

Abstract
Chronic overeating of obesogenic diets can lead to obesity, reduced dopamine signaling, and increased consumption of added sugars to compensate for blunted reward. However, the specific role of diet composition yet remains unknown. To study this, Sprague Dawley male rats were fed a high-energy diet with high fat and low carbohydrate content (HFHE), a fat-sugar combination high-energy diet (FCHE), or standard chow for 24 weeks. We found that both high-energy diets produced substantial body weight gain compared to chow-fed controls. To investigate dopamine control of short (2-hr) intake of palatable sucrose or fructose solutions, rats were pretreated peripherally (IP) with equimolar doses (0–600 nmol/kg) of the dopamine D1 (SCH23390) and D2 (raclopride) subtype-specific receptor antagonists. The results showed an overall increase in the efficacy of D1 and D2 receptor antagonists on suppression of intake in obese rats compared to lean rats, with effects differing based on diets and test solutions. Specifically, SCH23390 potently reduced both sucrose and fructose intake in all groups; however, lower doses were more effective in HFHE rats. In contrast, raclopride was most effective at reducing fructose intake in the obese FCHE rats. Thus, it appears that obesity due to the consumption of combinations of dietary fat and sugar rather than extra calories from dietary fat alone, may result in reduced D2 receptor signaling. Furthermore, such deficits seem to preferentially affect the control of fructose intake. These findings demonstrate for the first time a plausible interaction between diet composition and dopamine control of carbohydrate intake in diet-induced obese rats. It also provides additional evidence that sucrose and fructose intake is regulated differentially by the dopamine system.
Keywords: diet-induced obesity, sucrose, fructose, reward, SCH2330, raclopride
1. Introduction
Decades of research by Hoebel and his trainees have provided essential information on the role of the brain's dopaminergic system in the regulation of feeding, thus developing the concept of “food reward” [1-4]. Remarkably, Hoebel's early experiments established midbrain dopamine as a key factor in chronic overeating and resultant obesity [5-8], long before direct evidence was made available from imaging studies [9, 10].
The notion that food exerts control over eating, and in turn, that sustained or intermittent access to highly palatable meals (i.e. those high in sugars and fats) could cause lasting changes within feeding regulatory systems has long been central to Hoebel's theories on the development of binge-type behaviors. Early in his career, he also applied elements of this reasoning to obesity. In a 1977 review, Hoebel remarked that there may be “different kinds of obesities that require different treatments” [11]. Since then, a plethora of research on obesity has indeed identified various genetic, metabolic, and environmental factors that may explain the variation in the development, consequences, and treatment of obesity [12-15]. However, our understanding of the specific contributions of macronutrients to altered food reward functions is far from complete. The present paper summarizes data from a study that was inspired by Bart's research and intended to reduce this gap in our knowledge.
Within the multifaceted etiology of obesity, diet remains a key factor in obesity development. Obesogenic diets are diets high in caloric value, often palatable foods that lead to obesity after extended exposure [16]. However, the macronutrient composition of obesogenic diets can differ and this variation could impact neural systems altered in obesity, such as dopamine. Indeed, maintenance on an obesogenic diet has been shown to reduce dopamine levels in the accumbens, as well as shift the reactivity of the mesocorticolimbic system such that a more palatable diet is required to achieve similar food-induced increases in extracellular dopamine as seen in chow-fed controls [17]. One potential mechanism is an adaptive down-regulation due to augmented and chronic stimulation by palatable foods [18]. In fact, studies from our laboratory have shown that even orosensory stimulation by either sucrose or fat is sufficient to stimulate dopamine release in the nucleus accumbens [19, 20]. Of particular relevance, fat and sugars appear to affect reward systems differently, as it is inferred from the greater potency of sugars to produce addictive-like behaviors [21]. Other recent investigations have shown differential effects on the neuroendocrine system and later susceptibility to weight gain based on the ratio of fat and carbohydrates in obesogenic diets [22, 23]. In addition, increased attention has been devoted to potential particularities in the regulatory responses to a high-fructose corn syrup diet and the purported consequences of the apparent ease with which it can cause obesity and derangement of food regulation. Specifically, recent studies by Avena and Hoebel demonstrated that rats with access to high-fructose corn syrup (HFCS) for 12 hrs every day for 8 weeks gained significantly more body weight than animals given equal access to 10% sucrose, even though they consumed the same number of total calories, but fewer calories from HFCS than sucrose [24]. The rising incidence of obesity and potential for the discovery of novel treatments demands investigation of how the intake of common high-energy and palatable foods, such as sucrose and fructose, is controlled under dietary obesity conditions.
Therefore the current study investigated dopamine regulation of sucrose and fructose intake in rats that became obese as a result of extended maintenance on two standard high-energy diets widely used to produce dietary obesity in rats, and varying in fat and carbohydrate content. Specifically, we evaluated the involvement of the two major classes of dopamine receptors using peripheral (interperitoneal; i.p.) administration of the dopamine D1 receptor (D1R) antagonist SCH23390 or the dopamine D2 recpetor (D2R) antagonist raclopride in lean and dietary obese rats in a short (2-hr) one-bottle intake test of sucrose or fructose. These common carbohydrates are prevalent within human diets, are readily consumed by rats and have positive reinforcing properties [25-28]. Sucrose intake has previously been shown to stimulate dopamine release within the nucleus accumbens [3, 19, 29] and peripheral administration of both SCH23390 and raclopride reduces sucrose sham-feeding [30]. Although there is heightened interest by the science community, as well as the public media, similar effects of dopamine antagonists on fructose intake have only been investigated in the context of acquisition and expression of conditioned preferences, and these studies were also limited to lean rats [31-33]. Despite the potential implications, effects of dopamine receptor antagonists on carbohydrate intake in various obesity models and in the absence of homeostatic drive (i.e. following periods of food restriction) have not been investigated. Therefore, rats in the current study were kept sated to avoid confounding effects from hunger and energy deficit.
2. Methods
2.1 Animals and diets
Twenty-eight adult male Sprague-Dawley rats (Charles River, Wilmington, MA) weighing approximately 250 g at the onset of the study were housed in individual cages in a temperature-controlled vivarium and maintained on a 12:12 light-dark cycle, with lights on at 0700.
Animals were given ad libitum access to one of the following three diets: standard laboratory chow (Teklad #2018, 3.4 kcal/g, 18 kcal% fat, 58 kcal% carbohydrates, 24 kcal% protein; Teklad Diets, Somerville, NJ) or one of two high-energy diets (Research Diets, New Brunswick, NJ), one diet where the primary energy source was fat (high fat-high-energy, HFHE diet; Research Diets #D12492: 5.24 kcal/g, 60 kcal% fat, 20 kcal% carbohydrates, 20 kcal% protein) or a high-energy diet consisting of both fat and carbohydrates (fat-sugar combination high-energy, FCHE diet; Research Diets #D12266B; 4.41 kcal/g, 32 kcal% fat, 51 kcal% carbohydrates, 17 kcal% protein). At the initiation of the study, groups were weight matched to form statistically equal cohorts based on body weight and were then maintained on their respective diets for 24 weeks prior to and throughout the behavioral experiments. At 18 weeks and throughout the experiment, body weight and food intake were measured daily. Animals were tested in a sated state with no periods of food restriction throughout the experiment.
2.2 Body composition
In addition to significant increase in body weight, to demonstrate the presence of obesity 1H-NMR body composition analysis (Bruker LF90 proton-NMR Minispec; Brucker Optics, Woodlands, TX) was performed after 12 weeks of maintenance on the diets.
2.3 Dopamine Antagonists, Test Solutions, and Testing Procedure
The dopamine D1R antagonist SCH23390 (HFHE: n=6; FCHE: n=5; Chow: n=4) and the dopamine D2 receptor antagonist raclopride (HFHE: n=5; FCHE: n=6; Chow: n=4) were used. SCH23390 and raclopride (Tocris Biosciences, Ellisville, MO) were dissolved in sterile saline and administered intraperitoneally 10 minutes prior to 2-hr access to 0.3 M sucrose or 0.4 M fructose. These concentrations were chosen as they are highly palatable to rats and have therefore been commonly used in previous studies [3, 19, 32, 34]. Sucrose and fructose (Fisher-Scientific, Fair Lawn, NJ) were dissolved in filtered tap water no more than 24 hours prior to testing.
Animals were trained to drink test solutions during daily sessions where 2-hr access (beginning at 1000 hr) to sucrose or fructose was provided for 8 days prior to testing to achieve stable baseline intakes, i.e. familiarity with the orosensory and postingestive effects. Training and testing took place in the animals' home colony room, with 100 ml plastic bottles temporarily attached to the front of the home cage so that spouts extended into the cage. Administration of vehicle (saline) or dopamine antagonists began after 24 weeks of maintenance on the diets, at which point both obesogenic diet groups (HFHE and FCHE) had significantly higher body weights than chow controls (Figure 1). A minimum of 48 hours was given between injection days to allow drugs to completely metabolize. No changes to body weight or 24 hour food intake occurred following treatment with the dopamine antagonists.
Figure 1. Body weight during the period prior to and throughout the pharmacological testing period (grey bar).
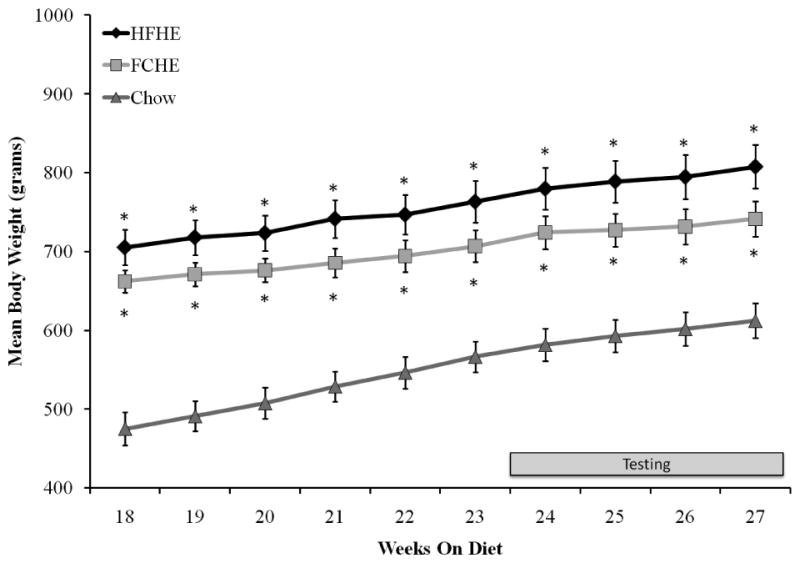
Rats maintained on HFHE or FCHE diets had significantly higher body weight than Chow rats at all times points. There was no difference in body weight between HFHE and FCHE groups at any time point. * p<0.001 compared to Chow.
2.4 Statistical analysis
Body weight and 1H-NMR data were analyzed using a one-way independent samples analysis of variance (ANOVA) with diet as the independent variable.
Intake was measured as ml consumed and is presented as mean ± SEM. Baseline intake (following vehicle, i.e. saline injection) was tested for differences between the diet groups in a three way ANOVA with diet, drug, and carbohydrate as the independent variables. There were no significant effects of diet (F(2,48)=0.3533, p=0.704), drug (F(1,48)=0.1482, p=0.701), nor were there significant interaction effects (diet × drug: F(2,48)=0.4144,p=0.66; diet × carbohydrate: F(2,48)=0.2759, p=0.76; drug × carbohydrate: F(1,48)=0.0062, p=0.73; diet × drug × carbohydrate: F(2,48)=0.3108, p=0.73). However, a significant effect of carbohydrate (F(1,48)=8.8974, p<0.01) was observed (Table 1). Therefore, for all subsequent analyzes intake was converted to a percent reduction from baseline (intake following dose × [ml]/intake following 0 μg/kg [ml]) and analyzed using repeated measures analysis of variance (ANOVA) with Diet (HFHE, FCHE, or Chow) and Drug (raclopride or SCH23390) as independent variables and dose (0, 50, 200, 400 or 600 nmol/kg SCH23390 or raclopride) as the repeated measure. The inhibitory dose (ID50) required to reduce intake to 50% of baseline (0 nmol/kg) was calculated as previously described [35]. Differences in ID50 were compared as a function of Diet and Drug using two-way ANOVA. All analyses were conducted using Statistica (v6.0, StatSoft® Inc., Tulsa, OK) and significant findings were further analyzed using Fischer's least significant difference (LSD) post-hoc tests. Differences were considered statistically significant if p<0.05.
Table 1.
Intake of sucrose and fructose in 2-h tests. Absolute intake values (in ml) of sucrose and fructose intake by diet groups following vehicle (0 nmol/kg) injections. No differences were observed in baseline intake between diet or drug groups. Baseline sucrose intake was significantly greater than baseline fructose intake (pb0.01). Therefore, for all further analyses of drug effects, changes were normalized and expressed as a percent change relative to vehicle baseline intakes.
| Carbohydrate | Drug | Diet Group | Mean Intake ± SEM (ml) |
|---|---|---|---|
| Sucrose | SCH23390 | HFHE | 20.33 + 3.54 |
| FCHE | 21.20 + 2.85 | ||
| Chow | 21.00 + 2.65 | ||
| Raclopride | HFHE | 21.00 + 2.35 | |
| FCHE | 21.00 + 4.14 | ||
| Chow | 22.25 + 2.02 | ||
|
| |||
| Fructose | SCH23390 | HFHE | 15.67 + 2.14 |
| FCHE | 17.2 + 2.85 | ||
| Chow | 12.5 + 2.02 | ||
| Raclopride | HFHE | 12.8 + 2.83 | |
| FCHE | 17.67 + 4.38 | ||
| Chow | 17.50 + 2.36 | ||
3. Results
3.1 Effects of diet on body weight and adiposity
After 12 weeks on the obesogenic diets, the groups differed in body weight (F(2,27)=27.25, p<0.001), percent fat mass (F(2,27)=14.96, p<0.001), and percent lean mass (F(2,27)=15.77, p<0.001). The post hoc tests showed that Chow rats weighed significantly less than both HFHE (p<0.001) and FCHE (p<0.001) rats. Comparison of body composition showed that HFHE and FCHE rats had a greater percentage of fat mass compared to Chow (p<0.05). At 18 weeks, at the beginning of testing (24 weeks) and throughout the testing period, there remained a significant effect of diet on body weight (Figure 1; week 18: F(2,27)=13.05, p<0.001; week 24: F(2,27)=16.96, p<0.001; week 26: F(2,27)=13.99, p<0.001; week 28: F(2,27)=13.05, p<0.001). Post hoc analysis revealed that HFHE and FCHE rats had significantly higher body weights than Chow controls (Figure 1; p<0.001, all time points). There were no statistical differences in body weight between the two obese groups at any time point.
3.2 Effects of dopamine D1R and D2R antagonism on sucrose intake
Sucrose intake was reduced by SCH23390 in all groups (Figure 2a). Raclopride reduced sucrose intake in HFHE rats, but was much less effective in Chow and FCHE rats (Figure 2b). The repeated measures ANOVA showed an overall effect of Drug (F(1,24)=8.8446, p<0.01), Dose (F(4,96)=27.1269, p<0.001), and a Dose by Drug interaction (F(4,96)=2.9799, p<0.05). Whereas the overall effect of Diet was not significant (F(1,24)=2.5787, p=0.09), post hoc comparisons did show significant differences of raclopride treatment between HFHE and Chow groups (p<0.05) and between HFHE and FCHE groups (p<0.05).
Figure 2. Changes in sucrose intake following dopamine receptor antagonists.

All data depicted as a reduction from baseline intake referring to vehicle injection (set at 1.0 or 100% on y-axis). A. Sucrose intake was significantly reduced by SCH23390 in all groups, with the most potent reduction occurring in HFHE rats. B. Raclopride reduced sucrose intake in HFHE rats at all doses tested and reduced intake by FCHE rats only at the highest tested dose. None of the doses significantly reduced sucrose intake in Chow rats. * p<0.05; **p<0.01; *** p<0.001 compared to vehicle.
Post hoc analysis revealed that SCH23390 was significantly more effective at reducing sucrose intake overall compared to raclopride (p<0.01). SCH23390 suppressed sucrose intake in HFHE rats at all doses tested and suppressed intake in FCHE and Chow rats at 200 nmol and higher doses (Figure 2a). Sucrose intake was suppressed in HFHE rats by all doses of raclopride, but only the highest dose reduced sucrose intake significantly in FCHE rats, while none of the doses suppressed sucrose intake by Chow rats (Figure 2b).
Analysis of the ID50 (Table 2) revealed no effect of Diet (F(2,24)=0.576, p=0.57) or Drug (F(1,24)=2.988, p=0.09), despite apparent differences in the ID50 for raclopride. This lack of an effect could be due to the substantial variance within groups.
Table 2.
Effectiveness of dopamine receptor antagonists expressed as by ID50. The ID50 represents the dose at which intake would be reduce to 50% of baseline (vehicle). No differences were observed between groups for SCH23390. In contrast, raclopride was more effective at reducing fructose intake in HFHE and FCHE rats compared to lean Chow controls.
| Diet effect on ID50 (nmol/kg) | ||||
|---|---|---|---|---|
| HFHE | FCHE | Chow | ||
| SCH 23390 (D1R) |
Sucrose | 384 ± 58 | 427 ± 160 | 325 ± 89 |
| Fructose | 175 ± 164 | 365 ± 54 | 307 ± 47 | |
|
| ||||
| Raclopride (D2R) |
Sucrose | 678 ± 233 | 2114 ± 1345 | 1436 ± 715 |
| Fructose | 608 ± 115* | 516 ± 99* | 1677 ± 1007 | |
3.3 Effects of dopamine D1R and D2R antagonism on fructose intake
SCH23390 reduced fructose intake in all groups (Figure 3a). Raclopride, on the other hand, only reduced intake significantly in the FCHE group (Figure 3b). The repeated measures ANOVA revealed an overall effect of Drug (F(1,24)=5.7400, p<0.05), Dose (F(4,96)=33.9351, p<0.001) and a significant Dose by Drug interaction (F(4,96)=3.0296, p<0.05) but no effect of Diet (F(2,24)=1.5205, p=0.24). Again, however, post hoc analyses showed a significant difference of raclopride treatment between HFHE and FCHE groups (p<0.05).
Figure 3. Changes in fructose intake following administration of dopamine receptor antagonists.
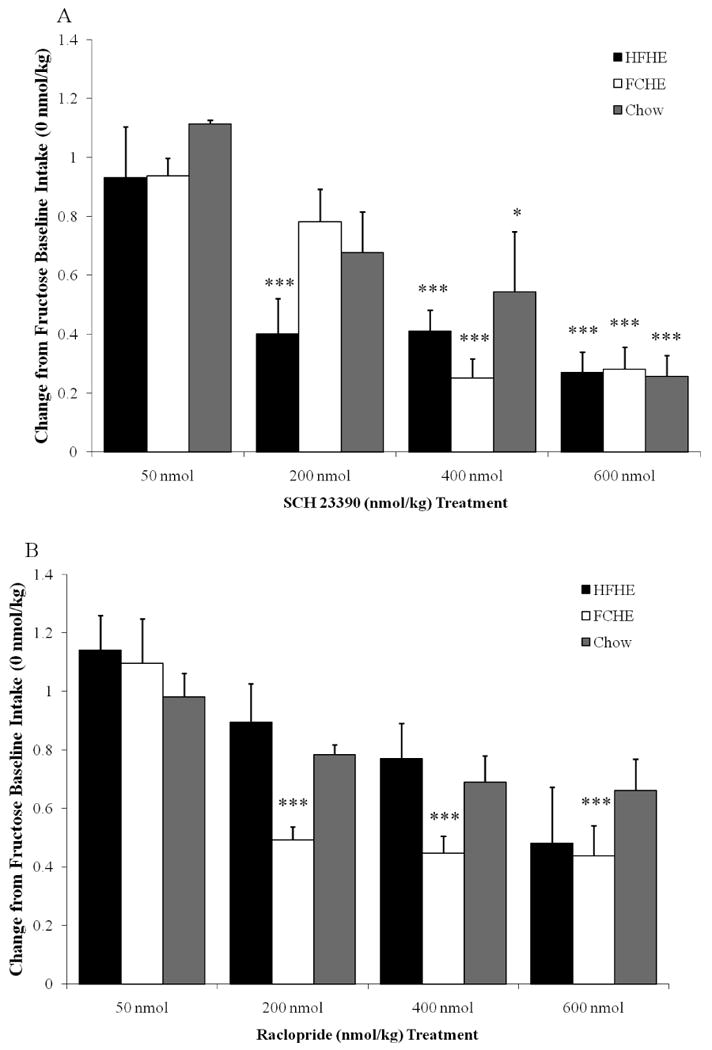
All data depicted as a reduction from baseline intake referring to vehicle injection (set at 1.0 or 100% on y-axis). A. Fructose intake was significantly reduced by SCH23390. B. Raclopride reduced fructose intake in FCHE rats, but failed to reduce intake in HFHE or Chow rats at any dose. * p<0.05; **p<0.01; *** p<0.001 compared to vehicle.
Post hoc analysis revealed that SCH23390 was overall more effective at suppressing fructose intake than raclopride (p<0.05), and did so in a dose-dependent manner (Figure 3). SCH23390 reduced intake in all diet groups at 400 and 600 nmol and reduced fructose intake as early as the dose of 200 nmol in HFHE rats (Figure 3a). Raclopride effects on fructose intake, however, were limited to FCHE rats with post hoc analysis revealing significant reductions in fructose consumption in FCHE rats at 200 nmol and higher doses, with none of the raclopride doses suppressing fructose intake in HFHE or Chow rats (Figure 3b).
ANOVA on the ID50 (Table 2) revealed an effect of Drug (F(1,24)=4.548, p<0.05) but not Diet (F(2,24)=1.495, p=0.25). SCH23390 required lower doses overall than raclopride to reduce intake to half of baseline (p<0.05). In line with the analysis on actual doses, post hoc analysis of ID50 also reveal a significantly increased sensitivity in both obese groups compared to Chow rats (p<0.05).
4. Discussion
The present study compared sensitivity to dopamine receptor blockade in reducing intake of two palatable carbohydrate solutions, sucrose or fructose, in two dietary obese animal models. We used two diets to mimic the chronic consumption of either a diet predominantly high in fat (HFHE), or a fat-sugar combination diet (FCHE), as seen in the Western diet [25]. As expected, both diets produced substantial weight gain and adiposity beginning at 12 weeks, with a continued increase in body weight throughout the experiment (Figure 1). The groups were then compared to age-matched chow-fed controls in their relative sensitivity to D1 and D2 receptor subtype-specific blockade with SCH23390 or raclopride, respectively. We found that blockade of the D1 receptors reduced both sucrose and fructose intake in all diet groups. Regardless of whether rats were consuming sucrose or fructose solutions, HFHE rats responded to slightly lower doses of SCH23390 compared to their obese FCHE or lean Chow counterparts (Figure 2a, 3a). This apparent increase in sensitivity to dopamine D1 receptor antagonism by HFHE rats was also observed following D2 receptor blockade during the sucrose test. Indeed, HFHE rats responded to all raclopride doses with reductions in sucrose intake, while FCHE rats only responded to the highest dose, and Chow rats showed no significant suppression of sucrose consumption following raclopride treatments (Figure 3b). Interestingly however, HFHE rats did not reduce fructose intake following raclopride treatment. Instead, raclopride significantly suppressed fructose intake only in FCHE rats. An increased sensitivity to the dopamine receptor antagonists is indicative of reduced dopamine signaling, i.e. due to fewer receptors, a reduced competition from endogenous DA at the receptor sites, or a combination of both. In fact there is evidence that either mechanism may be applicable to our model. For example, exposure to high fat diets even before birth may result in decreased D2Rs [36]. Furthermore, eating high fat food has shown to decrease natural or electrically evoked dopamine release, and attenuate dopamine turnover [37-39]. Whereas the underlying mechanism warrants further investigations, our data together with these and other previous observations support the notion that eating certain foods – potentially independent of obesity - may result in changes within the dopamine system reminiscent of neuroplasticity to drug of abuse [40]. In fact, recent research suggests that high fat diets increases sensitization to drugs acting on dopamine systems [41, 42].
Previous investigations in lean rats have shown differential efficacy of D1 and D2 receptor blockade to reduce carbohydrate intake using concentrations consistent with those used in the present study [31-33, 43]. These effects are believed to be partially mediated by areas of the brain involved in food reward, and D2 receptors in these areas may be especially susceptible to changes caused by obesity [31, 33, 44-46]. The present study expanded upon the findings of dopamine receptor modulation of carbohydrate intake in lean rats and compliments those studies showing lasting plasticity in the reward system in obesity. Whereas complexity of the systems and factors that may influence such interplay (acute control of intake by a chronically altered system) evidently increases individual variances and hence diminished interaction effects in the overall ANOVAs, the direct (post hoc) comparisons of dose-response effects did reveal differential sensitivity to isomolar doses of receptor antagonist between the diet groups. Changes affecting the D2Rs specifically appeared to be dependent upon the content of carbohydrates also present in the high fat diets, indicating the macronutrient content of the diets could differentially alter the reward system.
The differential effects of sensitivity to raclopride in the sucrose test could be due to the presence of sucrose in the diets. Although both obesogenic diets contained some sucrose, the FCHE diet contained 23% more sucrose than the HFHE diet. Thus the lack of a response to raclopride in the sucrose challenge by FCHE rats, but not HFHE rats, could have been due to the enhanced exposure to sucrose in the HFHE diet. However, neither obesogenic diet contained fructose, yet differences were observed in the responses of the obesogenic diet groups to raclopride in the fructose test as well. Furthermore, no sucrose was present in the Chow diet, yet responses by the Chow group to raclopride in the sucrose test were more akin to the responses made by FCHE than HFHE rats. This indicates that other factors may be underlying the differential responses to raclopride treatment as a function of diet and test carbohydrate.
Alternative explanations may include differential neural and hormonal postingestive effects exerted by fructose and sucrose. Whereas the exact mechanisms remain obscure, there is increasing evidence supporting this notion [47, 48]. In this context, the possibility that the two diets altered sucrose and fructose preferences differently as a result of their differential effects on oral and gastrointestinal signals upstream to the reward system cannot be excluded and warrants further investigation.
Obesity and palatable foods have independently been implied to alter dopamine signaling [3, 45, 49, 50], and therefore could also account for the differential responding observed in the present study. Indeed, our data support previous findings showing that dopamine D2R signaling is reduced in obesity [45, 50]. However, the novel finding of the present study was that the nature of this relationship may be dependent on the macronutrient content of the obesogenic diets rather than obesity or its associated complications. An additional major finding was the differences seen in the efficacy of D2R antagonists between test carbohydrates. We noted a trend in our data that fructose intake appeared to be more tightly controlled by D2Rs than sucrose intake, leading one to question how the intake of different carbohydrates may be differentially regulated, and if reward elicited by different carbohydrates may recruit varying mechanisms. Previous data has indicated that sucrose and fructose intake produce dissimilar physiological responses. Sucrose has been shown to produce conditioned effects based on both its taste and post-ingestive properties [28, 51, 52] while fructose appears to exert behaviorally relevant stimulation exclusively by its taste and not by reinforcing post-ingestive effects [27, 53]. Therefore, responsiveness of reward circuitries to fructose may remain intact even when feedback elicited by sucrose becomes compromised due to impairments secondary to obesity (e.g. reduced insulin/leptin sensitivity). The opposite may also be true: a counter-regulatory response to curb sucrose intake may fail to check fructose intake. Future studies in humans are needed to investigate whether preferences for foods rich in fructose would actually increase with obesity, or if relative sucrose and fructose preferences are different in obese patients who are also diabetic.
While effects of sucrose on dopamine have been extensively investigated [3, 19, 43, 50], less is known of the interaction between fructose and the dopamine reward system, although early reports from the Hoebel lab indicate that fructose may produce its own unique physiological responses [24]. The present study adds a further piece of information to this complex puzzle suggesting that diets of different macronutrient content may differentially alter dopamine control of fructose intake. Further investigation is required to fully understand the underlying mechanisms by which dietary fat and sugar may influence gut-brain signaling and elicit changes within the brain.
5. Conclusions
This study demonstrates that obesogenic (high-energy) diets varying in fat and carbohydrate content, rather than obesity itself, may differentially increase sensitivity to D1 and D2 receptor antagonists in reducing carbohydrate intake. This finding is compatible with the general notion that dopamine signaling in dietary obesity is blunted, and suggests a novel relationship between diets and central dopamine effects. An additional major finding was that the diets differentially altered the potency of dopamine receptor antagonists in suppressing sucrose and fructose intake. Compared to normal (low fat) or high fat, high carbohydrate diets, obesity produced by a very high fat but low sugar diet resulted in increased sensitivity to both D1 and D2 receptor antagonism in reducing sucrose intake, but D2 receptor control of fructose intake was preserved. In contrast, rats fed a high-energy diet with a combination of high dietary fat and carbohydrate demonstrated enhanced D2 receptor regulation of fructose intake. Thus, it appears that dietary history may alter the development of dopamine deficits previously attributed to obesity in general. The present data also suggest that these particularities of dopamine plasticity may influence how certain carbohydrates, such as fructose and sucrose, exert their rewarding effects. Such differences could explain some of the variation in the success rates of different anti-obesity treatments and therapies. Further studies are required to test the applicability of these findings to humans and investigate underlying mechanisms.
Highlights
High-energy diets independent of macronutrient content are potent to cause obesity.
Diet composition appears to differentially alter dopamine receptor sensitivity.
D1 receptor blockade reduced sucrose and fructose intake in lean and obese rats.
D2 receptor blockade reduced sucrose intake in high fat fed, but not lean rats.
D2 receptor blockade reduced fructose intake in fat-sugar fed rats only.
Acknowledgments
This research was supported by National Institute of Diabetes & Digestive & Kidney Diseases Grant DK080899, National Institute on Deafness and Other Communication Disorders Grant DC000240, and The Jane B. Barsumian Trust Fund. The authors thank Mr. N.K. Acharya for his excellent help with the maintenance of rats and performing the NMR assays.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Hernandez L, Hoebel BG. Feeding and hypothalamic stimulation increase dopamine turnover in the accumbens. Physiology & Behavior. 1988;44:599–606. doi: 10.1016/0031-9384(88)90324-1. [DOI] [PubMed] [Google Scholar]
- 2.Hernandez L, Hoebel BG. Food reward and cocaine increase extracellular dopamine in the nucleus accumbens as measured by microdialysis. Life Sciences. 1988;42:1705–12. doi: 10.1016/0024-3205(88)90036-7. [DOI] [PubMed] [Google Scholar]
- 3.Avena NM, Rada P, Moise N, Hoebel BG. Sucrose sham feeding on a binge schedule releases accumbens dopamine repeatedly and eliminates the acetylcholine satiety response. Neuroscience. 2006;139:813–20. doi: 10.1016/j.neuroscience.2005.12.037. [DOI] [PubMed] [Google Scholar]
- 4.Rada P, Avena NM, Hoebel BG. Daily bingeing on sugar repeatedly releases dopamine in the accumbens shell. Neuroscience. 2005;134:737–44. doi: 10.1016/j.neuroscience.2005.04.043. [DOI] [PubMed] [Google Scholar]
- 5.Ahlskog JE, Randall PK, Hernandez L, Hoebel BG. Diminished amphetamine anorexia and enhanced fenfluramine anorexia after midbrain 6-hydroxydopamine. Psychopharmacology. 1984;82:118–21. doi: 10.1007/BF00426393. [DOI] [PubMed] [Google Scholar]
- 6.Hernandez L, Hoebel BG. Overeating after midbrain 6-hydroxydopamine: Prevention by central injection of selective catecholamine reuptake blockers. Brain Research. 1982;245:333–43. doi: 10.1016/0006-8993(82)90816-2. [DOI] [PubMed] [Google Scholar]
- 7.Ahlskog J. Feeding response to regulatory challenges after 6-hydroxydopamine injection into the brain noradrenergic pathways. Physiology & Behavior. 1976;17:407–11. doi: 10.1016/0031-9384(76)90099-8. [DOI] [PubMed] [Google Scholar]
- 8.Hoebel BG, Hernandez L, Monaco A, Miller W. Amphetamine-induced overating and overweight in rats. Life Sciences. 1981;28:77–82. doi: 10.1016/0024-3205(81)90368-4. [DOI] [PubMed] [Google Scholar]
- 9.Volkow ND, Wang GJ, Baler RD. Reward, dopamine and the control of food intake: implications for obesity. Trends in Cognitive Sciences. 15:37–46. doi: 10.1016/j.tics.2010.11.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Stice E, Spoor S, Bohon C, Small DM. Relation Between Obesity and Blunted Striatal Response to Food Is Moderated by TaqIA A1 Allele. Science. 2008;322:449–52. doi: 10.1126/science.1161550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Hoebel BG. Phamacologic control of feeding. Ann Rev Pharmacol Toxicol. 1977;17 doi: 10.1146/annurev.pa.17.040177.003133. [DOI] [PubMed] [Google Scholar]
- 12.Bouchard C. Current understanding of the etiology of obesity: genetic and nongenetic factors. The American Journal of Clinical Nutrition. 1991;53:1561S–5S. doi: 10.1093/ajcn/53.6.1561S. [DOI] [PubMed] [Google Scholar]
- 13.Vogele C. Etiology of Obesity. In: Munsch S, Beglinger C, editors. Obesity and binge eating disorder. Switzerland: S. Karger; 2005. pp. 62–73. [Google Scholar]
- 14.Weinsier RL, Hunter GR, Heini AF, Goran MI, Sell SM. The etiology of obesity: relative contribution of metabolic factors, diet, and physical activity. The American Journal of Medicine. 1998;105:145–50. doi: 10.1016/s0002-9343(98)00190-9. [DOI] [PubMed] [Google Scholar]
- 15.Small DM. Individual differences in the neurophysiology of reward and the obesity epidemic. Int J Obes. 2009;33:S44–S8. doi: 10.1038/ijo.2009.71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Archer ZA, Mercer JG. Brain responses to obesogenic diets and diet-induced obesity. Proceedings of the Nutrition Society. 2007;66:124–30. doi: 10.1017/S0029665107005356. [DOI] [PubMed] [Google Scholar]
- 17.Geiger BM, Behr GG, Frank LE, Caldera-Siu AD, Beinfeld MC, Kokkotou EG, et al. Evidence for defective mesolimbic dopamine exocytosis in obesity-prone rats. FASEB J. 2008;22 doi: 10.1096/fj.08-110759. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Volkow ND, Wang GJ, Baler RD. Reward, dopamine and the control of food intake: implications for obesity. Trends in Cognitive Sciences. 2011;15:37–46. doi: 10.1016/j.tics.2010.11.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Hajnal A, Smith GP, Norgren R. Oral sucrose stimulation increases accumbens dopamine in the rat. Am J Physiol Regul Integr Comp Physiol. 2004;286:R31–7. doi: 10.1152/ajpregu.00282.2003. [DOI] [PubMed] [Google Scholar]
- 20.Liang NC, Hajnal A, Norgren R. Sham feeding corn oil increases accumbens dopamine in the rat. American Journal of Physiology - Regulatory, Integrative and Comparative Physiology. 2006;291:R1236–R9. doi: 10.1152/ajpregu.00226.2006. [DOI] [PubMed] [Google Scholar]
- 21.Avena NM, Rada P, Hoebel BG. Sugar and Fat Bingeing Have Notable Differences in Addictive-like Behavior. J Nutr. 2009;139:623–8. doi: 10.3945/jn.108.097584. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Shahkhalili Y, Mace K, Moulin J, Zbinden I, Acheson KJ. The Fat:Carbohydrate Energy Ratio of the Weaning Diet Programs Later Susceptibility to Obesity in Male Sprague Dawley Rats. Journal of Nutrition. 2011;141:81–6. doi: 10.3945/jn.110.126557. [DOI] [PubMed] [Google Scholar]
- 23.van den Heuvel JK, van Rozen AJ, Adan RAH, la Fleur SE. An overview on how components of the melanocortin system respond to different high energy diets. European Journal of Pharmacology. 2011 doi: 10.1016/j.ejphar.2010.12.019. Epub ahead of print. [DOI] [PubMed] [Google Scholar]
- 24.Bocarsly ME, Powell ES, Avena NM, Hoebel BG. High-fructose corn syrup causes characteristics of obesity in rats: Increased body weight, body fat and triglyceride levels. Pharmacology Biochemistry and Behavior. 2010;97:101–6. doi: 10.1016/j.pbb.2010.02.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Reedy J, Krebs-Smith SM. Dietary Sources of Energy, Solid Fats, and Added Sugars among Children and Adolescents in the United States. Journal of the American Dietetic Association. 2010;110:1477–84. doi: 10.1016/j.jada.2010.07.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Sclafani A. Carbohydrate taste, appetite, and obesity: An overview. Neuroscience & Biobehavioral Reviews. 1987;11:131–53. [PubMed] [Google Scholar]
- 27.Ackroff K, Touzani K, Peets TK, Sclafani A. Flavor preferences conditioned by intragastric fructose and glucose: differences in reinforcement potency. Physiology & Behavior. 2001;72:691–703. doi: 10.1016/s0031-9384(01)00442-5. [DOI] [PubMed] [Google Scholar]
- 28.Sclafani A, Thompson B, Smith JC. The Rat's Acceptance and Preference for Sucrose, Maltodextrin, and Saccharin Solutions and Mixtures. Physiology & Behaviour. 1998;63:499–503. doi: 10.1016/s0031-9384(97)00478-2. [DOI] [PubMed] [Google Scholar]
- 29.Hajnal A, Norgren R. Repeated access to sucrose augments dopamine turnover in the nucleus accumbens. Neuroreport. 2002;13:2213–6. doi: 10.1097/00001756-200212030-00010. [DOI] [PubMed] [Google Scholar]
- 30.Weatherford SC, Greenberg D, Gibbs J, Smith GP. The potency of D-1 and D-2 receptor antagonists is inversely related to the reward value of sham-fed corn oil and sucrose in rats. Pharmacology Biochemistry and Behavior. 1990;37:317–23. doi: 10.1016/0091-3057(90)90341-e. [DOI] [PubMed] [Google Scholar]
- 31.Bernal SY, Dostova I, Kest A, Abayev Y, Kandova E, Touzani K, et al. Role of dopamine D1 and D2 receptors in the nucleus accumbens shell on the acquisition and expression of fructose-conditioned flavor-flavor preferences in rats. Behavioural Brain Research. 2008;190:59–66. doi: 10.1016/j.bbr.2008.02.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Baker RM, Shah MJ, Sclafani A, Bodnar RJ. Dopamine D1 and D2 antagonists reduce the acquisition and expression of flavor-preferences conditioned by fructose in rats. Pharmacology Biochemistry and Behavior. 2003;75:55–65. doi: 10.1016/s0091-3057(03)00039-x. [DOI] [PubMed] [Google Scholar]
- 33.Bernal S, Miner P, Abayev Y, Kandova E, Gerges M, Touzani K, et al. Role of amygdala dopamine D1 and D2 receptors in the acquisition and expression of fructose-conditioned flavor preferences in rats. Behavioural Brain Research. 2009;205:183–90. doi: 10.1016/j.bbr.2009.06.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Smith GP. Accumbens dopamine mediates the rewarding effect of orosensory stimulation by sucrose. Appetite. 2004;43:11–3. doi: 10.1016/j.appet.2004.02.006. [DOI] [PubMed] [Google Scholar]
- 35.Hajnal A, De Jonghe BC, Covasa M. Dopamine D2 receptors contribute to increased avidity for sucrose in obese rats lacking CCK-1 receptors. Neuroscience. 2007;148:584–92. doi: 10.1016/j.neuroscience.2007.06.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Naef L, Moquin L, Dal Bo G, Giros B, Gratton A, Walker CD. Maternal high-fat intake alters presynaptic regulation of dopamine in the nucleus accumbens and increases motivation for fat rewards in the offspring. Neuroscience. 2010;176:225–36. doi: 10.1016/j.neuroscience.2010.12.037. [DOI] [PubMed] [Google Scholar]
- 37.Rada P, Bocarsly ME, Barson JR, Hoebel BG, Leibowitz SF. Reduced accumbens dopamine in Sprague-Dawley rats prone to overeating a fat-rich diet. Physiology & Behavior. 2010;101:394–400. doi: 10.1016/j.physbeh.2010.07.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Geiger BM, Haburcak M, Avena NM, Moyer MC, Hoebel BG, Pothos EN. Deficits of mesolimbic dopamine neurotransmission in rat dietary obesity. Neuroscience. 2009;159:1193–9. doi: 10.1016/j.neuroscience.2009.02.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Davis JF, Tracy AL, Schurdak JD, Tschöp MH, Lipton JW, Clegg DJ, et al. Exposure to Elevated Levels of Dietary Fat Attenuates Psychostimulant Reward and Mesolimbic Dopamine Turnover in the Rat. Behavioral Neuroscience. 2008;122:1257–63. doi: 10.1037/a0013111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Koob GF, Volkow ND. Neurocircuitry of Addiction. Neuropsychopharmacology. 2009;35:217–38. doi: 10.1038/npp.2009.110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Baladi MG, France CP. Eating high-fat chow increases the sensitivity of rats to quinpirole-induced discriminative stimulus effects and yawning. Behavioural Pharmacology. 2010;21:615–20. doi: 10.1097/FBP.0b013e32833e7e5a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.McGuire BA, Baladi MG, France CP. Eating high-fat chow enhances sensitization to the effects of methamphetamine on locomotion in rats. European Journal of Pharmacology. 2011;658:156–9. doi: 10.1016/j.ejphar.2011.02.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Tyrka A, Smith GP. SCH23390, but not raclopride, decreases intake of intraorally infused 10% sucrose in adult rats. Pharmacology Biochemistry and Behavior. 1993;45:243–6. doi: 10.1016/0091-3057(93)90113-8. [DOI] [PubMed] [Google Scholar]
- 44.Volkow ND, Wang GJ, Fowler JS, Telang F. Overlapping neuronal circuits in addiction and obesity: evidence of systems pathology. Philos Trans R Soc Lond B Biol Sci. 2008;363:3191–200. doi: 10.1098/rstb.2008.0107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Johnson PM, Kenny PJ. Dopamine D2 receptors in addiction-like reward dysfunction and compulsive eating in obese rats. Nat Neurosci. 2010;13:635–41. doi: 10.1038/nn.2519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Wang GJ, Volkow ND, Logan J, Pappas NR, Wong CT, Zhu W, et al. Brain dopamine and obesity. The Lancet. 2001;357:354–7. doi: 10.1016/s0140-6736(00)03643-6. [DOI] [PubMed] [Google Scholar]
- 47.Ackroff K, Sclafani A. Rats' preferences for high fructose corn syrup vs. sucrose and sugar mixtures. Physiology & Behavior. 2011;102:548–52. doi: 10.1016/j.physbeh.2011.01.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Glendinning JI, Breinager L, Kyrillou E, Lacuna K, Rocha R, Sclafani A. Differential effects of sucrose and fructose on dietary obesity in four mouse strains. Physiology & Behavior. 2010;101:331–43. doi: 10.1016/j.physbeh.2010.06.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Hajnal A, Margas WM, Covasa M. Altered dopamine D2 receptor function and binding in obese OLETF rat. Brain Res Bull. 2008;75:70–6. doi: 10.1016/j.brainresbull.2007.07.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Bello NT, Lucas L, Hajnal A. Repeated sucrose access influences dopamine D2 receptor density in the striatum. NeuroReport. 2002;13:1565–8. doi: 10.1097/00001756-200208270-00017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Ackroff K. Learned flavor preferences. The variable potency of post-oral nutrient reinforcers. Appetite. 2008;51:743–6. doi: 10.1016/j.appet.2008.05.059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Bonacchi KB, Ackroff K, Sclafani A. Sucrose taste but not Polycose taste conditions flavor preferences in rats. Physiology & Behavior. 2008;95:235–44. doi: 10.1016/j.physbeh.2008.06.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Sclafani A, Ackroff K. Glucose- and fructose-conditioned flavor preferences in rats: Taste versus postingestive conditioning. Physiology & Behavior. 1994;56:399–405. doi: 10.1016/0031-9384(94)90213-5. [DOI] [PubMed] [Google Scholar]