

Abstract
Urokinase-type plasminogen activator (uPA) and its receptor (uPAR) play an important role in cell guidance and chemotaxis during normal and pathological events. uPAR is GPI-anchored and the mechanism by which it transmits intracellular polarity cues across the plasma membrane during directional sensing has not been elucidated. The constitutively recycling endocytic receptor Endo180 forms a trimolecular complex with uPAR in the presence of uPA, hence its alternate name uPAR-associated protein. Here, we demonstrate that Endo180 is a general promoter of random cell migration and has a more specific function in cell chemotaxis up a uPA gradient. Endo180 expression was demonstrated to enhance uPA-mediated filopodia production and promote rapid activation of Cdc42 and Rac. Expression of a noninternalizing Endo180 mutant revealed that promotion of random cell migration requires receptor endocytosis, whereas the chemotactic response to uPA does not. From these studies, we conclude that Endo180 is a crucial link between uPA–uPAR and setting of the internal cellular compass.
Keywords: Cdc42; endocytosis; migration; Rac; uPA
Introduction
Chemotaxis is the mechanism that ensures guided relocalization of cells from one site to another and is pivotal within a plethora of biological and pathological processes (Parent and Devreotes, 1999; Jones, 2000; Baggiolini, 2001; Bourne and Weiner, 2002; Weiner, 2002), including the dissemination of tumor cells in malignant disease (Moore, 2001; Muller et al., 2001). For such polarized motility, cell surface receptors must sense directional signals in the external environment and activate the intracellular chemosensory machinery. If the necessary polarity cues are absent or not activated, a cell can only migrate in random directions.
Urokinase-type plasminogen activator (uPA) and its receptor uPAR are strongly linked to cancer cell chemotaxis during invasion and metastasis (Ossowski and Aguirre-Ghiso, 2000; Preissner et al., 2000; Mazar, 2001) and in this scenario uPAR has multiple functions. In particular, uPAR can focus and regulate the proteolytic activity of uPA at the plasma membrane. More intriguingly, uPAR acts as a central cell surface coordinator that transmits migratory and polarity cues into the cell. Because uPAR is a GPI-anchored receptor, its association with transmembrane coreceptors is required for this latter function as well as for the internalization of uPA–uPAR (Blasi and Carmeliet, 2002). Endo180 was recently identified in chemical cross-linking studies as part of a trimolecular cell surface complex with uPA and uPAR (Behrendt et al., 2000). This 180-kD transmembrane glycoprotein is one of the four members of the mannose receptor family (East and Isacke, 2002) and in common with other family members is a constitutively recycling endocytic receptor (Isacke et al., 1990; Howard and Isacke, 2002). Commensurate with its role as a uPAR coreceptor, Endo180 expression in normal tissue is restricted to stromal cells, macrophages, a subset of endothelial cells, and sites of chondrogenic/osteogenic activity with a marked increase in expression in tumor endothelium and breast cancer tissue (Blasi, 1993; East and Isacke, 2002; Schnack Nielsen et al., 2002). Here, we directly addressed the question as to whether Endo180 can regulate uPA–uPAR mediated cell chemotaxis.
Results and discussion
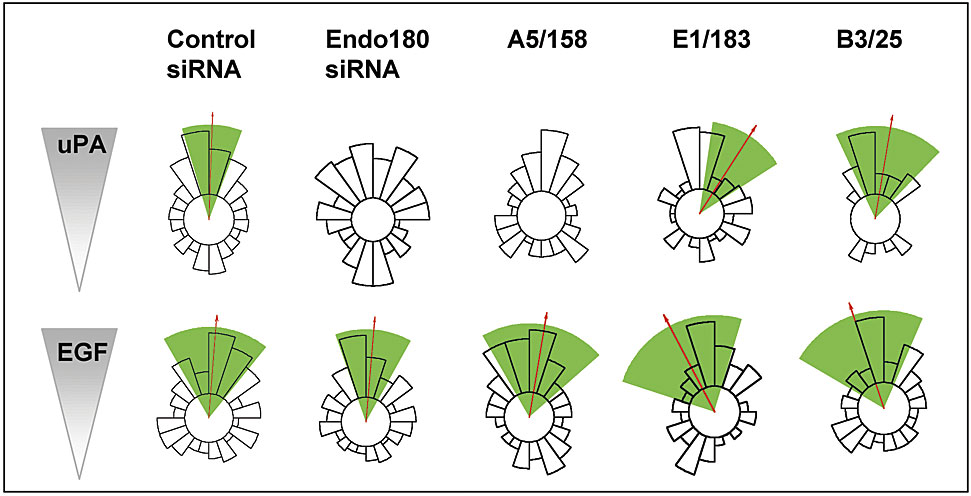
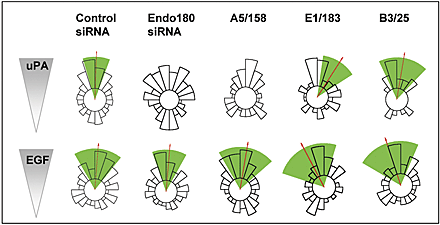
For initial studies, the MDA-MB-231 breast cancer cells were used as they express Endo180, uPAR and EGF receptor, and respond chemotactically to uPA and EGF (Sturge et al., 2002). The staining of Endo180 in MDA-MB-231 cells (Fig. 1) is consistent with previous reports that 10–30% of this receptor is in clathrin-coated pits at the cell surface, whereas 70–90% is located in endosomal compartments (Isacke et al., 1990; Howard and Isacke, 2002). uPAR is also detected in a clustered distribution, although it has a limited overlap with Endo180. Upon uPA stimulation, which is known to promote uPA–uPAR-Endo180 complex formation (Behrendt et al., 2000), Endo180 and uPAR are translocated to the leading edge of polarized cells where they display a substantial degree of colocalization (Fig. 1). To directly investigate the role of Endo180 in these cells, a small interfering RNA (siRNA) approach was taken. Endo180 siRNA treatment resulted in a marked reduction in Endo180 expression (Fig. 2 A; see Fig. 4 C) but did not alter uPAR levels (Fig. 2 A), indicating that expression of these two receptors is not coordinately regulated at the cell surface. This down-regulation of Endo180 had no effect on the basal migratory speed of unstimulated cells but significantly (P < 0.001) reduced the motility of cells in response to uPA or EGF, or when cultured in the presence of a growth factor cocktail (Fig. 2 B). This suggests a general role for Endo180 in the motility of cells stimulated by pro-migratory factors. The most striking alteration in the behavior of Endo180 siRNA treated cells was in their differential ability to respond directionally in chemotactic gradients of uPA and EGF. As expected from previous studies (Sturge et al., 2002), untreated (not depicted) or control siRNA treated MDA-MB-231 cells migrated directionally up gradients of uPA (Fig. 2 C; Video 1, available at http://www.jcb.org/cgi/content/full/jcb.200302124/DC1) and EGF (Fig. 2 C). Endo180 siRNA treatment significantly inhibited the chemotactic response to uPA (Fig. 2 C; Video 2, available at http://www.jcb.org/cgi/content/full/jcb.200302124/DC1) but had no impact on the chemotactic response to EGF (Fig. 2 C). To further investigate the structural interplay of Endo180 in the uPA–uPAR specific response, chemotaxis assays were performed in the presence of Endo180 mAbs E1/183 and A5/158. The four members of the mannose receptor family have a common structural organization of an NH2-terminal cysteine-rich domain, a fibronectin type II domain (FNII) and 8 or 10 C-type lectin-like domains (CTLDs), and single-particle electron microscopy has revealed that in Endo180 the NH2-terminal domains form a hairpin structure such that the cysteine-rich domain contacts CTLD2 (Rivera-Calzada et al., 2003). mAb A5/158 binds to CTLD2 and totally blocks directional migration up a uPA gradient, whereas mAb E1/183, which binds the cysteine-rich domain, partially blocks uPA-induced chemotaxis (Fig. 2 C; Fig. S1, see supplemental methods for a full definition of total and partial blockade, and available at http://www.jcb.org/cgi/content/full/jcb.200302124/DC1). The isotype matched mAb B3/25, which recognizes the endocytic transferrin receptor, had no effect on this response (Fig. 2 C). Together, these findings indicate that a specific biological function of Endo180 is to regulate uPA–uPAR mediated directionality and suggest that accessibility of CTLD2 is required for a full biological response.
Figure 1.

Endogenous Endo180 and uPAR colocalize in response to uPA stimulation. MDA-MB-231 cells were stimulated with or without uPA for 30 min, fixed, permeabilized, and stained. Notably the colocalization of uPAR and Endo180 rich clusters in uPA-stimulated cells occurs in areas of new actin polymerization and membrane ruffling (as determined by costaining of uPAR or Endo180 with phalloidin; not depicted). Bar, 20 μm. Asterisk indicates area of interest magnified five times and shown in inset.
Figure 2.
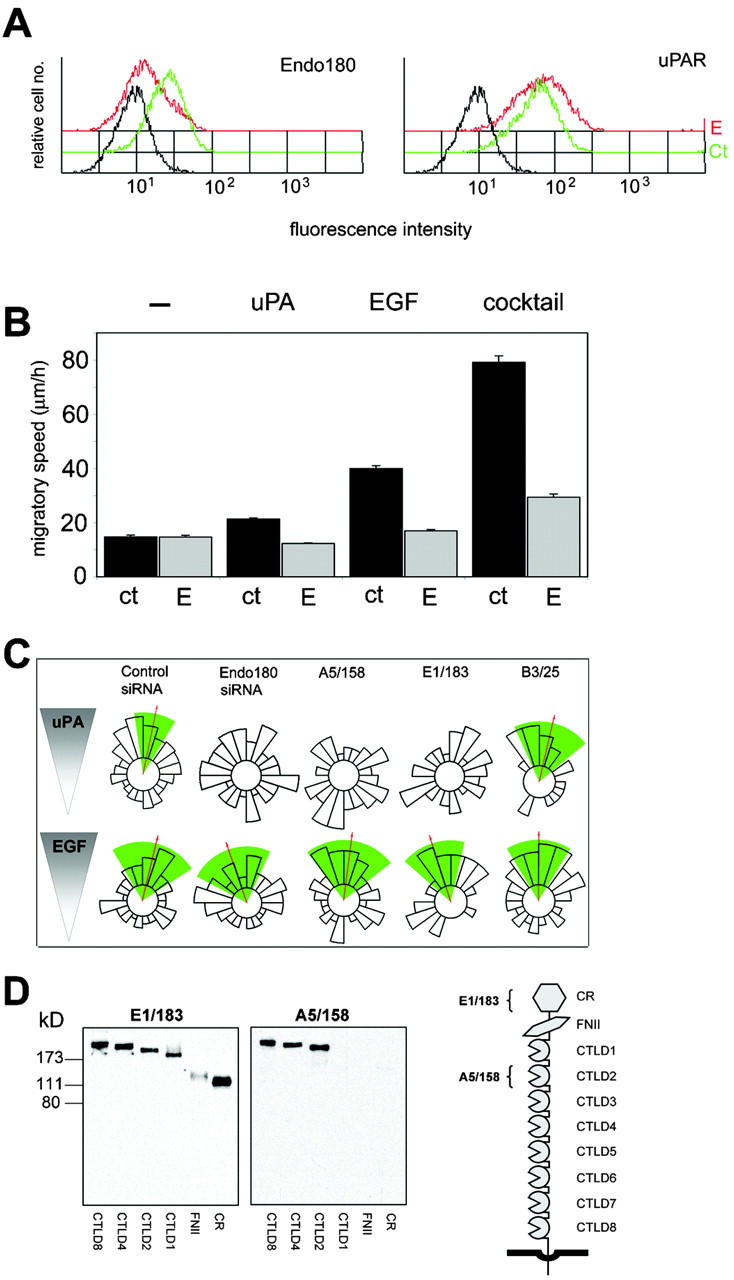
Endo180 is a general requirement for growth factor–stimulated cell motility but a specific promoter of directionality up a uPA gradient. MDA-MB-231 cells were incubated with control or Endo180 siRNA oligonucleotides or left untreated and analyzed after 48 h. (A) FACS® analysis of Endo180 and uPAR expression levels in control (green) and Endo180 (red) siRNA treated cells. Black lines show binding of second antibody alone to control siRNA treated cells. Endo180 expression in control siRNA treated cells was identical to that in untreated cells (not depicted). (B) Mean migratory speed of unstimulated, uPA- or EGF-stimulated cells, or cells cultured in growth factor cocktail (Opti-MEM medium). ct, control siRNA; E, Endo180 siRNA. Migratory speed values are given as the mean speed for all cells analyzed over the 5-h period ± SEM. (C) Chemotactic response of siRNA or mAb treated cells to a gradient of uPA or EGF. (B and C) Data shown includes analysis of >50 cells pooled from at least three separate experiments. (C) Cell directionality was determined using the Rayleigh test and a horizon distance, which included 50% of all cells assayed for each treatment group. Analysis of the same data using 85% of cells is included in Fig. S1. This highly stringent approach discounts any bias in the calculation of directionality that might be caused by alterations in cell speed. Mean direction of cell migration (red arrow)/95% confidence interval (green wedge). (D) Epitope mapping of anti-Endo180 mAbs. Endo180-Fc constructs were resolved by 10% nonreducing SDS-PAGE and subject to Western blotting. The schematic diagram of Endo180 indicates mAb E1/183 binding to the cysteine-rich domain (CR) and mAb A51/58 binding to CTLD2.
Figure 4.
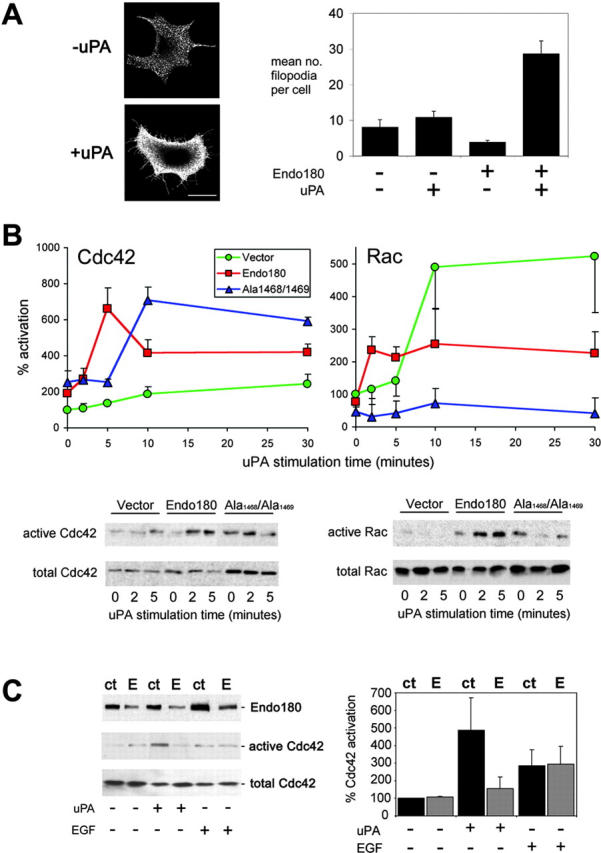
Endo180 promotes filopodia formation in response to uPA and promotes rapid uPA–uPAR mediated activation of Cdc42 and Rac. (A) Endo180 transfected MCF-7 cells were stimulated with or without uPA for 5 min, fixed and immunostained with mAb A5/158 and counterstained with Alexa 555-phalloidin. Bar, 10 μm. Quantitative values shown graphically represent mean ± SEM number of filopodia per cell from a total of 50 randomly chosen cells in two independent experiments. (B) Cdc42 and Rac activation time courses in MCF-7 cells transfected with vector alone (green), Endo180 (red) or Endo180(Ala1468/Ala1469) (blue) and stimulated with uPA. Values represent the mean ± SEM percent change from levels of vector alone cells at zero time point (n = 4–6 experiments). Bottom panels show representative immunoblots. (C) Endo180 expression and Cdc42 activation in siRNA treated MDA-MB-231 cells stimulated with uPA or EGF for 5 min ct, control siRNA; E, Endo180 siRNA. Left panel shows representative blot, right panel shows a graphical representation of mean ± SD percent change from levels in control siRNA treated cells (n = 2 experiments).
To confirm that these effects of Endo180 were not cell type specific or an artifact of the siRNA system, stable populations of the uPAR and EGF receptor positive/Endo180 negative MCF-7 breast cancer cell line transfected with vector alone or Endo180 were generated (Fig. 3 A). In agreement with the data obtained with MDA-MB-231 cells (Fig. 2 B), Endo180 expression promoted a statistically significant (P < 0.0001) twofold increase in random migration compared with vector-alone transfected cells (Fig. 3 B). Although unstimulated parental or vector-alone transfected MCF-7 cells produced small membrane protrusions, they did not translocate on a Matrigel surface (Video 3, available at http://www.jcb.org/cgi/content/full/jcb.200302124/DC1) or a variety of other substrata including fibronectin, collagens I and IV. In contrast, MCF-7 cells expressing Endo180 exhibited increased membrane protrusion formation and translocated randomly on Matrigel (Video 4, available at http://www.jcb.org/cgi/content/full/jcb.200302124/DC1) and the other substrata tested. These observations indicate that Endo180 is both necessary and sufficient to evoke motility in this cell type. Parental or vector-alone transfected MCF-7 cells placed in a gradient of uPA displayed increased but nondirectional migration (Fig. 3, B and C; Video 5, available at http://www.jcb.org/cgi/content/full/jcb.200302124/DC1). The inability of vector-alone cells to detect a chemotactic gradient was consistent at 1, 5, 10, 15, 20, and 25 nM uPA, a concentration range within which the directional migration of MDA-MB-231 cells is maximally stimulated (Sturge et al., 2002). In accordance with the MDA-MB-231 cell data, MCF-7 cells expressing Endo180 were endowed with a sense of direction up a uPA gradient (Fig. 3 C; Video 6, available at http://www.jcb.org/cgi/content/full/jcb.200302124/DC1; P < 0.0001). Furthermore, this directional migration was totally inhibited by the anti-Endo180 mAb A5/158 (Fig. 3 C) and was not associated with altered cell surface levels of uPAR (Fig. 3 A). Conversely, both vector-alone cells and cells expressing Endo180 displayed increased migratory speed (Fig. 3 B) and directionality up an EGF gradient that was unaffected by Endo180 mAbs (Fig. 3 D), further confirming that directional sensing of an EGF gradient is an Endo180-independent process.
Figure 3.

Expression of Endo180 is sufficient to confer a sense of direction up a concentration gradient of uPA. (A) FACS analysis of Endo180 and uPAR cell surface expression levels in MCF-7 cells transfected with vector alone (green), wild-type Endo180 (red), or Endo180(Ala1468/Ala1469) (blue). Profiles in black represent vector transfected cells incubated with secondary antibody alone. (B) Mean migratory speed of transfected MCF-7. The directional data are summarized in a circular histogram showing the number of cells lying within each 18° interval. The mean direction and its 95% confidence interval are represented as a red arrow and green sector, respectively. (C, D) Chemotactic response of untreated or mAb A5/158-treated transfected MCF-7 cells in a gradient of uPA or EGF. Data shown in B–D includes analysis of >70 cells pooled from at least three experiments. Cell directionality was determined using the Rayleigh test and a horizon distance which included 50% of all cells assayed for each treatment group. (C and D) The directional data are summarized in a circular histogram showing the number of cells lying within each 18 degree interval. The mean direction and its 95% confidence interval are represented as a red arrow and green sector, respectively.
Next, we addressed the mechanism by which Endo180 modulated migratory and chemotactic responses by examining MCF-7 cells transfected with an Endo180(Ala1468/Ala1469) construct. This mutant is expressed at the cell surface (Fig. 3 A) but is internalization defective (Howard and Isacke, 2002; Wienke et al., 2003). In contrast to wild-type Endo180, expression of Endo180(Ala1468/Ala1469) did not increase the migratory speed of cells on Matrigel (Fig. 3 B) or other substrata (not depicted), indicating that the random motility evoked by Endo180 is dependent on constitutive recycling of the receptor. Despite an impaired migratory speed (Fig. 3 B), cells expressing Endo180(Ala1468/Ala1469) still displayed strong directional sensing of a uPA gradient (P < 0.0001) and this directionality was totally blocked by mAb A5/158 (Fig. 3 C). Similar to cells expressing wild-type Endo180, there was little effect of Endo180(Ala1468/Ala1469) expression on their migratory speed (Fig. 3 B) or directionality (Fig. 3 D) up a gradient of EGF or on the cell surface expression of uPAR (Fig. 3 A). These results suggest that the ability of Endo180 to coordinate the directionality of a cell toward uPA can be transduced from its localization on the plasma membrane and is separate from its endocytic function.
The intracellular mechanisms for generation of polarity and migration in eukaryotic cells are beginning to be elucidated. Phospholipid and small Rho GTPase signaling pathways converge to regulate recruitment of Arp2/3 complex by WASP/WAVE family scaffolding proteins, which rapidly rearrange the actin cytoskeleton to produce structures that change the polarity and orientation of a cell (Franz et al., 2002; Weiner, 2002). Much less is understood about how extracellular signals are linked to this intracellular machinery. To further elucidate the mechanism of Endo180-dependent chemotaxis, the effect of altered Endo180 expression on the temporal activation of the Rho-family GTPases Cdc42, Rac and RhoA was examined. These molecular switches have fundamental roles in cell migration and chemotaxis (Nobes and Hall, 1995; Allen et al., 1998; Jones, 2000) and their deregulation is strongly implicated in metastasis (Jaffe and Hall, 2002; Sahai and Marshall, 2002). More importantly, Rac and Cdc42 can regulate cell motility downstream of uPAR (Kjoller and Hall, 2001; Sturge et al., 2002). In Endo180, but not vector-alone, expressing MCF-7 cells uPA stimulation increased filopodia production (Fig. 4 A) and caused rapid and sustained activation of Cdc42 (Fig. 4 B). Consistent with these findings, uPA-stimulated activation of Cdc42 in MDA-MB-231 cells was blocked by Endo180 siRNA, but not control siRNA, treatment (Fig. 4 C), whereas EGF-stimulated Cdc42 activation was unaffected by Endo180 siRNA treatment (Fig. 4 C). Despite an impaired activation of Cdc42, vector alone expressing cells show a substantial activation of Rac in response to uPA, which together may account for their increased migration without directional sensing in a uPA gradient. Although these data also suggest that Rac activation is not dependent on Endo180, the rapidity of uPA-dependent Rac activation in Endo180 transfected cells points to a regulatory role for this receptor in stimulating Rac activity. Endo180(Ala1468/Ala1469) expression resulted in delayed Cdc42 activation, but the level of activation stimulated by uPA was similar to that seen in Endo180 expressing cells. In contrast, Rac activation was impaired in Endo180(Ala1468/Ala1469) expressing cells providing a molecular explanation for the ability of these cells to sense a uPA gradient without increasing their migratory speed. Based on these results, we propose that Endo180 plays an important regulatory role in coordinating the activation of the Cdc42 and Rac GTPases during uPA-induced chemotaxis. In agreement with a previous report (Jo et al., 2002), RhoA was not significantly activated by uPA in either the MCF-7 cell lines or MDA-MB-231 cells (unpublished data). uPA–uPAR signaling has been shown to activate the MAPK pathway (Nguyen et al., 1999) and Akt (protein kinase B; Sturge et al., 2002; Chandrasekar et al., 2003). However, wild-type Endo180 or Endo180(Ala1468/Ala1469) expression did not alter the time course of uPA-mediated extracellular signal–regulated kinase 1/2 (ERK1/2) phosphorylation in MCF-7 cells or enhance Akt phosphorylation (Fig. 5 A).
Figure 5.

Endo189 expression does not alter ERK1/2 and Akt phosphorylation or cell surface LRP. (A) ERK1/2 and Akt phosphorylation in MCF-7 cells transfected with vector alone, Endo180 or Endo180(Ala1468/Ala1469) and stimulated with uPA. Blots show phosphorylated ERK1/2 and Akt together with total ERK1/2 and Akt as loading controls. (B) FACS® analysis of LRP cell surface expression levels in MCF-7 cells transfected with vector alone (green), wild-type Endo180 (red) or Endo180(Ala1468/Ala1469) (blue). Profiles in black represent vector transfected cells incubated with secondary antibody alone.
Finally, we considered the possibility that the effect of Endo180 on uPA-mediated cell chemotaxis might, at least in part, result from this constitutively recycling transmembrane receptor modulating the cell surface levels of other uPAR associated proteins. However, Endo180 expression was shown to have no effect on plasma membrane expression of either integrins (Wienke et al., 2003) or low density lipoprotein receptor–related protein (LRP; Fig. 5 B)–uPAR coreceptors, which have been previously demonstrated to promote cell signaling and migration in response to uPA stimulation.
The findings of this paper demonstrate that coexpression of Endo180 and uPAR promotes the guided migration of breast tumor cells. This significantly advances our understanding of how these receptors may participate in metastatic spread and tumor progression. The up-regulation of Endo180 expression in tumor endothelium (St. Croix et al., 2000) indicates a potential role for this receptor in the regulation of tumor neoangiogenesis, a process that is hallmarked by endothelial cell chemotaxis and has a strong dependence on the interaction between uPA and uPAR (Mazar, 2001; Pepper, 2001). In addition to its interaction with uPA–uPAR, Endo180 functions as a Ca2+-dependent mannose binding C-type lectin (East et al., 2002) and as a collagen binding/internalization receptor (Behrendt et al., 2000; Wienke et al., 2003), and binding of these ligands is mediated by CTLD2 and FNII, respectively. Furthermore, the functional interplay between the different ligand binding activities of Endo180 predicted in a recent 3D structural model (Rivera-Calzada et al., 2003), is evidenced by the finding that collagens block the association with uPAR (Behrendt et al., 2000), and as shown here, an antibody directed against the sugar binding CTLD2 blocks cellular responses to uPA. What is apparent from the data presented here is that role of Endo180 in promoting random “nondirectional” cell migration (Figs. 3 and 4; East et al., 2003; Engelholm et al., 2003) is dependent on receptor internalization and thereby mechanistically distinct from the unique function of Endo180 as a cell surface coordinator for setting up the internal cellular compass in a gradient of uPA.
Materials and methods
Reagents and cells
Rabbit anti-Endo180 polyclonal antibody and anti-Endo180 mAbs A5/158 and E1/183 have been described previously (Isacke et al., 1990; Sheikh et al., 2000). B3/25 antitransferrin receptor mAb was a gift from C. Hopkins (Imperial College, London, UK). Mouse anti–human uPAR antibody 3936, (America Diagnostica, Inc.), mouse anti–human-LRP (Serotec), mouse anti–human pan-ERK1/2 (Sigma-Aldrich), mouse anti–human phospho-ERK1/2, rabbit anti–human pan-PKB/AKT, and mouse anti–human phospho-AKT (Upstate Biotechnology). Alexa555 anti–rabbit Ig, Alexa488 anti–mouse Ig and Alexa555-phalloidin (Molecular Probes), and HRP anti–mouse Ig (Jackson ImmunoResearch Laboratories). Catalytically inactive uPA, as used previously (Sturge et al., 2002), was a gift from J. Hamelin (Institut National de la Santé et de la Recherche Medicale U461, France) and EGF (R&D Systems) were used at 10 nM and 15 ng/ml, respectively, in all assays. Extracellular domain Endo180 constructs truncated after the cysteine-rich domain, FNII, CTLD1, CTLD2, CTLD4, or CTLD8 fused in frame with the Fc portion of human Ig were generated and expressed in COS-1 cells as described previously (East et al., 2002). MCF-7 transfectants and siRNA treatment of MDA-MB-231 cells has been described previously (Wienke et al., 2003; see Online supplemental material). MCF-7 cells were maintained in DME + 10% FCS unless otherwise stated. MDA-MB-231 cells were cultured in DME + 10% FCS and starved in DME + 0.1% FCS for 24 h before analysis.
Cdc42, Rac, RhoA, ERK1/2, and Akt activation assays
MCF-7 or MDA-MB-231 cells were grown to 50% confluence and starved for 24 h in DME before treatment. Cells were lysed in ice-cold lysis buffer plus protease and phosphatase inhibitors. Active Cdc42, Rac, and RhoA were measured using assay kits according to the manufacturer's instructions (Upstate Biotechnology). ERK1/2 and Akt phosphorylation were measured by immunoblot analysis with phospho-specific antibodies.
Chemotaxis/migration assay
Chemotaxis and migration were measured by direct observation and recording of cell behavior in stable concentration gradients of 10 nM uPA or 15 ng/ml EGF using the Dunn chemotaxis chamber as described previously (Allen et al., 1998; Sturge et al., 2002). Full details are provided in Online supplemental material.
Online supplemental material
Materials and methods describing the Endo180 siRNA oligonucleotides and methods of transfections, a detailed account of how the Dunn chamber chemotaxis experiments were undertaken, and how the data is analyzed. Fig. S1 shows the same data as shown in Fig. 2 C but using a lower stringency analysis. Videos 1–6 show examples of Dunn chamber experiments. Online supplemental material is available at http://www.jcb.org/cgi/content/full/jcb.200302124/DC1.
Supplemental Material
Acknowledgments
We thank David Robertson for his help with confocal imaging, Dr. Jocelyne Hamelin for providing us with uPA, Dr. Graham Dunn for writing the Mathematica™ Notebooks used to analyze the chemotaxis data, and Prof. Alan Ashworth for his critical advice during the preparation of this manuscript.
This work was funded by Breakthrough Breast Cancer, the Association of International Cancer Research and The Wellcome Trust. L. East was supported by a Medical Research Council studentship.
Justin Sturge and Dirk Wienke contributed equally to this work.
The online version of this article includes supplemental material.
Abbreviations used in this paper: CTLD, C-type lectin-like domain; ERK1/2, extracellular signal–regulated kinase 1/2; FNII, fibronectin type II domain; LRP, low density lipoprotein receptor–related protein; siRNA, small interfering RNA; uPA, urokinase-type plasminogen activator; uPAR, uPA receptor.
References
- Allen, W.E., D. Zicha, A.J. Ridley, and G.E. Jones. 1998. A role for Cdc42 in macrophage chemotaxis. J. Cell Biol. 141:1147–1157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baggiolini, M. 2001. Chemokines in pathology and medicine. J. Intern. Med. 250:91–104. [DOI] [PubMed] [Google Scholar]
- Behrendt, N., O.N. Jensen, L.H. Engelholm, E. Mortz, M. Mann, and K. Dano. 2000. A urokinase receptor-associated protein with specific collagen binding properties. J. Biol. Chem. 275:1993–2002. [DOI] [PubMed] [Google Scholar]
- Blasi, F. 1993. Urokinase and urokinase receptor: a paracrine/autocrine system regulating cell migration and invasiveness. Bioessays. 15:105–111. [DOI] [PubMed] [Google Scholar]
- Blasi, F., and P. Carmeliet. 2002. uPAR: A versatile signalling orchestrator. Nat. Rev. Mol. Cell Biol. 3:932–943. [DOI] [PubMed] [Google Scholar]
- Bourne, H.R., and O. Weiner. 2002. A chemical compass. Nature. 419:21. [DOI] [PubMed] [Google Scholar]
- Chandrasekar, N., S. Mohanam, M. Gujrati, W.C. Olivero, D.H. Dinh, and J.S. Rao. 2003. Downregulation of uPA inhibits migration and PI3K/Akt signaling in glioblastoma cells. Oncogene. 22:392–400. [DOI] [PubMed] [Google Scholar]
- East, L., and C.M. Isacke. 2002. The mannose receptor family. Biochim. Biophys. Acta. 1572:364–386. [DOI] [PubMed] [Google Scholar]
- East, L., A. McCarthy, D. Wienke, J. Sturge, A. Ashworth, and C.M. Isacke. 2003. A targeted deletion in the endocytic receptor gene Endo180 results in a defect in collagen uptake. EMBO Rep. 4:710–716. [DOI] [PMC free article] [PubMed] [Google Scholar]
- East, L., S. Rushton, M.E. Taylor, and C.M. Isacke. 2002. Characterization of sugar binding by the mannose receptor family member, Endo180. J. Biol. Chem. 277:50469–50475. [DOI] [PubMed] [Google Scholar]
- Engelholm, L.H., K. List, S. Netzel-Arnett, E. Cukierman, D.J. Mitola, H. Aaronson, L. Kjoller, J.K. Larsen, K.M. Yamada, D.K. Strickland, et al. 2003. uPARAP/Endo180 is essential for cellular uptake of collagen and promotes fibroblast collagen adhesion. J. Cell Biol. 160:1009–1015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Franz, C.M., G.E. Jones, and A.J. Ridley. 2002. Cell migration in development and disease. Dev. Cell. 2:153–158. [DOI] [PubMed] [Google Scholar]
- Howard, M.J., and C.M. Isacke. 2002. The C-type lectin receptor Endo180 displays internalization and recycling properties distinct from other members of the mannose receptor family. J. Biol. Chem. 277:32320–32331. [DOI] [PubMed] [Google Scholar]
- Isacke, C.M., P. Vandergeer, T. Hunter, and I.S. Trowbridge. 1990. P180, a novel recycling transmembrane glycoprotein with restricted cell type expression. Mol. Cell. Biol. 10:2606–2618. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jaffe, A.B., and A. Hall. 2002. Rho GTPases in transformation and metastasis. Adv. Cancer Res. 84:57–80. [DOI] [PubMed] [Google Scholar]
- Jo, M., K.S. Thomas, A.V. Somlyo, A.P. Somlyo, and S.L. Gonias. 2002. Cooperativity between the Ras-ERK and Rho-Rho kinase pathways in urokinase-type plasminogen activator-stimulated cell migration. J. Biol. Chem. 277:12479–12485. [DOI] [PubMed] [Google Scholar]
- Jones, G.E. 2000. Cellular signaling in macrophage migration and chemotaxis. J. Leukoc. Biol. 68:593–602. [PubMed] [Google Scholar]
- Kjoller, L., and A. Hall. 2001. Rac mediates cytoskeletal rearrangements and increased cell motility induced by urokinase-type plasminogen activator receptor binding to vitronectin. J. Cell Biol. 152:1145–1157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mazar, A.P. 2001. The urokinase plasminogen activator receptor (uPAR) as a target for the diagnosis and therapy of cancer. Anticancer Drugs. 12:387–400. [DOI] [PubMed] [Google Scholar]
- Moore, M.A.S. 2001. The role of chemoattraction in cancer metastases. Bioessays. 23:674–676. [DOI] [PubMed] [Google Scholar]
- Muller, A., B. Homey, H. Soto, N.F. Ge, D. Catron, M.E. Buchanan, T. McClanahan, E. Murphy, W. Yuan, S.N. Wagner, et al. 2001. Involvement of chemokine receptors in breast cancer metastasis. Nature. 410:50–56. [DOI] [PubMed] [Google Scholar]
- Nguyen, D.H., A.D. Catling, D.J. Webb, M. Sankovic, L.A. Walker, A.V. Somlyo, M.J. Weber, and S.L. Gonias. 1999. Myosin light chain kinase functions downstream of Ras/ERK to promote migration of urokinase-type plasminogen activator-stimulated cells in an integrin-selective manner. J. Cell Biol. 146:149–164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nobes, C.D., and A. Hall. 1995. Rho, Rac, and Cdc42 Gtpases regulate the assembly of multimolecular focal complexes associated with actin stress fibers, lamellipodia, and filopodia. Cell. 81:53–62. [DOI] [PubMed] [Google Scholar]
- Ossowski, L., and J.A. Aguirre-Ghiso. 2000. Urokinase receptor and integrin partnership: coordination of signaling for cell adhesion, migration and growth. Curr. Opin. Cell Biol. 12:613–620. [DOI] [PubMed] [Google Scholar]
- Parent, C.A., and P.N. Devreotes. 1999. A cell's sense of direction. Science. 284:765–770. [DOI] [PubMed] [Google Scholar]
- Pepper, M.S. 2001. Role of the matrix metalloproteinase and plasminogen activator-plasmin systems in angiogenesis. Arterioscler Thromb. Vasc. Biol. 21:1104–1117. [DOI] [PubMed] [Google Scholar]
- Preissner, K.T., S.M. Kanse, and A.E. May. 2000. Urokinase receptor: a molecular organizer in cellular communication. Curr. Opin. Cell Biol. 12:621–628. [DOI] [PubMed] [Google Scholar]
- Rivera-Calzada, A., D. Robertson, J.R. MacFayden, J. Boskovic, C.M. Isacke, and O. Llorca. 2003. Three-dimensional interplay among the ligand binding domains of the uPAR-associated protein, Endo180. EMBO Rep. 4:807–812. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sahai, E., and C.J. Marshall. 2002. Rho-GTPases and cancer. Nat. Rev. Cancer. 2:133–142. [DOI] [PubMed] [Google Scholar]
- Schnack Nielsen, B., F. Rank, L.H. Engelholm, A. Holm, K. Dano, and N. Behrendt. 2002. Urokinase receptor-associated protein (uPARAP) is expressed in connection with malignant as well as benign lesions of the human breast and occurs in specific populations of stromal cells. Int. J. Cancer. 98:656–664. [DOI] [PubMed] [Google Scholar]
- Sheikh, H., H. Yarwood, A. Ashworth, and C.M. Isacke. 2000. Endo180, an endocytic recycling glycoprotein related to the macrophage mannose receptor is expressed on fibroblasts, endothelial cells and macrophages and functions as a lectin receptor. J. Cell Sci. 113:1021–1032. [DOI] [PubMed] [Google Scholar]
- St. Croix, B., C. Rago, V. Velculescu, G. Traverso, K.E. Romans, E. Montgomery, A. Lal, G.J. Riggins, C. Lengauer, B. Vogelstein, and K.W. Kinzer. 2000. Genes expressed in human tumor endothelium. Science. 289:1197–1202. [DOI] [PubMed] [Google Scholar]
- Sturge, J., J. Hamelin, and G.E. Jones. 2002. N-WASP activation by a beta 1-integrin-dependent mechanism supports P13K-independent chemotaxis stimulated by urokinase-type plasminogen activator. J. Cell Sci. 115:699–711. [DOI] [PubMed] [Google Scholar]
- Weiner, O.D. 2002. Regulation of cell polarity during eukaryotic chemotaxis: the chemotactic compass. Curr. Opin. Cell Biol. 14:196–202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wienke, D., J.R. MacFayden, and C.M. Isacke. 2003. Identification and characterisation of the endocytic transmembrane glycoprotein Endo180 as a novel collagen receptor. Mol. Biol. Cell. In press. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
{kind=link}
{kind=link}