

Abstract
Taste buds consist of at least three principal cell types that have different functions in processing gustatory signals — glial-like Type I cells, Receptor (Type II) cells, and Presynaptic (Type III) cells. Using a combination of Ca2+ imaging, single cell RT-PCR, and immunostaining, we show that γ-amino butyric acid (GABA) is an inhibitory transmitter in mouse taste buds, acting on GABA-A and GABA-B receptors to suppress transmitter (ATP) secretion from Receptor cells during taste stimulation. Specifically, Receptor cells express GABA-A receptor subunits β2, δ, π, as well as GABA-B receptors. In contrast, Presynaptic cells express the GABA-Aβ3 subunit and only occasionally GABA-B receptors. In keeping with the distinct expression pattern of GABA receptors in Presynaptic cells, we detected no GABAergic suppression of transmitter release from Presynaptic cells. We suggest that GABA may serve function(s) in taste buds in addition to synaptic inhibition. Finally, we also defined the source of GABA in taste buds: GABA is synthesized by GAD65 in Type I taste cells as well as by GAD67 in Presynaptic (Type III) taste cells and is stored in both those two cell types. We conclude that GABA is released during taste stimulation and possibly also during growth and differentiation of taste buds.
Introduction
Mammalian taste buds contain three morphologically and functionally distinct cell types (reviewed, Chaudhari and Roper, 2010). Type I cells appear to be supporting or glial-like cells (Bartel et al., 2006;Dvoryanchikov et al., 2009). Some of the Type I cells may also may play a role in salt (Na+) taste (Vandenbeuch et al., 2008;Chandrashekar et al., 2010). Type II (Receptor) cells are the primary detectors of sweet, bitter, and umami compounds; they express G protein-coupled taste receptors and effectors for these taste stimuli (Perez et al., 2002;Zhao et al., 2003;Clapp et al., 2004;DeFazio et al., 2006). Type III (Presynaptic) cells detect sour tastants. Presynaptic cells also are the only taste bud cells showing well-differentiated synapses and expressing synaptic proteins (Yee et al., 2001;DeFazio et al., 2006).
During taste stimulation and following the primary transduction response, the different types of cells in the taste bud interact and process gustatory signals via chemical signaling intrinsic to the taste bud. Taste stimulation triggers Receptor cells to secrete ATP and Presynaptic cells to release serotonin (5-HT) and norepinephrine (NE) (Dvoryanchikov et al., 2007;Huang et al., 2007;Romanov et al., 2007;Huang et al., 2008). ATP appears to be a transmitter between Receptor cells and primary afferent nerve fibers (Finger et al., 2005;Huang et al., 2007;Romanov et al., 2007). Both 5-HT and ATP play critical roles in cell-to-cell signaling within the taste bud, establishing positive- and negative feedback circuits that shape the afferent signal and may contribute to the coding of sensory information (Roper, 2007;Huang et al., 2009). Other transmitters such as glutamate and acetylcholine also serve in cell-cell communication within the taste bud (Ogura et al., 2007;Vandenbeuch et al., 2010). Additionally, cholecystokinin and neuropeptide Y may function in this capacity (Herness and Zhao, 2009).
In addition to the above transmitters, there is evidence that an inhibitory amino acid transmitter, γ-aminobutyric acid (GABA), figures in taste buds. Early immunocytochemical and autoradiography data revealed GABA in taste cells and gustatory nerve endings in amphibians and rodents (Jain and Roper, 1991;Obata et al., 1997;Nagai et al., 1998). Electrophysiological recordings from sensory ganglion cells that innervate taste buds showed that GABA mainly produces hyperpolarizing responses when applied to the cell body (Koga and Bradley, 2000). This was interpreted as a possible role for GABA as an afferent taste transmitter at the central and/or peripheral sensory endings of these ganglion cells. More recently, patch-clamp recordings have shown GABA hyperpolarizes cells in rat taste buds (Cao et al., 2009). Those workers proposed that GABA is involved in cell-to-cell communication within taste buds. Responses to GABA can be produced via ionotropic (GABA-A) and metabotropic (GABA-B) receptors. In different cells, responses to GABA may vary depending on the intracellular concentration of Cl−, the particular receptor subunits expressed, and the signaling pathways within cells.
Our understanding of the role of GABA in taste buds is very limited. The specific taste cells that synthesize and secrete this transmitter and the cells that respond to GABA in cell to cell communication are presently very incompletely defined. Importantly, the influence of GABA signaling on the sensory signal itself remains unexplored. Here, we begin to address these questions, focusing on the origin and cellular targets of GABA and its functional effects on the taste-evoked signal.
Material and Methods
Animals and Tissues
Adult mice of both sexes were used in this study, including C57BL/6J (wild type) mice and mice from two transgenic strains. In PLCβ2-GFP mice, GFP is expressed in over 95% of all PLCβ2-expressing (i.e. Receptor) cells (Kim et al., 2006); while in GAD1-GFP mice, GFP fluorescence is detected in ≈75% of Presynaptic cells of taste buds (Chattopadhyaya et al., 2004;Tomchik et al., 2007). Mice were killed by CO2 asphyxiation following National Institutes of Health guidelines, with procedures approved by the University of Miami Animal Care and Use Committee. Taste buds were obtained from lingual and palatal epithelia by enzymatic digestion as outlined below.
Biosensor cells
Chinese hamster ovary (CHO) cells expressing P2X2/P2X3, 5-HT2c receptors, or both served as ATP, 5-HT, or dual biosensors, respectively (Huang et al., 2005;Huang et al., 2007). Biosensors were loaded with Fura-2 AM for Ca2+ imaging. We verified that Ca2+ mobilization in biosensors was neither directly elicited nor altered by bath-applied KCl (up to 140 mM), by the taste stimuli used in this study, or by any of the GABA agonists and antagonists (Supplementary Figure S1).
Buffers, drugs and stimuli
During the dissection and recording, unless otherwise specified, we bathed tissues and cells in Tyrode's solution, composed of (in mM): 140 NaCl, 5 KCl, 2 CaCl2, 1 MgCl2, 10 HEPES, 10 glucose, 10 Na-pyruvate, 5 NaHCO3 (pH 7.2, 310-320 mOsm). Muscimol, baclofen and bicuculline were purchased from Sigma. CGP55845 was purchased from Tocris Bioscience. The taste stimulus mix consisted of cycloheximide (10 μM), denatonium (1 mM), saccharin (2 mM), SC45647 (0.1 mM), i.e., two “bitter” and two “sweet” compounds, respectively.
Ca2+ imaging
We used two different preparations in this study: (a) dispersed taste buds and single cells removed from vallate epithelium and imaged with Fura-2, and (b) semi-intact taste buds embedded in lingual tissue slices and imaged with Calcium Green dextran using confocal microscopy.
Dispersed taste buds or isolated taste cells were prepared as we described previously (Huang et al., 2007). Taste buds or taste cells were transferred to a shallow recording chamber, loaded with 5 μM Fura-2 AM, and superfused with buffer. The ratio of F340/F380 was converted to approximate [Ca2+]i as described by Grynkiewicz et al. (1985) using a Fura-2 Calibration Kit (Invitrogen). Taste buds and/or cells were stimulated by bath-perfusion of taste mix, ATP, or KCl. All solutions were made in Tyrode's buffer adjusted to pH 7.2. Stimuli were bath-applied for 30 s after which the perfusion was returned to Tyrode's solution. All experiments were conducted at room temperature.
For functional imaging in the lingual slice preparation, vallate taste cells were iontophoretically loaded with Calcium Green Dextran (CaGD) in blocks of tissue containing the vallate papilla. Tissue was sliced and imaged as previously detailed (Caicedo et al., 2000;Dando and Roper, 2009). Lingual slices were perfused at a rate of 2 ml/min at room temperature with Tyrode buffer containing elevated Ca2+ (8 mM) to improve signal/noise. Taste stimuli were focally applied to the taste pore for 2 sec via a focal ‘puffer’ micropipette; KCl and pharmacological agents were bath-applied. Images were captured with a confocal microscope (Olympus Fluoview FVx) at 2 s intervals and responses are presented as changes in relative fluorescence, ΔF/F (i.e. [F − F0)]/F0).
In both of the above cell/tissue preparations, we identified Receptor (Type II) or Presynaptic (Type III) taste cells by the expression of GFP in PLCβ2-GFP mice (Receptor cells) or GAD1-GFP mice (Presynaptic cells), or physiologically as follows. Cells that responded to taste stimulation but not to KCl-depolarization were classified as Receptor cells; cells that responded to KCl depolarization but not taste stimulation were identified as Presynaptic cells (DeFazio et al., 2006;Tomchik et al., 2007). The two methods for identifying cell types were mutually consistent.
RT-PCR analysis
Total RNA was isolated following DNase I digestion using the Absolutely RNA Nanoprep kit (Stratagene, La Jolla, CA). First strand cDNA synthesis was with Superscript III (Invitrogen, Carlsbad, CA) and PCR (Taq Polymerase, Qiagen, Valencia, CA) as detailed previously (Dvoryanchikov et al., 2009). Dispersed taste buds were washed individually in Tyrode's buffer to remove contaminating, adherent, non-taste cells. For analyzing single cells, taste buds were dissociated and GFP-labeled or GFP-lacking individual taste cells were collected. RNA was extracted from individual cells or from pools of 10 similar cells (e.g. GFP-positive). The RNA was subjected to T7 linear RNA amplification (Message BOOSTER cDNA kit for qPCR, Epicentre, Madison, WI) and the resulting cDNA was used in PCR to assess expression as previously detailed (Dvoryanchikov et al., 2009). PCR primers were designed for each of 19 GABA-A subunits currently recognized (Olsen and Sieghart, 2008). PCR primers and conditions are listed in Supplementary Table 1.
Quantitative RT-PCR was carried out on a BioRad iCycler (Hercules, CA) using cDNA from one taste bud in each reaction (Dvoryanchikov et al., 2009). SNAP25 mRNA in each tissue sample was used as a reference for normalization.
Immunostaining
Taste tissues were dissected from mice after perfusion-fixation with 4% paraformaldehyde and were processed for immunostaining as described (Dvoryanchikov et al., 2009). Antibodies used were rabbit anti-GABA (Sigma Chemical A2052, 1:1500), rabbit anti-GAD65 (Millipore AB5082, 1:500), rabbit anti-PLCβ2 (Santa Cruz Biotechnology, SC-1488, 1:1000), rabbit anti-NTPDase2 (J-P Sevigny, Univ. Laval, Canada, #N-1082, 1:1000) and guinea pig anti-GABA-B1 receptor (Millipore AB2256, 1:500). Secondary antibodies were goat anti-rabbit IgG-Alexa 488 (A-11034); goat anti-rabbit IgG-Alexa 568 (A-11011); goat anti-rabbit IgG Alexa 594 (A-11012); or goat anti-guinea pig IgG Alexa 594 (A-11076), all from Invitrogen, Carlsbad, CA. When two primary antibodies to different antigens were both raised in rabbit, we prelabeled one of the antibodies with Alexa 568 or Alexa 647 using a Zenon IgG labeling kit (Z-25306 or Z-25308, Invitrogen). For GABA-B1 immunostaining, antigen retrieval was performed in citrate buffer (Borg Decloaker RTU, BD1000 MM, Biocare Medical, Concord, CA) at 95°C for 30 min. Negative control sections incubated without primary antibody were included in each experiment.
Fluorescent and brightfield (with Nomarski differential interference contrast optics) micrographs were captured either on a Zeiss LSM510 or an Olympus Fluoview FV1000 confocal microscope. All images included in a given figure were adjusted in parallel for brightness and contrast.
Results
We examined whether and how GABA affected taste cells isolated from mouse vallate papillae. Isolated taste cells were identified as Receptor (Type II) or Presynaptic (Type III) cells based on their responses to taste stimulation and KCl depolarization, and on their green fluorescence if obtained from PLCβ2-GFP or GAD1-GFP transgenic mice (see Methods). Bath-applied GABA (10 μM) did not evoke Ca2+ responses in isolated Receptor cells (0 of 6) but did elicit modest changes of cytoplasmic Ca2+ in a minority of single Presynaptic cells (6 of 29). We reasoned, however, that if GABA was an inhibitory neurotransmitter, as is the case elsewhere in the nervous system and described for rat taste cells by Cao et al. (2009), one might not necessarily observe significant GABA-evoked Ca2+ mobilization (as indeed the case, shown below). Thus, we turned to other measures of taste bud function to measure the action of GABA.
GABA inhibits transmitter release from taste buds
Using highly-sensitive biosensors, we have previously shown that gustatory stimulation elicits 5-HT release from intact taste buds removed from lingual epithelium (Huang et al., 2005). Using that same methodology, we now asked whether GABA alters taste-evoked 5-HT secretion. Indeed, bath-applied GABA (10 μM) markedly suppressed the 5-HT release that was triggered when dispersed taste buds were stimulated with tastants (Figure 1). To classify the GABA receptor types involved, we repeated the experiments with muscimol and baclofen, agonists at ionotropic (GABA-A) or metabotropic (GABA-B) receptors, respectively (Figure 1 B,C). Each agonist (at 1 μM) significantly inhibited 5-HT secretion, suggesting that both GABA-A and −B receptors are present in taste buds (Figure 1 D). There was no significant difference between the depression produced by GABA, muscimol and baclofen (ANOVA with post-hoc Newman-Keuls multiple comparison test).
Figure 1. GABA inhibits taste-evoked serotonin release from mouse taste buds.

Taste buds were isolated from vallate papillae and recorded with biosensors to detect release of serotonin (5-HT). A, stimulating an isolated taste bud with a bitter-sweet taste mixture (arrows) containing cycloheximide (10 μM), denatonium (1 mM), saccharin (2 mM), and SC45647 (0.1 mM) elicited 5-HT secretion, as shown by the pronounced biosensor response (5-HT-bio]. 5-HT secretion was recorded as Δ[Ca2+]i in the serotonin biosensor, measured in nM (see Methods). Repeating the taste stimulation on the same taste bud in the presence of 10 μM GABA (shaded region, middle trace) showed a marked reduction of 5-HT secretion which was reversed upon washout of GABA. [GABA did not directly affect 5-HT biosensors, data not shown]. B, similar results were obtained by applying the GABA-A receptor agonist muscimol (1 μM), or C, the GABA-B receptor agonist baclofen. D, summary of data from several experiments such as shown in A-C. Taste-evoked serotonin release in the presence of GABA agonists was normalized to serotonin release under control conditions for the same taste bud / biosensor pair (paired Student's t-test; *, p<0.05; ** p<0.01). Bars show mean ± s.e.m.; mus, muscimol (1 μM); bac, baclofen (1 μM); be, control before drug application; af, after washout of drugs.
Taste stimulation also triggers ATP secretion from taste buds (Finger et al., 2005;Huang et al., 2007;Romanov et al., 2007). Thus, we tested whether GABA inhibits taste-evoked ATP release. We used ATP-biosensors to monitor ATP release from dispersed taste buds and found that GABA also significantly reduced taste-evoked ATP secretion (Figure 2A). Furthermore, both muscimol and baclofen were effective inhibitors of ATP secretion (Figure 2B,C), reinforcing the notion that GABA-A and −B receptors influence transduction and/or transmission pathways in mouse taste buds. The inhibitory action of muscimol was blocked by the GABA-A selective antagonist, bicuculline (10 μM) (Figure 2D). Conversely, the action of baclofen was antagonized by the GABA-B selective blocker, CGP55845 (10 μM) (Figure 2E). These findings confirm that GABA, muscimol, and baclofen act specifically on GABA-A and GABA-B receptors in mouse taste cells and are not depressing taste cell responses non-selectively.
Figure 2. GABA inhibits taste-evoked ATP release from taste buds.

Taste stimulation as in Figure 1 evokes ATP release that is reduced by 10 μM GABA (A), 1 μM muscimol (B), or 1 μM baclofen (C). Traces in A-C show responses from an ATP biosensor that was closely apposed to an isolated taste bud (3 different experiments). Arrows show application of taste mixture, gray shaded area shows bath-perfusion with agonists. D, E, inhibition of ATP release by muscimol or baclofen is eliminated by specific antagonists to the respective GABA receptors. Thus, 10 μM bicuculine, a GABA-A receptor antagonist, rescues taste-evoked ATP secretion even in the presence of muscimol (D), and 10 μM CGP55845, a GABA-B receptor antagonist, restores ATP release that was blocked by baclofen. Lightly shaded areas in D,E show bath-application of the GABA agonist, darker shaded area shows perfusion of agonist plus antagonist. Data in D,E verify that the GABA agonists were indeed acting on specific GABA receptors, not generally depressing taste bud cells. F, summary of data from several experiments as in A-E. Bars show means ± s.e.m. of ATP biosensor responses, normalized as in Figure 1. Paired Student's t-test compared each taste-evoked response in the presence of GABAergic drugs to the control evoked response in the same taste bud / biosensor pair (paired Student's t-test; ns, no significant difference; **, p<0.01). N, numbers of taste buds tested; mus, muscimol; bic, bicuculline; bac, baclofen; CGP, CGP55845.
Does GABA inhibit Receptor cells, Presynaptic cells, or both?
The above experiments were conducted on dispersed, but otherwise intact taste buds. To identify which taste cells are directly inhibited by GABA, we dissociated taste buds into isolated cells and used biosensors to monitor transmitter secretion from cells identified by type. GABA, muscimol, and baclofen significantly decreased taste-evoked ATP release from Receptor cells (Figure 3 A,B). ATP release was inhibited even though none of the agents (GABA, muscimol or baclofen) altered taste-evoked Ca2+ mobilization in Receptor cells. In contrast, transmitter release from Presynaptic cells (i.e., 5-HT secretion) was not directly affected by either GABA or GABA agonists (Figure 3C,D).
Figure 3. Muscimol and baclofen inhibit taste-evoked transmitter release from Receptor (Type II) but not Presynaptic (Type III) taste bud cells.

Traces in A, C, show recordings from an isolated Receptor cell (A) or Presynaptic cell (B) and its closely-apposed biosensor for detecting transmitter release. A, upper trace, recordings from a Receptor cell; lower trace, ATP biosensor. Repeated taste stimulation (arrows) produces responses in Receptor cells and triggers ATP secretion. ATP secretion is significantly and reversibly reduced in presence of 1 μM muscimol or 1 μM baclofen (shaded areas). B, summary of data from several experiments such as shown in A. Upper bars (black) show mean ± s.e.m. for Receptor cell responses; lower bars, concurrent measurements of ATP release (mean ± s.e.m.). Normalization, analyses and labels as in Figure 1. GABA reduced ATP release (to 34 ± 7%, N=3, not shown) similar to that seen for muscimol or baclofen. C, Sequential responses of a paired Presynaptic cell and 5-HT biosensor to repeated stimulation with 1 μM ATP (arrows). The Presynaptic cell repetitively releases 5-HT and this release is unaffected by muscimol or baclofen (shaded areas). D, summary of several experiments on Presynaptic cells, similar to summary shown in B for Receptor cells.
We infer from these results on isolated taste cells that in intact, dispersed taste buds, taste-evoked transmitter secretion (both ATP and 5-HT, Figures 1,2) is inhibited primarily by the actions of GABA on Receptor cells, as follows:
Consistent with this interpretation, GABA did not reduce 5-HT secretion from taste buds that were stimulated with ATP (Figure 4A), even though taste-evoked 5-HT release in the same taste bud was inhibited (Figure 4B,C).
Figure 4. In isolated taste buds, GABA interrupts communication between Receptor cells and Presynaptic cells.

Isolated taste bud were sequentially depolarized with KCl, stimulated with tastants and with ATP, in the absence or presence of 10 μM GABA. A, Traces show stimulus-evoked 5-HT release from Presynaptic cells within the taste bud (i.e., responses from a 5-HT biosensor apposed to the taste bud). All stimuli (arrows) elicit 5-HT release as shown previously (Huang et al., 2007). GABA has no effect on ATP-evoked 5-HT release. B, same taste bud, showing effects of GABA on taste-evoked 5-HT release. Here, Presynaptic cells are indirectly triggered to release 5-HT and GABA-mediated inhibition is consistent with GABA acting to reduce ATP release from Receptor cells, thereby interfering with cell-cell excitation of Presynaptic cells. C, summary of several experiments as those in A,B. Details of bars and analyses as in Figure 1.
Taste Receptor and Presynaptic cells express distinct GABA receptors
As an independent confirmation of GABAergic mechanisms in mouse taste buds, we used RT-PCR on cleanly isolated taste buds to evaluate the expression of all the known subunits of GABA-A and GABA-B receptors. Taste buds abundantly express mRNAs for several of GABA-A subunits, particularly β1, β2, β3, δ, and π, as well as for an obligate subunit of GABA-B receptors, GABA-B1 (Figure 5A,D). mRNAs for the remaining GABA-A subunits (α1, α2, α3, α4, α5, α6, γ1, γ2, γ3, ρ1, ρ2, ρ3, ε and θ) were either undetectable or appeared at very low levels and inconsistently across taste bud samples (Supplementary Figure S2). GABA receptor subunits were not expressed to any appreciable level in the non-taste epithelium adjacent to taste-buds (Figures 5A, S2).
Figure 5. Mouse taste buds express distinct GABA-A and GABA-B receptors in Receptor and Presynaptic cells.

A, RT-PCR on dissected vallate papillae (va), two preparations of cleanly isolated vallate taste buds (tb1, tb2), enzymatically peeled non-taste lingual epithelium (nt), and in parallel, brain (br) and no cDNA (-) control reactions. Expression was tested for several GABA-A receptor and one GABA-B receptor subunits. RT-PCR for β-actin and the taste-specific marker PLCβ2 on the same samples validated RNA quality and the presence of taste buds only in the first three samples. Data on additional GABA-A subunits are presented in Supplentary Figure S2. B, RT-PCR on pools of isolated Receptor (Type II) and Presynaptic cells (see Methods). Three pools of each class were validated by expression of either PLCβ2 or SNAP25 and then were tested for expression of the GABA receptor subunits shown in A. [Although GABA-A β1 is expressed in a taste-selective manner (see A), expression was not sufficiently high to permit detection in a few isolated cells]. Control PCRs are as in A. C, summary of data from expression profiles of Receptor and Presynaptic cells in B. Incidence of GABA receptor subunits is represented in a modified, overlapping Venn diagram. For example, all 3 pools of Receptor cells but only 1 of 3 pools of Presynaptic cells expressed GABA-B receptors (blue). Receptor cells expressed several different GABA-A subunits (warm colors), while Presynaptic cells express only the β3 isoform (see text for details). D, Immunostaining with anti-GABA-B1 shows that all Receptor cells (visualized as immunopositive for PLCβ2, green) co-express GABA-B1 receptors (red). Scale, 20 μM.
To assess which taste cell types express this limited set of GABA receptors, we tested cDNAs from Receptor cells (3 pools, each pool containing 10 cells) and Presynaptic cells (also 3 pools, each with 10 cells). These pools were obtained by harvesting GFP-positive taste cells after isolating and dissociating taste buds from PLCβ2-GFP mice (Receptor cells) or GAD67-GFP mice (Presynaptic cells). The homogeneity of each pool of cells was verified by RT-PCR for two diagnostic genes: PLCβ2 (for Receptor cells) and SNAP25 (for Presynaptic cells). Receptor cell pools consistently expressed GABA-B1. In addition, GABA-A subunits β2, δ, and π were detected in some but not all pools of Receptor cells. Of the three pools of Presynaptic cells, two expressed GABA-A β3; one of these also expressed GABA-B1 (Figure 5B). When expression levels for a particular gene are low, the mRNA is inconsistently detected by single-cell RT-PCR. Thus, the uneven pattern of expression of GABA-A subunits in both cell types may reflect either low mRNA abundance or heterogeneity in the particular subunits expressed in each cell. GABA-B subunits, on the other hand, were detected in all Receptor cell pools, suggesting that most Receptor cells express the metabotropic receptor for GABA. These RT-PCR data on the expression pattern of GABA subunits are summarized in Figure 5C.
We also carried out immunofluorescence on mouse taste tissue for the one subunit most consistently detected by RT-PCR, GABA-B1. Consistent with our single-cell RT-PCR data, we detected GABA-B1 in all Receptor cells, and also at lower intensity in additional, unidentified cells (arrow, Figure 5D).
In summary, Receptor cells express GABA-A and GABA-B receptor subunits, consistent with the calcium imaging data. GABA-B subunits are prominently expressed in Receptor cells but are also seen, though less frequently, in Presynaptic cells. GABA-A subunits are expressed at lower levels, or in subsets of cells: Receptor cells heterogeneously express β2, δ and π, while Presynaptic cells express β3 (Figure 5C).
GABA inhibits cell-cell communication in the lingual slice preparation
As a further investigation of GABAergic inhibition in mouse taste buds, and to verify the above findings in a more intact preparation, we tested whether GABA affects taste-evoked responses in lingual slices. For this, we recorded taste- and depolarization-evoked responses using confocal Ca2+ imaging. Receptor and Presynaptic taste cells were identified by their Ca2+ responses to a tastant mix or to KCl, as above (see (DeFazio et al., 2006;Tomchik et al., 2007). Consistent with the data from isolated taste cells (above), taste-evoked responses in Receptor cells were unaltered by bath-applied GABA (100 μM) (Figure 6A,B). However, Ca2+ responses in Presynaptic cells (i.e. “downstream” of Receptor cells) were significantly reduced by GABA (Figure 6C,D). These data strongly support the interpretation that GABA inhibits taste-triggered and ATP-mediated communication from Receptor to Presynaptic cells.
Figure 6. When tested in a semi-intact prepration, GABA inhibits communication between Receptor and Presynaptic taste bud cells.

Traces show Ca imaging from vallate taste cells in a lingual slice preparation. Receptor cells (A, B) were identified as responding to taste mixture but not 50mM KCl. Presynaptic cells (C,D) were identified as responding to both the taste mixture and KCl. A, focally applied taste mixture (arrow) evoked responses in a Receptor cell. Taste-evoked Receptor cell Ca responses were not significantly affected by bath-application of 100 μM GABA (shaded area). B, summary of data from 5 experiments as in A. Bars are means ± s.e.m., responses normalized to the group mean for taste responses before (be) GABA. C, in contrast, 100 μM GABA significantly reduces taste-evoked responses recorded from a Presynaptic cell. D, summary of 6 experiments such as shown in C. Details of bars as in B. Calibrations in A,C, 100 sec, 0.1 ΔF/F0
GABA is synthesized in Glial-like (Type I) and Presynaptic (Type III) taste bud cells
We next investigated potential sources of GABA in mouse taste buds. In neurons, GABA is synthesized by Glutamic Acid Decarboxylase (GAD), which exists as one of two isoforms--GAD65 (GAD2) or GAD67 (GAD1) (Soghomonian and Martin, 1998). Using RT-PCR, we found that both GAD65 and GAD67 are abundant in taste buds across the oral cavity, including vallate, foliate, fungiform and palate. In contrast, non-taste lingual epithelium did not express GAD65 at all, while GAD67 was detectable in some samples at low abundance (Figure 7A). The presence of GAD67 mRNA in non-taste samples may derive from small amounts of contaminating Von Ebner's glands, which have been reported to express GAD67 (Kosuge et al., 2009). With real-time RT-PCR, we found that in taste buds, both the synthetic enzymes are expressed at levels similar to SNAP25, a prominent mRNA selectively expressed in taste buds (Supplementary Figure S3).
Figure 7. GABA is synthesized and accumulates in both Type I and Presynaptic (Type III) taste bud cells.

A, RT-PCR for glutamic acid decarboxylase (GAD) subtypes in enzymatically peeled epithelium containing taste buds from four oral taste fields, vallate (va), foliate (fo), fungiform (fu) and palate (pa). Both GAD65 and GAD67 are expressed in all fields and little, if at all, in peeled non-taste lingual epithelium (nt). Control reactions include no cDNA (-) and brain cDNA (br). B, taste bud cells were subjected to single-cell RT-PCR, first to test for expression of NTPDase2, PLCβ2 and SNAP25. Of > 50 cells tested, none expressed more than one of these three cell-type markers. The expression of GAD67 and GAD65 in each cell was then tested. The gel shows three representative cells of each type. No expression of GAD65 or GAD67 was detected in Receptor cells (Plcβ2+). C, summary of single cell profiling of 45 cells as in B, shows that GAD65 is only expressed in Type I (Glial-like) cells, while GAD67 is only expressed in Presynaptic cells. Receptor (Type II) cells do not express either GABA-synthesizing enzyme. C, D-G, double immunofluorescence for GABA or its synthetic enzymes, and markers for Type I, Receptor, and Presynaptic cells in mouse vallate taste buds. D, GAD65 immunoreactivity (red) is in cells that express NTPDase2 (green), a marker for Type I cells. E. GFP fluorescence (green) in taste buds from GAD67-GFP transgenic mice is in cells distinct from cells that are immunoreactive for GAD65 (red). F, GABA immunoreactivity (red) in vallate taste buds from a GAD67-GFP mouse occurs in cells displaying GFP fluorescence (green) as well as in cells that are immunoreactive for NTPDase2 (blue). That is, GABA accumulates only in GAD67-positive Type III cells and in NTPDase2-positive Type I cells. G, In contrast, GFP fluorescence (green) in taste buds from PLCβ2-GFP transgenic mice does not co-localize with GABA immunostaining, confirming that Receptor (PLCβ2+) cells do not synthesize or store GABA.
To examine which taste cells express GAD isoforms and to test if both GAD65 and GAD67 are present in the same cells, we employed single-cell RT-PCR and immunofluorescence microscopy. Individually isolated cells were classified according to their expression of NTPDase2 (glial like, Type I), PLCβ2 (Receptor, Type II), or SNAP25 (Presynaptic, Type III). Type I glial-like cells (10 of 17, i.e. ≈ 60%) prominently expressed GAD65 while only 1 of 18 (≈6%) Presynaptic cells were positive for GAD65 (Figure 7B,C). In contrast, only Presynaptic cells (15 of 18, i.e. 83%) expressed GAD67. None of the Receptor cells tested expressed either isoform of GAD. These expression patterns of GAD67 and GAD65 across the three cell types were significantly different from each other (Chi-square test, p ≤ 0.0002).
We extended our single-cell RT-PCR results on GAD isoforms using immunofluorescence microscopy. We have previously shown (DeFazio et al., 2006;Tomchik et al., 2007) that GAD67 is expressed only in Type III Presynaptic cells. As shown in Figure 7D, GAD65 immunoreactivity substantially co-localized with NTPDase2, a marker for Type I glial-like taste cells. We further examined GAD65 expression in taste buds from two well-characterized strains of transgenic mice, PLCβ2-GFP (in which Receptor cells are labeled) and GAD67-GFP, in which most Presynaptic cells are labeled. GAD65 was lacking from GFP-positive taste cells in both GAD67-GFP tissue (i.e. from Presynaptic cells, Figure 7E) and PLCβ2-GFP tissue (i.e. Receptor cells, data not shown). These observations are fully consistent with the non-overlapping distribution of GAD65 and GAD67 seen in our single-cell RT-PCR data (Figure 7B).
Does GABA accumulate in the cells that express GAD65 or GAD67? Using anti-GABA antibodies, we observed GABA-immunofluorescence in several cells in vallate, fungiform, and palate taste buds (Figure 7F,G; Supplementary Figure S4). Using taste buds in which Receptor or Presynaptic cells are marked with GFP, and by immunostaining for cell-type markers, we determined that GABA-immunoreactive cells were either NTPDase2-expressing (i.e. Type I) or GAD67-expressing (i.e. Presynaptic cells, Figure 7F). Receptor cells (PLCβ2-positive or PLCβ2-GFP) were never seen to accumulate GABA (Figure 7G), consistent with their lack of GABA-synthesizing enzymes. We noted that occasional GAD67-GFP-positive Presynaptic cells did not accumulate GABA; we did not explore this further. In vallate taste buds, there are relatively large numbers of Presynaptic cells (Dvoryanchikov et al., 2007) and GABA may be secreted from either Type I or Presynaptic cells. Because fungiform and palatal taste buds contain few Presynaptic cells (see Supplemental Figure S4),Type I glial-like cells may constitute the main stores of GABA in these taste buds.
Discussion
Results presented here document that GABA is an inhibitory transmitter in mouse vallate taste buds. A prominent action of GABA is to reduce transmitter (ATP) secretion from Receptor cells by acting on GABA-A and GABA-B receptors expressed by those cells. GABA is synthesized by and found in Type I (glial-like) and Presynaptic (Type III) taste cells. These findings suggest that GABA is an important taste bud transmitter, likely released from Type I and Presynaptic taste cells during gustatory activation of taste buds and modulating the signal output from these peripheral sensory organs (Figure 8).
Figure 8. Schematic diagram of GABAergic inhibition in taste buds.
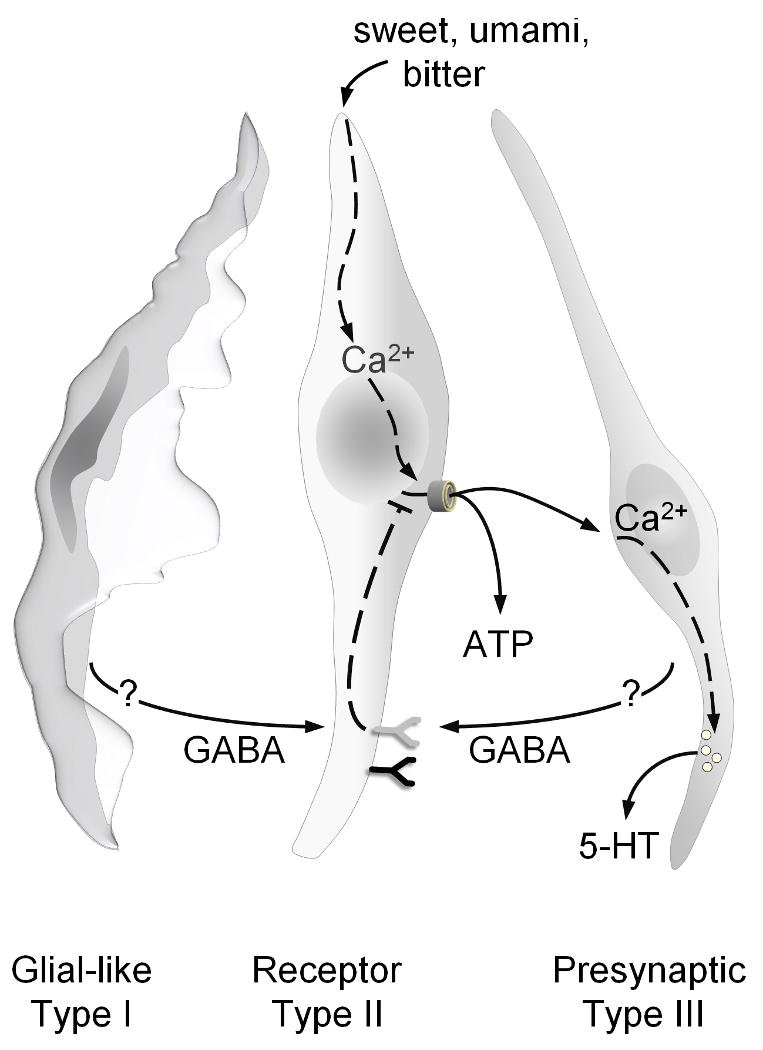
The three functional taste cell types are illustrated in grey (Type I glial-like), tan (Type II Receptor) and aqua (Type III Presynaptic). Sweet, bitter or umami stimuli evoke Ca2+ mobilization in Receptor cells, leading to ATP secretion. This ATP stimulates both afferent fibers (not shown) and adjacent Presynaptic cells that then secrete 5-HT. GABA, released from either Presynaptic or Type I cells or both, activates GABA-A and GABA-B receptors (light and dark green) on Receptor cells and reduces the taste-evoked secretion of ATP. The trigger(s) for GABA secretion and signaling events downstream of GABA receptor activation remain to be elucidated. For clarity, GABA receptors on non-Receptor cells are not shown.
ATP release from Receptor cells requires the combined actions of a membrane depolarization (mediated by TrpM5) concurrent with an increased intracellular Ca2+, mediated by IP3-triggered release from stores (Huang and Roper, 2010). These two factors act in concert to open Px1 gap junction hemichannels and allow ATP efflux. GABA presumably inhibits taste-evoked ATP secretion by hyperpolarizing Receptor cells (i.e., preventing TrpM5-mediated depolarization), an action not readily measured by Ca2+ imaging. This interpretation is entirely consistent with recent patch recordings in rat taste buds that showed that activating GABA receptors in taste cells increased membrane Cl− and K+ conductances, thereby stabilizing or even hyperpolarizing Receptor cells (Cao et al., 2009). Our study confirms some of the earlier findings and significantly extends them. Although Cao et al (2009) demonstrated the accumulation of GABA and the expression of GABA-A and -B receptors in rat taste buds, these entities were not specifically localized to functionally identified cellular populations. Importantly, because Cao et al (2009) did not discriminate GAD65 from GAD67, the two source cell populations for GABA were not apparent. The expression of GABAergic proteins in taste buds from mice and rats bear overall similarities. Importantly, our data now provide a more complete explanation for the role of GABA in taste buds—to inhibit taste-evoked transmitter secretion from Receptor cells. The significance of such inhibition in the intact system remains to be addressed. It is possible that GABAergic inhibition is employed in shaping responses to one or other of the taste qualities (sweet, bitter) in the mix we used. Alternatively, secreted GABA may tonically set the resting potential or responsivity of taste cells. In neurons, such tonic inhibition is mediated through extrasynaptic GABA-A receptors which typically include the δ subunit (Belelli et al., 2009). We note that we detected δ subunits in Receptor cells. Finally, because GABA may be secreted from Presynaptic or Type I cells (or both), it remains unclear which cell type is responsible for GABA secretion during taste stimulation. Our future experiments will address these questions.
Previous studies (Cao et al., 2009;Starostik et al., 2010) have examined GABA receptor subunits in taste tissues of rodents. Our semi-quantitative RT-PCR data suggest that expression levels for most GABA subunits are relatively or very low in taste cells. This may account for some of the apparent discrepancies across data in these three reports. Further, our comparisons between isolated taste buds and dissected taste papillae (Figures 5, S2) suggest that many of the GABA receptor subunits may be expressed in cells underneath the taste epithelium (e.g. nerves). Nevertheless, we and Starostik et al. (2010) both find that GABA-Aβ3 is a relatively abundant subunit in taste buds; all three reports show the presence of GABA-B receptors.
We note that muscimol, baclofen and GABA all produced similar levels of inhibition (Figures 1D, 3B). These results may suggest that signals from GABA-A and GABA-B receptors in taste cells converge on a common step that is upstream of the transmitter release. Thus, a saturating dose of any one agonist would produce maximum inhibition. Further, Receptor cells express both GABA-A and GABA-B subunits and ATP secretion from Receptor cells is the primary target of GABAergic inhibition (Figure 3A,B). That is, the taste cells that express GABA receptors prominently are the ones that are obviously affected by GABA.
Although both Receptor (Type II) and Presynaptic (Type III) cells express GABA-A receptors, we only detected GABAergic inhibition of transmitter secretion from Receptor cells. This may suggest that GABA has additional functions in mouse taste buds. For instance, Presynaptic cells express principally GABA-A β3 receptors. The β3 subunit has been shown to be important in the development of the palate (Hagiwara et al., 2003). Indeed, there is by now substantial evidence that GABA acts as a trophic factor and modulates cell proliferation and synaptic formation during neuronal development (Owens and Kriegstein, 2002). One might speculate that the action of GABA on Presynaptic cells is related to the development and maturation of these taste bud cells and their synapses, as is the case in the central nervous system (Wang and Kriegstein, 2009).
Our results in mice, combined with those of Cao et al (2009) in rats firmly establish GABA as an inhibitory transmitter in taste buds. However, the implication of this for how animals discriminate sweet, sour, salty, etc. and how taste behavior is affected by GABAergic mechanisms is unclear. Our data indicate that GABA is synthesized and stored in specific taste cells: GABA-biosynthetic enzymes are found in Type I glial-like cells (GAD65) and Presynaptic (Type III) cells (GAD67). While neurons have long been known to secrete GABA as a transmitter, recent evidence indicates that many glia also synthesize and secrete GABA (Jow et al., 2004) and indeed, GABA is recognized as a significant “gliotransmitter” (Angulo et al., 2008). Taste preference experiments have been conducted on GAD65 knockout mice (Shimura et al., 2004) but the findings have no straightforward interpretation. Namely, GAD65 KO mice did not differ from wild type mice in taste preferences for sucrose, NaCl, HCl, or quinine when these solutions were presented alone. However, GAD65 KO versus wild type mice responded differently to binary taste mixtures, notably sucrose plus quinine. The authors concluded that GABA (from GAD65) is not involved in basic taste discrimination, per se, but is instead involved in signal processing for more complex information such as taste mixtures. A major complication in interpreting these data is that knocking out GAD65 also interrupts GABAergic synapses in the central nervous system, not just GABA's actions (including trophic, if any) in taste buds. This may have profound effects on all behaviors, including taste preference and discrimination ability. Taste behavioral assays on GAD67 knockout mice have not been conducted; the genetic mutation is lethal at birth. In addition to the genetic studies, pharmacological investigations have revealed that GABAergic drugs, such as benzodiazepines, do indeed influence taste preferences (Cooper, 1989). Again, there is the caveat that these drugs are inevitably exerting powerful CNS actions. In short, given its inhibitory effects in taste buds, it is likely that GABA plays a distinct peripheral role in taste reception and signaling (in addition to the aforementioned role in development). However, pinpointing whether and how GABA is released from Type I glial-like cells, Presynaptic cells, or both during taste reception has not been undertaken and it remains to be determined what overall effects this inhibitory transmitter exerts in taste reception.
Supplementary Material
Figure S1. Biosensors do not respond to GABAergic agonists or antagonists. A. 5-HT biosensors show robust Ca2+ responses to bath-applied serotonin (3 nM), but not to GABA (10 μM), Bac (baclofen, 1 μM), Mus (muscimol, 1 μM), Bic (bicuculline, 10 μM), CGP (CGP55845, 10 μM). Similarly, responses to 5-HT itself were unaffected by these compounds (not shown). B. ATP biosensors respond to ATP (1 μM) but not to the GABAergic drugs as in A. Responses to ATP also were not affected by the presence of these drugs (not shown). For A and B alike, three representative, highly sensitive biosensor cells are shown.
Figure S2. RT-PCR for GABA receptor subunits that are expressed at low abundance or not at all in mouse taste buds. A, GABA-A receptor subunits α1, α5, ρ2 and γ3 are expressed in tissue dissected from the vallate papilla (va) and in some (but not all) samples of taste buds (tb1, tb2). The vallate sample contains muscle, connective tissue, and importantly, nerves. Thus, reaction product in some taste bud samples may represent either very low levels of expression in taste buds or contamination from other cells in the papilla. Controls as in Figures 5, 7 are non-taste lingual epithelium (nt), no cDNA (-), and brain (br). B, GABA-A receptor subunits α2, α4, γ2, θ, ε, and ρ3 are detectable only in vallate tissue pieces (va) and not in isolated taste buds (tb1, tb2). C, GABA-A receptor subunits α3, α6, γ1, and ρ1 were undetectable in all taste bud containing samples.
Figure S3. Relative expression of GAD 65 and GAD67 in vallate, foliate, fungiform, and palatal taste buds. Three independent samples of taste buds from each taste field were processed for qRT-PCR for for GAD65 (A), GAD67 (B) and SNAP25, a marker for taste bud cells. Bars are mean ± s.e.m. for relative expression of each GABA-synthetic enzyme, normalized to expression of SNAP25 in the same sample.
Figure S4. Immunoreactivity for GABA is also detected in mouse palatal and fungiform taste buds. Figure 7 in the manuscript demonstrates that GAD67-expressing (Presynaptic) taste cells in vallate taste buds accumulate GABA. Here we show similar results for taste buds from fungiform papillae and palate. GFP fluorescence (green) in taste cells from GAD67-GFP transgenic mice overlaps with, and is a subset of cells that are immunopositive for GABA (red). Scale, 20 μm.
Acknowledgments
This work was supported by NIH/NIDCD grants R01DC7630 (SDR), R01DC374 (SDR), and R01DC6308 (NC)
Footnotes
The authors have no conflicts of interest to declare
References
- Angulo MC, Le MK, Kozlov AS, Charpak S, Audinat E. GABA, a forgotten gliotransmitter. Prog Neurobiol. 2008;86:297–303. doi: 10.1016/j.pneurobio.2008.08.002. [DOI] [PubMed] [Google Scholar]
- Bartel DL, Sullivan SL, Lavoie EG, Sevigny J, Finger TE. Nucleoside triphosphate diphosphohydrolase-2 is the ecto-ATPase of type I cells in taste buds. J Comp Neurol. 2006;497:1–12. doi: 10.1002/cne.20954. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Belelli D, Harrison NL, Maguire J, Macdonald RL, Walker MC, Cope DW. Extrasynaptic GABAA receptors: form, pharmacology, and function. J Neurosci. 2009;29:12757–12763. doi: 10.1523/JNEUROSCI.3340-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caicedo A, Jafri MS, Roper SD. In situ Ca2+ imaging reveals neurotransmitter receptors for glutamate in taste receptor cells. J Neurosci. 2000;20:7978–7985. doi: 10.1523/JNEUROSCI.20-21-07978.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cao Y, Zhao FL, Kolli T, Hivley R, Herness S. GABA expression in the mammalian taste bud functions as a route of inhibitory cell-to-cell communication. Proc Natl Acad Sci U S A. 2009;106:4006–4011. doi: 10.1073/pnas.0808672106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chandrashekar J, Kuhn C, Oka Y, Yarmolinsky DA, Hummler E, Ryba NJ, Zuker CS. The cells and peripheral representation of sodium taste in mice. Nature. 2010;464:297–301. doi: 10.1038/nature08783. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chattopadhyaya B, Di CG, Higashiyama H, Knott GW, Kuhlman SJ, Welker E, Huang ZJ. Experience and activity-dependent maturation of perisomatic GABAergic innervation in primary visual cortex during a postnatal critical period. J Neurosci. 2004;24:9598–9611. doi: 10.1523/JNEUROSCI.1851-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chaudhari N, Roper SD. The cell biology of taste. J Cell Biol. 2010;190:285–296. doi: 10.1083/jcb.201003144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clapp TR, Yang R, Stoick CL, Kinnamon SC, Kinnamon JC. Morphologic characterization of rat taste receptor cells that express components of the phospholipase C signaling pathway. J Comp Neurol. 2004;468:311–321. doi: 10.1002/cne.10963. [DOI] [PubMed] [Google Scholar]
- Cooper SJ. Benzodiazepine receptor-mediated enhancement and inhibition of taste reactivity, food choice, and intake. Ann N Y Acad Sci. 1989;575:321–336. doi: 10.1111/j.1749-6632.1989.tb53253.x. [DOI] [PubMed] [Google Scholar]
- Dando R, Roper SD. Cell-to-cell communication in intact taste buds through ATP signalling from pannexin 1 gap junction hemichannels. J Physiol. 2009;587:5899–5906. doi: 10.1113/jphysiol.2009.180083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- DeFazio RA, Dvoryanchikov G, Maruyama Y, Kim JW, Pereira E, Roper SD, Chaudhari N. Separate populations of receptor cells and presynaptic cells in mouse taste buds. J Neurosci. 2006;26:3971–3980. doi: 10.1523/JNEUROSCI.0515-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dvoryanchikov G, Sinclair MS, Perea-Martinez I, Wang T, Chaudhari N. Inward rectifier channel, ROMK, is localized to the apical tips of glial-like cells in mouse taste buds. J Comp Neurol. 2009;517:1–14. doi: 10.1002/cne.22152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dvoryanchikov G, Tomchik SM, Chaudhari N. Biogenic amine synthesis and uptake in rodent taste buds. J Comp Neurol. 2007;505:302–313. doi: 10.1002/cne.21494. [DOI] [PubMed] [Google Scholar]
- Finger TE, Danilova V, Barrows J, Bartel DL, Vigers AJ, Stone L, Hellekant G, Kinnamon SC. ATP signaling is crucial for communication from taste buds to gustatory nerves. Science. 2005;310:1495–1499. doi: 10.1126/science.1118435. [DOI] [PubMed] [Google Scholar]
- Grynkiewicz G, Poenie M, Tsien RY. A new generation of Ca2+ indicators with greatly improved fluorescence properties. J Biol Chem. 1985;260:3440–3450. [PubMed] [Google Scholar]
- Hagiwara N, Katarova Z, Siracusa LD, Brilliant MH. Nonneuronal expression of the GABA(A) beta3 subunit gene is required for normal palate development in mice. Dev Biol. 2003;254:93–101. doi: 10.1016/s0012-1606(02)00030-1. [DOI] [PubMed] [Google Scholar]
- Herness S, Zhao FL. The neuropeptides CCK and NPY and the changing view of cell-to-cell communication in the taste bud. Physiol Behav. 2009;97:581–591. doi: 10.1016/j.physbeh.2009.02.043. [DOI] [PubMed] [Google Scholar]
- Huang YA, Dando R, Roper SD. Autocrine and paracrine roles for ATP and serotonin in mouse taste buds. J Neurosci. 2009;29:13909–13918. doi: 10.1523/JNEUROSCI.2351-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang YA, Maruyama Y, Roper SD. Norepinephrine is coreleased with serotonin in mouse taste buds. J Neurosci. 2008;28:13088–13093. doi: 10.1523/JNEUROSCI.4187-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang YA, Roper SD. Intracellular Ca(2+) and TRPM5-mediated membrane depolarization produce ATP secretion from taste receptor cells. J Physiol. 2010;588:2343–2350. doi: 10.1113/jphysiol.2010.191106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang YJ, Maruyama Y, Dvoryanchikov G, Pereira E, Chaudhari N, Roper SD. The role of pannexin 1 hemichannels in ATP release and cell-cell communication in mouse taste buds. Proc Natl Acad Sci U S A. 2007;104:6436–6441. doi: 10.1073/pnas.0611280104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang YJ, Maruyama Y, Lu KS, Pereira E, Plonsky I, Baur JE, Wu D, Roper SD. Mouse taste buds use serotonin as a neurotransmitter. J Neurosci. 2005;25:843–847. doi: 10.1523/JNEUROSCI.4446-04.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jain S, Roper SD. Immunocytochemistry of gamma-aminobutyric acid, glutamate, serotonin, and histamine in Necturus taste buds. J Comp Neurol. 1991;307:675–682. doi: 10.1002/cne.903070412. [DOI] [PubMed] [Google Scholar]
- Jow F, Chiu D, Lim HK, Novak T, Lin S. Production of GABA by cultured hippocampal glial cells. Neurochem Int. 2004;45:273–283. doi: 10.1016/j.neuint.2003.11.021. [DOI] [PubMed] [Google Scholar]
- Kim JW, Roberts C, Maruyama Y, Berg S, Roper S, Chaudhari N. Faithful expression of GFP from the PLCbeta2 promoter in a functional class of taste receptor cells. Chem Senses. 2006;31:213–219. doi: 10.1093/chemse/bjj021. [DOI] [PubMed] [Google Scholar]
- Koga T, Bradley RM. Biophysical properties and responses to neurotransmitters of petrosal and geniculate ganglion neurons innervating the tongue. J Neurophysiol. 2000;84:1404–1413. doi: 10.1152/jn.2000.84.3.1404. [DOI] [PubMed] [Google Scholar]
- Kosuge Y, Kawaguchi M, Sawaki K, Okubo M, Shinomiya T, Sakai T. Immunohistochemical study on GABAergic system in salivary glands. Eur J Pharmacol. 2009;610:18–22. doi: 10.1016/j.ejphar.2009.02.043. [DOI] [PubMed] [Google Scholar]
- Nagai T, Delay RJ, Welton J, Roper SD. Uptake and release of neurotransmitter candidates, [3H]serotonin, [3H]glutamate, and [3H]gamma-aminobutyric acid, in taste buds of the mudpuppy, Necturus maculosus. J Comp Neurol. 1998;392:199–208. [PubMed] [Google Scholar]
- Obata H, Shimada K, Sakai N, Saito N. GABAergic neurotransmission in rat taste buds: immunocytochemical study for GABA and GABA transporter subtypes. Brain Res Mol Brain Res. 1997;49:29–36. doi: 10.1016/s0169-328x(97)00118-6. [DOI] [PubMed] [Google Scholar]
- Ogura T, Margolskee RF, Tallini YN, Shui B, Kotlikoff MI, Lin W. Immuno-localization of vesicular acetylcholine transporter in mouse taste cells and adjacent nerve fibers: indication of acetylcholine release. Cell Tissue Res. 2007;330:17–28. doi: 10.1007/s00441-007-0470-y. [DOI] [PubMed] [Google Scholar]
- Olsen RW, Sieghart W. International Union of Pharmacology. LXX. Subtypes of gamma-aminobutyric acid(A) receptors: classification on the basis of subunit composition, pharmacology, and function. Update. Pharmacol Rev. 2008;60:243–260. doi: 10.1124/pr.108.00505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Owens DF, Kriegstein AR. Is there more to GABA than synaptic inhibition? Nat Rev Neurosci. 2002;3:715–727. doi: 10.1038/nrn919. [DOI] [PubMed] [Google Scholar]
- Perez CA, Huang L, Rong M, Kozak JA, Preuss AK, Zhang H, Max M, Margolskee RF. A transient receptor potential channel expressed in taste receptor cells. Nat Neurosci. 2002;5:1169–1176. doi: 10.1038/nn952. [DOI] [PubMed] [Google Scholar]
- Romanov RA, Rogachevskaja OA, Bystrova MF, Jiang P, Margolskee RF, Kolesnikov SS. Afferent neurotransmission mediated by hemichannels in mammalian taste cells. EMBO J. 2007;26:657–667. doi: 10.1038/sj.emboj.7601526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roper SD. Signal transduction and information processing in mammalian taste buds. Pflugers Arch. 2007;454:759–776. doi: 10.1007/s00424-007-0247-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shimura T, Watanabe U, Yanagawa Y, Yamamoto T. Altered taste function in mice deficient in the 65-kDa isoform of glutamate decarboxylase. Neurosci Lett. 2004;356:171–174. doi: 10.1016/j.neulet.2003.11.041. [DOI] [PubMed] [Google Scholar]
- Soghomonian JJ, Martin DL. Two isoforms of glutamate decarboxylase: why? Trends Pharmacol Sci. 1998;19:500–505. doi: 10.1016/s0165-6147(98)01270-x. [DOI] [PubMed] [Google Scholar]
- Starostik MR, Rebello MR, Cotter KA, Kulik A, Medler KF. Expression of GABAergic receptors in mouse taste receptor cells. PLoS One. 2010;5:e13639. doi: 10.1371/journal.pone.0013639. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tomchik SM, Berg S, Kim JW, Chaudhari N, Roper SD. Breadth of tuning and taste coding in mammalian taste buds. J Neurosci. 2007;27:10840–10848. doi: 10.1523/JNEUROSCI.1863-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vandenbeuch A, Clapp TR, Kinnamon SC. Amiloride-sensitive channels in type I fungiform taste cells in mouse. BMC Neurosci. 2008;9:1. doi: 10.1186/1471-2202-9-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vandenbeuch A, Tizzano M, Anderson CB, Stone LM, Goldberg D, Kinnamon SC. Evidence for a role of glutamate as an efferent transmitter in taste buds. BMC Neurosci. 2010;11:77. doi: 10.1186/1471-2202-11-77. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang DD, Kriegstein AR. Defining the role of GABA in cortical development. J Physiol. 2009;587:1873–1879. doi: 10.1113/jphysiol.2008.167635. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yee CL, Yang R, Bottger B, Finger TE, Kinnamon JC. “Type III” cells of rat taste buds: immunohistochemical and ultrastructural studies of neuron-specific enolase, protein gene product 9.5, and serotonin. J Comp Neurol. 2001;440:97–108. doi: 10.1002/cne.1372. [DOI] [PubMed] [Google Scholar]
- Zhao GQ, Zhang Y, Hoon MA, Chandrashekar J, Erlenbach I, Ryba NJ, Zuker CS. The receptors for mammalian sweet and umami taste. Cell. 2003;115:255–266. doi: 10.1016/s0092-8674(03)00844-4. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Figure S1. Biosensors do not respond to GABAergic agonists or antagonists. A. 5-HT biosensors show robust Ca2+ responses to bath-applied serotonin (3 nM), but not to GABA (10 μM), Bac (baclofen, 1 μM), Mus (muscimol, 1 μM), Bic (bicuculline, 10 μM), CGP (CGP55845, 10 μM). Similarly, responses to 5-HT itself were unaffected by these compounds (not shown). B. ATP biosensors respond to ATP (1 μM) but not to the GABAergic drugs as in A. Responses to ATP also were not affected by the presence of these drugs (not shown). For A and B alike, three representative, highly sensitive biosensor cells are shown.
Figure S2. RT-PCR for GABA receptor subunits that are expressed at low abundance or not at all in mouse taste buds. A, GABA-A receptor subunits α1, α5, ρ2 and γ3 are expressed in tissue dissected from the vallate papilla (va) and in some (but not all) samples of taste buds (tb1, tb2). The vallate sample contains muscle, connective tissue, and importantly, nerves. Thus, reaction product in some taste bud samples may represent either very low levels of expression in taste buds or contamination from other cells in the papilla. Controls as in Figures 5, 7 are non-taste lingual epithelium (nt), no cDNA (-), and brain (br). B, GABA-A receptor subunits α2, α4, γ2, θ, ε, and ρ3 are detectable only in vallate tissue pieces (va) and not in isolated taste buds (tb1, tb2). C, GABA-A receptor subunits α3, α6, γ1, and ρ1 were undetectable in all taste bud containing samples.
Figure S3. Relative expression of GAD 65 and GAD67 in vallate, foliate, fungiform, and palatal taste buds. Three independent samples of taste buds from each taste field were processed for qRT-PCR for for GAD65 (A), GAD67 (B) and SNAP25, a marker for taste bud cells. Bars are mean ± s.e.m. for relative expression of each GABA-synthetic enzyme, normalized to expression of SNAP25 in the same sample.
Figure S4. Immunoreactivity for GABA is also detected in mouse palatal and fungiform taste buds. Figure 7 in the manuscript demonstrates that GAD67-expressing (Presynaptic) taste cells in vallate taste buds accumulate GABA. Here we show similar results for taste buds from fungiform papillae and palate. GFP fluorescence (green) in taste cells from GAD67-GFP transgenic mice overlaps with, and is a subset of cells that are immunopositive for GABA (red). Scale, 20 μm.