

Two-component regulatory systems enable bacteria to adapt to changes in their environment during infection by altering gene expression and coordinating antihost responses. Despite the critical role of two-component systems in bacterial survival and pathogenesis, the activating signals for most of these regulators remain unidentified. This is exemplified by ArlRS, a Staphylococcus aureus global regulator that contributes to virulence and to resisting host-mediated restriction of essential nutrients, such as manganese. In this report, we demonstrate that manganese starvation and the absence of glycolytic substrates activate ArlRS. Further investigations revealed that ArlRS is activated when the latter half of glycolysis is disrupted, suggesting that S. aureus monitors flux through the second half of this pathway. Host-imposed manganese starvation also induced the expression of pore-forming toxins in an ArlRS-dependent manner. Cumulatively, this work reveals that ArlRS acts as a sensor that links nutritional status, cellular metabolism, and virulence regulation.
KEYWORDS: ArlRS, Staphylococcus aureus, calprotectin, glycolysis, manganese, nutritional immunity
ABSTRACT
During infection, bacteria use two-component signal transduction systems to sense and adapt to the dynamic host environment. Despite critically contributing to infection, the activating signals of most of these regulators remain unknown. This also applies to the Staphylococcus aureus ArlRS two-component system, which contributes to virulence by coordinating the production of toxins, adhesins, and a metabolic response that enables the bacterium to overcome host-imposed manganese starvation. Restricting the availability of essential transition metals, a strategy known as nutritional immunity, constitutes a critical defense against infection. In this work, expression analysis revealed that manganese starvation imposed by the immune effector calprotectin or by the absence of glycolytic substrates activates ArlRS. Manganese starvation imposed by calprotectin also activated the ArlRS system even when glycolytic substrates were present. A combination of metabolomics, mutational analysis, and metabolic feeding experiments revealed that ArlRS is activated by alterations in metabolic flux occurring in the latter half of the glycolytic pathway. Moreover, calprotectin was found to induce expression of staphylococcal leukocidins in an ArlRS-dependent manner. These studies indicated that ArlRS is a metabolic sensor that allows S. aureus to integrate multiple environmental stresses that alter glycolytic flux to coordinate an antihost response and to adapt to manganese starvation. They also established that the latter half of glycolysis represents a checkpoint to monitor metabolic state in S. aureus. Altogether, these findings contribute to understanding how invading pathogens, such as S. aureus, adapt to the host during infection and suggest the existence of similar mechanisms in other bacterial species.
INTRODUCTION
To successfully establish infection, pathogens must adapt to the dynamic host milieu. This need occurs as host defenses, such as nutrient limitation and antimicrobials, alter local environmental conditions. Among the mechanisms that bacteria use to sense and respond to these changes are the two-component signal transduction systems (TCSs). These regulators reprogram diverse aspects of microbial physiology in response to specific cues by controlling the expression of genes that are essential for bacterial survival, growth, and pathogenesis (1–3). The importance of these systems is exemplified in Staphylococcus aureus, where these regulators coordinate multiple cellular functions, including the expression of virulence determinants (4–8). Given that a wide range of virulence factors can be inhibited by targeting a common regulatory element, such as a TCS, these regulators are attractive targets for antibacterial therapy. To aid the design of novel and effective therapeutics, a better understanding of how TCSs allow bacteria to survive under competitive conditions within the human body is necessary. One of the least understood aspects of TCS signaling is their activation. Indeed, the identity of the signals that stimulate the activity of most of these regulators remains unknown.
S. aureus is a versatile pathogen that can cause devastating disease and that asymptomatically colonizes a third of the human population (9). The importance of adaptation to S. aureus pathogenesis is emphasized by the variety of the infections, ranging from skin and soft tissue infections to more-severe manifestations such as sepsis, endocarditis, and osteomyelitis, that it can cause (10–12). The remarkable capacity of S. aureus to cause disease is largely attributable to its ability to coordinate expression of a copious array of virulence determinants, such as toxins, immune-modulatory factors, and exoenzymes (13). To direct this antihost response, S. aureus uses a plethora of regulators, which include TCSs (14–16). S. aureus has 16 conserved TCSs, including ArlRS, which enable it to respond to its environment (17–21). ArlRS has been shown to contribute to the ability of S. aureus to cause skin infections, sepsis, and endocarditis in several animal models (8, 22–27). The arl locus, which encodes the sensor histidine kinase ArlS and the response regulator ArlR, modulates the expression of multiple toxins, exoenzymes, and immune modulators (26). Given that ArlRS also controls expression of various adhesins and surface proteins, it plays a critical role in the regulation of bacterial clumping and adherence (27). Moreover, ArlRS alters metabolism, enabling S. aureus to adapt to disruptions in glycolysis caused by manganese starvation (8). ArlRS regulates these processes directly or via the activity of other staphylococcal regulators, such as Agr, LytSR, MgrA, and Rot (26). Despite the clear importance of the ArlRS system, the molecular signals underlying its activation remain unknown.
During infection, invaders must obtain all of their nutrients from the host. As a countermeasure, the host restricts the availability of these essential nutrients to invaders, a form of defense referred to as nutritional immunity (28–30). The best-characterized example of nutritional immunity is the restriction of essential metals, including manganese (Mn), iron (Fe), and zinc (Zn) (28, 30, 31). As 30% of all proteins are predicted to use a metal (32, 33), this defense is pivotal with respect to the overall host immune response and the outcome of infection. The staphylococcal abscess is the prototypical example of a manganese-withholding response by the host (31, 34). A critical component of this response is the immune effector calprotectin (CP), which is the most abundant cytoplasmic protein in neutrophils and which can be found at foci of infection at concentrations exceeding 1 mg/ml (35, 36). A member of the S100 protein family, CP is a heterodimer comprised of S100A8 and S100A9 (31, 34, 37–39). It contains two metal-binding sites that are capable of sequestering Mn and Zn as well as other transition metals (38, 40–44). In addition to reducing bacterial growth, CP-imposed metal starvation inactivates metal-dependent enzymes and processes, including superoxide dismutases (SODs), which enable bacteria to resist the oxidative burst, and glucose utilization, rendering invading pathogens more sensitive to other host defenses (8, 31, 37, 45). Mice lacking CP have defects in Mn sequestration and are more susceptible to a variety of bacterial and fungal pathogens, including S. aureus, Klebsiella pneumoniae, Acinetobacter baumannii, and Candida albicans (31, 37, 38, 46–49).
Successful pathogens have developed strategies that enable them to overcome host-imposed Mn limitation (28). These adaptations include expression of Mn-sensing regulators and high-affinity Mn transporters, altering cellular metabolism and oxidative stress management (8, 34, 50–63). ArlRS contributes to the ability of S. aureus to resist Mn starvation during infection by promoting amino acid utilization and reducing the cellular demand for manganese that glucose utilization inflicts (8). The necessity of ArlRS for S. aureus to establish invasive disease and resist the restriction of Mn during infection further emphasizes the importance of this regulator (8). Despite the importance of ArlRS, the environmental cues responsible for its activation remain unknown. In this study, we sought to identify the signals that activate ArlRS. Our analysis indicated that disruption of glycolytic flux, resulting either from host-imposed Mn starvation or from growth of S. aureus in the absence of glycolytic substrates, activates this global virulence regulator. The identification of ArlRS as a sensor of glycolytic flux is significant since glucose is the preferred carbon source of S. aureus and multiple other pathogens, and it could explain how invading pathogens adapt to environments where availability of either Mn or glucose is limited. Moreover, analysis of toxin expression revealed that LukED and LukSF-PVL are produced in response to CP in an ArlRS-dependent manner. Overall, these findings reveal the existence of previously unknown links among nutritional immunity, glycolysis, and virulence factor expression.
RESULTS
ArlRS is activated upon reduction of intracellular manganese availability.
ArlRS enhances the ability of S. aureus to resist Mn starvation in culture and during infection (8). Therefore, this TCS could respond to changes in Mn availability. To evaluate this hypothesis, the activity of ArlRS was assessed in the presence and absence of CP. This was accomplished using a transcriptional reporter fusion under the control of the mgrA P2 promoter, which is controlled by ArlRS (23). In wild-type bacteria, but not the ΔarlR mutant, CP activated the mgrA promoter in a dose-dependent manner (Fig. 1A), indicating that metal starvation activates ArlRS. To resolve the issue of whether Mn or Zn sequestration by CP activated ArlRS, we utilized CP variants lacking the Mn/Zn-binding site (ΔSI), the Zn-only-binding site (ΔSII), or both sites (ΔSI/ΔSII) (37, 38, 64–66). ArlRS was activated only in the presence of wild-type CP and the ΔSII variant, indicating that Mn limitation activates the system (Fig. 1B). Altogether, these findings indicate that host-imposed Mn starvation activates ArlRS.
FIG 1.

ArlRS is activated in response to reduced levels of intracellular Mn. (A) Wild-type bacteria and a ΔarlR mutant were grown in TSB with various concentrations of CP. ArlRS activity was assessed by measuring the activity of the mgrA P2 promoter using a YFP-reporter construct at t = 6 h. *, P ≤ 0.05 (relative to the same strain without CP by two-way analysis of variance [ANOVA] with Dunnett’s multiple-comparison test). #, P ≤ 0.05 (relative to wild-type [WT] bacteria at the same CP concentration by two-way ANOVA with Sidak’s multiple-comparison test). RFU, relative fluorescence units. (B) Wild-type S. aureus and a ΔmntC ΔmntH mutant were grown in TSB with 480 μg/ml WT CP or the ΔSI, ΔSII, or ΔSIΔSII mutant. ArlRS activity was assessed by measuring the activity of the mgrA P2 promoter using a YFP-reporter construct at t = 9 h. *, P ≤ 0.05 (relative to wild-type CP in the same strain by two-way ANOVA with Dunnett’s multiple-comparison test). (C and D) Wild-type bacteria and a ΔmntC ΔmntH mutant were grown in TSB with increasing concentrations of CP. The activity of the mgrA P2 (C) and mntA (D) promoters was assessed using YFP-reporter plasmids at t = 9 h. *, P ≤ 0.05 (relative to wild-type bacteria at the same CP concentration by two-way ANOVA with Sidak’s multiple-comparison test). #, P ≤ 0.05 (relative to the same strain in the absence of CP by two-way ANOVA with Dunnett’s multiple-comparison test). (E and F) Wild-type S. aureus and the ΔmntC ΔmntH mutant were grown in chemically defined medium (CDM) supplemented with various concentrations of manganese, and activity of the mgrA P2 (E) and mntA (F) promoters was assessed using a YFP-reporter plasmid at t = 9 h. *, P ≤ 0.05 (relative to wild-type bacteria at the same Mn concentration by two-way ANOVA with Sidak’s multiple-comparison test). #, P ≤ 0.05 (relative to the same strain in the presence of 1 μM Mn by two-way ANOVA with Dunnett’s multiple-comparison test). n ≥ 3 (all panels). Error bars indicate standard errors of the means (SEM).
CP reduces both the extracellular and intracellular concentrations of bioavailable Mn. This raises the possibility that ArlRS could be responding to changes in the intracellular or extracellular availability of this metal. In wild-type bacteria, intracellular concentrations of Mn are tied to extracellular abundance. To separate these two compartments, we utilized a S. aureus mutant that lacks the Mn transporters MntABC and MntH (ΔmntC ΔmntH) (34, 50). Due to its defect in Mn acquisition, the ΔmntC ΔmntH mutant accumulates less intracellular Mn than wild-type bacteria (50). Hence, if extracellular levels of Mn modulate ArlRS activity, expression of mgrA should be equivalent in wild-type bacteria and the ΔmntC ΔmntH mutant. Conversely, if ArlRS responds to intracellular Mn availability, expression of mgrA should be higher in the ΔmntC ΔmntH mutant than in the wild-type bacteria under conditions of limited Mn availability. In the presence of CP, but not in the presence of metal-replete media, the ΔmntC ΔmntH mutant exhibited elevated ArlRS activity compared to the wild-type strain (Fig. 1C). Under all of the sets of conditions where ArlRS was activated, increased activity of the promoter for the mntABC locus was observed (Fig. 1D). As expression of the MntABC transporter is induced when intracellular manganese levels decrease (34), this indicates that the bacteria are experiencing Mn limitation. As expected in Mn-replete medium, activation of ArlRS was not observed in the ΔmntC ΔmntH mutant when the ΔSI CP variant was used (Fig. 1B). Cumulatively, these results indicate that ArlRS responded to a reduction in intracellular Mn concentrations. To evaluate whether this was generalizable or specific to CP, we evaluated the activity of ArlRS grown in metal-restricted defined medium supplemented with various concentrations of Mn. Consistent with the CP assays, ArlRS was more active in the ΔmntC ΔmntH mutant than in wild-type bacteria when no supplemental Mn was added to the medium (Fig. 1E). Surprisingly, activity was observed only in the absence of Mn supplementation and only in the ΔmntC ΔmntH mutant, although expression of mntA in the wild-type and mutant strains indicated that both were experiencing metal limitation (Fig. 1F). While confirming that ArlRS senses reduced intracellular Mn availability, the substantial limitation necessary to activate the system suggests that the system might be responding to disruption of a Mn-dependent process rather than directly sensing Mn availability.
Pyruvate indirectly activates ArlRS.
Previous work demonstrated that ArlRS is a critical component of pyruvate-mediated virulence regulation and suggested that this regulator might directly or indirectly sense pyruvate levels in S. aureus (67). In these studies, an ArlRS-dependent increase in the expression of staphylococcal toxins was observed in yeast complete (YC) medium supplemented with 2% pyruvate compared to unsupplemented medium (67). Mn starvation disrupts the ability of S. aureus to consume glucose (8), leading to the hypothesis that manganese starvation activates ArlRS via pyruvate. As the prior work relied on toxin expression (67), the activity of ArlRS following growth in the presence of pyruvate was initially assessed using the mgrA P2 promoter. Consistent with prior results (67), supplementing YC medium with 2% pyruvate, but not at a concentration of 1% or less, increased ArlRS activity (Fig. 2A). Building on this observation, pyruvate accumulation was assessed in S. aureus grown in the presence of concentrations of CP that activate ArlRS. These assays revealed that CP reduced intracellular pyruvate levels (Fig. 2B), suggesting that pyruvate indirectly activates ArlRS.
FIG 2.

Pyruvate indirectly activates ArlRS. (A) Wild-type S. aureus was grown in yeast complete (YC) medium supplemented with various concentrations of pyruvate (pyr) or glucose (glc). ArlRS activity was assessed by measuring the activity of the mgrA P2 promoter using a YFP-reporter plasmid at t = 6 h. *, P ≤ 0.05 (relative to bacteria grown in the absence of pyruvate by one-way ANOVA with Dunnett’s multiple-comparison test). (B) Wild-type S. aureus was grown in TSB medium in the presence and absence of 240 μg/ml CP, and pyruvate levels were assessed. *, P ≤ 0.05 (relative to bacteria grown without CP by unpaired two-tailed t test). (C) Wild-type S. aureus was grown in YC medium supplemented with 0.25% glucose or 2% pyruvate, and 2-phosphoglycerate levels were assessed. *, P ≤ 0.05 (relative to bacteria grown in YC medium alone by one-way ANOVA with Dunnett’s multiple-comparison test). n ≥ 3 (all panels). Error bars indicate SEM.
The absence of glycolytic substrates activates ArlRS.
The addition of 2% pyruvate increases 2-phosphoglycerate (2-PG) levels in S. aureus (Fig. 2C) and alters the expression of metabolic genes (67). Those observations and additional prior results (8) suggest a link between ArlRS activity and glycolysis. To test this idea, the impact of glucose on ArlRS activity was evaluated. Growth of S. aureus in tryptic soy broth (TSB) without glucose and in TSB with glucose (0.25%) revealed that ArlRS activity was ∼2 times higher in the cells grown in the absence of glucose than in those cultivated in its presence (Fig. 3A). Consistent with these observations, a similar change in ArlRS activity occurred when S. aureus was grown in Luria broth (LB) and LB supplemented with 0.25% glucose (Fig. 3B). The addition of 0.25% glucose to YC media also reduced the activity of ArlRS (Fig. 2A). The latter observation also indicates that the addition of 2% pyruvate to this medium leads to further activation of a system that is already on. Together, these observations indicate that the increased activity of ArlRS was due to the absence of glucose and not to the presence of or absence of other components of these media.
FIG 3.

The absence of glucose and manganese starvation activate ArlRS. Wild-type S. aureus was grown in TSB (A) or LB (B) supplemented with 0.25% glucose (+glc) or without glucose (no glc) as indicated. ArlRS activity was assessed by measuring the activity of the mgrA P2 promoter using a YFP-reporter plasmid at t = 6 h. *, P ≤ 0.05 (relative to bacteria grown without glucose at the same CP concentration by two-way ANOVA with Sidak’s multiple-comparison test). #, P ≤ 0.05 (relative to bacteria grown in the same medium without CP by two-way ANOVA with Dunnett’s multiple-comparison test). n ≥ 3. Error bars indicate SEM.
To evaluate the possibility that ArlRS was directly sensing the presence of glucose, we assessed the impact that other carbon sources had on its activity. Similarly to the results seen with glucose, ArlRS activation was repressed by fructose and glycerol, which feed into glycolysis at the level of fructose-6-phosphate and dihydroxyacetone phosphate (Fig. 4A), respectively (Fig. 4B). These observations indicate that lack of flux through the first half of glycolysis did not impact ArlRS activity and that the system was not sensing glucose directly. On the other hand, S. aureus grown in the presence of pyruvate (0.25%) exhibited increased ArlRS activity compared to that observed in cells grown in the presence of glucose (Fig. 4B). The elevated ArlRS activity seen under these conditions was presumably due to reduced glycolytic flux, as ArlRS activity in pyruvate-containing medium was repressed by reconstitution of glycolytic activity with either glucose or glycerol (Fig. 4C). Cumulatively, these results suggest that reduced glycolytic flux activates ArlRS.
FIG 4.

Disruption of the latter half of glycolysis activates ArlRS. (A) Simplified representation of glycolysis indicating where fructose and glycerol feed into the pathway. (B) Wild-type S. aureus Newman was grown in TSB without glucose and supplemented with carbon-balanced concentrations of glucose (glc), fructose (fru), glycerol (glyc), or pyruvate (pyr). (B to D) ArlRS activity was assessed by measuring the activity of the mgrA P2 promoter using a YFP-reporter plasmid at t = 6 h. (B) *, P ≤ 0.05 (relative to medium without glucose by one-way ANOVA with Dunnett’s multiple-comparison test). (C and D) Wild-type S. aureus Newman (C) and USA300 JE2 (D) were grown in TSB without glucose and without supplementation (−) or supplemented with carbon-balanced concentrations of glucose (glc), glycerol (glyc), and/or pyruvate (pyr) in the absence and presence of 480 μg/ml CP. *, P ≤ 0.05 (relative to growth in medium without glucose at the same CP concentration by two-way ANOVA with Dunnett’s multiple-comparison test). #, P ≤ 0.05 (relative to growth in the same carbon source without CP by two-way ANOVA with Sidak’s multiple-comparison test). n ≥ 3 (panels B to D). Error bars indicate SEM.
Manganese limitation activates ArlRS regardless of the glycolytic carbon source present.
To determine if the activities of the two stimuli that modulate ArlRS activation are interconnected, the activity of this TCS was assessed in S. aureus grown in TSB or LB with or without glucose in the presence of various concentrations of CP. Treatment with CP increased ArlRS activity in medium containing glucose in a dose-dependent manner (Fig. 3). However, CP treatment did not increase ArlRS activity when the bacteria were grown in the absence of glucose. CP also increased the activity of ArlRS when glycerol was provided as a carbon source (Fig. 4C). When grown in the presence of 0.25% pyruvate, a condition that results in activation comparable to that seen in the absence of glucose or glycerol (Fig. 4B), only minimal induction was observed when CP was added to the growth medium (Fig. 4C). Similar results were observed in the presence and absence of CP with the methicillin-resistant USA300 JE2 strain (Fig. 4D). Together with the observation that Mn starvation prevents S. aureus from using glucose as an energy source (8), these data suggest that disruption of glycolysis by CP activates ArlRS.
Disruption of the latter half of glycolysis activates ArlRS.
To understand how disruption of glycolytic flux activates ArlRS, the levels of glycolytic intermediates in bacteria grown in the absence of glucose were compared to the levels seen with those grown in the presence of glucose but treated with CP, using mass spectrometry. Bacterial cells grown in the absence of glucose contained lower levels of intracellular glucose, fructose 1,6-biphosphate, dihydroxyacetone phosphate, glyceraldehyde 3-phosphate, 2(3)-phosphoglycerate (the two forms were not differentiated by mass spectroscopy), phosphoenolpyruvate, and pyruvate than cells grown in the presence of this stimulus (Fig. 5). CP alters normal glycolytic activity, as it results in a substantial decrease in the levels of intracellular glucose and lactate coupled with an increase in the levels of dihydroxyacetone phosphate and 2(3)-phosphoglycerate (Fig. 5). These data are consistent with CP disrupting glycolytic flux as previously suggested (8). However, no common changes in glycolytic intermediates were observed between bacteria grown in the presence of CP and those grown in the absence of glucose when compared to bacteria grown in metal-replete medium containing glucose.
FIG 5.
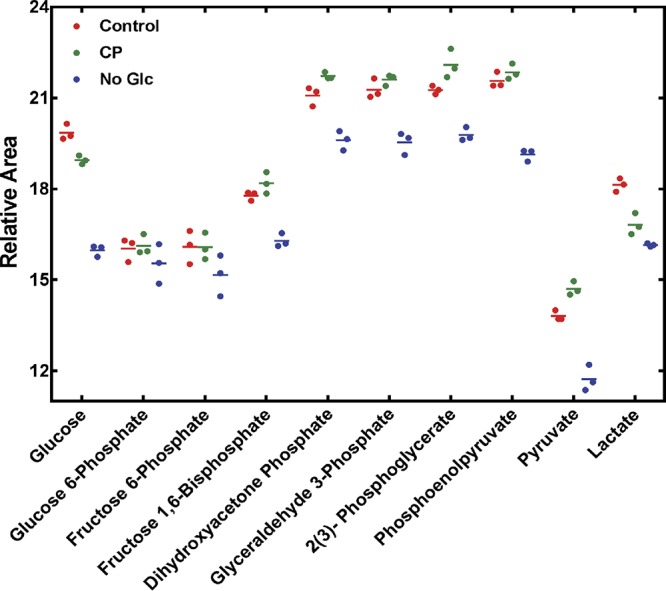
Relative levels of glycolytic intermediates in cells treated with CP or deprived of glucose. S. aureus was grown either in the presence of 120 μg/ml CP or without glucose, and the relative levels of glycolytic intermediates were measured using mass spectrometry as described in Materials and Methods. The level of each metabolite was normalized to the total ion current of the 86 metabolites detected. Each dot represents an independent measurement, and means are indicated by horizontal lines.
While metabolomics failed to identify a metabolite change that connected CP treatment and glucose withdrawal, the mass spectrometry approach utilized did not detect 1,3-bisphosphoglycerate and cannot distinguish 3-phosphoglycerate from 2-phosphoglycerate. 2-PG levels were assessed using enzymatic analysis to determine if a reduction in 2-PG correlated with activation of ArlRS. The abundance of this intermediate was assessed in S. aureus grown under conditions that activate this TCS, including 2% pyruvate-containing YC medium, TSB without glucose, and TSB with CP. Enzymatic analysis revealed that, although cells grown in the absence of glucose accumulated less 2-PG than cells grown in its presence, S. aureus grown in the presence of both CP and 2% pyruvate contained elevated 2-PG levels compared to bacteria grown in the absence of these stimuli (Fig. 2C; see also 6A and B). The divergent concentrations of 2-PG observed in these three conditions suggest that changes in this metabolite are not responsible for activating ArlRS.
The increase in 2-PG levels in the presence of pyruvate and CP led us to evaluate if accumulation rather than depletion of an intermediate activated ArlRS. In light of the mass spectrometry results, the possibility that accumulation of either 1,3-bisphosphoglycerate (1,3-BPG) or 3-phosphoglycerate (3-PG) might activate ArlRS was considered. S. aureus has two phosphoglycerate mutases, namely, GpmA and GpmI. GpmI requires Mn for activity, while GpmA does not (68, 69). We determined whether interfering with the conversion of 3-PG to 2-PG affects ArlRS activity by deleting each of these genes and growing the mutants in the presence of CP. In the presence of CP, loss of GpmA increased ArlRS activity compared to the levels seen with either wild-type bacteria or the ΔgpmI mutant (Fig. 6C). As the ΔgpmA mutant must use the Mn-dependent GpmI, which should be inhibited by CP, this indicates that disrupting phosphoglycerate mutase activity increases the activity of ArlRS. We also observed that ArlRS activity was modestly higher in the ΔgpmA mutant in Mn-replete medium. This observation suggests the GpmA is the main enzyme catalyzing this step of glycolysis when Mn is freely available. Together, these observations suggest that accumulation of either 3-PG or 1,3-bisphosphoglycerate could activate ArlRS.
FIG 6.

Accumulation of 3-PG or of other upstream intermediates activates ArlRS. (A and B) Wild-type S. aureus was grown in TSB without glucose (no glc) supplemented with 0.25% glucose (glc) (A) or in glucose-containing TSB in the absence and presence of 480 μg/ml CP (B), and 2-phosphoglycerate levels were assessed. *, P ≤ 0.05 (relative to bacteria grown in the absence of either glucose [A] or CP [B] by unpaired two-tailed t test). (C) Wild-type, ΔgpmI, and ΔgpmA S. aureus were grown in TSB in the absence and presence of 480 μg/ml CP. ArlRS activity was assessed by measuring the activity of the mgrA P2 promoter using a YFP-reporter plasmid at t = 6 h. *, P ≤ 0.05 (relative to wild-type bacteria at the same CP concentration by two-way ANOVA with Dunnett’s multiple-comparison test). #, P ≤ 0.05 (relative to the same strain in the absence of CP by two-way ANOVA with Sidak’s multiple-comparison test). n ≥ 3 (panels A to C). Error bars indicate SEM.
CP induces ArlRS-dependent expression of pore-forming leukocidins.
ArlRS coordinates the expression of virulence determinants, including the leukocidins LukED and LukSF-PVL (8, 22, 24, 26). Therefore, we investigated the impact that availability of glucose and Mn would have on the expression of these two leukocidins. Surprisingly, no induction of either toxin in either wild-type bacteria or a ΔarlR mutant was observed in glucose-free medium relative to medium containing glucose (Fig. 7A and B). However, in the presence of CP, the expression levels of lukED and lukSF-PVL increased ∼50-fold and ∼48-fold, respectively, in an ArlRS-dependent manner (Fig. 7C and D). Collectively, these observations indicate that ArlRS contributes to the expression of staphylococcal virulence factors in response to nutritional immunity.
FIG 7.

CP induces ArlRS-dependent expression of pore-forming leukocidins. (A to D) Wild-type and ΔarlR S. aureus USA300 JE2 were grown in TSB without glucose (no glc), supplemented with 0.25% glucose (glc) (A and B) or glucose-containing TSB in the presence of increasing concentrations of CP (C and D). Leukocidin expression was assessed by measuring the activity of the lukE (A and C) and lukS-PVL (B and D) promoters using a Lux-reporter plasmid at t = 6 h. (A to D) *, P ≤ 0.05 (relative to wild-type bacteria under each condition by two-way ANOVA with Sidak’s multiple-comparison test). (C and D) #, P ≤ 0.05 (relative to the same strain in the absence of CP by two-way ANOVA with Dunnett’s multiple-comparison test). n ≥ 3 (panels A to D). Error bars indicate SEM.
DISCUSSION
TCSs play an important role in allowing pathogens to adapt to the hostile host environment, as they enable bacteria to perceive environmental changes and reprogram their physiology in response to stress (1–3). Despite the importance of TCSs in bacterial pathogenesis, the identity of the activating signals for most of these regulators remains unclear. The current investigations revealed that the global virulence regulator ArlRS responds to alterations in metabolic flux occurring in the latter half of the glycolytic pathway (Fig. 8). This represents a new regulatory checkpoint for glycolysis. ArlRS is required for maximal toxin expression in response to CP-imposed disruption of glycolysis, and this sensor enables the integration of multiple inputs to coordinate an antihost response. Together, these findings highlight previously unappreciated connections among nutritional immunity, metabolism, and virulence and broaden our understanding of the signals utilized by invading pathogens to thrive within the host.
FIG 8.

Model of ArlRS activation. When glycolytic carbon sources (such as glucose and glycerol) and manganese are abundant in the host environment, ArlRS remains inactive. However, in an environment poor in glycolytic carbon sources or under conditions of host-imposed manganese starvation, both stimuli (which alter flux in the latter half of glycolysis) elicit the activation of ArlRS. Activation of this TCS allows S. aureus to coordinate an antihost response consisting of the production of virulence factors such as leukocidins and results in reprogramming of staphylococcal metabolism in response to nutritional immunity.
ArlRS was recently suggested to respond to pyruvate availability (67). However, the current investigations suggested that activation of ArlRS by pyruvate is indirect. The current studies also revealed that ArlRS responds to intracellular Mn availability. Notably, both CP and 2% pyruvate either disrupt glycolysis or alter the expression of glycolytic enzymes or both (8, 67). While the specific step in glycolysis that is disrupted by CP is unknown, excess pyruvate appears to drive staphylococcal metabolism toward gluconeogenesis (67). The latter idea is supported by the current observation that 2% pyruvate increased cellular levels of 2-PG. ArlRS is also activated by the absence of glycolytic intermediates that enter the pathway above 1,3-BPG, which would also trigger gluconeogenesis. Differing from the results seen with CP and 2% pyruvate, 2-PG levels decrease in the absence of glucose, suggesting that an upstream metabolite rather than 2-PG activates ArlRS. This idea is supported by the observation that ablation of staphylococcal phosphoglycerate mutase activity, which presumptively leads to the accumulation of 3-PG and other upstream metabolites, activates ArlRS. Combined with the results of metabolomics analyses, which failed to detect common changes in other glycolytic intermediates, these observations suggest that elevation of the level of 3-PG or 1,3-BPG results in ArlRS activation.
In S. aureus and other pathogens, metabolism and virulence factor expression are intertwined (70–74). In Gram-positive bacteria, the carbon catabolite repressor CcpA is important for the regulation of streptococcal capsular biosynthesis and the expression of a variety of toxins in organisms such as Clostridium difficile, Bacillus anthracis, and S. aureus (72, 75). Similarly, the branched-chain amino acid sensor CodY acts as a toxin gene regulator in S. aureus, C. difficile, B. anthracis, and Listeria monocytogenes (72, 75, 76). Rex, a redox-dependent transcriptional repressor, negatively regulates expression of genes involved in respiration and promotes toxin expression (72, 74, 75). The current investigations revealed that ArlRS responds to the abundance of intermediates formed in the second half of glycolysis, a novel node for regulatory control, and coordinates the expression of virulence factors such as adhesins and toxins (27, 67). Intriguingly, our observations suggest that ArlRS is part of a layered response. Both the absence of glycolytic substrates and the absence of CP activate the mgrA promoter, which is associated with controlling staphylococcal adherence and clumping (23). However, expression of LukED and LukSF-PVL changes only in response to CP, indicating that a second signal is necessary. This idea is further supported by the observation that in YC medium, ArlRS activation by pyruvate is also sufficient to induce staphylococcal toxin expression (67). CP enhances the activity of the SaeRS TCS, which regulates S. aureus toxin expression (21, 77, 78), suggesting that this system may provide the second signal. Importantly, the induction of LukED and LukSF-PVL in response to CP is not simply due to activation of the SaeRS system, as loss of ArlR ablates CP-mediated induction of the toxins. Both LukED and LukSF-PVL target neutrophils, which are the primary source of CP during staphylococcal infection (79). While increased toxin expression is potentially beneficial in the presence of neutrophils, it is difficult to envision how this would benefit S. aureus in the absence of glycolytic substrates. Thus, it is tempting to speculate that the multiple layers of regulation enable S. aureus to fine-tune a response triggered by a system that monitors a pathway that can be disrupted by multiple stressors.
Glycolysis is tightly regulated by the environment and flux through the pathway. The mechanisms by which both Gram-positive and Gram-negative organisms sense flux through the first half of glycolysis are well established (72, 80). In Gram-positive bacteria, including S. aureus, fructose bisphosphatase (FBP) regulates glycolysis by two mechanisms. First, FBP promotes the import of glucose and other sugars via the CcpA/Hpr pair, allowing bacteria to selectively use glycolytic substrates when diverse carbon sources are present (81, 82). Second, FBP allosterically stimulates pyruvate kinase activity, enabling S. aureus and other bacteria to ensure that the glucose brought into the cell is flowing through the first half of glycolysis (83, 84). And yet bacteria encounter stresses (including Mn limitation; copper, nickel, and fluoride intoxication; and nitrosative damage) that disrupt glycolysis after the import and conversion of glucose to FBP but before the production of pyruvate (8, 85–88). The current work revealed that the activity of ArlRS changes in response to disruptions in the second half of glycolysis triggered by host-imposed Mn limitation. This enables S. aureus to shift away from carbohydrate catabolism to amino acid degradation, enabling the bacterium to circumvent the impact of nutritional immunity (8). In other bacteria, including Escherichia coli, Salmonella enterica serovar Typhimurium, Bacillus stearothermophilus, and Bacillus subtilis, a step after the formation of FBP is known to be or is likely to be manganese dependent (89–93). Similarly to S. aureus, Streptococcus pneumoniae shifts metabolism away from glucose utilization under conditions of Mn starvation (62). Additionally, the ResDE two-component system in Bacillus subtilis has been suggested to respond to 1,3-bisphosphoglycerate levels enabling anaerobic growth (94). Although no obvious gene targets are shared between ArlRS and ResDE, both respond to disruptions in the latter half of glycolysis and enable adaptation to environmental stress (26, 95, 96). Together, these observations suggest that monitoring the second half of the glycolytic pathway may be a common strategy employed by bacteria.
Adaptation to a changing environment is critical for survival, and it is possible for multiple stresses to disrupt the same metabolic pathway. Sensing intracellular metabolic flux enables bacteria to monitor disruption of critical metabolic pathways, broadening their ability to respond to stressors. The current investigation established that ArlRS is activated by decreased sugar availability and nutritional immunity, both of which alter flux in the latter half of glycolysis, identifying this metabolic segment as a new regulatory node. Further investigations will increase our understanding of how S. aureus and potentially other pathogens leverage changes in the latter half of glycolysis to coordinate their response to nutrient abundance and the immune response.
MATERIALS AND METHODS
Bacterial strains and cloning.
S. aureus strains were grown at 37°C in tryptic soy broth (TSB) on a roller drum or on tryptic soy agar (TSA) plates for performing routine culturing or for genetic manipulation. E. coli strains were routinely cultivated at 37°C in Luria broth (LB) with shaking or on Luria agar plates. As needed for plasmid maintenance in E. coli and S. aureus, 100 μg/ml of ampicillin and 10 μg/ml of chloramphenicol were added to the growth medium, respectively. Both bacterial species were stored at −80°C in growth medium containing 30% glycerol.
S. aureus Newman and its derivatives were used for all experiments, unless otherwise indicated. For experiments using USA300 JE2 and the derivative USA300 JE2 arlR::erm, strains were obtained from the Nebraska library (97). The ΔarlR and ΔarlR ΔmntC ΔmntH mutants were generated by the use of phage transducing the arlR::erm allele from USA300 JE2 into the wild-type and ΔmntC ΔmntH backgrounds (Φ85 phage). For fluorescent reporters, the upstream promoter of mgrA P2 and the promoter of mntA were cloned into yellow fluorescent protein (YFP)-containing vector pAH5 (98) using the indicated primers (see Table S1 in the supplemental material). Plasmids were electroporated into S. aureus RN4220 (99) before being transferred into final recipient strains, as previously described (100). All constructs were verified by sequencing. The hemolytic activity of all staphylococcal strains was confirmed by plating on blood agar plates. See Tables S2 and S3 for the full lists of strains and plasmids utilized in this study.
Primers used in this study. Download Table S1, DOCX file, 0.01 MB (12.5KB, docx) .
Copyright © 2019 Párraga Solórzano et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Staphylococcus aureus strains used in this study. Download Table S2, DOCX file, 0.02 MB (19.2KB, docx) .
Copyright © 2019 Párraga Solórzano et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Plasmids used in this study. Download Table S3, DOCX file, 0.02 MB (21.6KB, docx) .
Copyright © 2019 Párraga Solórzano et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
ArlRS activity reporter assays.
For assays assessing ArlRS activity in the presence of CP, bacteria were grown as previously described with minor modifications (8, 37, 38, 101). Briefly, overnight cultures were back-diluted 1:100 in 96-well round-bottom plates containing 100 μl/well of growth medium, consisting of 38% of the indicated complex medium and 62% calprotectin buffer (20 mM Tris [pH 7.5], 100 mM NaCl, 3 mM CaCl2, 10 mM β-mercaptoethanol). This medium was supplemented with 1 μM MnCl2 and 1 μM ZnSO4. All cultures were incubated at 37°C with shaking at 180 rpm. Growth was assessed by measuring optical density at 600 nm (OD600). Expression of mgrA and mntA was determined by measuring yellow fluorescence (excitation and emission wavelengths of 505 nm and 535 nm, respectively) and normalizing to OD600, as previously described (63, 101, 102). Expression of lukE and lukS-PVL was determined by measuring luminescence and normalizing to OD600. When indicated, equimolar concentrations of carbon from glucose (5.34 mM), fructose (5.34 mM), glycerol (10.68 mM), or sodium pyruvate (10.68 mM) were provided.
For assays assessing ArlRS activation in the presence of different concentrations of manganese, S. aureus was grown overnight in 5 ml of NRPMI (Chelex-treated RPMI) (34) supplemented with 1 mM MgCl2, 100 μM CaCl2, 1 μM ZnSO4 and 1 μM FeCl2. The overnight cultures were back-diluted 1:100 in 96-well round-bottom plates containing 100 μl/well of growth medium, consisting of 38% chemically defined medium (2.6×) and 62% calprotectin buffer (20 mM Tris [pH 7.5], 100 mM NaCl, 1 mM CaCl2, 10 mM β-mercaptoethanol) (8). The defined medium (2.6×) consisted of 0.5 g/liter NaCl, 1.0 g/liter NH4Cl, 2 g/liter KH2PO4, 7 g/liter Na2HPO4, 0.228 μg/liter biotin, 0.228 mg/liter nicotinic acid, 0.228 mg/liter pyridoxine-HCl, 0.228 mg/liter thiamine-HCl, 0.114 mg/liter riboflavin, 0.684 mg/liter calcium pantothenate, 0.104 g/liter phenylalanine, 0.078 g/liter isoleucine, 0.130 g/liter tyrosine, 0.053 g/liter cysteine, 0.260 g/liter glutamic acid, 0.026 g/liter lysine, 0.182 g/liter methionine, 0.078 g/liter histidine, 0.026 g/liter tryptophan, 0.234 g/liter leucine, 0.234 g/liter aspartic acid, 0.182 g/liter arginine, 0.078 g/liter serine, 0.156 g/liter alanine, 0.078 g/liter threonine, 0.130 g/liter glycine, 0.208 g/liter valine, and 0.026 g/liter proline. Glucose (1.3%) was provided as the carbon source. This medium was supplemented with 2.3 mM MgSO4, 1 μM ZnSO4, 1 μM FeSO4, and the indicated concentration of MnCl2.
For assessing ArlRS activation in YC medium, overnight cultures grown in YC medium were back-diluted 1:100 in 96-well round-bottom plates containing 100 μl/well of YC medium supplemented with glucose (5.34 mM) or with 0.25%, 0.5%, 1%, or 2% sodium pyruvate or with both, as indicated.
Pyruvate assays.
Overnight cultures were back-diluted 1:100 in 96-well round-bottom plates containing 100 μl/well of growth medium consisting of 38% TSB (either with or without glucose) and 62% calprotectin buffer (20 mM Tris [pH 7.5], 100 mM NaCl, 3 mM CaCl2, 10 mM β-mercaptoethanol) and were grown in the absence or presence of 240 μg/ml CP. Bacteria were harvested during logarithmic-phase growth at similar optical densities (OD600 = 0.2 to 0.25). Bacterial culture (∼60 ml) was collected in a Millipore S-Pak membrane filter, washed twice with 0.5% NaCl, and stored at −80°C in 60% (vol/vol) high-performance liquid chromatography (HPLC)-grade ethanol. Prior to assay of pyruvate, cells were thawed, washed from the filter, and then lysed by mechanical disruption using a FastPrep-24 homogenizer (MP Biomedicals) for 45 s at 6.5 m/s. Cell lysates were clarified by centrifugation at 10,000 × g for 5 min at 4°C. The supernatants were collected and vacuum dried. The dehydrated pellets were resuspended in 100 μl of HPLC-grade water, and then pyruvate levels were measured using an EnzyChrom pyruvate assay kit (BioAssay Systems) as indicated by the manufacturer.
2-Phosphoglycerate assays.
For 2-phosphoglycerate determination, bacteria were grown and processed as previously described for pyruvate assays using either growth medium consisting of 38% TSB (either with or without glucose) and 62% calprotectin buffer in the absence or presence of 240 μg/ml CP or growth medium consisting of YC medium supplemented with 0.25% glucose or 2% sodium pyruvate or both, as indicated. The dehydrated pellets obtained after processing were resuspended in 100 μl of 2-PG assay buffer, and then 2-phosphoglycerate levels and total protein concentrations were measured using a 2-phosphoglycerate colorimetric/fluorometric assay kit (BioVision) and a Pierce bicinchoninic acid (BCA) protein assay kit, respectively.
Metabolomics analysis.
For metabolomics analysis, bacteria were grown and processed as described for the pyruvate assays using either growth medium consisting of 38% TSB (either with or without glucose) or 62% calprotectin buffer in the absence or presence of 120 μg/ml CP and were harvested during logarithmic-phase growth at an OD600 of 0.1 to 0.15. The dehydrated pellets were resuspended in 80% methanol. Metabolites were analyzed using a Shimadzu Prominence Ultra Fast Liquid Chromatograph (UFLC) attached to a Sciex QTrap 4500 system equipped with a Turbo V ion source (Sciex). Samples (5 μl) were injected into an XSelect HSS C18 column (2.5-μm pore size, 3.0 by 150 mm) using a flow rate of 0.3 ml/min. Solvent A contained 100 mM ammonium formate (pH 5.0), 2% acetonitrile, and 0.1% t-butanol. Solvent B was composed of 95% acetonitrile, 50 mM ammonium formate (pH 6.3), and 0.1% t-butanol. The HPLC program was as follows: starting solvent mixture of 0% solvent B, 0-to-2-min isocratic with 0% solvent B; 2-to-12-min linear gradient to 5% solvent B; 12-to-17-min linear gradient to 90% solvent B; 17-to-25-min isocratic with 90% solvent B; 25-to-27-min linear gradient to 0% solvent B; 27-to-30-min isocratic with 0% solvent B. The Sciex QTrap 4500 system was operated in the negative mode, and the ion source parameters were as follows: ion spray voltage, −4,500 V; curtain gas pressure, 40 lb/in2; temperature, 500°C; collision gas setting, high; ion source gas 1 pressure, 50 lb/in2; ion source gas 2 pressure, 50 lb/in2. Multiple-reaction monitoring (MRM) precursor and product ions were derived as described previously by Bajad et al. (103). The system was controlled by the use of Analyst software and analyzed with MultiQuant 3.0.2 software (Sciex, Inc.). The relative levels of individual metabolite were determined by normalizing each signal to the total current sum of ions of the 86 metabolites that were measured in the sample.
Statistical analysis.
All statistical analyses were performed using GraphPad Prism software, version 7.02. The specific statistical tests used are described in the figure legends.
ACKNOWLEDGMENTS
We thank the members of the Kehl-Fie laboratory for their critical reading of the manuscript and Matt Frank for mass spectrometry. We are also grateful to Victor J. Torres for the pXen1-lukE and pXen1-lukS reporter plasmids and for thoughtful discussions.
This work was supported by grants from the National Institutes of Health (K22 AI104805 and R01 AI 118880 to T.E.K.-F. and GM034496 to C.O.R.), a March of Dimes Basil O’Connor Award, a Vallee Scholar Award to T.E.K.-F., and an Alice Helm Graduate Student Excellence Award to P.K.P.S. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript. Its contents are solely our responsibility and do not necessarily represent the official views of the NIH, the March of Dimes, or Vallee Foundation.
Footnotes
Citation Párraga Solórzano PK, Yao J, Rock CO, Kehl-Fie TE. 2019. Disruption of glycolysis by nutritional immunity activates a two-component system that coordinates a metabolic and antihost response by Staphylococcus aureus. mBio 10:e01321-19. https://doi.org/10.1128/mBio.01321-19.
REFERENCES
- 1.Stock AM, Robinson VL, Goudreau PN. 2000. Two-component signal transduction. Annu Rev Biochem 69:183–215. doi: 10.1146/annurev.biochem.69.1.183. [DOI] [PubMed] [Google Scholar]
- 2.Gao R, Stock AM. 2009. Biological insights from structures of two-component proteins. Annu Rev Microbiol 63:133–154. doi: 10.1146/annurev.micro.091208.073214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Capra EJ, Laub MT. 2012. Evolution of two-component signal transduction systems. Annu Rev Microbiol 66:325–347. doi: 10.1146/annurev-micro-092611-150039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Sun F, Ji Q, Jones MB, Deng X, Liang H, Frank B, Telser J, Peterson SN, Bae T, He C. 2012. AirSR, a [2Fe-2S] cluster-containing two-component system, mediates global oxygen sensing and redox signaling in Staphylococcus aureus. J Am Chem Soc 134:305–314. doi: 10.1021/ja2071835. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Hall JW, Ji Y. 2013. Sensing and adapting to anaerobic conditions by Staphylococcus aureus. Adv Appl Microbiol 84:1–25. doi: 10.1016/B978-0-12-407673-0.00001-1. [DOI] [PubMed] [Google Scholar]
- 6.Yan M, Hall JW, Yang J, Ji Y. 2012. The essential yhcSR two-component signal transduction system directly regulates the lac and opuCABCD operons of Staphylococcus aureus. PLoS One 7:e50608. doi: 10.1371/journal.pone.0050608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Yan MY, Yu CX, Yang JS, Ji YD. 2011. The essential two-component system YhcSR is involved in regulation of the nitrate respiratory pathway of Staphylococcus aureus. J Bacteriol 193:1799–1805. doi: 10.1128/JB.01511-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Radin JN, Kelliher JL, Parraga Solorzano PK, Kehl-Fie TE. 2016. The two-component system ArlRS and alterations in metabolism enable Staphylococcus aureus to resist calprotectin-induced manganese starvation. PLoS Pathog 12:e1006040. doi: 10.1371/journal.ppat.1006040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Wertheim HF, Vos MC, Ott A, van Belkum A, Voss A, Kluytmans JA, van Keulen PH, Vandenbroucke-Grauls CM, Meester MHM, Verbrugh HA. 2004. Risk and outcome of nosocomial Staphylococcus aureus bacteraemia in nasal carriers versus non-carriers. Lancet 364:703–705. doi: 10.1016/S0140-6736(04)16897-9. [DOI] [PubMed] [Google Scholar]
- 10.Tong SY, Davis JS, Eichenberger E, Holland TL, Fowler VG Jr.. 2015. Staphylococcus aureus infections: epidemiology, pathophysiology, clinical manifestations, and management. Clin Microbiol Rev 28:603–661. doi: 10.1128/CMR.00134-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.WHO. 2014. Antimicrobial resistance global report on surveillance. World Health Organization, Geneva, Switzerland. [Google Scholar]
- 12.CDC. 2013. Antibiotic resistance threats in the United States, 2013. Centers for Disease Control and Prevention, Atlanta, GA. [Google Scholar]
- 13.Foster TJ, Geoghegan JA, Ganesh VK, Hook M. 2014. Adhesion, invasion and evasion: the many functions of the surface proteins of Staphylococcus aureus. Nat Rev Microbiol 12:49–62. doi: 10.1038/nrmicro3161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Novick RP. 2003. Autoinduction and signal transduction in the regulation of staphylococcal virulence. Mol Microbiol 48:1429–1449. doi: 10.1046/j.1365-2958.2003.03526.x. [DOI] [PubMed] [Google Scholar]
- 15.Cheung AL, Bayer AS, Zhang G, Gresham H, Xiong YQ. 2004. Regulation of virulence determinants in vitro and in vivo in Staphylococcus aureus. FEMS Immunol Med Microbiol 40:1–9. doi: 10.1016/S0928-8244(03)00309-2. [DOI] [PubMed] [Google Scholar]
- 16.Haag AF, Bagnoli F. 2017. The role of two-component signal transduction systems in Staphylococcus aureus virulence regulation. Curr Top Microbiol Immunol 409:145–198. doi: 10.1007/82_2015_5019. [DOI] [PubMed] [Google Scholar]
- 17.Kuroda M, Ohta T, Uchiyama I, Baba T, Yuzawa H, Kobayashi I, Cui L, Oguchi A, Aoki K, Nagai Y, Lian J, Ito T, Kanamori M, Matsumaru H, Maruyama A, Murakami H, Hosoyama A, Mizutani-Ui Y, Takahashi NK, Sawano T, Inoue R, Kaito C, Sekimizu K, Hirakawa H, Kuhara S, Goto S, Yabuzaki J, Kanehisa M, Yamashita A, Oshima K, Furuya K, Yoshino C, Shiba T, Hattori M, Ogasawara N, Hayashi H, Hiramatsu K. 2001. Whole genome sequencing of meticillin-resistant Staphylococcus aureus. Lancet 357:1225–1240. doi: 10.1016/s0140-6736(00)04403-2. [DOI] [PubMed] [Google Scholar]
- 18.Novick RP, Geisinger E. 2008. Quorum sensing in staphylococci. Annu Rev Genet 42:541–564. doi: 10.1146/annurev.genet.42.110807.091640. [DOI] [PubMed] [Google Scholar]
- 19.Voyich JM, Vuong C, DeWald M, Nygaard TK, Kocianova S, Griffith S, Jones J, Iverson C, Sturdevant DE, Braughton KR, Whitney AR, Otto M, DeLeo FR. 2009. The SaeR/S gene regulatory system is essential for innate immune evasion by Staphylococcus aureus. J Infect Dis 199:1698–1706. doi: 10.1086/598967. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Goerke C, Fluckiger U, Steinhuber A, Bisanzio V, Ulrich M, Bischoff M, Patti JM, Wolz C. 2005. Role of Staphylococcus aureus global regulators sae and sigmaB in virulence gene expression during device-related infection. Infect Immun 73:3415–3421. doi: 10.1128/IAI.73.6.3415-3421.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Cho H, Jeong DW, Liu Q, Yeo WS, Vogl T, Skaar EP, Chazin WJ, Bae T. 2015. Calprotectin increases the activity of the SaeRS two component system and murine mortality during Staphylococcus aureus infections. PLoS Pathog 11:e1005026. doi: 10.1371/journal.ppat.1005026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Benton BM, Zhang JP, Bond S, Pope C, Christian T, Lee L, Winterberg KM, Schmid MB, Buysse JM. 2004. Large-scale identification of genes required for full virulence of Staphylococcus aureus. J Bacteriol 186:8478–8489. doi: 10.1128/JB.186.24.8478-8489.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Crosby HA, Schlievert PM, Merriman JA, King JM, Salgado-Pabón W, Horswill AR. 2016. The Staphylococcus aureus global regulator MgrA modulates clumping and virulence by controlling surface protein expression. PLoS Pathog 12:e1005604. doi: 10.1371/journal.ppat.1005604. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Fournier B, Klier A, Rapoport G. 2001. The two-component system ArlS-ArlR is a regulator of virulence gene expression in Staphylococcus aureus. Mol Microbiol 41:247–261. doi: 10.1046/j.1365-2958.2001.02515.x. [DOI] [PubMed] [Google Scholar]
- 25.Fournier B, Hooper DC. 2000. A new two-component regulatory system involved in adhesion, autolysis, and extracellular proteolytic activity of Staphylococcus aureus. J Bacteriol 182:3955–3964. doi: 10.1128/jb.182.14.3955-3964.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Liang X, Zheng L, Landwehr C, Lunsford D, Holmes D, Ji Y. 2005. Global regulation of gene expression by ArlRS, a two-component signal transduction regulatory system of Staphylococcus aureus. J Bacteriol 187:5486–5492. doi: 10.1128/JB.187.15.5486-5492.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Walker JN, Crosby HA, Spaulding AR, Salgado-Pabon W, Malone CL, Rosenthal CB, Schlievert PM, Boyd JM, Horswill AR. 2013. The Staphylococcus aureus ArlRS two-component system is a novel regulator of agglutination and pathogenesis. PLoS Pathog 9:e1003819. doi: 10.1371/journal.ppat.1003819. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Kehl-Fie TE, Skaar EP. 2010. Nutritional immunity beyond iron: a role for manganese and zinc. Curr Opin Chem Biol 14:218–224. doi: 10.1016/j.cbpa.2009.11.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Weinberg ED. 2009. Iron availability and infection. Biochim Biophys Acta 1790:600–605. doi: 10.1016/j.bbagen.2008.07.002. [DOI] [PubMed] [Google Scholar]
- 30.Hood MI, Skaar EP. 2012. Nutritional immunity: transition metals at the pathogen-host interface. Nat Rev Microbiol 10:525–537. doi: 10.1038/nrmicro2836. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Corbin BD, Seeley EH, Raab A, Feldmann J, Miller MR, Torres VJ, Anderson KL, Dattilo BM, Dunman PM, Gerads R, Caprioli RM, Nacken W, Chazin WJ, Skaar EP. 2008. Metal chelation and inhibition of bacterial growth in tissue abscesses. Science 319:962–965. doi: 10.1126/science.1152449. [DOI] [PubMed] [Google Scholar]
- 32.Andreini C, Bertini I, Cavallaro G, Holliday GL, Thornton JM. 2008. Metal ions in biological catalysis: from enzyme databases to general principles. J Biol Inorg Chem 13:1205–1218. doi: 10.1007/s00775-008-0404-5. [DOI] [PubMed] [Google Scholar]
- 33.Waldron KJ, Rutherford JC, Ford D, Robinson NJ. 2009. Metalloproteins and metal sensing. Nature 460:823–830. doi: 10.1038/nature08300. [DOI] [PubMed] [Google Scholar]
- 34.Kehl-Fie TE, Zhang Y, Moore JL, Farrand AJ, Hood MI, Rathi S, Chazin WJ, Caprioli RM, Skaar EP. 2013. MntABC and MntH contribute to systemic Staphylococcus aureus infection by competing with calprotectin for nutrient manganese. Infect Immun 81:3395–3405. doi: 10.1128/IAI.00420-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Clohessy PA, Golden BE. 1995. Calprotectin-mediated zinc chelation as a biostatic mechanism in host-defense. Scand J Immunol 42:551–556. doi: 10.1111/j.1365-3083.1995.tb03695.x. [DOI] [PubMed] [Google Scholar]
- 36.Gebhardt C, Nemeth J, Angel P, Hess J. 2006. S100A8 and S100A9 in inflammation and cancer. Biochem Pharmacol 72:1622–1631. doi: 10.1016/j.bcp.2006.05.017. [DOI] [PubMed] [Google Scholar]
- 37.Kehl-Fie TE, Chitayat S, Hood MI, Damo S, Restrepo N, Garcia C, Munro KA, Chazin WJ, Skaar EP. 2011. Nutrient metal sequestration by calprotectin inhibits bacterial superoxide defense, enhancing neutrophil killing of Staphylococcus aureus. Cell Host Microbe 10:158–164. doi: 10.1016/j.chom.2011.07.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Damo SM, Kehl-Fie TE, Sugitani N, Holt ME, Rathi S, Murphy WJ, Zhang Y, Betz C, Hench L, Fritz G, Skaar EP, Chazin WJ. 2013. Molecular basis for manganese sequestration by calprotectin and roles in the innate immune response to invading bacterial pathogens. Proc Natl Acad Sci U S A 110:3841–3846. doi: 10.1073/pnas.1220341110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Korndorfer IP, Brueckner F, Skerra A. 2007. The crystal structure of the human (S100A8/S100A9)2 heterotetramer, calprotectin, illustrates how conformational changes of interacting alpha-helices can determine specific association of two EF-hand proteins. J Mol Biol 370:887–898. doi: 10.1016/j.jmb.2007.04.065. [DOI] [PubMed] [Google Scholar]
- 40.Nakashige TG, Zygiel EM, Drennan CL, Nolan EM. 2017. Nickel sequestration by the host-defense protein human calprotectin. J Am Chem Soc 139:8828–8836. doi: 10.1021/jacs.7b01212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Nakashige TG, Zhang B, Krebs C, Nolan EM. 2015. Human calprotectin is an iron-sequestering host-defense protein. Nat Chem Biol 11:765–771. doi: 10.1038/nchembio.1891. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Brophy MB, Nakashige TG, Gaillard A, Nolan EM. 2013. Contributions of the S100A9 C-terminal tail to high-affinity Mn(II) chelation by the host-defense protein human calprotectin. J Am Chem Soc 135:17804–17817. doi: 10.1021/ja407147d. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Radin JN, Zhu J, Brazel EB, McDevitt CA, Kehl-Fie TE. 19 December 2018, posting date. Synergy between nutritional immunity and independent host defenses contributes to the importance of the MntABC manganese transporter during Staphylococcus aureus infection. Infect Immun doi: 10.1128/IAI.00642-18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Andrews SC, Robinson AK, Rodríguez-Quiñones F. 2003. Bacterial iron homeostasis. FEMS Microbiol Rev 27:215–237. doi: 10.1016/S0168-6445(03)00055-X. [DOI] [PubMed] [Google Scholar]
- 45.Juttukonda LJ, Chazin WJ, Skaar EP. 2016. Acinetobacter baumannii coordinates urea metabolism with metal import to resist host-mediated metal limitation. mBio 7:e01475-16. doi: 10.1128/mBio.01475-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Urban CF, Ermert D, Schmid M, Abu-Abed U, Goosmann C, Nacken W, Brinkmann V, Jungblut PR, Zychlinsky A. 2009. Neutrophil extracellular traps contain calprotectin, a cytosolic protein complex involved in host defense against Candida albicans. PLoS Pathog 5:e1000639. doi: 10.1371/journal.ppat.1000639. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Bianchi M, Niemiec MJ, Siler U, Urban CF, Reichenbach J. 2011. Restoration of anti-Aspergillus defense by neutrophil extracellular traps in human chronic granulomatous disease after gene therapy is calprotectin-dependent. J Allergy Clin Immunol 127:1243–1252.e7. doi: 10.1016/j.jaci.2011.01.021. [DOI] [PubMed] [Google Scholar]
- 48.Achouiti A, Vogl T, Urban CF, Rohm M, Hommes TJ, van Zoelen MA, Florquin S, Roth J, van't Veer C, de Vos AF, van der Poll T. 2012. Myeloid-related protein-14 contributes to protective immunity in gram-negative pneumonia derived sepsis. PLoS Pathog 8:e1002987. doi: 10.1371/journal.ppat.1002987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Hood MI, Mortensen BL, Moore JL, Zhang Y, Kehl-Fie TE, Sugitani N, Chazin WJ, Caprioli RM, Skaar EP. 2012. Identification of an Acinetobacter baumannii zinc acquisition system that facilitates resistance to calprotectin-mediated zinc sequestration. PLoS Pathog 8:e1003068. doi: 10.1371/journal.ppat.1003068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Horsburgh MJ, Wharton SJ, Cox AG, Ingham E, Peacock S, Foster SJ. 2002. MntR modulates expression of the PerR regulon and superoxide resistance in Staphylococcus aureus through control of manganese uptake. Mol Microbiol 44:1269–1286. doi: 10.1046/j.1365-2958.2002.02944.x. [DOI] [PubMed] [Google Scholar]
- 51.Que Q, Helmann JD. 2000. Manganese homeostasis in Bacillus subtilis is regulated by MntR, a bifunctional regulator related to the diphtheria toxin repressor family of proteins. Mol Microbiol 35:1454–1468. [DOI] [PubMed] [Google Scholar]
- 52.Hendriksen WT, Bootsma HJ, van Diepen A, Estevao S, Kuipers OP, de Groot R, Hermans P. 2009. Strain-specific impact of PsaR of Streptococcus pneumoniae on global gene expression and virulence. Microbiology 155:1569–1579. doi: 10.1099/mic.0.025072-0. [DOI] [PubMed] [Google Scholar]
- 53.Kehres DG, Janakiraman A, Slauch JM, Maguire ME. 2002. Regulation of Salmonella enterica serovar Typhimurium mntH transcription by H2O2, Fe2+, and Mn2+. J Bacteriol 184:3151–3158. doi: 10.1128/JB.184.12.3151-3158.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Patzer SI, Hantke K. 2001. Dual repression by Fe2+-Fur and Mn2+-MntR of the mntH gene, encoding an NRAMP-like Mn2+ transporter in Escherichia coli. J Bacteriol 183:4806–4813. doi: 10.1128/JB.183.16.4806-4813.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Posey JE, Hardham JM, Norris SJ, Gherardini FC. 1999. Characterization of a manganese-dependent regulatory protein, TroR, from Treponema pallidum. Proc Natl Acad Sci U S A 96:10887–10892. doi: 10.1073/pnas.96.19.10887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Pandey R, Russo R, Ghanny S, Huang XJ, Helmann J, Rodriguez GM. 2015. MntR(Rv2788): a transcriptional regulator that controls manganese homeostasis in Mycobacterium tuberculosis. Mol Microbiol 98:1168–1183. doi: 10.1111/mmi.13207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Schmitt MP. 2002. Analysis of a DtxR-like metalloregulatory protein, MntR, from Corynebacterium diphtheriae that controls expression of an ABC metal transporter by an Mn2+-dependent mechanism. J Bacteriol 184:6882–6892. doi: 10.1128/JB.184.24.6882-6892.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Papp-Wallace KM, Maguire ME. 2006. Manganese transport and the role of manganese in virulence. Annu Rev Microbiol 60:187–209. doi: 10.1146/annurev.micro.60.080805.142149. [DOI] [PubMed] [Google Scholar]
- 59.Agranoff D, Monahan IM, Mangan JA, Butcher PD, Krishna S. 1999. Mycobacterium tuberculosis expresses a novel pH-dependent divalent cation transporter belonging to the Nramp family. J Exp Med 190:717–724. doi: 10.1084/jem.190.5.717. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Kehres DG, Zaharik ML, Finlay BB, Maguire ME. 2000. The NRAMP proteins of Salmonella Typhimurium and Escherichia coli are selective manganese transporters involved in the response to reactive oxygen. Mol Microbiol 36:1085–1100. doi: 10.1046/j.1365-2958.2000.01922.x. [DOI] [PubMed] [Google Scholar]
- 61.Janulczyk R, Ricci S, Bjorck L. 2003. MtsABC is important for manganese and iron transport, oxidative stress resistance, and virulence of Streptococcus pyogenes. Infect Immun 71:2656–2664. doi: 10.1128/IAI.71.5.2656-2664.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Ogunniyi AD, Mahdi LK, Jennings MP, McEwan AG, McDevitt CA, Van der Hoek MB, Bagley CJ, Hoffmann P, Gould KA, Paton JC. 2010. Central role of manganese in regulation of stress responses, physiology, and metabolism in Streptococcus pneumoniae. J Bacteriol 192:4489–4497. doi: 10.1128/JB.00064-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Garcia YM, Barwinska-Sendra A, Tarrant E, Skaar EP, Waldron KJ, Kehl-Fie TE. 2017. A superoxide dismutase capable of functioning with iron or manganese promotes the resistance of Staphylococcus aureus to calprotectin and nutritional immunity. PLoS Pathog 13:e1006125. doi: 10.1371/journal.ppat.1006125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Nakashige TG, Stephan JR, Cunden LS, Brophy MB, Wommack AJ, Keegan BC, Shearer JM, Nolan EM. 2016. The hexahistidine motif of host-defense protein human calprotectin contributes to zinc withholding and its functional versatility. J Am Chem Soc 138:12243–12251. doi: 10.1021/jacs.6b06845. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Hayden JA, Brophy MB, Cunden LS, Nolan EM. 2013. High-affinity manganese coordination by human calprotectin is calcium-dependent and requires the histidine-rich site formed at the dimer interface. J Am Chem Soc 135:775–787. doi: 10.1021/ja3096416. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Brophy MB, Hayden JA, Nolan EM. 2012. Calcium ion gradients modulate the zinc affinity and antibacterial activity of human calprotectin. J Am Chem Soc 134:18089–18100. doi: 10.1021/ja307974e. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Harper L, Balasubramanian D, Ohneck EA, Sause WE, Chapman J, Mejia-Sosa B, Lhakhang T, Heguy A, Tsirigos A, Ueberheide B, Boyd JM, Lun DS, Torres VJ. 2018. Staphylococcus aureus responds to the central metabolite pyruvate to regulate virulence. mBio 9:e02272-17. doi: 10.1128/mBio.02272-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Omelchenko MV, Galperin MY, Wolf YI, Koonin EV. 2010. Non-homologous isofunctional enzymes: a systematic analysis of alternative solutions in enzyme evolution. Biol Direct 5:31. doi: 10.1186/1745-6150-5-31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Foster JM, Davis PJ, Raverdy S, Sibley MH, Raleigh EA, Kumar S, Carlow CK. 2010. Evolution of bacterial phosphoglycerate mutases: non-homologous isofunctional enzymes undergoing gene losses, gains and lateral transfers. PLoS One 5:e13576. doi: 10.1371/journal.pone.0013576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Pohl K, Francois P, Stenz L, Schlink F, Geiger T, Herbert S, Goerke C, Schrenzel J, Wolz C. 2009. CodY in Staphylococcus aureus: a regulatory link between metabolism and virulence gene expression. J Bacteriol 191:2953–2963. doi: 10.1128/JB.01492-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Geiger T, Goerke C, Fritz M, Schafer T, Ohlsen K, Liebeke M, Lalk M, Wolz C. 2010. Role of the (p)ppGpp synthase RSH, a RelA/SpoT homolog, in stringent response and virulence of Staphylococcus aureus. Infect Immun 78:1873–1883. doi: 10.1128/IAI.01439-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Richardson AR, Somerville GA, Sonenshein AL. 2015. Regulating the intersection of metabolism and pathogenesis in gram-positive bacteria. Microbiol Spectr 3(3). doi: 10.1128/microbiolspec.MBP-0004-2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Seidl K, Müller S, François P, Kriebitzsch C, Schrenzel J, Engelmann S, Bischoff M, Berger-Bächi B. 2009. Effect of a glucose impulse on the CcpA regulon in Staphylococcus aureus. BMC Microbiol 9:95. doi: 10.1186/1471-2180-9-95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Pagels M, Fuchs S, Pane-Farre J, Kohler C, Menschner L, Hecker M, McNamarra PJ, Bauer MC, von Wachenfeldt C, Liebeke M, Lalk M, Sander G, von Eiff C, Proctor RA, Engelmann S. 2010. Redox sensing by a Rex-family repressor is involved in the regulation of anaerobic gene expression in Staphylococcus aureus. Mol Microbiol 76:1142–1161. doi: 10.1111/j.1365-2958.2010.07105.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Somerville GA, Proctor RA. 2009. At the crossroads of bacterial metabolism and virulence factor synthesis in staphylococci. Microbiol Mol Biol Rev 73:233–248. doi: 10.1128/MMBR.00005-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Berman N, Rettger LF. 1918. The influence of carbohydrate on the nitrogen metabolism of bacteria. J Bacteriol 3:389–402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Geiger T, Goerke C, Mainiero M, Kraus D, Wolz C. 2008. The virulence regulator Sae of Staphylococcus aureus promoter activities and response to phagocytosis-related signals. J Bacteriol 190:3419–3428. doi: 10.1128/JB.01927-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Novick RP, Jiang D. 2003. The staphylococcal saeRS system coordinates environmental signals with agr quorum sensing. Microbiology 149:2709–2717. doi: 10.1099/mic.0.26575-0. [DOI] [PubMed] [Google Scholar]
- 79.Spaan AN, van Strijp JAG, Torres VJ. 2017. Leukocidins: staphylococcal bi-component pore-forming toxins find their receptors. Nat Rev Microbiol 15:435–447. doi: 10.1038/nrmicro.2017.27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Shimizu K. 2013. Regulation systems of bacteria such as Escherichia coli in response to nutrient limitation and environmental stresses. Metabolites 4:1–35. doi: 10.3390/metabo4010001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Titgemeyer F, Hillen W. 2002. Global control of sugar metabolism: a gram-positive solution. Antonie Van Leeuwenhoek 82:59–71. doi: 10.1023/A:1020628909429. [DOI] [PubMed] [Google Scholar]
- 82.Warner JB, Lolkema JS. 2003. CcpA-dependent carbon catabolite repression in bacteria. Microbiol Mol Biol Rev 67:475–490. doi: 10.1128/mmbr.67.4.475-490.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Zoraghi R, See RH, Gong H, Lian T, Swayze R, Finlay BB, Brunham RC, McMaster WR, Reiner NE. 2010. Functional analysis, overexpression, and kinetic characterization of pyruvate kinase from methicillin-resistant Staphylococcus aureus. Biochemistry 49:7733–7747. doi: 10.1021/bi100780t. [DOI] [PubMed] [Google Scholar]
- 84.Munoz ME, Ponce E. 2003. Pyruvate kinase: current status of regulatory and functional properties. Comp Biochem Physiol B Biochem Mol Biol 135:197–218. doi: 10.1016/S1096-4959(03)00081-2. [DOI] [PubMed] [Google Scholar]
- 85.Tarrant E, Riboldi GP, McIlvin MR, Stevenson J, Barwinska-Sendra A, Stewart LJ, Saito MA, Waldron KJ. 2019. Copper stress in Staphylococcus aureus leads to adaptive changes in central carbon metabolism. Metallomics 11:183–200. doi: 10.1039/C8MT00239H. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Marquis RE. 1995. Antimicrobial actions of fluoride for oral bacteria. Can J Microbiol 41:955–964. doi: 10.1139/m95-133. [DOI] [PubMed] [Google Scholar]
- 87.Macomber L, Elsey SP, Hausinger RP. 2011. Fructose-1,6-bisphosphate aldolase (class II) is the primary site of nickel toxicity in Escherichia coli. Mol Microbiol 82:1291–1300. doi: 10.1111/j.1365-2958.2011.07891.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Bourret TJ, Boylan JA, Lawrence KA, Gherardini FC. 2011. Nitrosative damage to free and zinc-bound cysteine thiols underlies nitric oxide toxicity in wild-type Borrelia burgdorferi. Mol Microbiol 81:259–273. doi: 10.1111/j.1365-2958.2011.07691.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Hohle TH, O'Brian MR. 2012. Manganese is required for oxidative metabolism in unstressed Bradyrhizobium japonicum cells. Mol Microbiol 84:766–777. doi: 10.1111/j.1365-2958.2012.08057.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Watabe K, Freese E. 1979. Purification and properties of the manganese-dependent phosphoglycerate mutase of Bacillus subtilis. J Bacteriol 137:773–778. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Kehres DG, Maguire ME. 2003. Emerging themes in manganese transport, biochemistry and pathogenesis in bacteria. FEMS Microbiol Rev 27:263–290. doi: 10.1016/S0168-6445(03)00052-4. [DOI] [PubMed] [Google Scholar]
- 92.Chander M, Setlow B, Setlow P. 1998. The enzymatic activity of phosphoglycerate mutase from gram-positive endospore-forming bacteria requires Mn2+ and is pH sensitive. Can J Microbiol 44:759–767. doi: 10.1139/w98-060. [DOI] [PubMed] [Google Scholar]
- 93.Fraser HI, Kvaratskhelia M, White MF. 1999. The two analogous phosphoglycerate mutases of Escherichia coli. FEBS Lett 455:344–348. doi: 10.1016/s0014-5793(99)00910-2. [DOI] [PubMed] [Google Scholar]
- 94.Nakano MM, Zhu Y, Haga K, Yoshikawa H, Sonenshein AL, Zuber P. 1999. A mutation in the 3-phosphoglycerate kinase gene allows anaerobic growth of Bacillus subtilis in the absence of ResE kinase. J Bacteriol 181:7087–7097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Hulett FM. 1996. The signal-transduction network for Pho regulation in Bacillus subtilis. Mol Microbiol 19:933–939. doi: 10.1046/j.1365-2958.1996.421953.x. [DOI] [PubMed] [Google Scholar]
- 96.Nakano MM. 2002. Induction of ResDE-dependent gene expression in Bacillus subtilis in response to nitric oxide and nitrosative stress. J Bacteriol 184:1783–1787. doi: 10.1128/jb.184.6.1783-1787.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Fey PD, Endres JL, Yajjala VK, Widhelm TJ, Boissy RJ, Bose JL, Bayles KW. 2013. A genetic resource for rapid and comprehensive phenotype screening of nonessential Staphylococcus aureus genes. mBio 4:e00537-12. doi: 10.1128/mBio.00537-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Malone CL, Boles BR, Lauderdale KJ, Thoendel M, Kavanaugh JS, Horswill AR. 2009. Fluorescent reporters for Staphylococcus aureus. J Microbiol Methods 77:251–260. doi: 10.1016/j.mimet.2009.02.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Novick RP. 1991. Genetic systems in staphylococci. Methods Enzymol 204:587–636. doi: 10.1016/0076-6879(91)04029-N. [DOI] [PubMed] [Google Scholar]
- 100.Schenk S, Laddaga RA. 1992. Improved method for electroporation of Staphylococcus aureus. FEMS Microbiol Lett 73:133–138. doi: 10.1016/0378-1097(92)90596-g. [DOI] [PubMed] [Google Scholar]
- 101.Grim KP, San Francisco B, Radin JN, Brazel EB, Kelliher JL, Parraga Solorzano PK, Kim PC, McDevitt CA, Kehl-Fie TE. 2017. The metallophore staphylopine enables Staphylococcus aureus to compete with the host for zinc and overcome nutritional immunity. mBio 8:e01281-17. doi: 10.1128/mBio.01281-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Kelliher JL, Radin JN, Grim KP, Parraga Solorzano PK, Degnan PH, Kehl-Fie TE. 2018. Acquisition of the phosphate transporter NptA enhances Staphylococcus aureus pathogenesis by improving phosphate uptake in divergent environments. Infect Immun 86:e00631-17. doi: 10.1128/IAI.00631-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Bajad SU, Lu W, Kimball EH, Yuan J, Peterson C, Rabinowitz JD. 2006. Separation and quantitation of water soluble cellular metabolites by hydrophilic interaction chromatography-tandem mass spectrometry. J Chromatogr A 1125:76–88. doi: 10.1016/j.chroma.2006.05.019. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Primers used in this study. Download Table S1, DOCX file, 0.01 MB (12.5KB, docx) .
Copyright © 2019 Párraga Solórzano et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Staphylococcus aureus strains used in this study. Download Table S2, DOCX file, 0.02 MB (19.2KB, docx) .
Copyright © 2019 Párraga Solórzano et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Plasmids used in this study. Download Table S3, DOCX file, 0.02 MB (21.6KB, docx) .
Copyright © 2019 Párraga Solórzano et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.