

Abstract
Current models of parent–offspring communication do not explicitly predict the effect of parental food supply on offspring demand (ESD). However, existing theory is frequently interpreted as predicting a negative ESD, such that offspring beg less when parental supply is high. While empirical evidence largely supports this interpretation, several studies have identified the opposite case, with well-fed offspring begging more than those in poorer condition. Here, we show that signalling theory can give rise to either a negative or a positive ESD depending on the precise form of costs and benefits. Introducing variation among parents in the cost of care, we show that the ESD may change sign depending upon the quantitative relation between two effects: (i) decreased supply leads to increased begging because of an increase in marginal fitness benefit of additional resources to offspring, (ii) decreased supply leads to reduced begging because it is associated with a decrease in parental responsiveness, rendering begging less effective. To illustrate the interplay between these two effects, we show that Godfray's seminal model of begging yields a negative ESD when care is generally cheap, because the impact of supply on the marginal benefits of additional resources then outweighs the associated changes in parental responsiveness to begging. By contrast, the same model predicts a positive ESD when care is generally costly, because the impact of care costs on parental responsiveness then outweighs the change in marginal benefits.
Keywords: parent–offspring conflict, animal communication, parental care, signalling, supply and demand
1. Introduction
In past decades, the study of parent–offspring communication has exploited game theory to model the evolution of parental and offspring behaviour (e.g. [1,2]) in the face of the conflict between them [3]. Godfray's seminal ‘honest-signalling’ model [2] assumed that offspring in better or worse ‘condition’ will enjoy smaller or greater fitness gains from receiving resources, respectively, and predicted that begging should consequently be a reliable signal of their condition. Since then, many models have extended the honest-signalling framework to incorporate more offspring and different assumptions about the control of resource delivery and allocation, and have explored its limitations [4–9]. One of the basic predictions from these models is that parents should respond positively to their offspring's demands, i.e. that there will be a positive effect of demand on supply (EDS). Indeed, many empirical studies corroborate this prediction ([10–15], reviewed in [16]), and there is recent evidence for the benefits to parents of responding to begging in this way [17,18]. Here, we examine the complementary effect, namely the effect of parental supply on offspring demand (hereby ESD), which has also received much empirical attention.
Many experiments have shown that feeding offspring causes a reduction in begging levels, while food deprivation causes offspring to increase their begging (reviewed in [13,14,16]). However, some studies, usually considering longer periods of time, have failed to find such a negative ESD, and have even suggested a positive effect such that offspring demand levels increase with parental supply [19–23]. To explain such counterintuitive results, some authors have suggested explanations outside of the honest-signalling framework. For example, low-ranked nestlings, in poorer condition, may learn that their begging is less effective and therefore reduce their begging level, whereas nestlings receiving a more generous supply learn to increase their efforts [13,21–24]. By contrast, in an experimental study designed to test this suggestion with house-sparrow nestlings, the data supported a different learning mechanism, one leading to a negative effect of the level of feeding on begging intensity [25]. Rather than learning to reduce their begging levels after high begging levels had proved unsuccessful, these nestlings first required positive experience with low begging levels in order to do so. Since alternative learning mechanisms have been suggested, leading either to a positive or a negative effect of supply on demand, and given the conflicting evidence regarding the effect of supply on demand itself, we were interested in examining what current theory predicts. As is stressed below, existing analyses of parent–offspring communication have provided very few predictions regarding the ESD, and therefore could not be used to predict what the adaptive effect of learning would be in this case.
Classical models of scramble competition assume a negative ESD (e.g. [1,26,27]), but do not allow for evolution of this aspect of behaviour. More recently, the impact of the ESD (as well as the EDS) has been explored in behavioural models studying the proximate process of parent–offspring interaction ([28–30], reviewed in [31]). However, these effects have once again been included as fixed parameters rather than as evolving strategies [30,31]. Finally, another type of (parent–offspring conflict) ‘resolution models’, the ‘honest-signalling’ models mentioned above, allow for the evolution of both parental and offspring behavioural strategies [2,4,7,8]. The parental strategy specifies supply given demand; but, since parental supply does not affect ‘condition’ (which influences begging demand), the offspring strategy is ‘the effect of condition on demand’ rather than ‘the effect of supply on demand’ (but see [32]). Exploring how a parent should divide a fixed amount of resources between its two offspring, Godfray [4] predicted that higher fixed amounts lead to lower begging intensities (i.e. a negative ESD). However, it is unclear whether this result continues to hold if parents can also adjust the amount of resources supplied. Thus, we are left with no clear theoretical prediction as to what the ESD should be. Instead, the reasonable (implicit) assumption that supply will positively affect any imaginable aspect of ‘condition’ has led to an interpretation of the models according to which they predict a negative effect of parental feeding supply on offspring demand, i.e. reduced begging when feeding supply is high [13,14,16,33].
Our objective here was to determine what the ‘honest-signalling’ modelling framework predicts regarding the effect of supply on demand, when variation in supply can be attributed to variation in the costs of care to parents. To that end, we compared evolutionarily stable strategy (ESS) levels of supply and demand under different conditions, assuming that differences in supply reflect underlying differences in parental costs of care (i.e. in the extent to which parents' future reproductive prospects are damaged by provisioning current young [34]). We also assume that offspring are informed about such differences by prenatal cues that reflect parental quality or postnatal environmental cues that reflect local foraging opportunities. We first demonstrate, under general assumptions, that there are two distinct effects of parental costs and consequent provisioning on begging, and then show, using Godfray's seminal model [2] as an illustrative case, that the balance between these contrasting effects can lead to either a positive or a negative ESD, depending on the precise form of costs and benefits.
2. The model
Following Godfray's model [2], we consider one parent rearing a single offspring, with opportunities for future reproduction. The future fitness of the offspring, denoted Wo, is assumed to depend on its investment in costly solicitation, x, on the amount of resources its parent provides, y, and on its condition c (which is unknown to the parent). We assume that
| 2.1 |
where 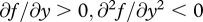 and
and 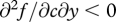 . Increasing quantities of food thus lead to greater future fitness but yield diminishing returns, and these returns diminish faster for offspring in better condition. Begging, on the other hand, entails an additive cost that is independent of condition.
. Increasing quantities of food thus lead to greater future fitness but yield diminishing returns, and these returns diminish faster for offspring in better condition. Begging, on the other hand, entails an additive cost that is independent of condition.
Parental future fitness, denoted Wp, depends on resources invested in the current offspring and on the costs of care, determined by the parameter m, which may vary from one parent to another, but which we assume is known to both parent and offspring. This parameter subsumes the parent's foraging ability, its level of energetic reserves, the quality of its territory, the current state of the environment and any other factor that influences how current provisioning of resources damages parental future reproduction. Consequently, offspring might acquire their information about the costs of care through prenatal cues of parental quality [35] or through postnatal environmental cues such as weather or predator presence that reflect the likely success of parental foraging efforts. We will suppose that
| 2.2 |
where 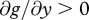 ,
, 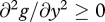 , and
, and 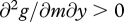 . G thus denotes expected future reproductive success in the absence of provisioning, and g(m,y) the fitness cost incurred through feeding the current offspring; this cost increases with the amount supplied, in a linear or accelerating manner, but does so less rapidly when the cost of care m is low.
. G thus denotes expected future reproductive success in the absence of provisioning, and g(m,y) the fitness cost incurred through feeding the current offspring; this cost increases with the amount supplied, in a linear or accelerating manner, but does so less rapidly when the cost of care m is low.
Lastly, we assume that offspring are related to future offspring of the parent by coefficient r (for simplicity, we assume that costs incurred by the parent do not impact on the fitness of its mate).
We focus on behaviour at a separating equilibrium, at which the begging intensity of the offspring is a strictly decreasing function of its condition, denoted x*(m,c) (where 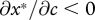 ), and where the quantity of food supplied by the parent is consequently an increasing function of offspring begging, denoted y*(m,x) (where
), and where the quantity of food supplied by the parent is consequently an increasing function of offspring begging, denoted y*(m,x) (where 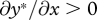 ). Both begging and provisioning at such an equilibrium also depend upon the cost of care to the parent m. It is easy to show (appendix A) that the equilibrium level of provisioning by a parent with cost of care m to an offspring in condition c, denoted
). Both begging and provisioning at such an equilibrium also depend upon the cost of care to the parent m. It is easy to show (appendix A) that the equilibrium level of provisioning by a parent with cost of care m to an offspring in condition c, denoted 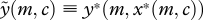 , is a decreasing function of m. That is, parents who experience higher costs of care provide less food. The question is, whether offspring of such parents, which receive a smaller supply, beg more or less strongly.
, is a decreasing function of m. That is, parents who experience higher costs of care provide less food. The question is, whether offspring of such parents, which receive a smaller supply, beg more or less strongly.
(a). Two distinct effects of parental costs of care on offspring begging
At equilibrium, offspring must beg at an optimal level that maximizes their inclusive fitness payoff given the parental response strategy. Formally,
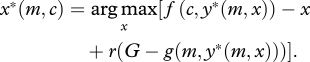 |
This implies that the marginal inclusive fitness benefits and costs of begging for offspring must balance one another.
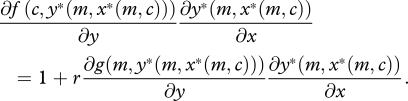 |
2.3 |
The left-hand side of equation (2.3) corresponds to the marginal benefit of begging, which is equal to the marginal benefit of provisioning (to the offspring) multiplied by the marginal change of provisioning in response to begging. The right-hand side of equation (2.3) gives the marginal cost of begging, which is equal to the direct marginal cost incurred by the offspring plus r times the indirect marginal cost incurred by the parent (owing to increased provisioning).
At the same time, the marginal fitness benefits and costs of provisioning for parents must balance one another, so that
| 2.4 |
where the left-hand side of equation (2.4) corresponds to the marginal benefit of provisioning in terms of survival of the current offspring, and the right-hand side to the marginal cost of provisioning in terms of lost future offspring. Using equation (2.4), we can simplify equation (2.3) by substituting the marginal benefit of additional provisioning to the offspring in place of the marginal cost to the parent, to obtain
| 2.5 |
i.e. the marginal benefit of begging, when multiplied by the factor (1 − r), must balance the direct marginal cost incurred by the offspring. In turn, this implies (given our assumptions) that
| 2.6 |
or, in words, that offspring beg at a level that maximizes their direct fitness payoff, when the benefits of begging are devalued by a factor of (1 − r) (reflecting the indirect cost of imposing a greater burden of care on the parent).
We want to know whether offspring beg more intensely when parents pay a lower cost for care. From equation (2.6), it follows that the impact of a small increase in parental cost of care on begging, 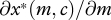 , is of the same sign as
, is of the same sign as
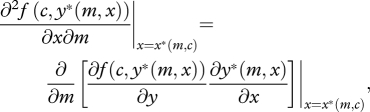 |
2.7 |
the impact of parental cost of care on the marginal direct benefit of begging (evaluated at the equilibrium level of begging intensity), which is equal to the marginal benefit of provisioning (to the offspring) multiplied by the marginal change in provisioning in response to begging.
Now, as stated above (and shown in appendix A), as parental cost of care increases, the parent supplies less food, so that the first term in the bracketed product in equation (2.7), the marginal benefit of provisioning (to the offspring), increases. This tends to give rise to a negative correlation between supply and demand—because reduced supply by parent who experiences higher costs puts offspring (of all condition levels) in a state in which extra resources are worth more, increasing the marginal benefit of begging. However, this effect alone does not necessarily guarantee a negative correlation between supply and demand, because we must also consider the second term in the bracketed product in equation (2.5), the marginal change in provisioning in response to begging. As we show below, this may decrease with parental cost of care, tending to give rise to a positive correlation between supply and demand—when parents who experience higher costs are less responsive to a change in offspring demand, the marginal benefit of begging may decrease with the cost of care. Whether the model predicts a negative or a positive correlation overall between supply and demand may thus depend on the quantitative relations between these two effects, as demonstrated below.
(b). An illustrative example
In this section, we focus on the particular cost and benefit functions adopted by Godfray [2], which we modify by introducing variation among parents in the cost of provisioning, That is, we assume that
(a specific functional form that, over appropriately chosen ranges of c and y, satisfies the general conditions stated above: 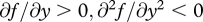 and
and 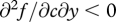 ), and that
), and that
implying that the cost of provisioning is proportional to the level of resources supplied, with the parental cost parameter, m (equivalent to γ in Godfray [2]), specifying the constant of proportionality. This illustrative case serves to show that the opposing effects described above can give rise to both positive or negative correlations between supply and demand, depending on the precise details of the model.
Figure 1 shows the ESS levels of parental supply, plotted against the ESS levels of offspring demand, as offspring condition varies, for several different levels of parental cost of care. For low costs of care (upper lines), there is a strong positive relation between parental supply and offspring demand, showing that begging is highly effective in eliciting extra resources under these circumstances. However, parental responsiveness to begging lowers as parental costs of care increase, reaching very low responsiveness for parents who experience high costs (lower lines). This makes begging less effective, and favours reduced begging intensity. Thus, high costs of care favour both reduced supply on the part of the parent and reduced demand on the part of the offspring, leading to a positive ESD.
Figure 1.

ESS levels of parental supply in our exploration of Godfray [2] plotted against ESS levels of offspring demand. Each line shows results for a specific value of the cost of parental care m, across all offspring condition levels (assuming that c is evenly distributed between 4 and 5). As the cost of parental care increases, parental supply levels decrease, but so does parental responsiveness to begging intensity.
Parental responsiveness is reduced as costs of care increase, accompanying the general reduction in supply (figure 1), and the question is which of the effects is stronger in each case. When costs of care are generally low, the increase in offspring marginal fitness gain associated with lower supply levels overcomes the decrease in begging effectiveness. Together, this leads to a net negative ESD when parental costs are relatively low, illustrated in the right side of figure 2. By contrast, when parental costs are high enough, parental responsiveness is so low (figure 1, lower lines) that the reduction in begging effectiveness associated with reduced supply now overcomes the increase in marginal gains. This results in a net positive ESD for those conditions (left side of figure 2).
Figure 2.

Mean ESS offspring begging intensity (averaging across offspring condition values, assuming that c is evenly distributed between 4 and 5) plotted against mean ESS parental supply of resources, for different values of the cost of parental care, m (ranging from 0.067 to 1.48). When parental care is cheap (right side of the graph), the model predicts a negative effect of supply on demand, while a positive effect is predicted when parental care is more costly (left side of the graph).
3. Discussion
Given the paucity of theoretical predictions about the ESD, and the conflicting empirical evidence for both positive and negative effects, we set out to explore what honest-signalling theory predicts. We have shown that increased costs of parental care will, unsurprisingly, cause a decrease in parental supply levels. Moreover, as long as offspring fitness increases with resources at a decelerating rate, this reduction in supply will cause the marginal fitness value of receiving extra resources to increase. This would give rise to a negative ESD, as has been the common interpretation of honest-signalling theory, but it is not the only effect of parental cost of care. Our results show that as the cost of care increases, parents may also become less responsive to offspring begging. In other words, in addition to the overall reduction in supply by parents who experience high costs of care, increased begging may be less effective in eliciting extra resources from such parents. Taken alone, the latter effect would lead to a positive ESD, as offspring receiving low supply should reduce their begging because it has become ineffective. Such an effect is suggested by some authors to come about through learning [13,19,22–24], while others have suggested that a positive correlation between begging and provisioning levels on a population scale may arise as a result of parent–offspring genetic co-adaptation [30].
Confusingly, therefore, we can have two contrasting general influences of the cost of parental care: (i) parental costs of care influence overall supply levels, and hence the marginal value of additional resources to the offspring, leading to a negative ESD and (ii) parental costs of care influence parental responsiveness to begging (or begging effectiveness), possibly leading to a positive ESD. Whether we should expect a negative or positive ESD will ultimately depend on which of these two factors outweighs the other. Indeed, for different ranges of parental costs of care, Godfray's original model [2] yields qualitatively different predictions regarding the ESD: when costs of care are generally low, Godfray's model predicts a negative ESD, as commonly assumed [13,14,16,33]. By contrast, when costs are generally high, the same model predicts a positive ESD. As discussed above, this positive ESD arises because begging becomes less effective as the cost of parental care increases.
Our analysis suggests that data on the sensitivity of parents to begging might help to predict or explain the effect of supply on demand. A recent study shows that banded mongoose pups that are fed by responsive escorts in good condition increase their begging in response to food deprivation more than do pups fed by less responsive escorts in worse condition [36]. These results confirm that costs of care can affect the responsiveness of the carer (in this case not a parent) to begging, and that this in turn can lead offspring to adjust the ESD (in this case probably through learning). Similarly, detailed studies that simultaneously consider all of the components in our model (i.e. costs of care, responsiveness to begging and the ESD) are still scarce, but there are a few more examples of how ecological factors likely to modify the costs of care affect responsiveness of parents to begging. In line with our predictions, crimson rosella fathers seem to be less responsive to begging when the brood is hungrier, which might suggest worse environmental conditions, and food-deprived nestlings do not increase their begging intensity [20]. By contrast, two contrasting studies of red-winged blackbirds suggest that parents may actually be more responsive to begging under worse environmental conditions [37,38], which runs counter to the predictions of our model.
One possible explanation for the contradictory findings described above is that better environmental conditions may not always be associated with lower costs of care, as illustrated in a recent study [39]. When the propensity of hihi (Notiomystis cincta) parents to breed again was increased by supplementing them with carotenoids, they supplied less food than did control parents to offspring with enhanced begging displays. This is consistent with our model because the costs of caring for the current brood, in terms of loss of future reproduction, are greater when the probability of future reproduction is higher. There is also an important implication for future studies of parent–offspring communication: in order to use the honest-signalling framework to predict how parental condition should affect provisioning, it is advisable to consider also how condition changes the relative importance of current and future reproduction. Parental costs of care and responsiveness are likely to vary for parents at different stages in their lives, or of different sexes [40,41]), for parents under different environmental conditions, and for parents from species with different life histories (determining the importance of current offspring relative to future ones, e.g. [42]).
It is also worth emphasizing that our analysis, like most previous signalling models, treats offspring condition as fixed and independent of parental supply. In such models, parents and offspring engage in a ‘one-shot’ interaction in which offspring, whose level of condition is drawn from some random distribution, advertise this by means of begging, parents respond by supplying food, and the game then ends (although see [32] for an exception). A more realistic, dynamic model would allow for repeated interactions, with provision of food during one step influencing offspring condition in subsequent steps. However, a dynamic model of this kind would be very complex, and is beyond the scope of the present paper. It seems likely that changes in condition would tend to lead to a more negative ESD, since greater supply will lead to improved condition for offspring, which favours reduced demand. However, no firm prediction is possible in the absence of a full, dynamic analysis.
Offspring may use cues and signals given by parents or siblings [43] to obtain information about parental cost of care and shape their begging levels, and to adjust the ESD accordingly. From the offspring's point of view, it is important whether supply levels drop because of (i) increased costs of parental care or (ii) increased sibling competition, as selection may favour different responses to these two factors. Here, we have focused only on the former (see [44] for a discussion of the latter). Prenatal signals help offspring adjust begging to foreseeable environmental conditions [35,45], whereas the level of competition in the nest is known to affect begging directly [12,46,47]. In addition, learning has been posited to explain results showing positive ESDs [19,22,23], but existing learning mechanisms may not allow offspring to reduce their begging when supply is low [25,48,49], possibly to prevent them from being outcompeted by siblings. In any case, we show here that both positive and negative ESDs are explicable within the honest-signalling framework. But this does not mean that it will not be beneficial for offspring to learn.
Learning may offer advantages over and above any response to prenatal cues, because it also allows offspring to adjust their response to parental supply in light of postnatal environmental changes. Of course, this presupposes that offspring can detect any such changes. We have made the simplistic assumption that offspring have full information about the costs of parental care, and can adjust their begging behaviour accordingly. In reality, their information is likely to be imperfect. A fuller model of offspring learning would allow for dynamic changes in the cost of care, which offspring cannot always detect directly, but must infer from changes in parental feeding behaviour. Such a model is, however, beyond the scope of the present paper. What we can say, on the basis of our current results, is that since both negative and positive responses to supply levels may be favoured under different conditions, learning of some kind is necessary if offspring are to respond appropriately to changes in parental supply.
Acknowledgements
We thank Matt Bell, Arnon Lotem and Miguel Rodríguez-Gironés for valuable and stimulating discussions, and two anonymous reviewers for their constructive comments. U.G. is funded by a fellowship from the Human Frontier Science Programme Organization.
Appendix A
At a separating equilibrium, begging provides full information about offspring condition, and the marginal costs and benefits of provisioning for parents must balance, just as the marginal costs and benefits of begging must balance for offspring. We therefore have
| A 1 |
We can solve equation (A 1) to find 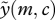 without any need to derive
without any need to derive 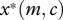 first. Since equation (A 1) holds for all values of m and c, we can also differentiate to obtain
first. Since equation (A 1) holds for all values of m and c, we can also differentiate to obtain
| A 2 |
and
| A 3 |
implying (given our assumptions that ∂2f/∂y2 < 0, ∂2g/∂y2 ≥ 0, ∂2f/∂c∂y < 0 and ∂2g/∂m∂y > 0) that provisioning decreases with both parental cost of care and with offspring condition.
References
- 1.Parker G. A., Macnair M. R. 1979. Models of parent–offspring conflict. 4. Suppression—evolutionary retaliation by the parent. Anim. Behav. 27, 1210–1235 10.1016/0003-3472(79)90068-X (doi:10.1016/0003-3472(79)90068-X) [DOI] [Google Scholar]
- 2.Godfray H. C. J. 1991. Signaling of need by offspring to their parents. Nature 352, 328–330 10.1038/352328a0 (doi:10.1038/352328a0) [DOI] [Google Scholar]
- 3.Trivers R. L. 1974. Parent–offspring conflict. Am. Zool. 14, 249–264 [Google Scholar]
- 4.Godfray H. C. J. 1995. Signaling of need between parents and young—parent–offspring conflict and sibling rivalry. Am. Nat. 146, 1–24 10.1086/285784 (doi:10.1086/285784) [DOI] [Google Scholar]
- 5.Nöldeke G., Samuelson L. 1999. How costly is the honest signaling of need? J. Theor. Biol. 197, 527–539 10.1006/jtbi.1998.0893 (doi:10.1006/jtbi.1998.0893) [DOI] [PubMed] [Google Scholar]
- 6.Godfray H. C. J., Johnstone R. A. 2000. Begging and bleating: the evolution of parent–offspring signalling. Phil. Trans. R. Soc. Lond. B 355, 1581–1591 10.1098/rstb.2000.0719 (doi:10.1098/rstb.2000.0719) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Johnstone R. A. 2004. Begging and sibling competition: how should offspring respond to their rivals? Am. Nat. 163, 388–406 10.1086/375541 (doi:10.1086/375541) [DOI] [PubMed] [Google Scholar]
- 8.Johnstone R. A., Godfray H. C. J. 2002. Models of begging as a signal of need. In The evolution of begging: competition, cooperation and communication (eds Wright J., Leonard M. L.), pp. 1–20 Dordrecht, The Netherlands: Kluwer [Google Scholar]
- 9.Rodríguez-Gironés M. A., Cotton P. A., Kacelnik A. 1996. The evolution of begging: signaling and sibling competition. Proc. Natl Acad. Sci. USA 93, 14 637–14 641 10.1073/pnas.93.25.14637 (doi:10.1073/pnas.93.25.14637) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Smiseth P. T., Moore A. J. 2008. Parental distribution of resources in relation to larval hunger and size rank in the burying beetle nicrophorus vespilloides. Ethology 114, 789–796 10.1111/j.1439-0310.2008.01516.x (doi:10.1111/j.1439-0310.2008.01516.x) [DOI] [Google Scholar]
- 11.Leonard M. L., Horn A. G. 2001. Begging calls and parental feeding decisions in tree swallows (tachycineta bicolor). Behav. Ecol. Sociobiol. 49, 170–175 10.1007/s002650000290 (doi:10.1007/s002650000290) [DOI] [Google Scholar]
- 12.Leonard M. L., Horn A. G. 1998. Need and nestmates affect begging in tree swallows. Behav. Ecol. Sociobiol. 42, 431–436 10.1007/s002650050457 (doi:10.1007/s002650050457) [DOI] [Google Scholar]
- 13.Budden A. E., Wright J. 2001. Begging in nestling birds. In Current ornithology. (eds Nolan V., Ketterson E.), pp. 83–118 New York, NY: Kluwer Academic/Plenum [Google Scholar]
- 14.Wells J. C. K. 2003. Parent–offspring conflict theory, signaling of need, and weight gain in early life. Q. Rev. Biol. 78, 169–202 10.1086/374952 (doi:10.1086/374952) [DOI] [PubMed] [Google Scholar]
- 15.Hinde C. A., Kilner R. M. 2007. Negotiations within the family over the supply of parental care. Proc. R. Soc. B 274, 53–60 10.1098/rspb.2006.3692 (doi:10.1098/rspb.2006.3692) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Kilner R., Johnstone R. A. 1997. Begging the question: are offspring solicitation behaviours signals of needs. Trends Ecol. Evol. 12, 11–15 10.1016/S0169-5347(96)10061-6 (doi:10.1016/S0169-5347(96)10061-6) [DOI] [PubMed] [Google Scholar]
- 17.Grodzinski U., Lotem A. 2007. The adaptive value of parental responsiveness to nestling begging. Proc. R. Soc. B 274, 2449–2456 10.1098/rspb.2007.0658 (doi:10.1098/rspb.2007.0658) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Grodzinski U., Hauber M. E., Lotem A. 2009. The role of feeding regularity and nestling digestive efficiency in parent–offspring communication: an experimental test. Funct. Ecol. 23, 569–577 10.1111/j.1365-2435.2008.01536.x (doi:10.1111/j.1365-2435.2008.01536.x) [DOI] [Google Scholar]
- 19.Stamps J., Clark A., Arrowood P., Kus B. 1985. Parent–offspring conflict in budgerigars. Behaviour 94, 1–40 10.1163/156853985X00253 (doi:10.1163/156853985X00253) [DOI] [Google Scholar]
- 20.Krebs E. A. 2001. Begging and food distribution in crimson rosella (Platycercus elegans) broods: why don't hungry chicks beg more? Behav. Ecol. Sociobiol. 50, 20–30 10.1007/s002650100339 (doi:10.1007/s002650100339) [DOI] [Google Scholar]
- 21.Stamps J., Clark A., Arrowood P., Kus B. 1989. Begging behavior in budgerigars. Ethology 81, 177–192 10.1111/j.1439-0310.1989.tb00765.x (doi:10.1111/j.1439-0310.1989.tb00765.x) [DOI] [Google Scholar]
- 22.Lotem A. 1998. Differences in begging behaviour between barn swallow, hirundo rustica, nestlings. Anim. Behav. 55, 809–818 10.1006/anbe.1997.0675 (doi:10.1006/anbe.1997.0675) [DOI] [PubMed] [Google Scholar]
- 23.Mock D. W., Schwagmeyer P. L., Parker G. A. 2005. Male house sparrows deliver more food to experimentally subsidized offspring. Anim. Behav. 70, 225–236 10.1016/j.anbehav.2004.10.020 (doi:10.1016/j.anbehav.2004.10.020) [DOI] [Google Scholar]
- 24.Wright J., Hinde C., Fazey I., Both C. 2002. Begging signals more than just short-term need: cryptic effects of brood size in the pied flycatcher (Ficedula hypoleuca). Behav. Ecol. Sociobiol. 52, 74–83 10.1007/s00265-002-0478-y (doi:10.1007/s00265-002-0478-y) [DOI] [Google Scholar]
- 25.Grodzinski U., Erev I., Lotem A. 2008. Can hungry nestlings be trained to reduce their begging? Behav. Ecol. 19, 116–125 10.1093/beheco/arm107 (doi:10.1093/beheco/arm107) [DOI] [Google Scholar]
- 26.Macnair M. R., Parker G. A. 1979. Models of parent–offspring conflict.3. Intra-brood conflict. Anim. Behav. 27, 1202–1209 10.1016/0003-3472(79)90067-8 (doi:10.1016/0003-3472(79)90067-8) [DOI] [Google Scholar]
- 27.Mock D. W., Parker G. A. 1997. The evolution of sibling rivalry. Oxford, UK: Oxford University Press [Google Scholar]
- 28.Hussell D. J. T. 1988. Supply and demand in tree swallow broods—a model of parent–offspring food-provisioning interactions in birds. Am. Nat. 131, 175–202 10.1086/284785 (doi:10.1086/284785) [DOI] [Google Scholar]
- 29.Kölliker M. 2003. Estimating mechanisms and equilibria for offspring begging and parental provisioning. Proc. R. Soc. Lond. B 270, S110–S113 10.1098/rsbl.2003.0032 (doi:10.1098/rsbl.2003.0032) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Kölliker M., Brodie E. D., Moore A. J. 2005. The coadaptation of parental supply and offspring demand. Am. Nat. 166, 506–516 10.1086/491687 (doi:10.1086/491687) [DOI] [PubMed] [Google Scholar]
- 31.Smiseth P. T., Wright J., Kölliker M. 2008. Parent–offspring conflict and co-adaptation: behavioural ecology meets quantitative genetics. Proc. R. Soc. B 275, 1823–1830 10.1098/rspb.2008.0199 (doi:10.1098/rspb.2008.0199) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Johnstone R. A. 1996. Begging signals and parent–offspring conflict: do parents always win? Proc. R. Soc. Lond. B 263, 1677–1681 10.1098/rspb.1996.0245 (doi:10.1098/rspb.1996.0245) [DOI] [Google Scholar]
- 33.Royle N. J., Hartley I. R., Parker G. A. 2002. Begging for control: when are offspring solicitation behaviours honest? Trends Ecol. Evol. 17, 434–440 10.1016/S0169-5347(02)02565-X (doi:10.1016/S0169-5347(02)02565-X) [DOI] [Google Scholar]
- 34.Parker G. A., Royle N. J., Hartley I. R. 2002. Intrafamilial conflict and parental investment: a synthesis. Phil. Trans. R. Soc. Lond. B 357, 295–307 10.1098/rstb.2001.0950 (doi:10.1098/rstb.2001.0950) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Hinde C. A., Johnstone R. A., Kilner R. M. 2010. Parent–offspring conflict and coadaptation. Science 327, 1373–1376 10.1126/science.1186056 (doi:10.1126/science.1186056) [DOI] [PubMed] [Google Scholar]
- 36.Bell M. B. V. 2008. Strategic adjustment of begging effort by banded mongoose pups. Proc. R. Soc. Lond. B 275, 1313–1319 10.1098/rspb.2008.0173 (doi:10.1098/rspb.2008.0173) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Clark A. B., Lee W. H. 1998. Red-winged blackbird females fail to increase feeding in response to begging call playbacks. Anim. Behav. 56, 563–570 10.1006/anbe.1998.0831 (doi:10.1006/anbe.1998.0831) [DOI] [PubMed] [Google Scholar]
- 38.Burford J. E., Friedrich T. J., Yasukawa K. 1998. Response to playback of nestling begging in the red-winged blackbird, Agelaius phoeniceus. Anim. Behav. 56, 555–561 10.1006/anbe.1998.0830 (doi:10.1006/anbe.1998.0830) [DOI] [PubMed] [Google Scholar]
- 39.Thorogood R., Ewen J. G., Kilner R. M. 2011. Sense and sensitivity: responsiveness to offspring signals varies with the parents' potential to breed again. Proc. R. Soc. B 278, 2638–2645 Proc. R. Soc. B. 10.1098/rspb.2010.2594 (doi:10.1098/rspb.2010.2594) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.English S., Kunc H. P., Madden J. R., Clutton-Brock T. H. 2008. Sex differences in responsiveness to begging in a cooperative mammal. Biol. Lett. 4, 334–337 10.1098/rsbl.2008.0200 (doi:10.1098/rsbl.2008.0200) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Macgregor N. A., Cockburn A. 2002. Sex differences in parental response to begging nestlings in superb fairy-wrens. Anim. Behav. 63, 923–932 10.1006/anbe.2001.1991 (doi:10.1006/anbe.2001.1991) [DOI] [Google Scholar]
- 42.Ghalambor C. K., Martin T. E. 2000. Parental investment strategies in two species of nuthatch vary with stage-specific predation risk and reproductive effort. Anim. Behav. 60, 263–267 10.1006/anbe.2000.1472 (doi:10.1006/anbe.2000.1472) [DOI] [PubMed] [Google Scholar]
- 43.Kilner R. M., Hinde C. A. 2008. Information warfare and parent–offspring conflict. Adv. Study Behav. 38, 283–336 10.1016/S0065-3454(08)00006-5 (doi:10.1016/S0065-3454(08)00006-5) [DOI] [Google Scholar]
- 44.Johnstone R. A. 1999. Signaling of need, sibling competition, and the cost of honesty. Proc. Natl Acad. Sci. USA 96, 12 644–12 649 10.1073/pnas.96.22.12644 (doi:10.1073/pnas.96.22.12644) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Hinde C. A., Buchanan K. L., Kilner R. M. 2009. Prenatal environmental effects match offspring begging to parental provisioning. Proc. R. Soc. B 276, 2787–2794 10.1098/rspb.2009.0375 (doi:10.1098/rspb.2009.0375) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Smith H. G., Montgomerie R. 1991. Nestling American robins compete with siblings by begging. Behav. Ecol. Sociobiol. 29, 307–312 10.1007/BF00163989 (doi:10.1007/BF00163989) [DOI] [Google Scholar]
- 47.Price K., Harvey H., Ydenberg R. 1996. Begging tactics of nestling yellow-headed blackbirds, Xanthocephalus xanthocephalus, in relation to need. Anim. Behav. 51, 421–435 10.1006/anbe.1996.0039 (doi:10.1006/anbe.1996.0039) [DOI] [Google Scholar]
- 48.Rodríguez-Gironés M. A., Zuniga J. M., Redondo T. 2002. Feeding experience and relative size modify the begging strategies of nestlings. Behav. Ecol. 13, 782–785 (doi:10.1093/beheco/13.6.782) [Google Scholar]
- 49.Kedar H., Rodríguez-Gironés M. A., Yedvab S., Winkler D. W., Lotem A. 2000. Experimental evidence for offspring learning in parent–offspring communication. Proc. R. Soc. Lond. B 267, 1723–1727 10.1098/rspb.2000.1201 (doi:10.1098/rspb.2000.1201) [DOI] [PMC free article] [PubMed] [Google Scholar]