

Abstract
The recent identification of cannabinoid receptors and their endogenous lipid ligands has triggered an exponential growth of studies exploring the endocannabinoid system and its regulatory functions in health and disease. Such studies have been greatly facilitated by the introduction of selective cannabinoid receptor antagonists and inhibitors of endocannabinoid metabolism and transport, as well as mice deficient in cannabinoid receptors or the endocannabinoid-degrading enzyme fatty acid amidohydrolase. In the past decade, the endocannabinoid system has been implicated in a growing number of physiological functions, both in the central and peripheral nervous systems and in peripheral organs. More importantly, modulating the activity of the endocannabinoid system turned out to hold therapeutic promise in a wide range of disparate diseases and pathological conditions, ranging from mood and anxiety disorders, movement disorders such as Parkinson’s and Huntington’s disease, neuropathic pain, multiple sclerosis and spinal cord injury, to cancer, atherosclerosis, myocardial infarction, stroke, hypertension, glaucoma, obesity/metabolic syndrome, and osteoporosis, to name just a few. An impediment to the development of cannabinoid medications has been the socially unacceptable psychoactive properties of plant-derived or synthetic agonists, mediated by CB1 receptors. However, this problem does not arise when the therapeutic aim is achieved by treatment with a CB1 receptor antagonist, such as in obesity, and may also be absent when the action of endocannabinoids is enhanced indirectly through blocking their metabolism or transport. The use of selective CB2 receptor agonists, which lack psychoactive properties, could represent another promising avenue for certain conditions. The abuse potential of plant-derived cannabinoids may also be limited through the use of preparations with controlled composition and the careful selection of dose and route of administration. The growing number of preclinical studies and clinical trials with compounds that modulate the endocannabinoid system will probably result in novel therapeutic approaches in a number of diseases for which current treatments do not fully address the patients’ need. Here, we provide a comprehensive overview on the current state of knowledge of the endocannabinoid system as a target of pharmacotherapy.
I. Introduction
Marijuana, or cannabis, is the most widely used illicit drug in Western societies and also the one with the longest recorded history of human use. The popularity of marijuana as a recreational drug is due to its ability to alter sensory perception and cause elation and euphoria, most vividly described by the 19th century French poet, Charles Baudelaire, in his book Les Paradis Artificiels (Iversen, 2000). However, the ability of extracts of the hemp plant (Cannabis sativa) to cause a variety of medicinal effects unrelated to its psychoactive properties had been recognized as early as the third millennium BC, when Chinese texts described its usefulness in the relief of pain and cramps (Mechoulam, 1986). In ancient India, the anxiety-relieving effect of bhang (the Indian term for marijuana ingested as food) had been recorded more than 3000 years ago. The use of cannabis or hashish as a psychoactive substance reached Europe and the Americas through the Arab world in the 19th century. During the same period, cannabis extracts had gained widespread use for medicinal purposes until 1937, when concern about the dangers of abuse led to the banning of marijuana for further medicinal use in the United States. The rather turbulent history of marijuana and the recent resurgence of interest in its medicinal properties have been the subject of excellent reviews (Mechoulam, 1986; Iversen, 2000; Di Marzo et al., 2004; Howlett et al., 2004; Pertwee, 2005a; Piomelli, 2005; Di Marzo and Petrocellis, 2006; Mackie, 2006; Pagotto et al., 2006). Added to this interest is the emergence of the endocannabinoid system, offering not only new insights into the mechanisms underlying the therapeutic actions of plant-derived phytocannabinoids but also novel molecular targets for pharmacotherapy. In this overview, we will briefly summarize current thoughts about the role of endocannabinoids in a given physiological or pathological process and then survey attempts to exploit this role for therapeutic gain.
II. The Pharmacology of Cannabinoids
A. Cannabinoid Receptors and Ligands
Up until the last two decades, marijuana research was a rather esoteric field, of interest to a small number of scientists. A contributory factor was the highly lipophilic nature of the biologically active ingredients, which led to the notion that marijuana elicits its effects nonspecifically by perturbing membrane lipids (Lawrence and Gill, 1975). The first important breakthrough that ultimately led to a rejection of this concept was the identification by Gaoni and Mechoulam (1964) of the correct chemical structure of the main psychoactive ingredient of marijuana, Δ9-tetrahydrocannabinol (THC1), and the subsequent demonstration that bioactivity resides in the l-stereoisomer of this compound (Mechoulam and Gaoni, 1967), which is one of approximately 60 cannabinoids present in the plant (Dewey, 1986). This discovery stimulated the generation of a whole range of synthetic analogs in the 1970s that included not only compounds structurally similar to phytocannabinoids (Fig. 1A) but also analogs with different chemical structures, including classic and nonclassic cannabinoids and aminoalkylindoles (Fig. 1B) (Howlett et al., 2002), as well as the subsequently discovered endogenous arachidonic acid derivatives or endocannabinoids (Fig. 1C), which are discussed in more detail below. Studies of the biological effects of THC and its synthetic analogs revealed strict structural selectivity (Hollister, 1974) as well as stereo-selectivity (Jones et al., 1974), telltale signs of drug-receptor interactions. Definitive evidence for the existence of specific cannabinoid receptors was followed soon by the demonstration of high-affinity, saturable, stereospecific binding sites for the synthetic cannabinoid agonist [3H]CP-55,940 in mouse brain plasma membranes, which correlated with both the in vitro inhibition of adenylate cyclase and the in vivo analgesic effect of the compound (Devane et al., 1988). The availability of a radioligand also allowed the mapping of cannabinoid receptors in the brain by receptor autoradiography (Herkenham et al., 1991b). This mapping turned out to be of key importance in the subsequent identification of an orphan G protein-coupled receptor (GPCR) as the brain receptor for cannabinoids (Matsuda et al., 1990), later named CB1 receptor, based on the overlapping regional distribution of the mRNA for this GPCR and [3H]CP-55,940 binding sites. CB1 receptors are the most abundant receptors in the mammalian brain but are also present at much lower concentrations in a variety of peripheral tissues and cells. A second cannabinoid GPCR, CB2, is expressed primarily in cells of the immune and hematopoietic systems (Munro et al., 1993) but recently were found to be present in the brain (Van Sickle et al., 2005; Gong et al., 2006), in nonparenchymal cells of the cirrhotic liver (Julien et al., 2005), in the endocrine pancreas (Juan-Pico et al., 2005), and in bone (Karsak et al., 2004; Idris et al., 2005; Ofek et al., 2006). Two splice variants of CB1 receptors have been also identified: CB1A, which has an altered amino-terminal sequence (Shire et al., 1995), and CB1B, which has an in-frame deletion of 33 amino acids at the amino terminus (Ryberg et al., 2005). The mRNAs of both splice variants are expressed at much lower levels than the CB1 mRNA and, although the receptors expressed from the cDNAs have unique pharmacology (Ryberg et al., 2005), evidence for their natural expression has not been reported.
Fig. 1.
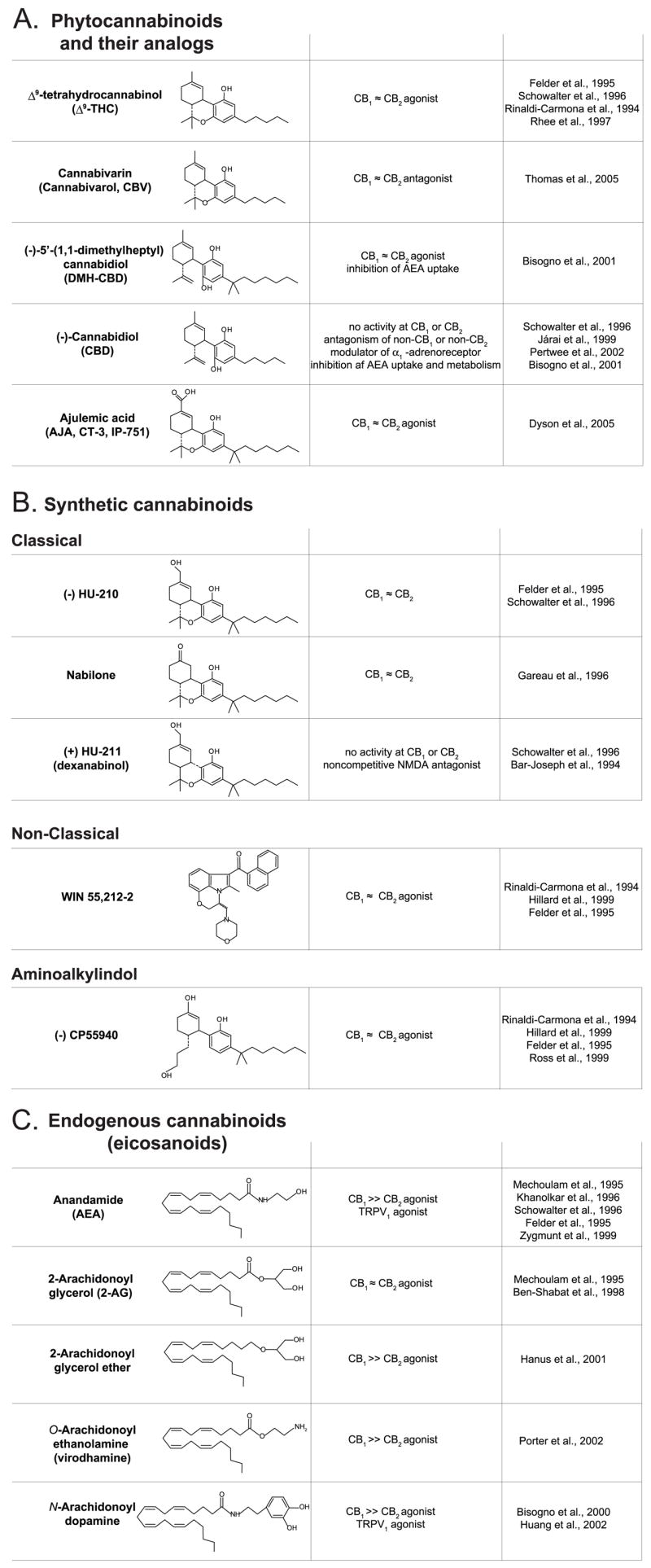
The chemical structure and pharmacological activity of selected plant derived (A), synthetic (B), and endogenous cannabinoids (C).
An interesting twist on the steric selectivity of cannabinoid receptors has emerged through recent studies of the behaviorally inactive phytocannabinoid (−)-cannabidiol (CBD) and its synthetic analogs, which have negligible affinity for either CB1 or CB2 receptors. Paradoxically, some of the synthetic (+)-(+)-stereoisomers of these compounds were found to bind potently to both CB1 and CB2 receptors (Bisogno et al., 2001) but to display only peripheral and not centrally mediated cannabinoid-like bioactivity, suggesting that they may act as antagonists rather than agonists at central, but not peripheral, CB1 receptors (Fride et al., 2005).
Another ligand that displays central versus peripheral selectivity is ajulemic acid, a metabolite of THC that was found to have potent anti-inflammatory and analgesic properties without any overt behavioral or psychoactive effects (Burstein et al., 1992; Dyson et al., 2005; Mitchell et al., 2005). Ajulemic acid was reported to bind to both CB1 and CB2 receptors with reasonably high affinity (Kd 100–200 nM) but only to activate the latter (Rhee et al., 1997), which may explain its unique and therapeutically attractive pharmacological profile. A more recent study indicated even higher affinities for CB1 (Ki 6 nM) and CB2 receptors (Ki 56 nM) and specified the role of CB1 in mediating its antihyperalgesic activity in neuropathic pain (Dyson et al., 2005). This article also documented limited brain penetration of ajulemic acid compared with other cannabinoids, which may account for its favorable therapeutic profile. Ajulemic acid also binds to peroxisome proliferator-activated receptor γ receptors with low (micromolar) affinity, which was proposed to account for its effect on adipocyte differentiation (Liu et al., 2003b).
Among the 60 or so cannabinoids present in marijuana, only THC is psychoactive. However, some of the other constituents, such as cannabidiol, have well-documented biological effects of potential therapeutic interest, such as antianxiety, anticonvulsive, antinausea, anti-inflammatory and antitumor properties (Mechoulam et al., 2002c; Grotenhermen, 2004; Vaccani et al., 2005). Cannabidiol does not significantly interact with CB1 or CB2 receptors, and its actions have been attributed to inhibition of anandamide degradation or its antioxidant properties (Mechoulam and Hanus, 2002; Mechoulam et al., 2002c), or an interaction with as yet unidentified cannabinoid receptors (see below). Another marijuana constituent of potential therapeutic interest is tetrahydrocannabivarin (Markus, 1971), which has recently been shown to have CB1 antagonist properties (Thomas et al., 2005).
In addition to CB1 and CB2 receptors, pharmacological evidence has been accumulating over the years to support the existence of one or more additional receptors for cannabinoids (reviewed in Begg et al., 2005). Two of these possibilities have been more extensively explored: an endothelial site involved in vasodilation and endothelial cell migration (Járai et al., 1999; Begg et al., 2003; Mo et al., 2004), and a presynaptic site on glutamatergic terminals in the hippocampus mediating inhibition of glutamate release (Hájos et al., 2001). Responses elicited at both of these sites were reported to survive genetic ablation of CB1 receptors, yet be sensitive to inhibition by the CB1 antagonist SR141716 or by pertussis toxin but not by the CB1 antagonist AM251 (Járai et al., 1999; Hájos and Freund, 2002; Ho and Hiley, 2003; Offertáler et al., 2003; O’Sullivan et al., 2004a,b). However, the two sites are apparently different. The aminoalkylindol WIN 55,212-2 was found to be an agonist and capsazepine an antagonist at the hippocampal (Hájos and Freund, 2002) but not at the endothelial receptor (Wagner et al., 1999; Mukhopadhyay et al., 2002). On the other hand, certain atypical cannabinoids with no affinity for CB1 or CB2 receptors behave as agonists (abnormal cannabidiol, O-1602) or antagonists at the endothelial receptor (cannabidiol, O-1918) but not at the hippocampal receptor (Begg et al., 2005). Arachidonoyl-L-serine, an endogenous lipid discovered in rat brain, has been found to be a vasodilator acting at the endothelial cannabinoid receptor (Milman et al., 2006), although its activity at the hippocampal receptor has not yet been evaluated. The existence of this latter receptor has recently been called into question, as the ability of WIN 55,212-2 to suppress the same excitatory synapse as studied by Hájos et al. (2001) was found to be absent in two different strains of CB1 knockout mice, yet present in their respective wild-type controls (Takahashi and Castillo, 2006). Atypical cannabinoid receptors with pharmacological properties similar to those of the endothelial receptor have been postulated to exist on microglia, where they mediate microglial migration (Walter et al., 2003), and on neurons of the mouse vas deferens (Pertwee et al., 2002, 2005c). Activation of this latter receptor by the CBD analog 7-OH-dimethylheptyl CBD, which is inactive at CB1, CB2, or transient receptor potential vanilloid type 1 (TRPV1) receptors, inhibits electrically evoked contractions of the vas deferens, and the effect is selectively inhibited by CBD itself. A brain cannabinoid receptor distinct from CB1 was also indicated by the ability of anandamide and WIN 55,212-2, but not other agonists, to stimulate GTPγS binding in brain plasma membranes from CB1 knockout mice (Breivogel et al., 2001).
Of interest are recent findings reported in the patent literature that the orphan receptor GPR-55 (Sawzdargo et al., 1999) recognizes a variety of cannabinoid ligands, but not WIN 55,212-2 (Brown and Wise, 2003; Drmota et al., 2004). However, GPR-55 is apparently not expressed in the vascular endothelium and is sensitive to HU-210 (Drmota et al., 2004), a potent synthetic cannabinoid devoid of vasorelaxant properties (Wagner et al., 1999). Furthermore, it couples to G12/G13 and ρ kinase, which have been linked to vasoconstrictor rather than vasodilator responses. This suggests that GPR-55 is not the abnormal cannabidiol-sensitive endothelial receptor. Mice deficient in GPR-55 will help in defining the biological functions of this novel cannabinoid-sensitive receptor.
Anandamide has been found to be an agonist ligand for the TRPV1 ion channel, although its affinity in the low micromolar range is lower than its affinity for CB1 receptors (reviewed by van der Stelt and Di Marzo, 2004). An in vitro study in rat mesenteric arteries provided evidence that the endothelium-independent component of anandamide-induced vasodilation is mediated via activation of capsaicin-sensitive TRPV1 in sensory nerve terminals. This triggers the release of CGRP, which then dilates the artery by activation of calcitonin gene-related peptide receptors on the vascular smooth muscle (Zygmunt et al., 1999). However, this mechanism does not contribute to the in vivo hypotensive action of anandamide, which is similar in wild-type and TRPV1−/− mice (Pacher et al., 2004).
Both CB1 and CB2 receptors are G protein-coupled receptors. Surprisingly, they share little sequence homology, only 44% at the protein level or 68% in the transmembrane domains, which are thought to contain the binding sites for cannabinoids (Lutz, 2002). Despite this, THC and most synthetic cannabinoids have similar affinities for the two receptors, and only recently did synthetic ligands that discriminate between CB1 and CB2 receptors emerge. These include agonists as well as antagonists, as listed in Fig. 2. The development of potent and highly selective CB1 and CB2 receptor antagonists (Rinaldi-Carmona et al., 1994, 1998) is particularly noteworthy as it provided critically important tools to explore the physiological functions of endocannabinoids. For example, as it will be discussed later in this review, the appetite-reducing effects of the CB1 antagonist SR141716 in various rodent models was the first sign to suggest that endocannabinoids may be tonically active orexigenic agents, representing the endogenous counterpart of the “munchies” caused by marijuana smoking.
Fig. 2.
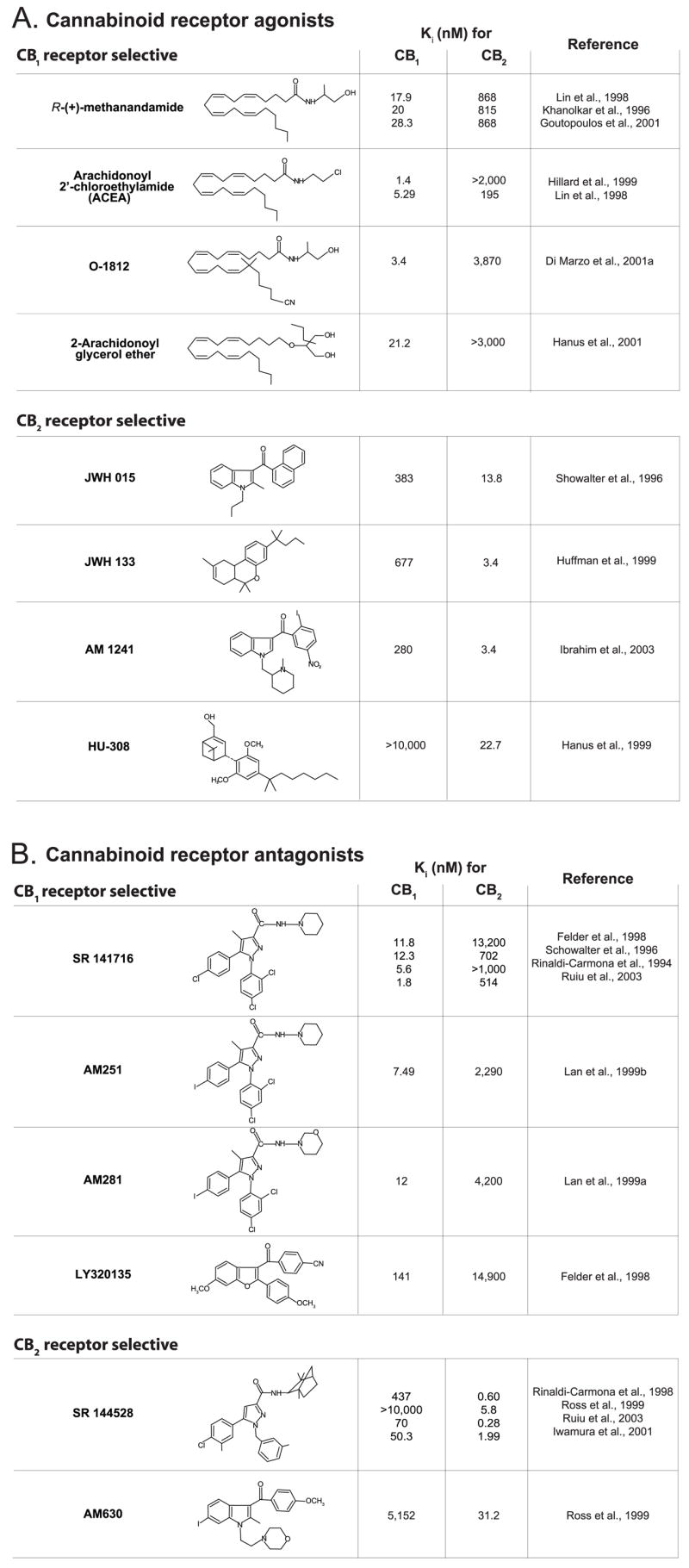
Selective agonists (A) and antagonists (B) of CB1 and CB2 receptors.
However, these antagonists, as well as most of the other CB1 and CB2 antagonists developed to date, have inverse agonist properties (Bouaboula et al., 1997, 1999), so their effects do not necessarily reflect reversal of the tonic action of an endocannabinoid. For this reason, the development of CB1 and CB2 receptor-deficient mouse strains (Ledent et al., 1999; Zimmer et al., 1999; Buckley et al., 2000; Marsicano et al., 2002b; Robbe et al., 2002) was similarly important, as the use of these animals in combination with receptor antagonists can reinforce the putative regulatory roles of endocannabinoids. More recently, the development of conditional mutant mice that lack the expression of CB1 receptors only in certain types of neurons represents another milestone, as it allows linking of specific neuronal populations with a well-defined cannabinoid-modulated behavior (Marsicano et al., 2003).
B. Cannabinoid Receptor Signaling
CB1 and CB2 receptors couple primarily to the Gi/o subtypes of G protein, and their signaling is remarkably complex. Although coupling to adenylate cyclase through Gi/o usually results in inhibition of cyclase activity through the release of Giα isoforms, cannabinoids can also stimulate isoforms 2, 4, or 7 of adenylate cyclase via the release of βγ subunits (Rhee et al., 1998). Activation of adenylate cyclase also occurs when CB1 and dopamine D2 receptors are simultaneously activated (Glass and Felder, 1997), probably as a result of heterodimerization of these two types of receptors (Kearn et al., 2005). Although direct evidence for the coupling of CB1 receptors to Gq/11 had until recently been lacking (Howlett, 2004), the agonist WIN 55,212-2, but not other cannabinoids, was recently reported to increase intracellular calcium in cultured hippocampal neurons and in human embryonic kidney 293 cells via coupling to Gq/11 proteins (Lauckner et al., 2005). Receptor dimerization may facilitate such coupling, which may account for CB1-mediated mobilization of intracellular calcium in NG108-15 neuroblastoma glioma cells (Sugiura et al., 1999). Cannabinoids can also inhibit different types of calcium channels (Mackie and Hille, 1992; Gebremedhin et al., 1999) and activate certain potassium channels (Mackie et al., 1995) via G protein βγ subunits (Ikeda, 1996). Cannabinoids can activate members of all three families of multifunctional mitogen-activated protein kinases, including p44/42 MAP kinase (Wartmann et al., 1995; Davis et al., 2003), p38 kinase (Liu et al., 2000; Derkinderen et al., 2001), and JUN-terminal kinase (Liu et al., 2000; Rueda et al., 2000) and activate the phosphatidylinositol-3-kinase pathway (Gómez Del Pulgar et al., 2002a). These effects could be via G protein activation (Galve-Roperh et al., 2002; Davis et al., 2003) or pathways independent of G proteins via other adaptor proteins (Sánchez et al., 2001b). Another G protein-independent pathway activated by cannabinoids involves G protein-coupled receptor kinase-3 and β-arrestin-2, which are required for desensitization, but not for internalization, of CB1 receptors, and the related development of tolerance (Jin et al., 1999). Cannabinoids can also regulate the activity of phosphatases, as ex-emplified the CB1-mediated regulation of calcineurin (protein phosphatase 2b) (Cannich et al., 2004) or the activation of mitogen-activated protein kinase phosphatase 1, which plays an important role in the anti-inflammatory action of anandamide (Eljaschewitsch et al., 2006).
Different structural classes of cannabinoid receptor agonists have the unique ability to activate different signaling cascades which, in turn, influences agonist efficacy. Using an in situ receptor/G protein reconstitution technique, CB1 receptors were found to efficiently couple and activate both Gi and Go, whereas CB2 receptors only activated Go. Furthermore, the efficacy of a given agonist was different whether CB1 receptors coupled to Gi or Go, demonstrating agonist-selective G protein signaling (Glass and Northup, 1999). Prather et al. (2000) found that the aminoalkylindol agonist WIN 55,212-2 activated different Giα subunits with markedly different potencies. Even more striking is the recent finding that demonstrates cannabinoid agonist-selective activation of different Giα subunits (Mukhopadhyay and Howlett, 2005). A possible practical implication of such findings is that unique therapeutic profiles may be achieved through the use of different agonists for the same receptor, and such profiles may differ from one target tissue to the other, depending on the pattern of G protein subunit expression.
At least part of this agonist selectivity in G protein activation may be related to the existence of distinct binding sites on CB1 receptors for different classes of ligands, as documented by site-directed mutagenesis and molecular modeling studies (see Reggio, 2003). These studies indicate that a K3.28A mutation in the third transmembrane domain caused a more than 1000-fold loss in affinity and loss of efficacy for anandamide and nonclassic cannabinoids, without affecting the affinity for WIN 55,212-2 (Song and Bonner, 1996). In contrast, mutations at different sites in the third, fifth, and sixth transmembrane helices (F3.36A, W5.43A, and W6.48A) affected the binding of WIN 55,212-2 and SR141716, but not anandamide (McAllister et al., 2003).
Another important feature of cannabinoid signaling in the brain is the lack of correlation between the density of CB1 receptors in a given brain region and the efficiency of receptor coupling, as determined by GTPγS binding (Breivogel et al., 1997), which may explain why functionally important responses can be triggered in brain regions with very sparse CB1 receptor expression, such as the brainstem (Rademacher et al., 2003) or the hypothalamus (Jamshidi and Taylor, 2001). Selley et al. (2001) have shown that the reduction in CB1 receptor density in CB1 heterozygote mice was compensated for by an increase in receptor/G protein coupling efficiency for some, but not other, agonists. Although the underlying mechanisms for such compensation are not clear, differences in the degree of receptor multimerization (Mackie, 2005), or changes in signal amplification are possibilities. Recent observations indicate that a considerable proportion of the psychomotor effect of cannabinoids can be accounted for by a signaling cascade in striatal projection neurons involving protein kinase A-dependent phosphorylation of DARPP-32, achieved via modulation of dopamine D2 and adenosine A2A transmission (Andersson et al., 2005). This represents a unique form of amplification of CB1 signaling, as phosphorylation of DARPP-32 at Thr-34 amplifies downstream signaling via inhibition of protein phosphatase-1 (Greengard, 2001). It would be interesting to test whether the efficiency of CB1 coupling to DARPP-32 is affected by cellular receptor density.
C. Endocannabinoids
The existence of specific receptors in mammalian cells that recognize a plant-derived substance rekindled the question raised two decades earlier, after brain receptors for morphine had been first described, i.e., is there an endogenous ligand? A positive answer was provided in 1992 by the report by Devane et al. describing the isolation from porcine brain of the lipid arachidonoyl ethanolamide, named anandamide, which bound to the brain cannabinoid receptor with reasonably high affinity and mimicked the behavioral actions of THC when injected into rodents (Devane et al., 1992). Three years later a second endocannabinoid, 2-arachidonoylglycerol (2-AG), was discovered independently by Mechoulam et al. (1995) and Sugiura et al. (1995). Since then, a number of related endogenous lipids with endocannabinoid-like activity have been reported (Fig. 1c), but follow-up studies about biosynthesis, cellular transport, metabolism, and biological function have focused on anandamide and 2-AG, with much less information available about the other compounds with endocannabinoid-like properties. The biochemical aspects of endocannabinoids have been recently reviewed by Bisogno et al. (2005).
Anandamide is a partial or full agonist of CB1 receptors, depending on the tissue and biological response measured. Although it also binds CB2 receptors, it has very low efficacy and may act as an antagonist (Gonsiorek et al., 2000). The in vivo biosynthesis of anandamide (Fig. 4) is believed to occur through the enzymatic hydrolysis catalyzed by a phospholipase D of a membrane lipid precursor, N-arachidonoyl phosphatidylethanolamide (NAPE) (Schmid et al., 1983), which itself is generated by the enzymatic transfer of arachidonic acid in the sn-1 position in phosphatidylcholine to the amide group of PE (Di Marzo et al., 1994; Cadas et al., 1997). Although a specific transacylase for the latter reaction has not yet been identified, a NAPE-specific PLD has recently been cloned (Okamoto et al., 2004). It is not yet known, however, whether NAPE-PLD is obligatory for the biosynthesis of anandamide, which could make it an attractive target of drug therapy when reduction of tissue anandamide would be of benefit. Indeed, there may be parallel pathways for the generation of anandamide from NAPE. A secretory PLA2 that can catalyze the hydrolysis of N-acyl-PE to N-acyl-lysoPE, which is then acted on by a lysoPLD to generate N-acyl-ethanolamides, including anandamide, was recently identified in the stomach (Sun et al., 2004). An alternative parallel pathway has been identified in our laboratory in RAW246.7 macrophages. This involves hydrolysis of NAPE to phosphoanandamide by a PLC, followed by dephosphorylation through a phosphatase (Liu et al., 2006). This latter pathway rather than PLD is the target of regulation by bacterial endotoxin, which increases anandamide synthesis in macrophages (Varga et al., 1998; Liu et al., 2003a). The existence of this pathway may also account for the recent finding that anandamide tissue levels are unchanged in NAPE-PLD knockout compared with wild-type mice (Leung et al., 2006).
Fig. 4.

Schematic representation of the endocannabinoid system in pre- and postsynaptic neurons. The presynaptic terminal is located in the top, whereas the postsynaptic neuron is located in the bottom. EMT, endocannabinoid membrane transporter; MAGL, monoacylglyceride lipase; DAGL, DAG lipase; AEA, anandamide; NArPE, N-arachidonyl phosphatidylethanolamine; NAT, N-acyltransferase.
2-AG is generated from diacylglycerol (DAG) by DAG lipase selective for the sn-1 position (Fig. 4). DAG, an intracellular second messenger that activates protein kinase C, can be generated from phosphoinositides by a phosphoinositide-specific PLC or from phosphatidic acid by phosphatidic acid phosphohydrolase (Bisogno et al., 2005). Two DAG lipase isozymes, α and β, have been cloned (Bisogno et al., 2003). In the adult brain they are localized in the postsynaptic plasma membrane, in line with their putative role in generating 2-AG involved in retrograde transmission.
Basal levels of 2-AG in the brain are approximately 2 orders of magnitude higher than the levels of anandamide. Despite this, stimulus-induced release resulting in detectable extracellular levels could be demonstrated only for anandamide and not for 2-AG in an in vivo microdialysis study (Giuffrida et al., 1999). This finding illustrates that, despite growing interest in endocannabinoids and their roles as retrograde neurotransmitters (Wilson and Nicoll, 2002; Chevaleyre et al., 2006), the mechanism of their release is not well understood. Like prostanoids, endocannabinoids are not stored but generated on demand in response to a depolarization-induced rise in intracellular calcium or activation of various metabotropic receptors (Varma et al., 2001; Kim et al., 2002; Witting et al., 2004; Di et al., 2005a,b). A putative membrane endocannabinoid transporter involved in the cellular uptake of endocannabinoids (see below) may also be involved in their release. This is suggested by the ability of a transport inhibitor to prevent the release of intracellularly applied anandamide (Maccarrone et al., 2000a; Gerdeman et al., 2002).
Anandamide present in the extracellular space is accumulated by neurons and other cells by facilitated diffusion. This process is driven by its transmembrane concentration gradient, is saturable and temperature-dependent, and does not require ATP or sodium ions. Most importantly for the topic of the present review, anandamide uptake is selectively inhibited by a variety of structural analogs, which suggests the existence of a saturable cellular component involved in anandamide transport (Beltramo et al., 1997; Bisogno et al., 1997; Hillard and Jarrahian, 2000; Maccarrone et al., 2000a). However, a specific anandamide transporter protein has yet to be cloned, and it has been proposed that intracellular degradation of anandamide by fatty acid amide hydrolase (FAAH) is sufficient to account for anandamide uptake in long incubation periods (Glaser et al., 2003). Studies with cells isolated from FAAH+/+ and FAAH−/− mice did not resolve this issue, as the absence of FAAH was found not to affect anandamide uptake (Fegley et al., 2004) or to reduce it substantially (Ortega-Gutierrez et al., 2004), albeit under different experimental conditions. Nevertheless, a FAAH-independent component of anandamide uptake, inhibited by the compound UCM707, was detected in the latter study, supporting the notion of a protein other than FAAH being involved. This notion is also supported by the emergence of a number of synthetic transport inhibitors, the potencies of which to inhibit anandamide uptake does not correlate with their affinities for CB1, CB2, or TRPV1 receptors or their potencies to inhibit FAAH (Fig. 3). However, in view of the important role of FAAH in generating the transmembrane concentration gradient for anandamide, the possibility that a noncatalytic region of FAAH or a FAAH-associated protein may act as anandamide transporter cannot be excluded. Interestingly, the elucidation of the crystal structure of FAAH revealed several channel-like regions in the enzyme, granting it simultaneous access to both the cytosolic and membrane domains (Bracey et al., 2002). Against this possibility, however, is the recent report that the novel, high affinity anandamide transport inhibitor LY2318912 binds with similar Kd and bmax values to membranes from HeLa cells devoid of FAAH or transfected with FAAH, pointing to a binding site independent of the FAAH molecule (Moore et al., 2005). Arguments for and against the existence of a bidirectional anandamide transporter have been recently reviewed (Hillard and Jarrahian, 2003; Fowler et al., 2004; Mc-Farland and Barker, 2004; Glaser et al., 2005).
Fig. 3.

The structure and pharmacological specificity of inhibitors of FAAH and of endocannabinoid membrane transport.
In some in vivo studies, treatment with transport inhibitors unmasked cannabinoid-like tonic effects on pain sensitivity, anxiety-like behaviors, locomotor activity, and muscle spasticity, which is an indication of the potential therapeutic usefulness of such compounds (Moore et al., 2005; Bortolato et al., 2006; La Rana et al., 2006). Similar and more pronounced effects have been reported in response to treatment with FAAH inhibitors, as discussed below.
In contrast to the unsettled status of anandamide transport and a putative transporter protein, the unique role of FAAH in the in vivo degradation of anandamide has been extensively documented (reviewed in McKinney and Cravatt, 2005). Initial evidence for a membrane-associated enzyme in the liver that hydrolyzes N-N-acyl ethanolamides (Schmid et al., 1985) was followed by the cloning of FAAH (Cravatt et al., 1996) and the identification of its crystal structure in complex with an active site-directed inhibitor (Bracey et al., 2002). The unique role of FAAH in terminating signaling by anandamide was indicated by the phenotype of FAAH knockout mice, which displayed 10 to 15 times elevated levels of anandamide across the brain, supersensitivity to the actions of exogenous anandamide, and the appearance of tonic signaling by endogenous anandamide, resulting in CB1 receptor-mediated hypoalgesia (Cravatt et al., 2001; Lichtman et al., 2004b), reduced anxiety (Kathuria et al., 2003), antidepressant activity (Gobbi et al., 2006), and lowering of blood pressure in different models of experimental hypertension (Bátkai et al., 2004b). Cravatt et al. (2004) were able to resolve the relative roles of central versus peripheral fatty acid amides by generating mice deficient in FAAH in peripheral tissues only. These mice did not display the hypoalgesia observed in mice with global deficiency in FAAH, but had a similar anti-inflammatory phenotype, indicating that the latter was mediated by elevated fatty acid amides in peripheral tissues (Cravatt et al., 2004). Interestingly, another amidohydrolase catalyzing the same reaction as FAAH but at acidic pH was recently identified and cloned (Tsuboi et al., 2005). This lysosomal enzyme is structurally unrelated to FAAH and is widely distributed in tissues, with highest levels in the lung, and has been recently shown to contribute to the physiological degradation of anandamide in macrophages but not in the brain (Sun et al., 2005).
Although 2-AG is also hydrolyzed by FAAH under in vitro conditions (Goparaju et al., 1998; Lang et al., 1999), in vivo it is not a substrate of FAAH, as indicated by the unchanged brain levels of 2-AG in wild-type and FAAH−/− mice (Osei-Hyiaman et al., 2005a). 2-AG is hydrolyzed in vivo by a monoacylglyceride lipase (MGL) (Dinh et al., 2002a,b; Saario et al., 2004). A study of the ultrastructural distribution of FAAH and MGL revealed that in the hippocampus, cerebellum, and amygdala, FAAH is located postsynaptically, whereas MGL is localized in presynaptic axon terminals, including terminals of GABAergic interneurons (Gulyas et al., 2004). Correspondingly, functional studies in hippocampus indicate that depolarization-induced suppression of inhibition (DSI) is unaffected by pharmacological blockade of FAAH (Kim and Alger, 2004), but it is potentiated by blocking MGL (Kim and Alger, 2004; Makara et al., 2005), in agreement with an earlier study implicating 2-AG rather than anandamide in synaptic plasticity in the hippocampus (Stella et al., 1997). Further evidence supporting the role of 2-AG as the retrograde transmitter involved in synaptic plasticity is the preferential postsynaptic distribution of the major 2-AG biosynthetic enzyme, diacylglycerol lipase α, in hippocampus and cerebellum (Katona et al., 2006; Yoshida et al., 2006).
However, the behavioral consequences of DSI and its modulation remain unclear: selective knockout of CB1 receptors from GABAergic interneurons was found to abolish DSI and long-term depression (LTD) of inhibitory synapses, whereas the classic behavioral responses to THC remained unaffected in these animals (Monory et al., 2005). Therefore, at this point it is difficult to predict the potential therapeutic usefulness of selective MGL inhibitors.
III. The Endocannabinoid System as Therapeutic Target in Pathophysiological Conditions
A. Diseases of Energy Metabolism
1. Endocannabinoids and Appetite Regulation
It has been known since antiquity that use of cannabis in its various forms increases appetite, particularly for palatable foods, and can also result in significant weight gain (Donovan, 1845; Berry and Mechoulam, 2002). Following the identification of THC as the main psychoactive principle in marijuana, the appetite-promoting effect of smoked marijuana could be attributed to THC even before the identification of specific cannabinoid receptors (Hollister, 1971; Greenberg et al., 1976). Animal studies also documented the ability of THC to promote food intake, although consistent effects were only seen with relatively low doses (Abel, 1975), most likely because the significant sedation and motor impairment seen with higher doses interferes with the animals’ ability to initiate feeding. Variability in the observed changes in THC-induced food intake may also relate to the feeding state of the animal, the orexigenic effect being optimal in presatiated animals with low basal levels of food intake (Williams et al., 1998). After the discovery of specific cannabinoid receptors and the introduction of selective antagonists, the increase in food intake caused by THC could be linked to CB1 receptors, as it was blocked by the selective CB1 antagonist SR141716, but not by the CB2 antagonist SR144528 (Williams and Kirkham, 2002).
The discovery of endocannabinoids has raised the question of their potential involvement in the physiological control of appetite and energy metabolism. This subject has been the topic of a number of recent reviews (Di Marzo and Matias, 2005; Kirkham, 2005; Sharkey and Pittman, 2005; Pagotto et al., 2006), and only a brief summary is provided here. The first indication of a role for endocannabinoids in appetite control was the documented ability of low doses of anandamide to increase food intake, when administered either systemically (Williams and Kirkham, 1999; Hao et al., 2000) or into the ventromedial hypothalamus (Jamshidi and Taylor, 2001), and this effect could be attributed to stimulation of CB1 receptors (Williams and Kirkham, 1999). Similar increases in food intake can be elicited by 2-AG administered systemically or into the nucleus accumbens shell region (Kirkham et al., 2002) or into the lateral hypothalamus (Kirkham and Williams, 2001a). Sites for the orexigenic actions of endocannabinoids in both the hypothalamus and the limbic forebrain suggest their involvement in both the homeostatic and hedonic control of eating (Harrold and Williams, 2003; Vickers and Kennett, 2005). Interestingly, endocannabinoid activation of hypothalamic centers, such as the paraventricular nucleus, may also occur indirectly via CB1 receptors on peripheral afferent nerve terminals (Gomez et al., 2002), most likely located in the gastrointestinal tract. Such an “indirect” pathway is compatible with recent findings that CB1 mRNA is present in cholecystokinin-containing neurons in the nodose ganglion, where CB1 mRNA expression is up-regulated by fasting and down-regulated by refeeding (Burdyga et al., 2004).
Studies with antagonists provide more direct support for a regulatory function of endocannabinoids on feeding. Treatment of rats with SR141716 and the closely related CB1 antagonist AM251 reduced food intake under free-feeding (Arnone et al., 1997; Colombo et al., 1998a; Simiand et al., 1998; Chambers et al., 2003; Shearman et al., 2003) or operant conditions (Freedland et al., 2000; McLaughlin et al., 2003), suggesting antagonism of the tonic orexigenic effect of an endocannabinoid. However, SR141716 and AM251 are inverse agonists (Gifford and Ashby, 1996; Bouaboula et al., 1997), which may be an alternative mechanism by which they reduce food intake.
Definitive evidence for the involvement of endocannabinoids in the control of food intake has been provided through the use of CB1 receptor-deficient mice. In a study from our laboratory, food-deprived CB1 knockout mice were found to eat less than their wild-type littermates, and their food intake was unaffected by SR141716 treatment, whereas in wild-type mice SR141716 reduced food intake to the levels seen in the knockout mice (Di Marzo et al., 2001b). Similar findings have been subsequently reported by others (Wiley et al., 2005). This indicates that part of the hunger-induced increase in food intake is mediated by endocannabinoids acting at CB1 receptors. CB1 knockout mice are also resistant to overeating caused by neuropeptide Y (NPY) (Poncelet et al., 2003), and SR141716 inhibits the hyperphagia of leptin-deficient mice even in the absence of temporary food deprivation (Di Marzo et al., 2001b). This latter finding suggests that the absence of leptin results in increased endocannabinoid activity. Indeed, hypothalamic levels of endocannabinoids were elevated in leptin-deficient mice and rats and reduced after leptin treatment, suggesting that endocannabinoids are part of the leptin-regulated neural circuitry involved in appetite regulation (Di Marzo et al., 2001b). Endogenous leptin may similarly suppress endocannabinoid levels, as indicated by our recent unpublished findings using mice with obesity induced by a high-fat diet, which have elevated plasma leptin levels proportional to their increased fat mass. Anandamide levels were significantly lower in the obese mice compared with their lean controls in the hypothalamus, limbic forebrain, and amygdala, with no difference in the cerebellum. Furthermore, there was a significant inverse correlation between plasma leptin levels and anandamide levels in the above three brain regions involved in appetite control but not in the cerebellum.
A possible hypothalamic site for an interaction between leptin and endocannabinoids is the lateral hypothalamus, where CB1 receptors are present in orexin-and melanin-concentrating hormone (MCH)-containing neurons (Cota et al., 2003), which also express functional leptin receptors (Hübschle et al., 2001; Iqbal et al., 2001). These neurons project to dopaminergic neurons in the ventral tegmental area (Fadel and Deutch, 2002), where they modulate the mesolimbic dopaminergic pathway involved in food reward. Thus, they could also represent a site of integration of hypothalamic and extrahypothalamic structures involved in the orexigenic effect of endocannabinoids. The MCH-containing neurons are tonically inhibited by GABAergic interneurons. Jo et al. (2005) recently demonstrated that this inhibitory tone can be suppressed by the depolarization-induced release of endocannabinoids from the MCH neurons and their retrograde activation of presynaptic CB1 receptors on the GABAergic interneurons. The resulting increase in the activity of MCH neurons may contribute to the in vivo appetitive effect of endocannabinoids. Furthermore, this DSI could be blocked by leptin through inhibition of voltage-gated calcium channels in the MCH neurons, whereas it was increased 6-fold in leptin-deficient mice (Jo et al., 2005), mirroring the changes in hypothalamic endocannabinoid content by leptin and leptin deficiency reported earlier (Di Marzo et al., 2001b). Another hypothalamic site where a leptin/endocannabinoid interaction may occur is the paraventricular nucleus. At this site, glucocorticoids have been shown to induce endocannabinoid synthesis and endocannabinoid-induced suppression of synaptic excitation via a cAMP-dependent mechanism, and leptin was found to block these effects by a phosphodiesterase 3B-mediated decrease in intracellular cAMP (Malcher-Lopes et al., 2006). These effects may underlie the orexigenic action of glucocorticoids.
Another recent study indicates the importance of lateral hypothalamic orexin neurons in reward-seeking behavior in general (Harris et al., 2005), suggesting that they may also be targets of the effects of endocannabinoids on drug reward (see section III. B.11.). Additionally, cannabinoids can increase the intake of palatable foods by acting at sites in the brainstem (Miller et al., 2004), which also have reciprocal neural connections with forebrain limbic structures (Saper, 2002). From a behavioral point of view, cannabinoids are involved in both the appetitive and consummatory aspects of feeding behavior (Chaperon et al., 1998; Thornton-Jones et al., 2005), in line with their multiple sites of action in the brain. Such multiple sites of action are also indicated by findings that in THC-naive rats, rimonabant suppressed food-maintained operant responses and metabolic activity in the limbic forebrain, measured by 2-deoxyglucose uptake, whereas in rats made tolerant to THC, an additional metabolic inhibition was detected in the hypothalamus (Freedland et al., 2003). Exposure of rats to a palatable diet containing sucrose and condensed milk resulted in down-regulation of CB1 receptors in limbic structures involved in the hedonic aspects of feeding, but not in the hypothalamus (Harrold et al., 2002). In the hypothalamus, the very low density of CB1 receptors is offset by their increased coupling (Breivogel et al., 1997), which may be an alternative target of regulation (Basavarajappa and Hungund, 1999; Wang et al., 2003) that needs to be explored.
Within the appetitive neural circuitry, endocannabinoids have been shown to interact with both orexigenic factors such as endogenous opioids, NPY, orexins, and ghrelin, and anorexigenic factors including α-melanocyte-stimulating hormone (α-MSH), corticotropin-releasing hormone (CRH), and the peptide product of the cocaine and amphetamine-related transcript (CART). Inhibition of food intake by opioid μ receptor antagonists and CB1 receptor antagonists is supra-additive (Kirkham and Williams, 2001b; Rowland et al., 2001; Chen et al., 2004), suggesting a synergism between the endogenous opioid and cannabinoid systems in mediating the reinforcing effect of food (Solinas and Goldberg, 2005). Indeed, CB1-deficient mice fail to self-administer morphine (Ledent et al., 1999; Cossu et al., 2001) or to release dopamine in the nucleus accumbens in response to morphine (Mascia et al., 1999), suggesting that the site of this synergism is in the mesolimbic dopaminergic pathway, which is involved in both drug and food reward (Le Foll and Goldberg, 2005). The observation that SR141716 inhibits the orexigenic effect of morphine microinjected into the hypothalamic paraventricular nucleus but not the nucleus accumbens shell suggests additional interactions between the two systems, unrelated to the hedonic aspects of feeding (Verty et al., 2003). A further intriguing parallel between the two systems is that opiate μ receptor knockout mice, just as CB1−/− mice (see below), are resistant to diet-induced obesity (Tabarin et al., 2005).
As for interactions with NPY, the similar effectiveness of SR141716 to inhibit food intake in wild-type and NPY−/− mice indicates that endocannabinoids are unlikely to be the primary compensatory factor that accounts for the lack of a lean phenotype in NPY−/− mice (Di Marzo et al., 2001b). However, anandamide was found to increase and AM251 to decrease depolarization-induced NPY release in rat hypothalamic explants, suggesting that NPY may contribute to the orexigenic effects of cannabinoids (Gamber et al., 2005). A possible role of orexins in the appetitive effects of endocannabinoids is suggested by the finding that coexpression of the CB1 and orexin 1 receptors results in a marked potentiation of orexin A-induced signaling (Hilairet et al., 2003). An important site of action of the orexigenic peptide ghrelin is the hypothalamic paraventricular nucleus, where its hyperphagic effect can be blocked by SR141716, suggesting that ghrelin may act via the release of endocannabinoids (Tucci et al., 2004). Endocannabinoids, in turn, may be involved in ghrelin release, at least in the periphery, as suggested by an SR141716-induced decrease in plasma ghrelin levels in rats (Cani et al., 2004).
The proopiomelanocortin-derived peptide α-MSH acting at MC-4 melanocortin receptors is part of the leptin-regulated appetitive circuitry as a major anorectic mediator. The observations that SR141716 inhibits the feeding response induced by blocking MC-4 receptors, whereas α-MSH does not affect THC-induced feeding, suggest that CB1 receptors are downstream from MC-4 receptors and have an obligatory role in α-MSH effects on food intake (Verty et al., 2004). The peptide product of CART is also a tonically active anorectic mediator (Kristensen et al., 1998) and, unlike α-MSH, may be a downstream mediator of the effect of endocannabinoids. Such an arrangement is suggested by the finding that SR141716 loses its ability to reduce food intake in CART−/− mice (Osei-Hyiaman et al., 2005a). Furthermore, mice deficient in FAAH have reduced levels of CART immunoreactivity in various hypothalamic and extrahypothalamic regions involved in appetite control, which is returned to normal levels by chronic SR141716 treatment (Osei-Hyiaman et al., 2005a). These findings suggest that inhibition of CART release by CB1 activation may be involved in the orexigenic effect of anandamide. Finally, an interaction between endocannabinoids and CRH is indirectly suggested by coexpression of the mRNA for the CB1 receptor with the mRNA for CRH (Cota et al., 2003) or the CRH type 1 receptor (Hermann and Lutz, 2005).
2. Endocannabinoids and Peripheral Energy Metabolism
It is generally accepted that energy intake and utilization are regulated in a coordinated fashion, and factors involved in the central regulation of appetite may also affect peripheral energy metabolism (Seeley and Woods, 2003). The first indirect indication that cannabinoids may affect energy homeostasis through a mechanism other than food intake came from a study of marijuana smokers tested in a hospital inpatient setting (Greenberg et al., 1976). In this study, the marijuana-induced increase in caloric intake leveled off after a few days, whereas weight gain continued throughout the rest of the 21-day observation period, suggesting independent effects on appetite and peripheral energy metabolism. After the introduction of SR141716 as the first selective CB1 receptor antagonist (Rinaldi-Carmona et al., 1994), a similar conclusion was reached in normal rats treated with SR141716 for 14 days. Tolerance to the anorectic effect of SR141716 developed within 5 days, whereas the reduction in body weight was maintained throughout the treatment period (Colombo et al., 1998a). Later, similar observations were reported in mice with diet-induced obesity, in which food intake was reduced transiently whereas the reduction in body weight was maintained when the animals were chronically treated with SR141716 (Ravinet Trillou et al., 2003) or AM251 (Hildebrandt et al., 2003). These results suggested that factors other than appetite must be involved in the weight-reducing effect of CB1 antagonists.
Peripheral targets of endocannabinoids include adipocytes, which express CB1 receptors (Bensaid et al., 2003; Cota et al., 2003). Stimulation of CB1 receptors on adipocytes can affect lipid metabolism through regulating the level of adiponectin production (Bensaid et al., 2003), by increasing lipoprotein lipase activity (Cota et al., 2003), or by inhibiting AMP-activated protein kinase (AMPK) (Kola et al., 2005), which leads to increased lipogenesis and decrease in fatty acid β-oxidation through reducing the phosphorylation and thus disinhibiting acetyl CoA carboxylase-1 (ACC1), the rate-limiting enzyme in fatty acid synthesis. The work by Cota et al. (2003) provided the first clear evidence of peripheral metabolic targets of endocannabinoids in vivo in a mouse model of diet-induced obesity. By careful analysis of body composition, they were able to establish the lean phenotype of CB1-deficient mice that had escaped earlier attention. Furthermore, the use of a pair-feeding paradigm revealed that hypophagia accounts for the lean phenotype only in young and not in adult animals, which clearly indicated the involvement of peripheral metabolic target(s) in the latter. The additional documentation of functional CB1 receptors in primary cultured adipocytes and their role in regulating lipogenesis provided one of the likely peripheral targets for the anabolic effects of endocannabinoids. The lean phenotype of CB1−/− mice in this study was more prominent in male than in female animals, which could suggest that endocannabinoid regulation of adiposity may be subject to modulation by sex hormones.
Although earlier studies failed to detect CB1 receptors in the liver, more recently they have been identified in the mouse liver using a combination of methods including reverse transcription-polymerase chain reaction, in situ hybridization, immunohistochemistry, and Western blotting. In the same study, treatment of mice with the cannabinoid agonist HU-210 increased de novo lipogenesis and the expression of the transcription factor sterol regulatory element binding protein 1c (SREBP1c) as well as of its targets, ACC1 and fatty acid synthase (Osei-Hyiaman et al., 2005b). The role of CB1 receptors in these effects was indicated by the ability of SR141716 to block them and by their absence in CB1 knockout mice (Osei-Hyiaman et al., 2005b). The hepatic lipogenic pathway may be also directly activated through a cannabinoid-induced decrease in AMPK phosphorylation and activity in the liver (Kola et al., 2005). CB1 receptors have been also detected in rat hepatocytes (Michalopoulos et al., 2003), in whole mouse liver (Biecker et al., 2004), and in rat and human hepatic stellate cells (Siegmund et al., 2005; Teixeira-Clerc et al., 2006).
Fatty acid metabolism in hypothalamic neurons acts as a sensor of nutrient availability (Obici et al., 2003), and its pharmacological modulation influences food intake (Kim et al., 2004). CB1 activation was reported to increase SREBP1c and FAS gene expression in the hypothalamus, and the increased expression of these genes by fasting/refeeding (Paulaskis and Sul, 1988) could be inhibited by SR141716 treatment at the beginning of the refeeding period, which also reduced food intake (Osei-Hyiaman et al., 2005b). Although fatty acid synthesis was not measured directly in the hypothalamus, these findings suggest that the increase in food intake after fasting may involve a CB1-mediated modulation of the fatty acid synthetic pathway. Modulation of AMPK activity by cannabinoids was documented not only in liver and adipose tissue but also in hypothalamus (Kola et al., 2005), where it has been linked to appetite control (Minokoshi et al., 2004). Thus, the AMPK/ACC1/FAS pathway may represent a common molecular pathway involved in both the central appetitive and the peripheral metabolic effects of endocannabinoids.
Because total caloric intake is similar in wild-type and CB1−/− mice on a high-fat diet (Ravinet Trillou et al., 2004; Osei-Hyiaman et al., 2005b), the resistance of CB1-deficient mice to diet-induced obesity must be associated with increased energy expenditure. Exposing wild-type C57BL6/J mice to a high-fat diet decreases energy expenditure, as documented by indirect calorimetry (Hu et al., 2004), which may account for the increase in feed efficiency observed in such animals, whereas in CB1−/− mice feed efficiency was unaffected by a high-fat diet (Osei-Hyiaman et al., 2005b). This suggests that the high-fat diet-induced decrease in energy expenditure is mediated by endocannabinoid activation of CB1 receptors. Accordingly, HU-210 treatment of wild-type mice decreased and SR141716 treatment increased the activity of carnitine palmitoyl transferase-1, the rate-limiting enzyme in fatty acid β-oxidation (D. Osei-Hyiaman and G. Kunos, unpublished observations).
One of the factors involved in this effect in vivo could be adiponectin, the adipocyte-derived hormone that promotes fatty acid β-oxidation (Yamauchi et al., 2002). Indeed, exposure to a high-fat diet resulted in a significant decline in plasma adiponectin in wild-type but not in CB1−/− mice (Osei-Hyiaman et al., 2005a), and CB1 receptor activation in isolated adipocytes was found to suppress adiponectin expression (Perwitz et al., 2005; Matias et al., 2006). Expression of the thermogenic uncoupling protein-1 was also down-regulated by CB1 activation, whereas the expression of the insulin-mimetic adipokine visfatin was increased (Perwitz et al., 2005). Conversely, rimonabant increases adiponectin secretion by adipocytes (Bensaid et al., 2003) and adiponectin plasma levels in obese human subjects (Després et al., 2005), which should lead to increased lipid β-oxidation and thermogenesis in vivo. Chronic treatment of ob/ob mice with SR141716 increased thermogenesis, as indicated by increased oxygen consumption at a thermoneutral temperature measured by whole body calorimetry (Liu et al., 2005). Glucose uptake, subsequently measured in the isolated soleus muscle of these animals, was significantly increased in the SR141716-pretreated group. A similar effect in humans may account for the increased glucose tolerance observed in obese patients treated with rimonabant (Van Gaal et al., 2005). These observations could suggest the presence of CB1 receptors in skeletal muscle, which was recently documented (Pagotto et al., 2006). Alternatively, increased glucose tolerance may be secondary to an effect of SR141716 on CB1 receptors in the liver. It has been proposed that increased lipid synthesis in the liver may produce insulin resistance in other tissues such as muscle (McGarry, 1992), and CB1 receptor activation has been shown to contribute to the development of hepatic steatosis in diet-induced obesity (Osei-Hyiaman et al., 2005b). Endocannabinoids may also influence insulin secretion directly in islet β-cells via CB1 (Matias et al., 2006) or CB2 receptors (Juan-Pico et al., 2005).
The ability of rimonabant to increase energy expenditure may not be limited to an effect on adiponectin secretion, as indicated by an analysis of the effect of rimonabant treatment on gene expression profiles in lean and diet-induced obese mice as well as CB1−/− mice (Jbilo et al., 2005). Rimonabant-induced decreases in body weight and adipose tissue mass in obese mice was accompanied by a near-complete reversal of obesity-induced changes in the expression of a wide range of genes. These included genes involved in adipocyte differentiation, lipolysis, generation of futile cycles, and glycolysis. These broad-based targets may underlie the ability of rimonabant to correct symptoms of the metabolic syndrome, as discussed below. They also raise the intriguing possibility that if a CB1 antagonist that does not cross the blood-brain barrier were available, it could be effective in the treatment of the metabolic syndrome without the risk of adverse CNS side effects (Horvath, 2006).
3. Obesity and Associated Metabolic Abnormalities
Genetic manipulation of the expression of endogenous proteins has been instrumental in uncovering their regulatory role in normal and pathological phenotypes. When CB1 knockout mice were first introduced, no change in body mass or feeding pattern had been noted (Ledent et al., 1999; Zimmer et al., 1999). However, in a subsequent study, CB1 knockout mice were found to have a life-long, small, but significant, weight deficit compared with their wild-type littermates, which could be attributed to a selective deficit in adipose tissue mass (Cota et al., 2003) and was confirmed by others (Ravinet Trillou et al., 2004; Osei-Hyiaman et al., 2005b). Parallel to their decreased fat mass, CB1−/− mice have lower plasma leptin levels and an increased sensitivity to the anorectic effect of exogenous leptin (Ravinet Trillou et al., 2004).
The possibility that an increase in the activity of the endocannabinoid system may contribute to at least some forms of obesity was suggested by three sets of findings. First, CB1 antagonists were significantly more efficacious in reducing caloric intake and body weight in rodents with diet-induced or genetic obesity than in their respective lean controls (Di Marzo et al., 2001b; Hildebrandt et al., 2003; Ravinet Trillouet et al., 2003; Vickers et al., 2003).
Second, CB1−/− mice are resistant to diet-induced obesity (Ravinet Trillou et al., 2004; Osei-Hyiaman et al., 2005b). In both of these studies, overall caloric intake was not different between wild-type compared with CB1−/− mice receiving the high-fat diet, suggesting that peripheral mechanisms play a dominant role in the control of body weight by CB1 receptors. CB1−/− mice are also resistant to the metabolic changes that accompany diet-induced obesity in normal mice, including hypertriglyceridemia and elevated plasma leptin and insulin levels, indicative of leptin and insulin resistance, respectively (Ravinet Trillou et al., 2004; Osei-Hyiaman et al., 2005b). These metabolic changes, collectively defined by some as the “metabolic syndrome”, could also be reversed by SR141716 treatment (Ravinet Trillou et al., 2004; Poirier et al., 2005).
As a third line of evidence, recent findings indicate that endocannabinoids and CB1 receptors are up-regulated in the liver and adipose tissue in various forms of experimental as well as in human obesity. In wild-type mice on a high-fat diet for 3 weeks, the basal rate of de novo hepatic fatty acid synthesis was markedly increased, and the increase was partially reversed by SR141716 treatment (Osei-Hyiaman et al., 2005b). After 3 weeks of diet, the mice were not yet overweight but showed significant hepatic steatosis. Their hepatic content of anandamide was increased 3-fold, and the level of CB1 receptor protein in liver plasma membranes was also markedly increased (Osei-Hyiaman et al., 2005b). These findings indicate that intake of a high-fat diet activates the hepatic endocannabinoid system, which contributes to increased lipogenesis and the subsequent development of hepatic steatosis and, ultimately, the development of obesity. Exposure of C57BL6/J mice to a high-fat diet has been reported to induce changes characteristic of the metabolic syndrome and also to rapidly induce the expression of SREBP1c and its downstream target lipogenic enzymes (Biddinger et al., 2005). CB1 receptor knockout mice are resistant to these diet-induced changes, which indicates that endocannabinoids have a major role in mediating them (Osei-Hyiaman et al., 2005b).
An up-regulation of CB1 receptors has been also reported in adipose tissue of genetically obese compared with lean mice (Bensaid et al., 2003), and elevated endocannabinoid levels have been detected in adipose tissue of obese compared with lean patients (Matias et al., 2006). In a study involving 40 women (Engeli et al., 2005), circulating levels of anandamide and 2-AG were significantly increased in 20 obese versus 20 lean subjects, and remained elevated after a 5% diet-induced weight reduction. Although these plasma levels were much too low to exert hormone-like activity, they probably originate from overflow from tissues and thus may reflect functionally relevant changes in endocannabinoid content at or near sites of action. In the same study, FAAH expression was markedly reduced in the adipose tissue of obese subjects and correlated negatively with circulating endocannabinoid levels. Furthermore, the expression of both CB1 and FAAH increased in mature adipocytes compared with preadipocytes. These findings suggest that the endocannabinoid system is activated in human obesity (Engeli et al., 2005).
A genetic missense polymorphism in the FAAH gene predicting a proline to threonine substitution at position 129, which was reported to result in reduced cellular expression and activity of the enzyme (Chiang et al., 2004), had been earlier found to be significantly associated with problem drug use (Sipe et al., 2002). The same polymorphism has been linked to overweight and obesity in both Caucasian and African-American subjects (Sipe et al., 2005). Interestingly, the elevated hepatic levels of anandamide in mice receiving a high-fat diet could be attributed to a decrease in FAAH activity (Osei-Hyiaman et al., 2005b), suggesting that FAAH may play a key role in regulating endocannabinoid “tone” in both experimental and human obesity. Although this finding could suggest the targeting of FAAH in the treatment of eating/metabolic disorders, such an approach will be complicated by the fact that oleylethanolamide, an anorectic lipid that acts on the peroxisome proliferator-proliferator-activated receptor α (PPARα) (Fu et al., 2003), is also a substrate for FAAH. The opposing effects of elevated levels of both anandamide and oleylethanolamide after pharmacological blockade of FAAH may therefore result in no net change in appetite and energy metabolism.
That increased endocannabinoid activity may also contribute to obesity and its metabolic consequences in humans was indicated by the highly promising results of recent clinical trials with rimonabant. As in the animal models of diet-induced obesity, rimonabant was effective both in reducing body weight and in reversing many of the associated metabolic abnormalities in obese subjects. In a multicenter, phase III study involving 1507 obese European subjects with a body mass index >30 kg/m2 or a body mass index >27 kg/m2 with dyslipidemia and moderate hypertension, rimonabant (20 mg/day) treatment for 1 year, combined with a moderately hypocaloric diet, not only reduced body weight but also reduced plasma triglycerides, increased HDL cholesterol, and decreased plasma insulin and insulin resistance (Van Gaal et al., 2005). Blood pressure was not significantly affected. The parallel reduction in body weight and waist circumference suggested that the weight loss was predominantly due to loss of visceral fat, which is known to be a predisposing factor for the metabolic syndrome. Rimonabant was well tolerated, with mild to moderate nausea, diarrhea, and mood disorders occurring slightly more in the treatment group than in the placebo group (Van Gaal et al., 2005).
Essentially similar findings were reported in another large-scale, phase III study (RIO-North America) involving 3045 randomized, obese or overweight subjects. At the end of the 1st year, rimonabant-treated subjects were re-randomized to receive rimonabant or placebo, whereas the placebo group continued onto receive the placebo. During the 2nd year, rimonabant-treated patients retained the improvements achieved during the 1st year, whereas those who switched to placebo regained their original weight (Pi-Sunyer et al., 2006).
In a third study (RIO-Lipids) involving 1036 overweight/obese subjects, 20 mg/day rimonabant taken for 1 year significantly reduced body weight (−6.3 ± 0.5 kg), weight circumference (−5.7 ± 0.6 cm), and plasma triglycerides (−12.4 ± 3.2%), increased HDL cholesterol by 8.1 ± 1.5% and increased LDL particle size, improved glucose tolerance, and significantly elevated plasma adiponectin levels, resulting in a 50% decrease in the prevalence of the metabolic syndrome in the study population (Després et al., 2005). In contrast with the other two studies, a statistically significant, small decrease in systolic and diastolic blood pressure was evident in the group receiving 20 mg of rimonabant, and the decrease was greater for patients with initial hypertension (blood pressure >140/90 mm Hg). Although the reason for the lack of a blood pressure change in the other studies is not clear, the proportion of females was lower in RIO-Lipids (~60%) than in the other two studies where they represented ~80% of subjects. It is possible that a modest reduction in blood pressure by rimonabant occurs preferentially in males. The cumulative finding that blood pressure reduction, if present, is less than expected based on a similar level of weight reduction alone (Appel et al., 2003), is noteworthy. As discussed in section D.1., rimonabant at an i.v. dose of 3 mg/kg causes a pressor response in anesthetized, hypertensive rats, which are supersensitive to the hypotensive effect of endogenous or exogenous anandamide (Bátkai et al., 2004). Although the pressor effect is much smaller at lower doses of rimonabant comparable with the 20-mg oral dose used in humans or in the absence of anesthesia (S. Bátkai, P. Pacher, and G. Kunos, unpublished observations), careful monitoring of blood pressure, particularly in the early stages of rimonabant treatment, may be advisable. A polymorphism in the FAAH gene is associated with obesity (Sipe et al., 2005), and because of the reduced enzyme activity resulting from this polymorphism, some of the affected individuals may have an elevated endocannabinoid tone, reversal of which by rimonabant could increase blood pressure.
It is noteworthy that part of the rimonabant-induced improvements in the hormonal and lipid abnormalities in the three clinical studies appeared to be independent of weight reduction and, based on the preclinical findings discussed above, are most likely mediated via peripheral sites of action. An interesting alternative mechanism is suggested by the results of a recent meta-analysis of the effects of low carbohydrate, nonenergy-restricted diets on weight loss and cardiovascular risk factors (Nordmann et al., 2006). Such diets were found to lead to significant weight loss for up to 1 year. Surprisingly, they were more favorable than low-fat diets in reducing plasma triglycerides and increasing HDL cholesterol levels, without a favorable effect on total or LDL cholesterol. The pattern of these metabolic changes is similar to that of those caused by 20 mg of rimonabant in the three clinical trials. Rimonabant has been shown to preferentially suppress the preference for sweet compared with normal (Simiand et al., 1998) or high-fat reinforcers (Ward and Dykstra, 2005) and can cause longer lasting suppression of intake of sweet compared with normal food (Gessa et al., 2006). It is very possible that obese subjects treated with rimonabant unwittingly altered their diet by reducing carbohydrate intake, which may have contributed to the observed effects on triglycerides and HDL cholesterol. Detailed analyses of the effects of rimonabant on dietary habits are warranted.
Overall, the findings in these three large, multicenter clinical trials strongly support a pathogenic role of increased endocannabinoid activity in obesity and the associated metabolic abnormalities and highlight the unique therapeutic potential of CB1 blockade. Additional benefits may be gained by combination therapies. The efficacy of statins to preferentially lower LDL cholesterol may be effectively complemented by the ability of rimonabant to increase HDL cholesterol. In the case of insulin, the ability of rimonabant ability to increase insulin sensitivity could reduce the dose requirement for insulin in obese diabetic subjects and could also counteract the tendency of insulin treatment to cause weight gain. Nevertheless, further large-scale studies are warranted in view of the high nonadherence rate observed in the three clinical trials to date, which may have resulted in overestimation of the benefits of treatment (Simons-Morton et al., 2006).
4. Cachexia and Anorexia
A negative energy balance resulting from decreased appetite and food intake and increased energy expenditure, leading to weight loss, can be the consequence of wasting diseases such as AIDS or metastatic cancer, or it could be associated with aging, chemotherapy of cancer, or neuropsychiatric conditions such as anorexia nervosa or various forms of dementia including Alzheimer’s disease. Although there is a growing body of evidence documenting the therapeutic effectiveness of synthetic THC or even smoked marijuana as appetite boosters in some of these conditions (Regelson et al., 1976; Gorter et al., 1992; Nelson et al., 1994; Beal et al., 1995, 1997; Timpone et al., 1997) (Table 1), there is only limited information on the potential involvement of the endocannabinoid system in their pathogenesis.
TABLE 1.
Clinical trials with cannabinoid-related medications in human disease
| Disease/Condition | Sample Size, Design, Target Symptoms | Compound (Dose) | Parameters Studied | Results | Adverse Effects | Reference |
|---|---|---|---|---|---|---|
| MS and SCI | ||||||
| MS | Nine patients, DB, PL, spasticity | THC (5- and 10-mg single-dose p.o.) | EMG, clinical | Improved spasticity score (objective) | Minimal | Petro and Ellenberger (1981) |
| MS | Eight patients, SB, PL, tremor, ataxia | THC (5 mg/6 h max three doses p.o.) | Clinical | Improved coordination and sense of well being, decreased tremor (subjective) | Subjective “high” in all patients | Clifford (1983) |
| MS | 13 patients, DB, PL, C, spasticity | THC (2.5–15 mg daily for 5 days p.o.) | Clinical, questionnaire | Reduced spasticity (subjective); objective function tests not improved | Common | Ungerleider et al. (1987) |
| MS | One patient, OL, spastic tetraparesis | Cigarette smoke marijuana (one cigarette) | Clinical, tremor recording, EMG | Reduced ataxia and spasticity (objective) | None | Meinck et al. (1989) |
| MS | 10 patients, DB, C, spasticity | Cigarette smoke marijuana (one cigarette; 1.54% THC) | Dynamic posturography, objective balance | Impaired posture and balance | Subjective unpleasant “high” in all patients | Greenberg et al. (1994) |
| MS | One patient, DB, PL, C, spasticity | Nabilone (1 mg/2 days for 16 wk p.o.) | Visual analog scales | Improved painful muscle spasms, mood and well being (subjective); reduced frequency of nocturia | Mild sedation | Martyn et al. (1995) |
| MS and SCI | Two patients, OL, spasticity | THC (10 or 15 mg p.o. or rectal) | Clinical | Improved walking ability and passive mobility, reduced rigidity, slight pain relief | Temporal deterioration in ability to concentrate and in mood | Brenneisen et al. (1996) |
| MS | One patient, PL, nystagmus | Cigarette smoke marijuana (inhaled) | Eye movement recording | Reduced nystagmus amplitude and improved visual acuity | None | Schon et al. (1999) |
| MS | 16 patients, DB, PL, C, spasticity | Plant extract of THC (2.5–5 mg b.i.d. for 4 wk p.o.) | Clinical, questionnaires, Ashworth score | No improvement in Ashworth scale, worsening global impression | 41 adverse events in 16 patients during plant extract treatment | Killestein et al. (2002) |
| MS and SCI | 24 patients, DB, PL, C, heterogeneous | Plant extract of THC and CBD 1:1 (2.5–120 mg/day for 2 wk sublingual) | Clinical, questionnaires | Improvement of bladder control, muscle spasms, and spasticity and pain relief (subjective) but no in Ashworth scale | 4 dropouts due to adverse events | Wade et al. (2003) |
| MS | 630 patients, DB, R, PL, spasticity | Cannabis extract (Cannador: 2.5 mg Δ9-THC + 1.25 mg CBD/capsule; Marinol: THC max 25 mg/day for 15 wk p.o.) | Clinical, questionnaires Ashworth score, Rivermead Mobility Index | No change in the Ashworthscore, but improvement in the patient-reported spasticity, pain, and sleep quality; unexpected reduction in hospital admission for relapse in the treatment groups; in 12-mo follow-up, THC improved muscle spasticity measured by the Ashworth scale and the Rivermead Mobility Index | Minimal, similar to placebo | Zajicek et al. (2003, 2004) |
| MS | 57 patients, DB, R, PL, C, spasticity, various | Cannabis-based capsules (2.5 mg THC and 0.9 mg CBD; max dose 30 mg/day THC p.o.) | Self-report of spasm frequency and symptoms, Ashworth Scale, Rivermead Mobility Index, 10-m timed walk | Improved spasm frequency and mobility in the 37 patients who received at least 90% of their prescribed dose | Minor adverse events were slightly more frequent in treated group | Vaney et al. (2004) |
| MS | 14 patients, DB, PL, tremor | Cannabis extract (Cannador: 2.5 mg Δ9-THC + 1.25 mg CBD/capsule p.o. for 2 wk) | Tremor index, measured using a validated tremor rating scale | No effects on tremor | Minimal | Fox et al. (2004) |
| MS | 57 patients, DB, R, PL, C, spasticity, various | Cannabis-based capsules (2.5 mg THC and 0.9 mg CBD; max dose 30 mg/day THC p.o.) | Self-report of spasm frequency and symptoms, Ashworth Scale, Rivermead Mobility Index, 10-m timed walk | Improved spasm frequency and mobility in the 37 patients who received at least 90% of their prescribed dose | Minor adverse events were slightly more frequent in treated group | Vaney et al. (2004) |
| MS | 14 patients, DB, PL, tremor | Cannabis extract (Cannador: 2.5 mg Δ9-THC + 1.25 mg CBD/capsule p.o. for 2 wk) | Tremor index, measured using a validated tremor rating scale | No effects on tremor | Minimal | Fox et al. (2004) |
| MS | 160 patients, DB, PL, R, M, VAS score for each patient’s most troublesome symptom | GW-1000 (Sativex) delivered by oromucosal spray (2.7 mg Δ9-THC and 2.5 mg CBD at each actuation) | VAS score for each patient’s most troublesome symptom, Ashworth Scale | No significant difference in the Ashworth scale, tremor, and pain at 6 wk between the Sativex and placebo groups; improved VAS scores for spasticity | Minimal | Wade et al. (2004) |
| Pain (see also MS above) | ||||||
| Cancer | 10 patients, P, non-R, non-DB, pain | THC (5, 10, 15, or 20 mg p.o.) | Cancer-associated pain | Superior to PL | Common at higher doses | Noyes et al. (1975a) |
| Cancer | 34 patients, P, non-R, C, pain | THC (20 mg, codeine 120 mg p.o.) | Cancer-associated pain | Both superior to PL | Common with THC | Noyes et al. (1975b) |
| Cancer | 45 patients, DB, C, P, pain | NIB (4 mg, codeine 50 mg, secobarbital 50 mg) | Cancer associated pain | NIB equal to codeine, superior to secobarbital and PL | Common | Staquet et al. (1978) |
| Dental extraction | 10 patients, DB, R, PL | THC (0.022, 0044 mg, diazepam 0.157 mg/kg i.v.) | Surgical pain | THC superior to PL, inferior to diazepam | Not discussed | Raft et al. (1977) |
| FMF | One patient, DB, R, C | THC (50 mg daily p.o.) | Gastrointestinal pain | Superior to PL | Not discussed | Holdcroft et al. (1997) |
| MS | One patient, OL, pain | Nabilone (1 mg b.i.d. p.o.) | Questionnaire, various | Complete pain relief | None | Hamann and di Vadi (1999) |
| MS | 66 patients, DB, R, PL, pain, sleep disturbances | Sativex delivered by oromucosal spray (2.7 mg Δ9-THC and 2.5 mg CBD at each actuation) | Pain, sleep disturbances, numerical rating scale | Improved central neuropathic pain and sleep disturbances | Minimal | Rog et al. (2005) |
| Neuropathy of varying etiologies | 21 patients, DB, R, C, PL, pain | Ajulemic acid (CT-3, IP-751: 4 or 10 mg p.o. two times daily) | Neuropathic pain, VAS | Significant reduction of chronic neuropathic pain | Minimal | Karst et al. (2003) |
| HIV | 523 patients, cross-sectional questionnaire study | Cannabis | Questionnaire, various | In most patients who used cannabis to treat symptoms (143/523); reduction in muscle and neuropathic pain | Not discussed | Woolridge et al. (2005) |
| Anorexia-cachexia in patients with cancer, HIV, or AIDS | ||||||
| Cancer | 54 patients, R, DB, weight | THC (three doses of 0.1 mg/kg/day p.o.) | Appetite, weight | Improved appetite and increased weight | Dizziness, sedation, confusion | Regelson et al. (1976) |
| Cancer | 19 patients, OL, non-R, weight | THC (three doses of 5 mg/day p.o.) | Appetite, weight | Improved appetite, trends for weight increase | Common, but well-tolerated | Nelson et al. (1994) |
| HIV/AIDS | 10 patients, non-R, weight | THC (three doses of 2.5 mg/day p.o.) | Weight | Increased/stabilized weight | Mild | Gorter et al. (1992) |
| AIDS | 139 patients, R, PL, weight | THC (two doses of 2.5 mg/day p.o.) | VAS for hunger, weight | Improved VAS for hunger but not weight | Mild | Beal et al. (1995) |
| AIDS | 94 patients, non-R, OL, weight | THC (two doses of 2.5 mg/day p.o.) | VAS for hunger, weight | Improved VAS for hunger and weight (only for 1 month) | Sedation, psychosis, dysphoria | Beal et al. (1997) |
| AIDS | 52 patients, weight | THC (two doses of 2.5 mg/day p.o. +/− Megace) | VAS for hunger, weight | Less effective than Megace | Anxiety, euphoria, psychosis, confusion | Timpone et al. (1997) |
| Chemotherapy-induced nausea and vomiting | ||||||
| Chemotherapy-induced nausea and vomiting | Review of 30 randomized trials involving 1366 patients, nausea, vomiting | THC | Nausea, vomiting | Across all trials, cannabinoids were more effective than placebo | Various | Tramèr et al. (2001) |
| Traumatic brain injury | ||||||
| Closed head injury | 67 patients, R, DB, PL, phase II, M, neurological outcome | HU-211 (dexanabinol: 48 or 150 mg i.v.) | Intracranial, cerebral perfusion and blood pressure, Glasgow scale | Better intracranial pressure/cerebral perfusion pressure control, trends towards better neurological outcome | Similar in all groups, related to severe head trauma | Knoller et al. (2002) |
| Traumatic brain injury | 861 patients, R, PL, phase III, M, neurological outcome | HU-211 (dexanabinol: 150 mg i.v.) | Extended Glasgow scale at 6 months | No improvement | Similar in all groups, related to severe head trauma | Maas et al. (2006) |
| Parkinson’s disease, levodopa-induced dyskinesia | ||||||
| Parkinson’s disease | 24 patients, R, DB, PL, motor disability | SR141716 (0.3 mg/kg p.o.); antagonists of NK3 R (SR142801) and neurotensin receptors (SR48692) | Motor symptoms and levodopa-induced dyskinesias after a single dose of levodopa | No improvement in parkinsonian motor disability with any of drugs tested | Minimal | Mesnage et al. (2004) |
| Parkinson’s disease | Seven patients, R, DB, PL, C, motor disability | Nabilone | Motor symptoms | Reduces levodopa-induced dyskinesia in PD | Minimal | Sieradzan et al. (2001) |
| Alzheimer’s disease, dementia | ||||||
| Alzheimer’s disease, dementia | Six patients, OL, pilot, neuropsychiatric symptoms | Dronabinol (2.5 mg/day for 2 wk) | Neuropsychiatric Inventory score, subscores for agitation, aberrant motor, and nighttime behaviors | Significant improvement in Neuropsychiatric Inventory total score, subscores for agitation, aberrant motor, and nighttime behaviors | Minimal | Walther et al. (2006) |
| Primary dystonia | ||||||
| Primary dystonia | 15 patients, DB, R, PL, C, dystonia | Nabilone (0.03 mg/kg) | Motor symptoms | No improvement | Minimal | Fox et al. (2002b) |
| Tourette’s syndrome | ||||||
| Tourette’s syndrome | 12 patients, R, DB, PL, C, behavioral disorders | THC (one dose of 5, 7.5, or 10 mg) | Vocal and motor tics, various clinical scales | Significant improvement of vocal and motor tics | Minimal | Müller-Vahl et al. (2002) |
| Tourette’s syndrome | 24 patients, R, DB, PL, behavioral disorders | THC (5–10 mg) | Vocal and motor tics, various clinical scales | Significant improvement of vocal and motor tics | Minimal | Müller-Vahl et al. (2003a) |
| Psychosis/schizophrenia/schizoaffective disorder | ||||||
| Schizophrenia/schizoaffective disorder | 481 patients, PL, DB, positive and negative symptoms | SR141716; SR 142801 (NK3 R antagonist); SR46349B (5-HT2A/2C R antagonist) | Various symptom scales | No improvement with CB1 antagonist, slight improvement with NK3 R and 5-HT2A/2C R antagonist | Minimal | Meltzer et al. (2004) |
| Obesity | ||||||
| Obesity, metabolic syndrome | 1507 patients, R, DB, PL, weight | Rimonabant (5, 20 mg, 12 mo) | Weight, WC, BP, lipids, insulin, glucose | Weight and WC reduction, improved lipids, glucose tolerance, metabolic syndrome | Mild (nausea, arthralgia, diarrhea) | Van Gaal et al. (2005) |
| Obesity, metabolic syndrome | 1036 patients, R, DB, PL, weight | Rimonabant (5, 20 mg, 12 m) | Weight, waist c, lipids, glucose, insulin, leptin | Weight and WC reduction, improved cardiovascular risk, reduced metabolic syndrome; decreased BP, increased plasma adiponectin | Mild (nausea, anxiety, diarrhea, insomnia) | Després et al. (2005) |
| Obesity, metabolic syndrome | 3045 patients, R, DB, PL, M, weight, cardiometabolic risk factors | Rimonabant (5, 20 mg, 2 yr) | Weight, WC, lipids, glucose, insulin, Leptin | Weight and WC reduction, improved cardiovascular risk, reduced metabolic syndrome; weight regained in 2nd year with placebo | Mild (nausea, anxiety, diarrhea, insomnia) | Pi-Sunyer et al. (2006) |
DB, double blind; PL, placebo-controlled; EMG, electromyography; SB, single blind; C, crossover; OL, open-label; max, maximum; R, randomized; M, multicenter; VAS, visual analog scale; P, prospective; PL, placebo; FMF, familial Mediterranean fever; NIB, nitrogen analog of THC; NK3 R, neurokinin 3 receptor; SR142801, (R)-(N)-[1-[3-[1-benzoyl-3-(3,4-dichlorophenyl)piperidin-3-yl]propyl]-4-phenylpiperidin-4-yl]-N-methylacetamide; SR48692, 2-[(1-(7-chloro-4-quinolinyl)-5-(2,6-dimethoxyphenyl)pyrazol-3-yl)carbonylamino]tricycle(3.3.1.1)decan-2-carboxylic acid}; SR463493B, 1(Z)-[2-(dimethylamino)ethoxyimino]-1(2-fluorophenyl)-3-(4-hydroxyphenyl)-2(E)-propene; 5-HT2A/2C R, serotonin 2A/2C receptor, WC, waist circumference; BP, blood pressure.
A few studies have reported the effectiveness of THC in stimulating appetite and weight gain in cancer patients, but these therapeutic effects have been more extensively documented in AIDS patients (reviewed by Kirkham, 2004; Martin and Wiley, 2004; Hall et al., 2005) (see also Table 1). Although concerns have been voiced about the potential immunosuppressive effect of cannabinoids in immunocompromised individuals (Klein et al., 1998), repeated THC administration in a randomized, prospective, controlled trial was found to have few if any consistent effects on various immune functions in AIDS patients receiving antiviral treatment (Bredt et al., 2002).
Anorexia may also be associated with normal aging. A number of hormonal factors have been implicated in the loss of appetite in the elderly, including growth hormone, cholecystokinin, leptin, and various cytokines (Morley, 2001). In a recent study in mice, an age-related decline in food and alcohol intake was accompanied by the loss of ability of the CB1 antagonist SR141716 to reduce food and alcohol intake and a decrease in CB1 receptor-stimulated GTPγS labeling in the limbic forebrain (Wang et al., 2003). These findings suggest that, at least in this animal model, an age-dependent decrease in CB1 receptor signaling in the limbic forebrain may be related to the parallel decline in appetite for both food and alcohol. Anorexia can also accompany debilitating diseases such as Alzheimer’s disease, in which the effectiveness of THC to stimulate appetite has been documented (Volicer et al., 1997). Anorexia nervosa is a psychiatric condition that occurs predominantly in younger women and is characterized by self-starvation, weight loss, and a disturbed body image. Plasma anandamide levels have been reported to increase in patients with restricting anorexia nervosa, which may be secondary to a marked decrease in plasma leptin levels in such patients (Monteleone et al., 2002). Although the relationship between brain and plasma levels of anandamide is not clear, a parallel increase in anandamide in brain regions involved in reward may mediate the rewarding effect of self-starvation in anorexic patients (Monteleone et al., 2005). A recent family-based study examined the possible association of a CB1 receptor gene polymorphism consisting of differences in a trinucleotide repeat with anorexia nervosa. Although no difference was found between parental alleles transmitted or not transmitted to the affected siblings, preferential transmission of different alleles could be established when the patients were subdivided into restricting and binging/purging subgroups (Siegfried et al., 2004).
Endocannabinoids have been also implicated in a unique form of food intake: milk suckling in newborn animals. In an elegant series of studies, Fride et al. (2005) have proposed a role for 2-AG in the brain to stimulate the suckling response in mouse pups. In their model, endogenous 2-AG in the pup’s brain initiated the suckling response via CB1 receptors, with continued suckling depending on milk-derived 2-AG (Fride, 2004). As predicted by this model, treatment of pups with SR141716 inhibits suckling and leads to death due to failure to thrive, a condition analogous to a human condition known as nonorganic failure to thrive, in which an oral motor defect resulting in deficient suckling (Reilly et al., 1999) is similar to the condition in mouse produced by pharmacological blockade or genetic ablation of CB1 (Fride et al., 2005). The relatively high dose of SR141716 to inhibit suckling and its residual effectiveness in CB1 knockout mice suggested the additional involvement of a receptor distinct from CB1 or CB2 (Fride et al., 2003).
B. Pain and Inflammation
One of the earliest uses of cannabis was to treat pain. Historical documents reveal the use of cannabis for surgical anesthesia in ancient China and to relieve pain of diverse origin in ancient Israel, Greece, Rome, and India (reviewed in Mechoulam, 1986; Iversen, 2000; Mechoulam and Hanus, 2000). Numerous early studies have also demonstrated beneficial effects of cannabinoids in animal models of pain (reviewed in Walker and Huang, 2002; Fox and Bevan, 2005). In acute pain, anandamide, THC, cannabidiol, and synthetic cannabinoids such as CP55,940 and WIN 55,212-2 are effective against chemical (Sofia et al., 1973; Formukong et al., 1988; Calignano et al., 1998; Costa et al., 2004a,b; Guindon et al., 2006; Ulugol et al., 2006), mechanical (Sofia et al., 1973; Martin et al., 1996; Smith et al., 1998; Guindon and Beaulieu, 2006), and thermal pain stimuli (Buxbaum, 1972; Bloom et al., 1977; Lichtman and Martin, 1991a,b; Fride and Mechoulam, 1993; Guindon and Beaulieu, 2006). Recent animal studies indicate that anandamide and cannabinoid ligands are also very effective against chronic pain of both neuropathic (Herzberg et al., 1997; Bridges et al., 2001; Fox et al., 2001; Guindon and Beaulieu, 2006) and inflammatory origin (Tsou et al., 1996; Richardson et al., 1998a,b,c; Li et al., 1999; Martin et al., 1999b; Guindon et al., 2006). Moreover, endocannabinoids and synthetic cannabinoids exert synergistic antinociceptive effects when combined with commonly used nonsteroid anti-inflammatory drugs, which may have utility in the pharmacotherapy of pain (Guindon and Beaulieu, 2006; Guindon et al., 2006; Ulugol et al., 2006). Interestingly, a recent study has implicated the endocannabinoid system in the analgesic activity of paracetamol (acetaminophen), the most widely used painkiller (Ottani et al., 2006), and there is also evidence for endocannabinoid involvement in the action of some general anesthetics, such as propofol (Patel et al., 2003; Schelling et al., 2006).
Cannabinoids exert their antinociceptive effects by complex mechanisms involving effects on the central nervous system (Martin et al., 1993; Hohmann et al., 1995, 1998, 1999; Martin et al., 1995, 1996, 1998, 1999a,b; Richardson et al., 1997, 1998a,b; Meng et al., 1998; Strangman et al., 1998; Hohmann and Walker, 1999; Fox et al., 2001), spinal cord (Yaksh, 1981; Lichtman and Martin, 1991a,b; Welch and Stevens, 1992; Richardson et al., 1997, 1998a,b; Hohmann et al., 1998; Chapman, 1999; Drew et al., 2000; Naderi et al., 2005; Suplita et al., 2006), and peripheral sensory nerves (Calignano et al., 1998; Richardson et al., 1998c; Hohmann and Herkenham, 1999; Fox et al., 2001; Johanek et al., 2001; Kelly et al., 2003; Johanek and Simone, 2004; Jordt et al., 2004; Amaya et al., 2006). This is consistent with the anatomical location of CB1 receptors in areas relevant to pain in the brain, spinal dorsal horn, dorsal root ganglia, and peripheral afferent neurons (Herkenham et al., 1990, 1991a; Hohmann and Herkenham, 1998, 1999; Hohmann et al., 1999; SañudoPeña et al., 1999a).
In addition to the role of CB1 receptors, there is recent evidence implicating CB2 receptors in the antihyperalgesic activity of cannabinoids in models of acute and chronic, neuropathic pain, especially of inflammatory origin (Calignano et al., 1998; Hanus et al., 1999; Malan et al., 2001; Clayton et al., 2002; Ibrahim et al., 2003, 2005; Nackley et al., 2003a,b, 2004; Quartilho et al., 2003; Elmes et al., 2004; Hohmann et al., 2004; Scott et al., 2004; Whiteside et al., 2005; Ibrahim et al., 2006). Cannabinoid agonists may also release endogenous opioids, and a functional interplay between the endocannabinoid and opioid systems in modulating analgesic responses has been suggested by numerous studies (Pugh et al., 1997; Manzanares et al., 1999a,b; Houser et al., 2000; Ibrahim et al., 2005; Tham et al., 2005; Vigano et al., 2005a,b; Williams et al., 2006).
As discussed before, anandamide is also a ligand for TRPV1 receptors, albeit with an affinity lower than its affinity for CB1 receptors. The potential involvement of TRPV1 in the analgesic effect of endogenous anandamide has been raised by the findings that the analgesic response to microinjection of a FAAH antagonist into the periaqueductal gray of rats could be inhibited by a similar local microinjection of 6 nmol of capsazepine (Maione et al., 2006). However, others reported that systemic administration of 10 mg/kg capsazepine, which blocked capsaicin-induced analgesia, failed to inhibit endocannabinoid-mediated, stress-induced analgesia, which could be enhanced by a FAAH inhibitor and completely blocked by the CB1 antagonist rimonabant (Suplita et al., 2006).
The analgesic response to exogenous cannabinoids suggested a role for the endocannabinoid system in regulating pain sensitivity, which has received experimental support (reviewed in Walker et al., 2000, 2002; Cravatt et al., 2004; Boger et al., 2005). For example, Walker et al. (1999) have demonstrated increased anandamide levels in some brain areas involved in nociception after peripheral nociceptive input in the rat. The functional role of endogenous anandamide was further supported by the predominantly CB1-mediated analgesic response to FAAH or endocannabinoid transport inhibitors in animal models of acute and chronic pain (Lichtman et al., 2004a; Chang et al., 2006; Jayamanne et al., 2006; La Rana et al., 2006; Suplita et al., 2006). Similarly, FAAH knockout mice had elevated brain levels of anandamide and displayed analgesic behavior in acute inflammatory, but not in chronic neuropathic models of pain (Lichtman et al., 2004b). Formation of anandamide and 2-AG is also increased in response to stress in the periaqueductal gray matter, in which inhibition of endocannabinoid degradation was found to enhance stress-induced analgesia in a CB1 receptor-dependent manner (Hohmann et al., 2005; Suplita et al., 2006), confirming and extending an earlier finding that implicated CB1 receptors and endocannabinoids in stress-induced analgesia (Valverde et al., 2000).
In humans, the analgesic activity of THC and other cannabinoids is less clear-cut. There are numerous case reports on the beneficial effects of cannabis or synthetic derivatives of THC in pain associated with multiple sclerosis, cancer, neuropathies, and HIV infection (Noyes et al., 1975a,b; Martyn et al., 1995; Consroe et al., 1997; Hamann and di Vadi, 1999; Ware et al., 2003; Rudich et al., 2003; Ware and Beaulieu, 2005; Ware et al., 2005; Berlach et al., 2006; reviewed in Burns and Ineck, 2006) (Table 1). The results of randomized studies conducted before 1999 on the analgesic effect of orally administered synthetic cannabinoids in patients with postoperative, post-traumatic, cancer, or spastic pain had been subjected to a meta-analysis. The authors concluded that cannabinoids were not more effective than codeine in controlling pain, and their use was associated with numerous undesirable, dose-limiting CNS side effects (Campbell et al., 2001).
Recent clinical trials with THC or cannabis extracts containing a 1:1 mixture of THC and cannabidiol (Sativex, GW-1000) have provided mixed results. In a randomized, double-blind, placebo-controlled crossover study of 48 patients suffering from central neuropathic pain due to brachial plexus avulsion, oromucosally administered THC or Sativex was ineffective in reducing the pain severity score recorded during the last 7 days of treatment (Berman et al., 2004). Similarly, oral THC (dronabinol) did not improve postoperative (Buggy et al., 2003) and neuropathic pain (Attal et al., 2004) in trials involving small numbers of patients. However, numerous lessons have been learned from these initial human studies on optimal trial design, dose and route of administration of cannabinoids, and more recent larger-scale studies allow reason for more optimism, as outlined below.
THC or Sativex reduced neuropathic pain in patients with traumatic nerve injury or multiple sclerosis in randomized, double-blind, placebo-controlled, crossover trials (Wade et al., 2003; Notcutt et al., 2004). Modest, but clinically relevant analgesic effects were reported in 21 multiple sclerosis patients treated with dronabinol, in a randomized, controlled clinical trial (Svendsen et al., 2004). Effective pain relief by orally administered cannabis extract or THC was also reported in a randomized, controlled, multicenter trial involving 611 multiple sclerosis patients (Zajicek et al., 2003). Moreover, in a recent study of 66 multiple sclerosis patients, Sativex was effective in reducing central neuropathic pain (Rog et al., 2005). A preview of as-yet-unpublished human studies gave an account of a significant benefit of Sativex over placebo in peripheral neuropathic pain characterized by allodynia, in central pain associated with multiple sclerosis, and in opiate-resistant, intractable pain due to cancer (Russo, 2006). A multicenter dose-escalation study of the analgesic and adverse effects of an oral cannabis extract (Cannador) in patients with postoperative pain demonstrated significant dose-related improvements in rescue analgesia requirements and significant trends across the escalating dose groups for decreasing pain intensity (Holdcroft et al. 2006). THC (Marinol) was found to suppress otherwise intractable cholestatic pruritus in a case report (Neff et al., 2002). An analysis of pain questionnaires from 523 patients with HIV infections revealed that 90 to 94% of the subjects using cannabis experienced improvement in muscle and neuropathic pain (Woolridge et al., 2005). The therapeutic potential of cannabinoids in pain associated with trigeminal neuralgia and migraine has also been the subject of several recent reviews (Liang et al., 2004; Russo, 2004, 2006). Preclinical studies (Burstein et al., 1998, 2004; Burstein, 2000, 2005; Dyson et al., 2005; Mitchell et al., 2005; Salim et al., 2005) and a recent clinical trial of 24 patients with neuropathic pain of varying etiologies demonstrated that ajulemic acid, a major metabolite of THC with CB1 agonist activity, was effective in reducing pain without causing cannabinoid-like CNS side effects, the first evidence for the separability of the psychotropic and analgesic effects of a THC analog in humans (Karst et al., 2003). Numerous additional human studies are ongoing to determine the effectiveness of THC or cannabis-based extracts against various forms of pain (reviewed in Ware and Beaulieu, 2005 reviewed in Ware and Beaulieu, 2006).
Multiple lines of evidence support the important role of the cannabinoid signaling system in the modulation of immune function and inflammation (reviewed in Klein et al., 1998, 2003; Walter and Stella, 2004; Klein, 2005). First, cannabinoid receptors are present on immune cells, where their expression is modulated by microbial antigens or other stimuli that induce immune activation. Second, stimulation of immune cells by bacterial toxins such as lipopolysaccharide (LPS) increases the cellular levels of endocannabinoids and their degrading enzyme(s). Third, cannabinoid agonists modulate immune function both in vitro and in vivo via cannabinoid receptor-dependent and -independent mechanisms.
The anti-inflammatory effects of cannabinoids are complex and may involve modulation of cytokine (e.g., TNF-α, IL-12, IL-1, IL-6, and IL-10) and chemokine production (e.g., CCL2, CCL5, CXCL8, and CXCL10), modulation of adenosine signaling (Carrier et al., 2006), expression of adhesion molecules (e.g., ICAM-1, P- intercellular adhesion molecule-1 and P-selectin), and the migration, proliferation, and apoptosis of inflammatory cells (reviewed in Klein et al., 1998, 2003; Walter and Stella, 2004; Klein, 2005). To the extent that pain and inflammation accompany many of the disorders discussed in the rest of this review, cannabinoids would be expected to provide significant benefit due to their analgesic and anti-inflammatory properties.
C. Central Nervous System Disorders
The emerging role of the endocannabinoid system in a variety of CNS disorders should not come as a surprise given the very high level of expression of CB1 receptors in the brain. The particularly high density of CB1 receptors in the cortex, cerebellum, hippocampus, and basal ganglia had drawn early attention to diseases affecting movement, mood and anxiety disorders, and conditions related to altered brain reward mechanisms, as well as processes of memory and learning. The classic behavioral effects of marijuana also provided early clues about potential therapeutic targets, such as the control of pain or appetite. The role of the endocannabinoid system in the pathogenesis and treatment of specific CNS diseases is discussed below.
1. Neurotoxicity and Neurotrauma
The endocannabinoid system plays an important role in neuroprotection both in acute neuronal injury (e.g., traumatic brain injury, stroke, and epilepsy) and in chronic neurodegenerative disorders, such as multiple sclerosis, Parkinson’s disease, Huntington’s disease, amyotrophic lateral sclerosis, and Alzheimer’s disease (reviewed in Glass, 2001; Mechoulam et al., 2002a,b; Grundy, 2002; Croxford, 2003; Drysdale and Platt, 2003; Jackson et al., 2005a; Ramos et al., 2005). Although the underlying mechanisms are not fully understood, multiple cannabinoid receptor-dependent as well as receptor-independent processes have been implicated. These include, but are not limited to 1) modulation of excitatory glutamatergic transmissions and synaptic plasticity via presynaptic CB1 receptors (Molina-Holgado et al., 1997; Marsicano and Lutz, 1999; Gerdeman et al., 2002; reviewed in Alger, 2002; Robbe et al., 2002; Azad et al., 2003; Freund et al., 2003; Gerdeman and Lovinger, 2003; Piomelli, 2003; Mato et al., 2004), 2) modulation of immune responses and the release of inflammatory mediators by CB1, CB2, and non CB1/CB2 receptors on neurons, astrocytes, microglia, macrophages, neutrophils and lymphocytes (Watzl et al., 1991; Zheng et al., 1992; Fischer-Stenger et al., 1993; Cabral and Fischer-Stenger, 1994; Kusher et al., 1994; Burnette-Curley and Cabral, 1995; Cabral et al., 1995; reviewed in Friedman et al., 1995; Zheng and Specter, 1996; Shohami et al., 1997; Newton et al., 1998; Srivastava et al., 1998; Gallily et al., 2000; Klein et al., 2000a,b, 2003; Smith et al., 2000; Carlisle et al., 2002; Germain et al., 2002; Killestein et al., 2003; Kaplan et al., 2005; Ramirez et al., 2005; reviewed in Friedman et al., 1995; Stella, 2004; Walter and Stella, 2004; Correa et al., 2005; Croxford and Yamamura, 2005; Klein, 2005;), 3) activation of cytoprotective signaling pathways (Grigorenko et al., 2002), such as protein kinase B/Akt (Molina-Holgado et al., 2002), protein kinase A (Kim et al., 2005), or neurotrophic factors (Khaspekov et al., 2004), 4) modulation of excitability and calcium homeostasis via effects on Ca2+, K+, and Na+ channels, N-methyl D-aspartate (NMDA) receptors, gap junctions, and intracellular Ca2+ stores (Caulfield and Brown, 1992; Mackie and Hille, 1992; Mackie et al., 1993; Nadler et al., 1995; Venance et al., 1995; Shohami et al., 1997; Hampson et al., 2000b; Oz et al., 2000, 2004; Chemin et al., 2001; Maingret et al., 2001; Nogueron et al., 2001; Robbe et al., 2001; Wilson and Nicoll, 2001; Wilson et al., 2001; Nicholson et al., 2003; Guo and Ikeda, 2004; del Carmen et al., 2005; del Carmen Godino et al., 2005; Zhuang et al., 2005), 5) antioxidant properties of cannabinoids (Eshhar et al., 1995; Hampson et al., 1998; Chen and Buck, 2000; reviewed in Hampson et al., 2000a; Marsicano et al., 2002a), and 6) CB1 receptor-mediated hypothermia, possibly by reducing metabolic rate and oxygen demand (Leker et al., 2003).
Excitotoxicity, the toxic effects of an overactivation of glutamate receptors, and the resulting oxidative stress may contribute to the pathological processes eventually leading to cellular dysfunction or death in both acute and chronic forms of neurodegeneration (Coyle and Puttfarcken, 1993; McNamara, 1999; Lutz, 2004). Dexanabinol (HU-211), a behaviorally inactive cannabinoid and noncompetitive antagonist of NMDA receptors, protects primary rat neuronal cultures against NMDA and glutamate exposure in vitro (Eshhar et al., 1993; Nadler et al., 1993a,b). THC protects primary cultured neurons against kainate-mediated toxicity in a CB1-dependent manner (Abood et al., 2001), similar to protectin by WIN 55,212-2 against low extracellular magnesium-induced cell death (Shen and Thayer, 1998). Palmitoylethanolamide also improves neuronal survival in a glutamate-induced cell death model (Skaper et al., 1996). Intracerebral injection of NMDA in neonatal rats results in a 13-fold increase of cortical anandamide concentrations (Hansen et al., 2001a,b). Both THC and anandamide exerted CB1-mediated neuroprotective effects in an ouabain-induced rat model of in vivo excitotoxicity (van de Stelt et al., 2001a,b). Anandamide and synthetic agonists of CB1 receptors also protected the newborn brain against AMPA-kainate receptor-mediated excitotoxic damage in mice (Shouman et al., 2006).
Traumatic brain injury (TBI) is one of the leading causes of disability and mortality in young individuals (Holm et al., 2005), yet the available therapy is very limited (Faden, 2002; Maas et al., 2004). TBI is characterized by cerebral edema, axonal and neuronal injury, increased permeability of the blood-brain barrier, and post-traumatic changes in cognitive and neurological functions (Bayir et al., 2003). TBI can trigger glutamate-induced excitotoxicity, oxidative stress, release of inflammatory cytokines from brain-resident cells (microglia, neurons, and astrocytes), programmed cell death, and cortical blood flow dysregulation (reviewed in Wang and Feuerstein, 2000; Gentleman et al., 2004).
The protective effect of cannabinoids in traumatic brain injury was first indicated in studies with the nonpsychotropic cannabinoid dexanabinol (HU-211) (Fig. 1b). These studies have demonstrated reduced brain damage and improved motor and cognitive function in HU-211-treated animals in a rat model of TBI. The favorable effects of a single injection of HU-211 on learning and neurological deficits lasted up to 30 days and could be achieved within a therapeutic window of 6 h (Shohami et al., 1993, 1995). Beneficial effects of HU-211 were also demonstrated in an axonal injury model (Yoles et al., 1996; Zalish and Lavie, 2003). These protective effects were attributed, at least in part, to NMDA receptor blockade, attenuation of Ca2+ influx and decreased TNF-α levels (Nadler et al., 1995; Shohami et al., 1997; reviewed in Mechoulam et al., 2002a,b; Biegon, 2004). In mice with closed head injury, brain levels of 2-AG increased, and exogenous 2-AG administered 1 h after the head injury reduced infarct size and improved neurological outcome (Panikashvili et al., 2001). Neuroprotection by 2-AG was attributed to CB1 receptor-mediated inhibition of nuclear factor-κB and of the early expression of proinflammatory cytokines TNF-α, IL-1β, and IL-6 (Panikashvili et al., 2005, 2006). In a rat model of TBI, BAY 38-7271, a CB1/CB2 agonist with predominant action at CB1 receptors, caused a marked, 70% reduction in infarct volume when administered as a 4-h infusion immediately after induction of subdural hematoma, and even when it was applied with a 3-h delay, infarct volume was reduced by 59% (Mauler et al., 2002).
A multicenter, double-blind, randomized, placebo-controlled phase II trial conducted in 67 patients with severe closed head injury found dexanabinol to be safe and well tolerated. The treated patients achieved significantly better intracranial pressure/cerebral perfusion pressure control without jeopardizing blood pressure, and a trend toward faster improvement and better neurological outcome was also observed (Knoller et al., 2002). However, a double-blind, randomized, placebo-controlled phase III clinical trial of dexanabinol, conducted in 15 countries in 86 specialized centers and involving 861 patients failed to demonstrate any favorable effects (Maas et al., 2006).
2. Stroke
Ischemic stroke is the most common form of stroke, mostly caused by a transient interruption of blood supply to the brain by thrombotic occlusion of blood vessels. It is an important cause of death and disability in industrialized countries, affecting up to 0.2% of the population each year (Klijn and Hankey, 2003; Pinto et al., 2004). One in six patients die in the 1st month after ischemic stroke, and half of the survivors are permanently disabled despite the best efforts to rehabilitate them and to prevent complications (Klijn and Hankey, 2003).
One of the first indications of the neuroprotective effect of cannabinoids came from the field of stroke research, using various in vitro and in vivo models of ischemic injury. Anandamide, 2-AG, and WIN 55,212-2 protected cultured cortical neurons against hypoxia and glucose deprivation (Nagayama et al., 1999; Sinor et al., 2000). The effects of various cannabinoid ligands were also investigated in in vivo models of global cerebral ischemia induced by two-vessel occlusion with hypotension or by four-vessel occlusion, or in focal ischemia induced by occlusion of the middle cerebral artery (MCAo), with or without reperfusion. Dexanabinol at doses of 2 to 10 mg/kg decreased infarct size and histological damage and improved neurological score in rat and gerbil models of both global and focal cerebral ischemia (Bar-Joseph et al., 1994; Vered et al., 1994 a,b; Belayev et al., 1995a,b,c; Leker et al., 1999; Lavie et al., 2001; Teichner et al., 2003). Importantly, this protective effect was observed even when the drug was administered 60 to 180 min after the insult (Vered et al., 1994; Belayev et al., 1995a,b,c; Leker et al., 1999; Lavie et al., 2001; Teichner et al., 2003).
WIN 55,212-2, at doses of 0.03 and 1 mg/kg but not 3 mg/kg decreased hippocampal neuronal loss after transient global cerebral ischemia in rats. It also reduced infarct size after permanent focal cerebral ischemia induced by MCAo, when given 40 min before 30 min after the occlusion, and these effects were prevented by SR141716 (Nagayama et al., 1999). WIN 55,212-2 also protected cultured cerebral cortical neurons from in vitro hypoxia and glucose deprivation, but in contrast to the receptor-mediated neuroprotection observed in vivo, this in vitro effect was not stereoselective and was insensitive to CB1 and CB2 receptor antagonists (Nagayama et al., 1999). BAY38-7271 also decreased infarct size in rats with permanent MCAo even when given intravenously 4 h after the occlusion (Mauler et al., 2002). Similarly, HU-210 reduced infarct size by up to 77% and improved motor disability in a rat model of permanent MCAo (Leker et al., 2003). The protective effect of HU-210 was partially reversed by pretreatment with SR141716, indicating CB1 receptor involvement. Surprisingly, all protection could be abolished by warming the animals to the body temperature of controls, indicating that CB1-mediated hypothermia contributed to the neuroprotection (Leker et al., 2003). Likewise, CB1-mediated hypothermia was responsible for the neuroprotective effects of THC in a mouse transient MCAo model (Hayakawa et al., 2004) and perhaps also in a rat model of global cerebral ischemia (Louw et al., 2000). Consistent with these findings, CB1 knockout mice had increased mortality from permanent focal cerebral ischemia, increased infarct size, more severe neurological deficits after transient focal cerebral ischemia, and decreased cerebral blood flow in the ischemic penumbra during reperfusion, compared with wild type controls subjected to the same insult (Parmentier-Batteur et al., 2002). NMDA neurotoxicity was also increased in CB1−/− mice compared with wild-type littermates (Parmentier-Batteur et al., 2002). Further evidence for a role of CB1 receptors is their increased expression on neurons in the arterial boundary zone of cortical infarction (Jin et al., 2000). Finally, brain levels of endocannabinoids are increased during ischemic (Schmid et al., 1995; Schabitz et al., 2002; Berger et al., 2004; Muthian et al., 2004) and other types of brain injury (Sugiura et al., 2000; Hansen et al., 2001a,b; Panikashvili et al., 2001).
Other studies do not support the neuroprotective role of CB1 receptor activation. For example, CB1 antagonists by themselves had no effect on the outcome of injury, and in two recent reports, SR141716 and LY320135 were found to actually reduce infarct size and to improve neurological function in a rat model of MCAo (Berger et al., 2004; Muthian et al., 2004), whereas low doses of WIN 55,212-2 had no protective effect (Muthian et al., 2004). Thus, it appears that both CB1 agonists and antagonists can be neuroprotective in cerebral ischemia. The reason for the opposite effects of pharmacological blockade versus genetic knockout of CB1 receptors is not clear and may be related to the CB1 receptor-independent effects of antagonists (Begg et al., 2005; Pertwee, 2005b,c). Clearly, evaluating the potential usefulness of cannabinoid ligands in the treatment of stroke warrants future studies.
3. Multiple Sclerosis and Spinal Cord Injury
Multiple sclerosis (MS) is a complex, immune-mediated, inflammatory disease of the white matter of the brain, which compromises impulse conduction due to the loss of the myelin sheath of neurons and the secondary axonal loss (Sospedra and Martin, 2005). MS affects 2 to 5 million people worldwide and commonly presents with an unpredictable, relapsing-remitting course and a range of clinical symptoms depending on where the demyelination and axonal loss have occurred (Compston and Coles, 2002). Some patients become disabled within a short period of time, whereas others can live their entire lives with only negligible or no disability. The symptoms of MS typically involve tremor, ataxia, visual loss, double vision, weakness or paralysis, difficulty in speaking, loss of bladder control and constipation, cognitive impairment, and painful muscle spasms. Muscle spasticity often leads to reduced mobility, considerable distress from pain, and interference with daily living activities. Spasticity, neuropathic and nociceptive pain, and some of the above symptoms are also common in spinal cord injury (SCI). Although there are numerous drugs available that target the immune system to slow down the progression of the disease, they are only moderately effective, and the treatment of MS remains mostly symptomatic and far from satisfactory (Killestein and Polman, 2005).
Cannabis had been used in ancient Greece, Rome, China, and India for relieving muscle cramps, spasm, and pain (reviewed in Mechoulam, 1986, Mechoulam et al., 1998; Mechoulam and Hanus, 2000) and its therapeutic application in MS is a topic of recent lively debate (Grundy, 2002; Pertwee, 2002; Baker and Pryce, 2003; Croxford, 2003; Killestein et al., 2004; Sirven and Berg, 2004; Jackson et al., 2005a; Pryce and Baker, 2005; Robson, 2005; Smith, 2005). Lyman et al. (1989) examined the effects of parenteral THC in experimental autoimmune encephalomyelitis (EAE) in rats, a laboratory model of MS. THC treatment not only reduced CNS inflammation and improved neurological outcome but also improved survival compared with placebo. Δ8-THC, a less psychotropic and more stable analog of THC, also reduced the severity and incidence of neurological deficits in rats with EAE (Wirguin et al., 1994). The nonpsychotropic dexanabinol also suppressed inflammatory responses in the brain and spinal cord of rats with EAE and improved their neurological symptoms (Achiron et al., 2000). Although CB1 receptor density is decreased in the striatum and cortex of EAE rats, this is compensated for by increased coupling to G protein-mediated signaling, ensuring the effectiveness of treatment with cannabinoid agonists (Berrendero et al., 2001).
In a mouse model of chronic relapsing EAE, intravenous administration of THC, WIN 55,212-2, JWH-133, or methanandamide reduced spasticity and tremor, whereas the same symptoms were exacerbated by treatment with either CB1 or CB2 antagonists (Baker et al., 2000). These mice with EAE had increased levels of anandamide, 2-AG, and palmitoylethanolamide (PEA) in areas associated with nerve damage (Baker et al., 2001). Furthermore, spasticity could be relieved not only by administration of exogenous anandamide, 2-AG, or PEA but also by selective inhibitors of endocannabinoid transport or hydrolysis, which suggests tonic control of muscle tone by the endocannabinoid system in EAE (Baker et al., 2001; Ligresti et al., 2006a). Additional evidence for this has emerged through the use of CB1-deficient mice, which tolerated inflammatory and excitotoxic insults poorly and developed substantial neurodegeneration after the induction of EAE (Pryce et al., 2003). Jackson et al. (2005b) reported that the absence of CB1 receptors was associated with increased caspase activation and a greater loss of myelin and axonal/neuronal proteins after the induction of chronic EAE. Interestingly, CB1 knockout mice had increased caspase 3 levels before the induction of EAE, suggesting a neuroprotective tone mediated by CB1 receptors (Jackson et al., 2005a,b). In mice with EAE, WIN 55,212-2 inhibited leukocyte/endothelial interactions via activation of CB2 receptors (Ni et al., 2004). Interestingly, a recent study suggests that the high levels of IFN-γ present in the CNS of mice with EAE can counteract endocannabinoid-mediated neuroprotection by disrupting P2X7 purinergic receptor signaling, a key step in endocannabinoid production by glia (Witting et al., 2006).
Another murine model of MS is Theiler’s murine encephalomyelitis virus-induced demyelinating disease. In mice with Theiler’s murine encephalomyelitis virus-induced demyelinating disease, treatment with WIN 55,212-2 slowed the progression of symptoms, down-regulated delayed-type hypersensitivity reactions and interferon-γ production, and inhibited the expression of proinflammatory cytokines in the CNS (Croxford and Miller, 2003). In another study using this model, treatment with WIN 55,212-2, ACEA, or JWH-015 caused long-lasting improvements in neurological deficits in the established disease and reduced microglial activation, abrogated major histocompatibility complex class II antigen expression, and decreased the number of CD4+ infiltrating T cells in the spinal cord. These changes were paralleled by extensive remyelination (Arevalo-Martin et al., 2003). Treatment of Theiler’s murine encephalomyelitis virus-infected mice with the transport inhibitors OMDM1 and OMDM2 enhanced anandamide levels, down-regulated inflammatory responses in the spinal cord, and ameliorated motor symptoms (Mestre et al., 2005), and similar findings were reported using the transport inhibitor UCM707 (Ortega-Gutierrez et al., 2005). In these two studies, the treatments were also shown to reduce the surface expression of major histo-compatibility complex class II molecules, the production of the proinflammatory cytokines (TNFα, IL-1β, and IL-6), and the expression of inducible NO synthase.
Consistent with the animal data, cannabinoids have shown promise in the treatment of MS in humans (Table 1). A possible underlying mechanism is suggested by a recent study in which the endocannabinoid system was found to be highly activated during CNS inflammation in MS patients and to protect neurons from inflammatory damage by activating a negative feedback loop in microglial cells via CB1/2-mediated epigenetic regulation of mitogen-activated protein kinase phosphatase 1 expression (Eljaschewitsch et al., 2006).
There have been anecdotal reports of the effectiveness of marijuana smoking in relieving symptoms of MS and SPI (Grinspoon and Bakalar, 1993, 1998), which were supported by the results of early open or single-blind observations with orally given THC or smoked marijuana, involving small numbers of patients (Dunn and Davis, 1974; Petro, 1980; Petro and Ellenberger, 1981; Clifford, 1983; Meinck et al., 1989; Brenneisen et al., 1996; Schon et al., 1999). The most consistent finding was a subjective improvement in spasticity, although benefits for mobility, tremor, nystagmus, mood, and bladder control were also reported. In a double-blind crossover study of a single MS patient, nabilone treatment improved muscle spasms, nocturia, and general well-being (Martyn, 1995). In contrast, Greenberg et al. (1994) reported impairments of both balance and posture after a single dose of smoked cannabis in a placebo-controlled study of 10 MS patients and 10 normal subjects. In an anonymous survey of 112 MS patients who self-medicated with cannabis, 30 to 97% of the subjects reported relief from a wide variety of symptoms by smoking marijuana (Consroe et al., 1997). These encouraging reports have triggered numerous larger, population-based clinical trials of cannabis-based medicines in MS, which have yielded mixed results.
Using a randomized, double-blind, placebo-controlled, crossover design, Killestein et al. (2002) have evaluated the effects of oral THC, two doses of 2.5 to 5 mg/day or a Cannabis sativa plant extract administered over a 4-week period, in 16 MS patients with severe spasticity. Spasticity and disability, quantified using the objective Ashworth scale (Ashworth, 1964) and the Expanded Disability Status Scale were not improved. However, a significant improvement in the subjective rating of spasm frequency and trends toward improved mobility were noted, with no effect on tremor, sleep quality, or lower urinary tract symptoms. Both THC and the plant extract worsened the patients’ global impression of their conditions, with plant extracts causing more adverse side effects. It should be mentioned, however, that the dose of THC used was lower than that in subsequent studies with more positive outcome, and as was noted in an accompanying editorial (Thompson and Baker, 2002), the study was not powered to detect efficacy.
A large multicenter study involving 33 clinical centers and 660 MS patients in the United Kingdom and United States and supported by the UK Medical Council aimed to explore the effects of cannabis extract (Cannador) or synthetic THC (Marinol) versus placebo on spasticity, pain, tremor, bladder function, and cognitive function [Cannabinoids in Multiple Sclerosis (CAMS) study; Zajicek et al., 2003, 2004]. There was no change in Ashworth score, tremor, irritability, depression, or tiredness after 15 weeks of treatment with Marinol or Cannador. However, there were significant improvements in patient-reported spasticity, pain, and sleep quality. Unexpectedly, there was also a reduction in hospital admissions for relapse in the two active treatment groups. Adverse side effects were generally minor and similar to those with placebo. Remarkably, in the 12-month follow-up of the original CAMS study of 657 patients, muscle spasticity measured by the Ashworth scale was significantly improved in the THC-treated group. The Rivermead Mobility Index was also improved, indicative of reduced disability. The effect of Cannador on tremor was also studied in a randomized, double-blind, placebo-controlled, crossover trial in 14 patients with MS. Consistent with an earlier report (Zajicek et al., 2003), no significant therapeutic effects were noted (Fox et al., 2004). In another study of similar design, administration of oral capsules containing 2.5 mg of Δ9-THC plus 0.9 mg of CBD (maximal dose of 30 mg of Δ9-THC/day) caused improvements in spasm frequency and mobility in 37 MS patients who received at least 90% of their prescribed dose (Vaney et al., 2004).
In a double-blind, placebo-controlled study involving 18 patients with MS, THC and CBD decreased self-reported spasticity and pain and improved bladder symptoms, whereas spasticity measured by the Ashworth scale was not significantly improved (Wade et al., 2003). The therapeutic effect of Sativex delivered by oromucosal spray (2.7 mg of THC and 2.5 mg of CBD at each actuation) was evaluated in 160 outpatients with MS (Smith, 2004). Patients were allowed to self-titrate the dose to achieve optimal effects, up to a maximal daily dose of 120 mg of THC and CBD. Efficacy was assessed by using a modified Ashworth scale to assess spasticity, whereas daily living, mobility, cognitive function, and tremor were quantified through the use of various scales and questionnaires (Wade et al., 2004). There was no significant difference in the Ashworth scale, tremor, and pain at 6 weeks between the Sativex and placebo groups. However, visual analog scales showed significant improvement in patients whose primary symptom had been spasticity (Wade et al., 2004). Sativex was well tolerated and effective against central neuropathic pain and sleep disturbances associated with MS in a randomized, controlled trial involving 66 patients (Rog et al., 2005). Sativex was approved and launched in Canada in 2005 for the treatment of neuropathic pain associated with MS and is currently being investigated for various other therapeutic indications (Russo, 2004, 2006).
In a recent case report, a 46-year-old woman was diagnosed with MS after having entered treatment with the CB1 receptor antagonist rimonabant for obesity, and recovery to near normal was noted within weeks after discontinuation of the treatment (van Oosten et al., 2004). This report, coupled with the more severe neurodegenerative process when MS is induced in CB1 knock-out mice or in mice treated with a CB1 receptor antagonist, could suggest that CB1 antagonism may exacerbate inflammatory demyelinating diseases in humans (van Oosten et al., 2004). However, the occurrence of MS in this one patient may have been purely coincidental.
Although the results of the above clinical studies (Table 1) are somewhat equivocal, patients treated with cannabis experienced improvements in the most disturbing symptoms including pain and spasticity compared with those receiving placebo, without experiencing significant side effects. These studies also suggest that the Ashworth scale as a primary measure of spasticity in MS does not accurately assess the complex collection of symptoms associated with spasticity, which may be more accurately evaluated using subjective measures. Indeed, the use of the Ashworth scale as a primary measure of spasticity in MS has often being criticized, and many commonly used antispasticity medications have also failed to generate statistically significant improvements according to this scale (Hinderer and Gupta, 1996; Shakespeare et al., 2003). Accurate assessment of the clinical effectiveness of cannabinoids in MS may be complicated by the difficulty in achieving the most appropriate individual oral dose (Table 1). Peak plasma concentrations and their timing vary greatly because of the low water solubility of cannabis components and the large variability in their absorption from the gastrointestinal tract. An additional disadvantage of oral administration is the hepatic first-pass effect. This can result in the formation of large quantities of the psychoactive metabolite 11-OH-THC, which may be responsible for some of the side effects observed (Table 1). Delivery of cannabis-based extracts as an oromucosal spray may minimize these drawbacks and may allow patients to better optimize their individual daily dose by self-titration (Russo, 2006).
In conclusion, controlled clinical trials with cannabinoids have demonstrated their efficacy in eliciting symptomatic improvements in MS patients. These results suggest that there is place for the use of cannabis in the treatment of MS, which should be confirmed in further larger-scale clinical trials.
4. Movement Disorders (Basal Ganglia Disorders)
Endocannabinoid involvement in the central regulation of motor functions and in movement disorders is based on multiple lines of evidence. First, CB1 receptors are highly expressed in the basal ganglia, especially in the substantia nigra and in the cerebellum (Herkenham et al., 1990, 1991a,b; Mailleux and Vanderhaeghen, 1992; Tsou et al., 1998; Hohmann and Herkenham, 2000; Moldrich and Wenger, 2000; Howlett et al., 2002), areas involved in motor control. Second, endocannabinoids are also abundant in these brain regions (Bisogno et al., 1999a; Di Marzo et al., 2000). Third, endogenous, plant-derived, and synthetic cannabinoids have potent, mostly inhibitory, effects on motor activity (Crawley et al., 1993; Fride and Mechoulam, 1993; Wickens and Pertwee, 1993; Smith et al., 1994; Romero et al., 1995a,b, 2002b; reviewed in Sañudo-Peñ a et al., 1999b). Fourth, CB1 receptor and endocannabinoid levels are altered in the basal ganglia both in experimental models (Zeng et al., 1999; Page et al., 2000; Romero et al., 2000; Lastres-Becker et al., 2001a,b, 2002a,b; Gonzalez et al., 2006) and human forms of movement disorders (Glass et al., 1993, 2000, 2004; Lastres-Becker et al., 2001a; reviewed in Romero et al., 2002b). Fifth, the endocannabinoid system interacts with several neurotransmitter pathways at various levels of the basal ganglia circuitry (Glass et al., 1997a; Miller et al., 1998; Sañudo-Peñ a and Walker, 1998; Giuffrida et al., 1999; Rodriguez De Fonseca et al., 2001; Brotchie, 2003; van der Stelt and Di Marzo, 2003; de Lago et al., 2004a).
a. Parkinson’s disease and levodopa-induced dyskinesia
Parkinson’s disease (PD) is the second most common neurodegenerative disease of adult onset, with incidence of 16 to 19/100,000 people worldwide (Twelves et al., 2003). PD is caused by a severe loss of dopaminergic neurons in the substantia nigra pars reticulata (SNr), resulting in reduced dopamine levels and a loss of dopaminergic neurotransmission in the striatum, which interferes with motor function and coordination. Although excitotoxicity, oxidative stress, inflammation, mitochondrial dysfunction, and environmental and hereditary factors have all been implicated in the pathogenesis of PD, the exact cause of the loss of dopaminergic neurons remains elusive (Hattori and Mizuno, 2004; Eriksen et al., 2005). Clinically, PD is characterized by the classic triad of resting tremor, muscular rigidity, and bradykinesia/akinesia (slowness of movement or postural immobility). Current therapies include oral dopamine replacement via the dopamine precursor levodopa, anti-cholinergic agents, and monoamine oxidase B inhibitors (Horn and Stern, 2004). Although dopamine replacement therapy can be effective in most patients by controlling the symptoms in the short term, their long-term use is associated with diminishing efficacy and severe side effects such as levodopa-induced dyskinesia (LID) (involuntary movements), which often lead to treatment discontinuation and severe disability.
In PD, there are secondary abnormalities in nondopaminergic transmission within the basal ganglia that are thought to contribute to the inhibition of motor function. Inhibitory GABAergic transmission from the striatum to the external region of the globus pallidus (GPe) is increased, making the GPe hypoactive. This results in decreased GABAergic input from the GPe to the subthalamic nucleus which, together with increased activity of glutamatergic efferents to this nucleus, results in its hyperactivity. In turn, the hyperactive subthalamic nucleus increases the activity of the SNr and internal globus pallidus (GPi) through glutamatergic efferents. Because both the SNr and GPi provide inhibitory output to motor nuclei outside the basal ganglia (e.g., motor thalamus and brain stem locomotor regions), this mechanism is thought to contribute to the excessive motor inhibition in PD (Obeso et al., 2000; Bezard et al., 2001). In general, changes opposite to those described above are likely to be involved in LID. The final outcomes of the dysregulation of neuronal circuits are abnormal patterning, firing rate, and synchronization of basal ganglia outputs (Obeso et al., 2000; Bezard et al., 2001). Importantly, nondopaminergic mechanisms may counterbalance the loss of dopamine and are probably responsible for the lack of parkinsonian symptoms until the loss of >80% of striatal dopamine. They may also attenuate the severity of symptoms once symptoms develop. As discussed below, the endocannabinoid system may play an important regulatory role in PD,PD and LID as well as in the compensatory mechanisms.
Overactivity of endocannabinoid transmission, as reflected by increased tissue levels of endocannabinoids and CB1 receptors as well as decreased rates of endocannabinoid transport and degradation by FAAH, have been found in the basal ganglia in the 6-hydroxydopamine-lesioned or reserpine-treated rat models of PD (Mailleux and Vanderhaeghen, 1993; Romero et al., 2000; Gubellini et al., 2002; Centonze et al., 2005; Fernandez-Espejo et al., 2005; Gonzalez et al., 2006). In basal ganglia from 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-lesioned marmosets, a primate model of PD, and in basal ganglia of PD patients, the density of striatal CB1 receptors and CB1 receptor-G-protein coupling were found to be increased (Lastres-Becker et al., 2001a). The above changes in 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-treated marmosets and 6-hydroxydopamine-lesioned rats were reversible by chronic L-dopa treatment, which indicates that the similar changes observed in PD patients were unlikely to have been induced by the replacement therapy (Lastres-Becker et al., 2001a; Maccarrone et al., 2003). There is broad agreement that the endocannabinoid system becomes overactive in the basal ganglia in PD (reviewed in Brotchie, 2003), although some studies report a reduction (Silverdale et al., 2001) or no change in CB1 receptor expression (Herkenham et al., 1991a) or a dependence on L-DOPA cotreatment of the increased CB1 receptor expression in the basal ganglia of animals with experimental PD (Zeng et al., 1999).
If the enhanced CB1 receptor signaling in the striatum is viewed as an attempt of the dopamine-deficient brain to normalize striatal function, the pharmacological amplification of this signaling might alleviate symptoms of PD, e.g., by reducing striatal glutamate release (Gerdeman and Lovinger, 2001; Gerdeman et al., 2002; Gubellini et al., 2002). On the other hand, enhanced CB1 receptor signaling, if focused on the other part of the circuitry (e.g., GPe), can enhance GABA transmission, leading to inhibition of GPe and thereby contributing to the symptoms of PD. Likewise, CB1 antagonism could have either pro-parkinsonian effects, if it targets the striatum, or antiparkinsonian effects, if it targets the GPe. Accordingly, both agonists and antagonists might have therapeutic potential, both in PD and LID (reviewed in Brotchie, 2003).
Treatment with CB1 receptor agonists can decrease the tremor associated with overactivity of the subthalamic nucleus (Sañudo-Peñ a et al., 1998, 1999b), improve motor impairment seen with dopaminergic agonists (Anderson et al., 1995; Maneuf et al., 1997; Sañudo-Peñ a et al., 1998), protect against dopaminergic cell death (Lastres-Becker et al., 2005), and delay or reduce the incidence of LID (Sieradzan et al., 2001; Fox et al., 2002a; Ferrer et al., 2003; Gilgun-Sherkiet et al., 2003). However, cannabinoid agonists are unlikely to be used for reducing bradykinesia in PD because of their hypokinetic profile both in primates and humans (Consroe, 1998; Müller-Vahl et al., 1999a; Romero et al., 2002; Brotchie, 2003; Croxford, 2003; Croxford and Miller, 2003).
On the other hand, dysfunction of nigrostriatal dopaminergic neurons can be associated with overactivity of endocannabinoid transmission in the basal ganglia (see above). CB1 receptor antagonists may therefore be useful for alleviating the bradykinesia of PD or LID, because they attenuate CB1 signaling in GPe or GPi. (Mailleux and Vanderhaeghen, 1993; Di Marzo et al., 2000; Lastres-Becker et al., 2001a,b; Gubellini et al., 2002; reviewed in Brotchie, 2003; Fernandez-Espejo et al., reviewed in Brotchie, 2003; 2005; Fernandez-Ruiz and Gonzalez, 2005). Notwithstanding the above, studies using SR141716 in rat (Di Marzo et al., 2000) and primate models of PD or LID (Meschler et al., 2001; van der Stelt et al., 2005) provided conflicting results. Rimonabant treatment also failed to influence dyskinesia in the first small-scale, randomized, double-blind, placebo-controlled human study (Mesnage et al., 2004). However, the dose used in this human study was approximately 10-fold lower (0.3 mg/kg versus. 3 mg/kg), than in a recent primate study with positive outcome (van der Stelt et al., 2005). As suggested by a recent report (Fernandez-Espejo et al., 2005), it is also possible that CB1 receptor blockade is effective only at the very advanced stages of the disease. More recently, using Park-2 knockout mice, a genetic model of early PD, Gonzalez et al. (2005) observed gender-dependent differences for both the levels of CB1 receptors and motor responses to agonists or antagonists, extending earlier data obtained in humans and in animal models of PD.
Taken together, although the above studies do not offer a complete understanding of the role of endocannabinoids and cannabinoid receptors in PD and LID, they support the notion that the endocannabinoid system plays an important role in movement disorders, including PD, and may provide the framework for novel therapeutic approaches in the future.
b. Huntington’s disease
Huntington’s disease (HD) is an inherited, autosomal dominant, progressive neuropsychiatric disorder of the midlife, caused by an unstable expansion of a trinucleotide polyglutamine repeat in the N-terminal domain of a protein termed huntingtin, which leads to degeneration of neurons in the basal ganglia and cortical regions. The disease is characterized by motor disturbances, such as chorea (involuntary movements) and dystonia, psychiatric symptoms, and dementia (Melone et al., 2005). The prevalence of HD is similar to that of ALS (see below), but much lower than that of most of the other neurodegenerative illnesses discussed above or below. The therapy of HD is very limited and includes antidopaminergic drugs to reduce the hyperkinesias and antiglutamatergic agents to reduce excitotoxicity (Melone et al., 2005).
It has been clearly demonstrated, both in postmortem human tissue (Glass et al., 1993, 2000; Richfield and Herkenham, 1994) and in chemically induced and transgenic animal models (Denovan-Wright and Robertson, 2000; Page et al., 2000; Lastres-Becker et al., 2001, 2002a,b; Sieradzan and Mann, 2001; Behrens et al., 2002; Glass et al., 2004; McCaw et al., 2004) that a decrease in CB1 receptor level and signaling activity in the basal ganglia is one of the earliest changes in HD, preceding nerve loss and clinical symptoms. Furthermore, decreased levels of anandamide and 2-AG in the striatum and an increase of anandamide in the ventral mesencephalon, where the substantia nigra is located, have been documented in a rat model of HD (Lastres-Becker et al., 2001). Thus, it appears that endocannabinoid signaling in the basal ganglia is hypofunctional in HD, which probably contributes to the hyperkinesia associated with the disease. These studies also suggest that the endocannabinoid system is involved in the pathogenesis and/or progression of HD, and cannabinoid agonists could be of significant therapeutic benefit in HD because of their anthyperkinetic and neuroprotective effects (reviewed in Lastres-Becker et al., 2003b). A recent study identified a novel population of progenitor cells expressing CB1 receptors in the subependymal layer of the normal and Huntington’s diseased human brain. This finding raises the intriguing possibility that these cells could be a source of replacement of cells lost due to neurodegenerative disease (Curtis et al., 2006).
Indeed, data from animal models demonstrated that both CB1 agonists and inhibitors of endocannabinoid transport are able to reduce hyperkinesia (Lastres-Becker et al., 2002b, 2003a). Interestingly, direct agonists of CB1 receptors, such as CP55,940, only produced a very modest effect compared with the anandamide transport inhibitor AM404, which also exhibits affinity for the VR1 receptor (Zygmunt et al., 2000). This latter property of AM404 may account for its ability to reduce hyperkinesia (Lastres-Becker et al., 2002b, 2003a), as other transport inhibitors such as VDM11 and AM374, which are not active at TRPV1 receptors, were devoid of antihyperkinetic effects in HD rats (Lastres-Becker et al., 2003a), and the most potent transport inhibitor to date, UCM707, only produced a modest effect (de Lago et al., 2002, 2004b, 2006). Arvanil, a hybrid endocannabinoid and vanilloid compound, was also reported to alleviate hyperkinesias in a rat model of HD (de Lago et al., 2005). These results suggest that TRPV1 receptors alone, or in combination with CB1 receptors, might represent novel therapeutic targets in HD (reviewed in Lastres-Becker et al., 2003b).
There have been few human trials on the effects of cannabinoid agonists in HD, and the results do not live up to the promise of the animal data. Small trials with the synthetic THC analog nabilone and with the nonpsychoactive cannabidiol showed no efficacy or even increased choreic movements in HD patients (Consroe et al., 1991; Müller-Vahl et al., 1999b). These negative results could be related to dosing issues, to the lack of TRPV1 receptor activity of the compounds tested, or to the advanced stage of the disease. Nevertheless, further studies are warranted to explore the therapeutic potential of cannabinoids in HD.
c. Gilles de la Tourette’s syndrome, tardive dyskinesia, and dystonia
Based on its ubiquitous presence in motor regions of the brain, the endocannabinoid system might be involved in other extrapyramidal disorders such as Gilles de la Tourette’s syndrome (TS), tardive dyskinesia, and dystonia. TS is a neurological syndrome that becomes evident in early childhood and is characterized by multiple motor and vocal tics lasting for more than 1 year. Plant-derived cannabinoids have been found to be effective in the treatment of tics and behavioral problems in TS (Müller-Vahl et al., 1997, 1998, 1999c, 2002, 2003a,Müller-Vahl et al., b; Müller-Vahl, 2003). Beneficial effects of cannabinoids have been also reported in dystonia, both in animal models (Richter and Löscher, 1994, 2002) and in humans (Fox et al., 2002b; Jabusch et al., 2004). In addition, as described in the sections above, cannabinoids have potential in the management of the LID in PD and of the spasticity and tremor in MS. On the other hand, in patients chronically treated with neuroleptic drugs, a correlation between chronic cannabis use and the presence of tardive dyskinesia has been described previously (Zaretsky et al., 1993).
5. Amyotrophic Lateral Sclerosis
ALS (also known as Lou Gehrig’s disease) is the most common adult-onset human motor neuron disease with a prevalence of 5 to 7/100,000. It is characterized by rapid, progressive degeneration of motor neurons in the brain and spinal cord, which ultimately leads to progressive weakness, paralysis, and premature death (Rowland and Shneider, 2001). Although weak, patients are cognitively intact and thus are completely aware of their progressive disability. The disease strikes adults at any age, and most patients die within 3 to 5 years after the onset of symptoms. Although most cases of ALS are sporadic and are probably acquired, approximately 10% are familial, usually inherited in an autosomal dominant pattern. Despite a variety of putative underlying mechanisms, including oxidative stress, neuroinflammation, autoimmunity, a defect in neuronal glutamate transport and glutamate toxicity, neurofilament accumulation, exogenous factors (virusesor toxins), mitochondrial dysfunction, and mutations in the superoxide dismutase (SOD1) gene, the pathogenesis of ALS is incompletely understood (Barnham et al., 2004). Tragically, available treatment options are limited and do not prevent disease progression and death (Rowland and Shneider, 2001).
Based on the well-known protective effect of cannabinoids against oxidative cell damage and excitotoxicity (Hampson et al., 1998; Shen and Thayer, 1999; Abood et al., 2001; van der Stelt et al., 2001a), combined with their antispastic effect in MS, Carter and Rosen (2001) have proposed the use of marijuana for the pharmacological management of ALS. Indeed, in a pilot study of the safety and tolerability of THC in ALS patients, symptomatic benefits were seen for spasticity, insomnia, and appetite (Gelinas et al., 2002). Consistent with this clinical report, studies using transgenic mice expressing a mutant form of human SOD1 (hSOD1G93A mice) as an experimental model of ALS have demonstrated that either THC or WIN55,212-2 administered after the onset of the disease or genetic ablation of FAAH delayed disease progression (Raman et al., 2004; Bilsland et al., 2006). Furthermore, THC potently reduced oxidative and excitotoxic damage in spinal cord cultures in vitro and prolonged survival in SOD1 mutant mice (Raman et al., 2004). Surprisingly, neither WIN55,212-2 nor FAAH ablation affected the life span of SOD1(G93A) mice, whereas deletion of the CB1 receptor significantly extended life span without affecting the disease onset (Bilsland et al., 2006). These results suggest that cannabinoids have significant neuroprotective effects in a mouse model of ALS but that these beneficial effects may be mediated by non-CB1 receptor mechanisms.
6. Alzheimer’s Disease
Alzheimer’s disease (AD) is a progressive neurodegenerative disorder that accounts for the vast majority of age-related dementia and is one of the most serious health problems in the industrialized world. The disease is characterized by the formation of neuritic plaques rich in β-amyloid (Aβ) peptide, neurofibrillary tangles rich in hyperphosphorylated τ protein, gliosis, and a neuroinflammatory response involving astrocytes and microglia, inevitably leading to progressive global cognitive decline (Weksler et al., 2005). These studies have engendered new perspectives on the possible role of the endocannabinoid system in neurodegenerative processes associated with inflammation (reviewed in Walter and Stella, 2004), including those in AD (reviewed in Pazos et al., 2004).
In an in vitro cell culture model of AD, anandamide prevented Aβ-induced neurotoxicity through CB1-mediated activation of the mitogen-activated protein kinase pathway (Milton, 2002). In rat microglia cells in culture, CB1 receptor stimulation also dose dependently inhibited the release of NO, which had been implicated in the neurotoxic effects of Aβ peptide (Waksman et al., 1999). In PC12 cells, protection against Aβ-induced neurotoxicity was also observed with cannabidiol, which does not bind to CB1/CB2 receptors (Iuvone et al., 2004). Interestingly, CB1 receptor blockade by SR141716 improved the memory deficit induced by administration of Aβ peptide in mice, presumably by increasing hippocampal acetylcholine levels (Mazzola et al., 2003). However, analyses of brain tissue samples obtained from AD patients (Westlake et al., 1994) or animal models of AD (Romero et al., 1998; Benito et al., 2003) indicate that CB1 receptors are not dramatically affected. In contrast, CB2 receptors and FAAH are overexpressed in microglia associated with neuritic plaques in the brain of AD patients (Benito et al., 2003). Senile plaques in AD patients express both CB1 and CB2 receptors together with markers of microglial activation, and CB1-positive neurons, present in high numbers in control cases, are greatly reduced in areas of microglial activation (Ramirez et al., 2005). CB1 receptor protein levels and G protein coupling were also markedly decreased in AD brains, coupled with increased nitration of the CB1 and CB2 receptor proteins (Ramirez et al., 2005). Intracerebroventricular administration of WIN 55,212-2 to rats prevented Aβ-induced microglial activation, cognitive impairment and loss of neuronal markers. HU-210, WIN 55,212-2, and JWH-133 blocked Aβ-induced activation of cultured microglial cells, as judged by mitochondrial activity, cell morphology and TNF-α release, and these effects were independent of the antioxidant action of ligands. Furthermore, cannabinoids abrogated microglia-mediated neurotoxicity after addition of Aβ to rat cortical cocultures (Ramirez et al., 2005). Although there are no data available on the endocannabinoid content in AD brain tissue are available, increased levels have been reported in the brain after inflammatory events and in neurodegenerative disorders associated with inflammation (reviewed in Walter and Stella, 2004 and see also sections above).
Based on the above, one might hypothesize that Aβ deposition induces the release of endocannabinoids from neurons and glia, which activate CB1-mediated neuroprotective pathways and modulate the release of inflammatory mediators in microglia through CB2 receptors. If this hypothesis is confirmed by future studies, the beneficial effects of CB1/CB2 agonists and FAAH antagonists in AD could be explored. Intriguingly, in a recent open-label pilot study of six patients in the late stages of dementia (five patients with AD and one patient with vascular dementia), treatment with 2.5 mg of dronabinol daily for 2 weeks significantly improved the Neuropsychiatric Inventory total score and the subscores for agitation and aberrant motor and nighttime behaviors (Walther et al., 2006).
7. Epilepsy
If the balance between inhibitory and excitatory communications among neurons is disturbed, the intensity of excitatory transmission may exceed a certain threshold, leading to epileptic seizures. Stimulation of postsynaptic neurons is known to trigger the on-demand synthesis of endocannabinoids via an increase in intracellular calcium and/or stimulation of metabotropic receptors (reviewed in Piomelli, 2003; Lutz, 2004). Thereafter, endocannabinoids are released and reach presynaptic CB1 receptors retrogradely to modulate both inhibitory GABAergic and excitatory glutamatergic transmissions via multiple mechanisms (Marsicano and Lutz, 1999; Alger, 2002, 2004; Gerdeman et al., 2002; Robbe et al., 2002; Azad et al., 2003; Freund et al., 2003; Gerdeman and Lovinger, 2003; Kim and Alger, 2004; Isokawa and Alger, 2005).
Multiple pathways, eventually culminating in neuronal death, are triggered by excessive excitatory activity through a process known as excitotoxicity (McNamara, 1999). Excitotoxicity is believed to contribute to the progression of numerous degenerative central nervous system disorders such as Parkinson’s disease, Alzheimer’s disease, and various forms of epilepsy. More than 1% of the human population is affected by epilepsy and the incidence is highest in elderly persons or during the first years of life (reviewed in Holmes and Ben-Ari, 1998; McCormick and Contreras, 2001). Epileptic syndromes are classified as generalized seizures, which affect the entire forebrain, or partial seizures, which occur within localized brain regions. Conventional antiepileptic treatment is not fully effective in ~30% of patients, therefore justifying the search for new targets (LaRoche and Helmers, 2004).
Cannabis has been used to treat epilepsy for centuries. Hashish was reported to cure the sick son of the chamberlain of the Caliphate Council in Baghdad by the medieval Arab writer Ibn al-Badri (Mechoulam, 1986; Iversen, 2000). Almost four centuries later, W. B. O’Shaughnessy, an Irish physician and scientist working at the Medical College of Calcutta, confirmed the benefit of hashish for treating pain, emesis, muscle spasms, and convulsions (reviewed in Karler and Turkanis, 1981; Mechoulam, 1986). The benefit of cannabis in epilepsy was also reported by a British neurologist (Reynolds, 1890), but the medicinal use of cannabis was prohibited in the early 20th century in most countries.
After the identification of the structure of THC (Gaoni and Mechoulam, 1964), several groups investigated its antiepileptic effects (reviewed in Gordon and Devinsky, 2001; Lutz, 2004). THC was originally characterized as an anticonvulsant, but it has a variety of excitatory and depressant effects, ranging from convulsions to ataxia, depending on the dose, experimental model, and the animal species used (Karler and Turkanis, 1981; reviewed in Gordon and Devinsky, 2001; Lutz, 2004). Further complicating the picture, animal studies also document a rebound effect to THC with enhanced CNS excitability and increased sensitivity to convulsions (Chiu et al., 1979; Karler and Turkanis, 1981; Karler et al., 1986). This withdrawal hypersensitivity implies that in susceptible patients, the use of marijuana may be associated with withdrawal seizures (Karler and Turkanis, 1981).
Only case reports on the effects of THC in epileptic patients are currently available. Two reports described decreased seizure frequency after marijuana use (Consroe et al., 1975; Ellison et al., 1990) and an epidemiological study found that chronic marijuana use is protective against seizures (Ng et al., 1990). According to a questionnaire completed by 215 epileptic patients using marijuana regularly, 7.4% experienced a reduction, 2.3% an increase, and 90.2% no change in seizure frequency (Gordon and Devinsky, 2001). In contrast, marijuana smoking was associated with an increase in seizure frequency in another study (Keeler and Reifler, 1967). Small-scale clinical studies have shown that the nonpsychotropic cannabidiol either reduced seizure frequency or had no significant effect on it (Cunha et al., 1980; Ames and Cridland, 1986; Gordon and Devinsky, 2001).
As in human studies, cannabinoids were found to exert both anti- and proconvulsive activity in animal models of epilepsy, largely depending on the stimulus applied to induce seizures (chemical, electrical, light, or fever) and the species used (Johnson et al., 1975; Ten Ham et al., 1975; Wada et al., 1975a,b; Corcoran et al., 1978; Chiu et al., 1979; Duncan and Dagirmanjian, 1979; Fish et al., 1981; Karler and Turkanis, 1981; Colasanti et al., 1982; Fish and Consroe, 1983; Karler et al., 1984, 1986; Consroe and Mechoulam, 1987; Pertwee et al., 1991; Hayase et al., 2001a,b; reviewed in Gordon and Devinsky, 2001; Lutz, 2004).
Anandamide and its metabolically stable analog, O-1812, dose dependently inhibited electroshock-induced seizures in rats, and this effect was abolished by SR141716 (Wallace et al., 2002). In a rat model of febrile seizures, the expression of presynaptic CB1 receptors in hippocampal GABAergic interneurons was increased (Chen et al., 2003), and the CB1 receptor-mediated DSI was enhanced (Alger, 2002), suggesting that the endogenous cannabinoid system is protective. Remarkably, in a rat model of pilocarpine-induced status epilepticus, CB1 receptor agonists were more effective in reducing seizure frequency than clinically used anticonvulsants, such as phenytoin or phenobarbital. Consistently, CB1 receptor blockade increased seizure frequency, and the seizure activity was associated with increased brain levels of CB1 receptors and 2-AG (Wallace et al., 2003a).
With use of the kainic acid-induced excitotoxic epileptiform seizure model in wild type and CB1 knockout mice, recent studies have established that the seizure-induced increase of intracellular calcium, a hallmark of epilepsy (Raza et al., 2001), triggers the synthesis of anandamide, which activates CB1 receptors in glutamatergic neurons in the hippocampus and cerebral cortex (Marsicano et al., 2003; Khaspekov et al., 2004). Such “on-demand” activation of CB1 receptors was suggested to protect against excitotoxicity by various mechanisms, including inhibition of calcium channels and stimulation of potassium channels to decrease neuronal excitability and the activation of extracellular signal regulated kinases (Marsicano et al., 2003; Khaspekov et al., 2004). In contrast to these findings, FAAH knockout mice or mice treated with a CB1 agonist were found to have increased sensitivity to kainic acid-induced seizures (Clement et al., 2003). The lack of protection in this latter study may be related to the nonselective activation of CB1 receptors on both inhibitory (proconvulsive effect) and excitatory neurons (anticonvulsive effect) and by the life-long rather than on-demand activation of CB1 receptors present in FAAH knockout animals.
In summary, the use of cannabinoids for the treatment of epilepsy is still controversial, although recent experimental studies provide some new insight. To date, there have been no large-scale, controlled clinical trials to examine the beneficial effects of cannabinoids in various forms of epilepsy. The potential use of the nonpsychotropic cannabidiol and of inhibitors of anandamide transport or degradation warrants further investigation.
8. Mental Disorders
The well-known psychotropic effects of cannabinoids and the distribution of cannabinoid receptors across important emotional circuits in the brain suggest that the endocannabinoid system may be involved in various psychiatric disorders such as schizophrenia and mood disorders (reviewed in van der Stelt and Di Marzo, 2003; Hall et al., 2004; Leweke et al., 2004; Manzanares et al., 2004; Ujike and Morita, 2004; Ashton et al., 2005; Gambi et al., 2005; Semple et al., 2005; Vinod and Hungund, 2005).
a. Schizophrenia
Schizophrenia is the second most common mental disorder with a lifetime prevalence of approximately 0.2 to 2% worldwide (Ban, 2004). The disease usually begins in early adulthood or late adolescence and is characterized by psychotic episodes with positive symptoms including delusions and/or hallucinations, loose associations, and distortion of perception. The psychotic episodes are separated by periods with negative symptoms consisting of apathy, anhedonia, reduced social drive, loss of motivation, poverty of speech and thought, and blunting of affect. With disease progression, behavioral impairment can lead to complete social isolation. Although recent advances in the pharmacotherapy of schizophrenia produced great improvement in the clinical symptoms and the quality of life of patients, there is room for further improvements (Ban et al., 2004; Moller, 2005).
Numerous theories have been put forth regarding the etiology of schizophrenia, ranging from developmental or neurodegenerative processes, environmental factors, neurotransmitter abnormalities (dopamine or glutamate), and infectious or autoimmune processes, but also including the cannabinoid hypothesis (reviewed in Thaker and Carpenter, 2001; Lewis et al., 2005). It appears that hypoglutamatergic and hypodopaminergic transmission in the prefrontal cortex is involved in the negative symptoms, whereas hyperactivity of dopamine neurotransmission in the mesencephalic projections to the nucleus accumbens may underlie the positive symptoms (Thaker and Carpenter, 2001; Lewis et al., 2005).
According to the endocannabinoid hypothesis of schizophrenia, overactivity of the endocannabinoid system may lead to a hyperdopaminergic and hypoglutamatergic state, which may underlie some of the symptoms (Emrich et al., 1997, reviewed in Ujike and Morita, 2004; Laviolette and Grace, 2006). The endocannabinoid hypothesis is supported by multiple lines of evidence. First, the use of large amounts of cannabis and THC may produce psychotic symptoms in normal individuals, including delusions, hallucinations, and cognitive impairment, which resemble schizophrenia (Spencer, 1971; Halikas et al., 1972; Chopra and Smith, 1974; McGuire et al., 1994; Emrich et al., 1997; Johns, 2001; D’Souza et al., 2004). Second, cannabis and THC may worsen psychotic symptoms in schizophrenic patients, contribute to poor outcome, increase the possibility of relapse, and decrease the effectiveness of antipsychotic drugs (Breakey et al., 1974; Treffert, 1978; Negrete, 1989; Turner and Tsuang, 1990; Linszen et al., 1994; Martinez-Arevalo et al., 1994; Voruganti et al., 2001; D’Souza et al., 2005). Third, the use of cannabis may precipitate the onset of schizophrenia in individuals susceptible to psychosis (Andreasson et al., 1987; Miller et al., 2001). Fourth, postmortem radioligand studies document increased CB1 receptor density in the dorsolateral and anterior cingular regions and subregions of the prefrontal cortex in schizophrenia (Dean et al., 2001; Zavitsanou et al., 2004; Newell et al., 2006). Fifth, the levels of anandamide are increased in cerebrospinal fluid or blood from schizophrenic patients (Leweke et al., 1999; De Marchi et al., 2003; Giuffrida et al., 2004). Sixth, treatment with neuroleptics appears to normalize the imbalance in endocannabinoid signaling in blood cells in schizophrenic patients (De Marchi et al., 2003) and also decreases CB1 receptor binding in the rat nucleus accumbens (Sundram et al., 2005). Last, the hebephrenic type of schizophrenia shows a strong association with polymorphisms in the CNR1 gene encoding CB1 receptors (Leroy et al., 2001; Ujike et al., 2002).
Taken together, the above evidence suggests that the endocannabinoid system may be a novel therapeutic target in schizophrenia. It is also tempting to speculate that CB1 antagonists may be beneficial against some, most likely the negative, symptoms of the disease. Some preclinical and clinical evidence also suggests that cannabidiol may have antipsychotic potential (reviewed in Zuardi et al., 2006).
b. Anxiety and depression
Mood disorders such as generalized anxiety or panic disorder, major depressive disorder and bipolar disorder (manic depressive illness) are very common, often serious, and potentially life-threatening conditions. More than 20% of the adult population experiences a mood disorder at some point during their life. In up to 15% of individuals with major depressive disorder the cause of death is suicide. According to a World Health Organization forecast, by the year 2020 depression will become the second leading cause of premature death and disability worldwide (Pacher and Kecskeméti, 2004). Although significant advances have been made in the treatment of mood disorders during the past decades, ~30% of the population do not respond to current therapies, and the search for novel pharmacological approaches continues (reviewed in Pacher and Kecskeméti, 2004).
Many of the psychological effects of cannabis and THC are biphasic and bidirectional, depending on mode of administration, dose, personality, time frame, degree of tolerance, and various other environmental and individual factors (Paton and Pertwee, 1973; Ashton et al., 1981, 2005; Viveros et al., 2005). The acute effects in normal subjects can range from euphoria, relaxation, excitation, heightened perception, and increased motor activity to dysphoria, anxiety, sedation, perceptual distortion, and incoordination. THC, under certain conditions and at certain doses, exerts anxiolytic, antidepressant, and hypnotic effects in patients suffering from pain associated with cancer or multiple sclerosis and improves mood and general well-being in normal subjects (Regelson et al., 1976; Glass et al., 1980; Ashton et al., 1981; Fabre and McLendon, 1981; Ilaria et al., 1981; Martyn et al., 1995; Ashton, 1999; Wade et al., 2003). However, under different conditions and at higher doses, cannabis or THC can produce dysphoric reactions, anxiety, panic paranoia, and psychosis (Spencer, 1971; Halikas et al., 1972; Chopra and Smith, 1974; Ashton et al., 1981, 2005; McGuire et al., 1994; Emrich et al., 1997; Johns, 2001; Patton et al., 2002; Tournier et al., 2003; Dannon et al., 2004; D’Souza et al., 2004; reviewed in Hollister, 1986; Hall and Solowij, 1998).
CBD also possesses anxiolytic, antipsychotic and anticonvulsant properties, which are not mediated by classic cannabinoid receptors (Carlini et al., 1975; Consroe and Wolkin, 1977; Consroe et al., 1981; Zuardi et al., 1982, 1995, 2006; Ames and Cridland, 1986; Martin et al., 1987; Guimaraes et al., 1990, 1994; reviewed in Mechoulam et al., 2002c; Grotenhermen, 2003; Long et al., 2006). The mode of action of CBD is not completely understood; it may involve blockade of anandamide and serotonin reuptake (Bisogno et al., 2001; McPartland and Russo, 2001), inhibition of the enzymatic hydrolysis of anandamide (Mechoulam et al., 2002), or an interaction with as yet unidentified receptors (Járai et al., 1999; Pertwee et al., 2002).
Animal studies yielded further support to the biphasic and bidirectional effects of cannabinoids on anxiety, with low doses being anxiolytic and high doses being anxiogenic. Indeed, low doses of CP55,940 (Genn et al., 2003; Marco et al., 2004), nabilone (Onaivi et al., 1990), and THC (Berrendero and Maldonado, 2002) exerted anxiolytic-like effects in the light-dark crossing test and in the elevated plus-maze in adult rodents. Low-dose CP55,940 was also anxiolytic in other models of anxiety in adult, juvenile, or infant rodents (Romero et al., 2002a; Borcel et al., 2004; Genn et al., 2004). In contrast, at medium to high doses, CP55,940 or HU-210 displayed anxiogenic effects in the same or other experimental paradigms in adult as well as in juvenile or infant animals (McGregor et al., 1996a,b; Rodriguez de Fonseca et al., 1996; Giuliani et al., 2000; Arevalo et al., 2001; Marin et al., 2002; Romero et al., 2002; Genn et al., 2003;2003;2004; Marin Marco et al., 2004). Although several hypotheses have been proposed to explain the biphasic effects of cannabinoids on anxiety, including distinct receptors (Haller et al., 2004a,b) or neuroanatomically separated CB1 receptors with a differential sensitivity to the anxiolytic versus anxiogenic effects of cannabinoids, these need to be confirmed in future studies (reviewed in Viveros et al., 2005).
The high level of CB1 receptors in the hippocampus, amygdala, and prefrontal and anterior cingular cortex, key regions in the regulation of anxiety, may suggest that the endocannabinoid system plays a role in the control of anxiety (Herkenham et al., 1990, 1991a,b; Glass et al., 1997b; Katona et al., 2001; Hájos and Freund, 2002; Tzavara et al., 2003; Pistis et al., 2004). Further support of this theory came from studies using CB1 receptor antagonists or CB1 receptor knockout mice. SR141716 produced anxiogenic effects in the elevated plus-maze and the defensive withdrawal tests in adult rats (Navarro et al., 1997; Arevalo et al., 2001). Furthermore, SR141716 not only reversed the anxiolytic effects of the CB1 agonist CP55,940 but also was anxiogenic in the ultrasonic vocalization test in rat pups when administered alone (McGregor et al., 1996a). In contrast, Haller et al. (2002) found SR141716 to be anxiolytic in the plus-maze in mice, but this effect was not mediated by CB1 receptors as indicated by its presence in CB1 knockout mice. Furthermore, another selective CB1 receptor antagonist, AM251, increased anxiety-like behavior in wild-type mice but had no effect in the knockouts, in support of a CB1 receptor-mediated anxiolysis. As discussed before, SR141716, but not AM251, also inhibits a CB1-like receptor that mediates presynaptic inhibition of glutamate release in the hippocampus (Hájos and Freund, 2002). Thus, the findings of Haller et al. (2002) could suggest that the anxiolytic effect of SR141716 is mediated by such a CB1-like receptor, activation of which would be anxiogenic.
CB1 knockout mice displayed increased anxiogenic responses in the light-dark box, plus-maze, and social interaction tests, an increased aggressive response in the resident-intruder test, and marked alterations in the hypothalamic-pituitary-adrenal (HPA) axis coupled with impaired action of known anxiolytic drugs such as buspiron and bromazepam (Haller et al., 2002, 2004b; Martin et al., 2002; Urigüen et al., 2004). However, Marsicano et al. (2002) were unable to demonstrate anxiogenic-like response in CB1 knockout mice in the plus-maze. This may be related to differences in the genetic background of the CB1 knockout mice used and/or different experimental conditions. The importance of the latter is also indicated by the confounding effect of stress on anxiogenic behaviors and their modulation by endocannabinoids (Haller et al., 2004a; Patel et al., 2005). Stress-induced down-regulation of hippocampal endocannabinoid signaling may contribute to problems in behavioral flexibility and may play a role in the development of perseveratory and ruminatory behaviors in stress-related neuropsychiatric disorders (Hill et al., 2005). Collectively, a majority of evidence supports a role for CB1 receptors in the control of emotional behavior and suggests the existence of an anxiolytic endocannabinoid tone. Facilitation of such a tone by inhibiting the degradation of endocannabinoids in vivo may be therapeutically exploited, as indicated by the reduced anxiety-like behavior and potent antidepressant-like effects in mice and rats treated with a FAAH or anandamide transport inhibitor and the blockade of this effect by SR141716 or AM281 (Kathuria et al., 2003; Gobbi et al., 2005; Bortolato et al., 2006; Rutkowska et al., 2006).
The mechanisms responsible for the effects of cannabinoids on anxiety-related responses are complex and may involve modulation of numerous neurotransmitter systems. For example, stimulation of CB1 receptors in rodents activates the HPA axis through the release of CRH (Weidenfeld et al., 1994; Wenger et al., 1997; Martin-Calderon et al., 1998; Manzanares et al., 1999a; Marco et al., 2004), which could account for the anxiogenic effects of high doses of cannabinoids (Rodriguez de Fonseca et al., 1996; Marin et al., 2002). In contrast, there are also examples of negative modulation of HPA function by endocannabinoids (Di et al., 2003; Patel et al., 2004). Cannabinoids also modulate GABAergic transmission and the release of the peptide cholecystokinin, which may contribute to both anxiolytic and anxiogenic effects (Onaivi et al., 1990; Katona et al., 1999, 2001; Marsicano and Lutz, 1999; Tsou et al., 1999; Beinfeld and Connolly, 2001; Rotzinger and Vaccarino, 2003). Furthermore, cannabinoids enhance the release of endogenous opioids and a functional interplay between the endocannabinoid and opioid systems modulates analgesic responses and is involved in antidepressant-like effects and in various addiction-related processes (Pugh et al., 1997; Manzanares et al., 1999b; Houser et al., 2000; Zimmer et al., 2001; Ghozland et al., 2002). From studies with THC and CP55,940, it appears that μ- and δ-opioid receptors mediate certain anxiolytic effects, whereas activation of κ-opioid receptors leads to increased anxiety (Pugh et al., 1997; Houser et al., 2000; Zimmer et al., 2001; Berrendero and Maldonado, 2002; Ghozland et al., 2002; Marin et al., 2003). There are also interactions between the endocannabinoid and serotonergic systems (Arevalo et al., 2001; Malone and Taylor, 2001; Fride and Shohami, 2002; Marin et al., 2003; Marco et al., 2004; Steffens and Feuerstein, 2004; reviewed in Viveros et al., 2005), although their role in anxiety-like behaviors has not been explored
In contrast to earlier dogma, recent findings indicate that neurogenesis occurs in the adult brain. Furthermore, stress and depression decrease neurogenesis, particularly in the hippocampus, whereas electroconvulsive therapy and chronic treatment with conventional antidepressants increases this process (reviewed in Pacher et al., 2004a). It has been recently demonstrated that the endocannabinoid system drives neural progenitor cell proliferation (Aguado et al., 2005, 2006), and cannabinoids promote neurogenesis (Berghuis et al., 2005; Jiang et al., 2005). Furthermore, CB1 receptors appear to be required for neuronal survival in the hippocampus (Bilkei-Gorzo et al., 2005). These findings are particularly exciting, as they raise the possibility of a role for endocannabinoids in antidepressive drug action. Indeed, CB1 receptor density in the hippocampus and hypothalamus is increased by chronic tricyclic antidepressant treatment (Hill et al., 2006), and the amplification of the actions of endocannabinoids by the FAAH inhibitor URB597 was found to produce antidepressant-like effects in the mouse tail-suspension and rat forced-swim tests, without eliciting reward-related effects indicative of addictive potential (Gobbi et al., 2005). It should not be surprising, however, that based on the basis of the bimodal action of cannabinoids on mood and anxiety, a case could be made for the opposite, i.e., for the antidepressive potential of CB1 antagonism. CB1 antagonists were reported to elicit antidepressant-like behavioral effects in rodents and can increase the synaptic concentration of biogenic amines, much like antidepressants do (reviewed in Witkin et al., 2005). Thus, pharmacological modulation of the endocannabinoid system holds considerable promise in the treatment of both anxiety-related and mood disorders.
The results of a recent study implicated endocannabinoids and CB1 receptors in the extinction of aversive memories by demonstrating that CB1 knockout mice show impaired extinction in auditory fear-conditioning tests, and this could be mimicked in wild-type mice by treatment with SR141716 (Marsicano et al., 2002b). These exciting findings raise the possibility that pharmacological amplification of CB1 signaling, for example, by FAAH inhibitors, may have therapeutic value in obsessive-compulsive disorder or post-traumatic shock syndrome.
9. Insomnia
Insomnia, the most common sleep disorder, is defined as difficulty with the initiation, maintenance, duration, or quality of sleep that results in the impairment of daytime functioning, despite adequate opportunity and circumstances for sleep (Silber, 2005). The cause for insomnia is often not known, but frequently it may be a consequence of a chronic disease associated with pain or depression.
Early studies documented the fact that marijuana and THC affect sleep patterns both in humans (Freemon, 1972, 1982; Pivik et al., 1972; Barratt et al., 1974; Feinberg et al., 1975, 1976) and in experimental animals (Monti, 1977; Buonamici et al., 1982). More recently, Nicholson et al. (2004) have studied the effects of cannabis extracts on nocturnal sleep, early-morning performance, memory, and sleepiness in a placebo-controlled, double-blind, crossover study in eight healthy volunteers. They found that 15 mg of THC was sedative, whereas 15 mg of CBD had alerting properties as it increased wake activity during sleep and counteracted the residual sedative activity of THC (Nicholson et al., 2004).
Anandamide was also found to modulate sleep by increasing slow-wave sleep two and rapid eye movement sleep in a CB1 receptor-dependent manner in rats (Murillo-Rodriguez et al., 1998, 2001). Moreover, CB1 receptor expression in the pons of rats was modulated by the light/dark cycle and by sleep (Martinez-Vargas et al., 2003), and endocannabinoids and CB1 receptors were also implicated in rapid eye movement sleep rebound (Navarro et al., 2003). Interestingly, a recent study has demonstrated that anandamide not only induced sleep but also increased levels of the sleep-inducing substance adenosine in the basal forebrain, and both of these effects were blocked by SR141716 (Murillo-Rodriguez et al., 2003).
Oleamide is a fatty acid amide with a variety of in vitro effects, including inhibition of gap junction-mediated cell-cell communication (Boger et al., 1998a,b), modulation of 5-HT1, 5-HT2A,C, and 5-HT7 receptors (Thomas et al., 1997, 1999; Hedlund et al., 1999), and modulation of inhibitory ionotropic receptors such as the GABAA receptor (Coyne et al., 2002). Oleamide accumulates in the cerebrospinal fluid of sleep-deprived cats (Cravatt et al., 1995) and rats (Basile et al., 1999) and induces sleep, an effect which could be blocked by SR141716 (Mendelson and Basile, 1999). Initially, it was suggested that inhibition of anandamide degradation by FAAH rather than the activation of CB1 receptors was responsible for the sleep-inducing effect of oleamide (Boring et al., 1996; Mechoulam et al., 1997), but this is a matter of dispute (Fowler, 2004; Lees and Dougalis, 2004; Leggett et al., 2004).
Although little is known about the role of the endocannabinoid system in the pathophysiology of sleep disorders, clinical studies uniformly report significantly improved sleep quality in patients taking cannabinoids for symptomatic treatment of multiple sclerosis, cancer, chronic pain, or intractable pruritus. Although psychotropic cannabinoids are unlikely to gain acceptance for the treatment of insomnia, FAAH inhibitors were shown to enhance certain endocannabinoid-mediated behaviors without evidence for addictive properties (Kathuria et al., 2003). The sleep-inducing property of some potent FAAH inhibitors, such as the endogenous lipid 2-octyl γ-bromoacetoacetate (Boger et al., 1998a), could therefore be therapeutically exploited.
10. Nausea and Emesis
Nausea and vomiting can present as symptoms of a variety of diseases or as secondary consequences of chemotherapy or radiotherapy of cancer. It is for this latter indication that THC has gained acceptance as a highly efficacious therapeutic agent, often effective in cases resistant to other, more conventional, medications (reviewed by Martin and Wiley, 2004; Aapro, 2005; Hall et al., 2005). Emesis is thought to involve activation of specific receptors on sensory nerve endings in the gut and also in brainstem regions including the medullary chemoreceptor trigger zone and the lateral reticular formation. Activation of 5-HT3 receptors appears to play a dominant role in acute emesis, whereas activation of NK1 (substance P) receptors is more important in the delayed emesis after chemotherapy, as indicated by the effectiveness of the respective receptor antagonists in controlling these different stages of the emetic response (Aapro, 2005). Although the mechanism of the antiemetic action of cannabinoids is not quite clear, an interaction with 5-HT3 is suggested by the colocalization of CB1 and 5-HT3 receptors on GABAergic neurons where they have opposite effects on GABA release (Morales et al., 2004). Also, cannabinoids may directly inhibit 5-HT3-gated ion currents by a mechanism not involving CB1 receptors (Fan, 1995; Barann et al., 2002). Such a CB1 receptor-independent effect is also suggested by the ability of cannabidiol, a natural constituent of marijuana which does not bind to the CB1 receptor, to reduce lithium-induced vomiting in the house musk shrew (Parker et al., 2004). Nevertheless, the involvement of CB1 receptors is clearly indicated by the ability of SR141716 to reverse the effects of THC and synthetic agonists in suppressing vomiting caused by cisplatin (Darmani, 2001b) or lithium chloride (Parker et al., 2004), or by the ability of these agonist to reverse the emesis elicited by SR141716 in the least shrew (Darmani, 2001a). These latter findings suggest that the emetic circuitry is tonically controlled by endocannabinoids.
In line with such a possibility, a recent human study found an association between chronic marijuana use, which probably results in desensitization of cannabinoid receptors, and cyclical hyperemesis: in the 19 subjects studied, the hyperemetic episodes subsided upon discontinuation of cannabis use and reappeared upon rechallenge with cannabis (Allen et al., 2005). A meta-analysis of 30 randomized comparisons of cannabis (nabilone, dronabinol, or levonantradol) with placebo or standard antiemetics, involving a total of 1366 patients, concluded that cannabinoids are slightly more effective than conventional antiemetics, and the patients prefer them because of their mood enhancing and sedative effects. However, they were also more toxic, with dizziness, dysphoria, hallucinations, and paranoia being the most prominent undesirable side effects (Tramér et al., 2001). This led to the recommendation to limit the use of cannabinoids as antiemetics to patients with chemotherapy-related sickness, in whom their mood-enhancing effects would be of added benefit.
11. Drug Addiction and Alcohol Disorders
The positive reinforcing effect of natural rewards, such as those derived from eating, drinking, work, or sexual activity, are mediated by the brain’s reward circuitry. Neuroanatomically, this circuitry consists of three series of coupled pathways. First-order neurons project from structures in the ventral limbic forebrain (orbitofrontal cortex and anterior cingulate area) to the mesencephalic ventral tegmental area (VTA) where they synapse onto dopaminergic neurons. These second-order neurons project primarily to neurons in the shell of the nucleus accumbens (nAc), but also to cortical areas and to the amygdala. Third-order neurons in the nAc, some of which are GABAergic, project to the ventral pallidum and other regions involved in mediating reward-related behaviors (recently reviewed by Lupica et al., 2004; Gardner, 2005). It is believed that addictive drugs activate or “hijack” the same pathway. Genetic vulnerability to drug addiction has been linked to a functional deficiency in the second-order dopaminergic neurons at their interface with third-order neurons in the nAc (Nestler, 2003). In human subjects prone to addiction, a deficiency in D2 dopamine receptors in the nAc could be documented by brain imaging (Volkow et al., 1997, 1999).
A common denominator among different addictive drugs interacting with distinct receptors is their ability to activate the mesolimbic dopaminergic reward pathway and increase dopamine levels in the nAc, which is believed to be responsible for their addictive properties (Koob, 1992; Wise, 2004). Similar to other drugs of abuse, THC increases extracellular dopamine levels in the nAc via activation of CB1 receptors (Chen et al., 1990; Tanda et al., 1999) and also lowers the reward threshold for electrical brain stimulation (Gardner et al., 1988), a phenomenon known to involve activation of the mesolimbic dopamine system. THC also increases the firing rate of the second-order VTA-nAc dopaminergic neurons via CB1 but not opiate receptors (French, 1997), and withdrawal from THC increases corticotropin-releasing factor levels in the central nucleus of the amygdala (Rodriguez de Fonseca et al., 1997), another hallmark of drugs of abuse (Koob, 1996).
THC and related synthetic cannabinoid agonists also fulfill the reward-related behavioral criteria for drugs of abuse: they support conditioned place preference (CPP) under appropriate conditions (Lepore et al., 1995; Valjent and Maldonado, 2000; Zangen et al., 2006), they are self-administered intravenously or intracerebrally in a CB1 antagonist-sensitive manner (Martellotta et al., 1998; Ledent et al., 1999; Braida et al., 2001; Zangen et al., 2006), and they reinstate cocaine-or heroine-seeking behavior in rats previously extinguished from self-administration (De Vries et al., 2001).
An issue of intense interest is the location of the CB1 receptors mediating these effects. Similar to cannabinoids, opiates also increase the activity of dopaminergic neurons in the VTA. This effect has been shown to result from μ receptor-mediated inhibition of GABA release from the terminals of inhibitory GABAergic interneurons, i.e., through a “disinhibitory” mechanism (Johnson and North, 1992). A similar mechanism has been postulated for cannabinoids by Cheer et al. (2000), who reported that local application of the cannabinoid agonist HU-210 to brain slices containing the VTA increased dopaminergic neuronal activity, which could be blocked by the GABAA antagonist bicuculline. In line with this, WIN 55,212-2 was found to suppress electrically evoked, but not muscimol-induced, inhibitory postsynaptic currents via CB1 receptors in brain slices containing the VTA (Szabo et al., 2002). However, cannabinoids also inhibit glutamate release in the VTA, which would have an opposite effect on dopaminergic activity (Melis et al., 2004a). There is evidence for additional sites of action, such as CB1 receptors on the terminals of GABAergic projection neurons that target GABAB receptors on VTA dopamine neurons resulting in their disinhibition (Riegel and Lupica, 2004). This pathway may be activated by ethanol, as indicated by the ability of the GABAB agonist baclofen to antagonize the increase in ethanol drinking caused by WIN 55,212-2 treatment of alcohol-preferring rats (Colombo et al., 2004). Activation of CB1 receptors on glutamatergic terminals in the nAc was reported to inhibit glutamate release onto GABAergic neurons in the nAc that project to the VTA, which may also result in disinhibition of VTA dopaminergic neurons (Robbe et al., 2001). Indeed, both the VTA and the nAc may be sites of the rewarding effects of cannabinoids, as documented by the propensity of rats to self-administer THC into either site (Zangen et al., 2006).
Regardless of the exact location of presynaptic CB1 receptors, their natural activation occurs through retrograde transmission, with their endogenous ligands being released from postsynaptic cells (Kreutzer and Regehr, 2001; Ohno-Shosaku et al., 2001; Wilson and Nicoll, 2001). This mechanism has also been implicated in LTD (Gerdeman et al., 2002; Robbe et al., 2002), a form of synaptic plasticity that can be initiated by drugs of abuse (Thomas et al., 2001), and may be involved in certain features of compulsive drug use (Gerdeman et al., 2003). A further indication that endocannabinoids may be involved in mechanisms of drug reward is findings that the neurochemical and behavioral responses to different classes of drugs of abuse can be inhibited by the CB1 receptor antagonists. These findings suggests that endocannabinoid activation of CB1 receptors in the mesolimbic reward pathway may be part of a “common pathway” of drug reward (reviewed in De Vries and Schoffelmeer, 2005; Maldonado et al., 2006). Examples of this are discussed below.
a. Opiates
There is a large body of evidence indicating a reciprocal relationship between the endocannabinoid and endogenous opioid systems in drug dependence (recently reviewed by Fattore et al., 2005; Vigano et al., 2005a,b). This fact is not surprising, given that opioids and cannabinoids have a similar pharmacological profile at both the behavioral level (e.g., analgesia, hypothermia, catalepsy, and motor impairment) and cellular/molecular levels (both CB1 and opiate μ receptors are predominantly presynaptic, they are coupled to and share the same pool of Gi/Go proteins, and have an overlapping brain distribution). There are numerous examples for opioid or cannabinoid reward-related effects being inhibited by both CB1 and opiate μ antagonists (Fattore et al., 2005; Gardner, 2005; Vigano et al., 2005a,b). The mechanisms underlying these reciprocal interactions are not clear, but they may involve heterodimerization of CB1 and μ opiate receptors, depletion of shared G protein pools and/or utilization of common postreceptor signaling pathways. In addition, the opiate/cannabinoid synergism observed in nAc/striatal neurons appears to require adenosine and A2a receptor signaling (Yao et al., 2006).
Here we will only review evidence that pertains to the potential involvement of endocannabinoids in the addictive, reward-related actions of opioids. Such evidence is based on the ability of pharmacological or genetic ablation of CB1 receptors to prevent or inhibit opioid effects. CB1 knockout mice were reported to be unable to acquire morphine self-administration (Ledent et al., 1999; Cossu et al., 2001), to have reduced morphine withdrawal symptoms (Ledent et al., 1999), and not to develop CPP for morphine (Martin et al., 2000). A possible neurochemical correlate of these changes is the lack of morphine-induced dopamine release in the nucleus accumbens of CB1 receptor knockout mice (Mascia et al., 1999), although more recently CB1 blockade was found to reverse the morphine-induced decrease in ventropallidal GABA overflow without affecting the morphine-induced increase in dopamine release in the nAc (Caillé and Parsons, 2006). Treatment of wild-type mice and rats with a CB1 antagonist elicits similar phenotypes (Rubino et al., 2000; Mas-Nieto et al., 2001; Navarro et al., 2001, 2004). These observations raise the therapeutic potential of chronic treatment with a CB1 receptor antagonist in preventing or reversing the development of opiate dependence.
b. Nicotine
Nicotine is the main neuroactive component in tobacco smoke and is responsible for its addictive properties. Nicotine’s rewarding effects are mediated by the same mesolimbic dopaminergic pathway that is involved in the rewarding effects of many other addictive drugs (Pontieri et al., 1996). Therefore, it should not be unexpected that there is a positive synergism between nicotine and THC in paradigms used to reveal reinforcing effects (Valjent and Maldonado, 2000). A role of endocannabinoids in the rewarding effects of nicotine is indicated by the absence of nicotine-induced CPP in CB1 knockout mice (Castane et al., 2002), although the acquisition of nicotine self-administration was not affected by the absence of CB1 receptors in another study using an acute reinforcement paradigm (Cossu et al., 2001). On the other hand, SR141716 was reported to decrease nicotine operant self-administration (Cohen et al., 2002) and nicotine-induced CPP in rats (Le Foll and Goldberg, 2004; Forget et al., 2006) and also to inhibit nicotine-induced dopamine release in the nucleus accumbens shell (Cohen et al., 2002). SR141716 also inhibited nicotine self-administration sustained by nicotine-associated cues in the absence of nicotine itself (Cohen et al., 2005), and chronic exposure to nicotine was reported to induce endocannabinoid release (Gonzalez et al., 2002). Furthermore, SR141716 abolished the anxiolytic effects of low-dose nicotine in mice and potentiated its anxiogenic effects at higher doses (Balerio et al., 2006). Together, these findings justified testing rimonabant in clinical trials to promote smoking abstinence. Indeed, the results of a recent multicenter phase III clinical trial in the United States indicate that a 10-week treatment of smokers with a daily oral dose of 20 mg of rimonabant with a follow-up period of 42 weeks doubled the odds of quitting smoking, was well tolerated, and also reduced the post-cessation weight gain by >80% (Dale and Anthenelli, 2004).
c. Cocaine
Unlike THC, opiates and nicotine, cocaine does not increase the activity of dopaminergic neurons in the VTA but elevates synaptic levels of dopamine in the nAc by blocking dopamine reuptake at the dopamine transporter (Giros et al., 1996). Therefore it is not surprising that cocaine-induced increases in dopamine in the nAc were found to be unaffected by genetic ablation of CB1 receptors (Soria et al., 2005). Accordingly, CB1 receptors do not appear to participate in the acute rewarding properties of cocaine, as indicated by the preserved acute cocaine self-administration and cocaine-induced CPP in CB1 knockout mice (Martin et al., 2000; Cossu et al., 2001; Lesscher et al., 2005; Soria et al., 2005) or in mice treated with SR141716 (Tanda et al., 2000; De Vries et al., 2001; Caillé and Parsons, 2006). SR141716 treatment also did not affect the threshold-lowering effect of cocaine in the intracranial self-stimulation paradigm, although treatment with WIN 55,212-2 was able to achieve this, suggesting that CB1 receptor stimulation might inhibit the reinforcing properties of cocaine (Fattore et al., 1999; Vlachou et al., 2003).
Other studies indicate, however, that endocannabinoid activation of CB1 receptors may mediate the reinforcing effects of cocaine. SR141716 treatment decreased the sensitivity of rats to the reinforcing effects of cocaine in an intracranial self-stimulation paradigm (Deroche-Gamonet et al., 2001). The ability to acquire operant self-administration of cocaine was reduced in CB1 knockout mice or in SR141716-treated wild-type mice, which also displayed a reduced maximal effort to obtain cocaine infusion in a progressive ratio schedule, compared with untreated wild-type mice (Martin et al., 2000; Soria et al., 2005). Furthermore, prior use of cannabis was found to enhance the “high” elicited by subsequent use of cocaine in humans (Foltin et al., 1993; Lukas et al., 1994) and also to hasten relapse in abstinent former cocaine users (Rawson et al., 1986). Furthermore, a recent genetic study found an association between an (AAT)n triplet repeat polymorphism in the CNR1 gene encoding the CB1 receptor with cocaine addiction in an African-Caribbean population (Ballon et al., 2006). Treatment with HU-210 promoted reinstatement of cocaine-seeking behavior in rats, whereas treatment with SR141716 prevented reinstatement (De Vries et al., 2001). Thus, the endocannabinoid system may be involved in the acquisition and consolidation of cocaine addiction as well as in relapse, through mechanisms other than an effect on the cocaine-induced increase in dopaminergic transmission in the nAc. These latter studies also predict the possible effectiveness of rimonabant in the treatment of cocaine addiction.
d. Alcohol
Several lines of evidence indicate the involvement of the endocannabinoid system in alcohol drinking behavior (recently reviewed by Colombo et al., 2005). Chronic alcohol intake increases endocannabinoid levels in the limbic forebrain (Gonzalez et al., 2002) and decreases CB1 receptor binding and signaling (Basavarajappa and Hungund, 2002). Studies in the late 1990s indicated the effectiveness of SR141716 in reducing voluntary ethanol intake in rodent models of ethanol drinking (Arnone et al., 1997; Colombo et al., 1998b; Freedland et al., 2001), whereas cannabinoid agonists promoted drinking (Gallate et al., 1999; Colombo et al., 2002). Operant self-administration of ethanol and relapse to drinking are also inhibited by SR141716 (Cippitelli et al., 2005; Economidou et al., 2006) and potentiated by chronic exposure to a cannabinoid agonist (Lopez-Moreno et al., 2005).
The possible role of the endocannabinoid system in ethanol preference was further indicated by observations of reduced voluntary ethanol drinking in CB1 knockout compared with wild-type mice (Hungund et al., 2003; Poncelet et al., 2003; Wang et al., 2003; Lallemand and de Witte, 2004; Naassila et al., 2004; Thanos et al., 2005), although no difference was noted in one study (Racz et al., 2003). Sensitivity to alcohol is inversely related to the chance of becoming an alcoholic among humans (Schuckit, 1997), and the same inverse relationship was noted in CB1 knockout mice and their wild-type littermates (Naassila et al., 2004). The reduced voluntary ethanol intake in CB1 knockout mice was associated with reduced alcohol-induced CPP (Houchi et al., 2004; Thanos et al., 2005), a further indication of the role of CB1 receptors in the rewarding effects of alcohol.
Similar to cannabinoids and other drugs of abuse, alcohol intake can also result in increased dopamine release in the nAc (Weiss et al., 1993; Campbell and McBride, 1995). The reported absence of such release in CB1 knockout mice and the ability of SR141716 to block ethanol-induced dopamine release in wild-type mice further suggest the involvement of endocannabinoids in the reinforcing effects of ethanol. However, the brain site where ethanol-induced endocannabinoid release and CB1 receptor activation occur is not yet known. The recent observation that microinjection of SR141716 into the prefrontal cortex of alcohol-preferring AA rats inhibited ethanol self-administration suggests that this region may be one of the sites involved (Hansson et al., 2006). In the same study, FAAH activity and CB1 signaling were both reduced in the same brain region of AA rats compared with their nonpreferring ANA counterparts, and microinjection of the FAAH inhibitor URB597 increased ethanol self-administration (Hansson et al., 2006). Analogous findings in female FAAH knockout mice are their increased voluntary ethanol intake and decreased alcohol sensitivity (Basavarajappa et al., 2006). These findings suggest that increased anandamide tone secondary to decreased FAAH activity in the prefrontal cortex may be causally linked to high alcohol preference. Such a scenario would be compatible with evidence for an association between problem drug and alcohol use and a missense mutation in the human FAAH gene (Sipe et al., 2002).
A number of mediators have been implicated in the control of appetite for both food and alcohol. In the case of endocannabinoids, the regulation is “unidirectional”, i.e., endocannabinoids promote both food intake (see section III.A.3.) and alcohol drinking. Because both food intake and alcohol drinking activate the brain reward pathways, one might postulate that the role of endocannabinoids in promoting drinking behavior would be most prominent in the type of alcoholics who drink for the rewarding effects of alcohol, such as young binge-drinkers. The high alcohol preference of C57BL6 mice and the role of the endocannabinoid system mediating it were found to be age-dependent (Wang et al., 2003), which is compatible with such a possibility. In contrast, the effects of NPY and CRH on food intake and ethanol consumption are bidirectional: NPY increases food intake (Clark et al., 1984) but reduces ethanol consumption (Thiele et al., 1998), whereas CRH is anoretic (Britton et al., 1982) but promotes ethanol drinking (George et al., 1990). The effects of NPY and CRH on alcohol preference correlate with their effects on anxiety-like behaviors, NPY being anxiolytic (Heilig et al., 1989) and CRH being anxiogenic (Koob and Thatcher-Britton, 1985). We would predict that CB1 antagonists will be more effective in reducing the drive to drink in younger people who drink for the rewarding effects of alcohol, whereas CRH antagonists or NPY agonists would be more effective in older, chronic alcoholics who more likely drink to suppress the negative affect and anxiety of alcohol withdrawal. This hypothesis may be tested by appropriately designed clinical trials. Studies to test the safety and efficacy of rimonabant in the treatment of alcoholism and alcohol abuse are currently underway at the National Institute on Alcohol Abuse and Alcoholism.
e. Psychostimulants
3,4-Methylenedioxymethamphetamine (MDMA, Ecstasy) is a psychostimulant abused for its euphorigenic and stimulant properties, and it is often used in combination with marijuana. Intracerebral self-administration of MDMA was found to be reduced in the presence of the cannabinoid agonist CP55,940 and increased after treatment with SR141716. These findings were interpreted to indicate synergism between the reinforcing effects of cannabinoids and MDMA and a reduction in the motivational value of MDMA by CB1 blockade (Braida and Sala, 2002). In another study, the authors found that SR141716 blocked MDMA-induced CPP (Braida et al., 2005). Amphetamine-induced long-term synaptic depression in the amygdala could be blocked by the CB1 antagonist AM251, mimicked by the agonist WIN 55,212-2, and occluded by the transport inhibitor AM404, suggesting that amphetamine-induced LTD and related behavioral effects may be mediated via endocannabinoid release (Huang et al., 2003). Together, these findings suggest that CB1 antagonists may be of value in the treatment of addiction to psychostimulants, including amphetamine and MDMA.
D. Cardiovascular and Respiratory Disorders
Besides their well known neurobehavioral and immunological actions, cannabinoids and their endogenous and synthetic analogs exert important cardiovascular effects. The underlying mechanisms are complex, involving direct effects on the vasculature (Gebremedhin et al., 1999; Járai et al., 1999; Wagner et al., 2001b; Wagner et al., 2005) and myocardium (Bonz et al., 2003; Maslov et al., 2004; Sterin-Borda et al., 2005), as well as modulation of autonomic outflow through sites of action in the central (Niederhoffer and Szabo, 2000; Pfitzer et al., 2004) and the peripheral nervous systems (Ishac et al., 1996; Malinowska et al., 1997; Szabo et al., 2001; Niederhoffer et al., 2003). As for endogenous cannabinoids, their effects are also complicated by their rapid metabolism, which liberates arachidonic acid that can be further metabolized into vasoactive prostanoids (reviewed in Mechoulam et al., 1998; Kunos et al., 2000; Randall et al., 2002; Ralevic et al., 2002).
Studies to date indicate that CB1 receptors are much more important than CB2 receptors in cardiovascular regulation, the latter so far being implicated only in ischemic preconditioning and ischemia/reperfusion (I/R) injury of the myocardium (see below). CB1 receptors have been detected in the human, rat, and mouse myocardium where they mediate negative inotropy (Bonz et al., 2003; Bátkai et al., 2004b; Pacher et al., 2004, b, 2005a,b,d; Engeli et al., 2005; Wagner et al., 2005) and also in vascular tissues (Gebremedhin et al., 1999; Liu et al., 2000), where their activation leads to vasodilation, and both of these effects appear to be involved in the hypotensive effect of anandamide (Wagner et al., 2001a,b; Bátkai et al., 2004a,b; Pacher et al., 2004, b, 2005a,b,d) in anesthetized rodents. Sympathetic nerve terminals contain presynaptic CB1 receptors, stimulation of which inhibits norepinephrine release (Ishac et al., 1996), which contributes to the bradycardic effects of anandamide in vivo (Wagner et al., 2001b). Anandamide-induced cardiovascular depressor effects are devoid of a centrally mediated component (Varga et al., 1996), in contrast to the effects of certain synthetic cannabinoids, which cause centrally mediated sympathoexcitation (Niederhoffer and Szabo, 2000; Gardiner et al., 2001, 2002b).
The vasorelaxant effect of endocannabinoids and synthetic cannabinoids in vitro are complex and display tissue and interspecies differences. They may involve CB1 and TRPV1 receptor- and NO-mediated or NO-independent mechanisms and also as yet undefined endothelial site(s) of action. A detailed discussion of these in vitro vasodilatory effects can be found in recent reviews (Hillard, 2000; Kunos et al., 2000, 2002; Ralevic et al., 2002; Randall et al., 2002, 2004; Begg et al., 2005; Pacher et al., 2005a,b) and is beyond the scope of this review.
Compared with the growing body of information on the vascular effects of cannabinoids, less is known about cannabinoid-induced direct cardiac effects. Anandamide, R-methanandamide, and HU-210 dose dependently decrease contractile performance in isolated, electrically paced human atrial muscle, an effect blocked by the potent CB1 antagonist AM251, whereas the involvement of CB2 receptors, NO, or prostanoids could be excluded (Bonz et al., 2003). HU-210 also decreased left ventricular developed pressure in isolated perfused rat hearts through CB1 receptor activation (Maslov et al., 2004; Krylatov et al., 2005). Another study using isolated, perfused, rat Langendorff heart preparations to study the effects of anandamide, R-methanandamide, and palmitoylethanolamide on coronary perfusion pressure and left ventricular developed pressure suggested the involvement of a cardiac site of action distinct from CB1 and CB2 receptors (Ford et al., 2002).
Several studies have examined the in vivo hemodynamic effects of endocannabinoids and their synthetic analogs in rodents (recently reviewed in Begg et al., 2005; Pacher et al., 2005a,b). Intravenous administration of anandamide causes a triphasic blood pressure response in anesthetized mice and rats, in which a prolonged hypotensive effect (phase III) is preceded by a transient, vagally mediated, fall in heart rate, cardiac and contractility, and blood pressure and an increase in total peripheral resistance (phase I) followed by a brief, pressor response (phase II) associated with increased cardiac contractility (Varga et al., 1995; Lake et al., 1997b; Pacher et al., 2004, b, 2005d). Inhibition of the phase I bradycardic response by TRPV1 receptor antagonists in rats (Malinowska et al., 2001) and the absence of both phase I and phase II responses in TRPV1−/− mice (Pacher et al., 2004) imply that these components are mediated by TRPV1 receptors. Additional central and vascular mechanisms may also be involved in the brief pressor response (phase II) in anesthetized rats (Kwolek et al., 2005). The third, prolonged hypotensive phase (phase III) is characterized by marked decreased cardiac contractility and slightly decreased total peripheral resistance, and it lasts up to 10 min in anesthetized mice (Pacher et al., 2004b, 2005d), similar to the hypotensive effect previously described in anesthetized but not conscious rats (Stein et al., 1996; Varga et al., 1996; Lake et al., 1997a,b; Gardiner et al., 2002a; Bátkai et al., 2004b) and also observed with synthetic cannabinoids (Vidrio et al., 1996; Lake et al., 1997a; Pacher et al., 2005d).
The anandamide-induced phase III hypotension and decreased cardiac contractility, as well as similar hemodynamic responses to synthetic cannabinoids, are mediated by CB1 receptors. First, these effects are prevented or reversed by selective CB1 antagonists both in normal rodents (Varga et al., 1995, 1996; Calignano et al., 1997; Pacher et al., 2004, b, 2005a,d) and in mice lacking FAAH, which exhibit increased sensitivity to hypotensive and cardiodepressant effects of anandamide (Pacher et al., 2005d). Second, there is a positive correlation between the concentrations of various cannabinoid agonists in producing half-maximal hypotensive and bradycardic responses (EC50) and in their affinity constants for binding to CB1 receptors in the brain (Lake et al., 1997a). Third, cannabinoid-induced hypotension and bradycardia are absent in mice lacking the CB1 receptor (Járai et al., 1999; Ledent et al., 1999). The involvement of the endocannabinoid system in various cardiovascular disorders is reviewed below.
1. Hypertension
Chronic use of cannabis in humans as well as both acute and prolonged administration of THC to experimental animals elicits a long-lasting decrease in blood pressure and heart rate (Rosenkratz and Braude, 1974; Benowitz and Jones, 1975), whereas the acute effect of smoking cannabis usually increases heart rate with no consistent change in blood pressure (Kanakis et al., 1976). In a recent study conducted in 63 male cannabis smokers, 22% of subjects experienced symptomatic hypotension, which could be reversed by the administration of 30 or 90 mg but not lower doses of rimonabant, indicating that CB1 receptors mediate the hypotensive effect of cannabis smoking in humans (Gorelick et al., 2006).
More than three decades ago, several studies explored the potential use of cannabinoids to treat hypertension (Birmingham, 1973; Archer, 1974; Varma and Goldbaum, 1975; Adams et al., 1977; Crawford and Merritt, 1979; Zaugg and Kyncl, 1983). Unfortunately, the initial high anticipation was tempered by a report of the development of rapid tolerance to the hypotensive and bradycardic effects of THC (Adams et al., 1976) and by the failure to separate the cardiovascular and neurobehavioral effects of cannabinoids. Albeit a later study in spontaneously hypertensive rats (SHR) demonstrated no tolerance to the same effects during a 10-day treatment period (Kosersky, 1978), interest in this issue had vanished for the next two decades.
As with many other effects of marijuana, the discovery of endocannabinoids has focused attention on their possible role in cardiovascular regulation. Studies with SR141716 indicated that the hypotensive/bradycardic effects of exogenous anandamide, THC, and potent synthetic cannabinoids are mediated by CB1 receptors (Varga et al., 1995; Lake et al., 1997a). CB1 receptor knockout mice have normal blood pressure (Járai et al., 1999; Ledent et al., 1999) and the blood pressure of normotensive mice and rats is unaffected or slightly reduced by CB1 antagonists (Varga et al., 1995; Lake et al., 1997a;Varga Bátkai et al., 2004b). In anesthetized rats, anandamide elicits only a modest and short-lasting hypotensive response (Varga et al., 1995; Lake et al., 1997a), whereas in conscious normotensive rats it has no hypotensive effect at all (Stein et al., 1996; Lake et al., 1997b; Gardiner et al., 2002). Furthermore, inhibitors of anandamide transport or FAAH do not lower blood pressure in normotensive animals (Calignano et al., 1997; Bátkai et al., 2004b), and mice deficient in FAAH have normal baseline hemodynamic characteristics and baroreceptor reflex function (Pacher et al., 2005d). As pointed out by a recent editorial (Awumey et al., 2005), these observations indicate a lack of involvement of endogenous cannabinoids in cardiovascular regulation under normal conditions.
In contrast, a number of observations indicate that endocannabinoids are involved in cardiovascular regulation in hypertension. Both THC (Kosersky, 1978) and anandamide (Lake et al., 1997b, Bátkai et al., 2004b) induce larger and longer lasting hypotension in anesthetized SHR compared with normotensive controls, and the hypotensive effect of anandamide is preserved in conscious SHR (Lake et al., 1997b). Interestingly, inhalation of THC also resulted in a greater and longer lasting decrease of arterial blood pressure in hypertensive compared with normotensive individuals (Crawford and Merritt, 1979). By using a sophisticated pressure-volume analysis system, the hemodynamic effects of cannabinoid agonists and antagonists were evaluated in three different models of experimental hypertension (Bátkai et al., 2004b). In anesthetized SHR, the CB1 antagonists AM251 and SR141716 both caused marked and sustained further increases in blood pressure and cardiac contractility (Fig. 5). Conversely, preventing the degradation or uptake of endogenous anandamide by treatment with the FAAH inhibitor URB597 or the transport inhibitor OMDM2 reduced blood pressure, cardiac contractility, and vascular resistance to levels observed in normotensive controls, and these effects were prevented by pretreatment with a CB1 antagonist. Similar effects were seen in Dahl salt-sensitive rats and rats with angiotensin II-induced hypertension, whereas in the respective normotensive controls the same parameters remained unaffected by any of these treatments (Bátkai et al., 2004b) (Fig. 5). Anandamide and HU-210 induced more pronounced and longer lasting hypotension in SHR than in WKY rats. Unexpectedly, decreased cardiac contractility rather than a reduction in peripheral resistance was primarily responsible for the antihypertensive effect of anandamide, which was fully prevented by CB1 antagonists, but was unaffected by the TRPV1 antagonist capsazepine. In the same study, the expression of CB1 receptors was found to be increased in the myocardium and the aortic endothelium of SHR compared with WKY rats.
Fig. 5.
Effects of anandamide, URB597, SR141716, and AM251 on left ventricular (LV) function in normotensive and spontaneously hypertensive rats. Representative left ventricular pressure-volume (PV) loops from WKY rats (a, d, and g) and SHR (b, c, e, f, h, and i) before (black) and after (red) treatment with indicated agents or their combinations. A leftward shift of PV loops and an increase in amplitude (pressure) indicate increased LV contractility, whereas a rightward shift and decrease in amplitude indicate decreased LV function. Experiments were repeated in three more animals in each treatment group with similar results. AEA, anandamide. Reproduced with permission from Bátkai et al. (2004) Circulation 110:1996–2002; © Lippincott Williams and Wilkins.
These findings point to the existence of an endocannabinoid tone in hypertension that limits the elevation of blood pressure and cardiac contractile performance through tonic activation of cardiac and probably vascular CB1. A possible underlying mechanism is the observed up-regulation of cardiac and vascular CB1 in SHR compared with their normotensive controls, although increased coupling of these CB1 receptors may also contribute to the augmented sensitivity to the cardiovascular effects of anandamide (Bátkai et al., 2004b). A proposed alternative mechanism would involve up-regulation of vascular TRPV1 receptors in hypertension, based on the reported ability of capsazepine to partially inhibit the hypotensive effect of anandamide and R-methanandamide in hypertensive but not in normotensive rats (Li et al., 2003; Wang et al., 2005). However, capsazepine is known to have nonspecific effects even at low concentrations (Ray et al., 2003), and up-regulation of TRPV1 cannot account either for the increased hypotensive potency of HU-210 (Bátkai et al., 2004b), which is not a ligand for TRPV1 receptors (Zygmunt et al., 1999), or for the dominant cardiac component in the hypotensive effect of exogenous or endogenous anandamide (Bátkai et al., 2004b). Also, physiological concentrations of endogenous anandamide are at least an order of magnitude lower than the micromolar concentrations required to activate TRPV1 receptors.
A practical implication of these findings is that enhancing endocannabinoid tone by blocking the enzymatic degradation or cellular uptake of anandamide could be a novel therapeutic approach in the treatment of hypertension. Such a strategy has a number of desirable features: 1) unlike the generalized activation of CB1 receptors by direct acting agonists, inhibition of FAAH causes a more restricted profile of cannabinoid-like effects with no indication of psychoactivity (Kathuria et al., 2003; Gobbi et al., 2005), probably related to the discrete distribution of FAAH in the brain; 2) FAAH or transport inhibitors have no hemodynamic effects under normotensive conditions, which predicts the absence of postural hypotension or other side effects; and 3) having a major effect on the inappropriately increased cardiac contractility, such treatment may be effective in reversing the cardiac hypertrophy that usually accompanies chronic hypertension.
2. Circulatory Shock
The profound hypotension that can be elicited through pharmacological activation of CB1 receptors (Lake et al., 1997a) triggered numerous studies to investigate the role of the endocannabinoid system in the hypotension associated with various forms of shock, including hemorrhagic (Wagner et al., 1997; Cainazzo et al., 2002), endotoxic (Varga et al., 1998; Wang et al., 2001; Liu et al., 2003a; Bátkai et al., 2004a; Gardiner et al., 2005; Kadoi et al., 2005), and cardiogenic shock (Wagner et al., 2001a, 2003), and the shock associated with necrotizing pancreatitis (Matsuda et al., 2005). Initial studies demonstrated that the putative CB1 receptor antagonist SR141716 prevented or reversed the hypotension associated with hemorrhagic, endotoxic, and cardiogenic shock (Wagner et al., 1997, 2001a,b; Varga et al., 1998). Likewise, SR141716 reversed the hypotension associated with advanced liver cirrhosis (Bátkai et al., 2001; Ros et al., 2002), which is possibly secondary to the endotoxemia frequently found in patients with late-stage cirrhosis (Lumsden et al., 1988). Observations that circulating macrophages and platelets from endotoxemic or cirrhotic animals or humans had elevated levels of endocannabinoids and, when isolated and injected into normal rats, these cells elicited SR141716-sensitive hypotension also pointed toward the involvement of CB1 receptors in many of these conditions (Wagner et al., 1997; Varga et al., 1998; Bátkai et al., 2001; Maccarrone et al., 2001, 2002; Ros et al., 2002; Liu et al., 2003a).
Several recent reports demonstrated that anandamide and some atypical cannabinoids can cause both cardiodepressant and vasodilatory effects via as-yet-undefined receptors sensitive to inhibition by SR141716 but not by AM251 (Járai et al., 1999; Ford et al., 2002; Ho and Hiley, 2003; O’Sullivan et al., 2004b), a selective CB1 antagonist equipotent with SR141716 (Gatley et al., 1997). A recent study compared the effects of SR141716 and AM251 in rats on the acute hypotensive effect of bacterial endotoxin (LPS) administered as an intravenous bolus. Hypotension in this model is fully attributable to the decreased cardiac contractility, whereas peripheral vascular resistance is increased, indicating vasoconstriction (Biber et al., 1988; Cheng et al., 2003). Using this model, the cardiodepressant and hypotensive effects of LPS were inhibited by SR141716 but not by AM251. Furthermore, LPS induced SR141716-sensitive hypotension in wild-type mice and in mice deficient in CB1 or both CB1 and CB2 receptors, suggesting that receptors distinct from CB1 or CB2 are primarily responsible for the observed hypotension (Bátkai et al., 2004a). Interestingly, another recent study has demonstrated that the CB1-selective cannabinoid antagonist AM281 prevented the hemodynamic changes induced by acute LPS injection in rats (Kadoi et al., 2005a). Other results indicate that endocannabinoids may also contribute to endotoxin-induced hypotension indirectly, through CB1-mediated prejunctional inhibition of sympathoexcitation (Godlewski et al., 2004). In a different shock model in which continuous infusion of LPS in conscious rats causes marked peripheral vasodilatation and increased cardiac output, AM251 attenuated the tachycardic and hind quarter vasodilator effects of LPS. This result was attributed to modulation of β-adrenergic vasodilation, rather than suppression of a direct vasodilator effect by endocannabinoids (Gardiner et al., 2005). Interestingly, in a recent study, Matsuda et al. (2005) demonstrated that AM251 improved mean arterial pressure and survival rate in models of severe acute necrotizing pancreatitis without affecting inflammatory changes, which suggests the involvement of cardiac or vascular CB1 receptors in the hypotension associated with this condition.
In hemorrhagic, cardiogenic, and endotoxic shock, the cannabinoid agonists HU-210, WIN 55,212-2, and THC improved endothelial function and/or survival (Wagner et al., 1997, 2001a, 2003; Varga et al., 1998; Smith et al., 2000, 2001). Surprisingly, the use of cannabinoid receptor antagonists, including SR141716, AM281, AM251, and SR144528, also leads to survival benefits in endotoxic and septic shock or necrotizing pancreatitis (Varga et al., 1998; Smith et al., 2000, 2001; Cainazzo et al., 2002; Kadoi et al., 2005a,b; Matsuda et al., 2005). In contrast, CB1 receptor blockade increased mortality in hemorrhagic (Wagner et al., 1997) and cardiogenic shock (Wagner et al., 2001a, 2003), despite the increase in blood pressure. In these latter conditions, endocannabinoid-mediated vasodilation may have survival value through improving tissue oxygenation by counteracting the excessive sympathetic vasoconstriction triggered by hemorrhage or myocardial infarction, and this would be removed by CB1 blockade. In contrast, CB1 blockade may improve survival in endotoxic shock by preventing the primary hypotensive response to LPS (reviewed in Kunos et al., 2000; Hiley and Ford, 2003, 2004; Pacher et al., 2005a,c).
It should also be kept in mind that in most of the above conditions, hemodynamic changes are triggered by overwhelming inflammatory reaction, increased oxidative stress, and activation of downstream effector pathways, eventually leading to cardiovascular dysfunction and failure (reviewed in Evgenov and Liaudet, 2005; Pacher et al., 2005e). Therefore, the well known immune-modulatory, anti-inflammatory, and antioxidant effects of cannabinoids should not be overlooked in these conditions. Indeed, endocannabinoids and synthetic cannabinoid agonists decrease inflammatory cytokine release in endotoxin-stimulated cells and in endotoxin-challenged animals (reviewed in Walter and Stella, 2004; Klein et al., 2005). Surprisingly, SR141716 and the CB2 antagonist SR144528 were also reported to have anti-inflammatory effects (Smith et al., 2000, 2001), which may be attributed to their inverse agonist properties or to CB1/2 receptor-independent mechanisms (reviewed in Begg et al., 2005; Pertwee, 2005b,c).
Collectively, it appears that both cannabinoids and antagonists of cannabinoid receptors may exert some beneficial effects in various rodent shock models. Further studies should establish the specificity of these effects and the relevance to various forms of circulatory shock in humans.
3. Myocardial Reperfusion Injury
The endocannabinoid system has been implicated in endotoxin-induced preconditioning against myocardial I/R injury (Lagneux and Lamontagne, 2001). In this study, the effects of 90 min of low-flow ischemia followed by 60 min of reperfusion at normal flow were compared in isolated hearts from rats pretreated with LPS or saline. Endotoxin pretreatment enhanced functional recovery on reperfusion and reduced infarct size compared with controls, and pretreatment with the CB2 antagonist SR144528 but not the CB1 antagonist SR141716 abolished the beneficial effects of preconditioning (Lagneux and Lamontagne, 2001). In a follow-up study, SR144528 but not SR141716 also abolished the infarct size-reducing effect of preconditioning induced by heat stress (Joyeux et al., 2002). These initial studies have suggested that the protection was mediated by endocannabinoids acting on CB2 receptors. In preconditioning induced by a brief period of ischemia (5 min), either CB2 or CB1 receptor blockade abolished the protection, and both CB1 and CB2 receptors were implicated in the preservation of endothelium-dependent, 5-HT-induced vasodilation by ischemic preconditioning (Bouchard et al., 2003). Perfusion of isolated rat hearts with PEA or 2-AG but not anandamide afforded protection against ischemia by improving myocardial recovery and decreasing myocardial damage and infarct size (Lepicier et al., 2003). The cardioprotective effect of both PEA and 2-AG were completely blocked by SR144528, whereas SR141716 partially inhibited the effect of 2-AG only (Lepicier et al., 2003). Likewise, the selective CB1 agonist ACEA and the selective CB2 agonist JWH-015 both reduced infarct size in this model, and the CB2 receptor-mediated cardioprotection by PEA involved activation of p38/extracellular signal-regulated kinases 1 and 2 and protein kinase C (Lepicier et al., 2003). In another study using isolated perfused rat hearts subjected to ischemia and reperfusion, reduction of the infarct size by anandamide could be equally well antagonized by CB1 or CB2 antagonists but could not be mimicked by selective CB1 or CB2 agonists, suggesting the involvement of a site distinct from CB1 or CB2 receptors (Underdown et al., 2005).
Others have used whole animal models of I/R injury induced by coronary occlusion/reocclusion in anesthetized rats. In this model, anandamide and HU-210 both decreased the incidence of ventricular arrhythmias and reduced infarct size through activation of CB2 but not CB1 receptors (Krylatov et al., 2001, 2002a,b,c; Ugdyzhekova et al., 2001, 2002). The moderately CB2-selective agonist WIN 55,212-2 also reduced the extent of leukocyte-dependent myocardial damage in a more recent mouse study of myocardial I/R in vivo. This effect was abolished by the selective CB2 receptor antagonist AM630 but was unaffected by AM251 (Di Filippo et al., 2004). In summary, evidence to date indicates that endocannabinoids protect against myocardial ischemic injury models predominantly via CB2 receptors.
4. Atherosclerosis
Chronic inflammation and the associated oxidative-nitrosative stress are key players in atherosclerosis and cardiovascular aging, and pharmacological modulation of these processes could be of therapeutic benefit (reviewed in Csiszar et al., 2005; Libby and Theroux, 2005). Using the apolipoprotein E knockout mouse model of atherosclerosis, Steffens et al. (2005) reported that orally administered THC significantly inhibited disease progression. Furthermore, CB2 receptor expressing immune cells were present both in human and mouse atherosclerotic plaques, lymphoid cells isolated from THC-treated mice had diminished proliferation capacity and decreased interferon-γ production, and THC inhibited macrophage chemotaxis in vitro. Most importantly, all of these effects were completely blocked by a selective CB2 receptor antagonist, suggesting that targeting CB2 receptors may offer a new approach in the treatment of atherosclerosis (Roth, 2005; Steffens et al., 2005).
5. Asthma
The effect of marijuana on airway functions was among the first to be explored for potential therapeutic benefit (reviewed in Lemberger, 1980; Tashkin et al., 2002). Smoking marijuana and ingesting THC were both found to increase airway conductance in normal, healthy subjects (Tashkin et al., 1973; Vachon et al., 1973), and these effects lasted longer than the bronchodilator effect of the β-adrenergic agonist isoproterenol. Bronchodilation induced by smoked marijuana and oral THC was also documented in subjects with mild to moderate asthma and in asthmatic patients with methacholine- or exercise-induced bronchoconstriction (Tashkin et al., 1974, 1975). Bronchodilation without side effects was observed in asthmatic patients after a low dose (0.2 mg) of nebulized THC (Williams et al., 1976; Hartley et al., 1978). In contrast, aerosols containing larger doses of THC (5–20 mg) caused paradoxical bronchoconstriction attributed to local irritation (Tashkin et al., 1977). In another study of normal and asthmatic subjects, orally administered THC elicited only minimal and inconsistent bronchodilation associated with significant CNS side effects (Abboud and Sanders, 1976). Nevertheless, most of these initial observations had suggested some therapeutic benefit of using cannabinoids in asthma.
As for the mechanisms underlying THC-induced bronchodilation, the potential involvement of β-adrenergic and muscarinic receptors on airway smooth muscle could be excluded (Kelly and Butcher, 1973; Shapiro et al., 1977; Lemberger, 1980). This conclusion was supported by the inability of THC to relax isolated rings of resting or precontracted human bronchioles (Orzelek-O’Neil et al., 1980a,b), suggesting a more proximal site of action in the lung (Cavero et al., 1972) or a central mechanism.
More recently, Calignano et al. (2000) reported that CB1 receptors are present on axon terminals innervating airway smooth muscle, and anandamide inhibited capsaicin-induced bronchospasm and cough in guinea pigs in an SR141716-sensitive manner. They also documented calcium-induced biosynthesis of anandamide in lung tissue, suggesting that locally generated anandamide participates in the intrinsic control of airway responsiveness by inhibiting prejunctional acetylcholine release. Indeed, SR141716 treatment was found to enhance capsaicin-evoked bronchospasm and cough. Interestingly, when airway smooth muscle was completely relaxed by vagotomy and atropine treatment, anandamide caused dose-dependent bronchoconstriction, which could be also prevented by CB1 blockade. This effect was tentatively attributed to direct stimulation of putative cannabinoid receptors on the airway smooth muscle or a CB1-mediated corelease of bronchoconstrictor neurotransmitters from nerve endings in the lung. In a follow-up study, presynaptic CB1 receptors in the guinea pig lung were only found on noradrenergic terminals where their stimulation by WIN 55,212-2 inhibited norepinephrine release (Vizi et al., 2001), consistent with the lack of a mediated CB1-mediated effect on acetylcholine release in guinea pig trachea (Spicuzza et al., 2000). In contrast to the findings of Calignano et al. (2000), Stengel et al. (1998) reported that anandamide given either intravenously or in aerosol did not affect airway resistance in guinea pigs, but possessed modest anti-inflammatory properties. It should be noted, however, that in this study bronchoconstriction was induced by a calcium ionophore rather than capsaicin. In an in vitro study of guinea pig airway smooth muscle (Yoshihara et al., 2005), anandamide and palmitoylethanolamide inhibited contractions elicited by electrical field stimulation but not by neurokinin A, and also blocked capsaicin-capsaicin-induced release of substance P-like immunoreactivity. These effects were selectively inhibited by a CB2 but not a CB1 antagonist, or by maxi-K+ channel blockers, suggesting that CB2 agonists may have therapeutic value in asthma (Yoshihara et al., 2005). In a recent study, inhibition of anandamide transport potently suppressed capsaicin-induced cough in mice, suggesting that the anandamide transporter may be a target for peripherally acting antitussive medications (Kamei et al., 2006). Diverse effects of endocannabinoids and synthetic agonist have also been reported on respiratory function and pulmonary circulation both in vivo and in vitro (Schmid et al., 2003; Wahn et al., 2005).
Allergic asthma is currently viewed as a complex inflammatory disorder characterized by recruitment of eosinophils into the lung, mucus hypersecretion by goblet cells, elevated serum IgE, and airway hyperresponsiveness (reviewed in Wills-Karp, 1999). Given the well known anti-inflammatory effects of cannabinoids, these effects could also be of therapeutic value. Indeed, in a murine model of allergic airway disease induced by ovalbumin sensitization, pretreatment with cannabinol or THC blunted the increase in IL-2, IL-4, IL-5, and IL-13 mRNA expression and decreased mucus overproduction and serum IgE levels (Jan et al., 2003). Anti-inflammatory effects of WIN 55,212-2, THC, anandamide, and palmitoylethanolamide were also reported in a mouse model of LPS-induced pulmonary inflammation (Berdyshev et al., 1998).
In conclusion, the effects of cannabinoids on respiratory function are rather complex, and evidence for their therapeutic potential in asthma is equivocal. The possibility remains that novel, nonpsychoactive cannabinoid analogs with long-lasting anti-inflammatory activity turn out to be useful adjuncts in the treatment of allergic asthma.
E. Eye Disorders (Glaucoma and Retinopathy)
Glaucoma, the leading cause of irreversible blindness in the United States, is characterized by an increase in intraocular pressure and consequent damage to the optic nerve. Despite the multitude of effective medications that can be used to decrease ocular hypertension (e.g., cholinergic agonists, β- and α2-adrenoceptor agonists, dopaminergic agonists, prostaglandins, and carbonic anhydrase inhibitors), some patients remain refractory to these drugs and may eventually become blind (reviewed in Alward, 1998; Crowston and Weinreb, 2005).
A decrease in intraocular pressure in a small number of healthy marijuana smokers was a serendipitous finding (Hepler and Frank, 1971), subsequently confirmed in a placebo-controlled, double-blind study of healthy volunteers who smoked either natural marijuana of known THC content or ingested synthetic THC (Hepler et al., 1972). THC or marijuana decreased intraocular pressure whether administered orally, topically, or intravenously, with no major tolerance to their effect reported (Shapiro, 1974; Purnell and Gregg, 1975; Cuendet et al., 1976; Hepler et al., 1976; Brown et al., 1977; Merritt et al., 1980, 1981a,b). Most of these studies also reported various systemic side effects, such as hypotension, tachycardia, euphoria, and dysphoria, as well as other ocular effects, such as changes in pupil size, decreased tear production, and conjunctival hyperemia. Endocannabinoids and synthetic cannabinoid ligands have also been reported to reduce intraocular pressure when given topically or systemically, both in animals and humans (Shapiro, 1974; ElSohly et al., 1981, 1984; Colasanti et al., 1984a,b,c; Pate et al., 1995; Porcella et al., 1998; Buchwald et al., 2002; Laine et al., 2002a,b; reviewed in Jarvinen et al., 2002; Chien et al., 2003; Tomida et al., 2004).
Early investigations into the mechanisms of the intraocular pressure-lowering effect of marijuana and THC implicated the sympathetic and central nervous systems in this effect (Green and Pederson, 1973; Green and Podos, 1974; Green et al., 1977a,b). However, in subsequent studies, the effect of a unilateral topical application of cannabinoids was limited to the treated eye, pointing toward a local site of action (Colasanti et al., 1984a,b,c). Indeed, a CNS site of action could be ruled out by the lack of change in intraocular pressure upon intracerebroventricular or ventriculocisternal application of THC in rabbits (Liu and Dacus, 1987).
Multiple lines of evidence suggest that endocannabinoids and cannabinoid receptors, in particular CB1, play an important role in the regulation of intraocular pressure, and topically applied cannabinoids and cannabinoid ligands may be of significant benefit in the treatment of glaucoma (reviewed in Jarvinen et al., 2002; Tomida et al., 2004). First, CB1 receptors are expressed in the rat ciliary body (Porcella et al., 1998), in human ciliary epithelium, ciliary muscle, ciliary body vessels, trabecular meshwork, Schlemm’s canal, and retina (Straiker et al., 1999a; Porcella et al., 2000; Stamer et al., 2001), and the retina of a variety of species (Straiker et al., 1999b; Yazulla et al., 1999, 2000). Second, ocular CB1 receptors are functionally active, as CB1 receptor agonists (CP55,940 and WIN 55,212-2) applied topically lower intraocular pressure both in animals and humans, and their effect can be antagonized by SR141716 (Pate et al., 1998; Song and Slowey, 2000; Porcella et al., 2001; Chien et al., 2003; Stumpff et al., 2005; reviewed in Jarvinen et al., 2002). The CB2 receptor agonist JWH-133 did not modify the intraocular pressure, suggesting that CB2 receptors may play only a minor, if any, role (Laine et al., 2003). CB1 receptor signaling is also operational in the ocular trabecular meshwork (Stumpff et al., 2005), and ciliary muscle (Lograno and Romano, 2004). Third, endocannabinoids are detectable in ocular tissues including the retina, ciliary body, and choroids plexus (Bisogno et al., 1999b; Straiker et al., 1999a,b; Stamer et al., 2001; Chen et al., 2005), and the levels of anandamide and especially 2-AG are significantly decreased in patients with glaucoma (Chen et al., 2005).
The cellular/molecular mechanisms responsible for the intraocular pressure-reducing effect of cannabinoids are not completely understood but may involve direct effects on ciliary processes such as vasodilation and reduction of capillary pressure and secretion and do not seem to be related to systemic reduction of arterial blood pressure (Green and Pederson, 1973; Korczyn, 1980). Cannabinoids may also inhibit calcium influx through presynaptic ion channels, thereby reducing norepinephrine release in the ciliary body, resulting in decreased aqueous humor production (Sugrue, 1997). In addition, cannabinoids may improve the uveoscleral outflow by dilating blood vessels of the anterior uvea (Porcella et al., 1998), most likely by induction of several outflow-facilitating mediators (Rosch et al., 2006). Some evidence implicates prostanoids in the intraocular pressure-reducing effect of certain cannabinoids and anandamide (Pate et al., 1996; Green et al., 2001; Rosch et al., 2006).
Endocannabinoids as well as functional CB1 receptors are present in the retina (Bisogno et al., 1999b; Straiker et al., 1999a,b; Fan and Yazulla, 2003; Savinainen and Laitinen, 2004). Cannabinoids exert neuroprotective effects against retinal neurotoxicity (El-Remessy et al., 2003), and cannabidiol helps to preserve the blood-retinal barrier in experimental diabetes (El-Remessy et al., 2006). These effects could suggest their usefulness in various retinopathies. Unlike CB1 receptors, CB2 receptors were undetectable in human retina, although they were found in the rat retina (Lu et al., 2000; Porcella et al., 2000).
Taken together, these findings indicate that cannabinoids may have great potential in the treatment of glaucoma, if the difficulty in formulating a stable and effective topical preparation and the problem of systemic side effects are conquered. Because of their well known neuroprotective, anti-inflammatory, and antiangiogenic effects, cannabinoid analogs may also be considered for the treatment of inflammatory and degenerative eye disorders and diabetic retinopathy.
F. Cancer
The palliative effects of cannabinoids in cancer patients are well known and may include appetite stimulation, inhibition of nausea and emesis associated with chemo- or radiotherapy, pain relief, mood elevation, and relief from insomnia (reviewed in Walsh et al., 2003; Hall et al., 2005) (Table 1). Δ9-THC (dronabinol, Marinol) and its synthetic derivative nabilone have been approved by the U.S. Food and Drug Administration to control nausea in cancer patients undergoing chemotherapy and to stimulate appetite in patients with AIDS (Walsh et al., 2003; Hall et al., 2005).
Numerous recent studies have suggested that cannabinoids might directly inhibit cancer growth (reviewed in Parolaro et al., 2002; Guzmán et al., 2002; Guzmán, 2003; Jones and Howl, 2003; Velasco et al., 2004; Patsos et al., 2005). The proposed mechanisms are complex and may involve induction of apoptosis in tumor cells, antiproliferative action, and an antimetastatic effect through inhibition of angiogenesis and tumor cell migration (reviewed in Bifulco and Di Marzo, 2002; Parolaro et al., 2002; Guzmán et al., 2002; Guzmán, 2003; Jones and Howl, 2003; Velasco et al., 2004; Patsos et al., 2005).
Various cannabinoids, including cannabidiol, anandamide, and 2-AG, and endocannabinoid transport inhibitors have been shown to induce apoptotic cell death and to inhibit proliferation and migration in numerous murine and human tumor cell lines including glioma (C6, U87, U373, and H4), oligodendroglioma (Gos3), glioblastoma multiforme, astrocytoma (U373-MG, U87MG, and human grade IV astrocytoma), neuroblastoma (N18 TG2 and CHP100), pheochromocytoma (PC12), breast cancer (MCF-7, EFM-19, T47D, TSA-E1, and MDA-MB-231), prostate cancer (LNCaP, DU145, and PC3), colon carcinoma (SW 480), uterine cervix carcinoma (CxCa), thyroid cancer (KiMol), leukemia (CEM, HEL-92, HL60, and Jurkat cell lines), and lymphoid tumors (EL-4 and P815) tumor cells via CB1/CB2- and VR1 receptor-dependent or independent (e.g., cyclooxygenase) mechanisms (De Petrocellis et al., 1998; Sánchez et al., 1998, 2003; Jacobsson et al., 2000; Maccarrone et al., 2000b; Sarker et al., 2000; McKallip et al., 2002a,b; Fowler et al., 2003; Jonsson et al., 2003; Mimeault et al., 2003; Bifulco et al., 2004; Contassot et al., 2004a,b; Hinz et al., 2004; Joseph et al., 2004; Kogan et al., 2004; Massi et al., 2004; Nithipatikom et al., 2004; Allister et al., 2005; Ellert-Miklaszewska et al., 2005; Herrera et al., 2005, 2006; Lombard et al., 2005; Powles et al., 2005; Sarfaraz et al., 2005; Vaccani et al., 2005; Carracedo et al., 2006; Grimaldi et al., 2006; Ligresti et al., 2006b). More importantly, systemic or local treatment with cannabinoids inhibited the growth of various types of tumor or tumor cell xenografts in vivo, including lung carcinoma (Munson et al., 1975), glioma (Galve-Roperh et al., 2000; Sánchez et al., 2001a; Massi et al., 2004), thyroid epithelioma (Bifulco et al., 2001), lymphoma (McKallip et al., 2002a), and skin carcinoma (Casanova et al., 2003) in mice.
The proapoptotic effect of cannabinoids in tumor cells is complex and may involve increased synthesis of the proapoptotic sphingolipid ceramide (Galve-Roperh et al., 2000; Gómez del Pulgar et al., 2002a,b), ceramide-dependent up-regulation of the stress protein p8 and several downstream stress-related genes expressed in the endoplasmic reticulum (ATF-4, CHOP, and TRB3; Carracedo et al., 2006), prolonged activation of the Raf-1/mitogen-activated protein kinase kinase/extracellular signal-regulated kinase signaling cascade (Galve-Roperh et al., 2000), and inhibition of Akt (Gómez del Pulgar et al., 2000; Ellert-Miklaszewska et al., 2005), c-Jun NH2-terminal kinase and p38 mitogen-activated protein kinase (Galve-Roperh et al., 2000; Sarker et al., 2003; Hinz et al., 2004; Powles et al., 2005). As mentioned above, cannabinoids also inhibit the proliferation of various tumor cells, possibly through inhibition of adenylyl cyclase and the cAMP/protein kinase A pathway (Melck et al., 1999), induction of the cyclin-dependent kinase inhibitor p27kip1 (Portella et al., 2003), a decrease in epidermal growth factor receptor expression and/or the attenuation of epidermal growth factor receptor tyrosine kinase activity (Casanova et al., 2003; Mimeault et al., 2003), and a decrease in the activity and/or expression of nerve growth factor or vascular endothelial growth factor tyrosine kinase receptors and prolactin (De Petrocellis et al., 1998; Melck et al., 2000; Portella et al., 2003). In addition to their proapoptotic and antiproliferative effects in tumor cells, cannabinoids also inhibit the expression of proangiogenic mediators or their receptors (e.g., vascular endothelial growth factor) and reduce vascular hyperplasia and cell migration, which play crucial roles in tumor growth and metastasis formation (Blázquez 2004;et al., 2003, 2004; Casanova et al., 2003; Portella et al., 2003).
In sharp contrast to the above, Hart et al. (2004) have demonstrated that treatment of lung cancer (NCI-H292), squamous cell carcinoma (SCC-9), bladder carcinoma (5637), glioblastoma (U373-MG), astrocytoma (1321N1), and kidney cancer (A498) cells with nanomolar concentrations of cannabinoids such as THC, anandamide, HU-210, and WIN 55,212-2 leads to rapid epidermal growth factor receptor- and metalloprotease-dependent cancer cell proliferation. However, the same study also documented that at micromolar concentrations cannabinoids induced cancer cell apoptosis, in agreement with previous reports (Hart et al., 2004). These results highlight the bimodal action of cannabinoids on cancer cell growth, with low concentrations being proproliferative and high concentrations having antiproliferative effects.
The key role of the immune system in controlling the development of cancers is supported by findings that immunosuppressed individuals are at increased risk for developing cancer. For example, there is increased incidence of non-Hodgkin’s lymphoma, Burkitt’s lymphoma, Kaposi’s sarcoma, and cervical cancer in AIDS patients and increased susceptibility to various lymphomas and solid tumors after organ transplantation (Bhatia et al., 2001; Scadden, 2003; Abu-Elmagd et al., 2004; Oruc et al., 2004). This concept is particularly important, because cannabinoids have well-known immunosuppressant effects (reviewed in Klein, 2005), which may compromise antitumor immune responses. Indeed, THC enhances breast and lung cancer growth and metastasis by suppressing CB2 receptor-mediated antitumor immune responses (Zhu et al., 2000; McKallip et al., 2005) and can also lead to increased susceptibility to infections with various pathogens such as herpes simplex virus, Legionella pneumophila, and Fried leukemia virus (Morahan et al., 1979; Cabral et al., 1986; Specter et al., 1991; Klein et al., 2000b).
Epidemiological studies investigating the relationship of cannabis smoking and various forms of cancer have yielded inconsistent results, thus failing to resolve the conflicting findings in animal models of cancer or in cancer cell lines (Taylor, 1988; Caplan and Brigham, 1990; Kuijten et al., 1992; Grufferman et al., 1993; Sidney et al., 1997; Barsky et al., 1998; Zhang et al., 1999; Efird et al., 2004; Llewellyn et al., 2004; Rosenblatt et al., 2004; reviewed in Hall et al., 2005). The variability of the effects of cannabinoids in different tumor models may be related to the differential expression of CB1 and CB2 receptors. Thus, cannabinoids may be effective in killing tumors that abundantly express cannabinoid receptors, such as gliomas, but may increase the growth and metastasis of other types of tumors, such as breast cancer, with no or low expression of cannabinoid receptors, due to the suppression of the antitumor immune response (McKallip et al., 2005). Nevertheless, the majority of the findings to date are encouraging and suggest that cannabinoids may be useful not only as palliative therapy but also because of their ability to inhibit tumor growth and metastasis.
G. Gastrointestinal and Liver Disorders
Cannabis has been used empirically for centuries to stimulate appetite and decrease emesis and diarrhea. Recent evidence indicates that the endocannabinoid system plays an important role in the control of gastrointestinal motility and secretion both under physiological conditions and in various gastrointestinal disorders (reviewed in Pertwee, 2001; Pinto et al., 2002a,b; Di Carlo and Izzo, 2003; Coutts and Izzo, 2004; Duncan et al., 2005; Massa et al., 2005). Unexpectedly, recent data also implicate endocannabinoids and their receptors in several aspects of acute and chronic liver disease, including hemodynamic changes, modulation of inflammatory processes, fibrosis, and altered brain function (reviewed in Gabbay et al., 2005; Jimenez, 2005).
Numerous studies using autoradiography, immunohistochemistry, and/or reverse transcription-polymerase chain reaction demonstrated colocalization of CB1 receptors with cholinergic neurons across the enteric nervous system, including sensory and interneuronal as well as motoneural cell bodies of the myenteric plexus, in mice (Mascolo et al., 2002; Pinto et al., 2002a,b; Casu et al., 2003; Izzo et al., 2003; Storr et al., 2004), rats (Adami et al., 2002; Coutts et al., 2002; Storr et al., 2002; Burdyga et al., 2004), guinea-pigs (Coutts et al., 2002; MacNaughton et al., 2004), and pigs (Kulkarni-Narla and Brown, 2000). CB1 receptors are also colocalized with neuropeptide Y and vasoactive intestinal peptide in a small population of submucous plexus neurons (Kulkarni-Narla and Brown, 2000; Coutts et al., 2002). CB1 receptor immunoreactivity was evident in normal human colonic epithelium, smooth muscle, and the sub-mucosal myenteric plexus (Wright et al., 2005). Both CB1 and CB2 receptors were found on plasma cells in the lamina propria, whereas only CB2 were detectable on macrophages (Wright et al., 2005). Endocannabinoids are also present in the gastrointestinal tact. Indeed, 2-AG was originally isolated from gut tissue (Mechoulam et al., 1995), and the intestinal content of anandamide was found to be regulated by feeding status (Gomez et al., 2002).
Although in earlier studies CB1 receptor expression was undetectable in the liver relative to the brain (Porcella et al., 2002), several recent studies revealed the presence of low levels of both CB1 mRNA (Bátkai et al., 2001; Michalopoulos et al., 2003; Biecker et al., 2004; Engeli et al., 2005; Osei-Hyiaman et al., 2005b; Teixeira-Clerc et al., 2006) and CB1 immunoreactivity (Osei-Hyiaman et al., 2005b) in whole liver or in various types of cells present in the liver, including hepatocytes (Michalopoulos et al., 2003; Osei-Hyiaman et al., 2005b), stellate cells (Siegmund et al., 2005; Teixeira-Clerc et al., 2006), and vascular endothelial cells (Bátkai et al., 2001). CB2 receptor mRNA was also detected in cirrhotic but not in normal liver tissue (Julien et al., 2005). Endocannabinoids are detectable in the liver or liver cells both in animals and humans at levels similar to those in the brain and play an important role under various physiological and pathophysiological conditions (Cravatt et al., 2004; Kurabayashi et al., 2005; Osei-Hyiaman et al., 2005b) (see also section III.A.3.).
A functional role for endocannabinoids and CB1 receptors in the gastrointestinal tract is supported by pharmacological studies demonstrating that anandamide and various CB1 agonists (WIN 55,212-2, CP55,940, and ACEA) but not the CB2-selective agonists JWH-133 inhibit gastrointestinal motility in rodents in vivo and in isolated ileum and colon from both experimental animals and humans (Shook and Burks, 1989; Pertwee et al., 1995, 1996; Coutts and Pertwee, 1997; McCallum et al., 1999; Mancinelli et al., 2001; Mang et al., 2001; Landi et al., 2002; Manara et al., 2002; Hinds et al., 2006). A similar role for endogenous substrates of FAAH is suggested by recent in vivo findings in mice, documenting inhibition of intestinal motility by the FAAH inhibitors N-arachidonoylserotonin and palmitoyliso-propylamide and by the FAAH substrates palmitoylethanolamide, oleamide, and oleoylethanolamide in wild-type but not in FAAH knockout mice (Capasso et al., 2005). Furthermore, the effect of N-arachidonoylserotonin was reduced either by the CB1 receptor antagonist SR141716 or by CB1 deficiency, but not by the TRPV1 receptor antagonist 5′-iodoresiniferatoxin (Capasso et al., 2005). Interestingly, in clinical trials using rimonabant for nicotine cessation or for the treatment of obesity, diarrhea was 2 to 2.4 times more frequent among subjects treated with the drug than with placebo, suggesting accelerated transit and/or enhanced secretion caused by CB1 blockade (Fernandez and Allison, 2004; Van Gaal et al., 2005). This and some of the above experimental reports suggest the existence of an inhibitory endocannabinoid tone in the gastrointestinal tract. Multiple mechanisms, including reduction of acetylcholine release from enteric nerves, inhibition of nonadrenergic/noncholinergic excitatory transmission, activation of apamin-sensitive K+ channels, and modulation of adenosine release have been proposed to explain the CB1-mediated reduction in enteric contractility and peristalsis (reviewed in Coutts and Izzo, 2004).
Activation of both CB1 and CB2 receptors may decrease the pathologically increased intestinal motility elicited by an inflammatory stimulus. In a mouse model of croton oil-induced intestinal inflammation, the increased efficacy of cannabinoids in inhibiting intestinal motility was attributed to up-regulation of intestinal CB1 receptors (Izzo et al., 2001a,b). Conversely, the accelerated gastrointestinal transit induced by bacterial endotoxin in rats could be inhibited by CB2 but not CB1 receptor agonists (Mathison et al., 2004). Interestingly, intestinal hypomotility in a mouse model of paralytic ileus has been linked, at least in part, to the enhancement of anandamide levels and CB1 expression in the gut, and it could be attenuated by CB1 receptor antagonists (Mascolo et al., 2002). Additionally, there is evidence that CB1 receptors are involved in the regulation of the lower esophageal sphincter, and CB1 activation might be beneficial in gastroesophageal reflux disease (reviewed in Coutts and Izzo, 2004; Massa et al., 2005).
The endocannabinoid system has also been implicated in the regulation of gastric acid and intestinal secretions. At high doses, THC decreased histamine-induced gastric acid secretion in isolated stomach preparations (Rivas-V and Garcia, 1980) and in pylorus-ligated rats (Sofia et al., 1978). Pentagastrin-induced gastric acid secretion was also inhibited by HU-210 and WIN 55,212-2, an effect that could be prevented by CB1 blockade (Coruzzi et al., 1999; Adami et al., 2002). These studies suggest a role for CB1 receptors located on preganglionic and postganglionic cholinergic pathways in the regulation of gastric acid secretion. The therapeutic relevance of this regulatory mechanism was highlighted by the CB1 receptor-mediated antiulcer activity of ACEA or WIN 55,212-2 treatment in a rat model of aspirin- and cold/restraint stress-induced gastric ulcers (Germano et al., 2001; Rutkowska and Fereniec-Goltbiewska, 2006). WIN 55,212-2 also reduced intestinal secretions evoked by electrical field stimulation or capsaicin (MacNaughton et al., 2004). Anandamide, the anandamide transport inhibitor VDM11, and the CB1 agonist ACEA all inhibited intestinal secretion and fluid accumulation in mice treated with cholera toxin, whereas SR141716 exerted opposite effects (Izzo et al., 2003). The ability of cannabinoids to inhibit gastrointestinal motility and secretion coupled with their anti-inflammatory properties strongly suggests that the modulation of this system could offer significant benefits in the treatment of various gastrointestinal pathological conditions, including inflammatory bowel disease (see below).
1. Inflammatory Bowel Disease
Idiopathic inflammatory bowel disease (IBD) includes ulcerative colitis and Crohn’s disease, and is characterized by intestinal inflammation presumably of autoimmune origin and a chronic relapsing course associated with local and systemic complications and affects >1 million people in the United States (Loftus, 2004). Although the etiology of IBD remains unclear, it may involve complex genetic, environmental, and immunological interactions. The most common symptoms of IBD are abdominal pain and diarrhea, which eventually lead to malabsorption and malnutrition, and in approximately half of patients surgery is eventually required to remove the affected bowel segment. Despite recent therapeutic advances, patients with IBD are often unresponsive to available treatment options.
As discussed above, the endocannabinoid system plays an important role in the control of gastrointestinal motility and secretion. Studies using animal models of IBD have suggested that targeting the endocannabinoid system may offer significant benefits in the treatment of IBD. Several studies have indicated that chemically induced intestinal inflammation is associated with the up-regulation of intestinal CB1 receptors, which may represent a compensatory, protective mechanism. For example, in croton oil-treated mice, the ability of CB1 agonists to inhibit intestinal motility is increased compared with that in control animals (Izzo et al., 2001a). More importantly, the anandamide transport inhibitor VDM11 was also shown to inhibit gastrointestinal motility and secretions in cholera toxin-treated mice, which implicates endocannabinoids in this action and holds out the promise of a nonpsychoactive form of treatment (Izzo et al., 2003). In a mouse model of colitis induced by 2,4-dinitrobenzene sulfonic acid and dextrane sulfate, Massa et al. (2004) have confirmed the up-regulation of CB1 receptors in experimental colitis. Furthermore, they demonstrated that the inflammation was more severe in mice deficient in CB1 receptors than in wild-type mice, whereas genetic ablation of FAAH resulted in protection against this chemically induced colitis (Massa et al., 2004). In a recent study, the anandamide reuptake inhibitor VDM11 afforded protection against colitis in mice, and elevated anandamide levels have been measured in biopsy material from patients with ulcerative colitis (D’Argenio et al., 2006). These findings strongly support the natural protective role of the endocannabinoid system in this form of experimental IBD. In contrast, Croci et al. (2003) have reported a CB1 receptor-independent protective effect of SR141716 against indomethacin-induced inflammation and ulcer formation in the small intestine of rats. Elevated levels of anandamide and desensitization of the presynaptic neural CB1 receptor found in colonic longitudinal muscle strips from patients undergoing surgery for complicated diverticulitis suggest that the endocannabinoid system may be also involved in the pathophysiology of this frequent complication of colitis and/or colon cancer (Guagnini et al., 2006).
Taken together, most of the above studies suggest that the endocannabinoid system in the gut is activated during inflammation, and endogenous anandamide may counteract inflammation (Kunos and Pacher, 2004) (Fig. 6). The findings of Massa et al. (2004) and D’Argenio et al. (2006) also suggest that inhibitors of FAAH or anandamide reuptake may amplify the natural protective action of endogenous anandamide, which warrants further studies to test such inhibitors in the treatment of experimental and, ultimately, human IBD (Kunos and Pacher, 2004). Future studies should further explore the mechanisms of the anti-inflammatory effects of cannabinoids and the potential role of CB2 receptors as therapeutic targets (Mathison et al., 2004; Wright et al., 2005).
Fig. 6.
Cellular source and proposed targets of anti-inflammatory endocannabinoids in inflammatory bowel disease. a, cross-section of inflamed bowel with leukocyte infiltration [polymorphonuclear leukocytes (PNM), lymphocytes (Ly), macrophages, and mast cells]. b, in macrophages, LPS induces the production of TNF-α and chemokines (such as MIP-2macrophage inflammatory protein-2 and CXCL-8) as well as anandamide. Anandamide is released to act as an autocrine mediator to inhibit TNF-α and chemokine production via CB1 or CB2 receptors or both. Activation of CB1 and CB2 receptors may similarly inhibit TNF-α production in mast cells, with these effects resulting in decreased leukocyte infiltration and inflammation. Paracrine activation of CB1 receptors on extrinsic and intrinsic enteric neurons inhibits acetylcholine (ACh) and tachykinin release, respectively, resulting in inhibition of gut motility. These effects are amplified by treatment with a FAAH inhibitor, which prevents the breakdown of anandamide. Reproduced with permission from Kunos and Pacher (2004) Nat Med 10:678–679. © Nature Publishing Group.
2. Acute and Chronic Liver Disease (Hepatitis and Liver Cirrhosis)
Endocannabinoids and CB1 receptors have been implicated in the systemic and portal vasodilation and hypotension associated with chronic liver cirrhosis (Bátkai et al., 2001; Garcia et al., 2001; Ros et al., 2002). These studies demonstrated that CB1 receptor blockade with SR141716 reversed the hypotension and low peripheral resistance and decreased the elevated mesenteric blood flow and portal pressure in rats with biliary and carbon tetrachloride-induced cirrhosis, whereas these hemodynamic parameters were unaffected by SR141716 in noncirrhotic control subjects (Bátkai et al., 2001; Ros et al., 2002). These findings suggested an increased endocannabinoid tone in cirrhosis, which could be attributed to both an up-regulation of CB1 receptors in hepatic vascular endothelial cells and an increased production of anandamide by circulating monocytes (Bátkai et al., 2001). Increased expression of CB1 receptors was also reported in whole liver from bile duct-ligated mice (Biecker et al., 2004). This increase was greater when bile duct ligation was performed in NO synthase-3 knockout compared with wild-type mice, which may account for the similar level of portal hypertension in the two strains despite the much higher systemic blood pressure in the knockout mice (Biecker et al., 2004). Increased anandamide-induced vasorelaxation mediated by CB1 and TRPV1 receptors was also reported in mesenteric arteries isolated from cirrhotic compared with control rats (Domenicali et al., 2005). The increase in anandamide in monocytes from cirrhotic rats or humans is functionally important, as these cells elicit SR141716-sensitive hypotension when injected into normal recipient rats (Bátkai et al., 2001; Ros et al., 2002). Plasma endotoxin levels progressively increase as liver function deteriorates in cirrhosis (Lumsden et al., 1988; Chan et al., 1997), and this effect is probably responsible for the elevated endocannabinoid production in plasma monocytes and platelets of cirrhotic animals and patients (Bátkai et al., 2001; Ros et al., 2002; Liu et al., 2003; Fernandez-Rodriguez et al., 2004). There is also recent experimental evidence implicating increased signaling through myocardial CB1 receptors in the pathogenesis of cirrhotic cardiomyopathy (Gaskari et al., 2005; Pacher et al., 2005c).
Beyond the vasculopathy of end-stage cirrhosis, the endocannabinoid system may also be involved in the pathogenesis of liver fibrosis. Siegmund et al. (2005) have recently reported that anandamide exerts antifibrogenic effects in vitro by inhibiting activated hepatic stellate cells at low micromolar concentrations and by inducing their necrosis at higher concentrations, via CB1/2- and TRPV1-independent mechanism(s). In a study by Julien et al. (2005), the liver fibrosis induced by carbon tetrachloride was more severe in CB2 knockout mice compared with their wild-type littermates. Also, the expression of CB2 receptors was found to be strongly induced in liver biopsy specimens from patients with active cirrhosis of various etiologies, particularly in non-parenchymal cells located within and at the edge of fibrous septa (Julien et al., 2005). Furthermore, CB2 receptor activation triggered growth inhibition and apoptosis in myofibroblasts and in activated hepatic stellate cells, highlighting the antifibrogenic role of CB2 receptors during chronic liver injury (Julien et al., 2005). However, chronic marijuana use has been associated with hepatotoxicity rather than hepatoprotection as expected from the above results (Borini et al., 2004), and results of a recent epidemiological study indicate that daily marijuana smoking is a risk factor for progression of fibrosis among people with chronic hepatitis C infection (Hezode et al., 2005). This finding has triggered an investigation into the possible profibrogenic role of CB1 receptor activation, which is supported by the results of a preliminary study showing that the progression of experimental liver fibrosis induced by carbon tetrachloride is slower in mice with genetic ablation of CB1 receptors or treated with CB1 receptor antagonist SR141716 (Teixeira-Clerc et al., 2006). These latter findings suggest a broader role of CB1 receptors in the pathogenesis of cirrhosis and forecast additional potential benefits from the therapeutic use of a CB1 antagonist in chronic liver disease.
In contrast to the hepatotoxicity associated with chronic marijuana use, a synthetic, nonpsychotropic cannabinoid derivative (PRS-211,092) was reported to inhibit acute hepatitis induced by concanavalin A via negative cytokine regulation in mice (Lavon et al., 2003). Interestingly, in animal models of acute hepatic failure-induced encephalopathy, both 2-AG and SR141716 have been reported to exert beneficial effects on neurological and cognitive function (Gabbay et al., 2005; Avraham et al., 2006). Cannabinoids may also be beneficial in intractable cholestatic pruritus (Neff et al., 2002), which is associated with severe forms of liver disease, presumably by increasing the nociceptive threshold (Gingold and Bergasa, 2003).
Collectively, the studies discussed in this section highlight the potential regulatory role of the endocannabinoid system in a variety of gastrointestinal and liver disorders, opening new avenues for their pharmacotherapy. It appears that CB1 agonists and perhaps FAAH antagonists might be beneficial in reducing increased gastrointestinal motility, bowel inflammation, and associated diarrhea, whereas CB1 antagonists could be used in the treatment of constipation. In chronic liver cirrhosis, CB1 antagonists may not only attenuate or reverse the adverse hemodynamic consequences of cirrhosis, thus extending life until a suitable liver becomes available for transplantation, but also could have additional benefits by slowing the progression of fibrosis and the neurological decline associated with hepatic encephalopathy. Selective CB2 receptor agonists might also be expected to protect against progression of liver fibrosis and perhaps against the chronic inflammation associated with IBD.
H. Musculoskeletal Disorders
1. Arthritis
Rheumatoid arthritis (RA) is a chronic, systemic, inflammatory autoimmune disease affecting ~0.8% of adults worldwide. RA is more common in women, and it leads to joint destruction, deformity, loss of function, chronic pain, and reduced quality of life. When unchecked, it leads to substantial disability and premature death (O’Dell, 2004). Current medications used to treat rheumatoid arthritis are divided into three main classes: nonsteroidal anti-inflammatory drugs, corticosteroids, and disease-modifying antirheumatic drugs such as methotrexate (O’Dell, 2004). A better understanding of the cytokine networks that are responsible for the ongoing inflammatory response in RA has led to the successful use of novel therapies that target TNF-α and IL-1.
The immunosuppressant and anti-inflammatory properties of cannabinoids are highly relevant for RA and other autoimmune disorders (e.g., systemic lupus erythematosus, autoimmune vasculitis, Sjögren’s syndrome, and ankylosing spondylitis). Indeed, ajulemic acid (THC-11-oic acid, CT-3, IP-751), a potent analog of the acid metabolites of THC (Burstein, 2000, 2005) and cannabidiol have been shown to have analgesic, anti-inflammatory, and immunosuppressive effects in animal models of arthritis (Zurier et al., 1998; Dajani et al., 1999; Malfait et al., 2000). Chronic administration of ajulemic acid attenuated joint inflammation in a murine model of adjuvant-induced arthritis and suppressed prostaglandin production in vitro to a greater extent than the potent nonsteroidal anti-inflammatory drug, indomethacin (Zurier et al., 1998). In another study, ajulemic acid caused less gastrointestinal ulcerations and was more effective in reducing adjuvant-induced arthritis than common nonsteroidal anti-inflammatory agents (Dajani et al., 1999). As discussed earlier in this review, ajulemic acid is a high-affinity agonist for human cannabinoid receptors and has CB1-mediated, potent antihyperalgesic activity in models of chronic neuropathic and inflammatory pain in the rat (Dyson et al., 2005). Ajulemic acid also induces apoptosis in human T lymphocytes (Bidinger et al., 2003) and suppresses IL-1β production in human monocytes (Zurier et al., 2003), which could contribute to its therapeutic effects in RA and other inflammatory disorders. Treatment with cannabidiol or its more potent dimethylheptyl derivative (HU-320) reduced an LPS-induced increase in serum TNF-α and immune function and effectively blocked the progression of collagen-induced arthritis in mice (Malfait et al., 2000; Sumariwalla et al., 2004). Other studies described the antinociceptive effects of anandamide and THC in rats with arthritis (Sofia et al., 1973; Smith et al., 1998; Cox and Welch, 2004). Mbvundula et al. (2005, 2006) have recently reported that WIN 55,212-2 and HU-210 inhibited IL-1-stimulated NO production in bovine articular chondrocytes, in contrast to AM281 and AM630, which elicited an opposite effect. Anandamide, WIN 55212-2, and HU-210 also inhibited the release of sulfated glycosaminoglycans in bovine cartilage explants and IL-1a stimulated proteoglycan and collagen degradation (Mbvundula et al., 2005, 2006).
In a survey of 2969 people using cannabis for medicinal purposes, ~25% of subjects mentioned relief of arthritis symptoms as the main reason for cannabis smoking, which was surpassed only by chronic pain, MS, and depression (Ware et al., 2003). Studies using cannabinoid-based extracts are also underway in patients with RA (Russo, 2006). The potential benefit of cannabinoids in fibromyalgia, a syndrome of widespread musculoskeletal pain, nonrestorative sleep, disturbed mood, and fatigue of unknown etiology, has also been reviewed (Russo, 2004).
2. Osteoporosis
Osteoporosis is a skeletal disorder characterized by low bone mass and microarchitectural deterioration of bone, leading to increased susceptibility to bone fractures. The associated fractures and the subsequent morbidity and mortality make osteoporosis an enormous public health concern. Osteoporosis is no longer considered an age-related disease, as it is increasingly recognized in children. Osteoporosis is thought to be a polygenic disorder, with vulnerability determined by multiple genes and environmental risk factors. It currently affects up to one in three women and 1 in 12 men worldwide (Keen, 2003). Treatment options include general measures on lifestyle, calcium and vitamin D supplements, hormone therapy, raloxifene, and bisphosphonates.
Cannabinoid receptors were first implicated in the regulation of bone mass by Karsak et al. (2004), who found that CB2 knockout mice had markedly accelerated age-related trabecular bone loss and cortical expansion accompanied by increased activity of trabecular osteoblasts, increased numbers of osteoclasts, and decreased numbers of diaphyseal osteoblast precursors (Ofek et al., 2006). CB2 receptors were expressed in osteoblasts, osteocytes, and osteoclasts. The selective CB2 agonist HU-308, but not the CB1 agonist noladine ether, attenuated ovariectomy-induced bone loss and markedly stimulated cortical thickness through the suppression of osteoclast number and stimulation of endocortical bone formation (Ofek et al., 2006). Furthermore, HU-308 dose dependently increased the number and activity of endocortical osteoblasts and restrained trabecular osteoclastogenesis by inhibiting proliferation of osteoclast precursors (Ofek et al., 2006). These results, coupled with CB2 but not CB1 receptor mRNA expression during osteoblastic differentiation, suggested a role for CB2 receptors in bone remodeling. Such a role of CB2 but not CB1 receptors is also supported by a recent genetic association study in human samples of postmenopausal osteoporosis patients and matched female control subjects (Karsak et al., 2005).
In contrast, Idris et al. (2005) have recently reported that CB1 receptor knockout mice or mice treated with antagonists of either CB1 or CB2 receptors were protected from ovariectomy-induced bone loss. Furthermore, cannabinoid antagonists promoted osteoclast apoptosis, inhibited osteoclast activity, and decreased the production of several osteoclast survival factors in vitro, suggesting that cannabinoid antagonists may be beneficial in the treatment of osteoporosis. Although the reason for the discrepancy between the above studies is not clear; they suggest a role for the endocannabinoid system in the regulation of bone mass.
I. Endocannabinoids and Reproductive Functions
There is abundant evidence that the endocannabinoid system is involved in reproductive functions in both males and females and in both animals and humans, as discussed in more detail in recent reviews (Fride, 2004; Park et al., 2004; Schuel and Burkman, 2005; Tranguch et al., 2005; Wang et al., 2006). In males, marijuana, synthetic cannabinoids, and anandamide adversely affect the fertilizing capacity of sperm, which express functional CB1 receptors (Rossato et al., 2005; Schuel and Burkman, 2005; Whan et al., 2006). On the other hand, there is preclinical evidence to suggest that blockade of CB1 may be useful in the treatment of erectile dysfunction (Melis et al., 2004b, 2006).
High levels of functional CB1 receptor, anandamide, and FAAH are present in the preimplantation embryo and/or in the uterus (Das et al., 1995; Paria et al., 1995, 2001; Schmid et al., 1997; Park et al., 2003; Guo et al., 2005). Anandamide synthesized in the uterus exerts dose- and stage-specific effects on embryo development and implantation. A temporary reduction of anandamide levels is essential for embryo implantation, and higher anandamide levels are associated with uterine nonreceptivity and impairment of blastocyst formation, zona hatching, and trophoblast outgrowth via CB1 receptors (Das et al., 1995; Paria et al., 1995, 2001, 2002; Schmid et al., 1997; Wang et al., 1999; Guo et al., 2005). Consequently, cannabinoids may retard the development of embryos, eventually leading to fetal loss and pregnancy failure (Bloch et al., 1978; Smith and Asch, 1987; Park et al., 2004). Anandamide levels in the uterus are regulated by FAAH activity (Paria et al., 1995, 1999; Schmid et al., 1997). Accordingly, pregnant women with low FAAH activity in lymphocytes were found to have an increased incidence of miscarriage (Maccarrone et al., 2000c), and low FAAH activity also correlated with failure to maintain pregnancy after in vitro fertilization (Maccarrone et al., 2002b). Finally, cannabinoids may also affect the levels of various hormones crucial for normal fertility and reproduction (Brown and Dobs, 2002; Park et al., 2004; Scorticati et al., 2004; Gammon et al., 2005). Although such findings may suggest the potential usefulness of CB1 antagonists in the treatment of infertility problems, a note of caution is warranted because CB1 knockout mice were reported to have impaired oviductal transport of embryos, leading to embryo retention. This suggests that treatment with CB1 antagonists may facilitate ectopic pregnancy (Wang et al., 2004).
IV. Future Directions
The length of this review, necessitated by the steady growth in the number of indications for the potential therapeutic use of cannabinoid-related medications, is a clear sign of the emerging importance of this field. This is further underlined by the quantity of articles in the public database dealing with the biology of cannabinoids, which numbered ~200 to 300/year throughout the 1970s to reach an astonishing 5900 in 2004. The growing interest in the underlying science has been matched by a growth in the number of cannabinoid drugs in pharmaceutical development from two in 1995 to 27 in 2004, with the most actively pursued therapeutic targets being pain, obesity, and multiple sclerosis (Hensen, 2005). As in any rapidly growing area of research, not all the leads will turn out to be useful or even valid. Nevertheless, it is safe to predict that new therapeutic agents that affect the activity of the endocannaboinoid system will emerge and become members of our therapeutic armamentarium. The plant-derived cannabinoid preparation Sativex has already gained regulatory approval in Canada for the treatment of spasticity and pain associated with multiple sclerosis, and the CB1 receptor antagonist rimonabant has been approved in Europe and is awaiting Food and Drug Administration approval in the United States for the treatment of the metabolic syndrome. Undoubtedly, these will be followed by new and improved compounds aimed at the same or additional targets in the endocannabinoid system. However, it may be only after the widespread therapeutic use of such compounds that some important side effects will emerge. Although this occurrence would be undesirable from a health care perspective, such side effects may shed further light on the biological functions of endocannabinoids in health and disease.
Acknowledgments
We are indebted to Kimberly Bochniak for help with the list of references. P.P. dedicates this review to his beloved mother, Iren.
Work from the authors’ laboratory was supported by the Intramural Research Program of the National Institutes of Health/National Institute on Alcohol Abuse and Alcoholism.
Footnotes
Abbreviations: THC or Δ9-THC, Δ9-tetrahydrocannabinol; CP-55,940, (1R,3R,4R)-3-[2-hydroxy-4-(1,1-dimethylheptyl)phenyl]-4-(3-hydroxypropyl)cyclohexan-1-ol; GPCR, G protein-coupled receptor; CB1 or CB2, cannabinoid 1 or 2; CBD, cannabidiol; SR141716, N-(piperidin-1-yl)-5-(4-chlorophenyl)-1-(2,4-dichlorophenyl)-4-methyl-1H-pyrazole-3-carboximide hydrochloride (rimonabant); AM251, N-(piperin-1-yl)-5-(4-iodophenyl)-1-(2,4-dichlorophenyl)-4-methyl-1H-pyrazole-3-carboxamide; TRPV1 or VR1, transient receptor potential vanilloid 1 or vanilloid 1; WIN 55,212-2, R-(+)-[2,3-dihydro-5-methyl-3-[(morpholinyl)methyl]pyrrolo-[1,2,3-de]-1,4-benzoxazinyl]-(1-naphthalenyl)methanone mesylate; GTPγS, guanosine 5′-O-(3-thio)triphosphate; HU-210, Δ8-tetrahydrocannabinol dimethyl heptyl; DARPP-32, dopamine- and cAMP-regulated phosphoprotein of 32 kDa; 2-AG, 2-arachidonoylglycerol; NAPE; N-arachidonoyl phosphatidylethanolamide; PE, phosphatidylethanolamine; PL, phospholipase; DAG, diacylglycerol; FAAH, fatty acid amide hydrolase; UCM707, N-(3-furanylmethyl)-5Z,8Z,11Z,14Z-eicosatetraenamide; LY2318912, 5-(4-azido-3-iodo-benzoylaminomethyl]-tetrazole-1-carboxylic acid dimethylamide; MGL, monoacylglyceride lipase; DSI, depolarization-induced suppression of inhibition; SR144528, N-((1S)-endo-1,3,3-trimethyl bicyclo heptan-2-yl]-5-(4chloro-3-methylphenyl)-1-(4-methylbenzyl)-pyrazole-3-carboxamide); NPY, neuropeptide Y; MCH, melanin concentrating hormone; α-MSH, α-melanocyte-stimulating hormone; CRH, corticotropin-releasing hormone; CART, cocaine- and amphetamine-related transcript; AMPK, AMP-activated protein kinase; ACC1, acetyl CoA carboxylase-1; SREBP1c, sterol response element binding protein 1c; HDL, high-density lipoprotein; LDL, low-density lipoprotein; CNS, central nervous system; HIV, human immunodeficiency virus; LPS, lipopolysaccharide or endotoxin; TNF-α, tumor necrosis factor-α; IL, interleukin; CXCL, CXC chemokine ligand; NMDA receptor, N-methyl-D-aspartate receptor; HU-211, dexanabinol; TBI, traumatic brain injury; BAY 38-7271, (−)-(R)-3-(2-hydroxymethylindanyl-4-oxy)phenyl-4,4,4-trifluoro-1-sulfonate; MCAo, middle cerebral artery occlusion; GABA, gamma-aminobutyric acid; GPe or GPi, external or internal globus pallidus; HD, Huntington’s disease; HPA axis, hypothalamic-pituitary-adrenal axis; HU-211, dexanabinol; ICAM-1, intercellular adhesion molecule-1; IL, interleukin; I/R, ischemia reperfusion; KA, kainic acid; LID, levodopa-induced dyskinesia; methyl-D-aspartate receptor; NO, nitric oxide; PD, Parkinson’s disease; LY320135, [6-methoxy-2-(4-methoxyphenyl)benzo[b]-thien-3-yl][4-cyanophenyl] methanone; MS, multiple sclerosis; SCI, spinal cord injury; EAE, experimental autoimmune encephalomyelitis; JWH-133, 1,1-dimethylbutyl-1-deoxy-Δ9-tetrahydrocannabinol; PEA, palmitoylethanolamide; ACEA, arachidonyl-2′-chloroethylamide/(all Z)-N-(2-cycloethyl)-5,8,11,14-eicosatetraenamide; JWH-015, (2-methyl-1-propyl-1H-indol-3-yl)-1-naphthalenylmethanone; OM-DM1, (R)-N-oleoyl-(1′-hydroxybenzyl)-2′-ethanolamine; OMDM2, (S)-N-oleoyl-(1′-hydroxybenzyl)-2′-ethanolamine; SNr, substantia nigra pars reticulata; LID, levodopa-induced dyskinesia; GPe or GPi, external or internal globus pallidus; HD, Huntington’s disease; ALS, amyotrophic lateral sclerosis; AM404, N-(4-hydroxyphenyl)-eicosa-5,8,11,14-tetraenamide; VDM11, N-(4-hydroxy-2-methylphenyl) arachidonoyl amide; AM374, palmitylsulfonyl fluoride; TS, Gilles de la Tourette’s syndrome; AD, Alzheimer’s disease; Aβ, β-amyloid; HPA, hypothalamic-pituitary-adrenal; URB597, cyclohexyl carbamic acid 3′-carbamoyl-biphenyl-3-yl ester; 5-HT, 5-hydroxytryptamine (serotonin); VTA, ventral tegmental area; nAc, nucleus accumbens; CPP, conditioned place preference; MDMA, 3,4-methylenedioxymethamphetamine (Ecstasy); SHR, spontaneously hypertensive rat(s); WKY, Wistar-Kyoto; AM281, N-(morpholin-4-yl)-1-(2,4-dichlorophenyl)-5-(4-iodophenyl)-4-methyl-1H-pyrazole-3-carboxamide; AM630, 6-iodo-2-methyl-1-[2-(4-morpholinyl)ethyl]-1-H-indol-3-yl(4-methoxyphenyl)-methanone; IBD, inflammatory bowel disease; PRS-211,092, [(+)-(6aS,10aS)-6,6-dimethyl-3-(1,1-dimethylheptyl)-1-hydroxy-9-(1H-imidazol-2-ylsulfanylmethyl]-6a-,7,10,10a-tetrahydro-6H-dibenzo[b,d]pyran; RA, rheumatoid arthritis; HU-320, cannabidiol-dimethylheptyl-7-oic acid; HU-308, (+)-(1-aH,3H,5aH)-4-[2,6-dimethoxy-4-(1,1-dimethylheptyl)phenyl]-6,6-dimethylbicyclo[3.1.1]hept-2-ene-2-carbinol.
References
- Aapro M. Optimising antiemetic therapy: what are the problems and how can they be overcome? Curr Med Res Opin. 2005;21:885–897. doi: 10.1185/030079905X46313. [DOI] [PubMed] [Google Scholar]
- Abboud RT, Sanders HD. Effect of oral administration of Δ-tetrahydrocannabinol on airway mechanics in normal and asthmatic subjects. Chest. 1976;70:480–485. doi: 10.1378/chest.70.4.480. [DOI] [PubMed] [Google Scholar]
- Abel EL. Cannabis: effects on hunger and thirst. Behav Biol. 1975;15:255–281. doi: 10.1016/s0091-6773(75)91684-3. [DOI] [PubMed] [Google Scholar]
- Abood ME, Rizvi G, Sallapudi N, McAllister SD. Activation of the CB1 cannabinoid receptor protects cultured mouse spinal neurons against excitotoxicity. Neurosci Lett. 2001;309:197–201. doi: 10.1016/s0304-3940(01)02065-1. [DOI] [PubMed] [Google Scholar]
- Abu-Elmagd KM, Zak M, Stamos JM, Bond GJ, Jain A, Youk AO, Ezzelarab M, Costa G, Wu T, Nalesnik MA, et al. De novo malignancies after intestinal and multivisceral transplantation. Transplantation. 2004;77:1719–1725. doi: 10.1097/01.tp.0000131164.43015.4b. [DOI] [PubMed] [Google Scholar]
- Achiron A, Miron S, Lavie V, Margalit R, Biegon A. Dexanabinol (HU-211) effect on experimental autoimmune encephalomyelitis: implications for the treatment of acute relapses of multiple sclerosis. J Neuroimmunol. 2000;102:26–31. doi: 10.1016/s0165-5728(99)00149-6. [DOI] [PubMed] [Google Scholar]
- Adami M, Frati P, Bertini S, Kulkarni-Narla A, Brown DR, de Caro G, Coruzzi G, Soldani G. Gastric antisecretory role and immunohistochemical localization of cannabinoid receptors in the rat stomach. Br J Pharmacol. 2002;135:1598–1606. doi: 10.1038/sj.bjp.0704625. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Adams MD, Chait LD, Earnhardt JT. Tolerance to the cardiovascular effects of Δ9-tetrahydrocannabinol in the rat. Br J Pharmacol. 1976;56:43–48. doi: 10.1111/j.1476-5381.1976.tb06957.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Adams MD, Earnhardt JT, Martin BR, Harris LS, Dewey WL, Razdan RK. A cannabinoid with cardiovascular activity but no overt behavioral effects. Experientia (Basel) 1977;33:1204–1205. doi: 10.1007/BF01922330. [DOI] [PubMed] [Google Scholar]
- Aguado T, Monory K, Palazuelos J, Stella N, Cravatt B, Lutz B, Marsicano G, Kokaia Z, Guzmán M, Galve-Roperh I. The endocannabinoid system drives neural progenitor proliferation. FASEB J. 2005;19:1704–1706. doi: 10.1096/fj.05-3995fje. [DOI] [PubMed] [Google Scholar]
- Aguado T, Palazuelos J, Monory K, Stella N, Cravatt B, Lutz B, Marsicano G, Kokaia Z, Guzman M, Galve-Roperh I. The endocannabinoid system promotes astroglial differentiation by acting on neural progenitor cells. J Neurosci. 2006;26:1551–1561. doi: 10.1523/JNEUROSCI.3101-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alger BE. Retrograde signaling in the regulation of synaptic transmission: focus on endocannabinoids. Prog Neurobiol. 2002;68:247–286. doi: 10.1016/s0301-0082(02)00080-1. [DOI] [PubMed] [Google Scholar]
- Alger BE. Endocannabinoids and their implications for epilepsy. Epilepsy Curr. 2004;4:169–173. doi: 10.1111/j.1535-7597.2004.04501.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Allen JH, de Moore GM, Heddle R, Twartz JC. Cannabinoid hyperemesis: cyclical hyperemesis in association with chronic cannabis abuse. Gut. 2005;53:1566–1570. doi: 10.1136/gut.2003.036350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Allister SD, Chan C, Taft RJ, Luu T, Abood ME, Moore DH, Aldape K, Yount G. Cannabinoids selectively inhibit proliferation and induce death of cultured human glioblastoma multiforme cells. J Neurooncol. 2005;74:31–40. doi: 10.1007/s11060-004-5950-2. [DOI] [PubMed] [Google Scholar]
- Alward WL. Medical management of glaucoma. N Engl J Med. 1998;339:1298–1307. doi: 10.1056/NEJM199810293391808. [DOI] [PubMed] [Google Scholar]
- Amaya F, Shimosato G, Kawasaki Y, Hashimoto S, Tanaka Y, Ji RR. Induction of CB(1) cannabinoid receptor by inflammation in primary afferent neurons facilitates antihyperalgesic effect of peripheral CB(1) agonist. Pain. 2006 doi: 10.1016/j.pain.2006.04.001. in press. [DOI] [PubMed] [Google Scholar]
- Ames FR, Cridland S. Anticonvulsant effect of cannabidiol. S Afr Med J. 1986;69:14. [PubMed] [Google Scholar]
- Anderson LA, Anderson JJ, Chase TN, Walters JR. The cannabinoid agonists WIN 55,212-2 and CP 55,940 attenuate rotational behavior induced by a dopamine D1 but not a D2 agonist in rats with unilateral lesions of the nigrostriatal pathway. Brain Res. 1995;691:106–114. doi: 10.1016/0006-8993(95)00645-7. [DOI] [PubMed] [Google Scholar]
- Andreasson S, Allebeck P, Engstrom A, Rydberg U. Cannabis and schizophrenia: a longitudinal study of Swedish conscripts. Lancet. 1987;2:1483–1486. doi: 10.1016/s0140-6736(87)92620-1. [DOI] [PubMed] [Google Scholar]
- Appel LJ, Champagne CM, Harsha DW, Cooper LS, Obarzanek E, Elmer PJ, Stevens VJ, Vollmer WM, Lin PH, Svetkey LP, et al. Effects of comprehensive lifestyle modification on blood pressure control: main results of the PREMIER clinical trial. J Am Med Assoc. 2003;289:2083–2093. doi: 10.1001/jama.289.16.2083. [DOI] [PubMed] [Google Scholar]
- Archer RA. The cannabinoids: therapeutic potentials. Annu Rev Med Chem. 1974;9:253–259. doi: 10.1016/s0065-7743(08)61448-7. [DOI] [PubMed] [Google Scholar]
- Arevalo C, de Miguel R, Hernandez-Tristan R. Cannabinoid effects on anxiety-related behaviours and hypothalamic neurotransmitters. Pharmacol Biochem Behav. 2001;70:123–131. doi: 10.1016/s0091-3057(01)00578-0. [DOI] [PubMed] [Google Scholar]
- Arevalo-Martin A, Vela JM, Molina-Holgado E, Borrell J, Guaza C. Therapeutic action of cannabinoids in a murine model of multiple sclerosis. J Neurosci. 2003;23:2511–2516. doi: 10.1523/JNEUROSCI.23-07-02511.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arnone M, Maruani J, Chaperon F, Thiebot MH, Poncelet M, Soubrie P, Le Fur G. Selective inhibition of sucrose and ethanol intake by SR141716, an antagonist of central cannabinoid (CB1) receptors. Psychopharmacology. 1997;132:104–106. doi: 10.1007/s002130050326. [DOI] [PubMed] [Google Scholar]
- Andersson M, Usiello A, Borgkvist A, Pozzi L, Dominguez C, Fienberg AA, Svenningsson P, Fredholm BB, Borrelli E, Greengard P, et al. Cannabinoid action depends on phosphorylation of dopamine- and cAMP-regulated phospho-protein of 32 kDa at the protein kinase A site in striatal projection neurons. J Neurosci. 2005;25:8432–8438. doi: 10.1523/JNEUROSCI.1289-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ashton CH. Adverse effects of cannabis and cannabinoids. Br J Anaesth. 1999;83:637–649. doi: 10.1093/bja/83.4.637. [DOI] [PubMed] [Google Scholar]
- Ashton CH, Moore PB, Gallagher P, Young AH. Cannabinoids in bipolar affective disorder: a review and discussion of their therapeutic potential. J Psychopharmacol. 2005;19:293–300. doi: 10.1177/0269881105051541. [DOI] [PubMed] [Google Scholar]
- Ashton H, Golding J, Marsh VR, Millman JE, Thompson JW. The seed and the soil: effect of dosage, personality and starting state on the response to Δ9 tetrahydrocannabinol in man. Br J Clin Pharmacol. 1981;12:705–720. doi: 10.1111/j.1365-2125.1981.tb01294.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ashworth B. Preliminary trial of carisoprodol in multiple sclerosis. Practitioner. 1964;192:540–542. [PubMed] [Google Scholar]
- Attal N, Brasseur L, Guirimand D, Clermond-Gnamien S, Atlami S, Bouhassira D. Are oral cannabinoids safe and effective in refractory neuropathic pain? Eur J Pain. 2004;8:173–177. doi: 10.1016/S1090-3801(03)00084-3. [DOI] [PubMed] [Google Scholar]
- Avraham Y, Israeli E, Gabbay E, Okun A, Zolotarev O, Silberman I, Ganzburg V, Dagon Y, Magen I, Vorobia L, et al. Endocannabinoids affect neurological and cognitive function in thioacetamide-induced hepatic encephalopathy in mice. Neurobiol Dis. 2006;1:237–245. doi: 10.1016/j.nbd.2005.07.008. [DOI] [PubMed] [Google Scholar]
- Awumey EM, Howlett AC, Diz DI. Is there a role for anandamide in cardiovascular regulation? Insights from studies of endocannabinoid metabolism. Am J Physiol. 2005;289:H520–H521. doi: 10.1152/ajpheart.00433.2005. [DOI] [PubMed] [Google Scholar]
- Azad SC, Eder M, Marsicano G, Lutz B, Zieglgänsberger W, Rammes G. Activation of the cannabinoid receptor type 1 decreases glutamatergic and GABAergic synaptic transmission in the lateral amygdala of the mouse. Learn Mem. 2003;10:116–128. doi: 10.1101/lm.53303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baker D, Pryce G. The therapeutic potential of cannabis in multiple sclerosis. Expert Opin Invest Drugs. 2003;12:561–567. doi: 10.1517/13543784.12.4.561. [DOI] [PubMed] [Google Scholar]
- Baker D, Pryce G, Croxford JL, Brown P, Pertwee RG, Huffman JW, Layward L. Cannabinoids control spasticity and tremor in a multiple sclerosis model. Nature (Lond) 2000;404:84–87. doi: 10.1038/35003583. [DOI] [PubMed] [Google Scholar]
- Baker D, Pryce G, Croxford JL, Brown P, Pertwee RG, Makriyannis A, Khanolkar A, Layward L, Fezza F, Bisogno T, et al. Endocannabinoids control spasticity in a multiple sclerosis model. FASEB J. 2001;15:300–302. doi: 10.1096/fj.00-0399fje. [DOI] [PubMed] [Google Scholar]
- Balerio GN, Aso E, Maldonado R. Role of the cannabinoid system in the effects induced by nicotine an anxiety-like behaviour in mice. Psychopharmacology. 2006;184:504–513. doi: 10.1007/s00213-005-0251-9. [DOI] [PubMed] [Google Scholar]
- Ballon N, Leroy S, Roy C, Bourdel MC, Charles-Nicolas A, Krebs MO, Poirier MF. (AAT)n repeat in the cannabinoid receptor gene (CNR1): association with cocaine addiction in an African-Caribbean population. Pharmacogenonics J. 2006;6:126–130. doi: 10.1038/sj.tpj.6500352. [DOI] [PubMed] [Google Scholar]
- Ban TA. Neuropsychopharmacology and the genetics of schizophrenia: a history of the diagnosis of schizophrenia. Prog Neuropsychopharmacol Biol Psychiatry. 2004;28:753–762. doi: 10.1016/j.pnpbp.2004.05.021. [DOI] [PubMed] [Google Scholar]
- Barann M, Molderings G, Brüss M, Bönisch H, Urban BW, Göthert M. Direct inhibition by cannabinoids of human 5-HT3A receptors: probable involvement of an allosteric modulatory site. Br J Pharmacol. 2002;137:589–596. doi: 10.1038/sj.bjp.0704829. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barnham KJ, Masters CL, Bush AI. Neurodegenerative diseases and oxidative stress. Nat Rev Drug Discov. 2004;3:205–214. doi: 10.1038/nrd1330. [DOI] [PubMed] [Google Scholar]
- Bar-Joseph A, Berkovitch Y, Adamchik J, Biegon A. Neuroprotective activity of HU-211, a novel NMDA antagonist, in global ischemia in gerbils. Mol Chem Neuropathol. 1994;23:125–135. doi: 10.1007/BF02815406. [DOI] [PubMed] [Google Scholar]
- Barratt ES, Beaver W, White R. The effects of marijuana on human sleep patterns. Biol Psychiatry. 1974;8:47–54. [PubMed] [Google Scholar]
- Barsky SH, Roth MD, Kleerup EC, Simmons M, Tashkin DP. Histopathologic and molecular alterations in bronchial epithelium in habitual smokers of marijuana, cocaine, and/or tobacco. J Natl Cancer Inst. 1998;90:1198–1205. doi: 10.1093/jnci/90.16.1198. [DOI] [PubMed] [Google Scholar]
- Basavarajappa BS, Hungund BL. Down-regulation of cannabinoid receptor agonist-stimulated [35S]GTPγS binding in synaptic plasma membrane from chronic ethanol exposed mice. Brain Res. 1999;815:89–97. doi: 10.1016/s0006-8993(98)01072-5. [DOI] [PubMed] [Google Scholar]
- Basavarajappa BS, Hungund BL. Neuromodulatory role of the endocannabinoid signaling system in alcoholism: an overview. Prostaglandins Leukotrienes Essent Fatty Acids. 2002;66:287–299. doi: 10.1054/plef.2001.0352. [DOI] [PubMed] [Google Scholar]
- Basavarajappa BS, Yalamanchili R, Cravatt BF, Cooper TB, Hungund BL. Increased ethanol consumption and preference and decreased sensitivity in female FAAH knockout mice. Neuropharmacology. 2006;50:834–844. doi: 10.1016/j.neuropharm.2005.12.005. [DOI] [PubMed] [Google Scholar]
- Basile AS, Hanus L, Mendelson WB. Characterization of the hypnotic properties of oleamide. Neuroreport. 1999;10:947–951. doi: 10.1097/00001756-199904060-00010. [DOI] [PubMed] [Google Scholar]
- Bátkai S, Járai Z, Wagner JA, Goparaju SK, Varga K, Liu J, Wang L, Mirshahi F, Khanolkar AD, Makriyannis A, et al. Endocannabinoids acting at vascular CB1 receptors mediate the vasodilated state in advanced liver cirrhosis. Nat Med. 2001;7:827–832. doi: 10.1038/89953. [DOI] [PubMed] [Google Scholar]
- Bátkai S, Pacher P, Járai Z, Wagner JA, Kunos G. Cannabinoid antagonist SR141716 inhibits endotoxic hypotension by a cardiac mechanism not involving CB1 or CB2 receptors. Am J Physiol. 2004a;287:H595–H600. doi: 10.1152/ajpheart.00184.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bátkai S, Pacher P, Osei-Hyiaman D, Radaeva S, Liu J, Harvey-White J, Offertáler L, Mackie K, Rudd A, Bukoski RD, et al. Endocannabinoids acting at CB1 receptors regulate cardiovascular function in hypertension. Circulation. 2004b;110:1996–2002. doi: 10.1161/01.CIR.0000143230.23252.D2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bayir H, Kochanek PM, Clark RS. Traumatic brain injury in infants and children: mechanisms of secondary damage and treatment in the intensive care unit. Crit Care Clin. 2003;19:529–549. doi: 10.1016/s0749-0704(03)00014-9. [DOI] [PubMed] [Google Scholar]
- Beal JE, Olson R, Laubenstein L, Morales JO, Bellman P, Yangco B, Lefkowitz L, Plasse TF, Shepard KV. Dronabinol as a treatment for anorexia associated with weight loss in patients with AIDS. J Pain Symptom Manage. 1995;10:89–97. doi: 10.1016/0885-3924(94)00117-4. [DOI] [PubMed] [Google Scholar]
- Beal JE, Olson R, Lefkowitz L, Laubenstein L, Bellman P, Yangco B, Morales JO, Murphy R, Powderly W, Plasse TF, et al. Long-term efficacy and safety of dronabinol for acquired immunodeficiency syndrome-associated anorexia. J Pain Symptom Manage. 1997;14:7–14. doi: 10.1016/S0885-3924(97)00038-9. [DOI] [PubMed] [Google Scholar]
- Begg M, Mo FM, Offertaler L, Bátkai S, Pacher P, Razdan RK, Lovinger DM, Kunos G. G protein-coupled endothelial receptor for atypical cannabinoid ligands modulates a Ca2+-dependent K+ current. J Biol Chem. 2003;278:46188–46194. doi: 10.1074/jbc.M307258200. [DOI] [PubMed] [Google Scholar]
- Begg M, Pacher P, Bátkai S, Osei-Hyiaman D, Offertáler L, Mo F-M, Liu J, Kunos G. Evidence for novel cannabinoid receptors. Pharmacol Ther. 2005;106:133–145. doi: 10.1016/j.pharmthera.2004.11.005. [DOI] [PubMed] [Google Scholar]
- Behrens PF, Franz P, Woodman B, Lindenberg KS, Landwehrmeyer GB. Impaired glutamate transport and glutamate-glutamine cycling: downstream effects of the Huntington mutation. Brain. 2002;125:1908–1922. doi: 10.1093/brain/awf180. [DOI] [PubMed] [Google Scholar]
- Beinfeld MC, Connolly K. Activation of CB1 cannabinoid receptors in rat hippocampal slices inhibits potassium-evoked cholecystokinin release, a possible mechanism contributing to the spatial memory defects produced by cannabinoids. Neurosci Lett. 2001;301:69–71. doi: 10.1016/s0304-3940(01)01591-9. [DOI] [PubMed] [Google Scholar]
- Belayev L, Bar-Joseph A, Adamchik J, Biegon A. HU-211, a nonpsychotropic cannabinoid, improves neurological signs and reduces brain damage after severe forebrain ischemia in rats. Mol Chem Neuropathol. 1995a;25:19–33. doi: 10.1007/BF02815084. [DOI] [PubMed] [Google Scholar]
- Belayev L, Busto R, Watson BD, Ginsberg MD. Post-ischemic administration of HU-211, a novel non-competitive NMDA antagonist, protects against blood-brain barrier disruption in photochemical cortical infarction in rats: a quantitative study. Brain Res. 1995b;702:266–270. doi: 10.1016/0006-8993(95)01127-9. [DOI] [PubMed] [Google Scholar]
- Belayev L, Busto R, Zhao W, Ginsberg MD. HU-211, a novel noncompetitive N-methyl-D-aspartate antagonist, improves neurological deficit and reduces infarct volume after reversible focal cerebral ischemia in the rat. Stroke. 1995c;26:2313–2320. doi: 10.1161/01.str.26.12.2313. [DOI] [PubMed] [Google Scholar]
- Beltramo M, Stella N, Calignano A, Lin SY, Makriyannis A, Piomelli D. Functional role of high-affinity anandamide transport, as revealed by selective inhibition. Science (Wash DC) 1997;277:1094–1097. doi: 10.1126/science.277.5329.1094. [DOI] [PubMed] [Google Scholar]
- Benito C, Nunez E, Tolon RM, Carrier EJ, Rabano A, Hillard CJ, Romero J. Cannabinoid CB2 receptors and fatty acid amide hydrolase are selectively overexpressed in neuritic plaque-associated glia in Alzheimer’s disease brains. J Neurosci. 2003;23:11136–11141. doi: 10.1523/JNEUROSCI.23-35-11136.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benowitz NL, Jones RT. Cardiovascular effects of prolonged delta-9-tetrahydrocannabinol ingestion. Clin Pharmacol Ther. 1975;18:287–297. doi: 10.1002/cpt1975183287. [DOI] [PubMed] [Google Scholar]
- Ben-Shabat S, Fride E, Sheskin T, Tamiri T, Rhee MH, Vogel Z, Bisogno T, De Petrocellis L, Di Marzo V, Mechoulam R. An entourage effect: inactive endogenous fatty acid glycerol esters enhance 2-arachidonoyl-glycerol cannabinoid activity. Eur J Pharmacol. 1998;353:23–31. doi: 10.1016/s0014-2999(98)00392-6. [DOI] [PubMed] [Google Scholar]
- Bensaid M, Gary-Bobo M, Esclangon A, Maffrand JP, Le Fur G, Oury-Donat F, Soubrie P. The cannabinoid CB1 receptor antagonist SR141716 increases Acrp30 mRNA expression in adipose tissue of obese fa/fa rats and in cultured adipocyte cells. Mol Pharmacol. 2003;63:908–914. doi: 10.1124/mol.63.4.908. [DOI] [PubMed] [Google Scholar]
- Berdyshev E, Boichot E, Corbel M, Germain N, Lagente V. Effects of cannabinoid receptor ligands on LPS-induced pulmonary inflammation in mice. Life Sci. 1998;63:PL125–PL129. doi: 10.1016/s0024-3205(98)00324-5. [DOI] [PubMed] [Google Scholar]
- Berger C, Schmid PC, Schabitz WR, Wolf M, Schwab S, Schmid HH. Massive accumulation of N-acylethanolamines after stroke: cell signalling in acute cerebral ischemia? J Neurochem. 2004;88:1159–1167. doi: 10.1046/j.1471-4159.2003.02244.x. [DOI] [PubMed] [Google Scholar]
- Berghuis P, Dobszay MB, Wang X, Spano S, Ledda F, Sousa KM, Schulte G, Ernfors P, Mackie K, Paratcha G, et al. Endocannabinoids regulate interneuron migration and morphogenesis by transactivating the TrkB receptor. Proc Natl Acad Sci USA. 2005;102:19115–19120. doi: 10.1073/pnas.0509494102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berlach DM, Shir Y, Ware MA. Experience with the synthetic cannabinoid nabilone in chronic noncancer pain. Pain Med. 2006;7:25–29. doi: 10.1111/j.1526-4637.2006.00085.x. [DOI] [PubMed] [Google Scholar]
- Berman JS, Symonds C, Birch R. Efficacy of two cannabis based medicinal extracts for relief of central neuropathic pain from brachial plexus avulsion: results of a randomised controlled trial. Pain. 2004;112:299–306. doi: 10.1016/j.pain.2004.09.013. [DOI] [PubMed] [Google Scholar]
- Berrendero F, Maldonado R. Involvement of the opioid system in the anxiolytic-like effects induced by Delta(9)-tetrahydrocannabinol.Δ9-tetrahydrocannabinol. Psychopharmacology (Berl) 2002;163:111–117. doi: 10.1007/s00213-002-1144-9. [DOI] [PubMed] [Google Scholar]
- Berrendero F, Sanchez A, Cabranes A, Puerta C, Ramos JA, Garcia-Merino A, Fernandez-Ruiz J. Changes in cannabinoid CB1 receptors in striatal and cortical regions of rats with experimental allergic encephalomyelitis, an animal model of multiple sclerosis. Synapse. 2001;41:195–202. doi: 10.1002/syn.1075. [DOI] [PubMed] [Google Scholar]
- Berry EM, Mechoulam R. Tetrahydrocannabinol and endocannabinoids in feeding and appetite. Pharmacol Ther. 2002;95:185–190. doi: 10.1016/s0163-7258(02)00257-7. [DOI] [PubMed] [Google Scholar]
- Bezard E, Brotchie JM, Gross CE. Pathophysiology of levodopa-induced dyskinesia: potential for new therapies. Nat Rev Neurosci. 2001;2:577–588. doi: 10.1038/35086062. [DOI] [PubMed] [Google Scholar]
- Bhatia S, Louie AD, Bhatia R, O’Donnell MR, Fung H, Kashyap A, Krishnan A, Molina A, Nademanee A, Niland JC, et al. Solid cancers after bone marrow transplantation. J Clin Oncol. 2001;19:464–471. doi: 10.1200/JCO.2001.19.2.464. [DOI] [PubMed] [Google Scholar]
- Biber B, Schaefer CF, Smolik MJ, Lerner MR, Brackett DJ, Wilson MF, Fagraeus L. Improved techniques for cardiovascular monitoring in rats as applied during endotoxemia. Am J Physiol. 1988;254:H181–H186. doi: 10.1152/ajpheart.1988.254.1.H181. [DOI] [PubMed] [Google Scholar]
- Biddinger SB, Almind K, Miyazaki M, Kokkotou E, Ntambi JM, Kahn CR. Effects of diet and genetic background on sterol regulatory element-binding protein-1c, stearoyl-CoA desaturase 1, and the development of the metabolic syndrome. Diabetes. 2005;54:1314–1323. doi: 10.2337/diabetes.54.5.1314. [DOI] [PubMed] [Google Scholar]
- Bidinger B, Torres R, Rossetti RG, Brown L, Beltre R, Burstein S, Lian JB, Stein GS, Zurier RB. Ajulemic acid, a nonpsychoactive cannabinoid acid, induces apoptosis in human T lymphocytes. Clin Immunol. 2003;108:95–102. doi: 10.1016/s1521-6616(03)00064-0. [DOI] [PubMed] [Google Scholar]
- Biecker E, Sagesser H, Reichen J. Vasodilator mRNA levels are increased in the livers of portal hypertensive NO-synthase 3-deficient mice. Eur J Clin Investig. 2004;34:283–289. doi: 10.1111/j.1365-2362.2004.01331.x. [DOI] [PubMed] [Google Scholar]
- Biegon A. Cannabinoids as neuroprotective agents in traumatic brain injury. Curr Pharm Des. 2004;10:2177–2183. doi: 10.2174/1381612043384196. [DOI] [PubMed] [Google Scholar]
- Bifulco M, Di Marzo V. Targeting the endocannabinoid system in cancer therapy: a call for further research. Nat Med. 2002;8:547–550. doi: 10.1038/nm0602-547. [DOI] [PubMed] [Google Scholar]
- Bifulco M, Laezza C, Portella G, Vitale M, Orlando P, De Petrocellis L, Di Marzo V. Control by the endogenous cannabinoid system of ras oncogene-dependent tumor growth. FASEB J. 2001;15:2745–2747. doi: 10.1096/fj.01-0320fje. [DOI] [PubMed] [Google Scholar]
- Bifulco M, Laezza C, Valenti M, Ligresti A, Portella G, Di Marzo V. A new strategy to block tumor growth by inhibiting endocannabinoid inactivation. FASEB J. 2004;18:1606–1608. doi: 10.1096/fj.04-1754fje. [DOI] [PubMed] [Google Scholar]
- Bilkei-Gorzo A, Racz I, Valverde O, Otto M, Michel K, Sastre M, Zimmer A. Early age-related cognitive impairment in mice lacking cannabinoid CB1 receptors. Proc Natl Acad Sci USA. 2005;102:15670–15675. doi: 10.1073/pnas.0504640102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bilsland LG, Dick JR, Pryce G, Petrosino S, Di Marzo V, Baker D, Greensmith L. Increasing cannabinoid levels by pharmacological and genetic manipulation delay disease progression in SOD1 mice. FASEB J. 2006;20:1003–1005. doi: 10.1096/fj.05-4743fje. [DOI] [PubMed] [Google Scholar]
- Birmingham MK. Reduction by 9-tetrahydrocannabinol in the blood pressure of hypertensive rats bearing regenerated adrenal glands. Br J Pharmacol. 1973;48:169–171. doi: 10.1111/j.1476-5381.1973.tb08236.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bisogno T, Berrendero F, Ambrosino G, Cebeira M, Ramos JA, Fernandez-Ruiz JJ, Di Marzo V. Brain regional distribution of endocannabinoids: implications for their biosynthesis and biological function. Biochem Biophys Res Commun. 1999a;256:377–380. doi: 10.1006/bbrc.1999.0254. [DOI] [PubMed] [Google Scholar]
- Bisogno T, Delton-Vandenbroucke I, Milone A, Lagarde M, Di Marzo V. Biosynthesis and inactivation of N-arachidonoylethanolamine (anandamide) and N-docosahexaenoylethanolamine in bovine retina. Arch Biochem Biophys. 1999b;370:300–307. doi: 10.1006/abbi.1999.1410. [DOI] [PubMed] [Google Scholar]
- Bisogno T, Hanus L, De Petrocellis L, Tchilibon S, Ponde DE, Brandi I, Moriello AS, Davis JB, Mechoulam R, Di Marzo V. Molecular targets for cannabidiol and its synthetic analogues: effect on vanilloid VR1 receptors and on the cellular uptake and enzymatic hydrolysis of anandamide. Br J Pharmacol. 2001;134:845–852. doi: 10.1038/sj.bjp.0704327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bisogno T, Howell F, Williams G, Minassi A, Cascio MG, Ligresti A, Matias I, Schiano-Moriello A, Paul P, Williams EJ, et al. Cloning of the first sn1-DAG lipases points to the spatial and temporal regulation of endocannabinoid signaling in the brain. J Cell Biol. 2003;163:463–468. doi: 10.1083/jcb.200305129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bisogno T, Ligresti A, Di Marzo V. The endocannabinoid signaling system: biochemical aspects. Pharmacol Biochem Behav. 2005;81:224–238. doi: 10.1016/j.pbb.2005.01.027. [DOI] [PubMed] [Google Scholar]
- Bisogno T, Maurelli S, Melck D, De Petrocellis L, Di Marzo V. Biosynthesis, uptake and degradation of anandamide and palmitoylethanolamide in leukocytes. J Biol Chem. 1997;272:3315–3323. doi: 10.1074/jbc.272.6.3315. [DOI] [PubMed] [Google Scholar]
- Bisogno T, Melck D, Bobrov MYu, Gretskaya NM, Bezuglov VV, De Petrocellis L, Di Marzo V. N-Acyl-dopamines: novel synthetic CB1 cannabinoid-receptor ligands and inhibitors of anandamide inactivation with cannabimimetic activity in vitro and in vivo. Biochem J. 2000;351:817–824. [PMC free article] [PubMed] [Google Scholar]
- Bisogno T, Melck D, De Petrocellis L, Bobrov MYu, Gretskaya NM, Bezuglov VV, Sitachitta N, Gerwick WH, Di Marzo V. Arachidonoylserotonin and other novel inhibitors of fatty acid amide hydrolase. Biochem Biophys Res Commun. 1998;248:515–522. doi: 10.1006/bbrc.1998.8874. [DOI] [PubMed] [Google Scholar]
- Blázquez C, Casanova ML, Planas A, Del Pulgar TG, Villanueva C, Fernandez-Acenero MJ, Aragones J, Huffman JW, Jorcano JL, Guzman M. Inhibition of tumor angiogenesis by cannabinoids. FASEB J. 2003;17:529–531. doi: 10.1096/fj.02-0795fje. [DOI] [PubMed] [Google Scholar]
- Blázquez C, Gonzalez-Feria L, Alvarez L, Haro A, Casanova ML, Guzman M. Cannabinoids inhibit the vascular endothelial growth factor pathway in gliomas. Cancer Res. 2004;64:5617–5623. doi: 10.1158/0008-5472.CAN-03-3927. [DOI] [PubMed] [Google Scholar]
- Bloch E, Thysen B, Morrill GA, Gardner E, Fujimoto G. Effects of cannabinoids on reproduction and development. Vitam Horm. 1978;36:203–258. doi: 10.1016/s0083-6729(08)60985-1. [DOI] [PubMed] [Google Scholar]
- Bloom AS, Dewey WL, Harris LS, Brosius KK. 9-Nor-9β-hydroxyhexahy-drocannabinol a cannabinoid with potent antinociceptive activity: comparisons with morphine. J Pharmacol Exp Ther. 1977;200:263–270. [PubMed] [Google Scholar]
- Boger DL, Henriksen SJ, Cravatt BF. Oleamide: an endogenous sleep-inducing lipid and prototypical member of a new class of biological signaling molecules. Curr Pharm Des. 1998a;4:303–314. [PubMed] [Google Scholar]
- Boger DL, Miyauchi H, Du W, Hardouin C, Fecik RA, Cheng H, Hwang I, Hedrick MP, Leung D, Acevedo O, et al. Discovery of a potent, selective, and efficacious class of reversible α-ketoheterocycle inhibitors of fatty acid amide hydrolase effective as analgesics. J Med Chem. 2005;48:1849–1856. doi: 10.1021/jm049614v. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boger DL, Patterson JE, Guan X, Cravatt BF, Lerner RA, Gilula NB. ) Chemical requirements for inhibition of gap junction communication by the biologically active lipid oleamide. Proc Natl Acad Sci USA. 1998b;95:4810–4815. doi: 10.1073/pnas.95.9.4810. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bonz A, Laser M, Kullmer S, Kniesch S, Babin-Ebell J, Popp V, Ertl G, Wagner JA. Cannabinoids acting on CB1 receptors decrease contractile performance in human atrial muscle. J Cardiovasc Pharmacol. 2003;41:657–664. doi: 10.1097/00005344-200304000-00020. [DOI] [PubMed] [Google Scholar]
- Borcel E, Perez-Alvarez L, de Ceballos ML, Ramirez BG, Marco EM, Fernandez B, Rubio M, Guaza C, Viveros MP. Functional responses to the cannabinoid agonist WIN 55,212-2 in neonatal rats of both genders: influence of weaning. Pharmacol Biochem Behav. 2004;78:593–602. doi: 10.1016/j.pbb.2004.02.020. [DOI] [PubMed] [Google Scholar]
- Boring DL, Berglund BA, Howlett AC. Cerebrodiene, arachidonylethanolamide, and hybrid structures: potential for interaction with brain cannabinoid receptors. Prostaglandins Leukotrienes Essent Fatty Acids. 1996;55:207–210. doi: 10.1016/s0952-3278(96)90100-3. [DOI] [PubMed] [Google Scholar]
- Borini P, Guimaraes RC, Borini SB. Possible hepatotoxicity of chronic marijuana usage. Sao Paulo Med J. 2004;122:110–116. doi: 10.1590/S1516-31802004000300007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bortolato M, Campolongo P, Mangieri RA, Scattoni ML, Frau R, Trezza V, La Rana G, Russo R, Calignano A, Gessa GL, et al. Anxiolytic-like properties of the anandamide transport inhibitor AM404. Neuropsychopharmacology. 2006 doi: 10.1038/sj.npp.1301061. in press. [DOI] [PubMed] [Google Scholar]
- Bouaboula M, Dussossoy D, Casellas P. Regulation of peripheral cannabinoid receptor CB2 phosphorylation by the inverse agonist SR 144528: implications for receptor biological responses. J Biol Chem. 1999;274:20397–20405. doi: 10.1074/jbc.274.29.20397. [DOI] [PubMed] [Google Scholar]
- Bouaboula M, Perrachon S, Milligan L, Canat X, Rinaldi-Carmona M, Portier M, Barth F, Calandra B, Pecceu F, Lupker J, et al. A selective inverse agonist for central cannabinoid receptor inhibits mitogen-activated protein kinase activation stimulated by insulin or insulin-like growth factor 1: evidence for a new model of receptor/ligand interactions. J Biol Chem. 1997;272:22330–22339. doi: 10.1074/jbc.272.35.22330. [DOI] [PubMed] [Google Scholar]
- Bouchard JF, Lepicier P, Lamontagne D. Contribution of endocannabinoids in the endothelial protection afforded by ischemic preconditioning in the isolated rat heart. Life Sci. 2003;72:1859–1870. doi: 10.1016/s0024-3205(02)02474-8. [DOI] [PubMed] [Google Scholar]
- Bracey MH, Hanson MA, Masuda KR, Stevens RC, Cravatt BF. Structural adaptations in a membrane enzyme that terminates endocannabinoid signaling. Science (Wash DC) 2002;298:1793–1796. doi: 10.1126/science.1076535. [DOI] [PubMed] [Google Scholar]
- Braida D, Iosue S, Pegorini S, Sala M. 3,4-Methylenedioxymethamphetamine-induced conditioned place preference (CPP) is mediated by endocannabinoid system. Pharmacol Res. 2005;51:177–182. doi: 10.1016/j.phrs.2004.07.009. [DOI] [PubMed] [Google Scholar]
- Braida D, Pozzi M, Parolaro D, Sala M. Intracerebral self-administration of the cannabinoid receptor agonist CP 55,940 in the rat: interaction with the opioid system. Neuroscience. 2001;413:227–234. doi: 10.1016/s0014-2999(01)00766-x. [DOI] [PubMed] [Google Scholar]
- Braida D, Sala M. Role of the endocannabinoid system in MDMA intra-cerebral self-administration in rats. Br J Pharmacol. 2002;136:1089–1092. doi: 10.1038/sj.bjp.0704825. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Breakey WR, Goodell H, Lorenz PC, McHugh PR. Hallucinogenic drugs as precipitants of schizophrenia. Psychol Med. 1974;4:255–261. doi: 10.1017/s0033291700042938. [DOI] [PubMed] [Google Scholar]
- Bredt BM, Higuera-Alhino D, Shade SB, Hebert SJ, McCune JM, Abrams DI. Short-term effects of cannabinoids on immune phenotype and function in HIV-1-infected patients. J Clin Pharmacol. 2002;42:82s–89s. doi: 10.1002/j.1552-4604.2002.tb06007.x. [DOI] [PubMed] [Google Scholar]
- Breivogel CS, Griffin G, Di Marzo V, Martin BR. Evidence for a new G protein-coupled cannabinoid receptor in mouse brain. Mol Pharmacol. 2001;60:155–163. [PubMed] [Google Scholar]
- Breivogel CS, Sim LJ, Childers SR. Regional differences in cannabinoid receptor/G-protein coupling in rat brain. J Pharmacol Exp Ther. 1997;282:1632–1642. [PubMed] [Google Scholar]
- Brenneisen R, Egli A, Elsohly MA, Henn V, Spiess Y. The effect of orally and rectally administered Δ9-tetrahydrocannabinol on spasticity: a pilot study with 2 patients. Int J Clin Pharmacol Ther. 1996;34:446–452. [PubMed] [Google Scholar]
- Bridges D, Ahmad K, Rice AS. The synthetic cannabinoid WIN55,212-2 attenuates hyperalgesia and allodynia in a rat model of neuropathic pain. Br J Pharmacol. 2001;133:586–594. doi: 10.1038/sj.bjp.0704110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Britton DR, Koob GF, Rivier J, Vale W. Intraventricular corticotropin-releasing factor enhances behavioral effects of novelty. Life Sci. 1982;31:363–367. doi: 10.1016/0024-3205(82)90416-7. [DOI] [PubMed] [Google Scholar]
- Brotchie JM. CB1 cannabinoid receptor signalling in Parkinson’s disease. Curr Opin Pharmacol. 2003;3:54–61. doi: 10.1016/s1471-4892(02)00011-5. [DOI] [PubMed] [Google Scholar]
- Brown A, Wise A. GlaxoSmithKline, assignee. Identification of modulators of gpr55 activity. US patent 20030113814. 2003 inventors. Jun 19, 2003. [Google Scholar]
- Brown B, Adams AJ, Haegerstrom-Portnoy G, Jones RT, Flom MC. Pupil size after use of marijuana and alcohol. Am J Ophthalmol. 1977;83:350–354. doi: 10.1016/0002-9394(77)90732-2. [DOI] [PubMed] [Google Scholar]
- Brown TT, Dobs AS. Endocrine effects of marijuana. J Clin Pharmacol. 2002;42:90S–96S. doi: 10.1002/j.1552-4604.2002.tb06008.x. [DOI] [PubMed] [Google Scholar]
- Buchwald A, Derendorf H, Ji F, Nagaraja NY, Wu WM, Bodor N. Soft cannabinoid analogues as potential anti-glaucoma agents. Pharmazie. 2002;57:108–114. [PubMed] [Google Scholar]
- Buckley NE, McCoy KL, Mezey E, Bonner T, Zimmer A, Felder CC, Glass M, Zimmer A. Immunomodulation by cannabinoids is absent in mice deficient for the cannabinoid CB2 receptor. Eur J Pharmacol. 2000;396:141–149. doi: 10.1016/s0014-2999(00)00211-9. [DOI] [PubMed] [Google Scholar]
- Buggy DJ, Toogood L, Maric S, Sharpe P, Lambert DG, Rowbotham DJ. Lack of analgesic efficacy of oral Δ-9-tetrahydrocannabinol in postoperative pain. Pain. 2003;106:169–172. doi: 10.1016/s0304-3959(03)00331-2. [DOI] [PubMed] [Google Scholar]
- Buonamici M, Young GA, Khazan N. Effects of acute Δ9-THC administration on EEG and EEG power spectra in the rat. Neuropharmacology. 1982;21:825–829. doi: 10.1016/0028-3908(82)90071-5. [DOI] [PubMed] [Google Scholar]
- Burdyga G, Lal S, Varro A, Dimaline R, Thompson DG, Dockray DG. Expression of cannabinoid CB1 receptors by vagal afferent neurons is inhibited by cholecystokinin. J Neurosci. 2004;24:2708–2715. doi: 10.1523/JNEUROSCI.5404-03.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burnette-Curley D, Cabral GA. Differential inhibition of RAW264.7 macrophage tumoricidal activity by Δ9tetrahydrocannabinol: differential inhibition of RAW264.7 macrophage tumoricidal activity by Δ9tetrahydrocannabinol. Proc Soc Exp Biol Med. 1995;210:64–76. doi: 10.3181/00379727-210-43926. [DOI] [PubMed] [Google Scholar]
- Burns TL, Ineck JR. Cannabinoid analgesia as a potential new therapeutic option in the treatment of chronic pain. Ann Pharmacother. 2006;40:251–260. doi: 10.1345/aph.1G217. [DOI] [PubMed] [Google Scholar]
- Burstein S. Ajulemic acid (IP-751): synthesis, proof of principle, toxicity studies, and clinical trials. AAPS J. 2005;7:E143–E148. doi: 10.1208/aapsj070115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burstein SH. Ajulemic acid (CT3): a potent analog of the acid metabolites of THC. Curr Pharm Des. 2000;6:1339–1345. doi: 10.2174/1381612003399473. [DOI] [PubMed] [Google Scholar]
- Burstein SH, Audette CA, Breuer A, Devane WA, Colodner S, Doyle SA, Mechoulam R. Synthetic nonpsychotropic cannabinoids with potent antiinflammatory, analgesic, and leukocyte antiadhesion activities. J Med Chem. 1992;35:3135–3141. doi: 10.1021/jm00095a007. [DOI] [PubMed] [Google Scholar]
- Burstein SH, Friderichs E, Kogel B, Schneider J, Selve N. Analgesic effects of 1′,1′ dimethylheptyl-Δ8-THC-11-oic acid (CT3) in mice. Life Sci. 1998;63:161–168. doi: 10.1016/s0024-3205(98)00256-2. [DOI] [PubMed] [Google Scholar]
- Burstein SH, Karst M, Schneider U, Zurier RB. Ajulemic acid: a novel cannabinoid produces analgesia without a “high. Life Sci. 2004;75:1513–1522. doi: 10.1016/j.lfs.2004.04.010. [DOI] [PubMed] [Google Scholar]
- Buxbaum DM. Analgesic activity of Δ9-tetrahydrocannabinol in the rat and mouse. Psychopharmacology. 1972;25:275–280. doi: 10.1007/BF00422507. [DOI] [PubMed] [Google Scholar]
- Cabral GA, Fischer-Stenger K. Inhibition of macrophage inducible protein expression by Δ-9-tetrahydrocannabinol. Life Sci. 1994;54:1831–1844. doi: 10.1016/0024-3205(94)90122-8. [DOI] [PubMed] [Google Scholar]
- Cabral GA, Mishkin EM, Marciano-Cabral F, Coleman P, Harris L, Munson AE. Effect of Δ9-tetrahydrocannabinol on herpes simplex virus type 2 vaginal infection in the guinea pig. Proc Soc Exp Biol Med. 1986;182:181–186. doi: 10.3181/00379727-182-42325. [DOI] [PubMed] [Google Scholar]
- Cabral GA, Toney DM, Fischer-Stenger K, Harrison MP, Marciano-Cabral F. Anandamide inhibits macrophage-mediated killing of tumor necrosis factor-sensitive cells. Life Sci. 1995;56:2065–2072. doi: 10.1016/0024-3205(95)00190-h. [DOI] [PubMed] [Google Scholar]
- Cadas H, di Tomaso E, Piomelli D. Occurrence and biosynthesis of endogenous cannabinoid precursor N-arachidonoyl phosphatidylethanolamine in rat brain. J Neurosci. 1997;17:1226–1242. doi: 10.1523/JNEUROSCI.17-04-01226.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caillé S, Parsons LH. Cannabinoid modulation of opiate reinforcement through the ventral striatopallidal pathway. Neuropsychopharmacology. 2006;31:804–813. doi: 10.1038/sj.npp.1300848. [DOI] [PubMed] [Google Scholar]
- Cainazzo MM, Ferrazza G, Mioni C, Bazzani C, Bertolini A, Guarini S. Cannabinoid CB1 receptor blockade enhances the protective effect of melanocortins in hemorrhagic shock in the rat. Eur J Pharmacol. 2002;441:91–97. doi: 10.1016/s0014-2999(02)01487-5. [DOI] [PubMed] [Google Scholar]
- Calignano A, Katona I, Desarnaud F, Giuffrida A, La Rana G, Mackie K, Freund TF, Piomelli D. Bidirectional control of airway responsiveness by endogenous cannabinoids. Nature (Lond) 2000;408:96–101. doi: 10.1038/35040576. [DOI] [PubMed] [Google Scholar]
- Calignano A, La Rana G, Beltramo M, Makriyannis A, Piomelli D. Potentiation of anandamide hypotension by the transport inhibitor: AM404. Eur J Pharmacol. 1997;337:R1–R2. doi: 10.1016/s0014-2999(97)01297-1. [DOI] [PubMed] [Google Scholar]
- Calignano A, La Rana G, Giuffrida A, Piomelli D. Control of pain initiation by endogenous cannabinoids. Nature (Lond) 1998;394:277–281. doi: 10.1038/28393. [DOI] [PubMed] [Google Scholar]
- Campbell AD, McBride WJ. Serotonin-3 receptor and ethanol-stimulated dopamine release in the nucleus accumbens. Pharmacol Biochem Behav. 1995;51:835–842. doi: 10.1016/0091-3057(95)00050-7. [DOI] [PubMed] [Google Scholar]
- Campbell FA, Tramer MR, Carroll D, Reynolds DJ, Moore RA, McQuay HJ. Are cannabinoids an effective and safe treatment option in the management of pain? A qualitative systematic review. BMJ. 2001;323:13–16. doi: 10.1136/bmj.323.7303.13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cani PD, Montoya ML, Neyrinck AM, Delzenne NM, Lambert DM. Potential modulation of plasma ghrelin and glucagon-like peptide-1 by anorexigenoc cannabinoid compounds, SR141716A (rimonabant) and oleoylethanolamide. Br J Nutr. 2004;92:757–761. doi: 10.1079/bjn20041256. [DOI] [PubMed] [Google Scholar]
- Cannich A, Wotjak CT, Kamprath K, Hermann H, Lutz B, Marsicano G. CB1 cannabinoid receptors modulate kinase and phosphatase activity during extinction of conditioned fear in mice. Learn Mem. 2004;11:625–632. doi: 10.1101/lm.77904. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Capasso R, Matias I, Lutz B, Borrelli F, Capasso F, Marsicano G, Mascolo N, Petrosino S, Monory K, Valenti M, et al. Fatty acid amide hydrolase controls mouse intestinal motility in vivo. Gastroenterology. 2005;129:941–951. doi: 10.1053/j.gastro.2005.06.018. [DOI] [PubMed] [Google Scholar]
- Caplan GA, Brigham BA. Marijuana smoking and carcinoma of the tongue: is there an association? Cancer. 1990;66:1005–1006. doi: 10.1002/1097-0142(19900901)66:5<1005::aid-cncr2820660535>3.0.co;2-h. [DOI] [PubMed] [Google Scholar]
- Carlini EA, Mechoulam R, Lander N. Anticonvulsant activity of four oxygenated cannabidiol derivatives. Res Commun Chem Pathol Pharmacol. 1975;12:1–15. [PubMed] [Google Scholar]
- Carlisle SJ, Marciano-Cabral F, Staab A, Ludwick C, Cabral GA. Differential expression of the CB2 cannabinoid receptor by rodent macrophages and macrophage-like cells in relation to cell activation. Int Immunopharmacol. 2002;2:69–82. doi: 10.1016/s1567-5769(01)00147-3. [DOI] [PubMed] [Google Scholar]
- Carracedo A, Lorente M, Egia A, Blazquez C, Garcia S, Giroux V, Malicet C, Villuendas R, Gironella M, Gonzalez-Feria L, et al. The stress-regulated protein p8 mediates cannabinoid-induced apoptosis of tumor cells. Cancer Cell. 2006;9:301–312. doi: 10.1016/j.ccr.2006.03.005. [DOI] [PubMed] [Google Scholar]
- Carrier EJ, Auchampach JA, Hillard CJ. Inhibition of an equilibrative nucleoside transporter by cannabidiol: a mechanism of cannabinoid immunosuppression. Proc Natl Acad Sci USA. 2006;103:7895–7900. doi: 10.1073/pnas.0511232103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carter GT, Rosen BS. Marijuana in the management of amyotrophic lateral sclerosis. Am J Hosp Palliat Care. 2001;18:264–270. doi: 10.1177/104990910101800411. [DOI] [PubMed] [Google Scholar]
- Casanova ML, Blazquez C, Martinez-Palacio J, Villanueva C, Fernandez-Acenero MJ, Huffman JW, Jorcano JL, Guzman M. Inhibition of skin tumor growth and angiogenesis in vivo by activation of cannabinoid receptors. J Clin Investig. 2003;111:43–50. doi: 10.1172/JCI16116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Castane A, Valjent E, Ledent C, Parmentier M, Maldonado R, Valverde O. Lack of CB1 cannabinoid receptors modifies nicotine behavioural responses, but not nicotine abstinence. Neuropharmacology. 2002;43:857–867. doi: 10.1016/s0028-3908(02)00118-1. [DOI] [PubMed] [Google Scholar]
- Casu MA, Porcella A, Ruiu S, Saba P, Marchese G, Carai MA, Reali R, Gessa GL, Pani L. Differential distribution of functional cannabinoid CB1 receptors in the mouse gastroenteric tract. Eur J Pharmacol. 2003;459:97–105. doi: 10.1016/s0014-2999(02)02830-3. [DOI] [PubMed] [Google Scholar]
- Caulfield MP, Brown DA. Cannabinoid receptor agonists inhibit Ca current in NG108–15 neuroblastoma cells via a pertussis toxin-sensitive mechanism. Br J Pharmacol. 1992;106:231–232. doi: 10.1111/j.1476-5381.1992.tb14321.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cavero I, Buckley JP, Jandhyala BS. Parasympatholytic activity of (−)-9-trans-tetrahydrocannabinol in mongrel dogs. Eur J Pharmacol. 1972;19:301–304. doi: 10.1016/0014-2999(72)90027-1. [DOI] [PubMed] [Google Scholar]
- Centonze D, Gubellini P, Rossi S, Picconi B, Pisani A, Bernardi G, Calabresi P, Baunez C. Subthalamic nucleus lesion reverses motor abnormalities and striatal glutamatergic overactivity in experimental parkinsonism. Neuroscience. 2005;133:831–840. doi: 10.1016/j.neuroscience.2005.03.006. [DOI] [PubMed] [Google Scholar]
- Chambers AP, Sharkey KA, Koopmans HS. Cannabinoid (CB)1 receptor antagonist, AM 251, causes a sustained reduction of daily food intake in the rat. Physiol Behav. 2003;82:863–869. doi: 10.1016/j.physbeh.2004.07.004. [DOI] [PubMed] [Google Scholar]
- Chan CC, Hwang SJ, Lee FY, Wang SS, Chang FY, Li CP, Chu CJ, Lu RH, Lee SD. Prognostic value of plasma endotoxin levels in patients with cirrhosis. Scand J Gastroenterol. 1997;32:942–946. doi: 10.3109/00365529709011206. [DOI] [PubMed] [Google Scholar]
- Chang L, Luo L, Palmer JA, Sutton S, Wilson SJ, Barbier AJ, Breitenbucher JG, Chaplan SR, Webb M. Inhibition of fatty acid amide hydrolase produces analgesia by multiple mechanisms. Br J Pharmacol. 2006;148:102–113. doi: 10.1038/sj.bjp.0706699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chaperon F, Soubrie P, Puech AJ, Theibot MH. Involvement of central cannabinoid (CB1) receptors in the establishment of place conditioning in rats. Psychopharmacology. 1998;135:324–332. doi: 10.1007/s002130050518. [DOI] [PubMed] [Google Scholar]
- Chapman V. The cannabinoid CB1 receptor antagonist, SR141716A, selectively facilitates nociceptive responses of dorsal horn neurones in the rat. Br J Pharmacol. 1999;127:1765–1767. doi: 10.1038/sj.bjp.0702758. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheer JF, Marsden CA, Kendall DA, Mason R. Lack of response suppression follows repeated ventral tegmental cannabinoid administration: an in vitro electrophysiological study. Neuroscience. 2000;99:661–667. doi: 10.1016/s0306-4522(00)00241-4. [DOI] [PubMed] [Google Scholar]
- Chemin J, Monteil A, Perez-Reyes E, Nargeot J, Lory P. Direct inhibition of T-type calcium channels by the endogenous cannabinoid anandamide. EMBO (Eur Mol Biol Organ) J. 2001;20:7033–7040. doi: 10.1093/emboj/20.24.7033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen J, Matias I, Dinh T, Lu T, Venezia S, Nieves A, Woodward DF, Di Marzo V. Finding of endocannabinoids in human eye tissues: implications for glaucoma. Biochem Biophys Res Commun. 2005;330:1062–1067. doi: 10.1016/j.bbrc.2005.03.095. [DOI] [PubMed] [Google Scholar]
- Chen JP, Paredes W, Li J, Smith D, Lowinson J, Gardner EL. Δ9-Tetrahydrocannabinol produces naloxone-blockable enhancement of presynaptic basal dopamine efflux in nucleus accumbens of conscious, freely moving rats, as measured by intracerebral microdialysis. Psychopharmacology. 1990;102:156–162. doi: 10.1007/BF02245916. [DOI] [PubMed] [Google Scholar]
- Chen K, Ratzliff A, Hilgenberg L, Gulyas A, Freund TF, Smith M, Dinh TP, Piomelli D, Mackie K, Soltesz I. Long-term plasticity of endocannabinoid signaling induced by developmental febrile seizures. Neuron. 2003;39:599–611. doi: 10.1016/s0896-6273(03)00499-9. [DOI] [PubMed] [Google Scholar]
- Chen RZ, Huang RR, Shen CP, MacNeil DJ, Fong TM. Synergistic effects of cannabinoid inverse agonist AM251 and opioid antagonist nalmefene on food intake in mice. Brain Res. 2004;999:227–230. doi: 10.1016/j.brainres.2003.12.004. [DOI] [PubMed] [Google Scholar]
- Chen Y, Buck J. Cannabinoids protect cells from oxidative cell death: a receptor-independent mechanism. J Pharmacol Exp Ther. 2000;293:807–812. [PubMed] [Google Scholar]
- Cheng X, Leung SW, Lo LS, Pang CC. Selective versus non-selective suppression of nitric oxide synthase on regional hemodynamics in rats with or without LPS-induced endotoxemia. Naunyn-Schmiedeberg’s Arch Pharmacol. 2003;367:372–379. doi: 10.1007/s00210-002-0684-1. [DOI] [PubMed] [Google Scholar]
- Chiang K, Gerber AL, Sipe JC, Cravatt BF. Reduced cellular expression and activity of the P129T mutant of human fatty acid amide hydrolase: evidence for a link between defects in the endocannabinoid system and problem drug use. Hum Mol Genet. 2004;13:1–7. doi: 10.1093/hmg/ddh216. [DOI] [PubMed] [Google Scholar]
- Chien FY, Wang RF, Mittag TW, Podos SM. Effect of WIN 55212-2, a cannabinoid receptor agonist, on aqueous humor dynamics in monkeys. Arch Ophthalmol. 2003;121:87–90. doi: 10.1001/archopht.121.1.87. [DOI] [PubMed] [Google Scholar]
- Chiu P, Olsen DM, Borys HK, Karler R, Turkanis SA. The influence of cannabidiol and Δ9-tetrahydrocannabinol on cobalt epilepsy in rats. Epilepsia. 1979;20:365–375. doi: 10.1111/j.1528-1157.1979.tb04816.x. [DOI] [PubMed] [Google Scholar]
- Chopra GS, Smith JW. Psychotic reactions following cannabis use in East Indians. Arch Gen Psychiatry. 1974;30:24–27. doi: 10.1001/archpsyc.1974.01760070014002. [DOI] [PubMed] [Google Scholar]
- Cippitelli A, Bilbao A, Hansson AC, del Arco I, Sommer W, Heilig M, Massi M, Bermudez-Silva FJ, Navarro M, Ciccocioppo R, et al. The Cannabinoid CB1 receptor antagonism reduces conditioned reinstatement of ethanol-seeking behavior in rats. Eur J Neurosci. 2005;21:2243–2251. doi: 10.1111/j.1460-9568.2005.04056.x. [DOI] [PubMed] [Google Scholar]
- Clark JT, Kalra PS, Crowley WR, Kalra SP. Neuropeptide Y and human pancreatic polypeptide stimulate feeding behavior in rats. Endocrinology. 1984;115:427–429. doi: 10.1210/endo-115-1-427. [DOI] [PubMed] [Google Scholar]
- Clayton N, Marshall FH, Bountra C, O’Shaughnessy CT. CB1 and CB2 cannabinoid receptors are implicated in inflammatory pain. Pain. 2002;96:253–260. doi: 10.1016/S0304-3959(01)00454-7. [DOI] [PubMed] [Google Scholar]
- Clement AB, Hawkins EG, Lichtman AH, Cravatt BF. Increased seizure susceptibility and proconvulsant activity of anandamide in mice lacking fatty acid amide hydrolase. J Neurosci. 2003;23:3916–3923. doi: 10.1523/JNEUROSCI.23-09-03916.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clifford DB. Tetrahydrocannabinol for tremor in multiple sclerosis. Ann Neurol. 1983;13:669–671. doi: 10.1002/ana.410130616. [DOI] [PubMed] [Google Scholar]
- Cohen A, Perrault G, Griebel G, Soubrie P. Nicotine-associated cues maintain nicotine-seeking behavior in rats several weeks after nicotine withdrawal: reversal by the cannabinoid (CB1) receptor antagonist rimonabant (SR141716) Neuropsychopharmacology. 2005;30:145–155. doi: 10.1038/sj.npp.1300541. [DOI] [PubMed] [Google Scholar]
- Cohen C, Perrault G, Voltz C, Steinberg R, Soubrie P. SR141716, a central cannabinoid (CB1) receptor antagonist, blocks the motivational and dopamine-releasing effects of nicotine in rats. Behav Pharmacol. 2002;13:451–463. doi: 10.1097/00008877-200209000-00018. [DOI] [PubMed] [Google Scholar]
- Colasanti BK, Lindamood C, 3rd, Craig CR. Effects of marihuana cannabinoids on seizure activity in cobalt-epileptic rats. Pharmacol Biochem Behav. 1982;16:573–578. doi: 10.1016/0091-3057(82)90418-x. [DOI] [PubMed] [Google Scholar]
- Colasanti BK, Brown RE, Craig CR. Ocular hypotension, ocular toxicity, and neurotoxicity in response to marihuana extract and cannabidiol. Gen Pharmacol. 1984a;15:479–484. doi: 10.1016/0306-3623(84)90202-7. [DOI] [PubMed] [Google Scholar]
- Colasanti BK, Craig CR, Allara RD. Intraocular pressure, ocular toxicity and neurotoxicity after administration of cannabinol or cannabigerol. Exp Eye Res. 1984b;39:251–259. doi: 10.1016/0014-4835(84)90013-7. [DOI] [PubMed] [Google Scholar]
- Colasanti BK, Powell SR, Craig CR. Intraocular pressure, ocular toxicity and neurotoxicity after administration of Δ9-tetrahydrocannabinol or cannabichromene. Exp Eye Res. 1984c;38:63–71. doi: 10.1016/0014-4835(84)90139-8. [DOI] [PubMed] [Google Scholar]
- Colombo G, Agabio R, Diaz G, Lobina C, Reali R, Gessa GL. Appetite suppression and weight loss after the cannabinoid antagonist SR 141716. Life Sci. 1998a;63:PL113–PL117. doi: 10.1016/s0024-3205(98)00322-1. [DOI] [PubMed] [Google Scholar]
- Colombo G, Agabio R, Fa M, Guano L, Lobina C, Loche A, Reali R, Gessa GL. Reduction of voluntary ethanol intake in ethanol preferring sP rats by the cannabinoid antagonist SR141716A. Alcohol Alcohol. 1998b;33:126–130. doi: 10.1093/oxfordjournals.alcalc.a008368. [DOI] [PubMed] [Google Scholar]
- Colombo G, Serra S, Brunetti G, Gomez R, Melis S, Vacca G, Carai MM, Gessa L. Stimulation of voluntary ethanol intake by cannabinoid receptor agonists in ethanol-preferring sP rats. Psychopharmacology. 2002;159:181–187. doi: 10.1007/s002130100887. [DOI] [PubMed] [Google Scholar]
- Colombo G, Serra S, Vacca G, Carai MAM, Gessa L. Endocannabinoid system and alcohol addiction: pharmacological studies. Pharmacol Biochem Behav. 2005;81:369–380. doi: 10.1016/j.pbb.2005.01.022. [DOI] [PubMed] [Google Scholar]
- Colombo G, Serra S, Vacca G, Gessa L, Carai MAM. Suppression by baclofen of the stimulation of alcohol intake induced by morphine and WIN 55,212-2 in alcohol-preferring rats. Eur J Pharmacol. 2004;492:189–193. doi: 10.1016/j.ejphar.2004.03.065. [DOI] [PubMed] [Google Scholar]
- Compston A, Coles A. Multiple sclerosis. Lancet. 2002;359:1221–1231. doi: 10.1016/S0140-6736(02)08220-X. [DOI] [PubMed] [Google Scholar]
- Consroe P. Brain cannabinoid systems as targets for the therapy of neurological disorders. Neurobiol Dis. 1998;5:534–551. doi: 10.1006/nbdi.1998.0220. [DOI] [PubMed] [Google Scholar]
- Consroe P, Laguna J, Allender J, Snider S, Stern L, Sandyk R, Kennedy K, Schram K. Controlled clinical trial of cannabidiol in Huntington’s disease. Pharmacol Biochem Behav. 1991;40:701–708. doi: 10.1016/0091-3057(91)90386-g. [DOI] [PubMed] [Google Scholar]
- Consroe P, Martin A, Singh V. Antiepileptic potential of cannabidiol analogs. J Clin Pharmacol. 1981;21:428S–436S. doi: 10.1002/j.1552-4604.1981.tb02623.x. [DOI] [PubMed] [Google Scholar]
- Consroe P, Mechoulam R. Anticonvulsant and neurotoxic effects of tetrahydrocannabinol stereoisomers. NIDA Res Monogr. 1987;79:59–66. [PubMed] [Google Scholar]
- Consroe P, Musty R, Rein J, Tillery W, Pertwee R. The perceived effects of smoked cannabis on patients with multiple sclerosis. Eur Neurol. 1997;38:44–48. doi: 10.1159/000112901. [DOI] [PubMed] [Google Scholar]
- Consroe P, Wolkin A. Cannabidiol-antiepileptic drug comparisons and interactions in experimentally induced seizures in rats. J Pharmacol Exp Ther. 1977;201:26–32. [PubMed] [Google Scholar]
- Consroe PF, Wood GC, Buchsbaum H. Anticonvulsant nature of marihuana smoking. J Am Med Assoc. 1975;234:306–307. [PubMed] [Google Scholar]
- Contassot E, Tenan M, Schnuriger V, Pelte MF, Dietrich PY. Arachidonyl ethanolamide induces apoptosis of uterine cervix cancer cells via aberrantly expressed vanilloid receptor-1. Gynecol Oncol. 2004a;93:182–188. doi: 10.1016/j.ygyno.2003.12.040. [DOI] [PubMed] [Google Scholar]
- Contassot E, Wilmotte R, Tenan M, Belkouch MC, Schnuriger V, de Tribolet N, Burkhardt K, Dietrich PY. Arachidonylethanolamide induces apoptosis of human glioma cells through vanilloid receptor-1. J Neuropathol Exp Neurol. 2004b;63:956–963. doi: 10.1093/jnen/63.9.956. [DOI] [PubMed] [Google Scholar]
- Corcoran ME, McCaughran JA, Jr, Wada JA. Antiepileptic and prophylactic effects of tetrahydrocannabinols in amygdaloid kindled rats. Epilepsia. 1978;19:47–55. doi: 10.1111/j.1528-1157.1978.tb05011.x. [DOI] [PubMed] [Google Scholar]
- Correa F, Mestre L, Molina-Holgado E, Arevalo-Martin A, Docagne F, Romero E, Molina-Holgado F, Borrell J, Guaza C. The role of cannabinoid system on immune modulation: therapeutic implications on CNS inflammation. Mini Rev Med Chem. 2005;5:671–675. doi: 10.2174/1389557054368790. [DOI] [PubMed] [Google Scholar]
- Coruzzi G, Adami M, Coppelli G, Frati P, Soldani G. Inhibitory effect of the cannabinoid receptor agonist WIN 55,212-2 on pentagastrin-induced gastric acid secretion in the anaesthetized rat. Naunyn-Schmiedeberg’s Arch Pharmacol. 1999;360:715–718. doi: 10.1007/s002109900135. [DOI] [PubMed] [Google Scholar]
- Cossu G, Ledent C, Fattore L, Imperato A, Bohme GA, Parmentier M, Fratta W. Cannabinoid CB1 receptor knockout mice fail to self-administer morphine but not other drugs of abuse. Behav Brain Res. 2001;118:61–65. doi: 10.1016/s0166-4328(00)00311-9. [DOI] [PubMed] [Google Scholar]
- Costa B, Colleoni M, Conti S, Parolaro D, Franke C, Trovato AE, Giagnoni G. Oral anti-inflammatory activity of cannabidiol, a non-psychoactive constituent of cannabis, in acute carrageenan-induced inflammation in the rat paw. Naunyn-Schmiedeberg’s Arch Pharmacol. 2004a;369:294–299. doi: 10.1007/s00210-004-0871-3. [DOI] [PubMed] [Google Scholar]
- Costa B, Colleoni M, Conti S, Trovato AE, Bianchi M, Sotgiu ML, Giagnoni G. Repeated treatment with the synthetic cannabinoid WIN 55,212-2 reduces both hyperalgesia and production of pronociceptive mediators in a rat model of neuropathic pain. Br J Pharmacol. 2004b;141:4–8. doi: 10.1038/sj.bjp.0705587. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cota D, Marsicano G, Tschop M, Grubler Y, Flachskamm C, Schubert M, Auer D, Yassouridis A, Thone-Reineke C, Ortmann S, et al. The endogenous cannabinoid system affects energy balance via central orexigenic drive and peripheral lipogenesis. J Clin Investig. 2003;112:423–431. doi: 10.1172/JCI17725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coutts AA, Irving AJ, Mackie K, Pertwee RG, Anavi-Goffer S. Localisation of cannabinoid CB1 receptor immunoreactivity in the guinea pig and rat myenteric plexus. J Comp Neurol. 2002;448:410–422. doi: 10.1002/cne.10270. [DOI] [PubMed] [Google Scholar]
- Coutts AA, Izzo AA. The gastrointestinal pharmacology of cannabinoids: an update. Curr Opin Pharmacol. 2004;4:572–579. doi: 10.1016/j.coph.2004.05.007. [DOI] [PubMed] [Google Scholar]
- Coutts AA, Pertwee RG. Inhibition by cannabinoid receptor agonists of acetylcholine release from the guinea-pig myenteric plexus. Br J Pharmacol. 1997;121:1557–1566. doi: 10.1038/sj.bjp.0701301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cox ML, Welch SP. The antinociceptive effect of Δ9-tetrahydrocannabinol in the arthritic rat. Eur J Pharmacol. 2004;493:65–74. doi: 10.1016/j.ejphar.2004.04.022. [DOI] [PubMed] [Google Scholar]
- Coyle JT, Puttfarcken P. Oxidative stress, glutamate, and neurodegenerative disorders. Science (Wash DC) 1993;262:689–695. doi: 10.1126/science.7901908. [DOI] [PubMed] [Google Scholar]
- Coyne L, Lees G, Nicholson RA, Zheng J, Neufield KD. The sleep hormone oleamide modulates inhibitory ionotropic receptors in mammalian CNS in vitro. Br J Pharmacol. 2002;135:1977–1987. doi: 10.1038/sj.bjp.0704651. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cravatt BF, Demarest K, Patricelli MP, Bracey MH, Giang DK, Martin BR, Lichtman AH. Supersensitivity to anandamide and enhanced endogenous cannabinoid signaling in mice lacking fatty acid amide hydrolase. Proc Natl Acad Sci USA. 2001;98:9371–9376. doi: 10.1073/pnas.161191698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cravatt BF, Giang DK, Mayfield SP, Boger DL, Lerner RA, Gilula NB. Molecular characterization of an enzyme that degrades neuromodulatory fatty-acid amides. Nature (Lond) 1996;384:83–87. doi: 10.1038/384083a0. [DOI] [PubMed] [Google Scholar]
- Cravatt BF, Prospero-Garcia O, Siuzdak G, Gilula NB, Henriksen SJ, Boger DL, Lerner RA. Chemical characterization of a family of brain lipids that induce sleep. Science (Wash DC) 1995;268:1506–1509. doi: 10.1126/science.7770779. [DOI] [PubMed] [Google Scholar]
- Cravatt BF, Saghatelian A, Hawkins EG, Clement AB, Bracey MH, Lichtman AH. Functional disassociation of the central and peripheral fatty acid amide signaling systems. Proc Natl Acad Sci USA. 2004;101:10821–10826. doi: 10.1073/pnas.0401292101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crawford WJ, Merritt JC. Effects of tetrahydrocannabinol on arterial and intraocular hypertension. Int J Clin Pharmacol Biopharm. 1979;17:191–196. [PubMed] [Google Scholar]
- Crawley JN, Corwin RL, Robinson JK, Felder CC, Devane WA, Axelrod J. Anandamide, an endogenous ligand of the cannabinoid receptor, induces hypomotility and hypothermia in vivo in rodents. Pharmacol Biochem Behav. 1993;46:967–972. doi: 10.1016/0091-3057(93)90230-q. [DOI] [PubMed] [Google Scholar]
- Croci T, Landi M, Galzin AM, Marini P. Role of cannabinoid CB1 receptors and tumor necrosis factor-α in the gut and systemic anti-inflammatory activity of SR 141716 (rimonabant) in rodents. Br J Pharmacol. 2003;140:115–122. doi: 10.1038/sj.bjp.0705412. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crowston JG, Weinreb RN. Glaucoma medication and aqueous humor dynamics. Curr Opin Ophthalmol. 2005;16:94–100. doi: 10.1097/01.icu.0000156136.20570.eb. [DOI] [PubMed] [Google Scholar]
- Croxford JL. Therapeutic potential of cannabinoids in CNS disease. CNS Drugs. 2003;17:179–202. doi: 10.2165/00023210-200317030-00004. [DOI] [PubMed] [Google Scholar]
- Croxford JL, Miller SD. Immunoregulation of a viral model of multiple sclerosis using the synthetic cannabinoid R+WIN55,212. J Clin Investig. 2003;111:1231–1240. doi: 10.1172/JCI17652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Croxford JL, Yamamura T. Cannabinoids and the immune system: potential for the treatment of inflammatory diseases? J Neuroimmunol. 2005;166:3–18. doi: 10.1016/j.jneuroim.2005.04.023. [DOI] [PubMed] [Google Scholar]
- Csiszar A, Pacher P, Kaley G, Ungvari Z. Role of oxidative and nitrosative stress, longevity genes and poly(ADP-ribose) polymerase in cardiovascular dysfunction associated with aging. Curr Vasc Pharmacol. 2005;3:285–291. doi: 10.2174/1570161054368616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cuendet JF, Shapiro D, Calanca A, Faggioni R, Ducrey N. Action of Δ-9-tetrahydrocannabinol on ophthalmotonus. Ophthalmologica. 1976;172:122–127. doi: 10.1159/000307682. [DOI] [PubMed] [Google Scholar]
- Cunha JM, Carlini EA, Pereira AE, Ramos OL, Pimentel C, Gagliardi R, Sanvito WL, Lander N, Mechoulam R. Chronic administration of cannabidiol to healthy volunteers and epileptic patients. Pharmacology. 1980;21:175–185. doi: 10.1159/000137430. [DOI] [PubMed] [Google Scholar]
- Curtis MA, Faull RL, Glass M. A novel population of progenitor cells expressing cannabinoid receptors in the subependymal layer of the adult normal and Huntington’s disease human brain. J Chem Neuroanat. 2006;31:210–215. doi: 10.1016/j.jchemneu.2006.01.005. [DOI] [PubMed] [Google Scholar]
- Dajani EZ, Larsen KR, Taylor J, Dajani NE, Shahwan TG, Neeleman SD, Taylor MS, Dayton MT, Mir GN. 1′,1′-Dimethylheptyl-Δ-8-tetrahydrocannabinol-11-oic acid: a novel, orally effective cannabinoid with analgesic and anti-inflammatory properties. J Pharmacol Exp Ther. 1999;291:31–38. [PubMed] [Google Scholar]
- Dale LC, Anthenelli RM. Rimonabant as an aid to smoking cessation in smokers motivated to quit: results from a U.S. Multicenter Study—STRATUS U.S. Trial. Proceedings of the Annual Scientific Session of the American College of Cardiology; 2004 March 7–10; New Orleans, LA. 2004. [Google Scholar]
- Dannon PN, Lowengrub K, Amiaz R, Grunhaus L, Kotler M. Comorbid cannabis use and panic disorder: short term and long term follow-up study. Hum Psychopharmacol. 2004;19:97–101. doi: 10.1002/hup.560. [DOI] [PubMed] [Google Scholar]
- D’Argenio G, Valenti M, Scaglione G, Cosenza V, Sorrentini I, Di Marzo V. Up-regulation of anandamide levels as an endogenous mechanism and a pharmacological strategy to limit colon inflammation. FASEB J. 2006;20:568–570. doi: 10.1096/fj.05-4943fje. [DOI] [PubMed] [Google Scholar]
- Darmani NA. Δ9-Tetrahydrocannabinol and synthetic cannabinoids prevent emesis produced by the cannabinoid CB1 receptor antagonist/inverse agonist SR 141716A. Neuropsychopharmacology. 2001a;24:198–203. doi: 10.1016/S0893-133X(00)00197-4. [DOI] [PubMed] [Google Scholar]
- Darmani NA. Δ-9-Tetrahydrocannabinol differentially suppresses cisplatin-induced emesis and indices of motor function via cannabinoid CB1 receptors in the least shrew. Pharmacol Biochem Behav. 2001b;69:239–249. doi: 10.1016/s0091-3057(01)00531-7. [DOI] [PubMed] [Google Scholar]
- Das SK, Paria BC, Chakraborty I, Dey SK. Cannabinoid ligand-receptor signaling in the mouse uterus. Proc Natl Acad Sci USA. 1995;92:4332–4336. doi: 10.1073/pnas.92.10.4332. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davis MI, Ronesi J, Lovinger DM. A predominant role for inhibition of the adenylate cyclase/protein kinase A pathway in ERK activation by cannabinoid receptor 1 in N1E-115 neuroblastoma cells. J Biol Chem. 2003;278:48973–48980. doi: 10.1074/jbc.M305697200. [DOI] [PubMed] [Google Scholar]
- Dean B, Sundram S, Bradbury R, Scarr E, Copolov D. Studies on [3H]CP-55940 binding in the human central nervous system: regional specific changes in density of cannabinoid-1 receptors associated with schizophrenia and cannabis use. Neuroscience. 2001;103:9–15. doi: 10.1016/s0306-4522(00)00552-2. [DOI] [PubMed] [Google Scholar]
- de Lago E, de Miguel R, Lastres-Becker I, Ramos JA, Fernandez-Ruiz J. Involvement of vanilloid-like receptors in the effects of anandamide on motor behavior and nigrostriatal dopaminergic activity: in vivo and in vitro evidence. Brain Res. 2004a;1007:152–159. doi: 10.1016/j.brainres.2004.02.016. [DOI] [PubMed] [Google Scholar]
- de Lago E, Fernandez-Ruiz J, Ortega-Gutierrez S, Cabranes A, Pryce G, Baker D, Lopez-Rodriguez M, Ramos JA. UCM707, an inhibitor of the anandamide uptake, behaves as a symptom control agent in models of Huntington’s disease and multiple sclerosis, but fails to delay/arrest the progression of different motor-related disorders. Eur Neuropsychopharmacol. 2006;16:7–18. doi: 10.1016/j.euroneuro.2005.06.001. [DOI] [PubMed] [Google Scholar]
- de Lago E, Fernandez-Ruiz J, Ortega-Gutierrez S, Viso A, Lopez-Rodriguez ML, Ramos JA. UCM707, a potent and selective inhibitor of endocannabinoid uptake, potentiates hypokinetic and antinociceptive effects of anandamide. Eur J Pharmacol. 2002;449:99–103. doi: 10.1016/s0014-2999(02)01996-9. [DOI] [PubMed] [Google Scholar]
- de Lago E, Ligresti A, Ortar G, Morera E, Cabranes A, Pryce G, Bifulco M, Baker D, Fernandez-Ruiz J, Di Marzo V. In vivo pharmacological actions of two novel inhibitors of anandamide cellular uptake. Eur J Pharmacol. 2004b;484:249–257. doi: 10.1016/j.ejphar.2003.11.027. [DOI] [PubMed] [Google Scholar]
- de Lago E, Urbani P, Ramos JA, Di Marzo V, Fernandez-Ruiz J. Arvanil, a hybrid endocannabinoid and vanilloid compound, behaves as an antihyperkinetic agent in a rat model of Huntington’s disease. Brain Res. 2005;1050:210–216. doi: 10.1016/j.brainres.2005.05.024. [DOI] [PubMed] [Google Scholar]
- De Marchi N, De Petrocellis L, Orlando P, Daniele F, Fezza F, Di Marzo V. Endocannabinoid signalling in the blood of patients with schizophrenia. Lipids Health Dis. 2003;2:5. doi: 10.1186/1476-511X-2-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Petrocellis L, Bisogno T, Davis JB, Pertwee RG, Di Marzo V. Overlap between the ligand recognition properties of the anandamide transporter and the VR1 vanilloid receptor: inhibitors of anandamide uptake with negligible capsaicin-like activity. FEBS Lett. 2000;483:52–56. doi: 10.1016/s0014-5793(00)02082-2. [DOI] [PubMed] [Google Scholar]
- De Petrocellis L, Melck D, Palmisano A, Bisogno T, Laezza C, Bifulco M, Di Marzo V. The endogenous cannabinoid anandamide inhibits human breast cancer cell proliferation. Proc Natl Acad Sci USA. 1998;95:8375–8380. doi: 10.1073/pnas.95.14.8375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Vries TJ, Schoffelmeer AN. Cannabinoid CB1 receptors control conditioned drug seeking. Trends Pharmacol Sci. 2005;26:420–426. doi: 10.1016/j.tips.2005.06.002. [DOI] [PubMed] [Google Scholar]
- De Vries TJ, Shaham Y, Homberg JR, Crombag H, Schuurman K, Dieben J, Vander-schuren LJ, Schoffelmeer AN. A cannabinoid mechanism of relapse to cocaine seeking. Nat Med. 2001;7:1151–1154. doi: 10.1038/nm1001-1151. [DOI] [PubMed] [Google Scholar]
- del Carmen Godino M, Torres M, Sanchez-Prieto J. The modulation of Ca2+ and K+ channels but not changes in cAMP signaling contribute to the inhibition of glutamate release by cannabinoid receptors in cerebrocortical nerve terminals. Neuropharmacology. 2005;48:547–557. doi: 10.1016/j.neuropharm.2004.11.012. [DOI] [PubMed] [Google Scholar]
- Denovan-Wright EM, Robertson HA. Cannabinoid receptor messenger RNA levels decrease in a subset of neurons of the lateral striatum, cortex and hippocampus of transgenic Huntington’s disease mice. Neuroscience. 2000;98:705–713. doi: 10.1016/s0306-4522(00)00157-3. [DOI] [PubMed] [Google Scholar]
- Derkinderen P, Ledent C, Parmentier M, Girault JA. Cannabinoids activate p38 mitogen-activated protein kinases through CB1 receptors in hippocampus. J Neurochem. 2001;77:957–960. doi: 10.1046/j.1471-4159.2001.00333.x. [DOI] [PubMed] [Google Scholar]
- Deroche-Gamonet V, Le Moal M, Piazza PV, Soubrie P. SR141716, a CB1 receptor antagonist, decreases the sensitivity to the reinforcing effects of electrical brain stimulation in rats. Psychoharmacology. 2001;157:254–259. doi: 10.1007/s002130100804. [DOI] [PubMed] [Google Scholar]
- Després J-P, Golay A, Sjöström L for the Rimonabant in Obesity-Lipids Study Group. Effects of rimonabant on metabolic risk factors in overweight patients with dyslipidemia. N Engl J Med. 2005;353:2121–2134. doi: 10.1056/NEJMoa044537. [DOI] [PubMed] [Google Scholar]
- Deutsch DG, Lin S, Hill WA, Morse KL, Salehani D, Arreaza G, Omeir RL, Makriyannis A. Fatty acid sulfonyl fluorides inhibit anandamide metabolism and bind to the cannabinoid receptor. Biochem Biophys Res Commun. 1997a;231:217–221. doi: 10.1006/bbrc.1997.6072. [DOI] [PubMed] [Google Scholar]
- Deutsch DG, Omeir R, Arreaza G, Salehani D, Prestwich GD, Huang Z, Howlett A. Methyl arachidonyl fluorophosphonate: a potent irreversible inhibitor of anandamide amidase. Biochem Pharmacol. 1997b;53:255–260. doi: 10.1016/s0006-2952(96)00830-1. [DOI] [PubMed] [Google Scholar]
- Devane WA, Dysarz FA, III, Johnson MR, Melvin LS, Howlett AC. Determination and characterization of a cannabinoid receptor in rat brain. Mol Pharmacol. 1988;34:605–613. [PubMed] [Google Scholar]
- Devane WA, Hanus L, Breuer A, Pertwee RG, Stevenson LA, Griffin G, Gibson D, Mandelbaum A, Etinger A, Mechoulame R. Isolation and structure of a brain constituent that binds to the cannabinoid receptor. Science (Wash DC) 1992;258:1946–1949. doi: 10.1126/science.1470919. [DOI] [PubMed] [Google Scholar]
- Dewey WL. Cannabinoid pharmacology. Pharmacol Rev. 1986;38:151–178. [PubMed] [Google Scholar]
- Di S, Boudaba C, Popescu R, Weng FJ, Harris C, Marcheselli VL, Bazan NG, Tasker JG. Activity-dependent release and actions of endocannabinoids in the rat hypothalamic supraoptic nucleus. J Physiol (Lond) 2005a;569:751–760. doi: 10.1113/jphysiol.2005.097477. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Di S, Malcher-Lopes R, Halmos KC, Tasker JG. Nongenomic glucocorticoid inhibition via endocannabinoid release in the hypothalamus: a fast feedback mechanism. J Neurosci. 2003;23:4850–4857. doi: 10.1523/JNEUROSCI.23-12-04850.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Di S, Malcher-Lopes R, Marcheselli VL, Bazan NG, Tasker JG. Rapid glucocorticoid-mediated endocannabinoid release and opposing regulation of glutamate and γ-aminobutyric acid inputs to hypothalamic magnocellular neurons. Endocrinology. 2005b;146:4292–4301. doi: 10.1210/en.2005-0610. [DOI] [PubMed] [Google Scholar]
- Di Carlo G, Izzo AA. Cannabinoids for gastrointestinal diseases: potential therapeutic applications. Expert Opin Investig Drugs. 2003;12:39–49. doi: 10.1517/13543784.12.1.39. [DOI] [PubMed] [Google Scholar]
- Di Filippo C, Rossi F, Rossi S, D’Amico M. Cannabinoid CB2 receptor activation reduces mouse myocardial ischemia-reperfusion injury: involvement of cytokine/chemokines and PMN. J Leukoc Biol. 2004;75:453–459. doi: 10.1189/jlb.0703303. [DOI] [PubMed] [Google Scholar]
- Di Marzo V, Bifulco M, De Petrocellis L. The endocannabinoid system and its therapeutic exploitation. Nat Rev Drug Discov. 2004;3:771–784. doi: 10.1038/nrd1495. [DOI] [PubMed] [Google Scholar]
- Di Marzo V, Bisogno T, De Petrocellis L, Brandi I, Jefferson RG, Winckler RL, Davis JB, Dasse O, Mahadevan A, Razdan RK, et al. Highly selective CB1 cannabinoid receptor ligands and novel CB1/VR1 vanilloid receptor “hybrid” ligands. Biochem Biophys Res Commun. 2001a;281:444–451. doi: 10.1006/bbrc.2001.4354. [DOI] [PubMed] [Google Scholar]
- Di Marzo V, Fontana A, Cadas H, Schinelli S, Cimino G, Schwartz JC, Piomelli D. Formation and inactivation of endogenous cannabinoid anandamide in central neurons. Nature (Lond) 1994;372:686–691. doi: 10.1038/372686a0. [DOI] [PubMed] [Google Scholar]
- Di Marzo V, Goparaju SK, Wang L, Liu J, Bátkai S, Járai Z, Fezza F, Miura GI, Palmiter RD, Sugiura T, et al. Leptin-regulated endocannabinoids are involved in maintaining food intake. Nature (Lond) 2001b;410:822–825. doi: 10.1038/35071088. [DOI] [PubMed] [Google Scholar]
- Di Marzo V, Hill MP, Bisogno T, Crossman AR, Brotchie JM. Enhanced levels of endogenous cannabinoids in the globus pallidus are associated with a reduction in movement in an animal model of Parkinson’s disease. FASEB J. 2000;14:1432–1438. doi: 10.1096/fj.14.10.1432. [DOI] [PubMed] [Google Scholar]
- Di Marzo V, Matias I. Endocannabinoid control of food intake and energy balance. Nat Neurosci. 2005;8:585–589. doi: 10.1038/nn1457. [DOI] [PubMed] [Google Scholar]
- Di Marzo V, Petrocellis LD. Plant, synthetic, and endogenous cannabinoids in medicine. Annu Rev Med. 2006;57:553–574. doi: 10.1146/annurev.med.57.011205.135648. [DOI] [PubMed] [Google Scholar]
- Dinh TP, Carpenter D, Leslie FM, Freund TF, Katona I, Sensi SL, Kathuria S, Piomelli D. Brain monoglyceride lipase participating in endocannabinoid inactivation. Proc Natl Acad Sci USA. 2002a;99:10819–10824. doi: 10.1073/pnas.152334899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dinh TP, Freund TF, Piomelli D. A role for monoglyceride lipase in 2-arachidonoylglycerol inactivation. Chem Phys Lipids. 2002b;121:149–158. doi: 10.1016/s0009-3084(02)00150-0. [DOI] [PubMed] [Google Scholar]
- Domenicali M, Ros J, Fernandez-Varo G, Cejudo-Martin P, Crespo M, Morales-Ruiz M, Briones AM, Campistol JM, Arroyo V, Vila E, et al. Increased anandamide induced relaxation in mesenteric arteries of cirrhotic rats: role of cannabinoid and vanilloid receptors. Gut. 2005;54:522–527. doi: 10.1136/gut.2004.051599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Donovan M. On the physical and medicinal qualities of Indian hemp (Cannabis indica) Dublin J Med Sci. 1845;26:368–402. [Google Scholar]
- Drew LJ, Harris J, Millns PJ, Kendall DA, Chapman V. Activation of spinal cannabinoid 1 receptors inhibits C-fibre driven hyperexcitable neuronal responses and increases [35S]GTPγS binding in the dorsal horn of the spinal cord of noninflamed and inflamed rats. Eur J Neurosci. 2000;12:2079–2086. doi: 10.1046/j.1460-9568.2000.00101.x. [DOI] [PubMed] [Google Scholar]
- Drmota T, Greasley P, Groblewski T. inventors, AstraZeneca, assignee. Screening assays for cannabinoid-ligand-type modulators of GPR55; World Intellectual Property Organization patent application PCT/GB2004/000571; 2004. 2004 Sept 2. [Google Scholar]
- Drysdale AJ, Platt B. Cannabinoids: mechanisms and therapeutic applications in the CNS. Curr Med Chem. 2003;10:2719–2732. doi: 10.2174/0929867033456387. [DOI] [PubMed] [Google Scholar]
- D’Souza DC, Abi-Saab WM, Madonick S, Forselius-Bielen K, Doersch A, Braley G, Gueorguieva R, Cooper TB, Krystal JH. Δ-9-Tetrahydrocannabinol effects in schizophrenia: implications for cognition, psychosis, and addiction. Biol Psychiatry. 2005;57:594–608. doi: 10.1016/j.biopsych.2004.12.006. [DOI] [PubMed] [Google Scholar]
- D’Souza DC, Perry E, MacDougall L, Ammerman Y, Cooper T, Wu YT, Braley G, Gueorguieva R, Krystal JH. The psychotomimetic effects of intravenous Δ-9-tetrahydrocannabinol in healthy individuals: implications for psychosis. Neuropsychopharmacology. 2004;29:1558–1572. doi: 10.1038/sj.npp.1300496. [DOI] [PubMed] [Google Scholar]
- Duncan GE, Dagirmanjian R. Δ9-Tetrahydrocannabinol sensitization of the rat brain to direct cholinergic stimulation. Psychopharmacology. 1979;60:237–240. doi: 10.1007/BF00426661. [DOI] [PubMed] [Google Scholar]
- Duncan M, Davison JS, Sharkey KA. Review article: endocannabinoids and their receptors in the enteric nervous system. Aliment Pharmacol Ther. 2005;22:667–683. doi: 10.1111/j.1365-2036.2005.02648.x. [DOI] [PubMed] [Google Scholar]
- Dunn M, Davis R. The perceived effects of marijuana on spinal cord injured males. Paraplegia. 1974;12:175. doi: 10.1038/sc.1974.28. [DOI] [PubMed] [Google Scholar]
- Dyson A, Peacock M, Chen A, Courade JP, Yaqoob M, Groarke A, Brain C, Loong Y, Fox A. Antihyperalgesic properties of the cannabinoid CT-3 in chronic neuropathic and inflammatory pain states in the rat. Pain. 2005;116:129–137. doi: 10.1016/j.pain.2005.03.037. [DOI] [PubMed] [Google Scholar]
- Economidou D, Mattioli L, Cifani C, Perfumi M, Massi M, Cuomo V, Trabace L, Ciccocioppo R. Effect of cannabinoid CB1 receptor antagonist SR-141716A on ethanol self-administration and ethanol-seeking behavior in rats. Psychopharmacology. 2006;183:394–403. doi: 10.1007/s00213-005-0199-9. [DOI] [PubMed] [Google Scholar]
- Efird JT, Friedman GD, Sidney S, Klatsky A, Habel LA, Udaltsova NV, Van den Eeden S, Nelson LM. The risk for malignant primary adult-onset glioma in a large, multiethnic, managed-care cohort: cigarette smoking and other lifestyle behaviors. J Neurooncol. 2004;68:57–69. doi: 10.1023/b:neon.0000024746.87666.ed. [DOI] [PubMed] [Google Scholar]
- Eljaschewitsch E, Witting A, Mawrin C, Lee T, Schmidt PM, Wolf S, Hoertnagl H, Raine CS, Schneider-Stock R, Nitsch R, et al. The endocannabinoid anandamide protects neurons during CNS inflammation by induction of MKP-1 in microglial cells. Neuron. 2006;49:67–79. doi: 10.1016/j.neuron.2005.11.027. [DOI] [PubMed] [Google Scholar]
- Ellert-Miklaszewska A, Kaminska B, Konarska L. Cannabinoids down-regulate PI3K/Akt and Erk signalling pathways and activate proapoptotic function of Bad protein. Cell Signal. 2005;17:25–37. doi: 10.1016/j.cellsig.2004.05.011. [DOI] [PubMed] [Google Scholar]
- Ellison JM, Gelwan E, Ogletree J. Complex partial seizure symptoms affected by marijuana abuse. J Clin Psychiatry. 1990;51:439–440. [PubMed] [Google Scholar]
- Elmes SJ, Jhaveri MD, Smart D, Kendall DA, Chapman V. Cannabinoid CB2 receptor activation inhibits mechanically evoked responses of wide dynamic range dorsal horn neurons in naive rats and in rat models of inflammatory and neuropathic pain. Eur J Neurosci. 2004;20:2311–2320. doi: 10.1111/j.1460-9568.2004.03690.x. [DOI] [PubMed] [Google Scholar]
- El-Remessy AB, Al-Shabrawey M, Khalifa Y, Tsai NT, Caldwell RB, Liou GI. Neuroprotective and blood-retinal barrier-preserving effects of cannabidiol in experimental diabetes. Am J Pathol. 2006;168:235–244. doi: 10.2353/ajpath.2006.050500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- El-Remessy AB, Khalil IE, Matragoon S, Abou-Mohamed G, Tsai NJ, Roon P, Caldwell RB, Caldwell RW, Green K, Liou GI. Neuroprotective effect of (−) Δ9-tetrahydrocannabinol and cannabidiol in N-methyl-D-aspartate-induced retinal neurotoxicity: involvement of peroxynitrite. Am J Pathol. 2003;163:1997–2008. doi: 10.1016/s0002-9440(10)63558-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- ElSohly MA, Harland E, Murphy JC, Wirth P, Waller CW. Cannabinoids in glaucoma: a primary screening procedure. J Clin Pharmacol. 1981;21:472S–478S. doi: 10.1002/j.1552-4604.1981.tb02627.x. [DOI] [PubMed] [Google Scholar]
- ElSohly MA, Harland EC, Benigni DA, Waller CW. Cannabinoids in glaucoma. II: the effect of different cannabinoids on intraocular pressure of the rabbit. Curr Eye Res. 1984;3:841–850. doi: 10.3109/02713688409000797. [DOI] [PubMed] [Google Scholar]
- Emrich HM, Leweke FM, Schneider U. Towards a cannabinoid hypothesis of schizophrenia: cognitive impairments due to dysregulation of the endogenous cannabinoid system. Pharmacol Biochem Behav. 1997;56:803–807. doi: 10.1016/s0091-3057(96)00426-1. [DOI] [PubMed] [Google Scholar]
- Engeli S, Bohnke J, Feldpausch M, Gorzelniak K, Janke J, Bátkai S, Pacher P, Harvey-White J, Luft FC, Sharma AM, et al. Activation of the peripheral endocannabinoid system in human obesity. Diabetes. 2005;54:2838–2843. doi: 10.2337/diabetes.54.10.2838. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eriksen JL, Wszolek Z, Petrucelli L. Molecular pathogenesis of Parkinson disease. Arch Neurol. 2005;62:353–357. doi: 10.1001/archneur.62.3.353. [DOI] [PubMed] [Google Scholar]
- Eshhar N, Striem S, Biegon A. HU-211, a non-psychotropic cannabinoid, rescues cortical neurones from excitatory amino acid toxicity in culture. Neuroreport. 1993;5:237–240. doi: 10.1097/00001756-199312000-00013. [DOI] [PubMed] [Google Scholar]
- Eshhar N, Striem S, Kohen R, Tirosh O, Biegon A. Neuroprotective and antioxidant activities of HU-211, a novel NMDA receptor antagonist. Eur J Pharmacol. 1995;283:19–29. doi: 10.1016/0014-2999(95)00271-l. [DOI] [PubMed] [Google Scholar]
- Evgenov OV, Liaudet L. Role of nitrosative stress and activation of poly(ADP-ribose) polymerase-1 in cardiovascular failure associated with septic and hemorrhagic shock. Curr Vasc Pharmacol. 2005;3:293–299. doi: 10.2174/1570161054368580. [DOI] [PubMed] [Google Scholar]
- Fabre LF, McLendon D. The efficacy and safety of nabilone (a synthetic cannabinoid) in the treatment of anxiety. J Clin Pharmacol. 1981;21:377S–382S. doi: 10.1002/j.1552-4604.1981.tb02617.x. [DOI] [PubMed] [Google Scholar]
- Fadel J, Deutch AY. Anatomical substrates of orexin-dopamine interactions: lateral hypothalamic projections to the ventral tegmental area. Neuroscience. 2002;111:379–387. doi: 10.1016/s0306-4522(02)00017-9. [DOI] [PubMed] [Google Scholar]
- Faden AI. Neuroprotection and traumatic brain injury: theoretical option or realistic proposition. Curr Opin Neurol. 2002;15:707–712. doi: 10.1097/01.wco.0000044767.39452.bf. [DOI] [PubMed] [Google Scholar]
- Fan P. Cannabinoid agonists inhibit the activation of 5-HT3 receptors in rat nodose ganglion neurons. J Neurophysiol. 1995;73:907–910. doi: 10.1152/jn.1995.73.2.907. [DOI] [PubMed] [Google Scholar]
- Fan SF, Yazulla S. Biphasic modulation of voltage-dependent currents of retinal cones by cannabinoid CB1 receptor agonist WIN 55212-2. Vis Neurosci. 2003;20:177–188. doi: 10.1017/s095252380320208x. [DOI] [PubMed] [Google Scholar]
- Fattore L, Deiana S, Spano SM, Cossu G, Fadda P, Scherma M, Fratta W. Endocannabinoid system and opioid addiction: behavioural aspects. Pharmacol Biochem Behav. 2005;81:343–359. doi: 10.1016/j.pbb.2005.01.031. [DOI] [PubMed] [Google Scholar]
- Fattore L, Martellotta MC, Cossu G, Mascia MS, Fratta W. CB1 cannabinoid receptor agonist WIN 55,212-2 decreases cocaine self-administration in rats. Behav Brain Res. 1999;104:141–146. doi: 10.1016/s0166-4328(99)00059-5. [DOI] [PubMed] [Google Scholar]
- Fegley D, Kathuria S, Mercier R, Li C, Goutopoulos A, Makriyannis A, Piomelli D. Anandamide transport is independent of fatty-acid amide hydrolase activity and is blocked by the hydrolysis-resistant inhibitor AM1172. Proc Natl Acad Sci USA. 2004;101:8756–8761. doi: 10.1073/pnas.0400997101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Felder CC, Joyce KE, Briley EM, Glass M, Mackie KP, Fahey KJ, Cullinan GJ, Hunden DC, Johnson DW, Chaney MO, et al. LY320135, a novel cannabinoid CB1 receptor antagonist, unmasks coupling of the CB1 receptor to stimulation of cAMP accumulation. J Pharmacol Exp Ther. 1998;284:291–297. [PubMed] [Google Scholar]
- Felder CC, Joyce KE, Briley EM, Mansouri J, Mackie K, Blond O, Lai Y, Ma AL, Mitchell RL. Comparison of the pharmacology and signal transduction of the human cannabinoid CB1 and CB2 receptors. Mol Pharmacol. 1995;48:443–450. [PubMed] [Google Scholar]
- Feinberg I, Jones R, Walker J, Cavness C, Floyd T. Effects of marijuana extract and tetrahydrocannabinol on electroencephalographic sleep patterns. Clin Pharmacol Ther. 1976;19:782–794. doi: 10.1002/cpt1976196782. [DOI] [PubMed] [Google Scholar]
- Feinberg I, Jones R, Walker JM, Cavness C, March J. Effects of high dosage Δ-9-tetrahydrocannabinol on sleep patterns in man. Clin Pharmacol Ther. 1975;17:458–466. doi: 10.1002/cpt1975174458. [DOI] [PubMed] [Google Scholar]
- Fernandez JR, Allison DB. Rimonabant Sanofi-Synthelabo. Curr Opin Investig Drugs. 2004;5:430–435. [PubMed] [Google Scholar]
- Fernandez-Espejo E, Caraballo I, de Fonseca FR, El Banoua F, Ferrer B, Flores JA, Galan-Rodriguez B. Cannabinoid CB1 antagonists possess antiparkinsonian efficacy only in rats with very severe nigral lesion in experimental parkinsonism. Neurobiol Dis. 2005;18:591–601. doi: 10.1016/j.nbd.2004.10.015. [DOI] [PubMed] [Google Scholar]
- Fernandez-Rodriguez CM, Romero J, Petros TJ, Bradshaw H, Gasalla JM, Gutierrez ML, Lledo JL, Santander C, Fernandez TP, Tomas E, et al. Circulating endogenous cannabinoid anandamide and portal, systemic and renal hemodynamics in cirrhosis. Liver Int. 2004;24:477–483. doi: 10.1111/j.1478-3231.2004.0945.x. [DOI] [PubMed] [Google Scholar]
- Fernandez-Ruiz J, Gonzalez S. Cannabinoid control of motor function at the basal ganglia. In: Pertwee R, editor. Cannabinoids. Springer; New York: 2005. pp. 479–509. [DOI] [PubMed] [Google Scholar]
- Ferrer B, Asbrock N, Kathuria S, Piomelli D, Giuffrida A. Effects of levodopa on endocannabinoid levels in rat basal ganglia: implications for the treatment of levodopa-induced dyskinesias. Eur J Neurosci. 2003;18:1607–1614. doi: 10.1046/j.1460-9568.2003.02896.x. [DOI] [PubMed] [Google Scholar]
- Fischer-Stenger K, Dove Pettit DA, Cabral GA, Fischer-Stenger K, Dove Pettit DA, Cabral GA. Δ9-Tetrahydrocannabinol inhibition of tumor necrosis factor-α: suppression of post-translational events. J Pharmacol Exp Ther. 1993;267:1558–1565. [PubMed] [Google Scholar]
- Fish BS, Consroe P. The ontogeny of Δ-9-tetrahydrocannabinol responsiveness in the rabbit. Dev Psychobiol. 1983;16:147–158. doi: 10.1002/dev.420160207. [DOI] [PubMed] [Google Scholar]
- Fish BS, Consroe P, Fox RR. Inheritance of Δ9-tetrahydrocannabinol seizure susceptibility in rabbits. J Hered. 1981;72:215–216. doi: 10.1093/oxfordjournals.jhered.a109475. [DOI] [PubMed] [Google Scholar]
- Foltin RW, Fishman MW, Pippen PA, Kelly TH. Behavioral effects of cocaine alone and in combination of ethanol and marijuana in humans. Drug Alcohol Depend. 1993;32:93–106. doi: 10.1016/0376-8716(93)80001-u. [DOI] [PubMed] [Google Scholar]
- Ford WR, Honan SA, White R, Hiley CR. Evidence of a novel site mediating anandamide-induced negative inotropic and coronary vasodilator responses in rat isolated hearts. Br J Pharmacol. 2002;135:1191–1198. doi: 10.1038/sj.bjp.0704565. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Forget B, Hamon M, Thiebot MH. Cannabinoid CB1 receptors are involved in motivational effects of nicotine in rats. Psychopharmacology. 2005;181:722–734. doi: 10.1007/s00213-005-0015-6. [DOI] [PubMed] [Google Scholar]
- Formukong EA, Evans AT, Evans FJ. Analgesic and antiinflammatory activity of constituents of Cannabis sativa L. Inflammation. 1988;12:361–371. doi: 10.1007/BF00915771. [DOI] [PubMed] [Google Scholar]
- Fowler CJ. Oleamide: a member of the endocannabinoid family? Br J Pharmacol. 2004;141:195–196. doi: 10.1038/sj.bjp.0705608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fowler CJ, Jonsson KO, Andersson A, Juntunen J, Jarvinen T, Vandevoorde S, Lambert DM, Jerman JC, Smart D. Inhibition of C6 glioma cell proliferation by anandamide, 1-arachidonoylglycerol, and by a water soluble phosphate ester of anandamide: variability in response and involvement of arachidonic acid. Biochem Pharmacol. 2003;66:757–767. doi: 10.1016/s0006-2952(03)00392-7. [DOI] [PubMed] [Google Scholar]
- Fowler CJ, Tiger G, Ligresti A, Lopez-Rodriguez ML, Di Marzo V. Selective inhibition of anandamide cellular uptake versus enzymatic hydrolysis—a difficult issue to handle. Eur J Pharmacol. 2004;492:1–11. doi: 10.1016/j.ejphar.2004.03.048. [DOI] [PubMed] [Google Scholar]
- Fox A, Bevan S. Therapeutic potential of cannabinoid receptor agonists as analgesic agents. Expert Opin Investig Drugs. 2005;14:695–703. doi: 10.1517/13543784.14.6.695. [DOI] [PubMed] [Google Scholar]
- Fox A, Kesingland A, Gentry C, McNair K, Patel S, Urban L, James I. The role of central and peripheral cannabinoid1 receptors in the antihyperalgesic activity of cannabinoids in a model of neuropathic pain. Pain. 2001;92:91–100. doi: 10.1016/s0304-3959(00)00474-7. [DOI] [PubMed] [Google Scholar]
- Fox P, Bain PG, Glickman S, Carroll C, Zajicek J. The effect of cannabis on tremor in patients with multiple sclerosis. Neurology. 2004;62:1105–1109. doi: 10.1212/01.wnl.0000118203.67138.3e. [DOI] [PubMed] [Google Scholar]
- Fox SH, Henry B, Hill M, Crossman A, Brotchie J. Stimulation of cannabinoid receptors reduces levodopa-induced dyskinesia in the MPTP-lesioned nonhuman primate model of Parkinson’s disease. Mov Disord. 2002a;17:1180–1187. doi: 10.1002/mds.10289. [DOI] [PubMed] [Google Scholar]
- Fox SH, Kellett M, Moore AP, Crossman AR, Brotchie JM. Randomised, double-blind, placebo-controlled trial to assess the potential of cannabinoid receptor stimulation in the treatment of dystonia. Mov Disord. 2002b;17:145–149. doi: 10.1002/mds.1280. [DOI] [PubMed] [Google Scholar]
- Freedland CS, Poston JS, Porrino LJ. Effects of Sr141716A, a central cannabinoid receptor antagonist, on food-maintained responding. Pharmacol Biochem Behav. 2000;67:265–270. doi: 10.1016/s0091-3057(00)00359-2. [DOI] [PubMed] [Google Scholar]
- Freedland CS, Sharpe AL, Samson HH, Porrino LJ. Effects of SR141716A on ethanol and sucrose self-administration. Alcohol Clin Exp Res. 2001;25:277–282. [PubMed] [Google Scholar]
- Freedland CS, Whitlow CT, Smith HR, Porrino LJ. Functional consequences of the acute administration of the cannabinoid receptor antagonist, SR141716A, in cannabinoid-naive and -tolerant animals: a quantitative 2-[14C]deoxyglucose study. Brain Res. 2003;962:169–179. doi: 10.1016/s0006-8993(02)03999-9. [DOI] [PubMed] [Google Scholar]
- Freemon FR. Effects of marihuana on sleeping states. J Am Med Assoc. 1972;220:1364–1365. [PubMed] [Google Scholar]
- Freemon FR. The effect of chronically administered Δ-9-tetrahydrocannabinol upon the polygraphically monitored sleep of normal volunteers. Drug Alcohol Depend. 1982;10:345–353. doi: 10.1016/0376-8716(82)90036-9. [DOI] [PubMed] [Google Scholar]
- French ED. Δ9-Tetrahydrocannabinol excites rat VTA dopamine neurons through activation of cannabinoid CB1 but not opioid receptors. Neurosci Lett. 1997;226:159–162. doi: 10.1016/s0304-3940(97)00278-4. [DOI] [PubMed] [Google Scholar]
- Freund TF, Katona I, Piomelli D. Role of endogenous cannabinoids in synaptic signaling. Physiol Rev. 2003;83:1017–1066. doi: 10.1152/physrev.00004.2003. [DOI] [PubMed] [Google Scholar]
- Fride E. The endocannabinoid-CB1 receptor system during gestation and postnatal development. Eur J Pharmacol. 2004;500:289–297. doi: 10.1016/j.ejphar.2004.07.033. [DOI] [PubMed] [Google Scholar]
- Fride E, Bregman T, Kirkham TC. Endocannabinoids and food intake: newborn suckling and appetite regulation in adulthood. Exp Biol Med. 2005;230:225–234. doi: 10.1177/153537020523000401. [DOI] [PubMed] [Google Scholar]
- Fride E, Foox A, Rosenberg E, Faigenboim M, Cohen V, Barda L, Blau H, Mechoulam R. Milk intake and survival in newborn cannabinoid CB1 receptor knockout mice: evidence for a “CB3” receptor. Eur J Pharmacol. 2003;461:27–34. doi: 10.1016/s0014-2999(03)01295-0. [DOI] [PubMed] [Google Scholar]
- Fride E, Mechoulam R. Pharmacological activity of the cannabinoid receptor agonist, anandamide, a brain constituent. Eur J Pharmacol. 1993;231:313–314. doi: 10.1016/0014-2999(93)90468-w. [DOI] [PubMed] [Google Scholar]
- Fride E, Shohami E. The endocannabinoid system: function in survival of the embryo, the newborn and the neuron. Neuroreport. 2002;13:1833–1841. doi: 10.1097/00001756-200210280-00001. [DOI] [PubMed] [Google Scholar]
- Friedman H, Klein TW, Newton C, Daaka Y. Marijuana, receptors and immunomodulation. Adv Exp Med Biol. 1995;373:103–113. doi: 10.1007/978-1-4615-1951-5_15. [DOI] [PubMed] [Google Scholar]
- Fu J, Gaetani S, Oveisi F, Lo Verme J, Serrano A, Rodriguez de Fonseca F, Rosengarth A, Luecke H, Di Giacomo B, Tarzia G, et al. Oleylethanolamide regulates feeding and body weight through activation of the nuclear receptor PPAR-α. Nature (Lond) 2003;425:90–93. doi: 10.1038/nature01921. [DOI] [PubMed] [Google Scholar]
- Gabbay E, Avraham Y, Ilan Y, Israeli E, Berry EM. Endocannabinoids and liver disease—review. Liver Int. 2005;25:921–926. doi: 10.1111/j.1478-3231.2005.01180.x. [DOI] [PubMed] [Google Scholar]
- Gallate JE, Saharov T, Mallet PE, McGregor IS. Increased motivation for beer in rats following administration of a cannabinoid CB1 receptor agonist. Eur J Pharmacol. 1999;370:233–240. doi: 10.1016/s0014-2999(99)00170-3. [DOI] [PubMed] [Google Scholar]
- Gallily R, Breuer A, Mechoulam R. 2-Arachidonylglycerol, an endogenous cannabinoid, inhibits tumor necrosis factor-α production in murine macrophages, and in mice. Eur J Pharmacol. 2000;406:R5–R7. doi: 10.1016/s0014-2999(00)00653-1. [DOI] [PubMed] [Google Scholar]
- Galve-Roperh I, Rueda D, Gómez Del Pulgar T, Velasco G, Guzman M. Mechanism of extracellular signal-regulated kinase activation by the CB1 cannabinoid receptor. Mol Pharmacol. 2002;62:1385–1392. doi: 10.1124/mol.62.6.1385. [DOI] [PubMed] [Google Scholar]
- Galve-Roperh I, Sanchez C, Cortes ML, del Pulgar TG, Izquierdo M, Guzman M. Anti-tumoral action of cannabinoids: involvement of sustained ceramide accumulation and extracellular signal-regulated kinase activation. Nat Med. 2000;6:313–319. doi: 10.1038/73171. [DOI] [PubMed] [Google Scholar]
- Gamber KM, Macarthur H, Westfall TC. Cannabinoids augment the release of neuropeptide Y in the rat hypothalamus. Neuropharmacology. 2005;49:646–652. doi: 10.1016/j.neuropharm.2005.04.017. [DOI] [PubMed] [Google Scholar]
- Gambi F, De Berardis D, Sepede G, Quartesan R, Calcagni E, Salerno RM, Conti CM, Ferro FM. Cannabinoid receptors and their relationships with neuropsychiatric disorders. Int J Immunopathol Pharmacol. 2005;18:15–19. doi: 10.1177/039463200501800103. [DOI] [PubMed] [Google Scholar]
- Gammon CM, Freeman GM, Jr, Xie W, Petersen SL, Wetsel WC. Regulation of gonadotropin-releasing hormone secretion by cannabinoids. Endocrinology. 2005;146:4491–4499. doi: 10.1210/en.2004-1672. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gaoni Y, Mechoulam R. Isolation, structure and partial synthesis of an active constituent of hashish. J Am Chem Soc. 1964;86:1646–1647. [Google Scholar]
- Garcia N, Jr, Jarai Z, Mirshahi F, Kunos G, Sanyal AJ. Systemic and portal hemodynamic effects of anandamide. Am J Physiol. 2001;280:G14–G20. doi: 10.1152/ajpgi.2001.280.1.G14. [DOI] [PubMed] [Google Scholar]
- Gardiner SM, March JE, Kemp PA, Bennett T. Regional haemodynamic responses to the cannabinoid agonist, WIN 55212-2, in conscious, normotensive rats, and in hypertensive, transgenic rats. Br J Pharmacol. 2001;133:445–453. doi: 10.1038/sj.bjp.0704100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gardiner SM, March JE, Kemp PA, Bennett T. Complex regional haemodynamic effects of anandamide in conscious rats. Br J Pharmacol. 2002a;135:1889–1896. doi: 10.1038/sj.bjp.0704649. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gardiner SM, March JE, Kemp PA, Bennett T. Influence of the CB1 receptor antagonist, AM 251, on the regional haemodynamic effects of WIN-55212-2 or HU 210 in conscious rats. Br J Pharmacol. 2002b;136:581–587. doi: 10.1038/sj.bjp.0704750. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gardiner SM, March JE, Kemp PA, Bennett T. Involvement of CB1-receptors and β-adrenoceptors in the regional hemodynamic responses to lipopoly-saccharide infusion in conscious rats. Am J Physiol. 2005;288:H2280–H2288. doi: 10.1152/ajpheart.00851.2004. [DOI] [PubMed] [Google Scholar]
- Gardner EL. Endocannabinoid signaling system and brain reward: emphasis on dopamine. Pharmacol Biochem Behav. 2005;81:263–284. doi: 10.1016/j.pbb.2005.01.032. [DOI] [PubMed] [Google Scholar]
- Gardner EL, Paredes W, Smith D, Donner A, Milling C, Cohen D, Morrison D. Facilitation of brain stimulation reward by 9-tetrahydrocannabinol.Δ9-tetrahydrocannabinol. Psychopharmacology (Berl) 1988;96:142–144. doi: 10.1007/BF02431546. [DOI] [PubMed] [Google Scholar]
- Gareau Y, Dufresne C, Gallant M, Rochette C, Sawyer N, Slipetz DM, Tremblay N, Weech PK, Metters KM, Labelle M. Structure activity relationships of tetrahydrocannabinol analogues on human cannabinoid receptors. Bioorg Med Chem Lett. 1996;6:189–194. [Google Scholar]
- Gaskari SA, Liu H, Moezi L, Li Y, Baik SK, Lee SS. Role of endocannabinoids in the pathogenesis of cirrhotic cardiomyopathy in bile duct-ligated rats. Br J Pharmacol. 2005;146:315–323. doi: 10.1038/sj.bjp.0706331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gatley SJ, Lan R, Pyatt B, Gifford AN, Volkow ND, Makriyannis A. Binding of the non-classical cannabinoid CP 55,940, and the diarylpyrazole AM251 to rodent brain cannabinoid receptors. Life Sci. 1997;61:191–197. doi: 10.1016/s0024-3205(97)00690-5. [DOI] [PubMed] [Google Scholar]
- Gebremedhin D, Lange AR, Campbell WB, Hillard CJ, Harder DR. Cannabinoid CB1 receptor of cat cerebral arterial muscle functions to inhibit L-type Ca2+ channel current. Am J Physiol. 1999;266:H2085–H2093. doi: 10.1152/ajpheart.1999.276.6.H2085. [DOI] [PubMed] [Google Scholar]
- Gelinas D, Miller R, Abood M. A pilot study of safety and tolerability of Δ9-THC (Marinol) treatment for ALS. Amyotroph Lateral Scler Other Motor Neuron Disord. 2002;3:23. [Google Scholar]
- Genn RF, Tucci S, Marco E, Viveros MP, File SE. Anxiolytic and anxiogenic effects of the cannabinoid agonist CP 55,940 in animal tests of anxiety. J Psychopharmacology. 2003;17:A27. [Google Scholar]
- Genn RF, Tucci S, Marco EM, Viveros MP, File SE. Unconditioned and conditioned anxiogenic effects of the cannabinoid receptor agonist CP 55,940 in the social interaction test. Pharmacol Biochem Behav. 2004;77:567–573. doi: 10.1016/j.pbb.2003.12.019. [DOI] [PubMed] [Google Scholar]
- Gentleman SM, Leclercq PD, Moyes L, Graham DI, Smith C, Griffin WS, Nicoll JA. Long-term intracerebral inflammatory response after traumatic brain injury. Forensic Sci Int. 2004;146:97–104. doi: 10.1016/j.forsciint.2004.06.027. [DOI] [PubMed] [Google Scholar]
- George SR, Fan T, Roldan L, Naranjo CA. Corticotropin-releasing factor is altered in brains of animals with high preference for ethanol. Alcohol Clin Exp Res. 1990;14:425–429. doi: 10.1111/j.1530-0277.1990.tb00499.x. [DOI] [PubMed] [Google Scholar]
- Gerdeman G, Lovinger DM. CB1 cannabinoid receptor inhibits synaptic release of glutamate in rat dorsolateral striatum. J Neurophysiol. 2001;85:468–471. doi: 10.1152/jn.2001.85.1.468. [DOI] [PubMed] [Google Scholar]
- Gerdeman GL, Lovinger DM. Emerging roles for endocannabinoids in long-term synaptic plasticity. Br J Pharmacol. 2003;140:781–789. doi: 10.1038/sj.bjp.0705466. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gerdeman GL, Partridge JG, Lupica CR, Lovinger DM. It could be habit forming: drugs of abuse and striatal synaptic plasticity. Trends Neurosci. 2003;26:184–192. doi: 10.1016/S0166-2236(03)00065-1. [DOI] [PubMed] [Google Scholar]
- Gerdeman GL, Ronesi J, Lovinger DM. Postsynaptic endocannabinoid release is critical to long-term depression in the striatum. Nat Neurosci. 2002;5:446–451. doi: 10.1038/nn832. [DOI] [PubMed] [Google Scholar]
- Germain N, Boichot E, Advenier C, Berdyshev EV, Lagente V. Effect of the cannabinoid receptor ligand, WIN 55,212-2, on superoxide anion and TNF-α production by human mononuclear cells. Int Immunopharmacol. 2002;2:537–543. doi: 10.1016/s1567-5769(01)00200-4. [DOI] [PubMed] [Google Scholar]
- Germano MP, D’Angelo V, Mondello MR, Pergolizzi S, Capasso F, Capasso R, Izzo AA, Mascolo N, De Pasquale R. Cannabinoid CB1-mediated inhibition of stress-induced gastric ulcers in rats. Naunyn-Schmiedeberg’s Arch Pharmacol. 2001;363:241–244. doi: 10.1007/s002100000360. [DOI] [PubMed] [Google Scholar]
- Gessa GL, Orru A, Lai P, Maccioni P, Lecca R, Lobina C, Carai MA, Colombo G. Lack of tolerance to the suppressing effect of rimonabant on chocolate intake in rats. Psychopharmacology. 2006;185:248–254. doi: 10.1007/s00213-006-0327-1. [DOI] [PubMed] [Google Scholar]
- Ghozland S, Matthes HW, Simonin F, Filliol D, Kieffer BL, Maldonado R. Motivational effects of cannabinoids are mediated by μ-opioid and κ-opioid receptors. J Neurosci. 2002;22:1146–1154. doi: 10.1523/JNEUROSCI.22-03-01146.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gifford AN, Ashby CR., Jr Electrically evoked acetylcholine release from hippocampal slices is inhibited by the cannabinoid receptor agonist, WIN 55212-2, and is potentiated by the cannabinoid antagonist, SR 141716A. J Pharmacol Exp Ther. 1996;277:1431–1436. [PubMed] [Google Scholar]
- Gilgun-Sherki Y, Melamed E, Mechoulam R, Offen D. The CB1 cannabinoid receptor agonist, HU-210, reduces levodopa-induced rotations in 6-hydroxy-dopamine-lesioned rats. Pharmacol Toxicol. 2003;93:66–70. doi: 10.1034/j.1600-0773.2003.930202.x. [DOI] [PubMed] [Google Scholar]
- Gingold AR, Bergasa NV. The cannabinoid agonist WIN 55,212-2 increases nociception threshold in cholestatic rats: implications for the treatment of the pruritus of cholestasis. Life Sci. 2003;73:2741–2747. doi: 10.1016/s0024-3205(03)00668-4. [DOI] [PubMed] [Google Scholar]
- Giros B, Jaber M, Jones SR, Wightman RM, Caron MG. Hyperlocomotion and indifference to cocaine and amphetamine in mice lacking the dopamine transporter. Nature (Lond) 1996;379:606–612. doi: 10.1038/379606a0. [DOI] [PubMed] [Google Scholar]
- Giuffrida A, Leweke FM, Gerth CW, Schreiber D, Koethe D, Faulhaber J, Klosterkotter J, Piomelli D. Cerebrospinal anandamide levels are elevated in acute schizophrenia and are inversely correlated with psychotic symptoms. Neuropsychopharmacology. 2004;29:2108–2114. doi: 10.1038/sj.npp.1300558. [DOI] [PubMed] [Google Scholar]
- Giuffrida A, Parsons LH, Kerr TM, Rodriguez de Fonseca F, Navarro M, Piomelli D. Dopamine activation of endogenous cannabinoid signaling in dorsal striatum. Nat Neurosci. 1999;2:358–363. doi: 10.1038/7268. [DOI] [PubMed] [Google Scholar]
- Giuliani D, Ferrari F, Ottani A. The cannabinoid agonist HU210 modifies behavioural responses to novelty and stress. Pharmacol Res. 2000;41:45–51. doi: 10.1006/phrs.1999.0560. [DOI] [PubMed] [Google Scholar]
- Glaser ST, Abumrad NA, Fatade F, Kaczocha M, Studholme KM, Deutsch DG. Evidence against the presence of an anandamide transporter. Proc Natl Acad Sci USA. 2003;100:4269–4274. doi: 10.1073/pnas.0730816100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Glaser ST, Kaczocha M, Deutsch DG. Anandamide transport: a critical review. Life Sci. 2005;77:1584–1604. doi: 10.1016/j.lfs.2005.05.007. [DOI] [PubMed] [Google Scholar]
- Glass M. The role of cannabinoids in neurodegenerative diseases. Prog Neuropsychopharmacol Biol Psychiatry. 2001;25:743–765. doi: 10.1016/s0278-5846(01)00162-2. [DOI] [PubMed] [Google Scholar]
- Glass M, Brotchie JM, Maneuf YP. Modulation of neurotransmission by cannabinoids in the basal ganglia. Eur J Neurosci. 1997a;9:199–203. doi: 10.1111/j.1460-9568.1997.tb01390.x. [DOI] [PubMed] [Google Scholar]
- Glass M, Dragunow M, Faull RL. Cannabinoid receptors in the human brain: a detailed anatomical and quantitative autoradiographic study in the fetal, neonatal and adult human brain. Neuroscience. 1997b;77:299–318. doi: 10.1016/s0306-4522(96)00428-9. [DOI] [PubMed] [Google Scholar]
- Glass M, Dragunow M, Faull RL. The pattern of neurodegeneration in Huntington’s disease: a comparative study of cannabinoid, dopamine, adenosine and GABA(A) receptor alterations in the human basal ganglia in Huntington’s disease. Neuroscience. 2000;97:505–519. doi: 10.1016/s0306-4522(00)00008-7. [DOI] [PubMed] [Google Scholar]
- Glass M, Faull RL, Dragunow M. Loss of cannabinoid receptors in the substantia nigra in Huntington’s disease. Neuroscience. 1993;56:523–527. doi: 10.1016/0306-4522(93)90352-g. [DOI] [PubMed] [Google Scholar]
- Glass M, Felder CC. Concurrent stimulation of cannabinoid CB1 and dopamine D2 receptors augments cAMP accumulation in striatal neurons: evidence for a Gs linkage to the CB1 receptor. J Neurosci. 1997;17:5327–5333. doi: 10.1523/JNEUROSCI.17-14-05327.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Glass M, Northup JK. Agonist selective regulation of G proteins by cannabinoid CB1 and CB2 receptors. Mol Pharmacol. 1999;56:1362–1369. doi: 10.1124/mol.56.6.1362. [DOI] [PubMed] [Google Scholar]
- Glass M, van Dellen A, Blakemore C, Hannan AJ, Faull RL. Delayed onset of Huntington’s disease in mice in an enriched environment correlates with delayed loss of cannabinoid CB1 receptors. Neuroscience. 2004;123:207–212. doi: 10.1016/s0306-4522(03)00595-5. [DOI] [PubMed] [Google Scholar]
- Glass RM, Uhlenhuth EH, Hartel FW, Schuster CR, Fischman MW. A single dose study of nabilone, a synthetic cannabinoid. Psychopharmacology. 1980;71:137–142. doi: 10.1007/BF00434401. [DOI] [PubMed] [Google Scholar]
- Gobbi G, Bambico FR, Mangieri R, Bortolato M, Campolongo P, Solinas M, Cassano T, Morgese MG, Debonnel G, Duranti A, et al. Antidepressant-like activity and modulation of brain monoaminergic transmission by blockade of anandamide hydrolysis. Proc Natl Acad Sci USA. 2005;102:18620–18625. doi: 10.1073/pnas.0509591102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Godino MC, Torres M, Sanchez-Prieto J. Inhibition of N- and P/Q-type Ca2+ channels by cannabinoid receptors in single cerebrocortical nerve terminals. FEBS Lett. 2005;579:768–772. doi: 10.1016/j.febslet.2004.12.065. [DOI] [PubMed] [Google Scholar]
- Godlewski G, Malinowska B, Schlicker E. Presynaptic cannabinoid CB1 receptors are involved in the inhibition of the neurogenic vasopressor response during septic shock in pithed rats. Br J Pharmacol. 2004;142:701–708. doi: 10.1038/sj.bjp.0705839. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gómez Del Pulgar T, De Ceballos ML, Guzman M, Velasco G. Cannabinoids protect astrocytes from ceramide-induced apoptosis through the phosphatidylinositol 3-kinase/protein kinase B pathway. J Biol Chem. 2002a;277:36527–36533. doi: 10.1074/jbc.M205797200. [DOI] [PubMed] [Google Scholar]
- Gómez Del Pulgar T, Velasco G, Guzman M. The CB1 cannabinoid receptor is coupled to the activation of protein kinase B/Akt. Biochem J. 2000;347:369–373. doi: 10.1042/0264-6021:3470369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gómez Del Pulgar T, Velasco G, Sanchez C, Haro A, Guzman M. De novo-synthesized ceramide is involved in cannabinoid-induced apoptosis. Biochem J. 2002b;363:183–188. doi: 10.1042/0264-6021:3630183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gomez R, Navarro M, Ferrer B, Trigo JM, Bilbao A, Del Arco I, Cippitelli A, Nava F, Piomelli D, Rodriguez de Fonseca F. A peripheral mechanism for CB1 cannabinoid receptor-dependent modulation of feeding. J Neurosci. 2002;22:9612–9617. doi: 10.1523/JNEUROSCI.22-21-09612.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gong J-P, Onaivi ES, Ishiguro H, Liu Q-R, Tagliaferro PA, Brusco A, Uhl GR. Cannabinoid CB2 receptors: immunohistochemical localization in rat brain. Brain Res. 2006;1071:10–23. doi: 10.1016/j.brainres.2005.11.035. [DOI] [PubMed] [Google Scholar]
- Gonsiorek W, Lunn C, Fan X, Narula S, Lundell D, Hipkin RW. Endocannabinoid 2-arachidonyl glycerol is a full agonist through human type 2 cannabinoid receptor: antagonism by anandamide. Mol Pharmacol. 2000;57:1045–1050. [PubMed] [Google Scholar]
- Gonzalez S, Cascio MG, Fernandez-Ruiz J, Sparpaglione V, Parolaro D, Ramos JA. Changes in endocannabinoid content in the brain of rats chronically exposed to nicotine, ethanol or cocaine. Brain Res. 2002;954:73–81. doi: 10.1016/s0006-8993(02)03344-9. [DOI] [PubMed] [Google Scholar]
- Gonzalez S, Mena MA, Lastres-Becker I, Serrano A, de Yebenes JG, Ramos JA, Fernandez-Ruiz J. Cannabinoid CB1 receptors in the basal ganglia and motor response to activation or blockade of these receptors in parkin-null mice. Brain Res. 2005;1046:195–206. doi: 10.1016/j.brainres.2005.04.010. [DOI] [PubMed] [Google Scholar]
- Gonzalez S, Scorticati C, Garcia-Arencibia M, de Miguel R, Ramos JA, Fernandez-Ruiz J. Effects of rimonabant, a selective cannabinoid CB1 receptor antagonist, in a rat model of Parkinson’s disease. Brain Res. 2006;1073–1074:209–219. doi: 10.1016/j.brainres.2005.12.014. [DOI] [PubMed] [Google Scholar]
- Goparaju SK, Ueda N, Taniguchi K, Yamamoto S. Enzymes of porcine brain hydrolyzing 2-arachidonoylglycerol, an endogenous ligand of cannabinoid receptors. Biochem Pharmacol. 1999;57:417–423. doi: 10.1016/s0006-2952(98)00314-1. [DOI] [PubMed] [Google Scholar]
- Goparaju SK, Ueda N, Yamaguchi H, Yamamoto S. Anandamide amidohydrolase reacting with 2-arachidonoylglycerol, another cannabinoid receptor ligand. FEBS Lett. 1998;422:69–73. doi: 10.1016/s0014-5793(97)01603-7. [DOI] [PubMed] [Google Scholar]
- Gordon E, Devinsky O. Alcohol and marijuana: effects on epilepsy and use by patients with epilepsy. Epilepsia. 2001;42:1266–1272. doi: 10.1046/j.1528-1157.2001.19301.x. [DOI] [PubMed] [Google Scholar]
- Gorelick DA, Heishman SJ, Preston KL, Nelson RA, Moolchan ET, Huestis MA. The cannabinoid CB1 receptor antagonist rimonabant attenuates the hypotensive effect of smoked marijuana in male smokers. Am Heart J. 2006;151:754.e1–754.e5. doi: 10.1016/j.ahj.2005.11.006. [DOI] [PubMed] [Google Scholar]
- Gorter R, Seefried M, Volberding P. Dronabinol effects on weight in patients with HIV infection. AIDS. 1992;6:127. doi: 10.1097/00002030-199201000-00018. [DOI] [PubMed] [Google Scholar]
- Goutopoulos A, Fan P, Khanolkar AD, Xie XQ, Lin S, Makriyannis A. Stereochemical selectivity of methanandamides for the CB1 and CB2 cannabinoid receptors and their metabolic stability. Bioorg Med Chem. 2001;9:1673–1684. doi: 10.1016/s0968-0896(01)00088-8. [DOI] [PubMed] [Google Scholar]
- Green K, Bigger JF, Kim K, Bowman K. Cannabinoid action on the eye as mediated through the central nervous system and local adrenergic activity. Exp Eye Res. 1977a;24:189–196. doi: 10.1016/0014-4835(77)90259-7. [DOI] [PubMed] [Google Scholar]
- Green K, Bigger JF, Kim K, Bowman K. Cannabinoid penetration and chronic effects in the eye. Exp Eye Res. 1977b;24:197–205. doi: 10.1016/0014-4835(77)90260-3. [DOI] [PubMed] [Google Scholar]
- Green K, Kearse EC, McIntyre OL. Interaction between Δ-9-tetrahydrocannabinol and indomethacin. Ophthalmic Res. 2001;33:217–220. doi: 10.1159/000055673. [DOI] [PubMed] [Google Scholar]
- Green K, Pederson JE. Effect of 1-tetrahydrocannabinol on aqueous dynamics and ciliary body permeability in the rabbit. Exp Eye Res. 1973;15:499–507. doi: 10.1016/0014-4835(73)90142-5. [DOI] [PubMed] [Google Scholar]
- Green K, Podos SM. Antagonism of arachidonic acid-induced ocular effects by Δ1-tetrahydrocannabinol. Investig Ophthalmol. 1974;13:422–429. [PubMed] [Google Scholar]
- Greenberg HS, Werness SA, Pugh JE, Andrus RO, Anderson DJ, Domino EF. Short-term effects of smoking marijuana on balance in patients with multiple sclerosis and normal volunteers. Clin Pharmacol Ther. 1994;55:324–328. doi: 10.1038/clpt.1994.33. [DOI] [PubMed] [Google Scholar]
- Greenberg I, Kuehnle J, Mendelson JH, Bernstein JG. Effects of marijuana use on body weight and energy intake in humans. Psychopharmacology. 1976;49:79–84. doi: 10.1007/BF00427475. [DOI] [PubMed] [Google Scholar]
- Greengard P. The neurobiology of slow synaptic transmission. Science (Wash DC) 2001;294:1024–1030. doi: 10.1126/science.294.5544.1024. [DOI] [PubMed] [Google Scholar]
- Grenard P, Julien B, Tran Van Nhieu J, Li L, Ledent C, Mallat A, Lotersztajn S. Reduced fibrosis in mice invalidated for CB1 receptor. Proceedings of the 2004 Symposium on the Cannabinoids; 2004 June 22–27; Peastum, Italy. Burlington, VT: International Cannabinoid Research Society; 2004. p. 60. [Google Scholar]
- Grigorenko E, Kittler J, Clayton C, Wallace D, Zhuang S, Bridges D, Bundey S, Boon A, Pagget C, Hayashizaki S, et al. Assessment of cannabinoid induced gene changes: tolerance and neuroprotection. Chem Phys Lipids. 2002;121:257–266. doi: 10.1016/s0009-3084(02)00161-5. [DOI] [PubMed] [Google Scholar]
- Grimaldi C, Pisanti S, Laezza C, Malfitano AM, Santoro A, Vitale M, Caruso MG, Notarnicola M, Iacuzzo I, Portella G, et al. Anandamide inhibits adhesion and migration of breast cancer cells. Exp Cell Res. 2006;312:363–373. doi: 10.1016/j.yexcr.2005.10.024. [DOI] [PubMed] [Google Scholar]
- Grinspoon L, Bakalar JB. Marihuana: The Forbidden Medicine. Yale University Press; New Haven, CT: 1993. The history of cannabis; pp. 1–23. [Google Scholar]
- Grinspoon L, Bakalar JB. The use of cannabis as a mood stabilizer in bipolar disorder: anecdotal evidence and the need for clinical research. J Psychoact Drugs. 1998;30:171–177. doi: 10.1080/02791072.1998.10399687. [DOI] [PubMed] [Google Scholar]
- Grotenhermen F. Pharmacokinetics and pharmacodynamics of cannabinoids. Clin Pharmacokinet. 2003;42:327–360. doi: 10.2165/00003088-200342040-00003. [DOI] [PubMed] [Google Scholar]
- Grotenhermen F. Pharmacology of cannabinoids. Neuro Endocrinol Lett. 2004;25:14–23. [PubMed] [Google Scholar]
- Grufferman S, Schwartz AG, Ruymann FB, Maurer HM. Parents’ use of cocaine and marijuana and increased risk of rhabdomyosarcoma in their children. Cancer Causes Control. 1993;4:217–224. doi: 10.1007/BF00051316. [DOI] [PubMed] [Google Scholar]
- Grundy RI. The therapeutic potential of the cannabinoids in neuroprotection. Expert Opin Investig Drugs. 2002;11:1365–1374. doi: 10.1517/13543784.11.10.1365. [DOI] [PubMed] [Google Scholar]
- Gubellini P, Picconi B, Bari M, Battista N, Calabresi P, Centonze D, Bernardi G, Finazzi-Agro A, Maccarrone M. Experimental parkinsonism alters endocannabinoid degradation: implications for striatal glutamatergic transmission. J Neurosci. 2002;22:6900–6907. doi: 10.1523/JNEUROSCI.22-16-06900.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guagnini F, Valenti M, Mukenge S, Matias I, Bianchetti A, Di Palo S, Ferla G, Di Marzo V, Croci T. Neural contractions in colonic strips from patients with diverticulosis: role of endocannabinoids and substance P. Gut. 2006;55:946–953. doi: 10.1136/gut.2005.076372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guimaraes FS, Chiaretti TM, Graeff FG, Zuardi AW. Antianxiety effect of cannabidiol in the elevated plus-maze. Psychopharmacology. 1990;100:558–559. doi: 10.1007/BF02244012. [DOI] [PubMed] [Google Scholar]
- Guimaraes FS, de Aguiar JC, Mechoulam R, Breuer A. Anxiolytic effect of cannabidiol derivatives in the elevated plus-maze. Gen Pharmacol. 1994;25:161–164. doi: 10.1016/0306-3623(94)90027-2. [DOI] [PubMed] [Google Scholar]
- Guindon J, Beaulieu P. Antihyperalgesic effects of local injections of anandamide, ibuprofen, rofecoxib and their combinations in a model of neuropathic pain. Neuropharmacology. 2006;50:814–823. doi: 10.1016/j.neuropharm.2005.12.002. [DOI] [PubMed] [Google Scholar]
- Guindon J, De Lean A, Beaulieu P. Local interactions between anandamide, an endocannabinoid, and ibuprofen, a nonsteroidal anti-inflammatory drug, in acute and inflammatory pain. Pain. 2006;121:85–93. doi: 10.1016/j.pain.2005.12.007. [DOI] [PubMed] [Google Scholar]
- Gulyas AI, Cravatt BF, Bracey MH, Dinh TP, Piomelli D, Boscia F, Freund TF. Segregation of two endocannabinoid-hydrolyzing enzymes into pre- and postsynaptic compartments in the rat hippocampus, cerebellum and amygdala. Eur J Neurosci. 2004;20:441–458. doi: 10.1111/j.1460-9568.2004.03428.x. [DOI] [PubMed] [Google Scholar]
- Guo J, Ikeda SR. Endocannabinoids modulate N-type calcium channels and G-protein-coupled inwardly rectifying potassium channels via CB1 cannabinoid receptors heterologously expressed in mammalian neurons. Mol Pharmacol. 2004;65:665–674. doi: 10.1124/mol.65.3.665. [DOI] [PubMed] [Google Scholar]
- Guo Y, Wang H, Okamoto Y, Ueda N, Kingsley PJ, Marnett LJ, Schmid HH, Das SK, Dey SK. N-Acylphosphatidylethanolamine-hydrolyzing phospholipase D is an important determinant of uterine anandamide levels during implantation. J Biol Chem. 2005;280:23429–23432. doi: 10.1074/jbc.C500168200. [DOI] [PubMed] [Google Scholar]
- Guzmán M. Cannabinoids: potential anticancer agents. Nat Rev Cancer. 2003;3:745–755. doi: 10.1038/nrc1188. [DOI] [PubMed] [Google Scholar]
- Guzmán M, Sanchez C, Galve-Roperh I. Cannabinoids and cell fate. Pharmacol Ther. 2002;95:175–184. doi: 10.1016/s0163-7258(02)00256-5. [DOI] [PubMed] [Google Scholar]
- Hájos N, Freund TF. Pharmacological separation of cannabinoid sensitive receptors on hippocampal excitatory and inhibitory fibers. Neuropharmacology. 2002;43:503–510. doi: 10.1016/s0028-3908(02)00157-0. [DOI] [PubMed] [Google Scholar]
- Hájos N, Ledent C, Freund TF. Novel cannabinoid-sensitive receptor mediates inhibition of glutamatergic synaptic transmission in the hippocampus. Neuroscience. 2001;106:1–4. doi: 10.1016/s0306-4522(01)00287-1. [DOI] [PubMed] [Google Scholar]
- Halikas JA, Goodwin DW, Guze SB. Marihuana use and psychiatric illness. Arch Gen Psychiatry. 1972;27:162–165. doi: 10.1001/archpsyc.1972.01750260016003. [DOI] [PubMed] [Google Scholar]
- Hall W, Solowij N. Adverse effects of cannabis. Lancet. 1998;352:1611–1616. doi: 10.1016/S0140-6736(98)05021-1. [DOI] [PubMed] [Google Scholar]
- Hall W, Christie M, Currow D. Cannabinoids and cancer: causation, remediation, and palliation. Lancet Oncol. 2005;6:35–42. doi: 10.1016/S1470-2045(04)01711-5. [DOI] [PubMed] [Google Scholar]
- Hall W, Degenhardt L, Teesson M. Cannabis use and psychotic disorders: an update. Drug Alcohol Rev. 2004;23:433–443. doi: 10.1080/09595230412331324554. [DOI] [PubMed] [Google Scholar]
- Haller J, Bakos N, Szirmay M, Ledent C, Freund TF. The effects of genetic and pharmacological blockade of the CB1 cannabinoid receptor on anxiety. Eur J Neurosci. 2002;16:1395–1398. doi: 10.1046/j.1460-9568.2002.02192.x. [DOI] [PubMed] [Google Scholar]
- Haller J, Varga B, Ledent C, Barna I, Freund TF. Context-dependent effects of CB1 cannabinoid gene disruption on anxiety-like and social behaviour in mice. Eur J Neurosci. 2004a;19:1906–1912. doi: 10.1111/j.1460-9568.2004.03293.x. [DOI] [PubMed] [Google Scholar]
- Haller J, Varga B, Ledent C, Freund TF. CB1 cannabinoid receptors mediate anxiolytic effects: convergent genetic and pharmacological evidence with CB1-specific agents. Behav Pharmacol. 2004b;15:299–304. doi: 10.1097/01.fbp.0000135704.56422.40. [DOI] [PubMed] [Google Scholar]
- Hamann W, di Vadi PP. Analgesic effect of the cannabinoid analogue nabilone is not mediated by opioid receptors. Lancet. 1999;353:560. doi: 10.1016/s0140-6736(98)05429-4. [DOI] [PubMed] [Google Scholar]
- Hampson AJ, Grimaldi M, Axelrod J, Wink D. Cannabidiol and (−) Δ9-tetrahydrocannabinol are neuroprotective antioxidants. Proc Natl Acad Sci USA. 1998;95:8268–8273. doi: 10.1073/pnas.95.14.8268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hampson AJ, Grimaldi M, Lolic M, Wink D, Rosenthal R, Axelrod J. Neuroprotective antioxidants from marijuana. Ann NY Acad Sci. 2000a;899:274–282. [PubMed] [Google Scholar]
- Hampson RE, Mu J, Deadwyler SA. Cannabinoid and κ opioid receptors reduce potassium K current via activation of Gs proteins in cultured hippocampal neurons. J Neurophysiol. 2000b;84:2356–2364. doi: 10.1152/jn.2000.84.5.2356. [DOI] [PubMed] [Google Scholar]
- Hansen HH, Ikonomidou C, Bittigau P, Hansen SH, Hansen HS. Accumulation of the anandamide precursor and other N-acylethanolamine phospholipids in infant rat models of in vivo necrotic and apoptotic neuronal death. J Neurochem. 2001a;76:39–46. doi: 10.1046/j.1471-4159.2001.00006.x. [DOI] [PubMed] [Google Scholar]
- Hansen HH, Schmid PC, Bittigau P, Lastres-Becker I, Berrendero F, Manzanares J, Ikonomidou C, Schmid HH, Fernandez-Ruiz JJ, Hansen HS. Anandamide, but not 2-arachidonoylglycerol, accumulates during in vivo neurodegeneration. J Neurochem. 2001b;78:1415–1427. doi: 10.1046/j.1471-4159.2001.00542.x. [DOI] [PubMed] [Google Scholar]
- Hansson AC, Bermudez-Silva FJ, Malinen H, Hyytia P, Sanchez-Vera I, Rimondini R, Rodriguez de Fonseca F, Kunos G, Sommer WH, Heilig M. Genetic impairment of frontocortical endocannabinoid degradation and high alcohol preference. Neuropsychopharmacology. 2006 doi: 10.1038/sj.npp.1301034. in press. [DOI] [PubMed] [Google Scholar]
- Hanus L, Abu-Lafi S, Fride E, Breuer A, Vogel Z, Shalev DE, Kustanovich I, Mechoulam R. 2-Arachidonyl glyceryl ether, an endogenous agonist of the cannabinoid CB1 receptor. Proc Natl Acad Sci USA. 2001;98:3662–3665. doi: 10.1073/pnas.061029898. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hanus L, Breuer A, Tchilibon S, Shiloah S, Goldenberg D, Horowitz M, Pertwee RG, Ross RA, Mechovlam R, Fride E. HU-308: a specific agonist for CB2, a peripheral cannabinoid receptor. Proc Natl Acad Sci USA. 1999;96:14228–14233. doi: 10.1073/pnas.96.25.14228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hao S, Avraham Y, Mechoulam R, Berry EM. Low dose anandamide affects food intake, cognitive function, neurotransmitter and corticosterone levels in diet-restricted mice. Eur J Pharmacol. 2000;392:147–156. doi: 10.1016/s0014-2999(00)00059-5. [DOI] [PubMed] [Google Scholar]
- Harris GC, Wimmer M, Aston-Jones G. A role for lateral hypothalamic orexin neurons in reward seeking. Nature (Lond) 2005;437:556–559. doi: 10.1038/nature04071. [DOI] [PubMed] [Google Scholar]
- Harrold JA, Elliott JC, King PJ, Widdowson PS, Williams G. Down-regulation of cannabinoid-1 (CB-1) receptors in specific extrahypothalamic regions of rats with dietary obesity: a role for endogenous cannabinoids in driving appetite for palatable food? Brain Res. 2002;952:232–238. doi: 10.1016/s0006-8993(02)03245-6. [DOI] [PubMed] [Google Scholar]
- Harrold JA, Williams G. The cannabinoid system: a role in both the homeostatic and hedonic control of eating? Br J Nutr. 2003;90:729–734. doi: 10.1079/bjn2003942. [DOI] [PubMed] [Google Scholar]
- Hart S, Fischer OM, Ullrich A. Cannabinoids induce cancer cell proliferation via tumor necrosis factor α-converting enzyme (TACE/ADAM17)-mediated transactivation of the epidermal growth factor receptor. Cancer Res. 2004;64:1943–1950. doi: 10.1158/0008-5472.can-03-3720. [DOI] [PubMed] [Google Scholar]
- Hartley JP, Nogrady SG, Seaton A. Bronchodilator effect of Δ1-tetrahydrocannabinol. Br J Clin Pharmacol. 1978;5:523–525. doi: 10.1111/j.1365-2125.1978.tb01667.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hattori N, Mizuno Y. Pathogenetic mechanisms of parkin in Parkinson’s disease. Lancet. 2004;364:722–724. doi: 10.1016/S0140-6736(04)16901-8. [DOI] [PubMed] [Google Scholar]
- Hayakawa K, Mishima K, Abe K, Hasebe N, Takamatsu F, Yasuda H, Ikeda T, Inui K, Egashira N, Iwasaki K, et al. Cannabidiol prevents infarction via the non-CB1 cannabinoid receptor mechanism. Neuroreport. 2004;15:2381–2385. doi: 10.1097/00001756-200410250-00016. [DOI] [PubMed] [Google Scholar]
- Hayase T, Yamamoto Y, Yamamoto K. Protective effects of cannabinoid receptor agonists against cocaine and other convulsant-induced toxic behavioural symptoms. J Pharm Pharmacol. 2001a;53:1525–1532. doi: 10.1211/0022357011777891. [DOI] [PubMed] [Google Scholar]
- Hayase T, Yamamoto Y, Yamamoto K. Protective effects of cannabinoid receptor ligands analogous to anandamide against cocaine toxicity. Nihon Arukoru Yakubutsu Igakkai Zasshi. 2001b;36:596–608. [PubMed] [Google Scholar]
- Hedlund PB, Carson MJ, Sutcliffe JG, Thomas EA. Allosteric regulation by oleamide of the binding properties of 5-hydroxytryptamine7 receptors. Biochem Pharmacol. 1999;58:1807–1813. doi: 10.1016/s0006-2952(99)00274-9. [DOI] [PubMed] [Google Scholar]
- Heilig M, Soderpalm B, Engel JA, Viderlow E. Centrally administered neuropeptide Y (NPY) produces anxiolytic-like effects in animal anxiety models. Psychopharmacology. 1989;98:524–529. doi: 10.1007/BF00441953. [DOI] [PubMed] [Google Scholar]
- Hensen B. Cannabinoid therapeutics: high hopes for the future. Drug Discov Today. 2005;10:459–462. doi: 10.1016/S1359-6446(05)03417-3. [DOI] [PubMed] [Google Scholar]
- Hepler RS, Frank IM, Petrus R. Ocular effects of marijuana. In: Braude M, Szara S, editors. Pharmacology of Cannabis. Raven; New York: 1976. [Google Scholar]
- Hepler RS, Frank IR. Marihuana smoking and intraocular pressure. J Am Med Assoc. 1971;217:1392. [PubMed] [Google Scholar]
- Hepler RS, Frank IM, Ungerleider JT. Pupillary constriction after marijuana smoking. Am J Ophthalmol. 1972;74:1185–1190. doi: 10.1016/0002-9394(72)90741-6. [DOI] [PubMed] [Google Scholar]
- Herkenham M, Groen BG, Lynn AB, De Costa BR, Richfield EK. Neuronal localization of cannabinoid receptors and second messengers in mutant mouse cerebellum. Brain Res. 1991a;552:301–310. doi: 10.1016/0006-8993(91)90096-e. [DOI] [PubMed] [Google Scholar]
- Herkenham M, Lynn AB, Johnson MR, Melvin LS, de Costa BR, Rice KC. Characterization and localization of cannabinoid receptors in rat brain: a quantitative in vitro autoradiographic study. J Neurosci. 1991b;11:563–583. doi: 10.1523/JNEUROSCI.11-02-00563.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herkenham M, Lynn AB, Little MD, Johnson MR, Melvin LS, de Costa BR, Rice KC. Cannabinoid receptor localization in brain. Proc Natl Acad Sci USA. 1990;87:1932–1936. doi: 10.1073/pnas.87.5.1932. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hermann H, Lutz B. Coexpression of the cannabinoid receptor type 1 with the corticotrophin-releasing hormone receptor type 1 in distinct regions of the adult mouse forebrain. Neurosci Lett. 2005;375:13–18. doi: 10.1016/j.neulet.2004.10.080. [DOI] [PubMed] [Google Scholar]
- Herrera B, Carracedo A, Diez-Zaera M, Gomez Del Pulgar T, Guzman M, Velasco G. The CB(2) cannabinoid receptor signals apoptosis via ceramide-dependent activation of the mitochondrial intrinsic pathway. Exp Cell Res. 2006;312:2121–2131. doi: 10.1016/j.yexcr.2006.03.009. [DOI] [PubMed] [Google Scholar]
- Herrera B, Carracedo A, Diez-Zaera M, Guzman M, Velasco G. p38 MAPK is involved in CB2 receptor-induced apoptosis of human leukaemia cells. FEBS Lett. 2005;579:5084–5088. doi: 10.1016/j.febslet.2005.08.021. [DOI] [PubMed] [Google Scholar]
- Herzberg U, Eliav E, Bennett GJ, Kopin IJ. The analgesic effects of R(+)-WIN 55,212-2 mesylate a high affinity cannabinoid agonist, in a rat model of neuropathic pain. Neurosci Lett. 1997;221:157–160. doi: 10.1016/s0304-3940(96)13308-5. [DOI] [PubMed] [Google Scholar]
- Hezode C, Roudot-Thoraval F, Nguyen S, Grenard P, Julien B, Zafrani ES, Pawlostky JM, Dhumeaux D, Lotersztajn S, Mallat A. Daily cannabis smoking as a risk factor for progression of fibrosis in chronic hepatitis C. Hepatology. 2005;42:63–71. doi: 10.1002/hep.20733. [DOI] [PubMed] [Google Scholar]
- Hilairet S, Bouaboula M, Carriere D, Le Fur G, Casellas P. Hypersensitization of the Orexin 1 receptor by the CB1 receptor: evidence for cross-talk blocked by the specific CB1 antagonist, SR141716. J Biol Chem. 2003;278:23731–23737. doi: 10.1074/jbc.M212369200. [DOI] [PubMed] [Google Scholar]
- Hildebrandt AL, Kelly-Sullivan DM, Black SC. Antiobesity effects of chronic cannabinoid CB1 receptor antagonist treatment in diet-induced obese mice. Eur J Pharmacol. 2003;462:125–132. doi: 10.1016/s0014-2999(03)01343-8. [DOI] [PubMed] [Google Scholar]
- Hiley CR, Ford WR. Endocannabinoids as mediators in the heart: a potential target for therapy of remodelling after myocardial infarction? Br J Pharmacol. 2003;138:1183–1184. doi: 10.1038/sj.bjp.0705155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hiley CR, Ford WR. Cannabinoid pharmacology in the cardiovascular system: potential protective mechanisms through lipid signaling. Biol Rev Camb Philos Soc. 2004;79:187–205. doi: 10.1017/s1464793103006201. [DOI] [PubMed] [Google Scholar]
- Hill MN, Ho WS, Sinopoli KJ, Viau V, Hillard CJ, Gorzalka BB. Involvement of the endocannabinoid system in the ability of long-term tricyclic antide-pressant treatment to suppress stress-induced activation of the hypothalamic-pituitary-adrenal axis. Neuropsychopharmacology. 2006 doi: 10.1038/sj.npp.1301092. in press. [DOI] [PubMed] [Google Scholar]
- Hill MN, Patel S, Carrier EJ, Rademacher DJ, Ormerod BK, Hillard CJ, Gorzalka BB. Downregulation of endocannabinoid signaling in the hippocampus following chronic unpredictable stress. Neuropsychopharmacology. 2005;30:508–515. doi: 10.1038/sj.npp.1300601. [DOI] [PubMed] [Google Scholar]
- Hillard CJ. Endocannabinoids and vascular function. J Pharmacol Exp Ther. 2000;294:27–32. [PubMed] [Google Scholar]
- Hillard CJ, Jarrahian A. The movement of N-arachidonolethanolamide (anandamide) across cellular membranes. Chem Phys Lipids. 2000;108:123–134. doi: 10.1016/s0009-3084(00)00191-2. [DOI] [PubMed] [Google Scholar]
- Hillard CJ, Jarrahian A. Cellular accumulation of anandamide: consensus and controversy. Br J Pharmacol. 2003;140:802–808. doi: 10.1038/sj.bjp.0705468. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hillard CJ, Manna S, Greenberg MJ, DiCamelli R, Ross RA, Stevenson LA, Murphy V, Pertwee RG, Campbell WB. Synthesis and characterization of potent and selective agonists of the neuronal cannabinoid receptor (CB1) J Pharmacol Exp Ther. 1999;289:1427–1433. [PubMed] [Google Scholar]
- Hinderer SR, Gupta S. Functional outcome measures to assess interventions for spasticity. Arch Phys Med Rehabil. 1996;77:1083–1089. doi: 10.1016/s0003-9993(96)90073-8. [DOI] [PubMed] [Google Scholar]
- Hinds NM, Ullrich K, Smid SD. Cannabinoid 1 (CB1 receptors coupled to cholinergic motorneurones inhibit neurogenic circular muscle contractility in the human colon. Br J Pharmacol. 2006;148:191–200. doi: 10.1038/sj.bjp.0706710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hinz B, Ramer R, Eichele K, Weinzierl U, Brune K. Up-regulation of cyclooxygenase-2 expression is involved in R(+)-methanandamide-induced apoptotic death of human neuroglioma cells. Mol Pharmacol. 2004;66:1643–1651. doi: 10.1124/mol.104.002618. [DOI] [PubMed] [Google Scholar]
- Ho WS, Hiley CR. Vasodilator actions of abnormal-cannabidiol in rat isolated small mesenteric artery. Br J Pharmacol. 2003;138:1320–1332. doi: 10.1038/sj.bjp.0705160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hohmann AG, Briley EM, Herkenham M. Pre- and postsynaptic distribution of cannabinoid and mu opioid receptors in rat spinal cord. Brain Res. 1999;822:17–25. doi: 10.1016/s0006-8993(98)01321-3. [DOI] [PubMed] [Google Scholar]
- Hohmann AG, Farthing JN, Zvonok AM, Makriyannis A. Selective activation of cannabinoid CB2 receptors suppresses hyperalgesia evoked by intradermal capsaicin. J Pharmacol Exp Ther. 2004;308:446–453. doi: 10.1124/jpet.103.060079. [DOI] [PubMed] [Google Scholar]
- Hohmann AG, Herkenham M. Regulation of cannabinoid and mu opioid receptors in rat lumbar spinal cord following neonatal capsaicin treatment. Neurosci Lett. 1998;252:13–16. doi: 10.1016/s0304-3940(98)00534-5. [DOI] [PubMed] [Google Scholar]
- Hohmann AG, Herkenham M. Cannabinoid receptors undergo axonal flow in sensory nerves. Neuroscience. 1999;92:1171–1175. doi: 10.1016/s0306-4522(99)00220-1. [DOI] [PubMed] [Google Scholar]
- Hohmann AG, Herkenham M. Localization of cannabinoid CB1 receptor mRNA in neuronal subpopulations of rat striatum: a double-label in situ hybridization study. Synapse. 2000;37:71–80. doi: 10.1002/(SICI)1098-2396(200007)37:1<71::AID-SYN8>3.0.CO;2-K. [DOI] [PubMed] [Google Scholar]
- Hohmann AG, Martin WJ, Tsou K, Walker JM. Inhibition of noxious stimulus-evoked activity of spinal cord dorsal horn neurons by the cannabinoid WIN 55,212-2. Life Sci. 1995;56:2111–2119. doi: 10.1016/0024-3205(95)00196-d. [DOI] [PubMed] [Google Scholar]
- Hohmann AG, Suplita RL, Bolton NM, Neely MH, Fegley D, Mangieri R, Krey JF, Walker JM, Holmes PV, Crystal JD, et al. An endocannabinoid mechanism for stress-induced analgesia. Nature (Lond) 2005;435:1108–1112. doi: 10.1038/nature03658. [DOI] [PubMed] [Google Scholar]
- Hohmann AG, Tsou K, Walker JM. Cannabinoid modulation of wide dynamic range neurons in the lumbar dorsal horn of the rat by spinally administered WIN55,212-2. Neurosci Let. 1998;257:1–4. doi: 10.1016/s0304-3940(98)00802-7. [DOI] [PubMed] [Google Scholar]
- Hohmann AG, Walker JM. Cannabinoid suppression of noxious heat-evoked activity in wide dynamic range neurons in the lumbar dorsal horn of the rat. J Neurophysiol. 1999;81:575–583. doi: 10.1152/jn.1999.81.2.575. [DOI] [PubMed] [Google Scholar]
- Holdcroft A, Smith M, Jacklin A, Hodgson H, Smith B, Newton M, Evans F. Pain relief with oral cannabinoids in familial Mediterranean fever. Anaesthesia. 1997;52:483–486. doi: 10.1111/j.1365-2044.1997.139-az0132.x. [DOI] [PubMed] [Google Scholar]
- Hollister LE. Hunger and appetite after single doses of marijuana, alcohol and dextroamphetamine. Clin Pharmacol Ther. 1971;12:45–49. doi: 10.1002/cpt197112144. [DOI] [PubMed] [Google Scholar]
- Hollister LE. Structure-activity relationships in man of cannabis constituents, and homologs and metabolites of Δ9-tetrahydrocannabinol. Pharmacology. 1974;11:3–11. doi: 10.1159/000136462. [DOI] [PubMed] [Google Scholar]
- Hollister LE. Health aspects of cannabis. Pharmacol Rev. 1986;38:1–20. [PubMed] [Google Scholar]
- Holdcroft A, Maze M, Dore C, Tebbs S, Thompson S. A multicenter dose-escalation study of the analgesic and adverse effects of an oral cannabis extract (Cannador) for postoperative pain management. Anesthesiology. 2006;104:1040–1046. doi: 10.1097/00000542-200605000-00021. [DOI] [PubMed] [Google Scholar]
- Holm L, Cassidy JD, Carroll LJ, Borg J. Neurotrauma Task Force on Mild Traumatic Brain Injury of the WHO Collaborating Centre: summary of the WHO Collaborating Centre for Neurotrauma Task Force on mild traumatic brain injury. J Rehabil Med. 2005;37:137–141. doi: 10.1080/16501970510027321. [DOI] [PubMed] [Google Scholar]
- Holmes GL, Ben-Ari Y. Seizures in the developing brain: perhaps not so benign after all. Neuron. 1998;21:1231–1234. doi: 10.1016/s0896-6273(00)80642-x. [DOI] [PubMed] [Google Scholar]
- Horn S, Stern MB. The comparative effects of medical therapies for Parkinson’s disease. Neurology. 2004;63:S7–S12. doi: 10.1212/wnl.63.7_suppl_2.s7. [DOI] [PubMed] [Google Scholar]
- Houchi H, Babovic D, Pierrefiche O, Ledent C, Daoust M, Naassila M. CB1 receptor knockout mice display reduced ethanol-induced conditioned place preference and increased striatal dopamine D2 receptors. Neuropsychopharmacology. 2004;30:339–349. doi: 10.1038/sj.npp.1300568. [DOI] [PubMed] [Google Scholar]
- Houser SJ, Eads M, Embrey JP, Welch SP. Dynorphin B and spinal analgesia: induction of antinociception by the cannabinoids CP55,940, Δ9-THC and anandamide. Brain Res. 2000;857:337–342. doi: 10.1016/s0006-8993(00)01981-8. [DOI] [PubMed] [Google Scholar]
- Howlett AC. Efficacy in CB1 receptor-mediated signal transduction. Br J Pharmacol. 2004;142:1209–1218. doi: 10.1038/sj.bjp.0705881. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Howlett AC, Barth F, Bonner TI, Cabral G, Casellas P, Devane WA, Felder CC, Herkenham M, Mackie K, Martin BR, et al. International Union of Pharmacology. XXVII. Classification of cannabinoid receptors. Pharmacol Rev. 2002;54:161–202. doi: 10.1124/pr.54.2.161. [DOI] [PubMed] [Google Scholar]
- Howlett AC, Breivogel CS, Childers SR, Deadwyler SA, Hampson RE, Porrino LJ. Cannabinoid physiology and pharmacology: 30 years of progress. Neuropharmacology. 2004;47 (Suppl 1):345–358. doi: 10.1016/j.neuropharm.2004.07.030. [DOI] [PubMed] [Google Scholar]
- Hu CC, Qing K, Chen Y. Diet-induced changes in stearoyl-CoA desaturase 1 expression in obesity-prone and -resistant mice. Obes Res. 2004;12:1264–1270. doi: 10.1038/oby.2004.160. [DOI] [PubMed] [Google Scholar]
- Huang SM, Bisogno T, Trevisani M, Al-Hayani A, De Petrocellis L, Fezza F, Tognetto M, Petros TJ, Krey JF, Chu CJ, et al. An endogenous capsaicin-like substance with high potency at recombinant and native vanilloid VR1 receptors. Proc Natl Acad Sci USA. 2002;99:8400–8405. doi: 10.1073/pnas.122196999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang YC, Wang SJ, Chiou LC, Gean PW. Mediation of amphetamine-induced long-term depression of synaptic transmission by CB1 cannabinoid receptors in the rat amygdala. J Neurosci. 2003;23:10311–10320. doi: 10.1523/JNEUROSCI.23-32-10311.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hübschle T, Thom E, Watson A, Roth J, Klaus S, Meyerhof W. Leptin-induced nuclear translocation of STAT3 immunoreactivity in hypothalamic nuclei involved in body weight regulation. J Neurosci. 2001;21:2413–2424. doi: 10.1523/JNEUROSCI.21-07-02413.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huffman JW, Liddle J, Yu S, Aung MM, Abood ME, Wiley JL, Martin BR. 3-(1′,1′-Dimethylbutyl)-1-deoxy-Δ8-THC and related compounds: synthesis of selective ligands for the CB2 receptor. Bioorg Med Chem. 1999;7:2905–2914. doi: 10.1016/s0968-0896(99)00219-9. [DOI] [PubMed] [Google Scholar]
- Hungund BL, Szakall I, Adam A, Basavarajappa BS, Vadasz C. Cannabinoid CB1 receptor knockout mice exhibit markedly reduced voluntary alcohol consumption and lack alcohol-induced dopamine release in the nucleus accumbens. J Neurochem. 2003;84:698–704. doi: 10.1046/j.1471-4159.2003.01576.x. [DOI] [PubMed] [Google Scholar]
- Ibrahim MM, Deng H, Zvonok A, Cockayne DA, Kwan J, Mata HP, Vanderah TW, Lai J, Porreca F, Makriyannis A, et al. Activation of CB2 cannabinoid receptors by AM1241 inhibits experimental neuropathic pain: pain inhibition by receptors not present in the CNS. Proc Natl Acad Sci USA. 2003;100:10529–10533. doi: 10.1073/pnas.1834309100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ibrahim MM, Porreca F, Lai J, Albrecht PJ, Rice FL, Khodorova A, Davar G, Makriyannis A, Vanderah TW, Mata HP, et al. CB2 cannabinoid receptor activation produces antinociception by stimulating peripheral release of endogenous opioids. Proc Natl Acad Sci USA. 2005;102:3093–3098. doi: 10.1073/pnas.0409888102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ibrahim MM, Rude ML, Stagg NJ, Mata HP, Lai J, Vanderah TW, Porreca F, Buckley NE, Makriyannis A, Malan TP., Jr CB2 cannabinoid receptor mediation of antinociception. Pain. 2006;122:36–42. doi: 10.1016/j.pain.2005.12.018. [DOI] [PubMed] [Google Scholar]
- Idris AI, van’t Hof RJ, Greig IR, Ridge SA, Baker D, Ross RA, Ralston SH. Regulation of bone mass, bone loss and osteoclast activity by cannabinoid receptors. Nat Med. 2005;11:774–779. doi: 10.1038/nm1255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ikeda SR. Voltage-dependent modulation of N-type calcium channels by G-protein βγsubunits. Nature (Lond) 1996;380:255–258. doi: 10.1038/380255a0. [DOI] [PubMed] [Google Scholar]
- Ilaria RL, Thornby JI, Fann WE. Nabilone, a cannabinol derivative, in the treatment of anxiety neurosis. Curr Ther Res. 1981;29:943–949. [Google Scholar]
- Iqbal J, Pompolo S, Murakami T, Grouzmann E, Sakurai T, Meister B, Clarke IJ. Immunohistochemical characterization of localization of long-form leptin receptor (OB-Rb) in neurochemically defined cells in the ovine hypothalamus. Brain Res. 2001;920:55–64. doi: 10.1016/s0006-8993(01)02932-8. [DOI] [PubMed] [Google Scholar]
- Ishac EJ, Jiang L, Lake KD, Varga K, Abood ME, Kunos G. Inhibition of exocytotic noradrenaline release by presynaptic cannabinoid CB1 receptors on peripheral sympathetic nerves. Br J Pharmacol. 1996;118:2023–2028. doi: 10.1111/j.1476-5381.1996.tb15639.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Isokawa M, Alger BE. Retrograde endocannabinoid regulation of GABAergic inhibition in the rat dentate gyrus granule cell. J Physiol (Lond) 2005;567 (Pt 3):1001–1010. doi: 10.1113/jphysiol.2005.094219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iuvone T, Esposito G, Esposito R, Santamaria R, Di Rosa M, Izzo AA. Neuroprotective effect of cannabidiol, a non-psychoactive component from Cannabis sativa, on β-amyloid-induced toxicity in PC12 cells. J Neurochem. 2004;89:134–141. doi: 10.1111/j.1471-4159.2003.02327.x. [DOI] [PubMed] [Google Scholar]
- Iversen LL. The Science of Marijuana. Oxford University Press; Oxford, UK: 2000. [Google Scholar]
- Iwamura H, Suzuki H, Ueda Y, Kaya T, Inaba T. In vitro and in vivo pharmacological characterization of JTE-907, a novel selective ligand for cannabinoid CB2 receptor. J Pharmacol Exp Ther. 2001;296:420–425. [PubMed] [Google Scholar]
- Izzo AA, Capasso F, Costagliola A, Bisogno T, Marsicano G, Ligresti A, Matias I, Capasso R, Pinto L, Borrelli F, et al. An endogenous cannabinoid tone attenuates cholera toxin-induced fluidaccumulation in mice. Gastroenterology. 2003;125:765–774. doi: 10.1016/s0016-5085(03)00892-8. [DOI] [PubMed] [Google Scholar]
- Izzo AA, Fezza F, Capasso R, Bisogno T, Pinto L, Iuvone T, Esposito G, Mascolo N, Di Marzo V, Capasso F. Cannabinoid CB1-receptor mediated regulation of gastrointestinal motility in mice in a model of intestinal inflammation. Br J Pharmacol. 2001a;134:563–570. doi: 10.1038/sj.bjp.0704293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Izzo AA, Mascolo N, Capasso F. The gastrointestinal pharmacology of cannabinoids. Curr Opin Pharmacol. 2001b;1:597–603. doi: 10.1016/s1471-4892(01)00102-3. [DOI] [PubMed] [Google Scholar]
- Jabusch HC, Schneider U, Altenmuller E. Δ9-Tetrahydrocannabinol improves motor control in a patient with musician’s dystonia. Mov Disord. 2004;19:990–991. doi: 10.1002/mds.20214. [DOI] [PubMed] [Google Scholar]
- Jackson SJ, Diemel LT, Pryce G, Baker D. Cannabinoids and neuroprotection in CNS inflammatory disease. J Neurol Sci. 2005a;233:21–25. doi: 10.1016/j.jns.2005.03.002. [DOI] [PubMed] [Google Scholar]
- Jackson SJ, Pryce G, Diemel LT, Cuzner ML, Baker D. Cannabinoid-receptor 1 null mice are susceptible to neurofilament damage and caspase 3 activation. Neuroscience. 2005b;134:261–266. doi: 10.1016/j.neuroscience.2005.02.045. [DOI] [PubMed] [Google Scholar]
- Jacobsson SO, Rongard E, Stridh M, Tiger G, Fowler CJ. Serum-dependent effects of tamoxifen and cannabinoids upon C6 glioma cell viability. Biochem Pharmacol. 2000;60:1807–1813. doi: 10.1016/s0006-2952(00)00492-5. [DOI] [PubMed] [Google Scholar]
- Jayamanne A, Greenwood R, Mitchell VA, Aslan S, Piomelli D, Vaughan CW. Actions of the FAAH inhibitor URB597 in neuropathic and inflammatory chronic pain models. Br J Pharmacol. 2006;147:281–288. doi: 10.1038/sj.bjp.0706510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jamshidi N, Taylor DA. Anandamide administration into the ventromedial hypothalamus stimulates appetite in rats. Br J Pharmacol. 2001;134:1151–1154. doi: 10.1038/sj.bjp.0704379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jan TR, Farraj AK, Harkema JR, Kaminski NE. Attenuation of the ovalbumin-induced allergic airway response by cannabinoid treatment in A/J mice. Toxicol Appl Pharmacol. 2003;188:24–35. doi: 10.1016/s0041-008x(03)00010-3. [DOI] [PubMed] [Google Scholar]
- Járai Z, Wagner JA, Varga K, Lake KD, Compton DR, Martin BR, Zimmer AM, Bonner TI, Buckley NE, Mezey E, et al. Cannabinoid-induced mesenteric vasodilation through an endothelial site distinct from CB1 or CB2 receptors. Proc Natl Acad Sci USA. 1999;96:14136–14141. doi: 10.1073/pnas.96.24.14136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jarrahian A, Manna S, Edgemond WS, Campbell WB, Hillard CJ. Structure-activity relationships among N-arachidonylethanolamine (Anandamide) head group analogues for the anandamide transporter. J Neurochem. 2000;74:2597–2606. doi: 10.1046/j.1471-4159.2000.0742597.x. [DOI] [PubMed] [Google Scholar]
- Jarvinen T, Pate DW, Laine K. Cannabinoids in the treatment of glaucoma. Pharmacol Ther. 2002;95:203–220. doi: 10.1016/s0163-7258(02)00259-0. [DOI] [PubMed] [Google Scholar]
- Jbilo O, Ravinet Trillou C, Arnone M, Buisson I, Bribes E, Péleraux A, Pénarier G, Soubrié P, Le Fur G, Galiège S, et al. The CB1 receptor antagonist rimonabant reverses the diet-induced obesity phenotype through the regulation of lipolysis and energy balance. FASEB J. 2005;19:1567–1569. doi: 10.1096/fj.04-3177fje. [DOI] [PubMed] [Google Scholar]
- Jiang W, Zhang Y, Xiao L, Van Cleemput J, Ji SP, Bai G, Zhang X. Cannabinoids promote embryonic and adult hippocampus neurogenesis and produce anxiolytic- and antidepressant-like effects. J Clin Investig. 2005;115:3104–3116. doi: 10.1172/JCI25509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jimenez W. Endocannabinoids and liver disease. Hepatology. 2005;41:983–985. doi: 10.1002/hep.20714. [DOI] [PubMed] [Google Scholar]
- Jin KL, Mao XO, Goldsmith PC, Greenberg DA. CB1 cannabinoid receptor induction in experimental stroke. Ann Neurol. 2000;48:257–261. [PubMed] [Google Scholar]
- Jin W, Brown S, Roche JP, Hsieh C, Celver JP, Kovoor A, Chavkin C, Mackie K. Distinct domains of the CB1 cannabinoid receptor mediate desensitization and internalization. J Neurosci. 1999;19:3773–3780. doi: 10.1523/JNEUROSCI.19-10-03773.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jo Y-H, Chen Y-JJ, Chua SC, Jr, Talmage DA, Role LW. Integration of endocannabinoid and leptin signaling in an appetite-related neural circuit. Neuron. 2005;48:1055–1066. doi: 10.1016/j.neuron.2005.10.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johanek LM, Heitmiller DR, Turner M, Nader N, Hodges J, Simone DA. Cannabinoids attenuate capsaicin-evoked hyperalgesia through spinal and peripheral mechanisms. Pain. 2001;93:303–315. doi: 10.1016/S0304-3959(01)00336-0. [DOI] [PubMed] [Google Scholar]
- Johanek LM, Simone DA. Activation of peripheral cannabinoid receptors attenuates cutaneous hyperalgesia produced by a heat injury. Pain. 2004;109:432–442. doi: 10.1016/j.pain.2004.02.020. [DOI] [PubMed] [Google Scholar]
- Johns A. Psychiatric effects of cannabis. Br J Psychiatry. 2001;178:116–122. doi: 10.1192/bjp.178.2.116. [DOI] [PubMed] [Google Scholar]
- Johnson SW, North RA. Opioids excite dopamine neurons by hyperpolarization of local interneurons. J Neurosci. 1992;12:483–488. doi: 10.1523/JNEUROSCI.12-02-00483.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnson DD, McNeill JR, Crawford RD, Wilcox WC. Epileptiform seizures in domestic fowl. V. The anticonvulsant activity of Δ9-tetrahydrocannabinol. Can J Physiol Pharmacol. 1975;53:1007–1013. doi: 10.1139/y75-140. [DOI] [PubMed] [Google Scholar]
- Jones G, Pertwee RG, Gill EW, Paton WD, Nilsson IM, Widman M, Agurell S. Relative pharmacological potency in mice of optical isomers of Δ1-tetrahydrocannabinol. Biochem Pharmacol. 1974;23:439–446. doi: 10.1016/0006-2952(74)90435-3. [DOI] [PubMed] [Google Scholar]
- Jones S, Howl J. Cannabinoid receptor systems: therapeutic targets for tumour intervention. Expert Opin Ther Targets. 2003;7:749–758. doi: 10.1517/14728222.7.6.749. [DOI] [PubMed] [Google Scholar]
- Jonsson KO, Andersson A, Jacobsson SO, Vandevoorde S, Lambert DM, Fowler CJ. AM404 and VDM 11 non-specifically inhibit C6 glioma cell proliferation at concentrations used to block the cellular accumulation of the endocannabinoid anandamide. Arch Toxicol. 2003;77:201–207. doi: 10.1007/s00204-002-0435-6. [DOI] [PubMed] [Google Scholar]
- Jordt SE, Bautista DM, Chuang HH, McKemy DD, Zygmunt PM, Hogestatt ED, Meng ID, Julius D. Mustard oils and cannabinoids excite sensory nerve fibres through the TRP channel ANKTM1. Nature (Lond) 2004;427:260–265. doi: 10.1038/nature02282. [DOI] [PubMed] [Google Scholar]
- Joseph J, Niggemann B, Zaenker KS, Entschladen F. Anandamide is an endogenous inhibitor for the migration of tumor cells and T lymphocytes. Cancer Immunol Immunother. 2004;53:723–728. doi: 10.1007/s00262-004-0509-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Joyeux M, Arnaud C, Godin-Ribuot D, Demenge P, Lamontagne D, Ribuot C. Endocannabinoids are implicated in the infarct size-reducing effect conferred by heat stress preconditioning in isolated rat hearts. Cardiovasc Res. 2002;55:619–625. doi: 10.1016/s0008-6363(02)00268-7. [DOI] [PubMed] [Google Scholar]
- Juan-Pico P, Fuentes E, Javier Bermudez-Silva F, Javier Diaz-Molina F, Ripoll C, Rodriguez de Fonseca F, Nadal A. Cannabinoid receptors regulate Ca2+ signals and insulin secretion in pancreatic β-cell. Cell Calcium. 2005;39:155–162. doi: 10.1016/j.ceca.2005.10.005. [DOI] [PubMed] [Google Scholar]
- Julien B, Grenard P, Teixeira-Clerc F, Van Nhieu JT, Li L, Karsak M, Zimmer A, Mallat A, Lotersztajn S. Antifibrogenic role of the cannabinoid receptor CB2 in the liver. Gastroenterology. 2005;128:742–755. doi: 10.1053/j.gastro.2004.12.050. [DOI] [PubMed] [Google Scholar]
- Kadoi Y, Hinohara H, Kunimoto F, Kuwano H, Saito S, Goto F. Effects of AM281, a cannabinoid antagonist, on systemic haemodynamics, internal carotid artery blood flow and mortality in septic shock in rats. Br J Anaesth. 2005a;94:563–568. doi: 10.1093/bja/aei106. [DOI] [PubMed] [Google Scholar]
- Kadoi Y, Hinohara H, Kunimoto F, Saito S, Goto F. Cannabinoid antagonist AM 281 reduces mortality rate and neurologic dysfunction after cecal ligation and puncture in rats. Crit Care Med. 2005b;33:2629–2636. doi: 10.1097/01.ccm.0000187010.14426.cc. [DOI] [PubMed] [Google Scholar]
- Kamei J, Yoshikawa Y, Saitoh A. Effect of N-arachidonoyl-(2-methyl-4-hydroxyphenyl) amine (VDM11), an anandamide transporter inhibitor, on capsaicin-induced cough in mice. Cough. 2006;2:2. doi: 10.1186/1745-9974-2-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kanakis C, Jr, Pouget JM, Rosen KM. The effects of delta-9-tetrahydrocannabinol (cannabis) on cardiac performance with and without beta blockade. Circulation. 1976;53:703–707. doi: 10.1161/01.cir.53.4.703. [DOI] [PubMed] [Google Scholar]
- Kaplan BL, Ouyang Y, Herring A, Yea SS, Razdan R, Kaminski NE. Inhibition of leukocyte function and interleukin-2 gene expression by 2-methyl-arachidonyl-(2′-fluoroethyl)amide, a stable congener of the endogenous cannabinoid receptor ligand anandamide. Toxicol Appl Pharmacol. 2005;205:107–115. doi: 10.1016/j.taap.2004.09.013. [DOI] [PubMed] [Google Scholar]
- Karler R, Calder LD, Sangdee P, Turkanis SA. Interaction between Δ-9-tetrahydrocannabinol and kindling by electrical and chemical stimuli in mice. Neuropharmacology. 1984;23:1315–1320. doi: 10.1016/0028-3908(84)90052-2. [DOI] [PubMed] [Google Scholar]
- Karler R, Calder LD, Turkanis SA. Prolonged CNS hyperexcitability in mice after a single exposure to Δ-9-tetrahydrocannabinol. Neuropharmacology. 1986;25:441–446. doi: 10.1016/0028-3908(86)90240-6. [DOI] [PubMed] [Google Scholar]
- Karler R, Turkanis SA. The cannabinoids as potential antiepileptics. J Clin Pharmacol. 1981;21:437S–448S. doi: 10.1002/j.1552-4604.1981.tb02624.x. [DOI] [PubMed] [Google Scholar]
- Karsak M, Cohen-Solal M, Freudenberg J, Ostertag A, Morieux C, Kornak U, Essig J, Erxlebe E, Bab I, Kubisch C, et al. The cannabinoid receptor type 2 (CNR2) gene is associated with human osteoporosis. Hum Mol Genet. 2005;14:3389–3396. doi: 10.1093/hmg/ddi370. [DOI] [PubMed] [Google Scholar]
- Karsak M, Ofek O, Fogel M, Wright K, Tam J, Gabet Y, Birenboim R, Attar-Namdar M, Müller R, Cohen-Solal M. The cannabinoid CB2 receptor: a potential target for the treatment of osteoporosis. J Bone Miner Res. 2004;19:S383. [Google Scholar]
- Karst M, Salim K, Burstein S, Conrad I, Hoy L, Schneider U. Analgesic effect of the synthetic cannabinoid CT-3 on chronic neuropathic pain: a randomized controlled trial. J Am Med Assoc. 2003;290:1757–1762. doi: 10.1001/jama.290.13.1757. [DOI] [PubMed] [Google Scholar]
- Kathuria S, Gaetani S, Fegley D, Valino F, Duranti A, Tontini A, Mor M, Tarzia G, La Rana G, Calignano A, et al. Modulation of anxiety through blockade of anandamide hydrolysis. Nat Med. 2003;9:76–81. doi: 10.1038/nm803. [DOI] [PubMed] [Google Scholar]
- Katona I, Rancz EA, Acsady L, Ledent C, Mackie K, Hajos N, Freund TF. Distribution of CB1 cannabinoid receptors in the amygdala and their role in the control of GABAergic transmission. J Neurosci. 2001;21:9506–9518. doi: 10.1523/JNEUROSCI.21-23-09506.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Katona I, Sperlagh B, Sik A, Kafalvi A, Vizi ES, Mackie K, Freund TF. Presynaptically located CB1 cannabinoid receptors regulate GABA release from axon terminals of specific hippocampal interneurons. J Neurosci. 1999;19:4544–4558. doi: 10.1523/JNEUROSCI.19-11-04544.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kearn CS, Blake-Palmer K, Daniel E, Mackie K, Glass M. Concurrent stimulation of cannabinoid CB1 and dopamine D2 receptors enhances heterodimer formation: a mechanism for receptor cross-talk? Mol Pharmacol. 2005;67:1697–1704. doi: 10.1124/mol.104.006882. [DOI] [PubMed] [Google Scholar]
- Keeler MH, Reifler CB. Grand mal convulsions subsequent to marijuana use: case report. Dis Nerv Syst. 1967;28:474–475. [PubMed] [Google Scholar]
- Keen RW. Burden of osteoporosis and fractures. Curr Osteoporos Rep. 2003;1:66–70. doi: 10.1007/s11914-003-0011-x. [DOI] [PubMed] [Google Scholar]
- Kelly LA, Butcher RW. The effects of Δ1-tetrahydrocannabinol on cyclic AMP levels in WI-38 fibroblasts. Biochim Biophys Acta. 1973;320:540–544. doi: 10.1016/0304-4165(73)90335-8. [DOI] [PubMed] [Google Scholar]
- Kelly S, Jhaveri MD, Sagar DR, Kendall DA, Chapman V. Activation of peripheral cannabinoid CB1 receptors inhibits mechanically evoked responses of spinal neurons in noninflamed rats and rats with hindpaw inflammation. Eur J Neurosci. 2003;18:2239–2243. doi: 10.1046/j.1460-9568.2003.02957.x. [DOI] [PubMed] [Google Scholar]
- Khanolkar AD, Abadji V, Lin S, Hill WA, Taha G, Abouzid K, Meng Z, Fan P, Makriyannis A. Head group analogs of arachidonylethanolamide, the endogenous cannabinoid ligand. J Med Chem. 1996;39:4515–4519. doi: 10.1021/jm960152y. [DOI] [PubMed] [Google Scholar]
- Khaspekov LG, Brenz Verca MS, Frumkina LE, Hermann H, Marsicano G, Lutz B. Involvement of brain-derived neurotrophic factor in cannabinoid receptor-dependent protection against excitotoxicity. Eur J Neurosci. 2004;19:1691–1698. doi: 10.1111/j.1460-9568.2004.03285.x. [DOI] [PubMed] [Google Scholar]
- Killestein J, Hoogervorst EL, Reif M, Blauw B, Smits M, Uitdehaag BM, Nagelkerken L, Polman CH. Immunomodulatory effects of orally administered cannabinoids in multiple sclerosis. J Neuroimmunol. 2003;137:140–143. doi: 10.1016/s0165-5728(03)00045-6. [DOI] [PubMed] [Google Scholar]
- Killestein J, Hoogervorst EL, Reif M, Kalkers NF, Van Loenen AC, Staats PG, Gorter RW, Uitdehaag BM, Polman CH. Safety, tolerability, and efficacy of orally administered cannabinoids in MS. Neurology. 2002;58:1404–1407. doi: 10.1212/wnl.58.9.1404. [DOI] [PubMed] [Google Scholar]
- Killestein J, Polman CH. Current trials in multiple sclerosis: established evidence and future hopes. Curr Opin Neurol. 2005;18:253–260. doi: 10.1097/01.wco.0000169741.29535.cc. [DOI] [PubMed] [Google Scholar]
- Killestein J, Uitdehaag BM, Polman CH. Cannabinoids in multiple sclerosis: do they have a therapeutic role? Drugs. 2004;64:1–11. doi: 10.2165/00003495-200464010-00001. [DOI] [PubMed] [Google Scholar]
- Kim EK, Miller I, Aja S, Landree LE, Pinn M, McFadden J, Kuhajda FP, Moran TH, Ronnett GV. C75, a fatty acid synthase inhibitor, reduces food intake via hypothalamic AMP-activated protein kinase. J Biol Chem. 2004;279:19970–19976. doi: 10.1074/jbc.M402165200. [DOI] [PubMed] [Google Scholar]
- Kim J, Alger BE. Inhibition of cyclooxygenase-2 potentiates retrograde endocannabinoid effects in hippocampus. Nat Neurosci. 2004;7:697–698. doi: 10.1038/nn1262. [DOI] [PubMed] [Google Scholar]
- Kim J, Isokawa M, Ledent C, Alger BE. Activation of muscarinic acetylcholine receptors enhances the release of endogenous cannabinoids in the hippocampus. J Neurosci. 2002;22:10182–10191. doi: 10.1523/JNEUROSCI.22-23-10182.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim SH, Won SJ, Mao XO, Jin K, Greenberg DA. Involvement of protein kinase A in cannabinoid receptor-mediated protection from oxidative neuronal injury. J Pharmacol Exp Ther. 2005;313:88–94. doi: 10.1124/jpet.104.079509. [DOI] [PubMed] [Google Scholar]
- Kirkham TC. Cannabinoids and medicine: eating disorders, nausea and emesis. In: Di Marzo V, editor. Cannabinoids. Landes Bioscience; Georgetown, TX: 2004. pp. 147–160. [Google Scholar]
- Kirkham TC. Endocannabinoids in the regulation of appetite and body weight. Behav Pharmacol. 2005;16:297–313. doi: 10.1097/00008877-200509000-00004. [DOI] [PubMed] [Google Scholar]
- Kirkham TC, Williams CM. Endogenous cannabinoids and appetite. Nutr Res Rev. 2001a;14:65–86. doi: 10.1079/NRR200118. [DOI] [PubMed] [Google Scholar]
- Kirkham TC, Williams CM. Synergistic effects of opioid and cannabinoid antagonists on food intake. Psychopharmacology. 2001b;159:267–270. doi: 10.1007/s002130000596. [DOI] [PubMed] [Google Scholar]
- Kirkham TC, Williams CM, Fezza M, Di Marzo V. Endocannabinoid levels in rat limbic forebrain and hypothalamus in relation to fasting, feeding and satiation: stimulation of eating by 2-arachidonoyl glycerol. Br J Pharmacol. 2002;136:550–557. doi: 10.1038/sj.bjp.0704767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klein TW. Cannabinoid-based drugs as anti-inflammatory therapeutics. Nat Rev Immunol. 2005;5:400–411. doi: 10.1038/nri1602. [DOI] [PubMed] [Google Scholar]
- Klein TW, Friedman H, Specter SC. Marijuana, immunity and infection. J Neuroimmunol. 1998;83:102–115. doi: 10.1016/s0165-5728(97)00226-9. [DOI] [PubMed] [Google Scholar]
- Klein TW, Lane B, Newton CA, Friedman H. The cannabinoid system and cytokine network. Proc Soc Exp Biol Med. 2000a;225:1–8. doi: 10.1177/153537020022500101. [DOI] [PubMed] [Google Scholar]
- Klein TW, Newton C, Larsen K, Lu L, Perkins I, Nong L, Friedman H. The cannabinoid system and immune modulation. J Leukoc Biol. 2003;74:486–496. doi: 10.1189/jlb.0303101. [DOI] [PubMed] [Google Scholar]
- Klein TW, Newton CA, Nakachi N, Friedman H. Δ9-Tetrahydrocannab-inol treatment suppresses immunity and early IFN-γ, IL-12, and IL-12 receptor β 2 responses to Legionella pneumophila infection. J Immunol. 2000b;164:6461–6466. doi: 10.4049/jimmunol.164.12.6461. [DOI] [PubMed] [Google Scholar]
- Klijn CJ, Hankey GJ. American Stroke Association and European Stroke Initiative: management of acute ischaemic stroke: new guidelines from the American Stroke Association and European Stroke Initiative. Lancet Neurol. 2003;2:698–701. doi: 10.1016/s1474-4422(03)00558-1. [DOI] [PubMed] [Google Scholar]
- Knoller N, Levi L, Shoshan I, Reichenthal E, Razon N, Rappaport ZH, Biegon A. Dexanabinol (HU-211) in the treatment of severe closed head injury: a randomized, placebo-controlled, phase II clinical trial. Crit Care Med. 2002;30:548–554. doi: 10.1097/00003246-200203000-00009. [DOI] [PubMed] [Google Scholar]
- Kogan NM, Rabinowitz R, Levi P, Gibson D, Sandor P, Schlesinger M, Mechoulam R. Synthesis and antitumor activity of quinonoid derivatives of cannabinoids. J Med Chem. 2004;47:3800–3806. doi: 10.1021/jm040042o. [DOI] [PubMed] [Google Scholar]
- Kola B, Hubina E, Tucci SA, Kirkham TC, Garcia EA, Mitchell SE, Williams LM, Hawley SA, Hardie DG, Grossman AB, et al. Cannabinoids and ghrelin have both central and peripheral metabolic effects via AMP-activated protein kinase. J Biol Chem. 2005;280:25196–25201. doi: 10.1074/jbc.C500175200. [DOI] [PubMed] [Google Scholar]
- Koob GF. Drugs of abuse: anatomy, pharmacology and function of reward pathways. Trends Pharmacol Sci. 1992;13:177–184. doi: 10.1016/0165-6147(92)90060-j. [DOI] [PubMed] [Google Scholar]
- Koob GF. Drug addiction: the yin and yang of hedonic homeostasis. Neuron. 1996;16:893–896. doi: 10.1016/s0896-6273(00)80109-9. [DOI] [PubMed] [Google Scholar]
- Koob GF, Thatcher-Britton K. Stimulant and anxiogenic effects of corticotropin releasing factor. Prog Clin Biol Res. 1985;192:499–506. [PubMed] [Google Scholar]
- Korczyn AD. The ocular effects of cannabinoids. Gen Pharmacol. 1980;11:419–423. doi: 10.1016/0306-3623(80)90026-9. [DOI] [PubMed] [Google Scholar]
- Kosersky DS. Antihypertensive effects of Δ9-tetrahydrocannabinol. Arch Int Pharmacodyn Ther. 1978;233:76–81. [PubMed] [Google Scholar]
- Koutek B, Prestwich GD, Howlett AC, Chin SA, Salehani D, Akhavan N, Deutsch DG. Inhibitors of arachidonoyl ethanolamide hydrolysis. J Biol Chem. 1994;269:22937–22940. [PubMed] [Google Scholar]
- Kreutzer AC, Regehr WG. Retrograde inhibition of presynaptic calcium influx by endogenous cannabinoids at excitatory synapses onto Purkinje cells. Neuron. 2001;29:717–727. doi: 10.1016/s0896-6273(01)00246-x. [DOI] [PubMed] [Google Scholar]
- Kristensen P, Judge ME, Thim L, Ribel U, Christjansen KN, Wulff BS, Clausen JT, Jensen PB, Madsen OD, Vrang N, et al. Hypothalamic CART is a new anorectic peptide regulated by leptin. Nature (Lond) 1998;393:72–76. doi: 10.1038/29993. [DOI] [PubMed] [Google Scholar]
- Krylatov AV, Bernatskaia NA, Maslov LN, Pertwee RG, Mechoulam R, Stefano GB, Sharaevskii MA, Sal’nikova OM. Increase of the heart arrhythmogenic resistance and decrease of the myocardialnecrosis zone during activation of cannabinoid receptors. Ross Fiziol Zh Im I M Sechenova. 2002a;88:560–567. [PubMed] [Google Scholar]
- Krylatov AV, Maslov LN, Lasukova OV, Pertwee RG. Cannabinoid receptor antagonists SR141716 and SR144528 exhibit properties of partial agonists in experiments on isolated perfused rat heart. Bull Exp Biol Med. 2005;139:558–561. doi: 10.1007/s10517-005-0344-9. [DOI] [PubMed] [Google Scholar]
- Krylatov AV, Ugdyzhekova DS, Bernatskaya NA, Maslov LN, Mekhoulam R, Pertwee RG, Stephano GB. Activation of type II cannabinoid receptors improves myocardial tolerance to arrhythmogenic effects of coronary occlusion and reperfusion. Bull Exp Biol Med. 2001;131:523–525. doi: 10.1023/a:1012381914518. [DOI] [PubMed] [Google Scholar]
- Krylatov AV, Uzhachenko RV, Maslov LN, Bernatskaya NA, Makriyannis A, Mechoulam R, Pertwee RG, Sal’nikova OM, Stefano JB, Lishmanov Y. Endogenous cannabinoids improve myocardial resistance to arrhythmogenic effects of coronary occlusion and reperfusion: a possible mechanism. Bull Exp Biol Med. 2002b;133:122–124. doi: 10.1023/a:1015574100494. [DOI] [PubMed] [Google Scholar]
- Krylatov AV, Uzhachenko RV, Maslov LN, Ugdyzhekova DS, Bernatskaia NA, Pertwee R, Stefano GB, Makriyannis A. Anandamide and R-(+)-methanandamide prevent development of ischemic and reperfusion arrhythmia in rats by stimulation of CB2-receptors. Eksp Klin Farmakol. 2002c;65:6–9. [PubMed] [Google Scholar]
- Kuijten RR, Bunin GR, Nass CC, Meadows AT. Parental occupation and childhood astrocytoma: results of a case-control study. Cancer Res. 1992;52:782–786. [PubMed] [Google Scholar]
- Kulkarni-Narla A, Brown DR. Localization of CB1-cannabinoid receptor immunoreactivity in the porcine enteric nervous system. Cell Tissue Res. 2000;302:73–80. doi: 10.1007/s004410000261. [DOI] [PubMed] [Google Scholar]
- Kunos G, Bátkai S, Offertáler L, Mo F, Liu J, Karcher J, Harvey-White J. The quest for a vascular endothelial cannabinoids receptor. Chem Phys Lipids. 2002;121:45–56. doi: 10.1016/s0009-3084(02)00145-7. [DOI] [PubMed] [Google Scholar]
- Kunos G, Járai Z, Bátkai S, Goparaju SK, Ishac EJ, Liu J, Wang L, Wagner JA. Endocannabinoids as cardiovascular modulators. Chem Phys Lipids. 2000;108:159–168. doi: 10.1016/s0009-3084(00)00194-8. [DOI] [PubMed] [Google Scholar]
- Kunos G, Pacher P. Cannabinoids cool the intestine. Nat Med. 2004;10:678–679. doi: 10.1038/nm0704-678. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kurabayashi M, Takeyoshi I, Yoshinari D, Matsumoto K, Maruyama I, Morishita Y. 2-Arachidonoylglycerol increases in ischemia-reperfusion injury of the rat liver. J Investig Surg. 2005;18:25–31. doi: 10.1080/08941930590905189. [DOI] [PubMed] [Google Scholar]
- Kwolek G, Zakrzeska A, Schlicker E, Gothert M, Godlewski G, Malinowska B. Central and peripheral components of the pressor effect of anandamide in urethane-anaesthetized rats. Br J Pharmacol. 2005;145:567–575. doi: 10.1038/sj.bjp.0706195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lagneux C, Lamontagne D. Involvement of cannabinoids in the cardioprotection induced by lipopolysaccharide. Br J Pharmacol. 2001;132:793–796. doi: 10.1038/sj.bjp.0703902. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laine K, Jarvinen K, Jarvinen T. Topically administered CB2-receptor agonist, JWH-133, does not decrease intraocular pressure (IOP) in normotensive rabbits. Life Sci. 2003;72:837–842. doi: 10.1016/s0024-3205(02)02339-1. [DOI] [PubMed] [Google Scholar]
- Laine K, Jarvinen K, Mechoulam R, Breuer A, Jarvinen T. Comparison of the enzymatic stability and intraocular pressure effects of 2-arachidonylglycerol and noladin ether, a novel putative endocannabinoid. Investig Ophthalmol Vis Sci. 2002a;43:3216–3222. [PubMed] [Google Scholar]
- Laine K, Jarvinen K, Pate DW, Urtti A, Jarvinen T. Effect of the enzyme inhibitor, phenylmethylsulfonyl fluoride, on the IOP profiles of topical anandamides. Investig Ophthalmol Vis Sci. 2002b;43:393–397. [PubMed] [Google Scholar]
- Lake KD, Compton DR, Varga K, Martin BR, Kunos G. Cannabinoid-induced hypotension and bradycardia in rats mediated by CB1-like cannabinoid receptors. J Pharmacol Exp Ther. 1997a;281:30–1037. [PubMed] [Google Scholar]
- Lake KD, Martin BR, Kunos G, Varga K. Cardiovascular effects of anandamide in anesthetized and conscious normotensive and hypertensive rats. Hypertension. 1997b;29:1204–1210. doi: 10.1161/01.hyp.29.5.1204. [DOI] [PubMed] [Google Scholar]
- Lallemand F, de Witte P. Ethanol induces higher BEC in CB1 cannabinoid receptor knockout mice while decreasing ethanol preference. Alcohol Alcohol. 2004;40:54–62. doi: 10.1093/alcalc/agh115. [DOI] [PubMed] [Google Scholar]
- Lan R, Gatley J, Lu Q, Fan P, Fernando SR, Volkow ND, Pertwee R, Makriyannis A design and synthesis of the CB1 selective cannabinoid antagonist AM281: a potential human SPECT ligand. AAPS PharmSci. 1999a;1:E4. doi: 10.1208/ps010204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lan R, Liu Q, Fan P, Lin S, Fernando SR, McCallion D, Pertwee R, Makriyannis A. Structure-activity relationships of pyrazole derivatives as cannabinoid receptor antagonists. J Med Chem. 1999b;42:769–776. doi: 10.1021/jm980363y. [DOI] [PubMed] [Google Scholar]
- Landi M, Croci T, Rinaldi-Carmona M, Maffrand JP, Le Fur G, Manara L. Modulation of gastric emptying and gastrointestinal transit in rats through intestinal cannabinoid CB1 receptors. Eur J Pharmacol. 2002;450:77–83. doi: 10.1016/s0014-2999(02)02053-8. [DOI] [PubMed] [Google Scholar]
- Lang W, Qin C, Lin S, Khanolkar AD, Goutopoulos A, Fan P, Abouzid K, Meng Z, Biegel D, Makriyannis A. Substrate specificity and stereoselectivity of rat brain microsomal anandamide amidohydrolase. J Med Chem. 1999;42:896–902. doi: 10.1021/jm980461j. [DOI] [PubMed] [Google Scholar]
- La Rana G, Russo R, Campolongo P, Bortolato M, Mangieri RA, Cuomo V, Iacono A, Mattace Raso G, Meli R, Piomelli D, et al. Modulation of neuropathic and inflammatory pain by the endocannabinoid transport inhibitor AM404. J Pharmacol Exp Ther. 2006;317:1365–1371. doi: 10.1124/jpet.105.100792. [DOI] [PubMed] [Google Scholar]
- LaRoche SM, Helmers SL. The new antiepileptic drugs: scientific review. J Am Med Assoc. 2004;291:605–614. doi: 10.1001/jama.291.5.605. [DOI] [PubMed] [Google Scholar]
- Lastres-Becker I, Berrendero F, Lucas JJ, Martin-Aparicio E, Yamamoto A, Ramos JA, Fernandez-Ruiz JJ. Loss of mRNA levels, binding and activation of GTP-binding proteins for cannabinoid CB1 receptors in the basal ganglia of a transgenic model of Huntington’s disease. Brain Res. 2002a;929:236–242. doi: 10.1016/s0006-8993(01)03403-5. [DOI] [PubMed] [Google Scholar]
- Lastres-Becker I, Cebeira M, de Ceballos ML, Zeng BY, Jenner P, Ramos JA, Fernandez-Ruiz JJ. Increased cannabinoid CB1 receptor binding and activation of GTP-binding proteins in the basal ganglia of patients with Parkinson’s syndrome and of MPTP-treated marmosets. Eur J Neurosci. 2001a;14:1827–1832. doi: 10.1046/j.0953-816x.2001.01812.x. [DOI] [PubMed] [Google Scholar]
- Lastres-Becker I, de Miguel R, De Petrocellis L, Makriyannis A, Di Marzo V, Fernandez-Ruiz J. Compounds acting at the endocannabinoid and/or endovanilloid systems reduce hyperkinesia in a rat model of Huntington’s disease. J Neurochem. 2003a;84:1097–1109. doi: 10.1046/j.1471-4159.2003.01595.x. [DOI] [PubMed] [Google Scholar]
- Lastres-Becker I, De Miguel R, Fernandez-Ruiz JJ. The endocannabinoid system and Huntington’s disease. Curr Drug Targets CNS Neurol Disord. 2003b;2:335–347. doi: 10.2174/1568007033482751. [DOI] [PubMed] [Google Scholar]
- Lastres-Becker I, Fezza F, Cebeira M, Bisogno T, Ramos JA, Milone A, Fernandez-Ruiz J, Di Marzo V. Changes in endocannabinoid transmission in the basal ganglia in a rat model of Huntington’s disease. Neuroreport. 2001b;12:2125–2129. doi: 10.1097/00001756-200107200-00017. [DOI] [PubMed] [Google Scholar]
- Lastres-Becker I, Gomez M, De Miguel R, Ramos JA, Fernandez-Ruiz J. Loss of cannabinoid CB1 receptors in the basal ganglia in the late akinetic phase of rats with experimental Huntington’s disease. Neurotox Res. 2002b;4:601–608. doi: 10.1080/10298420290030514. [DOI] [PubMed] [Google Scholar]
- Lastres-Becker I, Molina-Holgado F, Ramos JA, Mechoulam R, Fernandez-Ruiz J. Cannabinoids provide neuroprotection against 6-hydroxydopamine toxicity in vivo and in vitro: relevance to Parkinson’s disease. Neurobiol Dis. 2005;19:96–107. doi: 10.1016/j.nbd.2004.11.009. [DOI] [PubMed] [Google Scholar]
- Lauckner JE, Hille B, Mackie K. The cannabinoid agonist WIN55,212-2 increases intracellular calcium via CB1 receptor coupling to Gq/11 G proteins. Proc Natl Acad Sci USA. 2005;102:19144–19149. doi: 10.1073/pnas.0509588102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lavie G, Teichner A, Shohami E, Ovadia H, Leker RR. Long term cerebroprotective effects of dexanabinol in a model of focal cerebral ischemia. Brain Res. 2001;901:195–201. doi: 10.1016/s0006-8993(01)02356-3. [DOI] [PubMed] [Google Scholar]
- Laviolette SR, Grace AA. The roles of cannabinoid and dopamine receptor systems in neural emotional learning circuits: implications for schizophrenia and addiction. Cell Mol Life Sci. 2006 doi: 10.1007/s00018-006-6027-5. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lavon I, Sheinin T, Meilin S, Biton E, Weksler A, Efroni G, Bar-Joseph A, Fink G, Avraham A. A novel synthetic cannabinoid derivative inhibits inflammatory liver damage via negative cytokine regulation. Mol Pharmacol. 2003;64:1334–1341. doi: 10.1124/mol.64.6.1334. [DOI] [PubMed] [Google Scholar]
- Lawrence DK, Gill EW. The effects of Δ1-tetrahydrocannabinol and other cannabinoids on spin-labeled liposomes and their relationship to mechanisms of general anesthesia. Mol Pharmacol. 1975;11:595–602. [PubMed] [Google Scholar]
- Le Foll B, Goldberg SR. Rimonabant, a CB1 antagonist, blocks nicotine-conditioned place preferences. Neuroreport. 2004;15:2139–2143. doi: 10.1097/00001756-200409150-00028. [DOI] [PubMed] [Google Scholar]
- Le Foll B, Goldberg DR. Cannabinoid CB1 receptor antagonists as promising new medications for drug dependence. J Pharmacol Exp Ther. 2005;312:875–883. doi: 10.1124/jpet.104.077974. [DOI] [PubMed] [Google Scholar]
- Ledent C, Valverde O, Cossu G, Petitet F, Aubert JF, Beslot F, Bohme GA, Imperato A, Pedrazzini T, Roques BP, et al. Unresponsiveness to cannabinoids and reduced addictive effects of opiates in CB1 receptor knockout mice. Science (Wash DC) 1999;283:401–404. doi: 10.1126/science.283.5400.401. [DOI] [PubMed] [Google Scholar]
- Lees G, Dougalis A. Differential effects of the sleep-inducing lipid oleamide and cannabinoids on the induction of long-term potentiation in the CA1 neurons of the rat hippocampus in vitro. Brain Res. 2004;997:1–14. doi: 10.1016/j.brainres.2003.10.019. [DOI] [PubMed] [Google Scholar]
- Leggett JD, Aspley S, Beckett SR, D’Antona AM, Kendall DA, Kendall DA. Oleamide is a selective endogenous agonist of rat and human CB1 cannabinoid receptors. Br J Pharmacol. 2004;141:253–262. doi: 10.1038/sj.bjp.0705607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leker RR, Gai N, Mechoulam R, Ovadia H. Drug-induced hypothermia reduces ischemic damage: effects of the cannabinoid HU-210. Stroke. 2003;34:2000–2006. doi: 10.1161/01.STR.0000079817.68944.1E. [DOI] [PubMed] [Google Scholar]
- Leker RR, Shohami E, Abramsky O, Ovadia H. Dexanabinol; a novel neuroprotective drug in experimental focal cerebral ischemia. J Neurol Sci. 1999;162:114–119. doi: 10.1016/s0022-510x(98)00301-3. [DOI] [PubMed] [Google Scholar]
- Lemberger L. Potential therapeutic usefulness of marijuana. Annu Rev Pharmacol Toxicol. 1980;20:151–172. doi: 10.1146/annurev.pa.20.040180.001055. [DOI] [PubMed] [Google Scholar]
- Lepicier P, Bouchard JF, Lagneux C, Lamontagne D. Endocannabinoids protect the rat isolated heart against ischaemia. Br J Pharmacol. 2003;139:805–815. doi: 10.1038/sj.bjp.0705313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lepore M, Vorel SR, Lowinson J, Gardner EL. Conditioned place preference induced by Δ9-tetrahydrocannabinol: comparison with cocaine, morphine and food reward. Life Sci. 1995;56:2073–2080. doi: 10.1016/0024-3205(95)00191-8. [DOI] [PubMed] [Google Scholar]
- Leroy S, Griffon N, Bourdel MC, Olie JP, Poirier MF, Krebs MO. Schizophrenia and the cannabinoid receptor type 1 (CB1): association study using a single-base polymorphism in coding exon 1. Am J Med Genet. 2001;105:749–752. doi: 10.1002/ajmg.10038. [DOI] [PubMed] [Google Scholar]
- Lesscher HMB, Hoogveld E, Burbach PH, van Ree JM, Gerrits MAFM. Endogenous cannabinoids are not involved in cocaine reinforcement and development of cocaine-induced behavioral sensitization. Eur Neuropsychopharmacol. 2005;15:31–37. doi: 10.1016/j.euroneuro.2004.04.003. [DOI] [PubMed] [Google Scholar]
- Leung D, Saghatelian A, Simon GM, Cravatt BF. Inactivation of N-acyl phosphatidylethanolamine phospholipase D reveals multiple mechanisms for the biosynthesis of endocannabinoids. Biochemistry. 2006;45:4720–4725. doi: 10.1021/bi060163l. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leweke FM, Gerth CW, Klosterkotter J. Cannabis-associated psychosis: current status of research. CNS Drugs. 2004;18:895–910. doi: 10.2165/00023210-200418130-00005. [DOI] [PubMed] [Google Scholar]
- Leweke FM, Giuffrida A, Wurster U, Emrich HM, Piomelli D. Elevated endogenous cannabinoids in schizophrenia. Neuroreport. 1999;10:1665–1669. doi: 10.1097/00001756-199906030-00008. [DOI] [PubMed] [Google Scholar]
- Lewis DA, Hashimoto T, Volk DW. Cortical inhibitory neurons and schizophrenia. Nat Rev Neurosci. 2005;6:312–324. doi: 10.1038/nrn1648. [DOI] [PubMed] [Google Scholar]
- Li J, Kaminski NE, Wang DH. Anandamide-induced depressor effect in spontaneously hypertensive rats: role of the vanilloid receptor. Hypertension. 2003;41:757–762. doi: 10.1161/01.HYP.0000051641.58674.F7. [DOI] [PubMed] [Google Scholar]
- Liang YC, Huang CC, Hsu KS. Therapeutic potential of cannabinoids in trigeminal neuralgia. Curr Drug Targets CNS Neurol Disord. 2004;3:507–514. doi: 10.2174/1568007043336833. [DOI] [PubMed] [Google Scholar]
- Libby P, Theroux P. Pathophysiology of coronary artery disease. Circulation. 2005;111:3481–3488. doi: 10.1161/CIRCULATIONAHA.105.537878. [DOI] [PubMed] [Google Scholar]
- Lichtman AH, Leung D, Shelton CC, Saghatelian A, Hardouin C, Boger DL, Cravatt BF. Reversible inhibitors of fatty acid amide hydrolase that promote analgesia: evidence for an unprecedented combination of potency and selectivity. J Pharmacol Exp Ther. 2004a;311:441–448. doi: 10.1124/jpet.104.069401. [DOI] [PubMed] [Google Scholar]
- Lichtman AH, Martin BR. Cannabinoid-induced antinociception is mediated by a spinal α2-noradrenergic mechanism. Brain Res. 1991a;559:309–314. doi: 10.1016/0006-8993(91)90017-p. [DOI] [PubMed] [Google Scholar]
- Lichtman AH, Martin BR. Spinal and supraspinal components of cannabinoid-induced antinociception. J Pharmacol Exp Ther. 1991b;258:517–523. [PubMed] [Google Scholar]
- Lichtman AH, Shelton CC, Advani T, Cravatt BF. Mice lacking fatty acid amide hydrolase exhibit a cannabinoid receptor-mediated phenotypic hypoalgesia. Pain. 2004b;109:319–327. doi: 10.1016/j.pain.2004.01.022. [DOI] [PubMed] [Google Scholar]
- Ligresti A, Cascio MG, Pryce G, Kulasegram S, Beletskaya I, De Petrocellis L, Saha B, Mahadevan A, Visintin C, Wiley JL, et al. New potent and selective inhibitors of anandamide reuptake with antispastic activity in a mouse model of multiple sclerosis. Br J Pharmacol. 2006a;147:83–91. doi: 10.1038/sj.bjp.0706418. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ligresti A, Schiano Moriello A, Starowicz K, Matias I, Pisanti S, De Petrocellis L, Laezza C, Portella G, Bifulco M, Di Marzo V. Anti-tumor activity of plant cannabinoids with emphasis on the effect of cannabidiol on human breast carcinoma. J Pharmacol Exp Ther. 2006b doi: 10.1124/jpet.106.105247. in press. [DOI] [PubMed] [Google Scholar]
- Lin S, Khanolkar AD, Fan P, Goutopoulos A, Qin C, Papahadjis D, Makriyannis A. Novel analogues of arachidonylethanolamide (anandamide): affinities for the CB1 and CB2 cannabinoid receptors and metabolic stability. J Med Chem. 1998;41:5353–5361. doi: 10.1021/jm970257g. [DOI] [PubMed] [Google Scholar]
- Linszen DH, Dingemans PM, Lenior ME. Cannabis abuse and the course of recent-onset schizophrenic disorders. Arch Gen Psychiatry. 1994;51:273–279. doi: 10.1001/archpsyc.1994.03950040017002. [DOI] [PubMed] [Google Scholar]
- Liu J, Bátkai S, Pacher P, Harvey-White J, Wagner JA, Cravatt BF, Gao B, Kunos G. LPS induces anandamide synthesis in macrophages via CD14/MAPK/PI3K/NF-κB independently of platelet activating factor. J Biol Chem. 2003a;278:45034–45039. doi: 10.1074/jbc.M306062200. [DOI] [PubMed] [Google Scholar]
- Liu J, Gao B, Mirshahi F, Sanyal AJ, Khanolkar AD, Makriyannis A, Kunos G. Functional CB1 cannabinoid receptors in human vascular endothelial cells. Biochem J. 2000;346:835–840. [PMC free article] [PubMed] [Google Scholar]
- Liu J, Li H, Burstein SH, Zurier RB, Chen JD. Activation and binding of peroxisome proliferator-activated receptor gamma by synthetic cannabinoid ajulemic acid. Mol Pharmacol. 2003b;63:983–992. doi: 10.1124/mol.63.5.983. [DOI] [PubMed] [Google Scholar]
- Liu J, Wang L, Harvey-White J, Osei-Hyiaman D, Razdan RK, Zhou Z, Chen A, Huang B, Kim HY, Kunos G. Novel biosynthetic pathway for anandamide. Proc Natl Acad Sci USA. 2006;103:13345–13350. doi: 10.1073/pnas.0601832103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu JH, Dacus AC. Central nervous system and peripheral mechanisms in ocular hypotensive effect of cannabinoids. Arch Ophthalmol. 1987;105:245–248. doi: 10.1001/archopht.1987.01060020099037. [DOI] [PubMed] [Google Scholar]
- Liu YL, Connoley IP, Wilson CA, Stock MJ. Effects of the cannabinoid CB1 receptor antagonist SR141716 on oxygen consumption and soleus muscle glucose uptake in Lepob/Lepob mice. Int J Obes Relat Metab Disord. 2005;29:183–187. doi: 10.1038/sj.ijo.0802847. [DOI] [PubMed] [Google Scholar]
- Llewellyn CD, Linklater K, Bell J, Johnson NW, Warnakulasuriya S. An analysis of risk factors for oral cancer in young people: a case-control study. Oral Oncol. 2004;40:304–313. doi: 10.1016/j.oraloncology.2003.08.015. [DOI] [PubMed] [Google Scholar]
- Loftus EV., Jr Clinical epidemiology of inflammatory bowel disease: incidence, prevalence, and environmental influences. Gastroenterology. 2004;126:1504–1517. doi: 10.1053/j.gastro.2004.01.063. [DOI] [PubMed] [Google Scholar]
- Lograno MD, Romano MR. Cannabinoid agonists induce contractile responses through Gi/o-dependent activation of phospholipase C in the bovine ciliary muscle. Eur J Pharmacol. 2004;494:55–62. doi: 10.1016/j.ejphar.2004.04.039. [DOI] [PubMed] [Google Scholar]
- Lombard C, Nagarkatti M, Nagarkatti PS. Targeting cannabinoid receptors to treat leukemia: role of cross-talk between extrinsic and intrinsic pathways in Δ9-tetrahydrocannabinol (THC)-induced apoptosis of Jurkat cells. Leuk Res. 2005;29:915–922. doi: 10.1016/j.leukres.2005.01.014. [DOI] [PubMed] [Google Scholar]
- Long LE, Malone DT, Taylor DA. Cannabidiol reverses MK-801-induced disruption of prepulse inhibition in mice. Neuropsychopharmacology. 2006;31:795–803. doi: 10.1038/sj.npp.1300838. [DOI] [PubMed] [Google Scholar]
- Lopez-Moreno JA, Gonzalez-Cuevas G, Rodriguez de Fonseca F, Navarro M. Long-lasting increase of alcohol relapse by the cannabinoid receptor agonist WIN 55,212-2 during alcohol deprivation. J Neurosci. 2005;24:8245–8252. doi: 10.1523/JNEUROSCI.2179-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lopez-Rodriguez ML, Viso A, Ortega-Gutierrez S, Fowler CJ, Tiger G, de Lago E, Fernandez-Ruiz J, Ramos JA. Design, synthesis and biological evaluation of new endocannabinoid transporter inhibitors. Eur J Med Chem. 2003;38:403–412. doi: 10.1016/s0223-5234(03)00045-x. [DOI] [PubMed] [Google Scholar]
- Lopez-Rodriguez ML, Viso A, Ortega-Gutierrez S, Lastres-Becker I, Gonzalez S, Fernandez-Ruiz J, Ramos JA. Design, synthesis and biological evaluation of novel arachidonic acid derivatives as highly potent and selective endocannabinoid transporter inhibitors. J Med Chem. 2001;44:4505–4508. doi: 10.1021/jm015545y. [DOI] [PubMed] [Google Scholar]
- Louw DF, Yang FW, Sutherland GR. The effect of Δ-9-tetrahydrocannab-inol on forebrain ischemia in rat. Brain Res. 2000;857:183–187. doi: 10.1016/s0006-8993(99)02422-1. [DOI] [PubMed] [Google Scholar]
- Lu Q, Straiker A, Lu Q, Maguire G. Expression of CB2 cannabinoid receptor mRNA in adult rat retina. Vis Neurosci. 2000;17:91–95. doi: 10.1017/s0952523800171093. [DOI] [PubMed] [Google Scholar]
- Lukas SE, Sholar M, Kouri E, Fukuzako H, Mendelson JH. Marihuana smoking increases plasma cocaine levels and subjective reports of euphoria in male volunteers. Pharmacol Biochem Behav. 1994;48:715–721. doi: 10.1016/0091-3057(94)90338-7. [DOI] [PubMed] [Google Scholar]
- Lumsden AB, Henderson JM, Kutner MH. Endotoxin levels measured by a chromatographic assay in portal, hepatic and peripheral blood in patients with cirrhosis. Hepatology. 1988;8:232–236. doi: 10.1002/hep.1840080207. [DOI] [PubMed] [Google Scholar]
- Lupica CR, Riegel AC, Hoffman AF. Marijuana and cannabinoid regulation of brain reward circuits. Br J Pharmacol. 2004;143:227–234. doi: 10.1038/sj.bjp.0705931. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lutz B. Molecular biology of cannabinoid receptors. Prostaglandins Leukotrienes Essent Fatty Acids. 2002;66:123–142. doi: 10.1054/plef.2001.0342. [DOI] [PubMed] [Google Scholar]
- Lutz B. On-demand activation of the endocannabinoid system in the control of neuronal excitability and epileptiform seizures. Biochem Pharmacol. 2004;68:1691–1698. doi: 10.1016/j.bcp.2004.07.007. [DOI] [PubMed] [Google Scholar]
- Lyman WD, Sonett JR, Brosnan CF, Elkin R, Bornstein MB. Δ-9-Tetrahydrocannabinol: a novel treatment for experimental autoimmune encepha-lomyelitis. J Neuroimmunol. 1989;23:73–81. doi: 10.1016/0165-5728(89)90075-1. [DOI] [PubMed] [Google Scholar]
- Maas AI, Marmarou A, Murray GD, Steyerberg EW. Clinical trials in traumatic brain injury: current problems and future solutions. Acta Neurochir Suppl. 2004;89:113–118. doi: 10.1007/978-3-7091-0603-7_16. [DOI] [PubMed] [Google Scholar]
- Maas AI, Murray G, Henney H, 3rd, Kassem N, Legrand V, Mangelus M, Muizelaar JP, Stocchetti N, Knoller N Pharmos TBI Investigators. Efficacy and safety of dexanabinol in severe traumatic brain injury: results of a phase III randomised, placebo-controlled, clinical trial. Lancet Neurol. 2006;5:38–45. doi: 10.1016/S1474-4422(05)70253-2. [DOI] [PubMed] [Google Scholar]
- Maccarrone M, Bari M, Battista N, Finazzi-Agro A. Endocannabinoid degradation, endotoxic shock and inflammation. Curr Drug Targets Inflamm Allergy. 2002a;1:53–63. doi: 10.2174/1568010023344878. [DOI] [PubMed] [Google Scholar]
- Maccarrone M, Bari M, Lorenzon T, Bisogno T, Di Marzo V, Finazzi-Agro A. Anandamide uptake by human endothelial cells and is regulation by nitric oxide. J Biol Chem. 2000a;275:13484–13492. doi: 10.1074/jbc.275.18.13484. [DOI] [PubMed] [Google Scholar]
- Maccarrone M, Bisogno T, Valensise H, Lazzarin N, Fezza F, Manna C, Di Marzo V, Finazzi-Agro A. Low fatty acid amide hydrolase and high anandamide levels are associated with failure to achieve an ongoing pregnancy after IVF and embryo transfer. Mol Hum Reprod. 2002b;8:188–195. doi: 10.1093/molehr/8.2.188. [DOI] [PubMed] [Google Scholar]
- Maccarrone M, De Petrocellis L, Bari M, Fezza F, Salvati S, Di Marzo V, Finazzi-Agro A. Lipopolysaccharide downregulates fatty acid amide hydrolase expression and increases anandamide levels in human peripheral lymphocytes. Arch Biochem Biophys. 2001;393:321–328. doi: 10.1006/abbi.2001.2500. [DOI] [PubMed] [Google Scholar]
- Maccarrone M, Gubellini P, Bari M, Picconi B, Battista N, Centonze D, Bernardi G, Finazzi-Agro A, Calabresi P. Levodopa treatment reverses endocannabinoid system abnormalities in experimental parkinsonism. J Neurochem. 2003;85:1018–1025. doi: 10.1046/j.1471-4159.2003.01759.x. [DOI] [PubMed] [Google Scholar]
- Maccarrone M, Lorenzon T, Bari M, Melino G, Finazzi-Agro A. Anandamide induces apoptosis in human cells via vanilloid receptors: evidence for a protective role of cannabinoid receptors. J Biol Chem. 2000b;275:31938–31945. doi: 10.1074/jbc.M005722200. [DOI] [PubMed] [Google Scholar]
- Maccarrone M, Valensise H, Bari M, Lazzarin N, Romanini C, Finazzi-Agro A. Relation between decreased anandamide hydrolase concentrations in human lymphocytes and miscarriage. Lancet. 2000c;355:1326–1329. doi: 10.1016/S0140-6736(00)02115-2. [DOI] [PubMed] [Google Scholar]
- Mackie K. Cannabinoid receptor homo- and heterodimerization. Life Sci. 2005;77:1667–1673. doi: 10.1016/j.lfs.2005.05.011. [DOI] [PubMed] [Google Scholar]
- Mackie K. Cannabinoid receptors as therapeutic targets. Annu Rev Pharmacol Toxicol. 2006;46:101–122. doi: 10.1146/annurev.pharmtox.46.120604.141254. [DOI] [PubMed] [Google Scholar]
- Mackie K, Devane WA, Hille B. Anandamide, an endogenous cannabinoid, inhibits calcium currents as a partial agonist in N18 neuroblastoma cells. Mol Pharmacol. 1993;44:498–503. [PubMed] [Google Scholar]
- Mackie K, Hille B. Cannabinoids inhibit N-type calcium channels in neuroblastomaglioma cells. Proc Natl Acad Sci USA. 1992;89:3825–3829. doi: 10.1073/pnas.89.9.3825. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mackie K, Lai Y, Westenbroek R, Mitchell R. Cannabinoids activate an inwardly rectifying potassium conductance and inhibit Q-type calcium currents in AtT20 cells transfected with rat brain cannabinoid receptor. J Neurosci. 1995;15:6552–6561. doi: 10.1523/JNEUROSCI.15-10-06552.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- MacNaughton WK, Van Sickle MD, Keenan CM, Cushing K, Mackie K, Sharkey KA. Distribution and function of the cannabinoid-1 receptor in the modulation of ion transport in the guinea pig ileum: relationship to capsaicin-sensitive nerves. Am J Physiol. 2004;286:G863–G871. doi: 10.1152/ajpgi.00482.2003. [DOI] [PubMed] [Google Scholar]
- Mailleux P, Vanderhaeghen JJ. Localization of cannabinoid receptor in the human developing and adult basal ganglia: higher levels in the striatonigral neurons. Neurosci Lett. 1992;148:173–176. doi: 10.1016/0304-3940(92)90832-r. [DOI] [PubMed] [Google Scholar]
- Mailleux P, Vanderhaeghen JJ. Dopaminergic regulation of cannabinoid receptor mRNA levels in the rat caudate-putamen: an in situ hybridization study. J Neurochem. 1993;61:1705–1712. doi: 10.1111/j.1471-4159.1993.tb09807.x. [DOI] [PubMed] [Google Scholar]
- Maingret F, Patel AJ, Lazdunski M, Honore E. The endocannabinoid anandamide is a direct and selective blocker of the background K+ channel TASK-1. EMBO (Eur Mol Biol Organ) J. 2001;20:47–54. doi: 10.1093/emboj/20.1.47. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maione S, Bisogno T, de Novellis V, Palazzo E, Christino L, Valenti M, Petrosino S, Guglielmotti V, Rossi F, Di Marzo V. Elevation of endocannabinoid levels in the ventrolateral periaqueductal grey through inhibition of fatty acid amide hydrolase affects descending nociceptive pathways via both cannabinoid receptor type 1 and transient receptor potential vanilloid type-1 receptors. J Pharmacol Exp Ther. 2006;316:969–982. doi: 10.1124/jpet.105.093286. [DOI] [PubMed] [Google Scholar]
- Makara JK, Mor M, Fegley D, Szabo SI, Kathuria S, Astarita G, Duranti A, Tontini A, Tarzia G, Rivara S, Freund TF, Piomelli D. Selective inhibition of 2-AG hydrolysis enhances endocannabinoid signaling in hippocampus. Nat Neurosci. 2005;8:1139–1141. doi: 10.1038/nn1521. [DOI] [PubMed] [Google Scholar]
- Malan TP, Jr, Ibrahim MM, Deng H, Liu Q, Mata HP, Vanderah T, Porreca F, Makriyannis A. CB2 cannabinoid receptor-mediated peripheral antinociception. Pain. 2001;93:239–245. doi: 10.1016/S0304-3959(01)00321-9. [DOI] [PubMed] [Google Scholar]
- Malcher-Lopes R, Di S, Marcheselli VS, Weng F-J, Stuart CT, Bazan NG, Tasker JG. Opposing crosstalk between leptin and glucocorticoids rapidly modulates synaptic excitation via endocannabinoid release. J Neurosci. 2006;26:6643–6650. doi: 10.1523/JNEUROSCI.5126-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maldonado R, Valverde O, Berrendero F. Involvement of the endocannabinoid system in drug addiction. Trends Pharmacol Sci. 2006;29:225–232. doi: 10.1016/j.tins.2006.01.008. [DOI] [PubMed] [Google Scholar]
- Malfait AM, Gallily R, Sumariwalla PF, Malik AS, Andreakos E, Mechoulam R, Feldmann M. The nonpsychoactive cannabis constituent cannabidiol is an oral anti-arthritic therapeutic in murine collagen-induced arthritis. Proc Natl Acad Sci USA. 2000;97:9561–9566. doi: 10.1073/pnas.160105897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Malinowska B, Godlewski G, Boucher B, Schlicker E. Cannabinoid CB1 receptor-mediated inhibition of the neurogenic vasopressor response in the pithed rat. Naunyn-Schmiedeberg’s Arch Pharmacol. 1997;356:197–202. doi: 10.1007/pl00005041. [DOI] [PubMed] [Google Scholar]
- Malinowska B, Kwolek G, Gothert M. Anandamide and methanandamide induce both vanilloid VR1- and cannabinoids CB1 receptor-mediated changes in heart rate and blood pressure in anaesthetized rats. Naunyn-Schmiedeberg’s Arch Pharmacol. 2001;364:562–569. doi: 10.1007/s00210-001-0498-6. [DOI] [PubMed] [Google Scholar]
- Malone DT, Taylor DA. Involvement of somatodendritic 5-HT1A receptors in Δ9-tetrahydrocannabinol-induced hypothermia in the rat. Pharmacol Biochem Behav. 2001;69:595–601. doi: 10.1016/s0091-3057(01)00567-6. [DOI] [PubMed] [Google Scholar]
- Manara L, Croci T, Guagnini F, Rinaldi-Carmona M, Maffrand JP, Le Fur G, Mukenge S, Ferla G. Functional assessment of neuronal cannabinoid receptors in the muscular layers of human ileum and colon. Dig Liver Dis. 2002;34:262–269. doi: 10.1016/s1590-8658(02)80146-3. [DOI] [PubMed] [Google Scholar]
- Mancinelli R, Fabrizi A, Del Monaco S, Azzena GB, Vargiu R, Colombo GC, Gessa GL. Inhibition of peristaltic activity by cannabinoids in the isolated distal colon of mouse. Life Sci. 2001;69:101–111. doi: 10.1016/s0024-3205(01)01110-9. [DOI] [PubMed] [Google Scholar]
- Maneuf YP, Crossman AR, Brotchie JM. The cannabinoid receptor agonist WIN 55,212-2 reduces D2, but not D1, dopamine receptor-mediated alleviation of akinesia in the reserpine-treated rat model of Parkinson’s disease. Exp Neurol. 1997;148:265–270. doi: 10.1006/exnr.1997.6645. [DOI] [PubMed] [Google Scholar]
- Mang CF, Erbelding D, Kilbinger H. Differential effects of anandamide on acetylcholine release in the guinea-pig ileum mediated via vanilloid and non-CB1 cannabinoid receptors. Br J Pharmacol. 2001;134:161–167. doi: 10.1038/sj.bjp.0704220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Manzanares J, Corchero J, Fuentes JA. Opioid and cannabinoid receptor-mediated regulation of the increase in adrenocorticotropin hormone and corticosterone plasma concentrations induced by central administration of Δ9-tetrahydrocannabinol in rats. Brain Res. 1999a;839:173–179. doi: 10.1016/s0006-8993(99)01756-4. [DOI] [PubMed] [Google Scholar]
- Manzanares J, Corchero J, Romero J, Fernandez-Ruiz JJ, Ramos JA, Fuentes JA. Pharmacological and biochemical interactions between opioids and cannabinoids. Trends Pharmacol Sci. 1999b;20:287–294. doi: 10.1016/s0165-6147(99)01339-5. [DOI] [PubMed] [Google Scholar]
- Manzanares J, Uriguen L, Rubio G, Palomo T. Role of endocannabinoid system in mental diseases. Neurotox Res. 2004;6:213–224. doi: 10.1007/BF03033223. [DOI] [PubMed] [Google Scholar]
- Marco EM, Perez-Alvarez L, Borcel E, Rubio M, Guaza C, Ambrosio E, File SE, Viveros MP. Involvement of 5-HT1A receptors in behavioural effects of the cannabinoid receptor agonist CP 55,940 in male rats. Behav Pharmacol. 2004;15:21–27. doi: 10.1097/00008877-200402000-00003. [DOI] [PubMed] [Google Scholar]
- Marin S, Marco E, Biscaia M, Fernandez B, Rubio M, Guaza C, Schmidhammer H, Viveros MP. Involvement of the κ-opioid receptor in the anxiogenic-like effect of CP 55,940 in male rats. Pharmacol Biochem Behav. 2003;74:649–656. doi: 10.1016/s0091-3057(02)01041-9. [DOI] [PubMed] [Google Scholar]
- Markus FW. Cannabivarin and tetrahydrocannabivarin, two constituents of hashish. Nature (Lond) 1971;232:579–580. doi: 10.1038/232579a0. [DOI] [PubMed] [Google Scholar]
- Marsicano G, Lutz B. Expression of the cannabinoid receptor CB1 in distinct neuronal subpopulations in the adult mouse forebrain. Eur J Neurosci. 1999;11:4213–4225. doi: 10.1046/j.1460-9568.1999.00847.x. [DOI] [PubMed] [Google Scholar]
- Marsicano G, Goodenough S, Monory K, Hermann H, Eder M, Cannich A, Azad SC, Cascio MG, Gutiérrez SO, van der Stelt M, et al. CB1 cannabinoid receptors and on-demand defense against excitotoxicity. Science (Wash DC) 2003;302:84–88. doi: 10.1126/science.1088208. [DOI] [PubMed] [Google Scholar]
- Marsicano G, Moosmann B, Hermann H, Lutz B, Behl C. Neuroprotective properties of cannabinoids against oxidative stress: role of the cannabinoid receptor CB1. J Neurochem. 2002a;80:448–456. doi: 10.1046/j.0022-3042.2001.00716.x. [DOI] [PubMed] [Google Scholar]
- Marsicano G, Wotjak CT, Azad SC, Bisogno T, Rammes G, Cascio MG, Hermann H, Tang J, Hofmann C, Zieglgansberger W, et al. The endogenous cannabinoid system controls extinction of aversive memories. Nature (Lond) 2002b;418:530–534. doi: 10.1038/nature00839. [DOI] [PubMed] [Google Scholar]
- Martellotta MC, Cossu G, Fattore L, Gessa GL, Fratta G. Self-administration of the cannabinoid receptor agonist WIN 55,212-2 in drug-naïve rats. Neuroscience. 1998;85:327–330. doi: 10.1016/s0306-4522(98)00052-9. [DOI] [PubMed] [Google Scholar]
- Martin AR, Consroe P, Kane VV, Shah V, Singh V, Lander N, Mechoulam R, Srebnik M. Structure-anticonvulsant activity relationships of cannabidiol analogs. NIDA Res Monogr. 1987;79:48–58. [PubMed] [Google Scholar]
- Martin BR, Wiley JL. Mechanism of action of cannabinoids: how it may lead to treatment of cachexia, emesis, and pain. J Support Oncol. 2004;2:305–334. 314–316. [PubMed] [Google Scholar]
- Martin M, Ledent C, Parmentier M, Maldonado R, Valverde O. Cocaine, but not morphine, induces conditioned place preference and sensitization to locomotor responses in CB1 knockout mice. Eur J Neurosci. 2000;12:4038–4046. doi: 10.1046/j.1460-9568.2000.00287.x. [DOI] [PubMed] [Google Scholar]
- Martin M, Ledent C, Parmentier M, Maldonado R, Valverde O. Involvement of CB1 cannabinoid receptors in emotional behaviour. Psychopharmacology. 2002;159:379–387. doi: 10.1007/s00213-001-0946-5. [DOI] [PubMed] [Google Scholar]
- Martin WJ, Coffin PO, Attias E, Balinsky M, Tsou K, Walker JM. Anatomical basis for cannabinoid-induced antinociception as revealed by intracerebral microinjections. Brain Res. 1999a;822:237–242. doi: 10.1016/s0006-8993(98)01368-7. [DOI] [PubMed] [Google Scholar]
- Martin WJ, Hohmann AG, Walker JM. Suppression of noxious stimulus-evoked activity in the ventral posterolateral nucleus of the thalamus by the cannabinoid WIN 55,212-2:correlation between electrophysiological and antinociceptive effects. J Neurosci. 1996;16:6601–6611. doi: 10.1523/JNEUROSCI.16-20-06601.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin WJ, Lai NK, Patrick SL, Tsou K, Walker JM. Antinociceptive actions of WIN 55,212-2 following intraventricular administration in rats. Brain Res. 1993;629:300–304. doi: 10.1016/0006-8993(93)91334-o. [DOI] [PubMed] [Google Scholar]
- Martin WJ, Loo CM, Basbaum AI. Spinal cannabinoids are anti-allodynic in rats with persistent inflammation. Pain. 1999b;82:199–205. doi: 10.1016/S0304-3959(99)00045-7. [DOI] [PubMed] [Google Scholar]
- Martin WJ, Patrick SL, Coffin PO, Tsou K, Walker JM. An examination of the central sites of action of cannabinoid-induced antinociception in the rat. Life Sci. 1995;56:2103–2110. doi: 10.1016/0024-3205(95)00195-c. [DOI] [PubMed] [Google Scholar]
- Martin WJ, Tsou K, Walker JM. Cannabinoid receptor-mediated inhibition of the rat tail-flick reflex after microinjection into the rostral ventromedial medulla. Neurosci Let. 1998;232:33–36. doi: 10.1016/s0304-3940(98)00044-5. [DOI] [PubMed] [Google Scholar]
- Martin-Calderon JL, Munoz RM, Villanua MA, del Arco I, Moreno JL, de Fonseca FR, Navarro M. Characterization of the acute endocrine actions of (−)-11-hydroxy-Δ8-tetrahydrocannabinol-dimethylheptyl (HU-210), a potent synthetic cannabinoid in rats. Eur J Pharmacol. 1998;344:77–86. doi: 10.1016/s0014-2999(97)01560-4. [DOI] [PubMed] [Google Scholar]
- Martinez-Arevalo MJ, Calcedo-Ordonez A, Varo-Prieto JR. Cannabis consumption as a prognostic factor in schizophrenia. Br J Psychiatry. 1994;164:679–681. doi: 10.1192/bjp.164.5.679. [DOI] [PubMed] [Google Scholar]
- Martinez-Vargas M, Murillo-Rodriguez E, Gonzalez-Rivera R, Landa A, Mendez-Diaz M, Prospro-Garcia O, Navarro L. Sleep modulates cannabinoid receptor 1 expression in the pons of rats. Neuroscience. 2003;117:197–201. doi: 10.1016/s0306-4522(02)00820-5. [DOI] [PubMed] [Google Scholar]
- Martyn CN, Illis LS, Thom J. Nabilone in the treatment of multiple sclerosis. Lancet. 1995;345:579. doi: 10.1016/s0140-6736(95)90485-9. [DOI] [PubMed] [Google Scholar]
- Mascia MS, Obinu MC, Ledent C, Parmentier M, Böhme GA, Imperato A, Fratta W. lack of morphine-induced dopamine release in the nucleus accumbens of cannabinoid CB1 receptor knockout mice. Eur J Pharmacol. 1999;383:R1–R2. doi: 10.1016/s0014-2999(99)00656-1. [DOI] [PubMed] [Google Scholar]
- Mascolo N, Izzo AA, Ligresti A, Costagliola A, Pinto L, Cascio MG, Maffia P, Cecio A, Capasso F, Di Marzo V. The endocannabinoid system and the molecular basis of paralytic ileus in mice. FASEB J. 2002;16:1973–1975. doi: 10.1096/fj.02-0338fje. [DOI] [PubMed] [Google Scholar]
- Maslov LN, Lasukova OV, Krylatov AV, Uzhachenko RV, Pertwee R. Selective cannabinoid receptor agonist HU-210 decreases pump function of isolated perfused heart: role of cAMP and cGMP. Bull Exp Biol Med. 2004;138:550–553. doi: 10.1007/s10517-005-0123-7. [DOI] [PubMed] [Google Scholar]
- Mas-Nieto M, Pommier B, Tzavara ET, Caneparo A, Da Nascimento S, Le Fur G, Roques BP, Noble F. Reduction of opioid dependence by the CB1 antagonist SR141716A in mice: evaluation of the interest in pharmacotherapy of opioid addiction. Br J Pharmacol. 2001;132:1809–1816. doi: 10.1038/sj.bjp.0703990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Massa F, Marsicano G, Hermann H, Cannich A, Monory K, Cravatt BF, Ferri GL, Sibaev A, Storr M, Lutz B. The endogenous cannabinoid system protects against colonic inflammation. J Clin Investig. 2004;113:1202–1209. doi: 10.1172/JCI19465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Massa F, Storr M, Lutz B. The endocannabinoid system in the physiology and pathophysiology of the gastrointestinal tract. J Mol Med. 2005;83:944–954. doi: 10.1007/s00109-005-0698-5. [DOI] [PubMed] [Google Scholar]
- Massi P, Vaccani A, Ceruti S, Colombo A, Abbracchio MP, Parolaro D. Antitumor effects of cannabidiol, a nonpsychoactive cannabinoid, on human glioma cell lines. J Pharmacol Exp Ther. 2004;308:838–845. doi: 10.1124/jpet.103.061002. [DOI] [PubMed] [Google Scholar]
- Mathison R, Ho W, Pittman QJ, Davison JS, Sharkey KA. Effects of cannabinoid receptor-2 activation on accelerated gastrointestinal transit in lipopolysaccharide-treated rats. Br J Pharmacol. 2004;142:1247–1254. doi: 10.1038/sj.bjp.0705889. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matias I, Gonthier M-P, Monteleone P, Di Marzo V. Peripheral upregulation of the endocannabionoid system in obesity. Proceedings of the 2005 Symposium on the Cannabinoids; Burlington, VT: International Cannabinoid Research Society; 2005. p. 58. [Google Scholar]
- Matias I, Gonthier M-P, Orlando P, Martiadis V, De Petrocellis L, Cervino C, Petrosino S, Hoareau L, Festy F, Pasquali R, et al. Regulation, function and dysregulation of endocannabionoids in obesity and hyperglycemis. J Clin Endocr Metab. 2006 doi: 10.1210/jc.2005-2679. in press. [DOI] [PubMed] [Google Scholar]
- Mato S, Chevaleyre V, Robbe D, Pazos A, Castillo PE, Manzoni OJ. A single in-vivo exposure to Δ9THC blocks endocannabinoid-mediated synaptic plasticity. Nat Neurosci. 2004;7:585–586. doi: 10.1038/nn1251. [DOI] [PubMed] [Google Scholar]
- Matsuda K, Mikami Y, Takeda K, Fukuyama S, Egawa S, Sunamura M, Maruyama I, Matsuno S. The cannabinoid 1 receptor antagonist, AM251, prolongs the survival of rats with severe acute pancreatitis. Tohoku J Exp Med. 2005;207:99–107. doi: 10.1620/tjem.207.99. [DOI] [PubMed] [Google Scholar]
- Matsuda LA, Lolait SJ, Brownstein MJ, Young CA, Bonner TI. Structure of a cannabinoid receptor and functional expression of the cloned cDNA. Nature (Lond) 1990;346:561–564. doi: 10.1038/346561a0. [DOI] [PubMed] [Google Scholar]
- Mauler F, Mittendorf J, Horvath E, De Vry J. Characterization of the diarylether sulfonylester (−)-(R)-3-(2-hydroxymethylindanyl-4-oxy)phenyl-4,4,4-trifluoro-1-sulfonate (BAY 38-7271) as a potent cannabinoid receptor agonist with neuroprotective properties. J Pharmacol Exp Ther. 2002;302:359–368. doi: 10.1124/jpet.302.1.359. [DOI] [PubMed] [Google Scholar]
- Mazzola C, Micale V, Drago F. Amnesia induced by β-amyloid fragments is counteracted by cannabinoid CB1 receptor blockade. Eur J Pharmacol. 2003;477:219–225. doi: 10.1016/j.ejphar.2003.08.026. [DOI] [PubMed] [Google Scholar]
- Mbvundula EC, Bunning RA, Rainsford KD. Effects of cannabinoids on nitric oxide production by chondrocytes and proteoglycan degradation in cartilage. Biochem Pharmacol. 2005;69:635–640. doi: 10.1016/j.bcp.2004.11.018. [DOI] [PubMed] [Google Scholar]
- Mbvundula EC, Bunning RA, Rainsford KD. Arthritis and cannabinoids: HU-210 and Win-55,212-2 prevent IL-1α-induced matrix degradation in bovine articular chondrocytes in-vitro. J Pharm Pharmacol. 2006;58:351–358. doi: 10.1211/jpp.58.3.0009. [DOI] [PubMed] [Google Scholar]
- McAllister SD, Rizvi G, Anavi-Goffer S, Hurst DP, Barnett-Norris J, Lynch DL, Reggio PH, Abood ME. An aromatic microdomain at the cannabinoid CB1 receptor constitutes an agonist/inverse agonist binding region. J Med Chem. 2003;46:5139–5152. doi: 10.1021/jm0302647. [DOI] [PubMed] [Google Scholar]
- McCallum RW, Soykan I, Sridhar KR, Ricci DA, Lange RC, Plankey MW. Δ-9-Tetrahydrocannabinol delays the gastric emptying of solid food in humans: a double-blind, randomized study. Aliment Pharmacol Ther. 1999;13:77–80. doi: 10.1046/j.1365-2036.1999.00441.x. [DOI] [PubMed] [Google Scholar]
- McCaw EA, Hu H, Gomez GT, Hebb AL, Kelly ME, Denovan-Wright EM. Structure, expression and regulation of the cannabinoid receptor gene (CB1) in Huntington’s disease transgenic mice. Eur J Biochem. 2004;271:4909–4920. doi: 10.1111/j.1432-1033.2004.04460.x. [DOI] [PubMed] [Google Scholar]
- McCormick DA, Contreras D. On the cellular and network bases of epileptic seizures. Annu Rev Physiol. 2001;63:815–846. doi: 10.1146/annurev.physiol.63.1.815. [DOI] [PubMed] [Google Scholar]
- McFarland MJ, Barker EL. Anandamide transport. Pharmacol Ther. 2004;104:117–135. doi: 10.1016/j.pharmthera.2004.07.008. [DOI] [PubMed] [Google Scholar]
- McGarry JD. What if Minkowski had been ageusic? An alternative angle on diabetes. Science (Wash DC) 1992;258:766–770. doi: 10.1126/science.1439783. [DOI] [PubMed] [Google Scholar]
- McGregor IS, Dastur FN, McLellan RA, Brown RE. Cannabinoid modulation of rat pup ultrasonic vocalizations. Eur J Pharmacol. 1996a;313:43–49. doi: 10.1016/0014-2999(96)00511-0. [DOI] [PubMed] [Google Scholar]
- McGregor IS, Issakidis CN, Prior G. Aversive effects of the synthetic cannabinoid CP 55,940 in rats. Pharmacol Biochem Behav. 1996b;53:657–664. doi: 10.1016/0091-3057(95)02066-7. [DOI] [PubMed] [Google Scholar]
- McGuire PK, Jones P, Harvey I, Bebbington P, Toone B, Lewis S, Murray RM. Cannabis and acute psychosis. Schizophr Res. 1994;13:161–167. doi: 10.1016/0920-9964(94)90097-3. [DOI] [PubMed] [Google Scholar]
- McKallip RJ, Lombard C, Fisher M, Martin BR, Ryu S, Grant S, Nagarkatti PS, Nagarkatti M. Targeting CB2 cannabinoid receptors as a novel therapy to treat malignant lymphoblastic disease. Blood. 2002a;100:627–634. doi: 10.1182/blood-2002-01-0098. [DOI] [PubMed] [Google Scholar]
- McKallip RJ, Lombard C, Martin BR, Nagarkatti M, Nagarkatti PS. Δ9-Tetrahydrocannabinol-induced apoptosis in the thymus and spleen as a mechanism of immunosuppression in vitro and in vivo. J Pharmacol Exp Ther. 2002b;302:451–465. doi: 10.1124/jpet.102.033506. [DOI] [PubMed] [Google Scholar]
- McKallip RJ, Nagarkatti M, Nagarkatti PS. Δ-9-Tetrahydrocannabinol enhances breast cancer growth and metastasis by suppression of the antitumor immune response. J Immunol. 2005;174:3281–3289. doi: 10.4049/jimmunol.174.6.3281. [DOI] [PubMed] [Google Scholar]
- McKinney MK, Cravatt BF. Structure and function of fatty acid amide hydrolase. Annu Rev Biochem. 2005;74:411–434. doi: 10.1146/annurev.biochem.74.082803.133450. [DOI] [PubMed] [Google Scholar]
- McLaughlin PJ, Winston K, Swezey L, Wisniecki A, Aberman J, Tardif DJ, Betz AJ, Ishiwari K, Makriyannis A, Salamone JD. The cannabinoid CB1 antagonists SR141716A and AM 251 suppress food intake and food-reinforced behavior in a variety of tasks in rats. Behav Pharmacol. 2003;14:583–588. doi: 10.1097/00008877-200312000-00002. [DOI] [PubMed] [Google Scholar]
- McNamara JO. Emerging insights into the genesis of epilepsy. Nature (Lond) 1999;399:A15–A22. doi: 10.1038/399a015. [DOI] [PubMed] [Google Scholar]
- McPartland JM, Russo EB. Cannabis and cannabis extracts: greater than the sum of their parts? J Cannabis Ther. 2001;1:103–132. [Google Scholar]
- Mechoulam R. The pharmacohistory of cannabis sativa. In: Mechoulam R, editor. Cannabis as Therapeutic Agent. CRC Press; Boca Raton, FL: 1986. pp. 1–19. [Google Scholar]
- Mechoulam R, Benshabat S, Hanus L, Ligumsky M, Kaminski NE, Schatz AR, Gopher A, Sholomo A, Martin BR, Compton DR, et al. Identification of an endogenous 2-monoglyceride, present in canine gut, that binds to cannabinoid receptors. Biochem Pharmacol. 1995;50:83–90. doi: 10.1016/0006-2952(95)00109-d. [DOI] [PubMed] [Google Scholar]
- Mechoulam R, Fride E, Di Marzo V. Endocannabionoids. Eur J Pharmacol. 1998;359:1–18. doi: 10.1016/s0014-2999(98)00649-9. [DOI] [PubMed] [Google Scholar]
- Mechoulam R, Fride E, Hanus L, Sheskin T, Bisogno T, Di Marzo V, Bayewitch M, Vogel Z. Anandamide may mediate sleep induction. Nature (Lond) 1997;389:25–26. doi: 10.1038/37891. [DOI] [PubMed] [Google Scholar]
- Mechoulam R, Gaoni Y. The absolute configuration of Δ-1-tetrahydrocannabinol, the major active constituent of hashish. Tetrahedron Lett. 1967;12:1109–1111. doi: 10.1016/s0040-4039(00)90646-4. [DOI] [PubMed] [Google Scholar]
- Mechoulam R, Hanus L. A historical overview of chemical research on cannabinoids. Chem Phys Lipids. 2000;108:1–13. doi: 10.1016/s0009-3084(00)00184-5. [DOI] [PubMed] [Google Scholar]
- Mechoulam R, Hanus L. Cannabidiol: an overview of some chemical and pharmacological aspects. Part I: chemical aspects. Chem Phys Lipids. 2002;121:35–43. doi: 10.1016/s0009-3084(02)00144-5. [DOI] [PubMed] [Google Scholar]
- Mechoulam R, Spatz M, Shohami E. Endocannabinoids and neuroprotection. Sci STKE. 2002a;2002:129–RE5. doi: 10.1126/stke.2002.129.re5. [DOI] [PubMed] [Google Scholar]
- Mechoulam R, Panikashvili D, Shohami E. Cannabinoids and brain injury: therapeutic implications. Trends Mol Med. 2002b;8:58–61. doi: 10.1016/s1471-4914(02)02276-1. [DOI] [PubMed] [Google Scholar]
- Mechoulam R, Parker LA, Gallily R. Cannabidiol: an overview of some pharmacological aspects. J Clin Pharmacol. 2002c;42:11S–19S. doi: 10.1002/j.1552-4604.2002.tb05998.x. [DOI] [PubMed] [Google Scholar]
- Meinck HM, Schonle PW, Conrad B. Effect of cannabinoids on spasticity and ataxia in multiple sclerosis. J Neurol. 1989;236:120–122. doi: 10.1007/BF00314410. [DOI] [PubMed] [Google Scholar]
- Melck D, De Petrocellis L, Orlando P, Bisogno T, Laezza C, Bifulco M, Di Marzo V. Suppression of nerve growth factor Trk receptors and prolactin receptors by endocannabinoids leads to inhibition of human breast and prostate cancer cell proliferation. Endocrinology. 2000;141:118–126. doi: 10.1210/endo.141.1.7239. [DOI] [PubMed] [Google Scholar]
- Melck D, Rueda D, Galve-Roperh I, De Petrocellis L, Guzman M, Di Marzo V. Involvement of the cAMP/protein kinase A pathway and of mitogen-activated protein kinase in the anti-proliferative effects of anandamide in human breast cancer cells. FEBS Lett. 1999;463:235–240. doi: 10.1016/s0014-5793(99)01639-7. [DOI] [PubMed] [Google Scholar]
- Melis M, Pistis M, Perra S, Muntoni AL, Pillolla G, Gessa GL. Endocannabinoids mediate presynaptic inhibition of glutamatergic transmission in ventral tegmental area dopamine neurons through activation of CB1 receptors. J Neurosci. 2004a;24:53–62. doi: 10.1523/JNEUROSCI.4503-03.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Melis MR, Succu S, Mascia MS, Argiolas A. Antagonism of cannabinoid CB1 receptors in the paraventricular nucleus of male rats induces penile erection. Neurosci Lett. 2004b;359:17–20. doi: 10.1016/j.neulet.2004.01.025. [DOI] [PubMed] [Google Scholar]
- Melis MR, Succu S, Mascia MS, Sanna F, Melis T, Castelli MP, Argiolas A. The cannabinoid receptor antagonist SR-141716A induces penile erection in male rats: involvement of paraventricular glutamic acid and nitric oxide. Neuropharmacology. 2006;50:219–228. doi: 10.1016/j.neuropharm.2005.09.009. [DOI] [PubMed] [Google Scholar]
- Melone MA, Jori FP, Peluso G. Huntington’s disease: new frontiers for molecular and cell therapy. Curr Drug Targets. 2005;6:43–56. doi: 10.2174/1389450053344975. [DOI] [PubMed] [Google Scholar]
- Meltzer HY, Arvanitis L, Bauer D, Rein W Meta-Trial Study Group. Placebo-controlled evaluation of four novel compounds for the treatment of schizophrenia and schizoaffective disorder. Am J Psychiatry. 2004;161:975–984. doi: 10.1176/appi.ajp.161.6.975. [DOI] [PubMed] [Google Scholar]
- Mendelson WB, Basile AS. The hypnotic actions of oleamide are blocked by a cannabinoid receptor antagonist. Neuroreport. 1999;10:3237–3239. doi: 10.1097/00001756-199910190-00021. [DOI] [PubMed] [Google Scholar]
- Meng ID, Manning BH, Martin WJ, Fields HL. An analgesia circuit activated by cannabinoids. Nature (Lond) 1998;395:381–383. doi: 10.1038/26481. [DOI] [PubMed] [Google Scholar]
- Merritt JC, Crawford WJ, Alexander PC, Anduze AL, Gelbart SS. Effect of marihuana on intraocular and blood pressure in glaucoma. Ophthalmology. 1980;87:222–228. doi: 10.1016/s0161-6420(80)35258-5. [DOI] [PubMed] [Google Scholar]
- Merritt JC, Olsen JL, Armstrong JR, McKinnon SM. Topical Δ9-tetrahydrocannabinol in hypertensive glaucomas. J Pharm Pharmacol. 1981a;33:40–41. doi: 10.1111/j.2042-7158.1981.tb13699.x. [DOI] [PubMed] [Google Scholar]
- Merritt JC, Perry DD, Russell DN, Jones BF. Topical Δ9-tetrahydrocannabinol and aqueous dynamics in glaucoma. J Clin Pharmacol. 1981b;21:467S–471S. doi: 10.1002/j.1552-4604.1981.tb02626.x. [DOI] [PubMed] [Google Scholar]
- Meschler JP, Howlett AC, Madras BK. Cannabinoid receptor agonist and antagonist effects on motor function in normal and 1-methyl-4-phenyl-1,2,5,6-tetrahydropyridine (MPTP)-treated non-human primates. Psychopharmacology. 2001;156:79–85. doi: 10.1007/s002130100728. [DOI] [PubMed] [Google Scholar]
- Mesnage V, Houeto JL, Bonnet AM, Clavier I, Arnulf I, Cattelin F, Le Fur G, Damier P, Welter ML, Agid Y. Neurokinin B, neurotensin, and cannabinoid receptor antagonists and Parkinson disease. Clin Neuropharmacol. 2004;27:108–110. doi: 10.1097/00002826-200405000-00003. [DOI] [PubMed] [Google Scholar]
- Mestre L, Correa F, Arevalo-Martin A, Molina-Holgado E, Valenti M, Ortar G, Di Marzo V, Guaza C. Pharmacological modulation of the endocannabinoid system in a viral model of multiple sclerosis. J Neurochem. 2005;92:1327–1339. doi: 10.1111/j.1471-4159.2004.02979.x. [DOI] [PubMed] [Google Scholar]
- Michalopoulos GK, Bowen WC, Kule K, Luo J. HGF-, EGF-, and dexa-methasone-induced gene expression patterns during formation of tissue in hepatic organoid cultures. Gene Expr. 2003;11:55–75. doi: 10.3727/000000003108748964. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miller AS, Sanudo-Pena MC, Walker JM. Ipsilateral turning behavior induced by unilateral microinjections of a cannabinoid into the rat subthalamic nucleus. Brain Res. 1998;793:7–11. doi: 10.1016/s0006-8993(97)01475-3. [DOI] [PubMed] [Google Scholar]
- Miller CC, Murray TF, Freeman KG, Edwards GL. Cannabinoid agonist, CP 55,940, facilitates intake of palatable foods when injected into the hindbrain. Physiol Behav. 2004;80:611–616. doi: 10.1016/j.physbeh.2003.10.019. [DOI] [PubMed] [Google Scholar]
- Miller P, Lawrie SM, Hodges A, Clafferty R, Cosway R, Johnstone EC. Genetic liability, illicit drug use, life stress and psychotic symptoms: preliminary findings from the Edinburgh study of people at high risk for schizophrenia. Soc Psychiatry Psychiatr Epidemiol. 2001;36:338–342. doi: 10.1007/s001270170038. [DOI] [PubMed] [Google Scholar]
- Milman G, Maor Y, Abu-Lafi S, Horowitz M, Gallily R, Batkai S, Mo FM, Offertaler L, Pacher P, Kunos G, et al. N-arachidonoyl L-serine, an endocannabinoid-like brain constituent with vasodilatory properties. Proc Natl Sci USA. 2006;103:2428–2433. doi: 10.1073/pnas.0510676103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Milton NG. Anandamide and noladin ether prevent neurotoxicity of the human amyloid-β peptide. Neurosci Lett. 2002;332:127–130. doi: 10.1016/s0304-3940(02)00936-9. [DOI] [PubMed] [Google Scholar]
- Mimeault M, Pommery N, Wattez N, Bailly C, Henichart JP. Anti-proliferative and apoptotic effects of anandamide in human prostatic cancer cell lines: implication of epidermal growth factor receptor down-regulation and ceramide production. Prostate. 2003;56:1–12. doi: 10.1002/pros.10190. [DOI] [PubMed] [Google Scholar]
- Minokoshi Y, Alquier T, Furakawa N, Kim YB, Lee A, Xue B, Mu J, Foufelle F, Ferre P, Birnbaum MJ, et al. AMP-kinase regulates food intake by responding to hormonal and nutrient signals in the hypothalamus. Nature (Lond) 2004;428:569–574. doi: 10.1038/nature02440. [DOI] [PubMed] [Google Scholar]
- Mitchell VA, Aslan S, Safaei R, Vaughan CW. Effect of the cannabinoid ajulemic acid on rat models of neuropathic and inflammatory pain. Neurosci Lett. 2005;382:231–235. doi: 10.1016/j.neulet.2005.03.019. [DOI] [PubMed] [Google Scholar]
- Mo FM, Offertáler L, Kunos G. Atypical cannabinoid stimulates endothelial cell migration via Gi/Go-coupled receptor distinct from CB1, CB2 or EDG-1. Eur J Pharmacol. 2004;489:21–27. doi: 10.1016/j.ejphar.2004.02.034. [DOI] [PubMed] [Google Scholar]
- Moldrich G, Wenger T. Localization of the CB1 cannabinoid receptor in the rat brain: an immunohistochemical study. Peptides. 2000;21:1735–1742. doi: 10.1016/s0196-9781(00)00324-7. [DOI] [PubMed] [Google Scholar]
- Molina-Holgado E, Vela JM, Arevalo-Martin A, Almazan G, Molina-Holgado F, Borrell J, Guaza C. Cannabinoids promote oligodendrocyte progenitor survival: involvement of cannabinoid receptors and phosphatidylinositol-3 kinase/Akt signaling. J Neurosci. 2002;22:9742–9753. doi: 10.1523/JNEUROSCI.22-22-09742.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Molina-Holgado F, Lledo A, Guaza C. Anandamide suppresses nitric oxide and TNF-α responses to Theiler’s virus or endotoxin in astrocytes. Neuroreport. 1997;8:1929–1933. doi: 10.1097/00001756-199705260-00027. [DOI] [PubMed] [Google Scholar]
- Moller HJ. Antipsychotic agents: gradually improving treatment from the traditional oral neuroleptics to the first atypical depot. Eur Psychiatry. 2005;20:379–385. doi: 10.1016/j.eurpsy.2005.03.006. [DOI] [PubMed] [Google Scholar]
- Monory K, Massa F, Blaudzub H, Marsicano G, Lutz B. The role of different neuronal populations in the pharmacological actions of Δ9-tetrahydro-cannabinolin. Proceedings of the 2005 Symposium on the Cannabinoids; Burlington, VT: International Cannabinoid Research Society; 2005. p. 18. [Google Scholar]
- Monteleone P, Fabbrazzo M, Tortorella A, Fuschino A, Maj M. Opposite modifications in circulating leptin and soluble leptin receptor across the eating disorder spectrum. Mol Psychiatry. 2002;7:641–646. doi: 10.1038/sj.mp.4001043. [DOI] [PubMed] [Google Scholar]
- Monteleone P, Matias I, Martiadis V, De Petrocellis L, Maj M, Di Marzo V. Blood levels of the endocannabinoid anandamide are increased in anorexia nervosa and in binge-eating disorder, but not in bulimia nervosa. Neuropsychopharmacology. 2005;30:1216–1221. doi: 10.1038/sj.npp.1300695. [DOI] [PubMed] [Google Scholar]
- Monti JM. Hypnoticlike effects of cannabidiol in the rat. Psychopharmacology. 1977;55:263–265. doi: 10.1007/BF00497858. [DOI] [PubMed] [Google Scholar]
- Moore SA, Nomikos GG, Dickason-Chesterfield AK, Schober DA, Schaus JM, Ying B-P, Xu Y-C, Phebus L, Simmons RMA, Li D, et al. Identification of a high-affinity binding site involved in the transport of endocannabinoids. Proc Natl Acad Sci USA. 2005;102:17852–17857. doi: 10.1073/pnas.0507470102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morahan PS, Klykken PC, Smith SH, Harris LS, Munson AE. Effects of cannabinoids on host resistance to Listeria monocytogenes and herpes simplex virus. Infect Immun. 1979;23:670–674. doi: 10.1128/iai.23.3.670-674.1979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morales M, Wang S-D, Diaz-Ruiz O, Jho DH-J. Cannabinoid CB1 receptor and serotonin 3 receptor subunit A (5-HT3A) are co-expressed in GABA neurons in the rat telencephalon. J Comp Neurol. 2004;468:205–216. doi: 10.1002/cne.10968. [DOI] [PubMed] [Google Scholar]
- Morley JE. Anorexia, sarcopenia, and aging. Nutrition. 2001;17:660–663. doi: 10.1016/s0899-9007(01)00574-3. [DOI] [PubMed] [Google Scholar]
- Mukhopadhyay S, Chapnick BM, Howlett AC. Anandamide-induced vasorelaxation in rabbit aortic rings has two components: G protein dependent and independent. Am J Physiol. 2002;282:H2046–H2054. doi: 10.1152/ajpheart.00497.2001. [DOI] [PubMed] [Google Scholar]
- Mukhopadhyay S, Howlett AC. Chemically distinct ligands promote differential CB1 cannabinoid receptor-Gi protein interactions. Mol Pharmacol. 2005;67:2016–2024. doi: 10.1124/mol.104.003558. [DOI] [PubMed] [Google Scholar]
- Müller-Vahl KR. Cannabinoids reduce symptoms of Tourette’s syndrome. Expert Opin Pharmacother. 2003;4:1717–1725. doi: 10.1517/14656566.4.10.1717. [DOI] [PubMed] [Google Scholar]
- Müller-Vahl KR, Kolbe H, Dengler R. Gilles de la Tourette syndrome: effect of nicotine, alcohol and marihuana on clinical symptoms. Nervenarzt. 1997;68:985–989. doi: 10.1007/s001150050227. [DOI] [PubMed] [Google Scholar]
- Müller-Vahl KR, Kolbe H, Schneider U, Emrich HM. Cannabinoids: possible role in patho-physiology and therapy of Gilles de la Tourette syndrome. Acta Psychiatr Scand. 1998;98:502–506. doi: 10.1111/j.1600-0447.1998.tb10127.x. [DOI] [PubMed] [Google Scholar]
- Müller-Vahl KR, Kolbe H, Schneider U, Emrich HM. Cannabis in movement disorders. Forsch Komplementarmed. 1999a;6:23–27. doi: 10.1159/000057153. [DOI] [PubMed] [Google Scholar]
- Müller-Vahl KR, Prevedel H, Theloe K, Kolbe H, Emrich HM, Schneider U. Treatment of Tourette syndrome with Δ-9-tetrahydrocannabinol (Δ9-THC): no influence on neuropsychological performance. Neuropsychopharmacology. 2003a;28:384–388. doi: 10.1038/sj.npp.1300047. [DOI] [PubMed] [Google Scholar]
- Müller-Vahl KR, Schneider U, Emrich HM. Nabilone increases choreatic movements in Huntington’s disease. Mov Disord. 1999b;14:1038–1040. doi: 10.1002/1531-8257(199911)14:6<1038::aid-mds1024>3.0.co;2-7. [DOI] [PubMed] [Google Scholar]
- Müller-Vahl KR, Schneider U, Koblenz A, Jobges M, Kolbe H, Daldrup T, Emrich HM. Treatment of Tourette’s syndrome with Δ9-tetrahydrocannabinol (THC): a randomized crossover trial. Pharmacopsychiatry. 2002;35:57–61. doi: 10.1055/s-2002-25028. [DOI] [PubMed] [Google Scholar]
- Müller-Vahl KR, Schneider U, Kolbe H, Emrich HM. Treatment of Tourette’s syndrome with Δ-9-tetrahydrocannabinol. Am J Psychiatry. 1999c;156:495. [PubMed] [Google Scholar]
- Müller-Vahl KR, Schneider U, Prevedel H, Theloe K, Kolbe H, Daldrup T, Emrich HM. Δ9-Tetrahydrocannabinol (THC) is effective in the treatment of tics in Tourette syndrome: a 6-week randomized trial. J Clin Psychiatry. 2003b;64:459–465. doi: 10.4088/jcp.v64n0417. [DOI] [PubMed] [Google Scholar]
- Munro S, Thomas KL, Abu-Shaar M. Molecular characterization of a peripheral receptor for cannabinoids. Nature (Lond) 1993;365:61–65. doi: 10.1038/365061a0. [DOI] [PubMed] [Google Scholar]
- Munson AE, Harris LS, Friedman MA, Dewey WL, Carchman RA. Anti-neoplastic activity of cannabinoids. J Natl Cancer Inst. 1975;55:597–602. doi: 10.1093/jnci/55.3.597. [DOI] [PubMed] [Google Scholar]
- Murillo-Rodriguez E, Blanco-Centurion C, Sanchez C, Piomelli D, Shiromani PJ. Anandamide enhances extracellular levels of adenosine and induces sleep: an in vivo microdialysis study. Sleep. 2003;26:943–947. doi: 10.1093/sleep/26.8.943. [DOI] [PubMed] [Google Scholar]
- Murillo-Rodriguez E, Cabeza R, Mendez-Diaz M, Navarro L, Prospero-Garcia O. Anandamide-induced sleep is blocked by SR141716A, a CB1 receptor antagonist and by U73122, a phospholipase C inhibitor. Neuroreport. 2001;12:2131–2136. doi: 10.1097/00001756-200107200-00018. [DOI] [PubMed] [Google Scholar]
- Murillo-Rodriguez E, Sanchez-Alavez M, Navarro L, Martinez-Gonzalez D, Drucker-Colin R, Prospero-Garcia O. Anandamide modulates sleep and memory in rats. Brain Res. 1998;812:270–274. doi: 10.1016/s0006-8993(98)00969-x. [DOI] [PubMed] [Google Scholar]
- Muthian S, Rademacher DJ, Roelke CT, Gross GJ, Hillard CJ. Anandamide content is increased and CB1 cannabinoid receptor blockade is protective during transient, focal cerebral ischemia. Neuroscience. 2004;129:743–750. doi: 10.1016/j.neuroscience.2004.08.044. [DOI] [PubMed] [Google Scholar]
- Naassila M, Pierrefiche O, Ledent C, Daoust M. Decreased alcohol self-administration and increased alcohol sensitivity and withdrawal in CB1 receptor knockout mice. Neuropharmacology. 2004;46:243–253. doi: 10.1016/j.neuropharm.2003.09.002. [DOI] [PubMed] [Google Scholar]
- Nackley AG, Makriyannis A, Hohmann AG. Selective activation of cannabinoid CB2 receptors suppresses spinal fos protein expression and pain behavior in a rat model of inflammation. Neuroscience. 2003a;119:747–757. doi: 10.1016/s0306-4522(03)00126-x. [DOI] [PubMed] [Google Scholar]
- Nackley AG, Suplita RL, 2nd, Hohmann AG. A peripheral cannabinoid mechanism suppresses spinal fos protein expression and pain behavior in a rat model of inflammation. Neuroscience. 2003b;117:659–670. doi: 10.1016/s0306-4522(02)00870-9. [DOI] [PubMed] [Google Scholar]
- Nackley AG, Zvonok AM, Makriyannis A, Hohmann AG. Activation of cannabinoid CB2 receptors suppresses C-fiber responses and windup in spinal wide dynamic range neurons in the absence and presence of inflammation. J Neurophysiol. 2004;92:3562–3574. doi: 10.1152/jn.00886.2003. [DOI] [PubMed] [Google Scholar]
- Naderi N, Shafaghi B, Khodayar MJ, Zarindast MR. Interaction between γ-aminobutyric acid GABAB and cannabinoid CB1 receptors in spinal pain pathways in rat. Eur J Pharmacol. 2005;514:159–164. doi: 10.1016/j.ejphar.2005.03.037. [DOI] [PubMed] [Google Scholar]
- Nadler V, Biegon A, Beit-Yannai E, Adamchik J, Shohami E. 45Ca accumulation in rat brain after closed head injury; attenuation by the novel neuroprotective agent HU-211. Brain Res. 1995;685:1–11. doi: 10.1016/0006-8993(95)00367-y. [DOI] [PubMed] [Google Scholar]
- Nadler V, Mechoulam R, Sokolovsky M. Blockade of 45Ca2+ influx through the N-methyl-D-aspartate receptor ion channel by the non-psychoactive cannabinoid HU-211. Brain Res. 1993a;622:79–85. doi: 10.1016/0006-8993(93)90804-v. [DOI] [PubMed] [Google Scholar]
- Nadler V, Mechoulam R, Sokolovsky M. The non-psychotropic cannabinoid (+)-(3S,4S)-7-hydroxy-Δ6- tetrahydrocannabinol 1,1-dimethylheptyl (HU-211) attenuates N-methyl-D-aspartate receptor-mediated neurotoxicity in primary cultures of rat forebrain. Neurosci Lett. 1993b;162:43–45. doi: 10.1016/0304-3940(93)90555-y. [DOI] [PubMed] [Google Scholar]
- Nagayama T, Sinor AD, Simon RP, Chen J, Graham SH, Jin K, Greenberg DA. Cannabinoids and neuroprotection in global and focal cerebral ischemia and in neuronal cultures. J Neurosci. 1999;19:2987–2995. doi: 10.1523/JNEUROSCI.19-08-02987.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Navarro L, Martinez-vargas M, Murillo-rodriguez E, Landa A, Mendez-diaz M, Prospero-garcia O. Potential role of the cannabinoid receptor CB1 in rapid eye movement sleep rebound. Neuroscience. 2003;120:855–859. doi: 10.1016/s0306-4522(03)00339-7. [DOI] [PubMed] [Google Scholar]
- Navarro M, Carrera MR, Del Arco I, Trigo JM, Koob GF, Rodriguez de Fonseca F. Cannabinoid receptor antagonist reduces heroin self-administration only in dependent rats. Eur J Pharmacol. 2004;501:235–237. doi: 10.1016/j.ejphar.2004.08.022. [DOI] [PubMed] [Google Scholar]
- Navarro M, Carrera MR, Fratta W, Valverde O, Cossu G, Fattore L, Chowen JA, Gomez R, del Arco I, Villanua MA, et al. Functional interaction between opioid and cannabinoid receptors in drug self-administration. J Neurosci. 2001;21:5344–5350. doi: 10.1523/JNEUROSCI.21-14-05344.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Navarro M, Hernandez E, Munoz RM, del Arco I, Villanua MA, Carrera MR, Rodriguez de Fonseca F. Acute administration of the CB1 cannabinoid receptor antagonist SR 141716A induces anxiety-like responses in the rat. Neuroreport. 1997;8:491–496. doi: 10.1097/00001756-199701200-00023. [DOI] [PubMed] [Google Scholar]
- Neff GW, O’Brien CB, Reddy KR, Bergasa NV, Regev A, Molina E, Amaro R, Rodriguez MJ, Chase V, Jeffers L, et al. Preliminary observation with dronabinol in patients with intractable pruritus secondary to cholestatic liver disease. Am J Gastroenterol. 2002;97:2117–2119. doi: 10.1111/j.1572-0241.2002.05852.x. [DOI] [PubMed] [Google Scholar]
- Negrete JC. Cannabis and schizophrenia. Br J Addict. 1989;84:349–351. doi: 10.1111/j.1360-0443.1989.tb00576.x. [DOI] [PubMed] [Google Scholar]
- Nelson K, Walsh D, Deeter P, Sheehan F. A phase II study of Δ-9-tetrahydrocannabinol for appetite stimulation in cancer-associated anorexia. J Palliat Care. 1994;10:14–18. [PubMed] [Google Scholar]
- Nestler EJ. Molecular mechanism of drug addiction in the mesolimbic dopaminergic pathway. Semin Neurosci. 2003;5:369–376. [Google Scholar]
- Newell KA, Deng C, Huang XF. Increased cannabinoid receptor density in the posterior cingulate cortex in schizophrenia. Exp Brain Res. 2006;172:556–560. doi: 10.1007/s00221-006-0503-x. [DOI] [PubMed] [Google Scholar]
- Newton C, Klein T, Friedman H. The role of macrophages in THC-induced alteration of the cytokine network. Adv Exp Med Biol. 1998;437:207–214. doi: 10.1007/978-1-4615-5347-2_23. [DOI] [PubMed] [Google Scholar]
- Ng SK, Brust JC, Hauser WA, Susser M. Illicit drug use and the risk of new-onset seizures. Am J Epidemiol. 1990;132:47–57. doi: 10.1093/oxfordjournals.aje.a115642. [DOI] [PubMed] [Google Scholar]
- Ni X, Geller EB, Eppihimer MJ, Eisenstein TK, Adler MW, Tuma RF. Win 55212-2, a cannabinoid receptor agonist, attenuates leukocyte/endothelial interactions in an experimental autoimmune encephalomyelitis model. Mult Scler. 2004;10:158–164. doi: 10.1191/1352458504ms1009oa. [DOI] [PubMed] [Google Scholar]
- Nicholson AN, Turner C, Stone BM, Robson PJ. Effect of Δ-9-tetrahydrocannabinol and cannabidiol on nocturnal sleep and early-morning behavior in young adults. J Clin Psychopharmacol. 2004;24:305–313. doi: 10.1097/01.jcp.0000125688.05091.8f. [DOI] [PubMed] [Google Scholar]
- Nicholson RA, Liao C, Zheng J, David LS, Coyne L, Errington AC, Singh G, Lees G. Sodium channel inhibition by anandamide and synthetic cannabimimetics in brain. Brain Res. 2003;978:194–204. doi: 10.1016/s0006-8993(03)02808-7. [DOI] [PubMed] [Google Scholar]
- Niederhoffer N, Schmid K, Szabo B. The peripheral sympathetic nervous system is the major target of cannabinoids in eliciting cardiovascular depression. Naunyn-Schmiedeberg’s Arch Pharmacol. 2003;367:434–443. doi: 10.1007/s00210-003-0755-y. [DOI] [PubMed] [Google Scholar]
- Niederhoffer N, Szabo B. Cannabinoids cause central sympathoexcitation and bradycardia in rabbits. J Pharmacol Exp Ther. 2000;294:707–713. [PubMed] [Google Scholar]
- Nithipatikom K, Endsley MP, Isbell MA, Falck JR, Iwamoto Y, Hillard CJ, Campbell WB. 2-Arachidonoylglycerol: a novel inhibitor of androgen-independent prostate cancer cell invasion. Cancer Res. 2004;64:8826–8830. doi: 10.1158/0008-5472.CAN-04-3136. [DOI] [PubMed] [Google Scholar]
- Nogueron MI, Porgilsson B, Schneider WE, Stucky CL, Hillard CJ. Cannabinoid receptor agonists inhibit depolarization-induced calcium influx in cerebellar granule neurons. J Neurochem. 2001;79:371–381. doi: 10.1046/j.1471-4159.2001.00567.x. [DOI] [PubMed] [Google Scholar]
- Nordmann AJ, Nordmann A, Briel M, Keller U, Yancy WS, Brehm BJ, Bucher HC. Effects of low-carbohydrate vs low-fat diets on weight loss and cardiovascular risk factors: a meta-analysis of randomized controlled trials. Arch Intern Med. 2006;166:285–293. doi: 10.1001/archinte.166.3.285. [DOI] [PubMed] [Google Scholar]
- Notcutt W, Price M, Miller R, Newport S, Phillips C, Simmons S, Sansom C. Initial experiences with medicinal extracts of cannabis for chronic pain: results from 34 ‘N of 1′ studies. Anaesthesia. 2004;59:440–452. doi: 10.1111/j.1365-2044.2004.03674.x. [DOI] [PubMed] [Google Scholar]
- Noyes R, Jr, Brunk SF, Avery DH, Canter A. The analgesic properties of Δ-9-tetrahydrocannabinol and codeine. Clin Pharmacol Ther. 1975a;18:84–89. doi: 10.1002/cpt197518184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Noyes R, Jr, Brunk SF, Baram DA, Canter A. Analgesic effect of Δ-9-tetrahydrocannabinol. J Clin Pharmacol. 1975b;1:139–143. doi: 10.1002/j.1552-4604.1975.tb02348.x. [DOI] [PubMed] [Google Scholar]
- Obeso JA, Rodriguez-Oroz MC, Rodriguez M, DeLong MR, Olanow CW. Pathophysiology of levodopa-induced dyskinesias in Parkinson’s disease: problems with the current model. Ann Neurol. 2000;47:S22–S34. [PubMed] [Google Scholar]
- Obici S, Feng Z, Arduini A, Conti R, Rossetti R. Inhibition of hypothalamic carnitine palmitoyltransferase-1 decreases food intake and glucose production. Nat Med. 2003;9:756–761. doi: 10.1038/nm873. [DOI] [PubMed] [Google Scholar]
- O’Dell JR. Therapeutic strategies for rheumatoid arthritis. N Engl J Med. 2004;350:2591–2602. doi: 10.1056/NEJMra040226. [DOI] [PubMed] [Google Scholar]
- Ofek O, Karsak M, Leclerc N, Fogel M, Frenkel B, Wright K, Tam J, Attar-Namdar M, Kram V, Shohami E, et al. Peripheral cannabinoid receptor, CB2, regulates bone mass. Proc Natl Acad Sci USA. 2006;103:696–701. doi: 10.1073/pnas.0504187103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Offertáler L, Mo FM, Bátkai S, Liu J, Begg M, Razdan RK, Martin BR, Bukoski RD, Kunos G. Selective ligands and cellular effectors of a G protein-coupled endothelial cannabinoid receptor. Mol Pharmacol. 2003;63:699–705. doi: 10.1124/mol.63.3.699. [DOI] [PubMed] [Google Scholar]
- Ohno-Shosaku T, Maejima T, Kano M. Endogenous cannabinoids mediate retrograde signals from depolarized postsynaptic neurons to presynaptic terminals. Neuron. 2001;29:729–738. doi: 10.1016/s0896-6273(01)00247-1. [DOI] [PubMed] [Google Scholar]
- Okamoto Y, Morishita J, Tsuboi K, Tonai T, Ueda N. Molecular characterization of a phospholipase D generating anandamide and its congeners. J Biol Chem. 2004;279:5298–5305. doi: 10.1074/jbc.M306642200. [DOI] [PubMed] [Google Scholar]
- Onaivi ES, Green MR, Martin BR. Pharmacological characterization of cannabinoids in the elevated plus maze. J Pharmacol Exp Ther. 1990;253:1002–1009. [PubMed] [Google Scholar]
- Ortar G, Ligresti A, De Petrocellis L, Morera E, Di Marzo V. Novel selective and metabolically stable inhibitors of anandamide cellular uptake. Biochem Pharmacol. 2003;65:1473–1481. doi: 10.1016/s0006-2952(03)00109-6. [DOI] [PubMed] [Google Scholar]
- Ortega-Gutierrez S, Hawkins EG, Viso A, Lopez-Rodriguez ML, Cravatt BF. Comparison of anandamide transport in FAAH wild-type and knockout neurons: evidence for contributions by both FAAH and the CB1 receptor to anandamide uptake. Biochemistry. 2004;43:8184–8190. doi: 10.1021/bi049395f. [DOI] [PubMed] [Google Scholar]
- Ortega-Gutierrez S, Molina-Holgado E, Arevalo-Martin A, Correa F, Viso A, Lopez-Rodriguez ML, Di Marzo V, Guaza C. Activation of the endocannabinoid system as a therapeutic approach in a murine model of multiple sclerosis. FASEB J. 2005;19:1338–1343. doi: 10.1096/fj.04-2464fje. [DOI] [PubMed] [Google Scholar]
- Oruc MT, Soran A, Jain AK, Wilson JW, Fung J. De novo breast cancer in patients with liver transplantation: University of Pittsburgh’s experience and review of the literature. Liver Transplant. 2004;10:1–6. doi: 10.1002/lt.20025. [DOI] [PubMed] [Google Scholar]
- Orzelek-O’Neil RM, Goodman FR, Forney RB. Δ-9-Tetrahydrocannabinol on isolated human bronchioles. Arch Int Pharmacodyn Ther. 1980a;246:71–83. [PubMed] [Google Scholar]
- Orzelek-O’Neil RM, Goodman FR, Forney RB. The effects of Δ9-tetrahydrocannabinol and nabilone on the isolated guinea pig bronchus. Toxicol Appl Pharmacol. 1980b;54:493–500. doi: 10.1016/0041-008x(80)90176-3. [DOI] [PubMed] [Google Scholar]
- Osei-Hyiaman D, Depetrillo M, Harvey-White J, Bannon AW, Cravatt BF, Kuhar MJ, Mackie K, Palkovits M, Kunos G. Cocaine- and amphetamine-related transcript peptide is involved in the orexigenic effect of endogenous anandamide. Neuroendocrinology. 2005a;81:273–282. doi: 10.1159/000087925. [DOI] [PubMed] [Google Scholar]
- Osei-Hyiaman D, DePetrillo M, Pacher P, Liu J, Radaeva S, Batkai S, Harvey-White J, Mackie K, Offertaler L, Wang L, et al. Endocannabinoid activation at hepatic CB1 receptors stimulates fatty acid synthesis and contributes to diet-induced obesity. J Clin Investig. 2005b;115:1298–1305. doi: 10.1172/JCI23057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O’Sullivan SE, Kendall DA, Randall MD. Characterisation of the vasorelaxant properties of the novel endocannabinoid N-arachidonoyl-dopamine (NADA) Br J Pharmacol. 2004a;141:803–812. doi: 10.1038/sj.bjp.0705643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O’Sullivan SE, Kendall DA, Randall MD. Heterogeneity in the mechanisms of vasorelaxation to anandamide in resistance and conduit rat mesenteric arteries. Br J Pharmacol. 2004b;142:435–442. doi: 10.1038/sj.bjp.0705810. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ottani A, Leone S, Sandrini M, Ferrari A, Bertolini A. The analgesic activity of paracetamol is prevented by the blockade of cannabinoid CB1 receptors. Eur J Pharmacol. 2006;531:280–281. doi: 10.1016/j.ejphar.2005.12.015. [DOI] [PubMed] [Google Scholar]
- Oz M, Tchugunova Y, Dinc M. Differential effects of endogenous and synthetic cannabinoids on voltage-dependent calcium fluxes in rabbit T-tubule membranes: comparison with fatty acids. Eur J Pharmacol. 2004;502:47–58. doi: 10.1016/j.ejphar.2004.08.052. [DOI] [PubMed] [Google Scholar]
- Oz M, Tchugunova YB, Dunn SM. Endogenous cannabinoid anandamide directly inhibits voltage-dependent Ca2+ fluxes in rabbit T-tubule membranes. Eur J Pharmacol. 2000;404:13–20. doi: 10.1016/s0014-2999(00)00396-4. [DOI] [PubMed] [Google Scholar]
- Pacher P, Bátkai S, Kunos G. Haemodynamic profile and responsiveness to anandamide of TRPV1 receptor knock-out mice. J Physiol (Lond) 2004;558:647–657. doi: 10.1113/jphysiol.2004.064824. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pacher P, Bátkai S, Kunos G. Blood pressure regulation by endocannabinoids and their receptors. Neuropharmacology. 2005a;48:1130–1138. doi: 10.1016/j.neuropharm.2004.12.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pacher P, Bátkai S, Kunos G. Cardiovascular pharmacology of cannabinoids. In: Pertwee R, editor. Cannabinoids. Springer; New York: 2005b. pp. 599–627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pacher P, Bátkai S, Kunos G. Cirrhotic cardiomyopathy: an endocannabinoid connection? Br J Pharmacol. 2005c;146:313–314. doi: 10.1038/sj.bjp.0706332. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pacher P, Bátkai S, Osei-Hyiaman D, Offertaler L, Liu J, Harvey-White J, Brassai A, Jarai Z, Cravatt BF, Kunos G. Hemodynamic profile, responsiveness to anandamide, and baroreflex sensitivity of mice lacking fatty acid amide hydrolase. Am J Physiol. 2005d;289:H533–H541. doi: 10.1152/ajpheart.00107.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pacher P, Kecskeméti V. Trends in the development of new antidepressants: is there a light at the end of the tunnel? Curr Med Chem. 2004;11:925–943. doi: 10.2174/0929867043455594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pacher P, Nivorozhkin A, Szabo C. Therapeutic effects of xanthine oxidase inhibitors: renaissance half a century after the discovery of allopurinol. Pharmacol Rev. 2006;58:87–114. doi: 10.1124/pr.58.1.6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pacher P, Schulz R, Liaudet L, Szabo C. Nitrosative stress and pharmacological modulation of heart failure. Trends Pharmacol Sci. 2005e;26:302–310. doi: 10.1016/j.tips.2005.04.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Page KJ, Besret L, Jain M, Monaghan EM, Dunnett SB, Everitt BJ. Effects of systemic 3-nitropropionic acid-induced lesions of the dorsal striatum on cannabinoid and μ-opioid receptor binding in the basal ganglia. Exp Brain Res. 2000;130:142–150. doi: 10.1007/s002210050016. [DOI] [PubMed] [Google Scholar]
- Pagotto U, Marsicano G, Cota D, Lutz B, Pasquali R. The emerging role of the endocannabinoid system in endocrine regulation and energy balance. Endocr Rev. 2006;27:73–100. doi: 10.1210/er.2005-0009. [DOI] [PubMed] [Google Scholar]
- Panikashvili D, Mechoulam R, Beni SM, Alexandrovich A, Shohami E. CB1 cannabinoid receptors are involved in neuroprotection via NF-κB inhibition. J Cereb Blood Flow Metab. 2005;25:477–484. doi: 10.1038/sj.jcbfm.9600047. [DOI] [PubMed] [Google Scholar]
- Panikashvili D, Shein NA, Mechoulam R, Trembovler V, Kohen R, Alexandrovich A, Shohami E. The endocannabinoid 2-AG protects the blood-brain barrier after closed head injury and inhibits mRNA expression of proinflammatory cytokines. Neurobiol Dis. 2006;22:257–264. doi: 10.1016/j.nbd.2005.11.004. [DOI] [PubMed] [Google Scholar]
- Panikashvili D, Simeonidou C, Ben-Shabat S, Hanus L, Breuer A, Mechoulam R, Shohami E. An endogenous cannabinoid (2-AG) is neuroprotective after brain injury. Nature (Lond) 2001;413:527–531. doi: 10.1038/35097089. [DOI] [PubMed] [Google Scholar]
- Paria BC, Das SK, Dey SK. The preimplantation mouse embryo is a target for cannabinoid ligand-receptor signaling. Proc Natl Acad Sci USA. 1995;92:9460–9464. doi: 10.1073/pnas.92.21.9460. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paria BC, Song H, Wang X, Schmid PC, Krebsbach RJ, Schmid HH, Bonner TI, Zimmer A, Dey SK. Dysregulated cannabinoid signaling disrupts uterine receptivity for embryo implantation. J Biol Chem. 2001;276:20523–20528. doi: 10.1074/jbc.M100679200. [DOI] [PubMed] [Google Scholar]
- Paria BC, Wang H, Dey SK. Endocannabinoid signaling in synchronizing embryo development and uterine receptivity for implantation. Chem Phys Lipids. 2002;121:201–210. doi: 10.1016/s0009-3084(02)00156-1. [DOI] [PubMed] [Google Scholar]
- Paria BC, Zhao X, Wang J, Das SK, Dey SK. Fatty-acid amide hydrolase is expressed in the mouse uterus and embryo during the periimplantation period. Biol Reprod. 1999;60:1151–1157. doi: 10.1095/biolreprod60.5.1151. [DOI] [PubMed] [Google Scholar]
- Park B, Gibbons HM, Mitchell MD, Glass M. Identification of the CB1 cannabinoid receptor and fatty acid amide hydrolase (FAAH) in the human placenta. Placenta. 2003;24:990–995. doi: 10.1016/s0143-4004(03)00165-6. [DOI] [PubMed] [Google Scholar]
- Park B, McPartland JM, Glass M. Cannabis, cannabinoids and reproduction. Prostaglandins Leukotrienes Essent Fatty Acids. 2004;70:189–197. doi: 10.1016/j.plefa.2003.04.007. [DOI] [PubMed] [Google Scholar]
- Parker LA, Kwiatkowska M, Burton P, Mechoulam R. Effect of cannabinoids on lithium-induced vomiting in the Suncus murinus (house musk shrew) Psychopharmacology. 2004;171:156–161. doi: 10.1007/s00213-003-1571-2. [DOI] [PubMed] [Google Scholar]
- Parmentier-Batteur S, Jin K, Mao XO, Xie L, Greenberg DA. Increased severity of stroke in CB1 cannabinoid receptor knock-out mice. J Neurosci. 2002;22:9771–9775. doi: 10.1523/JNEUROSCI.22-22-09771.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parolaro D, Massi P, Rubino T, Monti E. Endocannabinoids in the immune system and cancer: endocannabinoids in the immune system and cancer. Prostaglandins Leukotrienes Essent Fatty Acids. 2002;66:319–332. doi: 10.1054/plef.2001.0355. [DOI] [PubMed] [Google Scholar]
- Pate DW, Jarvinen K, Urtti A, Jarho P, Fich M, Mahadevan V, Jarvinen T. Effects of topical anandamides on intraocular pressure in normotensive rabbits. Life Sci. 1996;58:1849–1860. doi: 10.1016/0024-3205(96)00169-5. [DOI] [PubMed] [Google Scholar]
- Pate DW, Jarvinen K, Urtti A, Jarho P, Jarvinen T. Ophthalmic arachidonylethanolamide decreases intraocular pressure in normotensive rabbits. Curr Eye Res. 1995;14:791–797. doi: 10.3109/02713689508995801. [DOI] [PubMed] [Google Scholar]
- Pate DW, Jarvinen K, Urtti A, Mahadevan V, Jarvinen T. Effect of the CB1 receptor antagonist, SR141716A, on cannabinoid-induced ocular hypotension in normotensive rabbits. Life Sci. 1998;63:2181–2188. doi: 10.1016/s0024-3205(98)00499-8. [DOI] [PubMed] [Google Scholar]
- Patel S, Cravatt BF, Hillard CJ. Synergistic interactions between cannabinoids and environmental stress in the activation of the central amygdala. Neuropsychopharmacology. 2005;30:497–507. doi: 10.1038/sj.npp.1300535. [DOI] [PubMed] [Google Scholar]
- Patel S, Roelke CT, Rademacher DJ, Cullinan WE, Hillard CJ. Endocannabinoid signaling negatively modulates stress-induced activation of the hypothalamic-pituitary-adrenal axis. Endocrinology. 2004;145:5431–5438. doi: 10.1210/en.2004-0638. [DOI] [PubMed] [Google Scholar]
- Patel S, Wohlfeil ER, Rademacher DJ, Carrier EJ, Perry LJ, Kundu A, Falck JR, Nithipatikom K, Campbell WB, Hillard CJ. The general anesthetic propofol increases brain N-arachidonylethanolamine (anandamide) content and inhibits fatty acid amide hydrolase. Br J Pharmacol. 2003;139:1005–1013. doi: 10.1038/sj.bjp.0705334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paton WDM, Pertwee RG. The actions of cannabis in man. In: Nahas GG, Paton WDM, editors. Marijuana: Chemistry, Pharmacology, Metabolism and Clinical Effects. Pergamon Press; Oxford: 1973. pp. 735–738. [Google Scholar]
- Patrick GB. Marijuana and the lung. Postgrad Med. 1980;67:110–113. 116–118. doi: 10.1080/00325481.1980.11715451. [DOI] [PubMed] [Google Scholar]
- Patsos HA, Hicks DJ, Greenhough A, Williams AC, Paraskeva C. Cannabinoids and cancer: potential for colorectal cancer therapy. Biochem Soc Trans. 2005;33:712–714. doi: 10.1042/BST0330712. [DOI] [PubMed] [Google Scholar]
- Patton GC, Coffey C, Carlin JB, Degenhardt L, Lynskey M, Hall W. Cannabis use and mental health in young people: cohort study. BMJ. 2002;325:1195–1198. doi: 10.1136/bmj.325.7374.1195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paulaskis JD, Sul HS. Cloning and expression of mouse fatty acid synthase and other specific mRNA: developmental and hormonal regulation in 3T3–L1 cells. J Biol Chem. 1988;263:7049–7054. [PubMed] [Google Scholar]
- Pazos MR, Nunez E, Benito C, Tolon RM, Romero J. Role of the endocannabinoid system in Alzheimer’s disease: new perspectives. Life Sci. 2004;75:1907–1915. doi: 10.1016/j.lfs.2004.03.026. [DOI] [PubMed] [Google Scholar]
- Pertwee RG. Cannabinoids and the gastrointestinal tract. Gut. 2001;48:859–867. doi: 10.1136/gut.48.6.859. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pertwee RG. Cannabinoids and multiple sclerosis. Pharmacol Ther. 2002;95:165–174. doi: 10.1016/s0163-7258(02)00255-3. [DOI] [PubMed] [Google Scholar]
- Pertwee RG. The therapeutic potential of drugs that target cannabinoid receptors or modulate the tissue levels or actions of endocannabinoids. AAPS J. 2005a;7:E625–E654. doi: 10.1208/aapsj070364. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pertwee RG. Inverse agonism and neutral antagonism at cannabinoid CB1 receptors. Life Sci. 2005b;76:1307–1324. doi: 10.1016/j.lfs.2004.10.025. [DOI] [PubMed] [Google Scholar]
- Pertwee RG. Pharmacological actions of cannabinoids. In: Pertwee R, editor. Cannabinoids. Springer; New York: 2005c. pp. 1–53. [Google Scholar]
- Pertwee RG, Browne SE, Ross TM, Stretton CD. An investigation of the involvement of GABA in certain pharmacological effects of Δ-9-tetrahydrocannabinol. Pharmacol Biochem Behav. 1991;40:581–585. doi: 10.1016/0091-3057(91)90366-a. [DOI] [PubMed] [Google Scholar]
- Pertwee RG, Fernando SR, Griffin G, Abadji V, Makriyannis A. Effect of phenylmethylsulphonyl fluoride on the potency of anandamide as an inhibitor of electrically evoked contractions in two isolated tissue preparations. Eur J Pharmacol. 1995;272:73–78. doi: 10.1016/0014-2999(94)00618-h. [DOI] [PubMed] [Google Scholar]
- Pertwee RG, Fernando SR, Nash JE, Coutts AA. Further evidence for the presence of cannabinoid CB1 receptors in guinea-pig small intestine. Br J Pharmacol. 1996;118:2199–2205. doi: 10.1111/j.1476-5381.1996.tb15663.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pertwee RG, Ross RA, Craib SJ, Thomas A. (−)-Cannabidiol antagonizes cannabinoid receptor agonists and noradrenaline in the mouse vas deferens. Eur J Pharmacol. 2002;456:99–106. doi: 10.1016/s0014-2999(02)02624-9. [DOI] [PubMed] [Google Scholar]
- Perwitz N, Fasshauer M, Klein J. Cannabinoid receptor signaling directly inhibits thermogenesis and alters expression of adiponectin and visfatin. Horm Metab Res. 2006;38:356–358. doi: 10.1055/s-2006-925401. [DOI] [PubMed] [Google Scholar]
- Petro DJ. Marihuana as a therapeutic agent for muscle spasm or spasticity. Psychosomatics. 1980;21:81–85. doi: 10.1016/S0033-3182(80)73129-8. [DOI] [PubMed] [Google Scholar]
- Petro DJ, Ellenberger C., Jr Treatment of human spasticity with Δ9-tetrahydrocannabinol. J Clin Pharmacol. 1981;21:413S–416S. doi: 10.1002/j.1552-4604.1981.tb02621.x. [DOI] [PubMed] [Google Scholar]
- Pfitzer T, Niederhoffer N, Szabo B. Central effects of the cannabinoid receptor agonist WIN55212-2 on respiratory and cardiovascular regulation in anaesthetised rats. Br J Pharmacol. 2004;142:943–952. doi: 10.1038/sj.bjp.0705874. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pi-Sunyer FX, Aronne LJ, Heshmati HM, Devin J, Rosenstock J for the RIO North America Group. Effect of rimonabant, a cannabinoid-1 receptor blocker, on weight and cardiometabolic risk factors in overweight or obese patients. J Am Med Assoc. 2006;295:761–775. doi: 10.1001/jama.295.7.761. [DOI] [PubMed] [Google Scholar]
- Pinto A, Tuttolomondo A, Di Raimondo D, Fernandez P, Licata G. Cerebrovascular risk factors and clinical classification of strokes. Semin Vasc Med. 2004;4:287–303. doi: 10.1055/s-2004-861497. [DOI] [PubMed] [Google Scholar]
- Pinto L, Capasso R, Di Carlo G, Izzo AA. Endocannabinoids and the gut. Prostaglandins Leukotrienes Essent Fatty Acids. 2002a;66:333–341. doi: 10.1054/plef.2001.0345. [DOI] [PubMed] [Google Scholar]
- Pinto L, Izzo AA, Cascio MG, Bisogno T, Hospodar-Scott K, Brown DR, Mascolo N, Di Marzo V, Capasso F. Endocannabinoids as physiological regulators of colonic propulsion in mice. Gastroenterology. 2002b;123:227–234. doi: 10.1053/gast.2002.34242. [DOI] [PubMed] [Google Scholar]
- Piomelli D. The molecular logic of endocannabinoid signaling. Nat Rev Neurosci. 2003;4:873–884. doi: 10.1038/nrn1247. [DOI] [PubMed] [Google Scholar]
- Pistis M, Perra S, Pillolla G, Melis M, Gessa GL, Muntoni AL. Cannabinoids modulate neuronal firing in the rat basolateral amygdala: evidence for CB1-and non-CB1-mediated actions. Neuropharmacology. 2004;46:115–125. doi: 10.1016/j.neuropharm.2003.08.003. [DOI] [PubMed] [Google Scholar]
- Pivik RT, Zarcone V, Dement WC, Hollister LE. Δ-9-Tetrahydrocannabinol and synhexl: effects on human sleep patterns. Clin Pharmacol Ther. 1972;13:426–435. doi: 10.1002/cpt1972133426. [DOI] [PubMed] [Google Scholar]
- Poirier B, Bidouard J-P, Cadrouvele C, Marniquet X, Staels B, O’Connor SE, Janiak P, Herbert J-M. The anti-obesity effect of rimonabant is associated with an improved serum lipid profile. Diabetes Obesity Metab. 2005;7:65–72. doi: 10.1111/j.1463-1326.2004.00374.x. [DOI] [PubMed] [Google Scholar]
- Poncelet M, Maruani J, Calassi R, Soubrié P. Overeating, alcohol and sucrose consumption decrease in CB1 receptor deleted mice. Neurosci Lett. 2003;343:216–218. doi: 10.1016/s0304-3940(03)00397-5. [DOI] [PubMed] [Google Scholar]
- Pontieri FE, Tanda G, Orzi F, Di Chiara G. Effects of nicotine on the nucleus accumbens and similarity to those of addictive drugs. Nature (Lond) 1996;382:255–257. doi: 10.1038/382255a0. [DOI] [PubMed] [Google Scholar]
- Porcella A, Casellas P, Gessa GL, Pani L. Cannabinoid receptor CB1 mRNA is highly expressed in the rat ciliary body: implications for the antiglaucoma properties of marihuana. Brain Res Mol Brain Res. 1998;58:240–245. doi: 10.1016/s0169-328x(98)00105-3. [DOI] [PubMed] [Google Scholar]
- Porcella A, Marchese G, Casu MA, Rocchitta A, Lai ML, Gessa GL, Pani L. Evidence for functional CB1 cannabinoid receptor expressed in the rat thyroid. Eur J Endocrinol. 2002;147:255–261. doi: 10.1530/eje.0.1470255. [DOI] [PubMed] [Google Scholar]
- Porcella A, Maxia C, Gessa GL, Pani L. The human eye expresses high levels of CB1 cannabinoid receptor mRNA and protein. Eur J Neurosci. 2000;12:1123–1127. doi: 10.1046/j.1460-9568.2000.01027.x. [DOI] [PubMed] [Google Scholar]
- Porcella A, Maxia C, Gessa GL, Pani L. The synthetic cannabinoid WIN55212-2 decreases the intraocular pressure in human glaucoma resistant to conventional therapies. Eur J Neurosci. 2001;13:409–412. doi: 10.1046/j.0953-816x.2000.01401.x. [DOI] [PubMed] [Google Scholar]
- Portella G, Laezza C, Laccetti P, De Petrocellis L, Di Marzo V, Bifulco M. Inhibitory effects of cannabinoid CB1 receptor stimulation on tumor growth and metastatic spreading: actions on signals involved in angiogenesis and metastasis. FASEB J. 2003;17:1771–1773. doi: 10.1096/fj.02-1129fje. [DOI] [PubMed] [Google Scholar]
- Porter AC, Sauer JM, Knierman MD, Becker GW, Berna MJ, Bao J, Nomikos GG, Carter P, Bymaster FP, Leese AB, et al. Characterization of a novel endocannabinoid, virodhamine, with antagonist activity at the CB1 receptor. J Pharmacol Exp Ther. 2002;301:1020–1024. doi: 10.1124/jpet.301.3.1020. [DOI] [PubMed] [Google Scholar]
- Powles T, te Poele R, Shamash J, Chaplin T, Propper D, Joel S, Oliver T, Liu WM. Cannabis-induced cytotoxicity in leukemic cell lines: the role of the cannabinoid receptors and the MAPK pathway. Blood. 2005;105:1214–1221. doi: 10.1182/blood-2004-03-1182. [DOI] [PubMed] [Google Scholar]
- Prather PL, Martin NA, Breivogel CS, Childers SR. Activation of cannabinoid receptors in rat brain by WIN 55212-2 produces coupling to multiple G protein α-subunits with different potencies. Mol Pharmacol. 2000;57:1000–1010. [PubMed] [Google Scholar]
- Pryce G, Baker D. Emerging properties of cannabinoid medicines in management of multiple sclerosis. Trends Neurosci. 2005;28:272–276. doi: 10.1016/j.tins.2005.03.006. [DOI] [PubMed] [Google Scholar]
- Pryce G, Ahmed Z, Hankey DJ, Jackson SJ, Croxford JL, Pocock JM, Ledent C, Petzold A, Thompson AJ, Giovannoni G, et al. Cannabinoids inhibit neurodegeneration in models of multiple sclerosis. Brain. 2003;126:2191–2202. doi: 10.1093/brain/awg224. [DOI] [PubMed] [Google Scholar]
- Pugh G, Jr, Mason DJ, Jr, Combs V, Welch SP. Involvement of dynorphin B in the antinociceptive effects of the cannabinoid CP55,940 in the spinal cord. J Pharmacol Exp Ther. 1997;281:730–737. [PubMed] [Google Scholar]
- Purnell WD, Gregg JM. Δ9-Tetrahydrocannabinol, euphoria and intraocular pressure in man. Ann Ophthalmol. 1975;7:921–923. [PubMed] [Google Scholar]
- Quartilho A, Mata HP, Ibrahim MM, Vanderah TW, Porreca F, Makriyannis A, Malan TP., Jr Inhibition of inflammatory hyperalgesia by activation of peripheral CB2 cannabinoid receptors. Anesthesiology. 2003;99:955–960. doi: 10.1097/00000542-200310000-00031. [DOI] [PubMed] [Google Scholar]
- Racz I, Bilkei-Gorzo A, Toth ZE, Michel K, Palkovits M, Zimmer A. A critical role for the cannabinoid CB1 receptors in alcohol dependence and stress- stimulated ethanol drinking. J Neurosci. 2003;23:2453–2458. doi: 10.1523/JNEUROSCI.23-06-02453.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rademacher DJ, Patel S, Hopp FA, Dean C, Hillard CJ, Seagard JL. Microinjection of a cannabinoid receptor antagonist into the NTS increases baroreflex duration in dogs. Am J Physiol. 2003;284:H1570–H1576. doi: 10.1152/ajpheart.00772.2002. [DOI] [PubMed] [Google Scholar]
- Raft D, Gregg J, Ghia J, Harris L. Effects of intravenous tetrahydrocannabinol on experimental and surgical pain: psychological correlates of the analgesic response. Clin Pharmacol Ther. 1977;21:26–33. doi: 10.1002/cpt197721126. [DOI] [PubMed] [Google Scholar]
- Ralevic V, Kendall DA, Randall MD, Smart D. Cannabinoid modulation of sensory neurotransmission via cannabinoid and vanilloid receptors: roles in regulation of cardiovascular function. Life Sci. 2002;71:2577–2594. doi: 10.1016/s0024-3205(02)02086-6. [DOI] [PubMed] [Google Scholar]
- Raman C, McAllister SD, Rizvi G, Patel SG, Moore DH, Abood ME. Amyotrophic lateral sclerosis: delayed disease progression in mice by treatment with a cannabinoid. Amyotroph Lateral Scler Other Motor Neuron Disord. 2004;5:33–39. doi: 10.1080/14660820310016813. [DOI] [PubMed] [Google Scholar]
- Ramirez BG, Blazquez C, Gómez del Pulgar T, Guzman M, de Ceballos ML. Prevention of Alzheimer’s disease pathology by cannabinoids: neuroprotection mediated by blockade of microglial activation. J Neurosci. 2005;25:1904–1913. doi: 10.1523/JNEUROSCI.4540-04.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ramos JA, Gonzalez S, Sagredo O, Gomez-Ruiz M, Fernandez-Ruiz J. Therapeutic potential of the endocannabinoid system in the brain. Mini Rev Med Chem. 2005;5:609–617. doi: 10.2174/1389557054368817. [DOI] [PubMed] [Google Scholar]
- Randall MD, Harris D, Kendall DA, Ralevic V. Cardiovascular effects of cannabinoids. Pharmacol Ther. 2002;95:191–202. doi: 10.1016/s0163-7258(02)00258-9. [DOI] [PubMed] [Google Scholar]
- Randall MD, Kendall DA, O’Sullivan The complexities of the cardiovascular actions of cannabinoids. Br J Pharmacol. 2004;142:20–26. doi: 10.1038/sj.bjp.0705725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ravinet Trillou C, Arnone M, Delgorge C, Gonalons N, Keane P, Maffrand JP, Soubrié P. Anti-obesity effect of SR141716, a CB1 receptor antagonist, in diet-induced obese mice. Am J Physiol. 2003;284:R345–R353. doi: 10.1152/ajpregu.00545.2002. [DOI] [PubMed] [Google Scholar]
- Ravinet Trillou C, Delgorge C, Menet C, Arnone M, Soubrié P. CB1 cannabinoid receptor knockout in mice leads to leanness, resistance to diet-induced obesity and enhanced leptin sensitivity. Int J Obes. 2004;24:640–648. doi: 10.1038/sj.ijo.0802583. [DOI] [PubMed] [Google Scholar]
- Rawson RA, Obert JL, McCann MJ, Mann AJ. Cocaine treatment outcome: cocaine use following inpatient, outpatient and no treatment. NIDA Res Monogr. 1986;67:271–277. [PubMed] [Google Scholar]
- Ray AM, Benham CD, Roberts JC, Gill CH, Lanneau C, Gitterman DP, Harries M, Davis JB, Davies CH. Capsazepine protects against neuronal injury caused by oxygen glucose deprivation by inhibiting Ih. J Neurosci. 2003;23:10146–10153. doi: 10.1523/JNEUROSCI.23-31-10146.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raza M, Pal S, Rafiq A, DeLorenzo RJ. Long-term alteration of calcium homeostatic mechanisms in the pilocarpine model of temporal lobe epilepsy. Brain Res. 2001;903:1–12. doi: 10.1016/s0006-8993(01)02127-8. [DOI] [PubMed] [Google Scholar]
- Regelson W, Butler JR, Schulz J, Kirk T, Peek L, Green ML, Zalis MO. Δ-9-THC as an effective antidepressant and appetite-stimulating agent in advanced cancer patients. In: Braude MC, Szara S, editors. The Pharmacology of Marijuana. Raven Press; New York: 1976. pp. 763–776. [Google Scholar]
- Reggio PH. Pharmacophores for ligand recognition and activation/inactivation of the cannabinoid receptors. Curr Pharm Des. 2003;9:1607–1633. doi: 10.2174/1381612033454577. [DOI] [PubMed] [Google Scholar]
- Reilly SM, Skuse DH, Wolke D, Stevenson J. Oral-motor dysfunction in children who fail to thrive: organic or non-organic? Dev Med Child Neurol. 1999;41:115–122. doi: 10.1017/s0012162299000225. [DOI] [PubMed] [Google Scholar]
- Reynolds JR. On therapeutic uses and toxic effects of cannabis indica. Lancet. 1890;1:637–638. [Google Scholar]
- Rhee MH, Beywitch M, Avidor-Reiss T, Levy R, Vogel Z. Cannabinoid receptor activation differentially regulates the various adenylyl cyclase isozymes. J Neurochem. 1998;71:1525–1534. doi: 10.1046/j.1471-4159.1998.71041525.x. [DOI] [PubMed] [Google Scholar]
- Rhee MH, Vogel Z, Barg J, Bayewitch M, Levy R, Hanus L, Breuer A, Mechoulam R. Cannabinol derivatives: binding to cannabinoid receptors and inhibition of adenylyl cyclase. J Med Chem. 1997;40:3228–3233. doi: 10.1021/jm970126f. [DOI] [PubMed] [Google Scholar]
- Richardson JD, Aanonsen L, Hargreaves KM. SR141716A, a cannabinoid receptor antagonist, produces hyperalgesia in untreated mice. Eur J Pharmacol. 1997;319:R3–R4. doi: 10.1016/s0014-2999(96)00952-1. [DOI] [PubMed] [Google Scholar]
- Richardson JD, Aanonsen L, Hargreaves KM. Antihyperalgesic effects of spinal cannabinoids. Eur J Pharmacol. 1998a;345:145–153. doi: 10.1016/s0014-2999(97)01621-x. [DOI] [PubMed] [Google Scholar]
- Richardson JD, Aanonsen L, Hargreaves KM. Hypoactivity of the spinal cannabinoid system results in NMDA-dependent hyperalgesia. J Neurosci. 1998b;18:451–457. doi: 10.1523/JNEUROSCI.18-01-00451.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Richardson JD, Kilo S, Hargreaves KM. Cannabinoids reduce hyperalgesia and inflammation via interaction with peripheral CB1 receptors. Pain. 1998c;75:111–119. doi: 10.1016/S0304-3959(97)00213-3. [DOI] [PubMed] [Google Scholar]
- Richfield EK, Herkenham M. Selective vulnerability in Huntington’s disease: preferential loss of cannabinoid receptors in lateral globus pallidus. Ann Neurol. 1994;36:577–584. doi: 10.1002/ana.410360406. [DOI] [PubMed] [Google Scholar]
- Richter A, Löscher W. (+)-WIN 55,212-2, a novel cannabinoid receptor agonist, exerts antidystonic effects in mutant dystonic hamsters. Eur J Pharmacol. 1994;264:371–377. doi: 10.1016/0014-2999(94)00490-0. [DOI] [PubMed] [Google Scholar]
- Richter A, Löscher W. Effects of pharmacological manipulations of cannabinoid receptors on severity of dystonia in a genetic model of paroxysmal dyskinesia. Eur J Pharmacol. 2002;454:145–151. doi: 10.1016/s0014-2999(02)02477-9. [DOI] [PubMed] [Google Scholar]
- Riegel AC, Lupica CR. Independent presynaptic and postsynaptic mechanisms regulate endocannabinoid signaling at multiple synapses in the ventral tegmental area. J Neurosci. 2004;24:11070–11078. doi: 10.1523/JNEUROSCI.3695-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rinaldi-Carmona M, Barth F, Heaulme M, Shire D, Calandra B, Congy C, Matinez S, Marvani J, Neliat G, Caput D, et al. SR141716A, a potent and selective antagonist of the brain cannabinoid receptor. FEBS Lett. 1994;350:240–244. doi: 10.1016/0014-5793(94)00773-x. [DOI] [PubMed] [Google Scholar]
- Rinaldi-Carmona M, Barth F, Millan J, Derocq JM, Casellas P, Congy C, Oustric D, Sarran M, Bouaboula M, Calandra B, et al. SR 144528, the first potent and selective antagonist of the CB2 cannabinoid receptor. J Pharmacol Exp Ther. 1998;284:644–650. [PubMed] [Google Scholar]
- Rios C, Gomes I, Devi LA. μ Opioid and CB1 cannabinoid receptor interactions: reciprocal inhibition of receptor signaling and neuritogenesis. Br J Pharmacol. 2006;148:387–395. doi: 10.1038/sj.bjp.0706757. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rivas-V JF, Garcia R. Inhibition of histamine-stimulated gastric acid secretion by −9-tetrahydrocannabinol in rat isolated stomach. Eur J Pharmacol. 1980;65:317–318. doi: 10.1016/0014-2999(80)90410-0. [DOI] [PubMed] [Google Scholar]
- Robbe D, Alonso G, Duchamp F, Bockaert J, Manzoni OJ. Localization and mechanisms of action of cannabinoid receptors at the glutamatergic synapses of the mouse nucleus accumbens. J Neurosci. 2001;21:109–116. doi: 10.1523/JNEUROSCI.21-01-00109.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robbe D, Kopf M, Remaury A, Bockaert J, Manzoni OJ. Endogenous cannabinoids mediate long-term synaptic depression in the nucleus accumbens. Proc Natl Acad Sci USA. 2002;99:8384–8388. doi: 10.1073/pnas.122149199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robson P. Human studies of cannabinoids and medicinal cannabis. In: Pertwee R, editor. Cannabinoids. Springer; New York: 2005. pp. 719–757. [DOI] [PubMed] [Google Scholar]
- Rodriguez de Fonseca F, Carrera MRA, Navarro M, Koob GF, Weiss F. Activation of corticotrophin-releasing factor in the limbic system during cannabinoid withdrawal. Science (Wash DC) 1997;276:2050–2054. doi: 10.1126/science.276.5321.2050. [DOI] [PubMed] [Google Scholar]
- Rodriguez De Fonseca F, Gorriti MA, Bilbao A, Escuredo L, Garcia-Segura LM, Piomelli D, Navarro M. Role of the endogenous cannabinoid system as a modulator of dopamine transmission: implications for Parkinson’s disease and schizophrenia. Neurotox Res. 2001;3:23–35. doi: 10.1007/BF03033228. [DOI] [PubMed] [Google Scholar]
- Rodriguez de Fonseca F, Rubio P, Menzaghi F, Merlo-Pich E, Rivier J, Koob GF, Navarro M. Corticotropin-releasing factor (CRF) antagonist [D-Phe12,Nle21,38yαMeLeu37]CRF attenuates the acute actions of the highly potent cannabinoid receptor agonist HU-210 on defensive-withdrawal behavior in rats. J Pharmacol Exp Ther. 1996;276:56–64. [PubMed] [Google Scholar]
- Rog DJ, Nurmikko TF, Friede T, Young CA. Randomized, controlled trial of cannabisbased medicine in central pain in multiple sclerosis. Neurology. 2005;65:812–819. doi: 10.1212/01.wnl.0000176753.45410.8b. [DOI] [PubMed] [Google Scholar]
- Romero EM, Fernandez B, Sagredo O, Gomez N, Uriguen L, Guaza C, De Miguel R, Ramos JA, Viveros MP. Antinociceptive, behavioural and neuroendocrine effects of CP 55,940 in young rats. Brain Res Dev Brain Res. 2002a;136:85–92. doi: 10.1016/s0165-3806(02)00306-1. [DOI] [PubMed] [Google Scholar]
- Romero J, Berrendero F, Garcia-Gil L, de la Cruz P, Ramos JA, Fernandez-Ruiz JJ. Loss of cannabinoid receptor binding and messenger RNA levels and cannabinoid agonist-stimulated [35S]guanylyl-5′O-(thio)-triphosphate binding in the basal ganglia of aged rats. Neuroscience. 1998;84:1075–1083. doi: 10.1016/s0306-4522(97)00552-6. [DOI] [PubMed] [Google Scholar]
- Romero J, Berrendero F, Perez-Rosado A, Manzanares J, Rojo A, Fernandez-Ruiz JJ, de Yebenes JG, Ramos JA. Unilateral 6-hydroxydopamine lesions of nigrostriatal dopaminergic neurons increased CB1 receptor mRNA levels in the caudate-putamen. Life Sci. 2000;66:485–494. doi: 10.1016/s0024-3205(99)00618-9. [DOI] [PubMed] [Google Scholar]
- Romero J, de Miguel R, Garcia-Palomero E, Fernandez-Ruiz JJ, Ramos JA. Time-course of the effects of anandamide, the putative endogenous cannabinoid receptor ligand, on extrapyramidal function. Brain Res. 1995a;694:223–232. doi: 10.1016/0006-8993(95)00835-e. [DOI] [PubMed] [Google Scholar]
- Romero J, Garcia L, Cebeira M, Zadrozny D, Fernandez-Ruiz JJ, Ramos JA. The endogenous cannabinoid receptor ligand, anandamide, inhibits the motor behavior: role of nigrostriatal dopaminergic neurons. Life Sci. 1995b;56:2033–2040. doi: 10.1016/0024-3205(95)00186-a. [DOI] [PubMed] [Google Scholar]
- Romero J, Lastres-Becker I, de Miguel R, Berrendero F, Ramos JA, Fernandez-Ruiz J. The endogenous cannabinoid system and the basal ganglia. Biochemical, pharmacological, and therapeutic aspects. Pharmacol Ther. 2002b;95:137–152. doi: 10.1016/s0163-7258(02)00253-x. [DOI] [PubMed] [Google Scholar]
- Ros J, Claria J, To-Figueras J, Planaguma A, Cejudo-Martin P, Fernandez-Varo G, Martin-Ruiz R, Arroyo V, Rivera F, Rodes J, et al. Endogenous cannabinoids: a new system involved in the homeostasis of arterial pressure in experimental cirrhosis in the rat. Gastroenterology. 2002;122:85–93. doi: 10.1053/gast.2002.30305. [DOI] [PubMed] [Google Scholar]
- Rosch S, Ramer R, Brune K, Hinz B. R(+)-Methanandamide and other cannabinoids induce the expression of cyclooxygenase-2 and matrix metalloproteinases in human nonpigmented ciliary epithelial cells. J Pharmacol Exp Ther. 2006;316:1219–1228. doi: 10.1124/jpet.105.092858. [DOI] [PubMed] [Google Scholar]
- Rosenblatt KA, Daling JR, Chen C, Sherman KJ, Schwartz SM. Marijuana use and risk of oral squamous cell carcinoma. Cancer Res. 2004;64:4049–4054. doi: 10.1158/0008-5472.CAN-03-3425. [DOI] [PubMed] [Google Scholar]
- Rosenkrantz H, Braude M. Acute, subacute and 23-day chronic marihuana inhalation toxicities in the rat. Taxicol Appl Pharmacol. 1974;28:428–441. doi: 10.1016/0041-008x(74)90228-2. [DOI] [PubMed] [Google Scholar]
- Ross RA, Brockie HC, Stevenson LA, Murphy VL, Templeton F, Makriyannis A, Pertwee RG. Agonist-inverse agonist characterization at CB1 and CB2 cannabinoid receptors of L759633, L759656, and AM630. Br J Pharmacol. 1999;126:665–672. doi: 10.1038/sj.bjp.0702351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rossato M, Ion Popa F, Ferigo M, Clari G, Foresta C. Human sperm express cannabinoid receptor Cb1, the activation of which inhibits motility, acrosome reaction, and mitochondrial function. J Clin Endocrinol Metab. 2005;90:984–991. doi: 10.1210/jc.2004-1287. [DOI] [PubMed] [Google Scholar]
- Roth MD. Pharmacology: marijuana and your heart. Nature (Lond) 2005;434:708–709. doi: 10.1038/434708a. [DOI] [PubMed] [Google Scholar]
- Rotzinger S, Vaccarino FJ. Cholecystokinin receptor subtypes: role in the modulation of anxiety-related and reward-related behaviours in animal models. J Psychiatry Neurosci. 2003;28:171–181. [PMC free article] [PubMed] [Google Scholar]
- Rowland LP, Shneider NA. Amyotrophic lateral sclerosis. N Engl J Med. 2001;344:1688–1700. doi: 10.1056/NEJM200105313442207. [DOI] [PubMed] [Google Scholar]
- Rowland NE, Mukherjee M, Roberston K. Effects of the cannabinoid receptor antagonist SR 141716, alone and in combination with dexfenfluramine or naloxone, on food intake in rats. Psychopharmacology. 2001;159:111–116. doi: 10.1007/s002130100910. [DOI] [PubMed] [Google Scholar]
- Rubino T, Massi P, Vigano D, Fuzio D, Parolaro D. Long-term treatment with SR141716A, the CB1 receptor antagonist, influences morphine withdrawal syndrome. Life Sci. 2000;66:2213–2219. doi: 10.1016/s0024-3205(00)00547-6. [DOI] [PubMed] [Google Scholar]
- Rudich Z, Stinson J, Jeavons M, Brown SC. Treatment of chronic intractable neuropathic pain with dronabinol: case report of two adolescents. Pain Res Manag. 2003;8:221–224. doi: 10.1155/2003/675976. [DOI] [PubMed] [Google Scholar]
- Rueda D, Galve-Roperh I, Haro A, Guzman M. The CB(1) cannabinoid receptor is coupled to the activation of c-Jun N-terminal kinase. Mol Pharmacol. 2000;58:814–820. doi: 10.1124/mol.58.4.814. [DOI] [PubMed] [Google Scholar]
- Ruiu S, Pinna GA, Marchese G, Mussinu JM, Saba P, Tambaro S, Casti P, Vargiu R, Pani L. Synthesis and characterization of NESS 0327: a novel putative antagonist of the CB1 cannabinoid receptor. J Pharmacol Exp Ther. 2003;306:363–370. doi: 10.1124/jpet.103.049924. [DOI] [PubMed] [Google Scholar]
- Russo E. A tale of two cannabinoids: the therapeutic rationale for combining tetrahydrocannabinol and cannabidiol. Med Hypotheses. 2006;66:234–246. doi: 10.1016/j.mehy.2005.08.026. [DOI] [PubMed] [Google Scholar]
- Russo EB. Clinical endocannabinoid deficiency (CECD): can this concept explain therapeutic benefits of cannabis in migraine, fibromyalgia, irritable bowel syndrome and other treatment-resistant conditions? Neuro Endocrinol Lett. 2004;25:31–39. [PubMed] [Google Scholar]
- Rutkowska M, Fereniec-Goltbiewska L. ACEA (arachidonyl-2-chloroethylamide), the selective cannabinoid CB1 receptor agonist, protects against aspirin-induced gastric ulceration. Pharmazie 2006. 2006;61:341–342. [PubMed] [Google Scholar]
- Rutkowska M, Jamontt J, Gliniak H. Effects of cannabinoids on the anxiety-like response in mice. Pharmacol Rep. 2006;58:200–206. [PubMed] [Google Scholar]
- Ryberg E, Vu HK, Larsson N, Groblewski T, Hjorth S, Elebring T, Sjogren S, Greasley PJ. Identification and characterisation of a novel splice variant of the human CB1 receptor. FEBS Lett. 2005;579:259–264. doi: 10.1016/j.febslet.2004.11.085. [DOI] [PubMed] [Google Scholar]
- Saario SM, Savinainen JR, Laitinen JT, Jarvinen T, Niemi R. Monoglyceride lipase-like enzymatic activity is responsible for hydrolysis of 2-arachidonoylglycerol in rat cerebellar membranes. Biochem Pharmacol. 2004;67:1381–1387. doi: 10.1016/j.bcp.2003.12.003. [DOI] [PubMed] [Google Scholar]
- Salim K, Schneider U, Burstein S, Hoy L, Karst M. Pain measurements and side effect profile of the novel cannabinoid ajulemic acid. Neuropharmacology. 2005;48:1164–1171. doi: 10.1016/j.neuropharm.2005.02.010. [DOI] [PubMed] [Google Scholar]
- Sánchez C, de Ceballos ML, del Pulgar TG, Rueda D, Corbacho C, Velasco G, Galve-Roperh I, Huffman JW, Ramon y Cajal S, Guzman M. Inhibition of glioma growth in vivo by selective activation of the CB2 cannabinoid receptor. Cancer Res. 2001a;61:5784–5789. [PubMed] [Google Scholar]
- Sánchez C, Galve-Roperh I, Canova C, Brachet P, Guzman M. Δ9-Tetrahydrocannabinol induces apoptosis in C6 glioma cells. FEBS Lett. 1998;436:6–10. doi: 10.1016/s0014-5793(98)01085-0. [DOI] [PubMed] [Google Scholar]
- Sánchez C, Rueda D, Segui B, Galve-Roperh I, Levade T, Guzman M. The CB1 cannabinoid receptor of astrocytes is coupled to sphingomyelin hydrolysis through the adaptor protein fan. Mol Pharmacol. 2001b;59:955–959. doi: 10.1124/mol.59.5.955. [DOI] [PubMed] [Google Scholar]
- Sánchez MG, Ruiz-Llorente L, Sanchez AM, Diaz-Laviada I. Activation of phosphoinositide 3-kinase/PKB pathway by CB1 and CB2 cannabinoid receptors expressed in prostate PC-3 cells: involvement in Raf-1 stimulation and NGF induction. Cell Signal. 2003;15:851–859. doi: 10.1016/s0898-6568(03)00036-6. [DOI] [PubMed] [Google Scholar]
- Sañudo-Peña MC, Patrick SL, Khen S, Patrick RL, Tsou K, Walker JM. Cannabinoid effects in basal ganglia in a rat model of Parkinson’s disease. Neurosci Lett. 1998;248:171–174. doi: 10.1016/s0304-3940(98)00368-1. [DOI] [PubMed] [Google Scholar]
- Sañudo-Peña MC, Strangman NM, Mackie K, Walker JM, Tsou K. CB1 receptor localization in spinal cord and roots, dorsal root ganglion, and peripheral nerve. Acta Pharmacol Sin. 1999a;12:1115–1120. [PubMed] [Google Scholar]
- Sañudo-Peña MC, Tsou K, Walker JM. Motor actions of cannabinoids in the basal ganglia output nuclei. Life Sci. 1999b;65:703–713. doi: 10.1016/s0024-3205(99)00293-3. [DOI] [PubMed] [Google Scholar]
- Sañudo-Peña MC, Walker JM. Effects of intrapallidal cannabinoids on rotational behavior in rats: interactions with the dopaminergic system. Synapse. 1998;28:27–32. doi: 10.1002/(SICI)1098-2396(199801)28:1<27::AID-SYN4>3.0.CO;2-E. [DOI] [PubMed] [Google Scholar]
- Saper CB. The central autonomic nervous system: conscious visceral perception and autonomic pattern generation. Annu Rev Neurosci. 2002;25:433–469. doi: 10.1146/annurev.neuro.25.032502.111311. [DOI] [PubMed] [Google Scholar]
- Sarfaraz S, Afaq F, Adhami VM, Mukhtar H. Cannabinoid receptor as a novel target for the treatment of prostate cancer. Cancer Res. 2005;65:1635–1641. doi: 10.1158/0008-5472.CAN-04-3410. [DOI] [PubMed] [Google Scholar]
- Sarker KP, Biswas KK, Yamakuchi M, Lee KY, Hahiguchi T, Kracht M, Kitajima I, Maruyama I. ASK1–p38 MAPK/JNK signaling cascade mediates anandamide-induced PC12 cell death. J Neurochem. 2003;85:50–61. doi: 10.1046/j.1471-4159.2003.01663.x. [DOI] [PubMed] [Google Scholar]
- Sarker KP, Obara S, Nakata M, Kitajima I, Maruyama I. Anandamide induces apoptosis of PC-12 cells: involvement of superoxide and caspase-3. FEBS Lett. 2000;472:1039–1044. doi: 10.1016/s0014-5793(00)01425-3. [DOI] [PubMed] [Google Scholar]
- Savinainen JR, Laitinen JT. Detection of cannabinoid CB1, adenosine A1, muscarinic acetylcholine, and GABAB receptor-dependent G protein activity in transducin-deactivated membranes and autoradiography sections of rat retina. Cell Mol Neurobiol. 2004;24:243–256. doi: 10.1023/B:CEMN.0000018619.18631.53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sawzdargo M, Nguyen T, Lee DK, Lynch KR, Cheng R, Heng HHQ, George SR, O’Dowd BF. Identification and cloning of three novel human G protein-coupled receptor genes GPR52, ΨGPR53 and GPR55: GPR55 is extensively expressed in human brain. Mol Brain Res. 1999;64:193–198. doi: 10.1016/s0169-328x(98)00277-0. [DOI] [PubMed] [Google Scholar]
- Scadden DT. AIDS-related malignancies. Annu Rev Med. 2003;54:285–303. doi: 10.1146/annurev.med.54.101601.152143. [DOI] [PubMed] [Google Scholar]
- Schabitz WR, Giuffrida A, Berger C, Aschoff A, Schwaninger M, Schwab S, Piomelli D. Release of fatty acid amides in a patient with hemispheric stroke: a microdialysis study. Stroke. 2002;33:2112–2114. doi: 10.1161/01.str.0000023491.63693.18. [DOI] [PubMed] [Google Scholar]
- Schelling G, Hauer D, Azad SC, Schmoelz M, Chouker A, Schmidt M, Hornuss C, Rippberger M, Briegel J, Thiel M, et al. Effects of general anesthesia on anandamide blood levels in humans. Anesthesiology. 2006;104:273–277. doi: 10.1097/00000542-200602000-00012. [DOI] [PubMed] [Google Scholar]
- Schmid K, Niederhoffer N, Szabo B. Analysis of the respiratory effects of cannabinoids in rats. Naunyn-Schmiedeberg’s Arch Pharmacol. 2003;368:301–308. doi: 10.1007/s00210-003-0787-3. [DOI] [PubMed] [Google Scholar]
- Schmid PC, Krebsbach RJ, Perry SR, Dettmer TM, Maasson JL, Schmid HH. Occurrence and postmortem generation of anandamide and other long-chain N-acylethanolamines in mammalian brain. FEBS Lett. 1995;375:117–120. doi: 10.1016/0014-5793(95)01194-j. [DOI] [PubMed] [Google Scholar]
- Schmid PC, Paria BC, Krebsbach RJ, Schmid HH, Dey SK. Changes in anandamide levels in mouse uterus are associated with uterine receptivity for embryo implantation. Proc Natl Acad Sci USA. 1997;94:4188–4192. doi: 10.1073/pnas.94.8.4188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schmid PC, Reddy PV, Natarajan V, Schmid HH. Metabolism on N-acylethanolamine phospholipids by a mammalian phosphodiesterase of the phospholipase D type. J Biol Chem. 1983;258:9302–9306. [PubMed] [Google Scholar]
- Schmid PC, Zuzarte-Augustin ML, Schmid HH. Properties of rat liver N-acylethanolamine amidohydrolase. J Biol Chem. 1985;260:14145–14149. [PubMed] [Google Scholar]
- Schon F, Hart PE, Hodgson TL, Pambakian AL, Ruprah M, Williamson EM, Kennard C. Suppression of pendular nystagmus by smoking cannabis in a patient with multiple sclerosis. Neurology. 1999;53:2209–2210. doi: 10.1212/wnl.53.9.2209-a. [DOI] [PubMed] [Google Scholar]
- Schuckit MA. Low level of response to alcohol as a predictor of future alcoholism. Am J Psychiatry. 1997;151:184–189. doi: 10.1176/ajp.151.2.184. [DOI] [PubMed] [Google Scholar]
- Schuel H, Burkman LJ. A tale of two cells: endocannabinoid-signaling regulates functions of neurons and sperm. Biol Reprod. 2005;73:1078–1086. doi: 10.1095/biolreprod.105.043273. [DOI] [PubMed] [Google Scholar]
- Scorticati C, Fernandez-Solari J, De Laurentiis A, Mohn C, Prestifilippo JP, Lasaga M, Seilicovich A, Billi S, Franchi A, McCann SM, et al. The inhibitory effect of anandamide on luteinizing hormone-releasing hormone secretion is reversed by estrogen. Proc Natl Acad Sci USA. 2004;101:11891–11896. doi: 10.1073/pnas.0404366101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scott DA, Wright CE, Angus JA. Evidence that CB-1 and CB-2 cannabinoid receptors mediate antinociception in neuropathic pain in the rat. Pain. 2004;109:124–131. doi: 10.1016/j.pain.2004.01.020. [DOI] [PubMed] [Google Scholar]
- Seeley RJ, Woods SC. Monitoring of stored and available fuel by the CNS: implications for obesity. Nat Neurosci. 2003;4:901–909. doi: 10.1038/nrn1245. [DOI] [PubMed] [Google Scholar]
- Selley DE, Rorrer WK, Breivogel CS, Zimmer AM, Zimmer A, Martin BR, Sim-Selley LJ. Agonist efficacy and receptor efficiency in heterozygous CB1 knockout mice: relationship of reduced CB1 receptor density to G-protein activation. J Neurochem. 2001;77:1048–1057. doi: 10.1046/j.1471-4159.2001.00308.x. [DOI] [PubMed] [Google Scholar]
- Semple DM, McIntosh AM, Lawrie SM. Cannabis as a risk factor for psychosis: systematic review. J Psychopharmacol. 2005;19:187–194. doi: 10.1177/0269881105049040. [DOI] [PubMed] [Google Scholar]
- Shakespeare DT, Boggild M, Young C. Anti-spasticity agents for multiple sclerosis. Cochrane Database Syst Rev. 2003;4:CD001332. doi: 10.1002/14651858.CD001332. [DOI] [PubMed] [Google Scholar]
- Shapiro BJ, Tashkin DP, Vachon L. Tetrahydrocannabinol as a bronchodilator: Why bother. Chest. 1977;71:558–560. doi: 10.1378/chest.71.4.558b. [DOI] [PubMed] [Google Scholar]
- Shapiro D. The ocular manifestations of the cannabinols. Ophthalmologica. 1974;168:366–369. doi: 10.1159/000307060. [DOI] [PubMed] [Google Scholar]
- Sharkey KA, Pittman QJ. Central and peripheral signaling mechanisms involved in endocannabinoid regulation of feeding: a perspective on the munchies. SciSTKE. 2005;277:pe15. doi: 10.1126/stke.2772005pe15. [DOI] [PubMed] [Google Scholar]
- Shearman LP, Rosko KM, Fleischer R, Wang J, Xu S, Tong XS, Rocha BA. Antidepressant-like and anorectic effects of the cannabinoid CB1 receptor inverse agonist AM251 in mice. Behav Pharmacol. 2003;14:573–582. doi: 10.1097/00008877-200312000-00001. [DOI] [PubMed] [Google Scholar]
- Shen M, Thayer SA. Cannabinoid receptor agonists protect cultured rat hippocampal neurons from excitotoxicity. Mol Pharmacol. 1998;54:459–462. doi: 10.1124/mol.54.3.459. [DOI] [PubMed] [Google Scholar]
- Shen M, Thayer SA. Δ9-Tetrahydrocannabinol acts as a partial agonist to modulate glutamatergic synaptic transmission between rat hippocampal neurons in culture. Mol Pharmacol. 1999;55:8–13. doi: 10.1124/mol.55.1.8. [DOI] [PubMed] [Google Scholar]
- Shire D, Carillon C, Kaghad M, Calandra B, Rinaldi-Carmona M, Le Fur G, Caput D, Ferara P. An amino-terminal variant of the central cannabinoid receptor resulting from alternative splicing. J Biol Chem. 1995;270:3726–3731. doi: 10.1074/jbc.270.8.3726. [DOI] [PubMed] [Google Scholar]
- Shohami E, Gallily R, Mechoulam R, Bass R, Ben-Hur T. Cytokine production in the brain following closed head injury: dexanabinol (HU-211) is a novel TNF-α inhibitor and an effective neuroprotectant. J Neuroimmunol. 1997;72:169–177. doi: 10.1016/s0165-5728(96)00181-6. [DOI] [PubMed] [Google Scholar]
- Shohami E, Novikov M, Bass R. Long-term effect of HU-211, a novel non-competitive NMDA antagonist, on motor and memory functions after closed head injury in the rat. Brain Res. 1995;674:55–62. doi: 10.1016/0006-8993(94)01433-i. [DOI] [PubMed] [Google Scholar]
- Shohami E, Novikov M, Mechoulam R. A nonpsychotropic cannabinoid, HU-211, has cerebroprotective effects after closed head injury in the rat. J Neurotrauma. 1993;10:109–119. doi: 10.1089/neu.1993.10.109. [DOI] [PubMed] [Google Scholar]
- Shook JE, Burks TF. Psychoactive cannabinoids reduce gastrointestinal propulsion and motility in rodents. J Pharmacol Exp Ther. 1989;249:444–449. [PubMed] [Google Scholar]
- Shouman B, Fontaine RH, Baud O, Schwendimann L, Keller M, Spedding M, Lelievre V, Gressens P. Endocannabinoids potently protect the newborn brain against AMPA-kainate receptor-mediated excitotoxic damage. Br J Pharmacol. 2006;148:442–451. doi: 10.1038/sj.bjp.0706755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Showalter VM, Compton DR, Martin BR, Abood ME. Evaluation of binding in a transfected cell line expressing a peripheral cannabinoid receptor (CB2): identification of cannabinoid receptor subtype selective ligands. J Pharmacol Exp Ther. 1996;278:989–999. [PubMed] [Google Scholar]
- Sidney S, Quesenberry CP, Jr, Friedman GD, Tekawa IS. Marijuana use and cancer incidence (California, United States) Cancer Causes Control. 1997;8:722–728. doi: 10.1023/a:1018427320658. [DOI] [PubMed] [Google Scholar]
- Siegfried Z, Kanyas K, Latzer Y, Karni O, Bloch M, Lerer B, Berry EM. Association study of cannabinoid receptor gene (CNR1) alleles and anorexia nervosa: differences between restricting and bingeing/purging subtypes. Am J Med Genetics. 2004;125B:126–130. doi: 10.1002/ajmg.b.20089. [DOI] [PubMed] [Google Scholar]
- Siegmund SV, Uchinami H, Osawa Y, Brenner DA, Schwabe RF. Anandamide induces necrosis in primary hepatic stellate cells. Hepatology. 2005;41:1085–1095. doi: 10.1002/hep.20667. [DOI] [PubMed] [Google Scholar]
- Sieradzan KA, Fox SH, Hill M, Dick JP, Crossman AR, Brotchie JM. Cannabinoids reduce levodopa-induced dyskinesia in Parkinson’s disease: a pilot study. Neurology. 2001;57:2108–2111. doi: 10.1212/wnl.57.11.2108. [DOI] [PubMed] [Google Scholar]
- Sieradzan KA, Mann DM. The selective vulnerability of nerve cells in Huntington’s disease. Neuropathol Appl Neurobiol. 2001;27:1–21. doi: 10.1046/j.0305-1846.2001.00299.x. [DOI] [PubMed] [Google Scholar]
- Silber MH. Clinical practice: chronic insomnia. N Engl J Med. 2005;353:803–810. doi: 10.1056/NEJMcp043762. [DOI] [PubMed] [Google Scholar]
- Silverdale MA, McGuire S, McInnes A, Crossman AR, Brotchie JM. Striatal cannabinoid CB1 receptor mRNA expression is decreased in the reserpine-treated rat model of Parkinson’s disease. Exp Neurol. 2001;169:400–406. doi: 10.1006/exnr.2001.7649. [DOI] [PubMed] [Google Scholar]
- Simiand J, Keane M, Keane PE, Soubrié P. SR 141716, a CB1 cannabinoid receptor antagonist, selectively reduces sweet food intake in marmoset. Behav Pharmacol. 1998;9:179–181. [PubMed] [Google Scholar]
- Simons-Morton DG, Obarzanek E, Cutler JA. Obesity research—limitations of methods, measurements, and medications. J Am Med Assoc. 2006;295:826–828. doi: 10.1001/jama.295.7.826. [DOI] [PubMed] [Google Scholar]
- Sinor AD, Irvin SM, Greenberg DA. Endocannabinoids protect cerebral cortical neurons from in vitro ischemia in rats. Neurosci Lett. 2000;278:157–160. doi: 10.1016/s0304-3940(99)00922-2. [DOI] [PubMed] [Google Scholar]
- Sipe JC, Chiang K, Gerber AL, Beutler E, Cravatt BF. A missense mutation in human fatty acid amide hydrolase associated with problem drug use. Proc Natl Acad Sci USA. 2002;99:8394–8399. doi: 10.1073/pnas.082235799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sipe JC, Waalen J, Gerber A, Beutler E. Overweight and obesity associated with a missense polymorphism in fatty acid amide hydrolase (FAAH) Int J Obes. 2005;29:755–759. doi: 10.1038/sj.ijo.0802954. [DOI] [PubMed] [Google Scholar]
- Sirven JI, Berg AT. Marijuana as a treatment for epilepsy and multiple sclerosis? A “grass roots” movement. Neurology. 2004;62:1924–1925. doi: 10.1212/01.wnl.0000129703.59857.d6. [DOI] [PubMed] [Google Scholar]
- Skaper SD, Buriani A, Dal Toso R, Petrelli L, Romanello S, Facci L, Leon A. The ALIAmide palmitoylethanolamide and cannabinoids, but not anandamide, are protective in a delayed postglutamate paradigm of excitotoxic death in cerebellar granule neurons. Proc Natl Acad Sci USA. 1996;93:3984–3989. doi: 10.1073/pnas.93.9.3984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith CG, Asch RH. Drug abuse and reproduction. Fertil Steril. 1987;48:355–373. doi: 10.1016/s0015-0282(16)59400-x. [DOI] [PubMed] [Google Scholar]
- Smith FL, Fujimori K, Lowe J, Welch SP. Characterization of Δ9-tetrahydrocannabinol and anandamide antinociception in nonarthritic and arthritic rats. Pharmacol Biochem Behav. 1998;60:183–191. doi: 10.1016/s0091-3057(97)00583-2. [DOI] [PubMed] [Google Scholar]
- Smith PB, Compton DR, Welch SP, Razdan RK, Mechoulam R, Martin BR. The pharmacological activity of anandamide, a putative endogenous cannabinoid, in mice. J Pharmacol Exp Ther. 1994;270:219–227. [PubMed] [Google Scholar]
- Smith PF. GW-1000: GW pharmaceuticals. Curr Opin Investig Drugs. 2004;5:748–754. [PubMed] [Google Scholar]
- Smith PF. The safety of cannabinoids for the treatment of multiple sclerosis. Expert Opin Drug Saf. 2005;4:443–456. doi: 10.1517/14740338.4.3.443. [DOI] [PubMed] [Google Scholar]
- Smith SR, Terminelli C, Denhardt G. Effects of cannabinoid receptor agonist and antagonist ligands on production of inflammatory cytokines and anti-inflammatory interleukin-10 in endotoxemic mice. J Pharmacol Exp Ther. 2000;293:136–150. [PubMed] [Google Scholar]
- Smith SR, Terminelli C, Denhardt G. Modulation of cytokine responses in Corynebacterium parvum-primed endotoxemic mice by centrally administered cannabinoid ligands. Eur J Pharmacol. 2001;425:73–83. doi: 10.1016/s0014-2999(01)01142-6. [DOI] [PubMed] [Google Scholar]
- Sofia RD, Diamantis W, Harrison JE, Melton J. Evaluation of antiulcer activity of Δ9-tetrahydrocannabinol in the Shay rat test. Pharmacology. 1978;17:173–177. doi: 10.1159/000136851. [DOI] [PubMed] [Google Scholar]
- Sofia RD, Nalepa SD, Harakal JJ, Vassar HB. Anti-edema and analgesic properties of Δ9-tetrahydrocannabinol (THC) J Pharmacol Exp Ther. 1973;186:646–655. [PubMed] [Google Scholar]
- Solinas M, Goldberg SR. Motivational effects of cannabinoids and opioids on food reinforcement depend on simultaneous activation of cannabinoid and opioid systems. Neuropsychopharmacology. 2005;30:2035–2045. doi: 10.1038/sj.npp.1300720. [DOI] [PubMed] [Google Scholar]
- Song ZH, Bonner TI. A lysine residue of the cannabinoid receptor is critical for receptor recognition by several agonists bu not WIN55212-2. Mol Pharmacol. 1996;49:891–896. [PubMed] [Google Scholar]
- Song ZH, Slowey CA. Involvement of cannabinoid receptors in the intraocular pressure-lowering effects of WIN55212-2. J Pharmacol Exp Ther. 2000;292:136–139. [PubMed] [Google Scholar]
- Soria G, Mendizabal V, Tourino C, Robledo P, Ledent C, Parmentier M, Maldonado R, Valverde O. Lack of CB1 cannabinoid receptor impairs cocaine self-administration. Neuropsychopharmacology. 2005;30:1670–1680. doi: 10.1038/sj.npp.1300707. [DOI] [PubMed] [Google Scholar]
- Sospedra M, Martin R. Immunology of multiple sclerosis. Annu Rev Immunol. 2005;23:683–747. doi: 10.1146/annurev.immunol.23.021704.115707. [DOI] [PubMed] [Google Scholar]
- Specter S, Lancz G, Westrich G, Friedman H. Δ-9-Tetrahydrocannabinol augments murine retroviral induced immunosuppression and infection. Int J Immunopharmacol. 1991;13:411–417. doi: 10.1016/0192-0561(91)90011-u. [DOI] [PubMed] [Google Scholar]
- Spencer DJ. Cannabis-induced psychosis. Int J Addict. 1971;6:323–326. doi: 10.3109/10826087109057790. [DOI] [PubMed] [Google Scholar]
- Spicuzza L, Haddad EB, Birrell M, Ling A, Clarke D, Venkatesan P, Barnes PJ, Belvisi MG. Characterization of the effects of cannabinoids on guinea-pig tracheal smooth muscle tone: role in the modulation of acetylcholine release from parasympathetic nerves. Br J Pharmacol. 2000;130:1720–1726. doi: 10.1038/sj.bjp.0703497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Srivastava MD, Srivastava BI, Brouhard B. Δ9 Tetrahydrocannabinol and cannabidiol alter cytokine production by human immune cells. Immunopharmacology. 1998;40:179–185. doi: 10.1016/s0162-3109(98)00041-1. [DOI] [PubMed] [Google Scholar]
- Stamer WD, Golightly SF, Hosohata Y, Ryan EP, Porter AC, Varga E, Noecker RJ, Felder CC, Yamamura HI. Cannabinoid CB1 receptor expression, activation and detection of endogenous ligand in trabecular meshwork and ciliary process tissues. Eur J Pharmacol. 2001;431:277–286. doi: 10.1016/s0014-2999(01)01438-8. [DOI] [PubMed] [Google Scholar]
- Staquet M, Gantt C, Machin D. Effect of a nitrogen analog of tetrahydrocannabinol on cancer pain. Clin Pharmacol Ther. 1978;23:397–401. doi: 10.1002/cpt1978234397. [DOI] [PubMed] [Google Scholar]
- Steffens M, Feuerstein TJ. Receptor-independent depression of DA and 5-HT uptake by cannabinoids in rat neocortex–involvement of Na+/K+-ATPase. Neurochem Int. 2004;44:529–538. doi: 10.1016/j.neuint.2003.08.009. [DOI] [PubMed] [Google Scholar]
- Steffens S, Veillard NR, Arnaud C, Pelli G, Burger F, Staub C, Karsak M, Zimmer A, Frossard JL, Mach F. Low dose oral cannabinoid therapy reduces progression of atherosclerosis in mice. Nature (Lond) 2005;434:782–786. doi: 10.1038/nature03389. [DOI] [PubMed] [Google Scholar]
- Stein EA, Fuller SA, Edgemond WS, Campbell WB. Physiological and behavioural effects of the endogenous cannabinoid, arachidonylethanolamide (anandamide), in the rat. Br J Pharmacol. 1996;119:107–114. doi: 10.1111/j.1476-5381.1996.tb15683.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stella N. Cannabinoid signaling in glial cells. Glia. 2004;48:267–277. doi: 10.1002/glia.20084. [DOI] [PubMed] [Google Scholar]
- Stella N, Schweitzer P, Piomelli D. A second endogenous cannabinoid that modulates long-term potentiation. Nature (Lond) 1997;388:773–778. doi: 10.1038/42015. [DOI] [PubMed] [Google Scholar]
- Stengel PW, Rippy MK, Cockerham SL, Devane WA, Silbaugh SA. Pulmonary actions of anandamide, an endogenous cannabinoid receptor agonist, in guinea pigs. Eur J Pharmacol. 1998;355:57–66. doi: 10.1016/s0014-2999(98)00472-5. [DOI] [PubMed] [Google Scholar]
- Sterin-Borda L, Del Zar CF, Borda E. Differential CB1 and CB2 cannabinoid receptor-inotropic response of rat isolated atria: endogenous signal transduction pathways. Biochem Pharmacol. 2005;69:1705–1713. doi: 10.1016/j.bcp.2005.03.027. [DOI] [PubMed] [Google Scholar]
- Storr M, Gaffal E, Saur D, Schusdziarra V, Allescher HD. Effect of cannabinoids on neural transmission in rat gastric fundus. Can J Physiol Pharmacol. 2002;80:67–76. doi: 10.1139/y02-005. [DOI] [PubMed] [Google Scholar]
- Storr M, Sibaev A, Marsicano G, Lutz B, Schusdziarra V, Timmermans JP, Allescher HD. Cannabinoid receptor type 1 modulates excitatory and inhibitory neurotransmission in mouse colon. Am J Physiol. 2004;286:G110–G117. doi: 10.1152/ajpgi.00148.2003. [DOI] [PubMed] [Google Scholar]
- Straiker A, Stella N, Piomelli D, Mackie K, Karten HJ, Maguire G. Cannabinoid CB1 receptors and ligands in vertebrate retina: localization and function of an endogenous signaling system. Proc Natl Acad Sci USA. 1999a;96:14565–14570. doi: 10.1073/pnas.96.25.14565. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Straiker AJ, Maguire G, Mackie K, Lindsey J. Localization of cannabinoid CB1 receptors in the human anterior eye and retina. Investig Ophthalmol Vis Sci. 1999b;40:2442–2448. [PubMed] [Google Scholar]
- Strangman NM, Patrick SL, Hohmann AG, Tsou K, Walker JM. Evidence for a role of endogenous cannabinoids in the modulation of acute and tonic pain sensitivity. Brain Res. 1998;813:323–328. doi: 10.1016/s0006-8993(98)01031-2. [DOI] [PubMed] [Google Scholar]
- Stumpff F, Boxberger M, Krauss A, Rosenthal R, Meissner S, Choritz L, Wiederholt M, Thieme H. Stimulation of cannabinoid (CB1) and prostanoid (EP2) receptors opens BKCa channels and relaxes ocular trabecular meshwork. Exp Eye Res. 2005;80:697–708. doi: 10.1016/j.exer.2004.12.003. [DOI] [PubMed] [Google Scholar]
- Sugiura T, Kodaka T, Nakane S, Miyashita T, Kondo S, Suhara Y, Takayama H, Waku K, Seki C, Baba N, et al. Evidence that the cannabinoid CB1 receptor is a 2-arachidonoylglycerol receptor: structure-activity relationship of 2-arachidonoylglycerol, ether-linked analogues, and related compounds. J Biol Chem. 1999;274:2794–2801. doi: 10.1074/jbc.274.5.2794. [DOI] [PubMed] [Google Scholar]
- Sugiura T, Kondo S, Sukagawa A, Nakane S, Shinoda A, Itoh K, Yamashita A, Waku K. 2-Arachidonoylglycerol: a possible endogenous cannabinoid receptor ligand in brain. Biochem Biophys Res Commun. 1995;215:89–97. doi: 10.1006/bbrc.1995.2437. [DOI] [PubMed] [Google Scholar]
- Sugiura T, Yoshinaga N, Kondo S, Waku K, Ishima Y. Generation of 2-arachidonoylglycerol, an endogenous cannabinoid receptor ligand, in picrotoxinin-administered rat brain. Biochem Biophys Res Commun. 2000;271:654–658. doi: 10.1006/bbrc.2000.2686. [DOI] [PubMed] [Google Scholar]
- Sugrue MF. New approaches to antiglaucoma therapy. J Med Chem. 1997;40:2793–2809. doi: 10.1021/jm9608725. [DOI] [PubMed] [Google Scholar]
- Sumariwalla PF, Gallily R, Tchilibon S, Fride E, Mechoulam R, Feldmann M. A novel synthetic, nonpsychoactive cannabinoid acid (HU-320) with anti-inflammatory properties in murine collagen-induced arthritis. Arthritis Rheum. 2004;50:985–998. doi: 10.1002/art.20050. [DOI] [PubMed] [Google Scholar]
- Sun YX, Tsuboi K, Okamoto Y, Tonai T, Murakami M, Kudo I, Ueda N. Biosynthesis of anandamide and N-palmitoylethanolamine by sequential actions of phospholipase A2 and lysophospholipase D. Biochem J. 2004;380:749–756. doi: 10.1042/BJ20040031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sun YX, Tsuboi K, Zhao LY, Okamoto Y, Lambert DM, Ueda N. Involvement of N-acylethanolamine-hydrolyzing acid amidase in the degradation of anandamide and other N-acylethanolamines in macrophages. Biochim Biophys Acta. 2005;1736:211–2120. doi: 10.1016/j.bbalip.2005.08.010. [DOI] [PubMed] [Google Scholar]
- Sundram S, Copolov D, Dean B. Clozapine decreases [3H] CP 55940 binding to the cannabinoid1 receptor in the rat nucleus accumbens. Naunyn-Schmiedeberg’s Arch Pharmacol. 2005;371:428–433. doi: 10.1007/s00210-005-1074-2. [DOI] [PubMed] [Google Scholar]
- Suplita RL, 2nd, Gutierrez T, Fegley D, Piomelli D, Hohmann AG. Inhibition of fatty-acid amide hydrolase enhances cannabinoid stress-induced analgesia: sites of action in the dorsolateral periaqueductal gray and rostral ventrome-dial medulla. Neuropharmacology. 2006;50:372–379. doi: 10.1016/j.neuropharm.2005.07.007. [DOI] [PubMed] [Google Scholar]
- Svendsen KB, Jensen TS, Bach FW. Does the cannabinoid dronabinol reduce central pain in multiple sclerosis? Randomised double blind placebo controlled crossover trial. BMJ. 2004;329:253. doi: 10.1136/bmj.38149.566979.AE. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Szabo B, Nordheim U, Niederhoffer N. Effects of cannabinoids on sympathetic and parasympathetic neuroeffector transmission in the rabbit heart. J Pharmacol Exp Ther. 2001;297:819–826. [PubMed] [Google Scholar]
- Szabo B, Siemes S, Wallmichrath I. Inhibition of GABAergic neurotransmission in the ventral tegmental area by cannabinoids. Eur J Neurosci. 2002;15:2057–2061. doi: 10.1046/j.1460-9568.2002.02041.x. [DOI] [PubMed] [Google Scholar]
- Tabarin A, Chaves YD, Carmona M del C, Catargi B, Zorrilla EP, Roberts AJ, Coscina DV, Rousset S, Redonnet A, Parker GC, et al. Resistance to diet-induced obesity in μ-opioid receptor-deficient mice: evidence for a “thrifty gene”. Diabetes. 2005;54:3510–3516. doi: 10.2337/diabetes.54.12.3510. [DOI] [PubMed] [Google Scholar]
- Takahashi KA, Castillo PE. The CB1 receptor mediates glutamatergic synaptic suppression in the hippocampus. Neuroscience. 2006;139:792–802. doi: 10.1016/j.neuroscience.2006.01.024. [DOI] [PubMed] [Google Scholar]
- Tanda G, Loddo P, Di Chiara G. Dependence of mesolimbic dopamine transmission on delta9-tetrahydrocannabinol. Eur J Pharmacol. 1999;376:23–26. doi: 10.1016/s0014-2999(99)00384-2. [DOI] [PubMed] [Google Scholar]
- Tanda G, Munzar P, Goldberg SR. Self-administration behavior is maintained by the psychoactive ingredient of marijuana in squirrel monkeys. Nat Neurosci. 2000;3:1073–1074. doi: 10.1038/80577. [DOI] [PubMed] [Google Scholar]
- Tashkin DP, Baldwin GC, Sarafian T, Dubinett S, Roth MD. Respiratory and immunologic consequences of marijuana smoking. J Clin Pharmacol. 2002;42:71S–81S. doi: 10.1002/j.1552-4604.2002.tb06006.x. [DOI] [PubMed] [Google Scholar]
- Tashkin DP, Reiss S, Shapiro BJ, Calvarese B, Olsen JL, Lodge JW. Bronchial effects of aerosolized Δ9-tetrahydrocannabinol in healthy and asthmatic subjects. Am Rev Respir Dis. 1977;115:57–65. doi: 10.1164/arrd.1977.115.1.57. [DOI] [PubMed] [Google Scholar]
- Tashkin DP, Shapiro BJ, Frank IM. Acute pulmonary physiologic effects of smoked marijuana and oral 9-tetrahydrocannabinol in healthy young men. N Engl J Med. 1973;289:336–341. doi: 10.1056/NEJM197308162890702. [DOI] [PubMed] [Google Scholar]
- Tashkin DP, Shapiro BJ, Frank IM. Acute effects of smoked marijuana and oral Δ9-tetrahydrocannabinol on specific airway conductance in asthmatic subjects. Am Rev Respir Dis. 1974;109:420–428. doi: 10.1164/arrd.1974.109.4.420. [DOI] [PubMed] [Google Scholar]
- Tashkin DP, Shapiro BJ, Lee YE, Harper CE. Effects of smoked marijuana in experimentally induced asthma. Am Rev Respir Dis. 1975;112:377–386. doi: 10.1164/arrd.1975.112.3.377. [DOI] [PubMed] [Google Scholar]
- Taylor FM., 3rd Marijuana as a potential respiratory tract carcinogen: a retrospective analysis of a community hospital population. South Med J. 1988;81:1213–1216. doi: 10.1097/00007611-198810000-00003. [DOI] [PubMed] [Google Scholar]
- Teichner A, Ovadia H, Lavie G, Leker RR. Combination of dexanabinol and tempol in focal cerebral ischemia: is there a ceiling effect? Exp Neurol. 2003;182:353–360. doi: 10.1016/s0014-4886(03)00083-9. [DOI] [PubMed] [Google Scholar]
- Teixeira-Clerc F, Julien B, Grenard P, Tran Van Nhieu J, Deveaux V, Li L, Serriere-Lanneau V, Ledent C, Mallat A, Lotersztajn S. CB1 cannabinoid receptor antagonism: a new strategy for the treatment of liver fibrosis. Nat Med. 2006;12:671–676. doi: 10.1038/nm1421. [DOI] [PubMed] [Google Scholar]
- Ten Ham M, Loskota WJ, Lomax P. Acute and chronic effects of β9-tetrahydrocannabinol on seizures in the gerbil. Eur J Pharmacol. 1975;31:148–152. doi: 10.1016/0014-2999(75)90087-4. [DOI] [PubMed] [Google Scholar]
- Thaker GK, Carpenter WT., Jr Advances in schizophrenia. Nat Med. 2001;7:667–671. doi: 10.1038/89040. [DOI] [PubMed] [Google Scholar]
- Tham SM, Angus JA, Tudor EM, Wright CE. Synergistic and additive interactions of the cannabinoid agonist CP55,940 with μ opioid receptor and α2-adrenoceptor agonists in acute pain models in mice. Br J Pharmacol. 2005;144:875–884. doi: 10.1038/sj.bjp.0706045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thanos PK, Dimitrakakis ES, Rice O, Gifford A, Volkow ND. Ethanol self-administration and ethanol conditioned place preference are reduced in mice lacking cannabinoid CB1 receptors. Behav Brain Res. 2005;164:206–213. doi: 10.1016/j.bbr.2005.06.021. [DOI] [PubMed] [Google Scholar]
- Thiele TE, Marsh DJ, Ste Marie L, Bernstein IL, Palmiter RD. Ethanol consumption and resistance are inversely related to neuropeptide Y levels. Nature (Lond) 1998;396:366–369. doi: 10.1038/24614. [DOI] [PubMed] [Google Scholar]
- Thomas A, Stevenson LA, Wease KN, Price MR, Baillie G, Ross RA, Pertwee RG. Evidence that the plant cannabinoid Δ9-tetrahydrocannabivarin is a cannabinoid CB1 and CB2 receptor antagonist. Br J Pharmacol. 2005;146:917–926. doi: 10.1038/sj.bjp.0706414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thomas EA, Carson MJ, Neal MJ, Sutcliffe JG. Unique allosteric regulation of 5-hydroxytryptamine receptor-mediated signal transduction by oleamide. Proc Natl Acad Sci USA. 1997;94:14115–14119. doi: 10.1073/pnas.94.25.14115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thomas EA, Cravatt BF, Sutcliffe JG. The endogenous lipid oleamide activates serotonin 5-HT7 neurons in mouse thalamus and hypothalamus. J Neurochem. 1999;72:2370–2378. doi: 10.1046/j.1471-4159.1999.0722370.x. [DOI] [PubMed] [Google Scholar]
- Thomas MJ, Beurrier C, Bonci A, Malenka RC. Long-term depression in the nucleus accumbens: a neural correlate of behavioral sensitization to cocaine. Nat Neurosc. 2001;4:1217–1223. doi: 10.1038/nn757. [DOI] [PubMed] [Google Scholar]
- Thompson AJ, Baker D. Cannabinoids in MS: potentially useful but not just yet! Neurology. 2002;58:1323–1324. doi: 10.1212/wnl.58.9.1323. [DOI] [PubMed] [Google Scholar]
- Thornton-Jones ZD, Vickers SP, Clifton PG. The cannabinoid CB1 receptor antagonist SR141716A reduces appetitive and consummatory responses to food. Psychopharmacology. 2005;179:452–460. doi: 10.1007/s00213-004-2047-8. [DOI] [PubMed] [Google Scholar]
- Timpone JG, Wright DJ, Li N, Egorin MJ, Enama ME, Mayers J, Galetto G. The safety and pharmacokinetics of single-agent and combination therapy with megestrol acetate and dronabinol for the treatment of HIV wasting syndrome: the DATRI 004 Study Group. Division of AIDS Treatment Research Initiative. AIDS Res Hum Retroviruses. 1997;13:305–315. doi: 10.1089/aid.1997.13.305. [DOI] [PubMed] [Google Scholar]
- Tomida I, Pertwee RG, Azuara-Blanco A. Cannabinoids and glaucoma. Br J Ophthalmol. 2004;88:708–713. doi: 10.1136/bjo.2003.032250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tournier M, Sorbara F, Gindre C, Swendsen JD, Verdoux H. Cannabis use and anxiety in daily life: a naturalistic investigation in a non-clinical population. Psychiatry Res. 2003;118:1–8. doi: 10.1016/s0165-1781(03)00052-0. [DOI] [PubMed] [Google Scholar]
- Tramèr MR, Carroll D, Campbell FA, Reynolds DJM, Moore RA, McQuay HJ. Cannabinoids for control of chemotherapy induced nausea and vomiting: quantitative systematic review. Br Med J. 2001;323:1–8. doi: 10.1136/bmj.323.7303.16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tranguch S, Daikoku T, Guo Y, Wang H, Dey SK. Molecular complexity in establishing uterine receptivity and implantation. Cell Mol Life Sci. 2005;62:1964–1973. doi: 10.1007/s00018-005-5230-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Treffert DA. Marijuana use in schizophrenia: a clear hazard. Am J Psychiatry. 1978;135:1213–1215. doi: 10.1176/ajp.135.10.1213. [DOI] [PubMed] [Google Scholar]
- Tsou K, Brown S, Sanudo-Pena MC, Mackie K, Walker JM. Immunohistochemical distribution of cannabinoid CB1 receptors in the rat central nervous system. Neuroscience. 1998;83:393–411. doi: 10.1016/s0306-4522(97)00436-3. [DOI] [PubMed] [Google Scholar]
- Tsou K, Lowitz KA, Hohmann AG, Martin WJ, Hathaway CB, Bereiter DA, Walker JM. Suppression of noxious stimulus-evoked expression of Fos protein-like immunoreactivity in rat spinal cord by a selective cannabinoid agonist. Neuroscience. 1996;70:791–798. doi: 10.1016/s0306-4522(96)83015-6. [DOI] [PubMed] [Google Scholar]
- Tsou K, Mackie K, Sanudo-Pena MC, Walker JM. Cannabinoid CB1 receptors are localized primarily on cholecystokinin-containing GABAergic inter-neurons in the rat hippocampal formation. Neuroscience. 1999;93:969–975. doi: 10.1016/s0306-4522(99)00086-x. [DOI] [PubMed] [Google Scholar]
- Tsuboi K, Sun YX, Okamoto Y, Araki N, Tonai T, Ueda N. Molecular characterization of N-acylethanolamine-hydrolyzing acid amidase, a novel member of the choloylglycine hydrolase family with structural and functional similarity to acid ceramidase. J Biol Chem. 2005;280:11082–11092. doi: 10.1074/jbc.M413473200. [DOI] [PubMed] [Google Scholar]
- Tucci SA, Rogers EK, Korbonits M, Kirkham TC. The cannabinoid CB1 receptor antagonist SR141716 blocks the orexigenic effects of intrahypothalamic ghrelin. Br J Pharmacol. 2004;143:520–523. doi: 10.1038/sj.bjp.0705968. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turner WM, Tsuang MT. Impact of substance abuse on the course and outcome of schizophrenia. Schizophr Bull. 1990;16:87–95. doi: 10.1093/schbul/16.1.87. [DOI] [PubMed] [Google Scholar]
- Twelves D, Perkins KS, Counsell C. Systematic review of incidence studies of Parkinson’s disease. Mov Disord. 2003;18:19–31. doi: 10.1002/mds.10305. [DOI] [PubMed] [Google Scholar]
- Tzavara ET, Wade M, Nomikos GG. Biphasic effects of cannabinoids on acetylcholine release in the hippocampus: site and mechanism of action. J Neurosci. 2003;23:9374–9384. doi: 10.1523/JNEUROSCI.23-28-09374.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ueda N, Kurahashi Y, Yamamoto S, Tokunaga T. Partial purification and characterization of the porcine brain enzyme hydrolyzing and synthesizing anandamide. J Biol Chem. 1995;270:23823–23827. doi: 10.1074/jbc.270.40.23823. [DOI] [PubMed] [Google Scholar]
- Ugdyzhekova DS, Krylatov AV, Bernatskaya NA, Maslov LN, Mechoulam R, Pertwee RG. Activation of cannabinoid receptors decreases the area of ischemic myocardial necrosis. Bull Exp Biol Med. 2002;133:125–126. doi: 10.1023/a:1015526217332. [DOI] [PubMed] [Google Scholar]
- Ugdyzhekova DS, Maslov LN, Krylatov AV, Lishmanov IuB, Tam SV. Specificity of the anti-arrhythmic effect of κ1-opioid receptor agonists. Eksp Klin Farmakol. 2001;64:17–20. [PubMed] [Google Scholar]
- Ujike H, Morita Y. New perspectives in the studies on endocannabinoid and cannabis: cannabinoid receptors and schizophrenia. J Pharmacol Sci. 2004;96:376–381. doi: 10.1254/jphs.fmj04003x4. [DOI] [PubMed] [Google Scholar]
- Ujike H, Takaki M, Nakata K, Tanaka Y, Takeda T, Kodama M, Fujiwara Y, Sakai A, Kuroda S. CNR1, central cannabinoid receptor gene, associated with susceptibility to hebephrenic schizophrenia. Mol Psychiatry. 2002;7:515–518. doi: 10.1038/sj.mp.4001029. [DOI] [PubMed] [Google Scholar]
- Ulugol A, Ozyigit F, Yesilyurt O, Dogrul A. The additive antinociceptive interaction between WIN 55,212-2, a cannabinoid agonist, and ketorolac. Anesth Analg. 2006;102:443–447. doi: 10.1213/01.ane.0000194587.94260.1d. [DOI] [PubMed] [Google Scholar]
- Underdown NJ, Hiley CR, Ford WR. Anandamide reduces infarct size in rat isolated hearts subjected to ischaemia-reperfusion by a novel cannabinoid mechanism. Br J Pharmacol. 2005;146:809–816. doi: 10.1038/sj.bjp.0706391. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ungerleider JT, Andyrsiak T, Fairbanks L, Ellison GW, Myers LW. Δ-9-THC in the treatment of spasticity associated with multiple sclerosis. Adv Alcohol Subst Abuse. 1987;7:39–50. doi: 10.1300/j251v07n01_04. [DOI] [PubMed] [Google Scholar]
- Urigüen L, Perez-Rial S, Ledent C, Palomo T, Manzanares J. Impaired action of anxiolytic drugs in mice deficient in cannabinoid CB1 receptors. Neuropharmacology. 2004;46:966–973. doi: 10.1016/j.neuropharm.2004.01.003. [DOI] [PubMed] [Google Scholar]
- Vaccani A, Massi P, Colombo A, Rubino T, Parolaro D. Cannabidiol inhibits human glioma cell migration through a cannabinoid receptor-independent mechanism. Br J Pharmacol. 2005;144:1032–1036. doi: 10.1038/sj.bjp.0706134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vachon L, FitzGerald MX, Solliday NH, Gould IA, Gaensler EA. Single-dose effects of marihuana smoke: bronchial dynamics and respiratory-center sensitivity in normal subjects. N Engl J Med. 1973;288:985–989. doi: 10.1056/NEJM197305102881902. [DOI] [PubMed] [Google Scholar]
- Valjent E, Maldonado R. A behavioural model to reveal place preference to Δ9-tetrahydrocannabinol in mice. Psychopharmacology. 2000;147:436–438. doi: 10.1007/s002130050013. [DOI] [PubMed] [Google Scholar]
- Valverde O, Ledent C, Beslot F, Parmentier M, Roques BP. Reduction of stress-induced analgesia but not of exogenous opioid effects in mice lacking CB1 receptors. Eur J Neurosci. 2000;12:533–539. doi: 10.1046/j.1460-9568.2000.00929.x. [DOI] [PubMed] [Google Scholar]
- van der Stelt M, Di Marzo V. The endocannabinoid system in the basal ganglia and in the mesolimbic reward system: implications for neurological and psychiatric disorders. Eur J Pharmacol. 2003;480:133–150. doi: 10.1016/j.ejphar.2003.08.101. [DOI] [PubMed] [Google Scholar]
- van der Stelt M, Di Marzo V. Endovanilloids: putative endogenous ligands of transient receptor potential vanilloid 1 channels. Eur J Biochem. 2004;271:1827–1834. doi: 10.1111/j.1432-1033.2004.04081.x. [DOI] [PubMed] [Google Scholar]
- van der Stelt M, Fox SH, Hill M, Crossman AR, Petrosino S, Di Marzo V, Brotchie JM. A role for endocannabinoids in the generation of parkinsonism and levodopa-induced dyskinesia in MPTP-lesioned non-human primate models of Parkinson’s disease. FASEB J. 2005;19:1140–1142. doi: 10.1096/fj.04-3010fje. [DOI] [PubMed] [Google Scholar]
- van der Stelt M, Veldhuis WB, Bar PR, Veldink GA, Vliegenthart JF, Nicolay K. Neuroprotection by Δ9-tetrahydrocannabinol, the main active compound in marijuana, against ouabain-induced in vivo excitotoxicity. J Neurosci. 2001a;21:6475–6479. doi: 10.1523/JNEUROSCI.21-17-06475.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van der Stelt M, Veldhuis WB, van Haaften GW, Fezza F, Bisogno T, Bar PR, Veldink GA, Vliegenthart JF, Di Marzo V, Nicolay K. Exogenous anandamide protects rat brain against acute neuronal injury in vivo. J Neurosci. 2001b;21:8765–8771. doi: 10.1523/JNEUROSCI.21-22-08765.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vaney C, Heinzel-Gutenbrunner M, Jobin P, Tschopp F, Gattlen B, Hagen U, Schnelle M, Reif M. Efficacy, safety and tolerability of an orally administered cannabis extract in the treatment of spasticity in patients with multiple sclerosis: a randomized, double-blind, placebo-controlled, crossover study. Mult Scler. 2004;10:417–424. doi: 10.1191/1352458504ms1048oa. [DOI] [PubMed] [Google Scholar]
- Van Gaal LF, Rissanen AM, Scheen AJ, Ziegler O, Rossner S for the RIO-Europe Study Group. Effects of the cannabinoid-1 receptor blocker rimonabant on weight reduction and cardiovascular risk factors in overweight patients: 1-year experience from the RIO-Europe study. Lancet. 2005;365:1389–1397. doi: 10.1016/S0140-6736(05)66374-X. [DOI] [PubMed] [Google Scholar]
- van Oosten BW, Killestein J, Mathus-Vliegen EM, Polman CH. Multiple sclerosis following treatment with a cannabinoid receptor-1 antagonist. Mult Scler. 2004;10:330–331. doi: 10.1191/1352458504ms1044oa. [DOI] [PubMed] [Google Scholar]
- Van Sickle MD, Duncan M, Kingsley PJ, Mouihate A, Urbani P, Mackie K, Stella N, Makriyannis A, Piomelli D, Davison JS, et al. Identification and functional characterization of brainstem cannabinoid CB2 receptors. Science (Wash DC) 2005;310:329–332. doi: 10.1126/science.1115740. [DOI] [PubMed] [Google Scholar]
- Varga K, Lake K, Martin BR, Kunos G. Novel antagonist implicates the CB1 cannabinoid receptor in the hypotensive action of anandamide. Eur J Pharmacol. 1995;278:279–283. doi: 10.1016/0014-2999(95)00181-j. [DOI] [PubMed] [Google Scholar]
- Varga K, Lake KD, Huangfu D, Guyenet PG, Kunos G. Mechanism of the hypotensive action of anandamide in anesthetized rats. Hypertension. 1996;28:682–686. doi: 10.1161/01.hyp.28.4.682. [DOI] [PubMed] [Google Scholar]
- Varga K, Wagner JA, Bridgen DT, Kunos G. Platelet- and macrophage-derived endogenous cannabinoids are involved in endotoxin-induced hypotension. FASEB J. 1998;12:1035–1044. doi: 10.1096/fasebj.12.11.1035. [DOI] [PubMed] [Google Scholar]
- Varma DR, Goldbaum D. Effect of Δ9-tetrahydrocannabinol on experimental hypertension in rats. J Pharm Pharmacol. 1975;27:790–791. doi: 10.1111/j.2042-7158.1975.tb09406.x. [DOI] [PubMed] [Google Scholar]
- Varma N, Carlson GC, Ledent C, Alger BE. Metabotropic glutamate receptors drive the endocannabinoid system in the hippocampus. J Neurosci. 2001;21:RC188. doi: 10.1523/JNEUROSCI.21-24-j0003.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Velasco G, Galve-Roperh I, Sanchez C, Blazquez C, Guzman M. Hypothesis: cannabinoid therapy for the treatment of gliomas. Neuropharmacology. 2004;47:315–323. doi: 10.1016/j.neuropharm.2004.04.016. [DOI] [PubMed] [Google Scholar]
- Venance L, Piomelli D, Glowinski J, Giaume C. Inhibition by anandamide of gap junctions and intercellular calcium signalling in striatal astrocytes. Nature (Lond) 1995;376:590–594. doi: 10.1038/376590a0. [DOI] [PubMed] [Google Scholar]
- Vered M, Bar-Joseph A, Belayev L, Berkovich Y, Biegon A. Anti-ischemia activity of HU-211, a non-psychotropic synthetic cannabinoid. Acta Neurochir Suppl. 1994;60:335–337. doi: 10.1007/978-3-7091-9334-1_90. [DOI] [PubMed] [Google Scholar]
- Verty ANA, McFarlane JR, McGregor IS, Mallet PE. Evidence for an interaction between CB1 cannabinoid and melanocortin NCR-4 receptors in regulating food intake. Endocrinology. 2004;145:3224–3231. doi: 10.1210/en.2004-0059. [DOI] [PubMed] [Google Scholar]
- Verty ANA, Singh ME, McGregor IS, Mallet PE. The cannabinoid receptor antagonist SR 141716 attenuates overfeeding induced by systemic or intracranial morphine. Psychopharmacology. 2003;168:314–323. doi: 10.1007/s00213-003-1451-9. [DOI] [PubMed] [Google Scholar]
- Vickers SP, Kennett GA. Cannabinoids and the regulation of ingestive behavior. Curr Drug Targets. 2005;6:215–223. doi: 10.2174/1389450053174514. [DOI] [PubMed] [Google Scholar]
- Vickers SP, Webster LJ, Wyatt A, Dourish CT, Kennett GA. Preferential effects of the cannabinoid CB1 receptor antagonist, SR 141716, on food intake and body weight gain of obese (fa/fa) compared to lean Zucker rats. Psychopharmacology. 2003;167:103–111. doi: 10.1007/s00213-002-1384-8. [DOI] [PubMed] [Google Scholar]
- Vidrio H, Sanchez-Salvatori MA, Medina M. Cardiovascular effects of (−)-11-OH-delta 8-tetrahydrocannabinol-dimethylheptyl in rats. J Cardiovasc Pharmacol. 1996;28:332–336. doi: 10.1097/00005344-199608000-00022. [DOI] [PubMed] [Google Scholar]
- Vigano D, Rubino T, Parolaro D. Molecular and cellular basis of cannabinoid and opioid interactions. Pharmacol Biochem Behav. 2005a;81:360–368. doi: 10.1016/j.pbb.2005.01.021. [DOI] [PubMed] [Google Scholar]
- Vigano D, Rubino T, Vaccani A, Bianchessi S, Marmorato P, Castiglioni C, Parolaro D. Molecular mechanisms involved in the asymmetric interaction between cannabinoid and opioid systems. Psychopharmacology. 2005b;182:527–536. doi: 10.1007/s00213-005-0114-4. [DOI] [PubMed] [Google Scholar]
- Vinod KY, Hungund BL. Endocannabinoid lipids and mediated system: Implications for alcoholism and neuropsychiatric disorders. Life Sci. 2005;77:1569–1583. doi: 10.1016/j.lfs.2005.05.041. [DOI] [PubMed] [Google Scholar]
- Viveros MP, Marco EM, File SE. Endocannabinoid system and stress and anxiety responses. Pharmacol Biochem Behav. 2005;81:331–342. doi: 10.1016/j.pbb.2005.01.029. [DOI] [PubMed] [Google Scholar]
- Vizi ES, Katona I, Freund TF. Evidence for presynaptic cannabinoid CB1 receptor-mediated inhibition of noradrenaline release in the guinea pig lung. Eur J Pharmacol. 2001;431:237–244. doi: 10.1016/s0014-2999(01)01413-3. [DOI] [PubMed] [Google Scholar]
- Vlachou S, Nomikos GG, Panagis G. WIN 55,212-2 decreases the reinforcing actions of cocaine through CB1 cannabinoid receptor stimulation. Behav Brain Res. 2003;141:215–222. doi: 10.1016/s0166-4328(02)00370-4. [DOI] [PubMed] [Google Scholar]
- Volicer L, Stelly M, Morris J, McLaughlin J, Volicer BJ. Effects of dronabinol on anorexia and disturbed behavior in patients with Alzheimer’s disease. J Geriat Psychiatry. 1997;12:913–919. [PubMed] [Google Scholar]
- Volkow ND, Wang G-J, Fowler JS, Logan J, Gatley SJ, Gifford A, Hitzemann R, Ding YS, Pappas N. Prediction of reinforcing responses to psychostimulants in humans by brain dopamine D2 receptor levels. Am J Psychiatry. 1999;156:1440–1443. doi: 10.1176/ajp.156.9.1440. [DOI] [PubMed] [Google Scholar]
- Volkow ND, Wang G-J, Fowler JS, Logan J, Gatley SJ, Hitzemann R, Chen AD, Dewey SL, Pappas N. Decreased striatal dopaminergic responsiveness in detoxified cocaine-dependent subjects. Nature (Lond) 1997;386:830–833. doi: 10.1038/386830a0. [DOI] [PubMed] [Google Scholar]
- Voruganti LN, Slomka P, Zabel P, Mattar A, Awad AG. Cannabis induced dopamine release: an in-vivo SPECT study. Psychiatry Res. 2001;107:173–177. doi: 10.1016/s0925-4927(01)00104-4. [DOI] [PubMed] [Google Scholar]
- Wada JA, Osawa T, Corcoran ME. Effects of tetrahydrocannabinols on kindled amygdaloid seizures and photogenic seizures in Senegalese baboons, Papio papio. Epilepsia. 1975a;16:439–448. doi: 10.1111/j.1528-1157.1975.tb06071.x. [DOI] [PubMed] [Google Scholar]
- Wada JA, Wake A, Sato M, Corcoran ME. Antiepileptic and prophylactic effects of tetrahydrocannabinols in amygdaloid kindled cats. Epilepsia. 1975b;16:503–510. doi: 10.1111/j.1528-1157.1975.tb06080.x. [DOI] [PubMed] [Google Scholar]
- Wade DT, Makela P, Robson P, House H, Bateman C. Do cannabis-based medicinal extracts have general or specific effects on symptoms in multiple sclerosis? A double-blind, randomized, placebo-controlled study on 160 patients. Mult Scler. 2004;10:434–441. doi: 10.1191/1352458504ms1082oa. [DOI] [PubMed] [Google Scholar]
- Wade DT, Robson P, House H, Makela P, Aram J. A preliminary controlled study to determine whether whole-plant cannabis extracts can improve intractable neurogenic symptoms. Clin Rehabil. 2003;17:21–29. doi: 10.1191/0269215503cr581oa. [DOI] [PubMed] [Google Scholar]
- Wagner JA, Abesser M, Karcher J, Laser M, Kunos G. Coronary vasodilator effects of endogenous cannabinoids in vasopressin-preconstricted unpaced rat isolated hearts. J Cardiovasc Pharmacol. 2005;46:348–355. doi: 10.1097/01.fjc.0000175437.87283.f2. [DOI] [PubMed] [Google Scholar]
- Wagner JA, Hu K, Bauersachs J, Karcher J, Wiesler M, Goparaju SK, Kunos G, Ertl G. Endogenous cannabinoids mediate hypotension after experimental myocardial infarction. J Am Coll Cardiol. 2001a;38:2048–2054. doi: 10.1016/s0735-1097(01)01671-0. [DOI] [PubMed] [Google Scholar]
- Wagner JA, Hu K, Karcher J, Bauersachs J, Schafer A, Laser M, Han H, Ertl G. CB1 cannabinoid receptor antagonism promotes remodeling and cannabinoid treatment prevents endothelial dysfunction and hypotension in rats with myocardial infarction. Br J Pharmacol. 2003;138:1251–1258. doi: 10.1038/sj.bjp.0705156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wagner JA, Járai Z, Bátkai S, Kunos G. Hemodynamic effects of cannabinoids: coronary and cerebral vasodilation mediated by cannabinoid CB1 receptors. Eur J Pharmacol. 2001b;423:203–210. doi: 10.1016/s0014-2999(01)01112-8. [DOI] [PubMed] [Google Scholar]
- Wagner JA, Varga K, Ellis EF, Rzigalinski BA, Martin BR, Kunos G. Activation of peripheral CB1 cannabinoid receptors in haemorrhagic shock. Nature (Lond) 1997;390:518–521. doi: 10.1038/37371. [DOI] [PubMed] [Google Scholar]
- Wagner JA, Varga K, Járai Z, Kunos G. Mesenteric vasodilation mediated by endothelial anandamide receptors. Hypertension. 1999;33:429–434. doi: 10.1161/01.hyp.33.1.429. [DOI] [PubMed] [Google Scholar]
- Wahn H, Wolf J, Kram F, Frantz S, Wagner JA. The endocannabinoid arachidonyl ethanolamide (anandamide) increases pulmonary arterial pressure via cyclooxygenase-2 products in isolated rabbit lungs. Am J Physiol. 2005;289:H2491–H2496. doi: 10.1152/ajpheart.00718.2005. [DOI] [PubMed] [Google Scholar]
- Waksman Y, Olson JM, Carlisle SJ, Cabral GA. The central cannabinoid receptor (CB1) mediates inhibition of nitric oxide production by rat microglial cells. J Pharmacol Exp Ther. 1999;288:1357–1366. [PubMed] [Google Scholar]
- Walker JM, Huang SM. Cannabinoid analgesia. Pharmacol Ther. 2002;95:127–135. doi: 10.1016/s0163-7258(02)00252-8. [DOI] [PubMed] [Google Scholar]
- Walker JM, Huang SM, Strangman NM, Sañudo-Peña MC. Identification of the role of endogenous cannabinoids in pain modulation: strategies and pitfalls. J Pain. 2000;1:20–32. [Google Scholar]
- Walker JM, Huang SM, Strangman NM, Tsou K, Sañudo-Peña MC. Pain modulation by release of the endogenous cannabinoid anandamide. Proc Natl Acad Sci USA. 1999;96:12198–12203. doi: 10.1073/pnas.96.21.12198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walker JM, Krey JF, Chu CJ, Huang SM. Endocannabinoids and related fatty acid derivatives in pain modulation. Chem Phys Lipids. 2002;121:159–172. doi: 10.1016/s0009-3084(02)00152-4. [DOI] [PubMed] [Google Scholar]
- Wallace MJ, Blair RE, Falenski KW, Martin BR, DeLorenzo RJ. The endogenous cannabinoid system regulates seizure frequency and duration in a model of temporal lobe epilepsy. J Pharmacol Exp Ther. 2003a;307:129–137. doi: 10.1124/jpet.103.051920. [DOI] [PubMed] [Google Scholar]
- Wallace MJ, Martin BR, DeLorenzo RJ. Evidence for a physiological role of endocannabinoids in the modulation of seizure threshold and severity. Eur J Pharmacol. 2002;452:295–301. doi: 10.1016/s0014-2999(02)02331-2. [DOI] [PubMed] [Google Scholar]
- Walsh D, Nelson KA, Mahmoud FA. Established and potential therapeutic applications of cannabinoids in oncology. Support Care Cancer. 2003;11:137–143. doi: 10.1007/s00520-002-0387-7. [DOI] [PubMed] [Google Scholar]
- Walter L, Franklin A, Witting A, Wade C, Xie Y, Kunos G, Mackie K, Stella N. Nonpsychotropic cannabinoid receptors regulate microglial cell migration. J Neurosci. 2003;23:1398–1405. doi: 10.1523/JNEUROSCI.23-04-01398.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walter L, Stella N. Cannabinoids and neuroinflammation. Br J Pharmacol. 2004;141:775–785. doi: 10.1038/sj.bjp.0705667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walther S, Mahlberg R, Eichmann U, Kunz D. Δ-9-Tetrahydrocannabinol for nighttime agitation in severe dementia. Psychopharmacology. 2006;185:524–528. doi: 10.1007/s00213-006-0343-1. [DOI] [PubMed] [Google Scholar]
- Wang H, Dey SK, Maccarrone M. Jekyll and Hyde: two faces of cannabinoid signaling in male and female fertility. Endocr Rev. 2006 doi: 10.1210/er.2006-0006. in press. [DOI] [PubMed] [Google Scholar]
- Wang H, Guo Y, Wang D, Kingsley PJ, Marnett LJ, Das SK, DuBois RN, Dey SK. Aberrant cannabinoid signaling impairs oviductal transport of embryos. Nat Med. 2004;10:1074–1079. doi: 10.1038/nm1104. [DOI] [PubMed] [Google Scholar]
- Wang J, Paria BC, Dey SK, Armant DR. Stage-specific excitation of cannabinoid receptor exhibits differential effects on mouse embryonic development. Biol Reprod. 1999;60:839–844. doi: 10.1095/biolreprod60.4.839. [DOI] [PubMed] [Google Scholar]
- Wang L, Liu J, Harvey-White J, Zimmer A, Kunos G. Endocannabinoid signaling via cannabinoid receptor 1 is involved in ethanol preference and its age-dependent decline in mice. Proc Natl Acad Sci USA. 2003;100:1393–1398. doi: 10.1073/pnas.0336351100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang X, Feuerstein GZ. Role of immune and inflammatory mediators in CNS injury. Drug News Perspect. 2000;13:133–140. doi: 10.1358/dnp.2000.13.3.657283. [DOI] [PubMed] [Google Scholar]
- Wang Y, Kaminski NE, Wang DH. VR1-mediated depressor effects during high-salt intake: role of anandamide. Hypertension. 2005;46:986–991. doi: 10.1161/01.HYP.0000174596.95607.fd. [DOI] [PubMed] [Google Scholar]
- Wang Y, Liu Y, Ito Y, Hashiguchi T, Kitajima I, Yamakuchi M, Shimizu H, Matsuo S, Imaizumi H, Maruyama I. Simultaneous measurement of anandamide and 2-arachidonoylglycerol by polymyxin B-selective adsorption and subsequent high-performance liquid chromatography analysis: increase in endogenous cannabinoids in the sera of patients with endotoxic shock. Anal Biochem. 2001;294:73–82. doi: 10.1006/abio.2001.5015. [DOI] [PubMed] [Google Scholar]
- Ward SJ, Dykstra LA. The role of CB1 receptors in sweet versus fat reinforcement: effect of CB1 receptor deletion, CB1 receptor antagonism (SR141716A) and CB1 receptor agonism (CP-55940) Behav Pharmacol. 2005;16:381–388. doi: 10.1097/00008877-200509000-00010. [DOI] [PubMed] [Google Scholar]
- Ware M, Beaulieu P. Cannabinoids for the treatment of pain: an update on recent clinical trials. Pain Res Manag. 2005;10:27A–30A. doi: 10.1155/2005/847562. [DOI] [PubMed] [Google Scholar]
- Ware MA, Adams H, Guy GW. The medicinal use of cannabis in the UK: results of a nationwide survey. Int J Clin Pract. 2005;59:291–295. doi: 10.1111/j.1742-1241.2004.00271.x. [DOI] [PubMed] [Google Scholar]
- Ware MA, Doyle CR, Woods R, Lynch ME, Clark AJ. Cannabis use for chronic non-cancer pain: results of a prospective survey. Pain. 2003;102:211–216. doi: 10.1016/s0304-3959(02)00400-1. [DOI] [PubMed] [Google Scholar]
- Wartmann M, Campbell D, Subramanian A, Burstein SH, Davis RJ. The MAP kinase signal transduction pathway is activated by the endogenous cannabinoid anandamide. FEBS Lett. 1995;359:133–136. doi: 10.1016/0014-5793(95)00027-7. [DOI] [PubMed] [Google Scholar]
- Watzl B, Scuderi P, Watson RR. Influence of marijuana components (THC and CBD) on human mononuclear cell cytokine secretion in vitro. Adv Exp Med Biol. 1991;288:63–70. doi: 10.1007/978-1-4684-5925-8_7. [DOI] [PubMed] [Google Scholar]
- Weidenfeld J, Feldman S, Mechoulam R. Effect of the brain constituent anandamide, a cannabinoid receptor agonist, on the hypothalamo-pituitary-adrenal axis in the rat. Neuroendocrinology. 1994;59:110–112. doi: 10.1159/000126646. [DOI] [PubMed] [Google Scholar]
- Weiss F, Lorang MT, Bloom FE, Koob GF. Oral alcohol self-administration stimulates dopamine release in the rat nucleus accumbens: genetic and motivational determinants. J Pharmacol Exp Ther. 1993;267:250–258. [PubMed] [Google Scholar]
- Weksler ME, Gouras G, Relkin NR, Szabo P. The immune system, amyloid-β peptide, and Alzheimer’s disease. Immunol Rev. 2005;205:244–256. doi: 10.1111/j.0105-2896.2005.00264.x. [DOI] [PubMed] [Google Scholar]
- Welch SP, Stevens DL. Antinociceptive activity of intrathecally administered cannabinoids alone and in combination with morphine in mice. J Pharmacol Exp Ther. 1992;262:10–18. [PubMed] [Google Scholar]
- Wenger T, Jamali KA, Juaneda C, Leonardelli J, Tramu G. Arachidonyl ethanolamide (anandamide) activates the parvocellular part of hypothalamic paraventricular nucleus. Biochem Biophys Res Commun. 1997;237:724–728. doi: 10.1006/bbrc.1997.7222. [DOI] [PubMed] [Google Scholar]
- Westlake TM, Howlett AC, Bonner TI, Matsuda LA, Herkenham M. Cannabinoid receptor binding and messenger RNA expression in human brain: an in vitro receptor autoradiography and in situ hybridization histochemistry study of normal aged and Alzheimer’s brains. Neuroscience. 1994;63:637–652. doi: 10.1016/0306-4522(94)90511-8. [DOI] [PubMed] [Google Scholar]
- Whan LB, West MC, McClure N, Lewis SE. Effects of Δ-9-tetrahydrocannabinol, the primary psychoactive cannabinoid in marijuana, on human sperm function in vitro. Fertil Steril. 2006;85:653–660. doi: 10.1016/j.fertnstert.2005.08.027. [DOI] [PubMed] [Google Scholar]
- Whiteside GT, Gottshall SL, Boulet JM, Chaffer SM, Harrison JE, Pearson MS, Turchin PI, Mark L, Garrison AE, Valenzano KJ. A role for cannabinoid receptors, but not endogenous opioids, in the antinociceptive activity of the CB2-selective agonist, GW405833. Eur J Pharmacol. 2005;528:65–72. doi: 10.1016/j.ejphar.2005.10.043. [DOI] [PubMed] [Google Scholar]
- Wickens AP, Pertwee RG. Δ9-Tetrahydrocannabinol and anandamide enhance the ability of muscimol to induce catalepsy in the globus pallidus of rats. Eur J Pharmacol. 1993;250:205–208. doi: 10.1016/0014-2999(93)90646-y. [DOI] [PubMed] [Google Scholar]
- Wiley JL, Burston JJ, Leggett DC, Alekseeva OO, Razdan RK, Mahadevan A, Martin BR. CB1 cannabinoid receptor-mediated modulation of food intake in mice. Br J Pharmacol. 2005;145:293–300. doi: 10.1038/sj.bjp.0706157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Williams CM, Kirkham TC. Anandamide induces overeating: mediation by central cannabinoid (CB1) receptors. Psychopharmacology (Berl) 1999;143:315–317. doi: 10.1007/s002130050953. [DOI] [PubMed] [Google Scholar]
- Williams CM, Kirkham TC. Reversal of Δ9-THC hyperphagia by SR141716 and naloxone but not dexfenfluramine. Pharmacol Biochem Behav. 2002;71:341–348. doi: 10.1016/s0091-3057(01)00694-3. [DOI] [PubMed] [Google Scholar]
- Williams CM, Rogers PJ, Kirkham TC. Hyperphagia in prefed rats following oral Δ9-THC. Physiol Behav. 1998;65:343–346. doi: 10.1016/s0031-9384(98)00170-x. [DOI] [PubMed] [Google Scholar]
- Williams IJ, Edwards S, Rubo A, Haller VL, Stevens DL, Welch SP. Time course of the enhancement and restoration of the analgesic efficacy of codeine and morphine by delta(9)-tetrahydrocannabinol. Eur J Pharmacol. 2006;539:57–63. doi: 10.1016/j.ejphar.2006.04.003. [DOI] [PubMed] [Google Scholar]
- Williams SJ, Hartley JP, Graham JD. Bronchodilator effect of Δ1-tetrahydrocannabinol administered by aerosol of asthmatic patients. Thorax. 1976;31:720–723. doi: 10.1136/thx.31.6.720. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wills-Karp M. Immunologic basis of antigen-induced airway hyperresponsiveness. Annu Rev Immunol. 1999;17:255–281. doi: 10.1146/annurev.immunol.17.1.255. [DOI] [PubMed] [Google Scholar]
- Wilson RI, Kunos G, Nicoll RA. Presynaptic specificity of endocannabinoid signalling in the hippocampus. Neuron. 2001;31:453–462. doi: 10.1016/s0896-6273(01)00372-5. [DOI] [PubMed] [Google Scholar]
- Wilson RI, Nicoll RA. Endogenous cannabinoids mediate retrograde signalling at hippocampal synapses. Nature (Lond) 2001;410:588–592. doi: 10.1038/35069076. [DOI] [PubMed] [Google Scholar]
- Wilson RI, Nicoll RA. Endocannabinoid signaling in the brain. Science (Wash DC) 2002;296:678–682. doi: 10.1126/science.1063545. [DOI] [PubMed] [Google Scholar]
- Wirguin I, Mechoulam R, Breuer A, Schezen E, Weidenfeld J, Brenner T. Suppression of experimental autoimmune encephalomyelitis by cannabinoids. Immunopharmacology. 1994;28:209–214. doi: 10.1016/0162-3109(94)90056-6. [DOI] [PubMed] [Google Scholar]
- Wise RA. Dopamine, learning and motivation. Nat Rev Neurosci. 2004;5:483–494. doi: 10.1038/nrn1406. [DOI] [PubMed] [Google Scholar]
- Witkin JM, Tzavara ET, Davis RJ, Li X, Nomikos GG. A therapeutic role for cannabinoid CB1 receptor antagonists in major depressive disorders. Trends Pharmacol Sci. 2005;26:609–617. doi: 10.1016/j.tips.2005.10.006. [DOI] [PubMed] [Google Scholar]
- Witting A, Chen L, Cudaback E, Straiker A, Walter L, Rickman B, Moller T, Brosnan C, Stella N. Experimental autoimmune encephalomyelitis disrupts endocannabinoid-mediated neuroprotection. Proc Natl Acad Sci USA. 2006;103:6362–6367. doi: 10.1073/pnas.0510418103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Witting A, Walter L, Wacker J, Moller T, Stella N. P2X7 receptors control 2-arachidonoylglycerol production by microglial cells. Proc Natl Acad Sci USA. 2004;101:3214–3219. doi: 10.1073/pnas.0306707101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woolridge E, Barton S, Samuel J, Osorio J, Dougherty A, Holdcroft A. Cannabis use in HIV for pain and other medical symptoms. J Pain Symptom Manage. 2005;29:358–367. doi: 10.1016/j.jpainsymman.2004.07.011. [DOI] [PubMed] [Google Scholar]
- Wright K, Rooney N, Feeney M, Tate J, Robertson D, Welham M, Ward S. Differential expression of cannabinoid receptors in the human colon: cannabinoids promote epithelial wound healing. Gastroenterology. 2005;129:437–453. doi: 10.1016/j.gastro.2005.05.026. [DOI] [PubMed] [Google Scholar]
- Yaksh TL. The antinociceptive effects of intrathecally administered levonantradol and desacetyllevonantradol in the rat. J Clin Pharmacol. 1981;21:334S–340S. doi: 10.1002/j.1552-4604.1981.tb02612.x. [DOI] [PubMed] [Google Scholar]
- Yamauchi T, Kamon J, Minokoshi Y, Ito Y, Waki H, Uchida S, Yamashita S, Noda M, Kita S, Ueki K, et al. Adiponectin stimulates glucose utilization and fatty-acid oxidation by activating AMP-activated protein kinase. Nat Med. 2002;8:1288–1295. doi: 10.1038/nm788. [DOI] [PubMed] [Google Scholar]
- Yao L, McFarland K, Fan P, Jiang Z, Ueda T, Diamond I. Adenosine A2a blockade prevents synergy between μ-opiate and cannabinoid CB1 receptors and eliminates heroin-seeking behavior in addicted rats. Proc Natl Acad Sci USA. 2006;103:7877–7882. doi: 10.1073/pnas.0602661103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yazulla S, Studholme KM, McIntosh HH, Deutsch DG. Immunocytochemical localization of cannabinoid CB1 receptor and fatty acid amide hydrolase in rat retina. J Comp Neurol. 1999;415:80–90. doi: 10.1002/(sici)1096-9861(19991206)415:1<80::aid-cne6>3.0.co;2-h. [DOI] [PubMed] [Google Scholar]
- Yazulla S, Studholme KM, McIntosh HH, Fan SF. Cannabinoid receptors on goldfish retinal bipolar cells: electron-microscope immunocytochemistry and whole-cell recordings. Vis Neurosci. 2000;17:391–401. doi: 10.1017/s0952523800173079. [DOI] [PubMed] [Google Scholar]
- Yoles E, Belkin M, Schwartz M. HU-211, a nonpsychotropic cannabinoid, produces short- and long-term neuroprotection after optic nerve axotomy. J Neurotrauma. 1996;13:49–57. doi: 10.1089/neu.1996.13.49. [DOI] [PubMed] [Google Scholar]
- Yoshida T, Fukaya M, Uchigashima M, Miura E, Kamiya H, Kano M, Watanabe M. Localization of diacylglycerol lipase-α around postsynaptic spine suggests close proximity between production site of an endocannabinoid, 2-arachidinoyl-glycerol, and presynaptic cannabinoid CB1 receptor. J Neurosci. 2006;26:4740–4751. doi: 10.1523/JNEUROSCI.0054-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoshihara S, Morimoto H, Ohori M, Yamada Y, Abe T, Arisaka O. Endogenous cannabinoid receptor agonists inhibit neurogenic inflammations in guinea pig airways. Int Arch Allergy Immunol. 2005;138:80–87. doi: 10.1159/000087361. [DOI] [PubMed] [Google Scholar]
- Zajicek J, Fox P, Sanders H, Wright D, Vickery J, Nunn A, Thompson A. The cannabinoids in MS study—final results from 12 months follow-up. Mult Scler. 2004;109:S115. [Google Scholar]
- Zajicek J, Fox P, Sanders H, Wright D, Vickery J, Nunn A, Thompson A UK MS Research Group. Cannabinoids for treatment of spasticity and other symptoms related to multiple sclerosis (CAMS study): multicentre randomised placebo-controlled trial. Lancet. 2003;362:1517–1526. doi: 10.1016/S0140-6736(03)14738-1. [DOI] [PubMed] [Google Scholar]
- Zalish M, Lavie V. Dexanabinol (HU-211) has a beneficial effect on axonal sprouting and survival after rat optic nerve crush injury. Vision Res. 2003;43:237–242. doi: 10.1016/s0042-6989(02)00494-7. [DOI] [PubMed] [Google Scholar]
- Zangen A, Solinas M, Ikemoto S, Goldberg SR, Wise RA. Two brain sites for cannabinoid reward. J Neurosci. 2006;26:4901–4907. doi: 10.1523/JNEUROSCI.3554-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zaretsky A, Rector NA, Seeman MV, Fornazzari X. Current cannabis use and tardive dyskinesia. Schizophr Res. 1993;11:3–8. doi: 10.1016/0920-9964(93)90032-e. [DOI] [PubMed] [Google Scholar]
- Zaugg HE, Kyncl J. New antihypertensive cannabinoids. J Med Chem. 1983;26:214–217. doi: 10.1021/jm00356a017. [DOI] [PubMed] [Google Scholar]
- Zavitsanou K, Garrick T, Huang XF. Selective antagonist [3H]SR141716A binding to cannabinoid CB1 receptors is increased in the anterior cingulate cortex in schizophrenia. Prog Neuropsychopharmacol Biol Psychiatry. 2004;28:355–360. doi: 10.1016/j.pnpbp.2003.11.005. [DOI] [PubMed] [Google Scholar]
- Zeng BY, Dass B, Owen A, Rose S, Cannizzaro C, Tel BC, Jenner P. Chronic L-DOPA treatment increases striatal cannabinoid CB1 receptor mRNA expression in 6-hydroxydopamine-lesioned rats. Neurosci Lett. 1999;276:71–74. doi: 10.1016/s0304-3940(99)00762-4. [DOI] [PubMed] [Google Scholar]
- Zhang ZF, Morgenstern H, Spitz MR, Tashkin DP, Yu GP, Marshall JR, Hsu TC, Schantz SP. Marijuana use and increased risk of squamous cell carcinoma of the head and neck. Cancer Epidemiol Biomarkers Prev. 1999;8:1071–1078. [PubMed] [Google Scholar]
- Zheng ZM, Specter S, Friedman H. Inhibition by Δ-9-tetrahydrocannab-inol of tumor necrosis factor α production by mouse and human macrophages. Int J Immunopharmacol. 1992;14:1445–1452. doi: 10.1016/0192-0561(92)90017-f. [DOI] [PubMed] [Google Scholar]
- Zheng ZM, Specter SC. Δ-9-Tetrahydrocannabinol suppresses tumor necrosis factor α maturation and secretion but not its transcription in mouse macrophages. Int J Immunopharmacol. 1996;18:53–68. doi: 10.1016/0192-0561(95)00107-7. [DOI] [PubMed] [Google Scholar]
- Zhu LX, Sharma S, Stolina M, Gardner B, Roth MD, Tashkin DP, Dubinett SM. Δ-9-Tetrahydrocannabinol inhibits antitumor immunity by a CB2 receptor-mediated, cytokine-dependent pathway. J Immunol. 2000;165:373–380. doi: 10.4049/jimmunol.165.1.373. [DOI] [PubMed] [Google Scholar]
- Zhuang SY, Bridges D, Grigorenko E, McCloud S, Boon A, Hampson RE, Deadwyler SA. Cannabinoids produce neuroprotection by reducing intracellular calcium release from ryanodine-sensitive stores. Neuropharmacology. 2005;48:1086–1096. doi: 10.1016/j.neuropharm.2005.01.005. [DOI] [PubMed] [Google Scholar]
- Zimmer A, Valjent E, Konig M, Zimmer AM, Robledo P, Hahn H, Valverde O, Maldonado R. Absence of Δ-9-tetrahydrocannabinol dysphoric effects in dynorphin-deficient mice. J Neurosci. 2001;21:9499–9505. doi: 10.1523/JNEUROSCI.21-23-09499.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zimmer A, Zimmer AM, Hohmann AG, Herkenham M, Bonner TI. Increased mortality, hypoactivity, and hypoalgesia in cannabinoid CB1 receptor knockout mice. Proc Natl Acad Sci USA. 1999;96:5780–5785. doi: 10.1073/pnas.96.10.5780. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zuardi AW, Crippa JA, Hallak JE, Moreira FA, Guimaraes FS. Cannabidiol, a Cannabis sativa constituent, as an antipsychotic drug. Braz J Med Biol Res. 2006;39:421–429. doi: 10.1590/s0100-879x2006000400001. [DOI] [PubMed] [Google Scholar]
- Zuardi AW, Morais SL, Guimaraes FS, Mechoulam R. Antipsychotic effect of cannabidiol. J Clin Psychiatry. 1995;56:485–486. [PubMed] [Google Scholar]
- Zuardi AW, Shirakawa I, Finkelfarb E, Karniol IG. Action of cannabidiol on the anxiety and other effects produced by Δ9-THC in normal subjects. Psychopharmacology. 1982;76:245–250. doi: 10.1007/BF00432554. [DOI] [PubMed] [Google Scholar]
- Zurier RB, Rossetti RG, Burstein SH, Bidinger B. Suppression of human monocyte interleukin-1β production by ajulemic acid, a nonpsychoactive cannabinoid. Biochem Pharmacol. 2003;65:649–655. doi: 10.1016/s0006-2952(02)01604-0. [DOI] [PubMed] [Google Scholar]
- Zurier RB, Rossetti RG, Lane JH, Goldberg JM, Hunter SA, Burstein SH. Dimethylheptyl-THC-11 oic acid: a nonpsychoactive antiinflammatory agent with a cannabinoid template structure. Arthritis Rheum. 1998;41:163–170. doi: 10.1002/1529-0131(199801)41:1<163::AID-ART20>3.0.CO;2-9. [DOI] [PubMed] [Google Scholar]
- Zygmunt PM, Chuang H, Movahed P, Julius D, Hogestatt ED. The anandamide transport inhibitor AM404 activates vanilloid receptors. Eur J Pharmacol. 2000;396:39–42. doi: 10.1016/s0014-2999(00)00207-7. [DOI] [PubMed] [Google Scholar]
- Zygmunt PM, Petersson J, Andersson DA, Chuang HH, Sorgard M, Di Marzo V, Julius D, Högestätt ED. Vanilloid receptors on sensory nerves mediate the vasodilator action of anandamide. Nature (Lond) 1999;400:452–457. doi: 10.1038/22761. [DOI] [PubMed] [Google Scholar]