

Abstract
The scurfy mutant mouse strain suffers from a fatal lymphoproliferative disease leading to early death within 3–4 wk of age. A frame-shift mutation of the forkhead box transcription factor Foxp3 has been identified as the molecular cause of this multiorgan autoimmune disease. Foxp3 is a central control element in the development and function of regulatory T cells (T reg cells), which are necessary for the maintenance of self-tolerance. However, it is unclear whether dysfunction or a lack of T reg cells is etiologically involved in scurfy pathogenesis and its human correlate, the IPEX syndrome. We describe the generation of bacterial artificial chromosome–transgenic mice termed “depletion of regulatory T cell” (DEREG) mice expressing a diphtheria toxin (DT) receptor–enhanced green fluorescent protein fusion protein under the control of the foxp3 gene locus, allowing selective and efficient depletion of Foxp3+ T reg cells by DT injection. Ablation of Foxp3+ T reg cells in newborn DEREG mice led to the development of scurfy-like symptoms with splenomegaly, lymphadenopathy, insulitis, and severe skin inflammation. Thus, these data provide experimental evidence that the absence of Foxp3+ T reg cells is indeed sufficient to induce a scurfy-like phenotype. Furthermore, DEREG mice will allow a more precise definition of the function of Foxp3+ T reg cells in immune reactions in vivo.
Naturally occurring CD25+CD4+ regulatory T cells (T reg cells) are currently intensively characterized because of their major importance in modulating host responses to tumors and infections, in preventing transplant rejection, and in inhibiting the development of autoimmunity and allergy (1–3). Originally, CD4+ T reg cells were identified exclusively by the constitutive expression of CD25, and many in vivo experiments have been performed using depleting antibodies directed against CD25 (4, 5). However, both the existence of CD25− T reg cells, especially within peripheral tissues (1, 6, 7), as well as the expression of CD25 on activated conventional T cells, which precludes discrimination between T reg cells and activated conventional T cells, limits the interpretation of data obtained by the use of anti-CD25 depleting antibodies (7–9). The most specific T reg cell marker currently known is the forkhead box transcription factor Foxp3, which has been shown to be expressed specifically in mouse CD4+ T reg cells and acts as a master switch in the regulation of their development and function (10). Moreover, activated conventional mouse CD4+ T cells or differentiated Th1/Th2 cells fail to induce Foxp3 expression (11, 12).
A key role of Foxp3 in the development of natural T reg cells has been suggested from the molecular characterization of the scurfy mouse mutant. These mice, which suffer from a fatal lymphoproliferative disorder, harbor a mutated foxp3 gene coding for a product that lacks the forkhead domain (13). Scurfy mice receiving CD25+CD4+ T reg cells from WT mice remained virtually disease free (11). Furthermore, transduction of mutant Foxp3, lacking the forkhead domain, failed to confer suppressive activity to naive CD4+ T cells, in contrast to full-length Foxp3 (12). Thus, it is widely accepted that an intrinsic T cell failure to generate functional CD4+ T reg cells is the main cause of the fatal autoimmune disease in scurfy mice, although an additional role of the scurfy mutation in nonhematopoietic cells has been suggested (14, 15). Moreover, it remains unknown whether the mere absence of functional Foxp3+ T reg cells is sufficient to provoke the development of the scurfy phenotype or whether those CD4+ T cells expressing the truncated Foxp3 protein play a more active role, because depletion of T reg cells with established protocols has failed to reproduce the fulminant natural disease (4, 5). The latter hypothesis is supported by the fact that scurfy disease could be transplanted into nude mice through adoptive transfer of CD4+ T cells (16). Furthermore, Foxp3+ T reg cells preferentially recognize self-antigens with high affinity (17, 18), and activated, not naive, T cells in Foxp3-deficient mice preferentially use TCRs found in the TCR repertoire of T reg cells in Foxp3-sufficient mice (19). Collectively, these data led to the hypothesis that these “would-be” T reg cells expressing self-reactive TCRs may contribute to the pathology of Foxp3-deficient mice (19).
To address the question of the in vivo role of T reg cells in immunopathology, we have generated bacterial artificial chromosome (BAC)–transgenic mice termed “depletion of regulatory T cell” (DEREG) mice, which express a diphtheria toxin receptor (DTR) enhanced GFP (eGFP) fusion protein under the control of the foxp3 locus, allowing both detection and inducible depletion of Foxp3+ T reg cells. Phenotypic characterization of DEREG mice not only revealed an eGFP expression pattern similar to that of previously published Foxp3 reporter mice (20, 21) but also showed that DT treatment, in contrast to conventional T reg cell depletion strategies, allows for efficient and selective depletion of Foxp3+ cells without affecting CD25+ effector T cells. Interestingly, ablation of Foxp3+ T reg cells in newborn DEREG mice led to the development of scurfy-like symptoms, thus providing direct evidence that Foxp3+ T reg cells are critically involved in controlling homeostasis of the immune system and in preventing the development of autoimmune diseases.
RESULTS AND DISCUSSION
Generation of BAC-transgenic DEREG mice
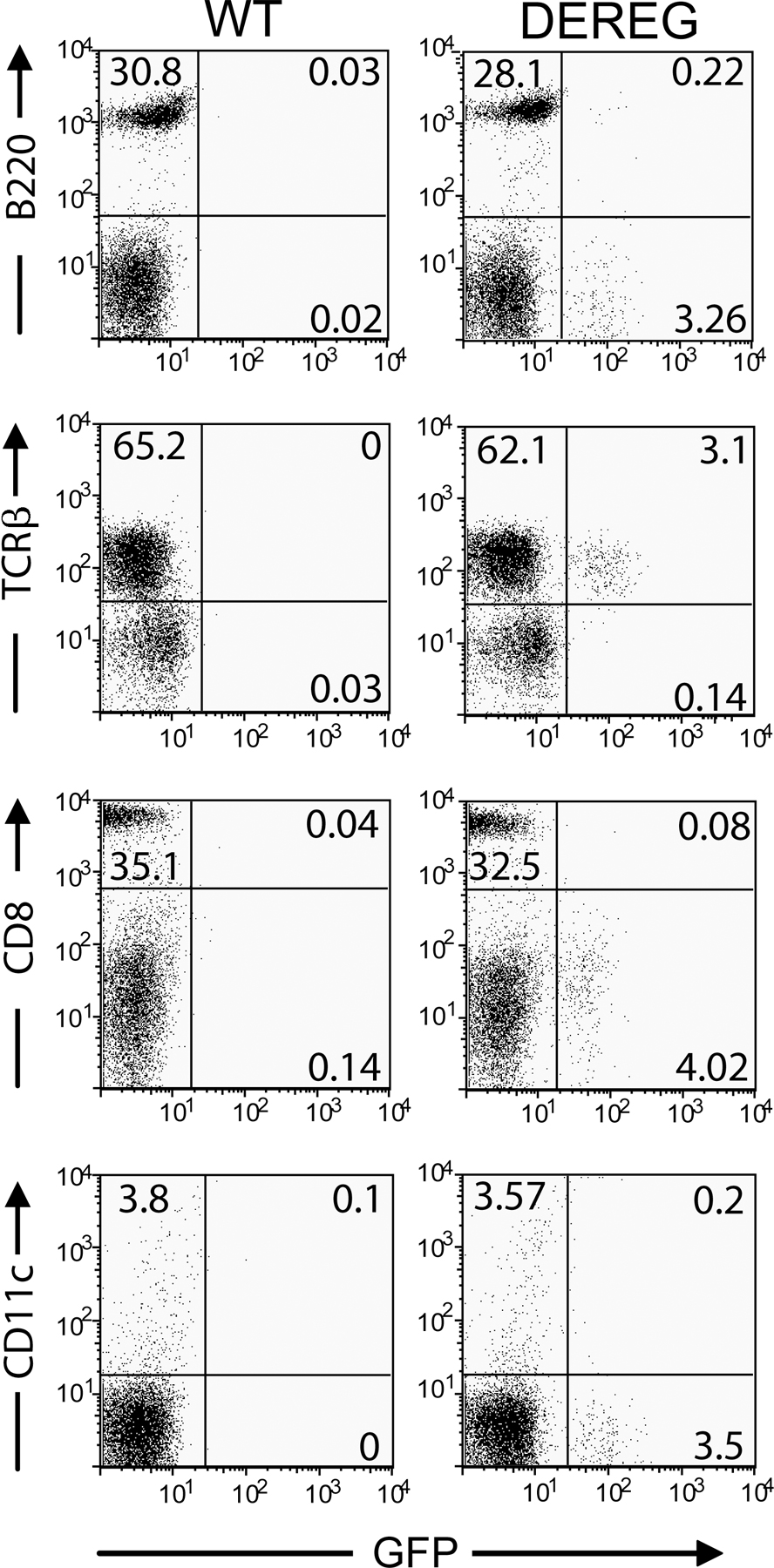
BACs are large fragments of genomic DNA cloned into bacterial vectors that allow for stable propagation in bacteria. Given the large size of BACs, most if not all regulatory sequences of a gene are present and can be used to direct faithful and tissue-specific expression of heterologous genes in vivo in BAC-transgenic mice. We modified a BAC containing the foxp3 locus by introducing the gene coding for a DTR-eGFP fusion protein into the first exon of the foxp3 gene (Fig. 1 A). The modified 150-kb BAC was injected into the pronuclei of fertilized C57BL/6 oocytes. Transgenic mouse lines with high transgene expression were established. The transgenic mice were termed DEREG mice. Analysis of DEREG mice revealed selective DTR-eGFP expression within the CD4+ T cell compartment, with the highest expression within the CD25+ subset (Fig. 1 B). Importantly, similar frequencies and total numbers of CD25+CD4+ T reg cells were observed between DEREG and WT mice (Fig. 1 C and not depicted), and Foxp3 levels in CD25+CD4+ T reg cells from DEREG mice were comparable to T reg cells from WT mice (Fig. 1 D), ruling out the possibility that the BAC transgene influences regulation of the endogenous foxp3 locus. Histological and flow cytometry analysis of spleen, thymus, and LN revealed that the BAC transgene, encoding the DTR-eGFP fusion protein, is faithfully expressed in Foxp3+CD4+ T cells and is absent from thymic epithelial as well as B220+ and CD8+ cells (Fig. 2 and Fig. S1, available at http://www.jem.org/cgi/content/full/jem.20061852/DC1). Furthermore, no gross alteration in the frequency and total number of CD4+ and CD8+ T cells, B220+ B cells, and CD11c+ DCs was observed in LNs and spleen of DEREG mice when compared with WT controls (Fig. S1 and not depicted). In addition, CD25+CD4+ T reg cells from DEREG and WT mice displayed a similar in vitro suppressive capacity (not depicted). Collectively, DEREG mice showed specific expression of the DTR-eGFP fusion protein in fully functional Foxp3+CD4+ T reg cells.
Figure 1.

DEREG mice specifically express DTR-eGFP in T reg cells. (A) Map of BAC construct used for generation of transgenic mice. I and XI indicate the positions of exons I and XI of the foxp3 gene. 24 bp of exon I were replaced by the gene coding for DTR-eGFPpA by homologous recombination with a 1-kb 5′ and 3′ homologous sequence (Boxes A and B). (B) Transgene expression is specific for CD25+CD4+ T cells in naive DEREG mice. Flow cytometric analysis of live-gated CD4+ LN cells of the indicated genotype reveals GFP expression mainly in CD25+CD4+ T cells. (C) Analysis of CD25+CD4+ T cell subsets in LNs from DEREG mice and WT littermates. Plots show similar frequencies of the cell subset in live-gated cells. (D) Foxp3 and GFP expression of LN cells. Flow cytometric analysis reveals a specific transgene expression in T reg cells. Cells are live gated. The percentage of cells in each quadrant (B–D) is indicated.
Figure 2.

Localization of Foxp3+ cells in the spleen and thymus. (A) Spleen sections were stained with αB220-allophycocyanin (blue), αCD3–Alexa Fluor 555 (red), and αGFP–Alexa Fluor 488 (green). (B) Thymus sections were stained with αCD3–Alexa Fluor 647 (blue), αFoxp3–Alexa Fluor 555 (red), and αGFP–Alexa Fluor 488 (green). Bars: (A and B, left) 500 μm; (A, right) 100 μm; (B, right) 50 μm.
DT injection allows selective depletion of Foxp3+ T reg cells
Next we assessed whether injection of DT leads to selective depletion of Foxp3+ T reg cells in vivo. To achieve this, 6-wk-old DEREG mice were injected with 1 μg DT per mouse for six consecutive days. On day 7, we analyzed Foxp3 expression in various lymphoid organs. Among splenocytes and LN-derived cells, we observed an almost complete obliteration in the frequency and absolute number of Foxp3+CD4+ T cells in DT-treated DEREG mice compared with untreated DEREG mice or WT control mice (Fig. 3 A). A similar degree of depletion was observed in the thymus and in Peyer's patches (Fig. 3 B and not depicted). The daily dose of 1 μg DT depleted up to 90% of Foxp3+ T reg cells after 7 d. Importantly, DT treatment did not affect other lymphocyte subsets (Fig. 3 and not depicted), suggesting that DT treatment of DEREG mice allows for the selective depletion of Foxp3+ cells.
Figure 3.

Depletion of Foxp3+ cells with DT in DEREG mice. After six consecutive days of DT injection (1 μg/day), mice were killed on day 7, and lymphoid organs were removed. (A) Flow cytometric analysis of LN-derived cells and splenocytes shows efficient depletion of Foxp3+ cells in DT-treated DEREG mice compared with DT-treated WT control mice. The percentage of cells in each quadrant is indicated. (B) Foxp3 staining (brown) of thymic and Peyer's patch (PP) sections shows effective depletion of Foxp3+ cells in DT-treated compared with untreated DEREG mice. Bar, 100 μm.
Selective depletion of T reg cells leads to an enhanced and prolonged delayed-type hypersensitivity (DTH) response
Previous attempts to analyze the role of CD4+ T reg cells in vivo have used anti-CD25 depleting antibodies (1). In those experiments, the antibodies remained in the circulation for a considerable time, and recent evidence has indicated that CD25+ effector T cells are also affected (unpublished data). In this report, we investigated whether selective depletion of Foxp3+ T reg cells in DEREG mice would provide a method to analyze the role of CD4+ T reg cells during an established immune response. We chose a DTH model in which T reg cells were depleted after hapten-specific sensitization during the phase when CD25+ effector T cells driving the DTH response were generated (22).
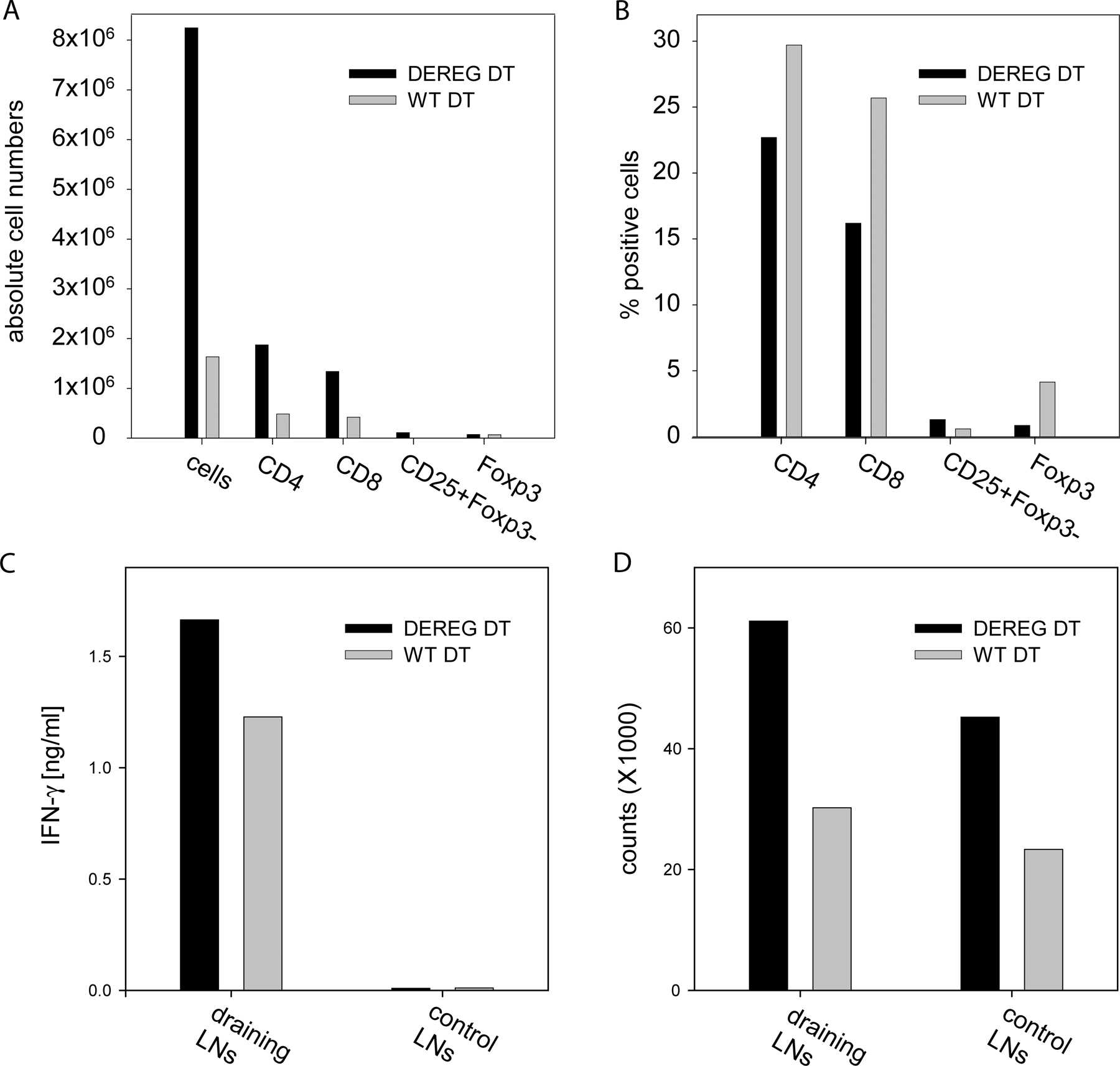
Both untreated DEREG and WT control mice developed a weak DTH response after challenge with the hapten, as indicated by footpad swelling. DT treatment resulted in a substantially increased DTH response in DEREG mice 24, 48, and 72 h after challenge, suggesting that Foxp3+ T reg cells are involved in the regulation of the local inflammatory response (Fig. 4 A and not depicted). The increased footpad swelling in DT-treated DEREG mice was accompanied by an increased cellular immune response within the draining LN (Fig. S2, available at http://www.jem.org/cgi/content/full/jem.20061852/DC1). In contrast to the increased footpad swelling after DT treatment in DEREG mice, WT control mice receiving anti-CD25 antibodies showed a weak DTH response comparable to challenged mice without T reg cell depletion (Fig. 4 A). Analysis of CD4+ T cells from peripheral blood before challenge revealed a very efficient depletion of both CD25+ and CD25−GFP+ T reg cells in DT-treated DEREG mice, whereas a sufficient fraction of CD25+GFP− cells remained (Fig. 4 B). Anti-CD25 antibody treatment completely removed CD25+ cells from circulation, suggesting that this treatment not only led to the depletion of CD25+CD4+ T reg cells but also removed CD25+ effector T cells (Fig. 4 C). Additionally, the same treatment has recently been shown to efficiently deplete CD25+CD4+ T reg cells but not CD25−Foxp3+ T reg cells (23). Therefore, our data suggest that selective removal of total Foxp3+ T reg cells by the use of DEREG mice represents a major progress in the analysis of T reg cell function during the effector phase of an ongoing immune response.
Figure 4.

Depletion of Foxp3+ cells in DEREG mice results in enhanced DTH reaction. (A) Differences in thickness between challenged and PBS-injected footpads in the indicated groups of mice 24 h after challenge. Horizontal lines represent the means. (B) Flow cytometric analysis of CD25 and eGFP expression in live-gated peripheral blood CD4+ T cells after DT treatment of DEREG mice (before challenge) and in untreated DEREG control mice reveals that, after depletion, most of the remaining CD25+CD4+ T cells are GFP negative. (C) Frequency of peripheral blood CD25+CD4+ T cells before challenge. Plots show the absence of double-positive cells in mice that received 500 μg αCD25 and a reduction of CD25+CD4+ T cells in DEREG mice injected with 5 × 1 μg DT (bottom). Control groups show the normal frequency of CD25+CD4+ T cells (top). Cells were live gated. Data are representative of one out of three independent experiments with five mice per group. The percentage of cells in each quadrant (B and C) is indicated.
Neonatal depletion of Foxp3+ T reg cells leads to the development of scurfy-like symptoms
Lack of functional T reg cells has been suggested to cause the fatal lymphoproliferative disease in scurfy mice. The DEREG mice described in this report have now allowed us to assess whether diminution of Foxp3+ T reg cells is sufficient to induce scurfy-like symptoms or whether dysfunctional T reg cells expressing the truncated Foxp3 protein contribute to pathology.
The scurfy mutation is manifested by several clinical disorders and is evident primarily in the lymphoid organs and skin 14 d after birth (24). The disease is characterized by runting; scaliness and crusting of the eyelids, ears, and tail; gross internal lesions including marked splenomegaly; enlarged LNs; and severe anemia. These symptoms result in a wasting syndrome that generally leads to death by 15–24 d of age (24). When newborn DEREG mice were treated with 500 ng DT i.p. (or 100 ng DT s.c.) followed by a second DT injection on day 7, which led to an almost complete depletion of Foxp3+CD4+ T cells (Fig. S3, available at http://www.jem.org/cgi/content/full/jem.20061852/DC1), these mice developed an aggressive lymphoproliferative autoimmune syndrome strongly resembling the observed pathology in scurfy mice. The spleens of DT-treated DEREG mice were significantly enlarged and resembled spleens from age-matched scurfy mice (Fig. 5 A). Histologically, they displayed a disrupted architecture with hyperplasia of the white and red pulp (not depicted). DT-treated DEREG mice showed markedly enlarged LNs as well as increased total cell numbers when compared with both DT-treated WT control and scurfy mice (Fig. 5 B). Histological analysis of DT-treated DEREG mice revealed massive inflammatory infiltrates in various organs (Fig. 5 C). The skin overlying the hyaline cartilage of the ear was extensively thickened with epidermal hyperplasia and a dense infiltrate of lymphocytes—including large numbers of CD3+ T cells (Fig. S4)—with admixed neutrophils and macrophages in the dermis compared with DT-treated WT control mice. The lungs of DT-treated DEREG mice showed similar infiltrates as scurfy mice with peribronchial and perivascular distribution when compared with the normal lung with thin and delicate alveolar walls in control mice. The acini of the pancreas displayed infiltrates with destruction of the islets (insulitis), and the liver exhibited portal aggregates (Fig. 5 C). Because lesions in DT-treated DEREG mice closely resembled those observed in scurfy mice and no significant differences in the pathology of those particular organs were observed, we concluded that diminution of CD4+ T reg cells is sufficient to induce the lethal lymphoproliferative autoimmune syndrome and that an active contribution of pathogenic autoreactive CD4+ T cells expressing the truncated Foxp3 protein is not necessarily required (19). Indeed, immunohistochemical staining for Foxp3 showed that in the thymi and spleens of DT-treated DEREG mice the vast majority of Foxp3+ T reg cells were absent (Fig. 5 D and not depicted).
Figure 5.

Depletion of Foxp3+ cells in neonate DEREG mice results in a scurfy-like phenotype. Neonates were injected i.p. on days 1 and 7 after birth with 500 ng DT and analyzed after 21 d. (A) Splenomegaly in scurfy mice (top) and depleted DEREG mice (middle) compared with a DT-injected WT littermate (bottom). (B) Enlargement and increase of LN cellularity (left, scapular LN; right, inguinal LN; the mean of total cell numbers from four pooled peripheral LN is shown) in scurfy and depleted DEREG mice compared with DT-treated WT littermates. (C) Histological analysis of different organs. Figures show hematoxylin and eosin staining of the indicated organs in the different mice. Bar, 100 μm. (D) Foxp3 staining (brown) of thymus sections shows remaining Foxp3+ expression only in WT littermates. Data are representative of 1 out of 10 independently analyzed mice in which a total of 12 organs each (spleen, LN, Peyer's patches, thymus, liver, pancreas, salivary gland, heart, lung, kidney, gastrointestinal tract, and skin) was evaluated. Bar, 50 μm.
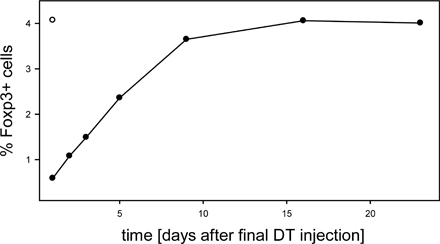
In contrast to depletion of Foxp3+ T reg cells by DT injection, which consistently resulted in the development of scurfy-like symptoms, T reg cell depletion in neonatal mice with anti-CD25 antibodies resulted in marginal development of autoimmune disease, if at all (4, 5). These discrepancies are again best elucidated by both the existence of CD25−Foxp3+ T reg cells, especially within peripheral tissues (1, 6, 7), as well as the induced expression of CD25 on activated conventional CD4+ T cells, which are critically involved in the development of the scurfy phenotype (16). Interestingly, in adult mice neither T reg cell depletion with anti-CD25 antibodies (8) nor depletion of total Foxp3+ T reg cells by DT treatment (not depicted) resulted in the development of autoimmune diseases. This finding can be explained by the lack of lymphopenia-induced proliferation preventing the activation of autoreactive T cells (5). Furthermore, within 2 wk after the last DT injection, we observed a significant rebound of Foxp3+ T reg cells in adult but not neonatal DEREG mice (Fig. 5 D; Fig. S5, available at http://www.jem.org/cgi/content/full/jem.20061852/DC1; and not depicted), which might participate in the control of pathogenic effector cells.
In summary, the novel BAC-transgenic DEREG mouse is a new tool to analyze Foxp3+ T reg cells in vivo. The previous limitations of antibody-based depletion protocols will be avoided because Foxp3+ T reg cells can be selectively depleted in DEREG mice at any time of the immune response. Thus, the model will allow more precise insights into the role of T reg cells in tumors, autoimmune disease, transplantation, and infection models. Furthermore, depletion of Foxp3+ T reg cells in neonatal DEREG mice also shed some light on the cellular players involved in the development of the fatal lymphoproliferative disease in scurfy mice, because our data unequivocally show that depletion of Foxp3+ cells is sufficient to induce a scurfy- like phenotype.
MATERIALS AND METHODS
Mice.
DEREG and WT C57BL/6 mice were bred at the animal facility of the Institut für Medizinische Mikrobiologie, Immunologie und Hygiene at the Technische Universität München. Scurfy mice (B6.Cg-Foxp3 sf/J) were purchased from Charles River Laboratories. All animal experiments were performed under specific pathogen-free conditions and in accordance with institutional, state, and federal guidelines.
Generation of DEREG mice.
Transgenic mice were generated using BAC technology, as previously described (25, 26), with the following modifications. As a transgene, the coding sequence of the DTR-eGFP fusion protein was used (27). The BAC encoding the complete mouse foxp3 gene locus (RP23-267C15) was obtained from the BACPAC Resources Center at Children's Hospital Oakland Research Institute. In contrast to the published overlap PCR strategy (26), homologous regions named Box A and Box B were ligated to DTR-eGFP via AscI (Box A) or PmeI sites (Box B). The polyA fragment was amplified from the TOPO Tools SV40 pA 3′ element kit (Promega) using primers, adding a SpeI site 5′ and a PmeI site 3′. The WT BAC was recombined using the pLD53.SC1 shuttle vector provided by N. Heintz (The Rockefeller University, New York, NY), gel purified, and injected into the pronuclei of fertilized C57BL/6 oocytes. Two transgenic mouse lines with high transgene expression were established (nos. 16.1 and 23.2). DEREG mice were genotyped by PCR using the primers P442 (5′-CCCAGGTTACCATGGAGAGA-3′) and P443 (5′-GAACTTCAGGGTCAGCTTGC-3′).
Antibodies, staining, and sorting reagents.
The following antibodies and secondary reagents were purchased from eBioscience: αCD4 (H129.19), αCD25 (PC61), αCD8 (53–6.7), αB220 (RA3-6B2), αTCR-β (H57-597), αCD11c (HL3), αCD3ɛ (500A2), αB220 (RA3-6B2), αFoxp3 (FJK-16s), a T reg cell staining kit, streptavidin, and appropriate isotype controls. Rabbit αGFP and FITC-conjugated goat α rabbit polyclonal antibodies were obtained from Invitrogen. The antibody αCD25 (PC61) was produced in our laboratory. For immunohistochemistry, the polyclonal αCD3 obtained from DakoCytomation was used.
Flow cytometry.
Cytometric analysis was performed using a CyAn (DakoCytomation) or a FACSCalibur (BD Biosciences) and FlowJo software (Tree Star, Inc.). Dead cells were excluded by propidium iodide staining or ethidium monoazide (Sigma-Aldrich). Intracellular Foxp3 staining was performed with the PE α mouse Foxp3 staining set (Bioscience), according to the manufacturer's instructions.
Depletion of T reg cells.
To deplete T reg cells, DEREG mice were injected with DT (Merck) diluted in endotoxin-free PBS. In our standard protocol, 1 μg DT was injected i.p. on a daily basis for four to six consecutive days unless otherwise indicated. Newborn mice were injected twice either i.p. with 500 ng or s.c. with 100 ng at days 1 and 7 after birth. Anti-CD25 (PC61) was administered i.p. at 500 μg/mouse.
Microscopy, immunofluorescence staining, histology, and immunohistochemistry.
LNs and spleens from scurfy and DT-treated mice were photographed individually with the same magnification using a stereo microscope (MZ-APO; Leica). Immunofluorescent stainings were performed as previously described (28). Slides were examined under a fluorescence microscope (AxioImager M1; Carl Zeiss MicroImaging, Inc.) equipped with a CCD camera (AxioCam MRm; Carl Zeiss MicroImaging, Inc.) and processed with Axiovision software (Carl Zeiss MicroImaging, Inc.). Hematoxylin and eosin stainings were performed on formalin-fixed and paraffin-embedded organs. Foxp3 and CD3 immunohistochemistry was performed as previously described (29).
DTH model.
DTH reactions were performed as previously described using 2% and 0.1% NP-O-Su (Biosearch Technologies) for sensitization and challenge, respectively (30). For depletion of T reg cells, mice were injected i.p. with 1 μg DT per mouse for five consecutive days, starting on day 2 after sensitization. Alternatively, CD25-expressing cells were depleted via i.p. injection of 500 μg anti-CD25 (PC61) on day 5 after sensitization. Mice were challenged on day 7 after sensitization, and the inflammatory response was determined 24 h later by the measurement of footpad thickness. For in vitro restimulation, mice were killed 48 h after challenge. 4 × 105 cells from draining popliteal and inguinal versus control LNs were stimulated with 3 × 105 modified bone marrow GM-CSF–cultured DCs from naive mice. For modification, DCs were incubated for 3 min at 37°C with 0.02% NP-O-Su. After 4 d, proliferation and IFN-γ production were measured using the CellTiter-Glo Luminescent Cell Viability Assay (Promega) and the DuoSet ELISA (R&D Systems), according to the manufacturer's instructions, respectively.
Online supplemental material.
In Fig. S1, flow cytometry analysis reveals that DTR-eGFP expression in DEREG mice is confined to TCRβ+ T cells and precluded from B220+, CD11c+, and CD8+ cells. Fig. S2 depicts the increased cellular immune response in the local draining LN after DT treatment in hapten-challenged mice. Fig. S3 depicts the degree of depletion of Foxp3+ T reg cells on day 8 in neonatal DEREG mice upon the second DT treatment on day 7. Fig. S4 shows CD3+ T cell infiltrates in different organs of DT-treated DEREG mice compared with scurfy and DT-treated WT control mice. Fig. S5 shows the rebound of Foxp3+ T reg cells in DT-treated DEREG mice. Online supplemental material is available at http://www.jem.org/cgi/content/full/jem.20061852/DC1.
Supplemental Material
Acknowledgments
We thank Karin Mink and Simone Spieckermann for technical assistance. We are grateful to Dr. Nathaniel Heintz for providing the pLD53.SC1 shuttle vector for BAC recombination. We would like to thank Drs. Vineet KewalRamani, Jürgen Ruland, Laura Layland, and Wilfried Ellmeier for their critical reading of the manuscript and scientific discussion.
This work was supported by the Wilhelm Sander Foundation, the Bundesministerium für Bildung und Forschung, and the Deutsche Forschungsgemeinschaft (grants SP 615/3-1; SFB/TR22; and SFB 421, 633, and 650).
The authors have no conflicting financial interests.
J. Huehn and T. Sparwasser contributed equally to this work.
References
- 1.Sakaguchi, S. 2005. Naturally arising Foxp3-expressing CD25+CD4+ regulatory T cells in immunological tolerance to self and non-self. Nat. Immunol. 6:345–352. [DOI] [PubMed] [Google Scholar]
- 2.von Boehmer, H. 2005. Mechanisms of suppression by suppressor T cells. Nat. Immunol. 6:338–344. [DOI] [PubMed] [Google Scholar]
- 3.Coombes, J.L., N.J. Robinson, K.J. Maloy, H.H. Uhlig, and F. Powrie. 2005. Regulatory T cells and intestinal homeostasis. Immunol. Rev. 204:184–194. [DOI] [PubMed] [Google Scholar]
- 4.Taguchi, O., and T. Takahashi. 1996. Administration of anti-interleukin-2 receptor alpha antibody in vivo induces localized autoimmune disease. Eur. J. Immunol. 26:1608–1612. [DOI] [PubMed] [Google Scholar]
- 5.McHugh, R.S., and E.M. Shevach. 2002. Cutting edge: depletion of CD4+CD25+ regulatory T cells is necessary, but not sufficient, for induction of organ-specific autoimmune disease. J. Immunol. 168:5979–5983. [DOI] [PubMed] [Google Scholar]
- 6.Leithauser, F., T. Meinhardt-Krajina, K. Fink, B. Wotschke, P. Moller, and J. Reimann. 2006. Foxp3-expressing CD103+ regulatory T cells accumulate in dendritic cell aggregates of the colonic mucosa in murine transfer colitis. Am. J. Pathol. 168:1898–1909. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Needham, D.J., J.X. Lee, and M.W. Beilharz. 2006. Intra-tumoural regulatory T cells: a potential new target in cancer immunotherapy. Biochem. Biophys. Res. Commun. 343:684–691. [DOI] [PubMed] [Google Scholar]
- 8.Onizuka, S., I. Tawara, J. Shimizu, S. Sakaguchi, T. Fujita, and E. Nakayama. 1999. Tumor rejection by in vivo administration of anti-CD25 (interleukin-2 receptor alpha) monoclonal antibody. Cancer Res. 59:3128–3133. [PubMed] [Google Scholar]
- 9.Sutmuller, R.P., L.M. van Duivenvoorde, A. van Elsas, T.N. Schumacher, M.E. Wildenberg, J.P. Allison, R.E. Toes, R. Offringa, and C.J. Melief. 2001. Synergism of cytotoxic T lymphocyte–associated antigen 4 blockade and depletion of CD25+ regulatory T cells in antitumor therapy reveals alternative pathways for suppression of autoreactive cytotoxic T lymphocyte responses. J. Exp. Med. 194:823–832. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Fontenot, J.D., and A.Y. Rudensky. 2005. A well adapted regulatory contrivance: regulatory T cell development and the forkhead family transcription factor Foxp3. Nat. Immunol. 6:331–337. [DOI] [PubMed] [Google Scholar]
- 11.Fontenot, J.D., M.A. Gavin, and A.Y. Rudensky. 2003. Foxp3 programs the development and function of CD4+CD25+ regulatory T cells. Nat. Immunol. 4:330–336. [DOI] [PubMed] [Google Scholar]
- 12.Hori, S., T. Nomura, and S. Sakaguchi. 2003. Control of regulatory T cell development by the transcription factor Foxp3. Science. 299:1057–1061. [DOI] [PubMed] [Google Scholar]
- 13.Brunkow, M.E., E.W. Jeffery, K.A. Hjerrild, B. Paeper, L.B. Clark, S.A. Yasayko, J.E. Wilkinson, D. Galas, S.F. Ziegler, and F. Ramsdell. 2001. Disruption of a new forkhead/winged-helix protein, scurfin, results in the fatal lymphoproliferative disorder of the scurfy mouse. Nat. Genet. 27:68–73. [DOI] [PubMed] [Google Scholar]
- 14.Godfrey, V.L., J.E. Wilkinson, E.M. Rinchik, and L.B. Russell. 1991. Fatal lymphoreticular disease in the scurfy (sf) mouse requires T cells that mature in a sf thymic environment: potential model for thymic education. Proc. Natl. Acad. Sci. USA. 88:5528–5532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Chang, X., J.X. Gao, Q. Jiang, J. Wen, N. Seifers, L. Su, V.L. Godfrey, T. Zuo, P. Zheng, and Y. Liu. 2005. The Scurfy mutation of FoxP3 in the thymus stroma leads to defective thymopoiesis. J. Exp. Med. 202:1141–1151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Blair, P.J., S.J. Bultman, J.C. Haas, B.T. Rouse, J.E. Wilkinson, and V.L. Godfrey. 1994. CD4+CD8− T cells are the effector cells in disease pathogenesis in the scurfy (sf) mouse. J. Immunol. 153:3764–3774. [PubMed] [Google Scholar]
- 17.Jordan, M.S., A. Boesteanu, A.J. Reed, A.L. Petrone, A.E. Holenbeck, M.A. Lerman, A. Naji, and A.J. Caton. 2001. Thymic selection of CD4+CD25+ regulatory T cells induced by an agonist self-peptide. Nat. Immunol. 2:301–306. [DOI] [PubMed] [Google Scholar]
- 18.Hsieh, C.S., Y. Liang, A.J. Tyznik, S.G. Self, D. Liggitt, and A.Y. Rudensky. 2004. Recognition of the peripheral self by naturally arising CD25+ CD4+ T cell receptors. Immunity. 21:267–277. [DOI] [PubMed] [Google Scholar]
- 19.Hsieh, C.S., Y. Zheng, Y. Liang, J.D. Fontenot, and A.Y. Rudensky. 2006. An intersection between the self-reactive regulatory and nonregulatory T cell receptor repertoires. Nat. Immunol. 7:401–410. [DOI] [PubMed] [Google Scholar]
- 20.Fontenot, J.D., J.P. Rasmussen, L.M. Williams, J.L. Dooley, A.G. Farr, and A.Y. Rudensky. 2005. Regulatory T cell lineage specification by the forkhead transcription factor foxp3. Immunity. 22:329–341. [DOI] [PubMed] [Google Scholar]
- 21.Wan, Y.Y., and R.A. Flavell. 2005. Identifying Foxp3-expressing suppressor T cells with a bicistronic reporter. Proc. Natl. Acad. Sci. USA. 102:5126–5131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Kelley, V.E., D. Naor, N. Tarcic, G.N. Gaulton, and T.B. Strom. 1986. Anti-interleukin 2 receptor antibody suppresses delayed-type hypersensitivity to foreign and syngeneic antigens. J. Immunol. 137:2122–2124. [PubMed] [Google Scholar]
- 23.Peffault de Latour, R., H.C. Dujardin, F. Mishellany, O. Burlen-Defranoux, J. Zuber, R. Marques, J. Di Santo, A. Cumano, P. Vieira, and A. Bandeira. 2006. Ontogeny, function, and peripheral homeostasis of regulatory T cells in the absence of interleukin-7. Blood. 108:2300–2306. [DOI] [PubMed] [Google Scholar]
- 24.Godfrey, V.L., J.E. Wilkinson, and L.B. Russell. 1991. X-linked lymphoreticular disease in the scurfy (sf) mutant mouse. Am. J. Pathol. 138:1379–1387. [PMC free article] [PubMed] [Google Scholar]
- 25.Yang, X.W., P. Model, and N. Heintz. 1997. Homologous recombination based modification in Escherichia coli and germline transmission in transgenic mice of a bacterial artificial chromosome. Nat. Biotechnol. 15:859–865. [DOI] [PubMed] [Google Scholar]
- 26.Sparwasser, T., S. Gong, J.Y. Li, and G. Eberl. 2004. General method for the modification of different BAC types and the rapid generation of BAC transgenic mice. Genesis. 38:39–50. [DOI] [PubMed] [Google Scholar]
- 27.Jung, S., D. Unutmaz, P. Wong, G. Sano, K. De los Santos, T. Sparwasser, S. Wu, S. Vuthoori, K. Ko, F. Zavala, et al. 2002. In vivo depletion of CD11c(+) dendritic cells abrogates priming of CD8(+) T cells by exogenous cell-associated antigens. Immunity. 17:211–220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Eberl, G., and D.R. Littman. 2004. Thymic origin of intestinal alphabeta T cells revealed by fate mapping of RORgammat+ cells. Science. 305:248–251. [DOI] [PubMed] [Google Scholar]
- 29.Loddenkemper, C., J. Maul, E. Berg, H. Stein, M. Zeitz, and R. Duchmann. 2006. Analysis of FOXP3 protein expression in human CD4(+)CD25(+) regulatory T cells at the single-cell level. Eur. J. Immunol. 36:245. [DOI] [PubMed] [Google Scholar]
- 30.Sunday, M.E., J.Z. Weinberger, B. Benacerraf, and M.E. Dorf. 1980. Hapten-specific T cell responses to 4-hydroxy-3-nitrophenyl acetyl. J. Immunol. 125:1601–1605. [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}