

Abstract
Lymphotoxin α (LTα) signals via tumor necrosis factor receptors (TNFRs) as a homotrimer and via lymphotoxin β receptor (LTβR) as a heterotrimeric LTα1β2 complex. LTα-deficient mice lack all lymph nodes (LNs) and Peyer's patches (PPs), and yet LTβ-deficient mice and TNFR-deficient mice have cervical and mesenteric LN. We now show that mice made deficient in both LTβ and TNFR type 1 (TNFR1) lack all LNs, revealing redundancy or synergism between TNFR1 and LTβ, acting presumably via LTβR. A complete lack of only PPs in mice heterozygous for both ltα and ltβ, but not ltα or ltβ alone, suggests a similar two-ligand phenomenon in PP development and may explain the incomplete lack of PPs seen in tnfr1 −/− mice.
Keywords: lymphotoxin beta, tumor necrosis factor receptor 1, knockout mice, mesenteric lymph nodes, Peyer's patches
Studies on mice genetically deficient in various secondary lymphoid organs are increasing our understanding of the requirement or otherwise for these highly organized structures in immune function, from antiviral immunity (1) to autoimmunity (2). Hox11 −/− mice lack a spleen (3), whereas aly/aly mutant mice lack LNs and have a disorganized spleen (4, 5). Also, mice made deficient in the putative chemokine receptor BLR1 lack inguinal lymph nodes and fail to form primary B cell follicles in the spleen (6). Our studies have involved members of the TNF receptor and ligand families (7, 8). Studies of TNF family members are not only providing insight into the intricate microarchitecture of immune cell responses in lymphoid organs but also of chronic inflammatory states (9), such as the phenomenon termed lymphoid neogenesis (10).
TNF-α and TNF-β (lymphotoxin α; LTα)1 are the archetypal ligands of a growing family, which includes CD30 ligand (L), CD40L, FasL, TRAIL, and lymphotoxin β (LTβ) (11, 12). LTβ was discovered by virtue of its ability to anchor LTα to the cell surface, without which LTα is secreted as a homotrimer (LTα3) (13, 14). LTα/β complex itself is a trimer with a predominant form (LTα1β2) that binds LTβR, and a minor form (LTα2β1) that binds TNF receptor type 1 (TNFR1) (15–17). Both forms of LTα are produced by activated lymphocytes and NK cells (12, 18).
Historically, LTα3 is known as a factor that causes cytotoxicity and inflammation, and signals via TNFR1 and TNFR2 (9, 19, 20). Although LTα/β complexes do not appear to mediate inflammation (21), pleiotropic effects of LTβR cross-linking are now emerging, including cytotoxicity (17, 22), chemokine induction (23), and integrin upregulation (21). Studies with ltα −/− mice and ltβ −/− mice are beginning to address the in vivo significance of these facets of LTα and LTβ biology (2). However, initial studies of ltα −/− mice were dominated by the unexpected observation of a complete lack of LNs and Peyer's patches (PPs), as well as a disorganized spleen lacking follicular dendritic cells and germinal centers (24–27). Since mice deficient in TNFR have LNs, it had been assumed that the LTα/β complexes were responsible rather than LTα3. However, we recently showed that this explanation was not entirely correct (28). Specifically, we determined that ltβ −/− mice retain mesenteric LNs (MLNs) and to a certain extent, cervical LNs, both of which drain mucosal surfaces. It was therefore a paradox that these LNs are absent in mice that lack the LTα3 ligand and yet they are present in mice that lack the known receptors TNFR1 and TNFR2.
We now report that mice made deficient in both TNFR1 and LTβ lack MLNs. We have thus revealed a redundancy or synergism between TNFR1 and LTβ (presumably signaling via LTβR) that warrants further investigation in other aspects of TNFR1 and LTβ biology. Ltα −/− mice and ltβ −/− mice were derived as littermates by interbreeding, and unambiguously confirmed the lack of MLNs in ltα −/− mice and their presence in ltβ −/− mice. Surprisingly, the latter studies also revealed a complete and specific lack of only PPs in ltα +/− ltβ +/− mice. This presents a unique mouse model for the study of gastrointestinal immunology and suggests that two LTα ligands are involved in PP as well as MLN development, and may explain the incomplete lack of PPs seen in tnfr1 −/− mice.
Materials and Methods
Mice.
ltβ −/− and ltβ +/+ wild-type mice (expanded from original littermates of ltβ −/− mice) are those described previously (28). A breeding pair of ltα −/− mice (24) was obtained from Nancy Ruddle (Yale University Department of Epidemiology and Public Health, Yale University), derived originally from David Chaplin (Washington University, St. Louis, MO). Mice deficient in both TNFR1 and TNFR2 (dtnfr −/−) represent mice derived by interbreeding tnfr1 −/− mice with tnfr2 −/− mice (29). Various other knockout combinations were obtained by interbreeding. All mice were on a mixed background of C57BL/6 and 129/Sv. Breeding pairs of C57BL/6 Ly5.1 (CD45.1) mice were purchased from Clarence Reeder (Frederick Cancer Research Institute, Frederick, MD). All mice were maintained at Yale University in specific pathogen-free conditions. All procedures were conducted in accordance with Yale animal care and use guidelines.
LTβ genotyping was by PCR using three oligonucleotides, yielding ∼120- and 330-bp products for the ltβ + and ltβ − alleles, respectively. The oligonucleotide sequences are: LTβfor, 5′-GAGACAGTCACACCTGTTG-3′; LTβrev, 5′-CCTGTAGTCCACCATGTCG-3′; and LTβneo, 5′-CTTGTTCAATGGCCGATCC-3′. TNFR1 and TNFR2 genotyping was by Southern blot analysis as described elsewhere (29).
Bone Marrow Chimeras.
Hosts were exposed to 950 rads at 6–8 wk of age and, 1 d later, were given 2 × 106 total nucleated bone marrow cells intravenously in 0.2 ml of PBS. Bone marrow was from sex-matched 8–12-wk-old C57BL/6 Ly5.1 mice. 8 wk after irradiation, the relative ratio of CD45.1+ donor cells versus CD45.2+ host cells in peripheral blood was determined by fluorocytometry. Both biotin-conjugated anti-CD45.1 and FITC-conjugated anti-CD45.2 were from PharMingen (San Diego, CA). The degree of chimerism was >95% in all cases. 9–10 wk after irradiation, recipients were challenged intraperitoneally with 0.1 mg of chicken γ globulin adsorbed to alum in 0.2 ml of PBS and were culled 12 d later.
Pathology.
Visualization of bracheal, axillary, inguinal, and popliteal LNs (30) was aided in some experiments by injecting 50 μl of india ink into each footpad of the mice 3–4 h before culling. The prominence of PPs was greatly increased by immersing the intestine in 10% (vol/vol) acetic acid for 5 min before preservation in 10% neutral-buffered formalin. Hematoxylin and eosin staining was done on paraffin sections using standard procedures.
Immunohistology.
Mice were challenged intraperitoneally at 6–8 wk of age with 0.1 mg of chicken γ globulin adsorbed to alum in 0.2 ml of PBS. Spleens and MLNs were harvested 12 d later and frozen in O.C.T. compound using a dry-ice/methylbutane bath. 5-μm thick sections were cut onto silanized glass slides and fixed in cold acetone for 5 min before storage at −70°C. For staining, sections were allowed to thaw for 10 min and then rehydrated in PBS for 20 min. Endogenous peroxidase was inactivated with 0.3% hydrogen peroxide for 5 min and the sections were then washed with PBS for 10 min. Blocking was with PBS/3% BSA/0.1% (vol/vol) Tween 20 for 30 min. Staining for IgD used rat anti–mouse IgD (Southern Biotechnology Associates, Birmingham, AL), followed by biotin-conjugated goat anti–rat IgG (Southern Biotechnology Associates) and then β-galactosidase– conjugated avidin (Vector Laboratories, Burlingame, CA). Washing between layers was with PBS/0.1% (vol/vol) Tween 20 before reblocking as above. Germinal centers were stained using horseradish peroxidase–conjugated peanut agglutinin (EY Laboratories, San Mateo, CA; reference 31). IgM detection was with alkaline phosphatase–conjugated goat anti–mouse IgM (Southern Biotechnology Associates). Follicular dendritic cells were revealed with biotin-conjugated anticomplement receptor 1 (PharMingen, San Diego, CA; reference 32), followed by alkaline phosphatase– conjugated streptavidin (Zymed, South San Francisco, CA). Substrates for β-galactosidase, horseradish peroxidase, and alkaline phosphatase were HistoMark X-Gal (Kirkegaard and Perry Labs., Inc., Gaithersburg, MD), diaminobenzidine-brown (Zymed), and HistoMark Red (Kirkegaard and Perry Labs., Inc.), respectively.
Results
Ltα− /− Littermates of ltβ− /− Mice Lack MLNs.
Initial reports of the phenotype of two independently generated ltα −/− mouse strains differed in that one indicated that MLNs were absent (24), whereas the other indicated that lymphoid structures were present in the mesentery of 4 out of 14 mice (25). Most recently, among ∼500 ltα −/− mice examined for MLNs, only 10 had a single MLN (33). It was thus suggested that the frequency of occurrence of MLNs in ltα −/− mice may vary depending on how the mice are housed (33). If true, this would perhaps apply equally to ltβ −/− mice, which we described as consistently having MLNs (28). Furthermore, Alimzhanov et al. independently generated ltβ −/− mice and found that only ∼75% of these mice have MLNs (34). It was therefore also conceivable that there are effects of background genes, although all mice examined were on a mixed background of 129/Sv and C57BL/6. The studies here were begun to examine these issues and determine why ltβ −/− mice have MLNs despite the fact that ltα −/− mice mostly do not.
The ltα and ltβ genes are separated by only ∼6 kbp in the MHC locus (12). Thus, we reasoned that it would be possible to generate ltα −/− mice and ltβ −/− mice as littermates by interbreeding mice which are heterozygous for both ltα and ltβ (ltα +/− ltβ +/− mice). In this way, 137 progeny were generated and genotyped as described in Materials and Methods. Ltα −/−, ltβ −/−, and ltα +/− ltβ +/− mice occurred in a relatively normal Mendelian fashion (n = 31, 40, and 66, respectively). Some of these mice were examined at 6–8 wk of age. Ltα −/− mice did not have MLNs (n = 14), whereas almost all of their ltβ −/− littermates did (n = 25). A single ltβ −/− mouse out of 25 appeared to lack MLNs.
Lymphotoxin Gene Dosage Effect in PP Development.
Unlike ltα −/− and ltβ −/− mice, the above heterozygous ltα +/− ltβ +/− mice had all LNs (n = 30), except that two mice had only one inguinal LN and one mouse had none. Surprisingly, however, ltα +/− ltβ +/− mice showed a complete lack of PPs (n = 30), whereas both ltα +/− mice (n = 13) and ltβ +/− mice (n = 14) have PPs as well as all LNs. Having made this observation, we examined ltα +/− ltβ +/− mice further. At 6–8 wk of age, the gross spleen architecture was normal by hematoxylin and eosin histology (data not shown). Immunohistology for complement receptor 1 in the spleen (done as previously described; reference 28) revealed the presence of follicular dendritic cells (data not shown). Also, splenic germinal centers were formed in discrete B cell follicles after intraperitoneal challenge, except there appeared to be some disorganization among IgM+IgDlo/− marginal zone B cells (Fig. 1 E). Ltα −/− mice (24–28) and ltβ −/− mice (28, 34) have severe defects in all of these aspects of lymphoid organogenesis.
Figure 1.
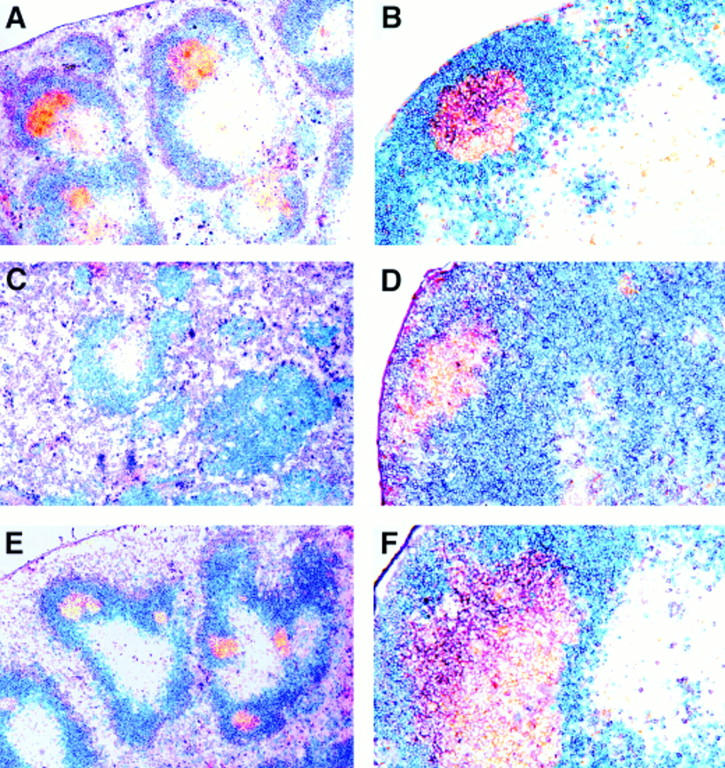
Ltα +/− ltβ +/− mice have relatively normal lymphoid organ architecture. Mice were challenged intraperitoneally with 0.1 mg of chicken γ globulin adsorbed to alum and culled 12 d later. Spleen (A, C, and E) and MLNs (B, D, and F) sections were stained for IgM (red), IgD (blue) and peanut agglutinin–binding germinal centers (brown). A and B, wild-type; C and D, ltβ −/−; E and F, ltα +/− ltβ +/−. Original magnification, ×65 and ×150 for spleen and MLN, respectively.
The organization of the MLNs of ltα +/− ltβ +/− mice was also relatively normal (Fig. 1 F). As previously noted (28), the organization of the MLNs of ltβ −/− mice is not normal in that there appears to be a generalized B cell infiltration, but B cell follicles are found around the rim of MLNs and germinal center B cell clusters are formed despite the absence of follicular dendritic cells (reference 28; Fig. 1 D).
The lack of PPs in ltα +/− ltβ +/− mice was confirmed in progeny from intercrossing ltα −/− mice with ltβ −/− mice (n = 4). Bone marrow chimeras were also generated using wild-type bone marrow, to examine whether or not the lack of PPs was reversible. None of the ltα +/− ltβ +/− recipients showed any sign of PPs 10–12 wk after irradiation, but they did have LNs (n = 9). None of the ltα −/− recipients had MLNs (n = 8), but all of the ltβ −/− recipients did (n = 11). None of the ltα −/− recipients or ltβ −/− recipients had PPs. Ltβ +/+ wild-type recipients had MLNs and PPs (n = 4).
TNFR1 Is Involved in MLN Development.
Both ltβ −/− mice and dtnfr −/− mice have MLNs, and yet ltα −/− mice do not. This led us to propose that LTα may act without LTβ (i.e., as LTα3) via an as yet unidentified receptor (28). To test this hypothesis, we generated mice lacking both LTβ and TNFR and examined them for the presence of MLNs. Since TNFR-deficient mice were originally obtained as dtnfr −/− mice, the first mice generated here were ltβ −/− dtnfr −/− mice. At 6–8 wk of age, ltβ −/− dtnfr −/− mice showed a complete lack of MLNs (n = 10), whereas ltβ +/− dtnfr −/− mice still had MLNs (n = 5).
In a similar way to ltα −/− mice, it is conceivable that the apparent absence of MLNs in ltβ −/− dtnfr −/− mice is due to a possible lack of immune competence and/or lymphocyte homing, and that this might be reversed after reconstitution with wild-type bone marrow. We therefore generated wild-type bone marrow chimeras. However, none of the bone marrow chimeras had MLNs 10–12 wk after reconstitution (n = 11).
In the meantime, we also generated ltβ −/− tnfr1 −/− and ltβ −/− tnfr2 −/− mice. The latter had MLNs (n = 4) but ltβ −/− tnfr1 −/− mice clearly did not (n = 5). Most ltβ −/− tnfr1 +/− littermates (n = 5) had one small MLN (Fig. 2). One ltβ −/− tnfr1 +/− littermate did not appear to have MLNs, whereas another had two small MLNs. This may be explained by the fact that tnfr1 heterozygosity is known to result in a partial phenotype at least in some respects (35), but at the same time ltβ +/− tnfr1 +/− mice had MLNs of a normal size (n = 13).
Figure 2.
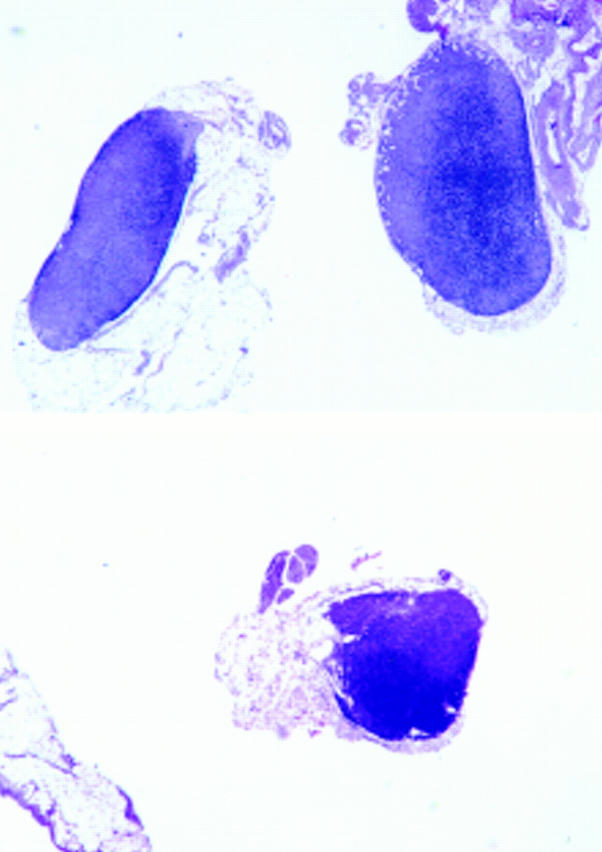
Ltβ −/− tnfr1 +/− mice have defective MLN development. ltβ −/− tnfr −/− mice completely lack MLNs and ltβ −/− tnfr1 +/+ littermates have MLN of apparently normal size and number (top), but ltβ −/− tnfr1 +/− littermates most often have only a single, small MLN (bottom). Hematoxylin and eosin histology; original magnification: ×15.
Discussion
The study reported here extends our knowledge of the roles of TNF ligand/receptor family members in lymphoid organogenesis (Table 1). Based on several observations, we had previously hypothesized that both TNFR1 and LTβR may be involved in PP development (28). First, both ltα −/− mice (24, 25) and ltβ −/− mice (28, 34) completely lack PPs. Second, Rennert et al. observed a complete lack of PPs in mice administered recombinant soluble LTβR in utero (36). Third, tnfr1 −/− mice lack PPs but have reduced numbers of residual lymphoid aggregates (37). Defective PP development was also reported recently with an independently generated tnfr1 −/− mouse strain (29). Others reported that tnfr1 −/− mice have PPs but that they appear flattened due to a lack of B cell follicle structures (38). However, even this study noted that tnfr1 −/− mice have on average only two to four such PPs compared with six to eight PPs in the wild-type control mice (38).
Table 1.
. Phenotypes of Mice Made Genetically Deficient in TNF Ligand/Receptor Family Members
| Family Members | ltα −/− | ltβ −/− | tnfα −/− | tnfr1 −/− | ltα +/− ltβ +/− | ltβ −/− tnfr1 −/− | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Spleen | ||||||||||||
| Primary B cell follicles | − | − | − | − | + | ND | ||||||
| Marginal zone | − | − | + | + | +/− | ND | ||||||
| Germinal centers | − | − | − | − | + | ND | ||||||
| Follicular dendritic cells | − | − | − | − | + | ND | ||||||
| MLNs | − | + | + | + | + | − | ||||||
| Primary B cell follicles | +/− | ND | − | + | ||||||||
| Germinal centers | +/− | ND | − | + | ||||||||
| Follicular dendritic cells | − | ND | − | + | ||||||||
| Cervical LN | − | +/−− | + | + | + | − | ||||||
| Inguinal LN | − | − | + | + | +/− | − | ||||||
| Other LN | − | − | + | + | + | − | ||||||
| PPs | − | − | +/− | +/− | − | − |
This table is based on citations in the text and others as summarized elsewhere (8), as well as the findings from this study. Unlike ltα +/− ltβ +/− double heterozygous mice, ltα +/− mice and ltβ +/− mice have PPs as well as all LNs. Other features of the phenotype of ltα +/− mice and ltβ +/− mice have not been determined. Also, although ltβ −/− tnfr1 −/− mice do not have MLNs, ltβ −/− tnfr2 −/− mice do.
In this study, we show the existence of a gene dosage effect that is consistent with a role for both TNFR1 and LTβR in PP development. That is, ltα +/− ltβ +/− mice specifically lack only PPs, but ltα +/− mice and ltβ +/− mice do not. If LTα and LTβ form a single species that signals via a single receptor, it might be expected that either LTα or LTβ would be the limiting factor and that heterozygosity in either ltα or ltβ alone should result in the lack of PPs seen in ltα +/− ltβ +/− mice. However, this is not the case. Only when both ltα and ltβ are heterozygous does insufficiency become evident. One interpretation would be that two ligands (e.g., LTα3 and LTα1β2 signaling via TNFR1 and LTβR, respectively) are involved in PP development, and that heterozygosity in either one or the other alone is not enough to cause a complete loss of PP development. This two-receptor model might therefore provide an explanation for the partial defect in PP development seen in tnfr1 −/− mice.
Clearly, our results show that both TNFR1 and LTβ are involved in MLN development, even though both tnfr1 −/− mice and ltβ −/− mice have MLNs. TNFR1 also functions independently of TNFR2 in this regard, as ltβ −/− tnfr2 −/− mice still have MLNs. We have thus revealed a previously unappreciated relationship between TNFR1 and LTβ (presumably acting via LTβR). An explanation for the lack of MLN in ltα −/− mice might therefore be that LTα deficiency actually eliminates both ligands of the relationship (i.e., LTα3 and LTα1β2 signaling via TNFR1 and LTβR, respectively). LTα3 itself is not believed to bind LTβR (16, 17).
However, having said this, it has been indicated that ltβr −/− mice lack MLNs (34). Thus, the relationship between TNFR1 and LTβR may be one of synergism with LTβR as the dominant partner. At the same time, the presence of MLN in ltβ −/− mice would imply that LTβR has a ligand besides the LTα/β complex. Indeed, Mauri et al. have very recently described a new LTβR ligand (LIGHT) as well as a new LTα3 receptor, the herpesvirus entry mediator, expressed by lymphocytes (39).
The molecular basis for the relationship between TNFR1 and LTβ (presumably via LTβR) remains to be determined. It is conceivable that TNFR1 and LTβR signaling in MLN development is simultaneous and that they interact at the level of intracellular signal transducers. Certainly, activation of LTβR has been shown to potentiate TNF-α cytotoxicity, possibly reflecting cross-talk between signaling pathways (17, 22). Ligation of LTβR causes recruitment of TNFR-associated factor family members (40–42), and activation of NF-κB and cell death by distinct signaling pathways (42, 43).
Thus far, our studies of ltβ −/− mice have evaluated the defects in lymphoid organogenesis (reference 28 and this study). We are now beginning to examine whether or not LTβ has roles in vivo in other respects. Certainly, in vitro studies have shown that signaling via LTβR causes cytotoxicity to some cell lines (17, 22), chemokine expression (23), and integrin upregulation (21). It remains to be seen whether or not the relationship between TNFR1 and LTβ (presumably via LTβR) in gut-associated lymphoid tissue development extends to any other facets of biology. With this in mind, caution is advised when interpreting the in vivo role (or rather, apparent lack thereof ) of LTβ and TNFR1 based on studies of ltβ −/− mice and tnfr1 −/− mice alone.
Finally, ltα +/− ltβ +/− mice may prove to be a useful PP-less mouse model, not only for the study of gastrointestinal infection, but also of oral tolerance, oral vaccination, and chronic disorders such as inflammatory bowel disease (44– 46). Ltα +/− ltβ +/− mice are being further characterized, particularly with respect to the subtle defect observed in splenic marginal zone organization. Although it remains possible that ltα +/− ltβ +/− mice have other as yet unidentified defects, unlike any other previously described mouse, these mice specifically and completely lack only PPs and do not appear to have any of the major abnormalities associated with ltα −/− and ltβ −/− mice.
Acknowledgments
We thank Jacques Peschon (Immunex Corp., Seattle, WA) for tnfr −/− mice; Frank Wilson, Cindy Hughes, and Debbie Butkus for technical assistance; and Fran Manzo for secretarial assistance.
This work was supported by the Howard Hughes Medical Institute (R.A. Flavell) with the aid of grants from the Human Frontiers Science Program (to P.A. Koni) and the American Diabetes Association (to R.A. Flavell). Richard A. Flavell is an Investigator of the Howard Hughes Medical Institute.
Abbreviations used in this paper: dtnfr
- −/−
mice deficient in both TNFR1 and TNFR2
- LT
lymphotoxin
- MLN
mesenteric lymph nodes
- PPs
Peyer's patches
References
- 1.Karrer U, Althage A, Odermatt B, Roberts CWM, Korsmeyer SJ, Miyamaki S, Hengartner H, Zinkernagel RM. On the key role of secondary lymphoid organs in antiviral immune responses studied in alymphoplastic (aly/ aly) and spleenless (Hox11 −/−) mutant mice. J Exp Med. 1997;185:2157–2170. doi: 10.1084/jem.185.12.2157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Suen WE, Bergman CM, Hjelmström P, Ruddle NH. A critical role for lymphotoxin in experimental allergic encephalomyelitis. J Exp Med. 1997;186:1233–1240. doi: 10.1084/jem.186.8.1233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Roberts CW, Shutter JR, Korsmeyer SJ. Hox11controls the genesis of the spleen. Nature. 1994;368:747–749. doi: 10.1038/368747a0. [DOI] [PubMed] [Google Scholar]
- 4.Miyawaki S, Nakamura Y, Suzuka H, Koba M, Yasumizu R, Ikehara S, Shibata Y. A new mutation, aly, that induces a generalized lack of lymph nodes accompanied by immunodeficiency in mice. Eur J Immunol. 1994;24:429–434. doi: 10.1002/eji.1830240224. [DOI] [PubMed] [Google Scholar]
- 5.Shinkura R, Matsuda F, Sakiyama T, Tsubata T, Hiai H, Paumen M, Miyawaki S, Honjo T. Defects of somatic hypermutation and class switching in alymphoplasia (aly)mutant mice. Int Immunol. 1996;8:1067–1075. doi: 10.1093/intimm/8.7.1067. [DOI] [PubMed] [Google Scholar]
- 6.Förster R, Mattis AE, Kremmer E, Wolf E, Brem G, Lipp M. A putative chemokine receptor, BLR1, directs B cell migration to defined lymphoid organs and specific anatomic compartments of the spleen. Cell. 1996;87:1037–1047. doi: 10.1016/s0092-8674(00)81798-5. [DOI] [PubMed] [Google Scholar]
- 7.Liu Y-J, Banchereau J. Mutant mice without B lymphocyte follicles. J Exp Med. 1996;184:1207–1211. doi: 10.1084/jem.184.4.1207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.von Boehmer H. Lymphotoxins: from cytotoxicity to lymphoid organogenesis. Proc Natl Acad Sci USA. 1997;94:8926–8927. doi: 10.1073/pnas.94.17.8926. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Sacca R, Cuff CA, Ruddle NH. Mediators of inflammation. Curr Opin Immunol. 1997;9:851–857. doi: 10.1016/s0952-7915(97)80189-6. [DOI] [PubMed] [Google Scholar]
- 10.Kratz A, Campos-Neto A, Hanson MS, Ruddle NH. Chronic inflammation caused by lymphotoxin is lymphoid neogenesis. J Exp Med. 1996;183:1461–1472. doi: 10.1084/jem.183.4.1461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Smith CA, Farrah T, Goodwin RG. The TNF receptor superfamily of cellular and viral proteins: activation, costimulation, and death. Cell. 1994;76:959–962. doi: 10.1016/0092-8674(94)90372-7. [DOI] [PubMed] [Google Scholar]
- 12.Ware CF, VanArsdale TL, Crowe PD, Browning JL. The ligands and receptors of the lymphotoxin system. Curr Top Microbiol Immunol. 1995;198:175–218. doi: 10.1007/978-3-642-79414-8_11. [DOI] [PubMed] [Google Scholar]
- 13.Androlewicz MJ, Browning JL, Ware CF. Lymphotoxin is expressed as a heteromeric complex with a distinct 33-kDa glycoprotein on the surface of an activated human T cell hybridoma. J Biol Chem. 1992;267:2542–2547. [PubMed] [Google Scholar]
- 14.Browning JL, Ngam-ek A, Lawton P, DeMarinis J, Tizard R, Chow EP, Hession C, O'Brine-Greco GB, Foley SF, Ware CF. Lymphotoxin β, a novel member of the TNF family that forms a heteromeric complex with lymphotoxin on the cell surface. Cell. 1993;72:847–856. doi: 10.1016/0092-8674(93)90574-a. [DOI] [PubMed] [Google Scholar]
- 15.Browning JL, Dougas I, Ngam-ek A, Bourdon PR, Ehrenfels BN, Miatkowski K, Zafari M, Yampaglia AM, Lawton P, Meier W, et al. Characterization of surface lymphotoxin forms: use of specific monoclonal antibodies and soluble receptors. J Immunol. 1995;154:33–46. [PubMed] [Google Scholar]
- 16.Crowe PD, VanArsdale TL, Walter BN, Ware CF, Hession C, Ehrenfels B, Browning JL, Din WS, Goodwin RG, Smith CA. A lymphotoxin-β–specific receptor. Science. 1994;264:707–710. [PubMed] [Google Scholar]
- 17.Mackay F, Bourdon PR, Griffiths DA, Lawton P, Zafari M, Sizing ID, Miatkowski K, Ngam-ek A, Benjamin CD, Hession C, et al. Cytotoxic activities of recombinant soluble murine lymphotoxin-α and lymphotoxin-αβ complexes. J Immunol. 1997;159:3299–3310. [PubMed] [Google Scholar]
- 18.Browning JL, Sizing ID, Lawton P, Bourdon PR, Rennert PD, Majeau GR, Ambrose CM, Hession C, Miatkowski K, Griffiths DA, et al. Characterization of lymphotoxin-αβ complexes on the surface of mouse lymphocytes. J Immunol. 1997;159:3288–3298. [PubMed] [Google Scholar]
- 19.Schoenfeld HJ, Poeschl B, Frey JR, Loetscher H, Hunziker W, Lustig A, Zulauf M. Efficient purification of recombinant human tumor necrosis factor β from Escherichia coliyields biologically active protein with a trimeric structure that binds to both tumor necrosis factor receptors. J Biol Chem. 1991;266:3863–3869. [PubMed] [Google Scholar]
- 20.Picarella D, Kratz A, Li C-B, Ruddle NH, Flavell RA. Insulitis in transgenic mice expressing TNF-β (lymphotoxin) in the pancreas. Proc Natl Acad Sci USA. 1992;89:10036–10040. doi: 10.1073/pnas.89.21.10036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Hochman PS, Majeau GR, Mackay F, Browning JL. Proinflammatory responses are efficiently induced by homotrimeric but not heterotrimeric lymphotoxin ligands. J Inflamm. 1996;46:220–234. [PubMed] [Google Scholar]
- 22.Browning JL, Miatkowski K, Sizing I, Griffiths D, Zafari M, Benjamin CD, Meier W, Mackay F. Signaling through the lymphotoxin β receptor induces the death of some adenocarcinoma tumor lines. J Exp Med. 1996;183:867–878. doi: 10.1084/jem.183.3.867. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Degli-Esposti MA, Davis-Smith T, Din WS, Smolak PJ, Goodwin RG, Smith CA. Activation of the lymphotoxin β receptor by cross-linking induces chemokine production and growth arrest in A375 melanoma cells. J Immunol. 1997;158:1756–1762. [PubMed] [Google Scholar]
- 24.De Togni P, Goellner J, Ruddle NH, Streeter PR, Fick A, Mariathasan S, Smith SC, Carlson R, Shornick LP, Strauss-Schoenberger J, et al. Abnormal development of peripheral lymphoid organs in mice deficient in lymphotoxin. Science. 1994;264:703–707. doi: 10.1126/science.8171322. [DOI] [PubMed] [Google Scholar]
- 25.Banks TA, Rouse BT, Kerley MK, Blair PJ, Godfrey VL, Kuklin NA, Bouley DM, Thomas J, Kanangat S, Mucenski ML. Lymphotoxin-α–deficient mice: effects on secondary lymphoid organ development and humoral immune responsiveness. J Immunol. 1995;155:1685–1693. [PubMed] [Google Scholar]
- 26.Matsumoto M, Mariathasan S, Nahm MH, Baranyay F, Peschon JJ, Chaplin DD. Role of lymphotoxin and the type I TNF receptor in the formation of germinal centers. Science. 1996;271:1289–1291. doi: 10.1126/science.271.5253.1289. [DOI] [PubMed] [Google Scholar]
- 27.Matsumoto M, Lo SF, Carruthers CJL, Min J, Mariathasan S, Huang G, Plas DR, Martin SM, Geha RS, Nahm MH, Chaplin DD. Affinity maturation without germinal centres in lymphotoxin-α–deficient mice. Nature. 1996;382:462–466. doi: 10.1038/382462a0. [DOI] [PubMed] [Google Scholar]
- 28.Koni PA, Sacca R, Lawton P, Browning JL, Ruddle NH, Flavell RA. Distinct roles in lymphoid organogenesis for lymphotoxins alpha α and β revealed in lymphotoxin β–deficient mice. Immunity. 1997;6:491–500. doi: 10.1016/s1074-7613(00)80292-7. [DOI] [PubMed] [Google Scholar]
- 29.Peschon JJ, Torrance DS, Stocking KL, Glaccum MB, Otten C, Willis CR, Charrier K, Morrissey PJ, Ware CB, Mohler KM. TNF receptor–deficient mice reveal divergent roles for p55 and p75 in several models of inflammation. J Immunol. 1998;160:943–952. [PubMed] [Google Scholar]
- 30.Hebel, R., and M.W. Stromberg. 1976. Anatomy of the Laboratory Rat. Williams & Wilkins Co., Baltimore. 112–118.
- 31.Rose ML, Birbeck MSC, Wallis VJ, Forrester JA, Davies AJS. Peanut lectin binding properties of germinal centres of mouse lymphoid tissue. Nature. 1980;284:364–366. doi: 10.1038/284364a0. [DOI] [PubMed] [Google Scholar]
- 32.Kinoshita T, Takeda J, Hong K, Kozono H, Sakai H, Inoue K. Monoclonal antibodies to mouse complement receptor type 1 (CR1). Their use in a distribution study showing that mouse erythrocytes and platelets are CR1-negative. J Immunol. 1988;140:3066–3072. [PubMed] [Google Scholar]
- 33.Fu Y-X, Huang G, Matsumoto M, Molina H, Chaplin DD. Independent signals regulate development of primary and secondary follicle structure in spleen and mesenteric lymph node. Proc Natl Acad Sci USA. 1997;94:5739–5743. doi: 10.1073/pnas.94.11.5739. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Alimzhanov MB, Kuprash DV, Kosco-Vilbois MH, Luz A, Turetskaya RL, Tarakhovsky A, Rajewski K, Nedospasov SA, Pfeffer K. Abnormal development of secondary lymphoid tissues in lymphotoxin β–deficient mice. Proc Natl Acad Sci USA. 1997;94:9302–9307. doi: 10.1073/pnas.94.17.9302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Sacca R, Cuff CA, Lesslauer W, Ruddle NH. Differential activities of secreted lymphotoxin-α3 and membrane lymphotoxin-α1β2in lymphotoxin-induced inflammation: critical role of TNF receptor 1 signaling. J Immunol. 1998;160:485–491. [PubMed] [Google Scholar]
- 36.Rennert PD, Browning JL, Mebius R, MacKay F, Hochman PS. Surface lymphotoxin α/β complex is required for the development of peripheral lymphoid organs. J Exp Med. 1996;184:1999–2006. doi: 10.1084/jem.184.5.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Neumann B, Luz A, Pfeffer K, Holzmann B. Defective Peyer's patch organogenesis in mice lacking the 55-kD receptor for tumor necrosis factor. J Exp Med. 1996;184:259–264. doi: 10.1084/jem.184.1.259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Pasparakis M, Alexopoulou L, Grell M, Pfizenmaier K, Bluethmann H, Kollias G. Peyer's patch organogenesis is intact yet formation of B lymphocyte follicles is defective in peripheral lymphoid organs of mice deficient for tumor necrosis factor and its 55-kDa receptor. Proc Natl Acad Sci USA. 1997;94:6319–6323. doi: 10.1073/pnas.94.12.6319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Mauri DN, Ebner R, Montgomery RI, Kochel KD, Cheung TC, Yu G-L, Ruben S, Murphy M, Eisenberg RJ, Cohen GH, et al. LIGHT, a new member of the TNF superfamily, and lymphotoxin α are ligands for herpesvirus entry mediator. Immunity. 1998;8:21–30. doi: 10.1016/s1074-7613(00)80455-0. [DOI] [PubMed] [Google Scholar]
- 40.Nakano H, Oshima H, Chung W, Williams-Abbott L, Ware CF, Yagita H, Okumura K. TRAF5, an activator of NF-κB and putative signal transducer for the lymphotoxin-β receptor. J Biol Chem. 1996;271:14661–14664. doi: 10.1074/jbc.271.25.14661. [DOI] [PubMed] [Google Scholar]
- 41.Force W R, Cheung TC, Ware CF. Dominant negative mutants of TRAF3 reveal an important role for the coiled coil domains in cell death signaling by the lymphotoxin-β receptor. J Biol Chem. 1997;272:30835–30840. doi: 10.1074/jbc.272.49.30835. [DOI] [PubMed] [Google Scholar]
- 42.VanArsdale TL, VanArsdale SL, Force WR, Walter BN, Mosialos G, Kieff E, Reed JC, Ware CF. Lymphotoxin-β receptor signaling complex: role of tumor necrosis factor receptor–associated factor 3 recruitment in cell death and activation of nuclear factor κB. Proc Natl Acad Sci USA. 1997;94:2460–2465. doi: 10.1073/pnas.94.6.2460. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Mackay F, Majeau GR, Hochman PS, Browning JL. Lymphotoxin β receptor triggering induces activation of the nuclear factor κB transcription factor in some cell types. J Biol Chem. 1996;271:24934–24938. doi: 10.1074/jbc.271.40.24934. [DOI] [PubMed] [Google Scholar]
- 44.Neutra MR, Pringault E, Kraehenbuhl J-P. Antigen sampling across epithelial barriers and induction of mucosal immune responses. Annu Rev Immunol. 1996;14:275–300. doi: 10.1146/annurev.immunol.14.1.275. [DOI] [PubMed] [Google Scholar]
- 45.Mowat AM, Viney JL. The anatomical basis of intestinal immunity. Immunol Rev. 1997;156:145–166. doi: 10.1111/j.1600-065x.1997.tb00966.x. [DOI] [PubMed] [Google Scholar]
- 46.Mayrhofer G. Peyer's patch organogenesis—cytokines rule, OK? . Gut. 1997;41:707–709. doi: 10.1136/gut.41.5.707. [DOI] [PMC free article] [PubMed] [Google Scholar]