

Abstract
Dendritic spines form the postsynaptic contact sites for most excitatory synapses in the brain. Spines occur in a wide range of different shapes that can vary depending on an animal’s experience or behavioral status. Recently we showed that spines on living neurons can change shape within seconds in a process that depends on actin polymerization. We have now found that this morphological plasticity is blocked by inhalational anesthetics at concentrations at which they are clinically effective. These volatile compounds also block actin-based motility in fibroblasts, indicating that their action is independent of neuron-specific components and thus identifying the actin cytoskeleton as a general cellular target of anesthetic action. These observations imply that inhibition of actin dynamics at brain synapses occurs during general anesthesia and that inhalational anesthetics are capable of influencing the morphological plasticity of excitatory synapses in the brain.
General anesthesia can be induced by inhaling one of a variety of gaseous compounds, ranging from the inert gas xenon to volatile clinical anesthetics such as isoflurane. It is remarkable that despite their chemical diversity, these simple molecules produce an anesthetized state that can otherwise be replicated only by a combination of several pharmacological agents. This observation has led to the suggestion that the anesthetics properties of these agents are produced by a unitary molecular mechanism (1). Consistent with this idea, it was initially believed that volatile anesthetics act by dissolving in the lipid plasma membrane of neurons and inhibiting conduction of the action potential (2, 3). However, subsequent studies have shown that these compounds bind to and influence a number of neuronal proteins, including ion channels and neurotransmitter receptors (4, 5). As a result, the source of the unitary characteristics of the anesthetized state has become less clear.
The binding sites for volatile anesthetics on several synaptic receptor proteins appear to be associated with hydrophobic pockets (6–8) and, consistent with the relatively unspecific nature of hydrophobic interactions, these compounds also bind to proteins not primarily associated with nervous system function (9), including cytochrome c oxidase, adenylate kinase, and luciferase (5, 10, 11). The general nature of this hydrophobic binding mechanism raises the possibility that volatile anesthetics may have additional, as yet unrecognized, neuronal targets. Here we show that such a target is associated with dendritic spines, where volatile anesthetics applied at clinically relevant concentrations antagonize their recently described actin-based morphological plasticity (12). The same concentrations of these agents also block actin-based motility in fibroblasts, indicating that their effect on the actin cytoskeleton is independent of neurotransmitter receptors or other neuron-specific cellular components. Because actin within neurons is concentrated in dendritic spines (13), these results implicate postsynaptic actin in the action of volatile anesthetics at excitatory synapses in the brain.
METHODS
Anesthetics.
Chloroform and diethylether were obtained from Sigma/Fluka, isoflurane (Forene) and enflurane (Ethrane), from Abbott, methoxyflurane (Metofane, Mallinkrodt), from Arovet, Zollikon, Switzerland, and halothane from Research Biochemicals, Buchs, Switzerland. For application to cells, all were diluted from saturated solutions in PBS with the following concentrations (14): chloroform, 66 mM; diethylether, 816 mM; methoxyflurane, 9.1 mM; isoflurane, 15.3 mM; and enflurane, 11.9 mM.
Cell Culture.
Cultures of primary rat hippocampal neurons were established from 19-day rat embryos and transfected to express actin tagged with green fluorescent protein (GFP-actin), as described (15). Alternatively, spines were visualized by using a membrane-targeted fusion construct between GFP and C-terminal 20 amino acids of Ki-ras 4B (A. Welman and J. Hagmann, personal communication). Transgenic mice expressing GFP-actin from a chicken beta-cytoplasmic actin promoter are described elsewhere (U. Wagner and A.M., unpublished data), and organotypic slice cultures from hippocampus were established according to protocols obtained from B. Gahwiler (Brain Research Institute, Zurich). Rat embryo fibroblast cells (REF52) were maintained in DMEM supplemented with 10% fetal calf serum. To test the effect of anesthetics, cells were serum starved for several hours until actin-based motility had ceased, then treated with phorbol 12-myristate 13-acetate [(PMA); Alexis, Laeufelfingen, Switzerland] in PBS in either the presence or absence of anesthetics. Cells fixed with 4% formaldehyde were stained with 10 μg/ml rhodamine-labeled phalloidin to visualize filamentous actin.
Microscopy and Image Analysis.
For live imaging of dissociated neuron cultures, coverslips were mounted in saline in closed observation chambers (Type 1, Life Imaging, Olten, Switzerland) at 37°C, and images were captured by using a cooled CCD camera and metamorph Imaging Software (Universal Imaging, Media, PA), as described (12). Anesthetics were diluted into prewarmed saline immediately before perfusion into the sealed observation chambers. For analysis of spine shape, 12-bit images were thresholded, binarized, and outlined by using metamorph functions for determining object profiles and calculating a shape factor from the area and the perimeter of the delineated object. Spines in organotypic slice cultures were imaged by using an UltraView microlens confocal system (LSR, Cambridge, U.K.) in closed observation chambers and perfused as described for the dissociated cultures. For analysis of differences in spine motility caused by the application of anesthetics, summed differences of the pixel values from successive frames of time-lapse recordings were compared for 10-min periods before and after perfusion with 0.35 mM isoflurane in saline.
RESULTS
Dendritic Spines Can Change Shape Rapidly.
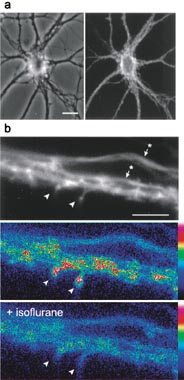
To examine the influence of volatile anesthetics on dendritic spine dynamics, we adapted a procedure previously developed for visualizing spine plasticity in cultured hippocampal neurons (12). This procedure takes advantage of the fact that cytoplasmic actins expressed in these cells by transfection are selectively targeted to spines (13). Expressing GFP-actin renders the spines of living neurons visible (Fig. 1 a and b) so that their dynamic activity can be captured in time-lapse recordings. To demonstrate in still images the time-dependent changes in spine shape that occur in such recordings, we used a computer routine to convert images of individual spines from single frames into profile outlines (Fig. 1c Left). Spine outlines for all the frames of a recording were then used to document the shape changes that had occurred, as illustrated by the selection of frames in Fig. 1c. A second computer routine was used to calculate shape factors for each frame, and these were plotted against time to provide a criterion for changes in spine shape throughout the duration of the recording (Fig. 1d).
Figure 1.

Assessment of actin-dependent dendritic spine motility in living neurons. (a) Primary rat hippocampal neurons expressing GFP-tagged γ-cytoplasmic actin by transfection develop normally [phase contrast of a living cell (Left)] and acquire numerous dendritic spines after 3 wk in culture (Right). Bar = 30 μm. (b) GFP-actin targets to spine heads (arrowheads) where it is present at higher concentrations than in the dendrite shaft. Bar = 2.5 μm. (c) Images of a single dendritic spines in individual frames from time-lapse recording were processed by using a computer algorithm to produce profile outlines [original image and derived profile shown (Left)]. Selected profiles, taken 10 sec apart, demonstrate changes in spine shape that occur during recording. (d) Changes in the shape of single spines during time-lapse recordings were followed by calculating a shape factor from the spine profiles shown in c and plotting them against time, as shown in this example. A perfect circle has a shape factor of one, whereas values for flat objects approach zero.
Effects of Isoflurane on Structural Spine Plasticity.
The procedure described above was used in the experiment shown in Fig. 2, in which spine motility was recorded in GFP-actin expressing cells before and after exposure to isoflurane, a widely used modern anesthetic. Fig. 2 Upper shows computer-generated profiles taken from the time-lapse recording of a single dendritic spine before (Fig. 2a) and after (Fig. 2b) the addition of isoflurane. Data for the entire experiment are shown in Fig. 2c, where the shape factors calculated from the spine profiles are plotted throughout the entire recording. The wide scatter of points before the addition of isoflurane indicates the rapid variation in spine shape that occurs under control conditions compared with the relative lack of change in this parameter in the presence of the anesthetic. This inhibition of plasticity was readily visualized and evident among all spines on the dendrites of hippocampal neurons exposed to isoflurane. The dynamic nature of these effects can be appreciated further from the supplemental video data to Figs. 2 and 3 (see the PNAS web site, www.pnas.org). The lack of spine motility in the presence of isoflurane was confirmed by imaging spine shape by using a membrane-targeted GFP construct indicating that the effects of volatile anesthetics reflect not merely the collapse of actin in the spine head but also a change in the overall morphology of the spine (supplemental Fig. 5; see www.pnas.org). To show that the inhibitory effect of isoflurane on spine motility is not an artifact of the dissociated neuronal culture system, we made recordings of hippocampal slice cultures established from transgenic mice expressing GFP-actin and confirmed that in this organotypic tissue, isoflurane likewise inhibits spine shape changes (supplemental Fig. 6; see www.pnas.org).
Figure 2.

Volatile anesthetics block dendritic spine motility. Outline profiles showing changes in the shape of a dendritic spine visualized in a time-lapse recording of GFP-actin in the spine head. Selected images taken 10 sec apart over a period of 5 min show changes in shape that occurred in control conditions (a) compared with the relative lack of shape change in the presence of clinically relevant concentrations of isoflurane (b). Data from all the frames recorded in this experiment are shown in the shape factor plot (c) in which the variation in spine shape (a Left) is blocked after medium containing isoflurane was perfused into the observation chamber (b Right).
Figure 3.

Effects of volatile anesthetics are reversible. (a) Profile outlines derived from individual frames taken from time-lapse recording made in presence of chloroform. (b) After washout, shape changes rapidly resume. (c) The shape factor plot shows the lack of change in the presence of chloroform (a) compared with the wide variation and high eccentricity that occurred after the anesthetic was removed (b).
Effects of Anesthetics Are Reversible on Washout.
Results similar to those for isoflurane were obtained with diverse volatile anesthetics including chloroform, enflurane, diethylether, and halothane. Fig. 3 shows data from one of a series of experiments in which the recording was started in the presence of an anesthetic to confirm that spine motility recovered when anesthetics were removed. The profile data in Fig. 3a show that spine motility was minimal in the presence of anesthetic, in this case chloroform, whereas Fig. 3b shows the resumption of rapid changes in spine shape after chloroform was removed. Data for this spine over the entire recording are shown in the shape factor plot (Fig. 3c).
Anesthetics Inhibit Spine Dynamics at Clinically Relevant Concentrations.
An important consideration in judging the physiological relevance of in vitro effects induced by general anesthetics is the concentration at which they take place. If they are biologically significant, then in vitro effects should occur at aqueous concentrations equivalent to the inhaled concentration at which they produce anesthesia in animals (5). In this respect, we found that the concentrations of volatile agents needed to inhibit spine motility were closely similar to those required for their anesthetic effects (Table 1). A further significant feature of the inhibition of spine motility by volatile anesthetics was the remarkably steep dose dependence of the effect: using isoflurane, we observed that whereas spines were motile at both 0.15 mM and 0.25 mM, their motility was fully inhibited at 0.30 mM, a dose-response relationship that corresponds in both steepness and effective concentration to the properties of this compound in clinical anesthesia (5).
Table 1.
Comparison of effective concentrations of anesthetics for inhibition of spine dynamics and anesthesia
Effects on Spine Dynamics Are Limited to Volatile Anesthetics.
In addition to these inhalational anesthetics, we also tested two nonvolatile compounds that are used to induce clinical anesthesia, ketamine and propofol. Ketamine, which acts as an antagonist at N-methyl-d-aspartate-type glutamate receptors with an IC50 of 0.43 μM (16), had no effect on spine motility at concentrations up to 10 μM. The clinically effective tissue concentration of propofol has been calculated to be 0.4 μM (5), but tested on hippocampal neurons, it had no effect on spine motility at concentrations up to 5 μM. These results suggest that the anesthetic properties of ketamine and propofol are the result of cellular mechanisms distinct from that of the volatile anesthetics, which despite their chemical heterogeneity share a common influence on spine motility.
Effects of Volatile Anesthetics on Actin Dynamics in Fibroblasts.
The results of these experiments on neurons posed the question whether volatile anesthetics act on dendritic spine motility directly through actin or indirectly through neurotransmitter receptors or neuronal ion channels, which are also known to be influenced by these agents at clinically relevant concentrations. To choose between these possibilities, we examined the influence of volatile anesthetics on actin-based motility in fibroblastic cells, where neurotransmitter receptors and other elements of the synaptic machinery are absent. Time-lapse recordings of GFP-actin transfected cells showed that each of the volatile anesthetics tested strongly inhibited actin motility in rat embryo fibroblasts [Fig. 4; see supplementary video material on the PNAS web site (www.pnas.org)]. To demonstrate this effect in still images, we used a modified experimental approach (Fig. 4). In this procedure, fibroblasts were first transferred to low serum medium until spontaneous actin-based ruffling at their periphery had ceased. Under these conditions, actin is predominantly organized in stress fibers (Fig. 4a). These quiescent cells were then treated with PMA, which causes the reorganization of actin into peripheral accumulations associated with transient surface motility (Fig. 4b). In parallel, some cultures were exposed to anesthetic before the addition of PMA and, under these conditions, actin rearrangement was blocked; the actin remained organized in stress fibers, and membrane ruffling failed to emerge (Fig. 4c). This blockade occurred at the same concentration of anesthetic required for their effects on dendritic spine motility.
Figure 4.

Volatile anesthetics inhibit actin-based dynamics in nonneuronal cells. Rat embryo fibroblast cultures were fixed and stained with rhodamine-labeled phalloidin to reveal polymerized actin. (a) Quiescent serum-starved fibroblasts contain abundant stress fibers but little actin at the cell periphery. (b) Treatment with 100 nM PMA for 15 min induces membrane ruffling, which is associated with high concentrations of filamentous actin. (c) Pretreatment with 0.3 mM isoflurane blocks PMA-induced actin motility, as shown by the absence of actin-rich membrane ruffles. Bar = 50 μm.
Volatile Anesthetics Do Not Influence Actin Polymerization in Vitro.
One explanation for the action of volatile anesthetics on actin motility that we considered was that they might bind to a hydrophobic site on the actin molecule itself and thus interfere with its assembly into filaments. To test this possibility, we examined the influence of isoflurane on actin filament assembly in a standard biochemical actin polymerization assay. Even at concentrations above those that block actin motility in cells, isoflurane had no effect on the polymerization of soluble actin or the depolymerization of preassembled actin filaments (data not shown). It thus seems that volatile anesthetics affect actin dynamics indirectly, for example by interacting with a component of one of the various pathways that influence the assembly state and arrangement of cytoplasmic actin (17, 18).
DISCUSSION
Volatile anesthetics have been shown to affect a variety of ion channels and neurotransmitter receptor subtypes (4, 6, 19–23), leading to the suggestion that they produce anesthesia by acting at many sites simultaneously (5, 24, 25). We describe here an action of volatile anesthetics whose effect is to inhibit actin-based dynamics either directly or indirectly in both neurons and nonneuronal cells. As a target for volatile anesthetics, the actin cytoskeleton offers not just one more putative site of action for these compounds but a mechanism quite different from those that have been proposed previously. In the central nervous system, actin is highly concentrated in dendritic spines (13, 26–28), which form the postsynaptic contact sites for more than 80% of all excitatory synapses (29–32). Current estimates for the human brain indicate that there are more than 1013 excitatory spine synapses in the cerebral cortex alone, with further large numbers in other areas including the cerebellum, basal ganglia, and olfactory bulb (33). Local concentrations of actin filaments are thus widely distributed throughout the brain at vast numbers of synapses while at the same time functioning as a single class of synaptic target sites for volatile anesthetics.
There are several ways in which volatile anesthetics might influence central nervous system activity by blocking spine motility. One possibility is through a feedback effect on neurotransmitter receptors, because drugs that modulate actin polymerization have, for example, been shown to influence N-methyl-d-aspartate-type glutamate receptor channels (34). By blocking changes in spine shape, anesthetics might interfere with mechanisms that regulate electrical or biochemical coupling between the spine head and the shaft of the dendrite, which modulate the postsynaptic effects of excitatory transmission (33, 35, 36). Finally, changes in spine morphology have been associated with alterations in the extent of the contact zone at the synaptic junction (37–40). As a target for volatile anesthetics, the actin cytoskeleton may also explain some of their undesirable side effects (41, 42), several of which involve actin-based mechanisms including cardiac muscle contractility (43), lymphocyte motility (44), and cell division (45).
Spine morphology has not previously been considered as potentially involved in anesthetic action because the rapidity with which it can change was only recently established (12). Nevertheless, it had been suggested that actin in dendritic spines might mediate twitching movements involved in a form of ultra-short-term memory (46). The sensitivity of spine motility to general anesthetics demonstrated here lends further support to the idea that rapid morphological changes at central synapses contribute to short-term brain function. The anesthetic sensitivity of this form of synaptic plasticity also reveals additional complexity in the mechanisms underlying the anesthetized state.
Supplementary Material
Acknowledgments
We are grateful to U. Wagner for the preparation of the organotypic slice cultures and to A. Welman and J. Hagmann for making the membrane-targeted GFP construct available.
ABBREVIATIONS
- GFP
green fluorescent protein
- GFP-actin
actin tagged with GFP
- PMA
phorbol 12-myristate 13-acetate
Footnotes
This paper was submitted directly (Track II) to the Proceedings Office.
References
- 1.Stoelting R K. Handbook of Pharmacology and Physiology in Anesthetic Practice. Philadelphia: Lippincott–Raven; 1995. [Google Scholar]
- 2.Miller K W. Int Rev Neurobiol. 1985;27:1–61. doi: 10.1016/s0074-7742(08)60555-3. [DOI] [PubMed] [Google Scholar]
- 3.Seeman P. Pharmacol Rev. 1972;24:583–655. [PubMed] [Google Scholar]
- 4.Scholz A, Appel N, Vogel W. Eur J Neurosci Suppl. 1998;10:2547–2556. [PubMed] [Google Scholar]
- 5.Franks N P, Lieb W R. Nature (London) 1994;367:607–614. doi: 10.1038/367607a0. [DOI] [PubMed] [Google Scholar]
- 6.Zhang L, Oz M, Stewart R R, Peoples R W, Weight F F. Br J Pharmacol. 1997;120:353–355. doi: 10.1038/sj.bjp.0700934. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Mihic S J, Ye Q, Wick M J, Koltchine V V, Krasowski M D, Finn S E, Mascia M P, Valenzuela C F, Hanson K K, Greenblatt E P, et al. Nature (London) 1997;389:385–389. doi: 10.1038/38738. [DOI] [PubMed] [Google Scholar]
- 8.Minami K, Wick M J, Stern-Bach Y, Dildy-Mayfield J E, Brozowski S J, Gonzales E L, Trudell J R, Harris R A. J Biol Chem. 1998;273:8248–8255. doi: 10.1074/jbc.273.14.8248. [DOI] [PubMed] [Google Scholar]
- 9.Eckenhof R G, Johansson J J. Pharmacol Rev. 1997;49:343–367. [PubMed] [Google Scholar]
- 10.Hasinoff B B, Davey J P. Biochem J. 1989;258:101–107. doi: 10.1042/bj2580101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Sachsenheimer W, Pai E F, Schulz G E, Schirmer R H. FEBS Lett. 1977;79:310–312. doi: 10.1016/0014-5793(77)80809-0. [DOI] [PubMed] [Google Scholar]
- 12.Fischer M, Kaech S, Knutti D, Matus A. Neuron. 1998;20:847–854. doi: 10.1016/s0896-6273(00)80467-5. [DOI] [PubMed] [Google Scholar]
- 13.Kaech S, Fischer M, Doll T, Matus A. J Neurosci. 1997;17:9565–9572. doi: 10.1523/JNEUROSCI.17-24-09565.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.McKenzie D, Franks N P, Lieb W R. Br J Pharmacol. 1995;115:275–282. doi: 10.1111/j.1476-5381.1995.tb15874.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Kaech S, Kim J B, Cariola M, Ralston E. Brain Res Mol Brain Res. 1996;35:344–348. doi: 10.1016/0169-328x(95)00238-n. [DOI] [PubMed] [Google Scholar]
- 16.Parsons C G, Panchenko V A, Pinchenko V O, Tsyndrenko A Y, Krishtal O A. Eur J Neurosci. 1996;8:446–454. doi: 10.1111/j.1460-9568.1996.tb01228.x. [DOI] [PubMed] [Google Scholar]
- 17.Mackay D J, Nobes C D, Hall A. Trends Neurosci. 1995;18:496–501. doi: 10.1016/0166-2236(95)92773-j. [DOI] [PubMed] [Google Scholar]
- 18.Hall A. Science. 1998;279:509–514. doi: 10.1126/science.279.5350.509. [DOI] [PubMed] [Google Scholar]
- 19.Krnjevic K. Gen Pharmacol. 1992;23:965–975. doi: 10.1016/0306-3623(92)90274-n. [DOI] [PubMed] [Google Scholar]
- 20.Pancrazio J J. J Physiol (London) 1996;494:91–103. doi: 10.1113/jphysiol.1996.sp021478. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Harrison N L, Kugler J L, Jones M V, Greenblatt E P, Pritchett D B. Mol Pharmacol. 1993;44:628–632. [PubMed] [Google Scholar]
- 22.Violet J M, Downie D L, Nakisa R C, Lieb W R, Franks N P. Anesthesiology. 1997;86:866–874. doi: 10.1097/00000542-199704000-00017. [DOI] [PubMed] [Google Scholar]
- 23.Jenkins A, Franks N P, Lieb W R. Br J Pharmacol. 1996;117:1507–1515. doi: 10.1111/j.1476-5381.1996.tb15314.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Langmoen I A, Larsen M, Berg-Johnsen J. Eur J Anaesthesiol. 1995;12:51–58. [PubMed] [Google Scholar]
- 25.Harrison N L. Anesthesiology. 1998;88:566–568. doi: 10.1097/00000542-199803000-00003. [DOI] [PubMed] [Google Scholar]
- 26.Matus A, Ackermann M, Pehling G, Byers H R, Fujiwara K. Proc Natl Acad Sci USA. 1982;79:7590–7594. doi: 10.1073/pnas.79.23.7590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Fifkova E. Brain Res. 1985;356:187–215. [PubMed] [Google Scholar]
- 28.Cohen R S, Chung S K, Pfaff D W. Cell Mol Neurobiol. 1985;5:271–284. doi: 10.1007/BF00711012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Gray E G. Nature (London) 1959;183:1592–1593. doi: 10.1038/1831592a0. [DOI] [PubMed] [Google Scholar]
- 30.Uchizono K. Nature (London) 1965;207:642–643. doi: 10.1038/207642a0. [DOI] [PubMed] [Google Scholar]
- 31.Harris K M, Kater S B. Annu Rev Neurosci. 1994;17:341–371. doi: 10.1146/annurev.ne.17.030194.002013. [DOI] [PubMed] [Google Scholar]
- 32.Shepherd G M. J Neurophysiol. 1996;75:2197–2210. doi: 10.1152/jn.1996.75.6.2197. [DOI] [PubMed] [Google Scholar]
- 33.Shepherd G M, Koch C. In: The Synaptic Organization of the Brain. Shepherd G M, editor. Oxford, U.K.: Oxford Univ. Press; 1998. pp. 1–36. [Google Scholar]
- 34.Rosenmund C, Westbrook G L. Neuron. 1993;10:805–814. doi: 10.1016/0896-6273(93)90197-y. [DOI] [PubMed] [Google Scholar]
- 35.Denk W, Yuste R, Svoboda K, Tank D W. Curr Opin Neurobiol. 1996;6:372–378. doi: 10.1016/s0959-4388(96)80122-x. [DOI] [PubMed] [Google Scholar]
- 36.Koch C, Zador A. J Neurosci. 1993;13:413–422. doi: 10.1523/JNEUROSCI.13-02-00413.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Carlin R K, Siekevitz P. Proc Natl Acad Sci USA. 1983;80:3517–3521. doi: 10.1073/pnas.80.11.3517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Siekevitz P. Proc Natl Acad Sci USA. 1985;82:3494–3498. doi: 10.1073/pnas.82.10.3494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Geinisman Y, Morrell F, de Toledo-Morrell L. Brain Res. 1989;480:326–329. doi: 10.1016/0006-8993(89)90201-1. [DOI] [PubMed] [Google Scholar]
- 40.Calverley R K, Jones D G. Brain Res Brain Res Rev. 1990;15:215–249. doi: 10.1016/0165-0173(90)90002-6. [DOI] [PubMed] [Google Scholar]
- 41.Wolfson B, Hetrick W D, Lake C L, Siker E S. Anesthesiology. 1978;48:187–190. [PubMed] [Google Scholar]
- 42.Nunn J F. Acta Anaesthesiol Belg. 1975;26:148–154. [PubMed] [Google Scholar]
- 43.Brown B R, Crout J R. Anesthesiology. 1971;34:236–245. doi: 10.1097/00000542-197103000-00007. [DOI] [PubMed] [Google Scholar]
- 44.Nunn J F, Sharp J A, Kimball K L. Nature (London) 1970;226:85–86. doi: 10.1038/226085a0. [DOI] [PubMed] [Google Scholar]
- 45.Sturrock J E, Nunn J F. Anesthesiology. 1975;43:21–33. doi: 10.1097/00000542-197507000-00004. [DOI] [PubMed] [Google Scholar]
- 46.Crick F. Trends Neurosci. 1982;5:44–46. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
{kind=link}
{kind=link}