

Abstract
Electrical recordings in humans and monkeys show attentional enhancement of evoked responses and gamma synchrony in ventral stream cortical areas. Does this synchrony result from intrinsic activity in visual cortex or from inputs from other structures? Using paired-recordings in the frontal eye field (FEF) and area V4, we found that attention to a stimulus in their joint receptive field leads to enhanced oscillatory coupling between the two areas, particularly at gamma frequencies. This coupling appeared to be initiated by FEF and was time-shifted by about 8-13 ms across a range of frequencies. Considering the expected conduction and synaptic delays between the areas, this time-shifted coupling at gamma frequencies may optimize the postsynaptic impact of spikes from one area upon the other, improving cross-area communication with attention.
A typical crowded scene contains many objects which cannot be processed simultaneously, thus requiring attentional mechanisms to select the ones most relevant to behavior. Electrophysiological studies in monkeys have shown that attention leads to enhanced responses of neurons in ventral stream areas important for object recognition, at the expense of responses to distracting stimuli (1). Moreover, attention increases neural synchrony, often in the gamma frequency range (2-5). Given that cells have limited integration times, increases in synchrony and firing rates may together have a larger impact on downstream neurons and thus increase the effectiveness of behaviorally relevant stimuli (6, 7). Areas in prefrontal (PFC) and parietal cortex may be sources of the top-down attentional feedback to ventral stream areas, which could enhance firing rates with attention (1, 4, 8). However, the mechanisms that cause increases in neural synchrony with attention in visual cortex are unknown.
We investigated if FEF, an area within the PFC, is a source of enhanced neural synchrony effects in area V4 during attention. The FEF has reciprocal connections with V4 (9-11) and electrical stimulation of FEF enhances V4 neuronal responses to a stimulus in the receptive field (RF) (12, 13). We recorded spikes (multi-unit) and local field potentials (LFPs) simultaneously from FEF and V4, in two monkeys trained in a covert attention task (Fig. 1A (14)). One grating stimulus appeared inside the shared RF, and two others appeared outside. Following a variable delay, the spot at fixation changed color (cue) to match the color of one of the three gratings, indicating the target stimulus to be attended. The monkey was rewarded for releasing a bar when the target stimulus changed color.
Fig. 1.
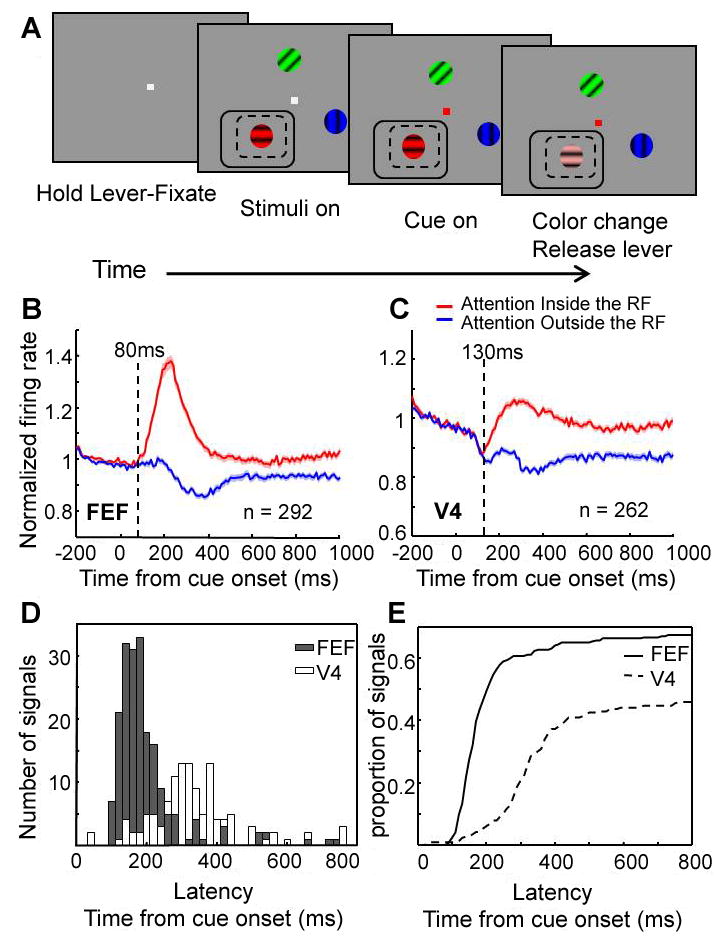
(A) Illustration of behavioral task. Dashed and solid line rectangles indicate hypothetical overlapping RFs for V4 and FEF sites, respectively. (B and C) Normalized firing rates averaged across the population of cells in FEF and V4, respectively. Standard error of the mean (±) at each time point is indicated by shading over the lines. Vertical dashed lines indicate latency of attentional effects at the population level. (D) Distribution of attentional latencies in the firing rates of FEF and V4 neurons. (E) Cumulative distribution of FEF and V4 latencies, represented as a proportion of recordings in which latencies could be reliably estimated.
We first verified that attention caused enhanced firing rates in FEF and V4. We recorded from 292 sites with visual responses in FEF and 262 sites in V4. The results were qualitatively similar (and statistically significant) in both monkeys, and were therefore combined. Figure 1B and C shows the average normalized response of the population of FEF and V4 neurons, respectively, for conditions with overlapping RFs. Neuronal responses were significantly increased by attention to the joint RF in both areas (average response 100-800 ms after cue onset; Wilcoxon sign-rank test p < 0.001), and remained significantly enhanced until the end of the trial (average response in a window 500 ms before the target's color change; Wilcoxon sign-rank test p < 0.001) (for distribution of attentional effects on firing rate, see S.O.M.).
Attentional effects on firing rates occurred significantly earlier in FEF than in V4 (at 80 ms post-cue in FEF, 130 ms in V4, Fig 1B,C dashed lines; p = 0.017 two-sided permutation test (see S.O.M.). The distribution of attentional latencies is shown separately for FEF and V4 in Figure 1D and E, and was similarly shifted earlier for FEF (Wilcoxon rank-sum test, p < 0.001) (for a table of latency measurements, see Table S1).
We next used multi-taper spectral methods to calculate the coherence between spikes and LFPs (14). Spike-field coherence in the gamma band significantly increased with attention within each area (paired t-test, p < 0.001 in both areas; coherence averaged between 40-60 Hz), whereas low frequencies were desynchronized (average coherence between 5-20 Hz: paired t-test, p < 0.001 in both areas) (Figs. 2A-B). At the population level, gamma band coherence increased by 14% in V4 (2, 3, 15) and by 22% in the FEF (for distributions of effects, see S.O.M.).
Fig. 2.
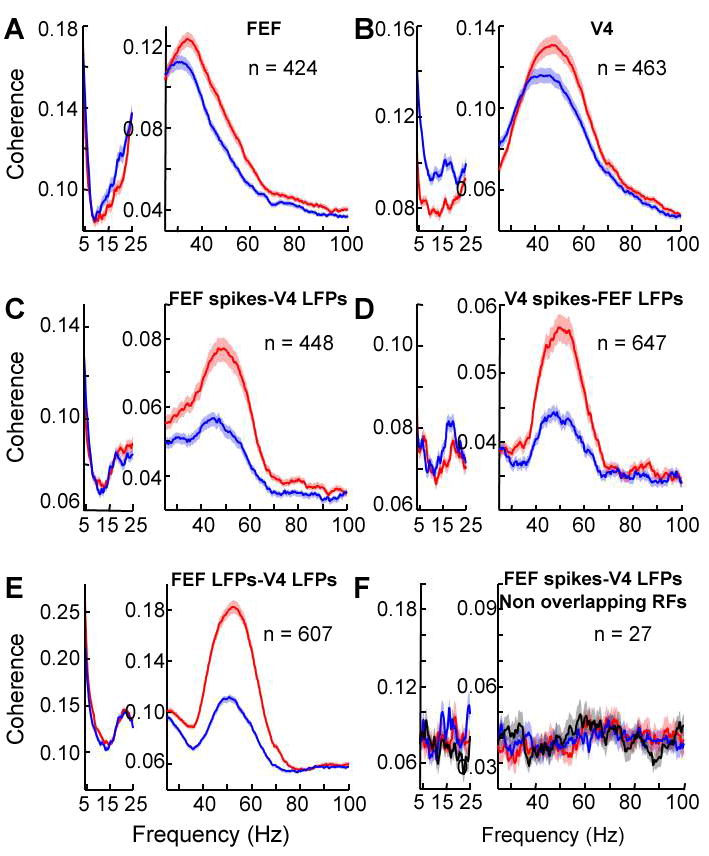
Attentional enhancement of synchronization. (A-D) Spike-field coherence: (A) within FEF, (B) within V4, (C) between spikes in FEF and LFPs in V4, and (D) between spikes in V4 and LFPs in FEF. (E) LFP-LFP coherence between FEF and V4 sites. Conventions as in Fig. 1. (F) Spike-LFP coherence between FEF spikes and V4 LFPs from recording sites with non-overlapping RFs (i) with attention inside the V4 RF in post-stimulus period (red line), (ii) with attention in right hemifield, outside both V4 and FEF RFs in the post-stimulus period (blue line) and (iii) in pre-stimulus period with no stimuli except for fixation spot (black line). Tapers providing an effective smoothing of ±10 Hz were used for spectral estimation of higher frequencies (25-100 Hz, right part of each graph) and tapers providing smoothing of ±3Hz were used for lower frequencies (<25 Hz, left part of each graph).
If FEF is the source of enhanced synchrony in V4, the critical question is whether attention increases coupled oscillations between the two areas. We found that the attentional effect on gamma frequency spike-field coherence between areas was even larger than the effects within areas (Fig. 2C and D). With attention, gamma coherence between V4 spikes and FEF LFPs increased by 26% at the population level (paired t-test, p < 0.001) and that between FEF spikes and V4 LFPs by 37% (paired t-test, p < 0.001), and remained enhanced through the end of the trial (paired t-test, p < 0.001 for all pair types). All of these effects were highly dependent on RF overlap at the locus of attention. For pairs of recordings with non-overlapping RFs, coherence in the two attention conditions did not differ from that in the pre-stimulus period (one way ANOVA, p = 0.86; Fig. 2F and S.O.M.).
Gamma frequency coherence between LFPs recorded across the two areas was enhanced 63% by attention (Fig. 2E) (paired t-test, p < 0.001), and gamma coherence between spike trains across areas was enhanced by 13% (paired t-test, p < 0.001). In general, spike-spike coherence across electrodes is smaller than spike-field and field-field coherence for statistical reasons (16). Another likely factor is that connections between FEF and V4 are patchy (9, 10) and LFPs sum signals over a wider area.
We considered whether the synchronous oscillations between V4 and FEF might have resulted from a common oscillatory input, which would be expected to result in zero phase-lag synchrony between the areas. To test for this, we computed the distribution of the coherence phase shifts within and across areas. Within areas, the distribution of the average (between 40 and 60 Hz) relative phase between the two recorded signals (Fig. 3) had a median close to zero (attend-in condition: Rayleigh test, FEF: p < 0.001, median = 7°; V4: p < 0.001, median = -26°), corresponding to a time delay of 0.5-1.5 ms between spikes and the phase of maximum depolarization in the LFP at 50 Hz (Fig. 3A). By contrast, the phase of spike-field coherence across areas was shifted approximately half a gamma cycle (attend-in condition: Rayleigh test, FEF spikes - V4 LFPs, p < 0.001, median phase = -142° (or 218°); V4 spikes - FEF LFPs, p < 0.001, median phase = 144° (or -216°)), corresponding to a time shift of ∼8 (or 12) ms (Fig. 3A). Likewise, the median phase of spike-spike coherence pairs having a maximum gamma coherence peak of at least 0.1 was about 120°, which corresponds to a time shift of 7 ms. Similar results were found by comparing spike-triggered averages of the LFP within and across areas (Fig. 3C-F).
Fig. 3.
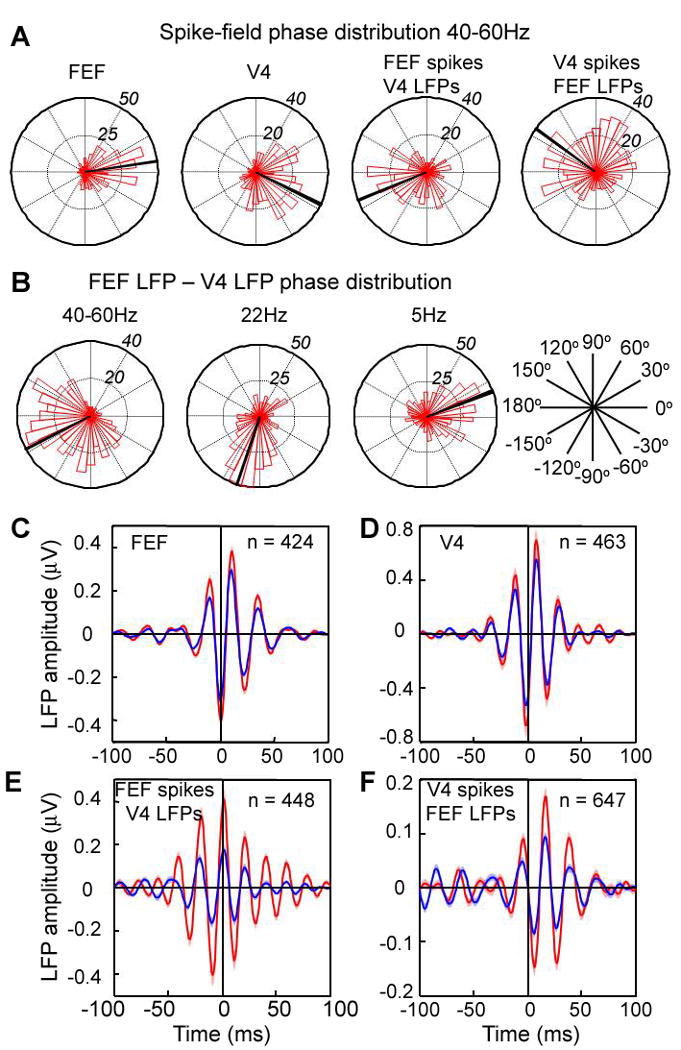
Relative phase. (A) Distribution of average relative phase (40-60 Hz) between spikes and LFPs within and across areas. (B) Distribution of relative phases between FEF and V4 LFPs at different frequencies (40-60 Hz, 22 Hz, 5 Hz). All phases from condition with attention inside the RF. (C-F) Spike triggered averages of LFPs filtered between 35 - 80 Hz with: (C) spikes and LFPs from FEF, (D) spikes and LFPs from V4, (E) spikes from FEF and LFPs from V4, and (F) spikes from V4 and LFPs from FEF. Conventions as in Fig. 1.
Although the peak coherence and largest attentional effects were in the gamma range, there was also coherence between FEF and V4 at other frequencies. We therefore tested whether the phase relationship at these other frequencies followed a fixed time shift of ∼8-12 ms or a fixed phase shift of half a cycle. The medians of the distributions for the gamma, beta and theta frequencies (40-60 Hz median = -152°; 22 Hz median = -105°; 5 Hz median = 20°) correspond to time delays of -8, -13 and 11 ms, or a relatively fixed time shift of 8-13 ms in either direction rather than a fixed phase shift (Fig. 3B). A comparable (∼10 ms) delay has been found between visual response latencies in anatomically connected areas along the ventral stream (for a review see (17)), suggesting that conduction times and synaptic delays account for the 8-13 ms shift in coupling.
The earlier latency of attentional effects on firing rates in FEF compared to V4 suggests that FEF may initiate the coupled oscillations between the two areas. To further test this idea, we used Granger causality analysis to test the relative strength of influence of V4 on FEF LFPs, and vice versa (14). Granger causality values for gamma increased with attention for both directions (paired t-test, p < 0.001 for both directions) and were significantly above chance (FEF→V4 peak = 0.010 at 46 Hz, V4→FEF peak = 0.025 at 55 Hz, permutation test p < 0.001 (14)), indicating that gamma activity in each area has a significant causal influence on the other area. However, the attentional effects on the Granger causality values appeared significantly earlier in the FEF to V4 direction than the reverse direction (Fig. 4A and B, FEF to V4: 110 ms, V4 to FEF: 160 ms, two-sided permutation test p < 0.05), consistent with the idea that FEF initiates the gamma frequency oscillations in V4. Interestingly, the causality relationship reversed a short time later, with the Granger values becoming significantly larger in the V4 to FEF direction around 300 ms after cue onset (average 400-1000 ms after cue onset, paired t-test, p < 0.001). In fact, the Granger values in the FEF to V4 direction greatly diminished across the trial.
Fig. 4.
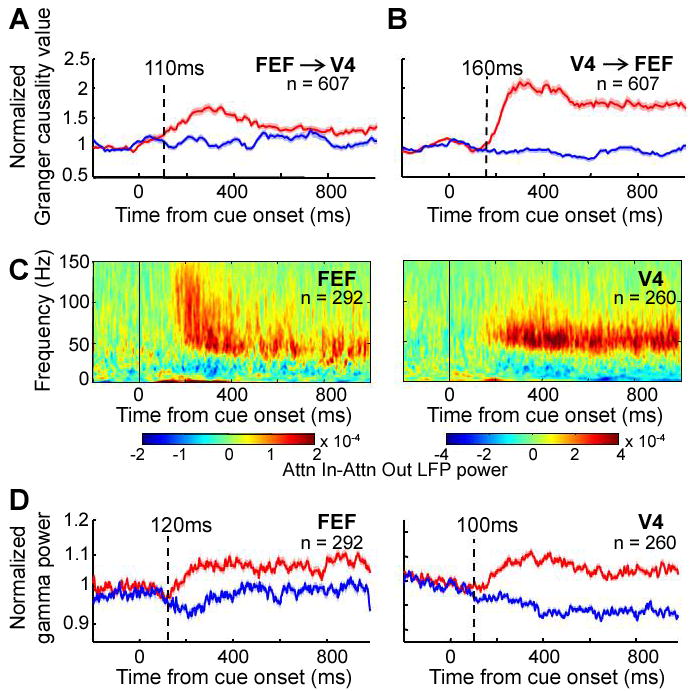
Time-Frequency synchrony measures and directional influences. (A and B) Population average of normalized Granger causality values averaged between 40-60 Hz across all pair-wise combinations of LFPs recorded in FEF and V4. Direction of influence indicated by arrows. (C) Population averages of attentional effects (attention inside RF-attention outside RF) on FEF and V4 power. (D) Normalized LFP gamma power in FEF and V4. Conventions as in Fig. 1.
We considered whether firing rate changes with attention in FEF preceded the attentional effects on synchrony or vice versa. We used the Hilbert-Huang transform method (18) to calculate instantaneous LFP power over time in FEF and V4 (Fig. 4C). At the population level, significant attentional enhancement of gamma power in the LFP in FEF and V4 occurred at 120 ms and 100 ms, respectively (Fig 4D), which was not a significant difference (two-sided permutation test, p = 0.84). To compare the relative latencies of attention effects on firing rate and LFP gamma power we calculated the distribution of latencies for attentional effects across all individual sites in the first 300 ms after cue onset. The distributions of attentional latencies in LFP gamma power in both FEF and V4 were significantly later than the distribution of latencies for attentional effects on firing rates in FEF (Wilcoxon rank-sum test p < 0.01 for both comparisons) and significantly earlier than the distribution of latencies for attentional effects on V4 firing rates (Wilcoxon rank-sum test, V4 LFP gamma power p < 0.05; FEF LFP gamma power p < 0.001, Table S2) Together, these results indicate that significant attentional effects on LFP gamma power in either area occur later than the earliest attentional effects on firing rates in the FEF. Rather than being caused by enhanced gamma oscillations, increases in firing rates in FEF with attention may initiate the coupled oscillations within and across areas. By contrast, firing rate changes in area V4 occur later and might result at least in part from enhanced gamma oscillations.
In summary, the results suggest that FEF is a major source of the attentional effects on gamma frequency synchrony in V4, and likely other ventral stream areas. The Granger causality analyses suggest that top-down inputs from FEF to V4 predominate at the onset of spatially directed attention, but the bottom-up inputs from V4 to FEF come to predominate over the course of sustained attention. The coupled oscillations across areas are shifted in time by about 8-13 ms, which may be the optimal time shift to allow for spikes initiated in one area to affect cells at a peak depolarization phase in the coupled area (17). Tight coupling between the inputs and outputs of cells in V4 and FEF may also allow for enhanced spike timing-dependent plasticity of the connections between the two areas (19), which might mediate learning effects with attention. For distracting stimuli, or for sites with non-overlapping RFs, these coupled oscillations are much smaller, which will reduce the impact of spikes in one area upon the other. We do not suggest that the attentional effects on gamma synchrony and firing rates in V4 are caused solely by inputs from FEF, as V4 receives inputs from several other structures which have been implicated in attention (see (1)). However, these other inputs may also need to be synchronized at compatible frequencies and with the appropriate time shifts to support effective communication.
It has been suggested that low frequency synchronization (e.g. beta) is more suitable for long-range or polysynaptic communication across distant brain areas, with gamma rhythms being used for local computations (20). Although there is evidence for such low frequency long-range synchronization (21-25), here, we show that two distant, but monosynaptically connected, areas can be synchronized at gamma frequencies, which is probably not caused simply by common input (see also (21)). Enhanced oscillatory coupling has now been reported across several brain structures in monkeys (4, 23, 26) and other species (21, 24, 27) in association with attention and other behaviors, at a variety of frequencies, and may therefore be a general mechanism for regulating communication across brain structures (6, 28).
Supplementary Material
References and Notes
- 1.Desimone R, Duncan J. Annu Rev Neurosci. 1995;18:193. doi: 10.1146/annurev.ne.18.030195.001205. [DOI] [PubMed] [Google Scholar]
- 2.Fries P, Reynolds JH, Rorie AE, Desimone R. Science. 2001;291:1560. doi: 10.1126/science.1055465. [DOI] [PubMed] [Google Scholar]
- 3.Bichot NP, Rossi AF, Desimone R. Science. 2005;308:529. doi: 10.1126/science.1109676. [DOI] [PubMed] [Google Scholar]
- 4.Saalmann YB, Pigarev IN, Vidyasagar TR. Science. 2007;316:1612. doi: 10.1126/science.1139140. [DOI] [PubMed] [Google Scholar]
- 5.Lakatos P, Karmos G, Mehta AD, Ulbert I, Schroeder CE. Science. 2008;320:110. doi: 10.1126/science.1154735. [DOI] [PubMed] [Google Scholar]
- 6.Borgers C, Epstein S, Kopell NJ. Proc Natl Acad Sci U S A. 2008;105:18023. doi: 10.1073/pnas.0809511105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Tiesinga P, Fellous JM, Sejnowski TJ. Nat Rev Neurosci. 2008;9:97. doi: 10.1038/nrn2315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Miller EK, Cohen JD. Annu Rev Neurosci. 2001;24:167. doi: 10.1146/annurev.neuro.24.1.167. [DOI] [PubMed] [Google Scholar]
- 9.Stanton GB, Bruce CJ, Goldberg ME. J Comp Neurol. 1995;353:291. doi: 10.1002/cne.903530210. [DOI] [PubMed] [Google Scholar]
- 10.Schall JD, Morel A, King DJ, Bullier J. J Neurosci. 1995;15:4464. doi: 10.1523/JNEUROSCI.15-06-04464.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Ungerleider LG, Galkin TW, Desimone R, Gattass R. Cereb Cortex. 2008;18:477. doi: 10.1093/cercor/bhm061. [DOI] [PubMed] [Google Scholar]
- 12.Moore T, Armstrong KM. Nature. 2003;421:370. doi: 10.1038/nature01341. [DOI] [PubMed] [Google Scholar]
- 13.Ekstrom LB, Roelfsema PR, Arsenault JT, Bonmassar G, Vanduffel W. Science. 2008;321:414. doi: 10.1126/science.1153276. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Materials and methods are available as supporting material on Science Online
- 15.Fries P, Womelsdorf T, Oostenveld R, Desimone R. J Neurosci. 2008;28:4823. doi: 10.1523/JNEUROSCI.4499-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Zeitler M, Fries P, Gielen S. Neural Comput. 2006;18:2256. doi: 10.1162/neco.2006.18.9.2256. [DOI] [PubMed] [Google Scholar]
- 17.Nowak LG, Bullier J. In: Cerebral Cortex. Rockland KS, Kaas JH, Peters A, editors. Vol. 12. Plenum Press; New York: 1997. pp. 205–241. [Google Scholar]
- 18.Huang NE, et al. Proceedings of the Royal Society of London Series a-Mathematical Physical and Engineering Sciences. 1998;454:903. [Google Scholar]
- 19.Dan Y, Poo MM. Neuron. 2004;44:23. doi: 10.1016/j.neuron.2004.09.007. [DOI] [PubMed] [Google Scholar]
- 20.Kopell N, Ermentrout GB, Whittington MA, Traub RD. Proc Natl Acad Sci U S A. 2000;97:1867. doi: 10.1073/pnas.97.4.1867. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.von Stein A, Chiang C, Konig P. Proc Natl Acad Sci U S A. 2000;97:14748. doi: 10.1073/pnas.97.26.14748. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Brovelli A, et al. Proc Natl Acad Sci U S A. 2004;101:9849. doi: 10.1073/pnas.0308538101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Pesaran B, Nelson MJ, Andersen RA. Nature. 2008;453:406. doi: 10.1038/nature06849. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Roelfsema PR, Engel AK, Konig P, Singer W. Nature. 1997;385:157. doi: 10.1038/385157a0. [DOI] [PubMed] [Google Scholar]
- 25.Sirota A, et al. Neuron. 2008;60:683. doi: 10.1016/j.neuron.2008.09.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Buschman TJ, Miller EK. Science. 2007;315:1860. doi: 10.1126/science.1138071. [DOI] [PubMed] [Google Scholar]
- 27.Siapas AG, Lubenov EV, Wilson MA. Neuron. 2005;46:141. doi: 10.1016/j.neuron.2005.02.028. [DOI] [PubMed] [Google Scholar]
- 28.Womelsdorf T, Fries P. Curr Opin Neurobiol. 2007;17:154. doi: 10.1016/j.conb.2007.02.002. [DOI] [PubMed] [Google Scholar]
- 29.We thank Grant Pielli, Donovan Stock and Courtney Alfes for help with the animal training and Zu-Xiang Liu for help with the Granger causality analysis. We also thank Narcisse Bichot, Rogier Landman, Grant Mulliken and Andy Mitz for helpful discussions. Supported by Grants EY017292 and EY017921 to R.D. S.J.G. was supported in part by MH64445 from the National Institutes of Health (USA).
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.