, 124, 3673–3682. [DOI] [PubMed] [Google Scholar]
- Ten Broek, C. M. A. , Bakker, A. J. , Varela‐Lasheras, I. , Bugiani, M. , Van Dongen, S. , & Galis, F. (2012). Evo‐devo of the human vertebral column: on homeotic transformations, pathologies and prenatal selection. Evolutionary Biology, 39, 456–471. 10.1007/s11692-012-9196-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thompson, T. (1908). Familial atropathy of the hand muscles. Brain, 31, 286–300. [Google Scholar]
- Tollis, M. , Schneider‐Utaka, A. K. , & Maley, C. C. (2020). The evolution of human cancer gene duplications across mammals. Molecular Biology and Evolution, 37, 2875–2886. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Toyokuni, S. , Okamoto, K. , Yodoi, J. , & Hiai, H. (1995). Persistent oxidative stress in cancer. FEBS Letters, 358, 1–3. [DOI] [PubMed] [Google Scholar]
- Valeix, M. , Fritz, H. , Loveridge, A. J. , Davidson, Z. , Hunt, J. E. , Murindagomo, F. , & Macdonald, D. W. (2009). Does the risk of encountering lions influence African herbivore behaviour at waterholes? Behavioral Ecology and Sociobiology, 63, 1483–1494. [Google Scholar]
- Varela‐Lasheras, I. , Bakker, A. J. , van der Mije, S. D. , Metz, J. A. , van Alphen, J. , & Galis, F. (2011). Breaking evolutionary and pleiotropic constraints in mammals: On sloths, manatees and homeotic mutations. EvoDevo, 2(1), 11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walker, A. (1969). The locomotion of the lorises, with special reference to the Potto. African Journal of Ecology, 7(1), 1–5. [Google Scholar]
- Weber, F. P. 1913. Bilateral cervical ribs with unilateral (right‐sided) atrophy of hand muscles. Proceedings Royal Society of Medicine 6 (Clinical Section), 52–54. [DOI] [PMC free article] [PubMed]
- Whittow, G. G. , Scammell, C. C. , Manuel, J. K. , Rand, D. , & Leong, M. (1977). Temperature regulation in a hypometabolic primate, the slow loris (Nycticebus coucang). Archives Internationales de Physiologie et de Biochimie, 85, 139–151. [DOI] [PubMed] [Google Scholar]
- Williams, S. A. , & Russo, G. A. (2015). Evolution of the hominoid vertebral column: The long and the short of it: Evolution of the hominoid vertebral column. Evolutionary Anthropology: Issues, News, and Reviews, 24, 15–32. [DOI] [PubMed] [Google Scholar]
- Woolfenden, G. E. (1961). Postcranial osteology of the waterfowl. Biological Sciences, 6, 1–129. [Google Scholar]
- Zeh, D. R. , Heupel, M. R. , Hamann, M. , Jones, R. , Limpus, C. J. , & Marsh, H. (2018). Evidence of behavioural thermoregulation by dugongs at the high latitude limit to their range in eastern Australia. Journal of Experimental Marine Biology and Ecology, 508, 27–34. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The data and the R script used for analysis are accessible via the Zenodo.org platform at https://doi.org/10.5281/zenodo.72210911, under licence CC BY‐NC‐SA 4.0.