

Abstract
Biosynthesis of membrane proteins requires that hydrophobic transmembrane (TM) regions be shielded from the cytoplasm while being directed to the correct membrane. Tail-anchored (TA) membrane proteins, characterized by a single C-terminal TM, pose an additional level of complexity because they must be post-translationally targeted. In eukaryotes, the GET pathway shuttles TA-proteins to the endoplasmic reticulum. The key proteins required in yeast (Sgt2 and Get1-5) have been under extensive structural and biochemical investigation during recent years. The central protein Get3 utilizes nucleotide linked conformational changes to facilitate substrate loading and targeting. Here we analyze this complex process from a structural perspective, as understood in yeast, and further postulate on similar pathways in other domains of life.
Introduction
Delivery of membrane proteins to the proper membrane is a critical process that is highly regulated. The field of protein targeting has recently added a new pathway involved in targeting the special class of tail-anchored membrane (TA) proteins to the ER. After the initial discovery of proteins involved in this pathway a wealth of genetics, biochemistry and structural information has rapidly elucidated a complex process of hand-offs of the TA-substrate. The pathway starts by transfer of the substrate to the Get4/Get5/Sgt2 sorting complex that then loads the TA onto the targeting chaperone Get3, which is subsequently released at the membrane by the Get1/Get2 receptor complex. While recent reviews describe these initial characterizations [1,2], in this review we will summarize the current state of the field from a structural perspective and more broadly comment on the possibility of similar pathways beyond fungi. Figure 1 is an overall model of TA-protein targeting, including the various ambiguities in certain steps, which will be referred to throughout the text.
Figure 1. A model for TA targeting by the GET pathway.
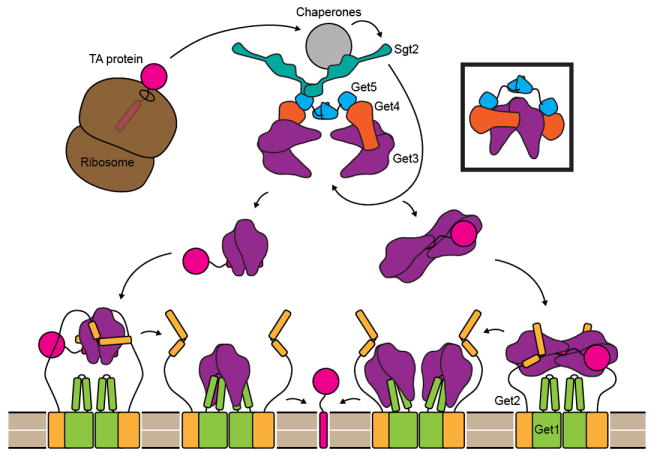
After protein synthesis is complete, a complex consisting of cellular chaperones, two copies each of Get4 and Get5 and at least one dimer of Sgt2 binds the TM helix of TA-proteins. The Get4/Get5 complex recruits Get3, and Sgt2 transfers ER destined TA-proteins to Get3. Each Get4 may bind a separate dimer of Get3, or the same dimer (boxed). The Get3/TA-protein complex may contain two or four copies of Get3. The stoichiometry of the Get1/Get2 complex within the ER membrane is unknown but is shown here as a dimer of 1:1 Get1 to Get2 dimers. Initially Get2 binds the Get3/TA-protein complex and is then displaced by Get1. Get1 binding is coupled to the opening of Get3, leading to release and integration of the TA-protein. In the case of a dimeric Get3/TA complex, Get1 and Get2 could bind the Get3 subunits symmetrically (left pathway). Alternatively, Get2 and Get1 could bind a tetramer of Get3 asymmetrically (right pathway).
Structural changes in the Get3 molecular machine
The general structure and mechanism of Get3, which we briefly summarize here, has been recently reviewed [2,3]. Get3 is a nucleotide hydrolase that modulates its conformation through nucleotide state. Similar to other SIMIBI class NTPases (named for representative members signal recognition particle, MinD and BioD), Get3 forms a homodimer through interactions between the nucleotide hydrolase domains (NHD) [4]. The subunits rotate relative to each other from an open state in apo form or bound to ADP to a more compact closed state bound to Mg2+ADP·AlF4−, Mg2+AMPPNP or Mg2+ADP, forming an intersubunit hydrophobic groove from α-helices connected to the NHD [5-8].
There are three models for how Get3 sequesters a TA-protein predominantly based on structures (Fig. 2A & B). The first two models are based on a dimer of Get3, which is the dominant form of purified fungal Get3. In the prevailing model, an α-helical TA is captured by the hydrophobic groove formed by the closed Get3 dimer [9] (Fig. 2D). The alternative dimer model posits that pairs of amphipathic α-helices extending out from either side of the groove bind the TA through a hydrophobic patch [8]. The third model correlates the observation that heterologously purified Get3/TA complexes contain four copies of Get3 with the structure of a tetrameric archaeal homolog. In this model, the TA would be sequestered in a hydrophobic chamber formed by the grooves of opposing dimers (Fig. 4E & S1) [10]. In all models, similar residues mediate potential TA interactions. The importance of these regions in TA binding has been shown through various mutagenesis experiments [7-9]. Additionally, changes in these regions upon TA binding have been demonstrated through hydrogen exchange mass spectrometry of Get3 complexed with TA-protein [5]. In general, results from biochemical experiments are compatible with both dimer and tetramer models. The simplicity of the dimer model is consistent with structures seen in complex with partners (see below) and binding of functionalized TA substrates [11]. The tetramer model allows for complete shielding of the hydrophobic domain but necessitates additional assembly and disassembly steps, yet agrees with complexes analyzed in solution [5,10,12]. The oligomeric state of Get3 in the native TA complex remains a subject for future study.
Figure 2. Conformations of Get3 and models for TA binding.
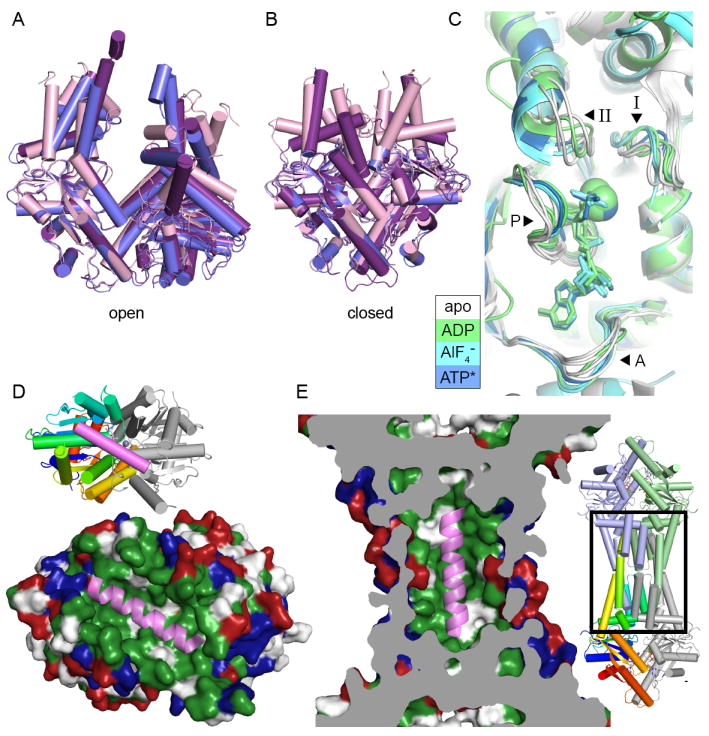
A. Open form structures of Get3 in a cartoon representation aligned by the right subunit, using PDB IDs 3A36 (purple), 3H84 (blue) and 2WOO (pink). B. Closed form structures shown as in A, using PDB IDs 3IQW (purple), 3IO3 (blue) and 2WOJ (pink). C. The NHD of aligned Get3 structures from one subunit. Motifs are indicated with arrowheads and labeled A (A-loop), P (P-loop), I (switch I) and II (switch II). Structures are colored according to the nucleotide bound with white for no nucleotide (PDB ID: 3A36, PDB ID: 2WOO, PDB ID: 3SJA and PDB ID: 3SJC), green for ADP (PDB ID: 3IQX and PDB ID: 3SJD), cyan for ADP·AlF4− (PDB ID: 2WOJ, PDB ID: 3ZQ6 and PDB ID: 3ZS9) and blue for AMPPNP (PDB ID: 3IQW). D. A model for TA binding by the Get3 dimer [9]. Get3 (PDB ID: 2WOJ) is shown in a surface representation colored hydrophobic (green), positively (blue) and negatively charged (red). A cartoon representation of a TA from Secβ (PDB ID: 1RHZ), colored pink, is shown in the groove formed in the closed structure of Get3. The inset shows a cartoon representation of Get3 with the left subunit colored ramped and the right subunit in gray. E. A TA binding model for a tetramer of Get3 [10] represented as in D. The structure of tetrameric Get3 (PDB ID: 3UG6) is cut away to show the central cavity with a TA modeled inside, similar to D. The area shown is indicated with a box on the overall structure in the inset.
Figure 4. Interactions between Get3 and other Get pathway members.
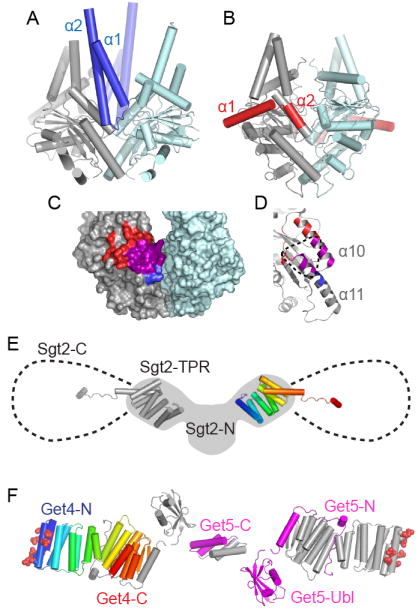
A. Crystal structure of Get3 in complex with the Get1 cytoplasmic domain (blue) (PDB ID 3ZS8). One Get3 subunit is gray and the other pale blue. B. Crystal structure of Get3 in complex with the Get2 cytoplasmic domain (red) (PDB ID: 3SJD). C. Surface rendering of open Get3 (PDB ID: 3A36). Regions where Get1 (blue) and Get2 (red) interact with the gray subunit are highlighted, with the overlapping region in magenta. D. Close up of Get3 α10 and α11, colored as in C. The expected Get4 binding interface is indicated with a dotted line. E. Model of the structure of Sgt2. Two copies of the TPR domain (PDB ID: 3SZ7) are displayed over a cartoon of a SAXS model of the N-terminal dimerization and TPR domains (gray, adapted from [35]). The additional 107 residues of the C-terminal domains rotate freely from the TPR and a potential range of motion is indicated with dotted lines. F. Structure of the Get4/Get5 heterotetramer. Residues shown to be critical for Get3 interaction are shown as spheres [35]. Subunits are arranged based upon SAXS models [31]. One copy of Get4 is color ramped from N- (blue) to C- (red) termini. One copy of Get5 is colored magenta and the remaining Get4 and Get5 subunits are gray.
Many nucleotide hydrolases are molecular machines that act as switches, utilizing nucleotide state and the energy of hydrolysis to alter their conformation with a broad spectrum of functionality from signaling interactions to actively driving processes. The Get3 homodimer uses ATP hydrolysis to switch between open and closed conformations during its targeting cycle (Fig. 2A & B). Related SIMIBI class NTPases in the MinD/Mrp family [4] switch by changes in dimer conformation like Get3 (NifH and ArsA pseudodimer) or convert between monomeric and dimeric states (MinD and Soj) [13]. Changes from open and closed dimer states were originally seen for NifH [14] and have been modeled for ArsA [15].
In SIMIBI proteins, nucleotide induced conformational changes control interactions with protein factors. For Get3, this involves ferrying the TA substrate from the sorting complex (Sgt2/Get4/5) to the membrane receptor (Get1/2) (see below). This is analogous to ArsA that receives arsenite from ArsD then exports through the membrane protein ArsB [16]. NifH switches between a MoFe bound state ferrying electrons along the nitrogenase cycle [17]. In the best characterized of these cases, in addition to rotation across the dimer interface, the switch II loop alters its conformation among different nucleotide states (Fig. 2C and Fig. S2). This loop connects to structural features mediating partner interactions. In the case of Get3, switch II is linked to α, which is connected to the putative substrate-binding groove (Fig. 2C). Therefore, the nucleotide state can modulate transitions associated with TA-binding. Similar switch II conformational changes occur during the NifH and ParA nucleotide cycle (Fig. S1).
The ArsA fold
Get3 belongs to a subset of the SIMIBI family of proteins characterized by the first member ArsA. ArsA contains two NHDs linked together forming a pseudo-dimer (Fig. 3A) [18]. In the first structure, substrate antimony atoms bind in a groove formed by two loops that extended from the NHD, linking the nucleotide-binding pocket to liganding cysteines and histidines (Fig. 3B). ATP hydrolysis is proposed to induce conformational changes that coordinate the release of the toxic metals to the ArsB exporter. Interestingly, the two halves have only 26% identity, and this asymmetry appears to play a functional role [19]. The substrate-binding groove appears to be a unique identifier for this family (Fig. 3A & B).
Figure 3. Members of the Get3/ArsA fold family.
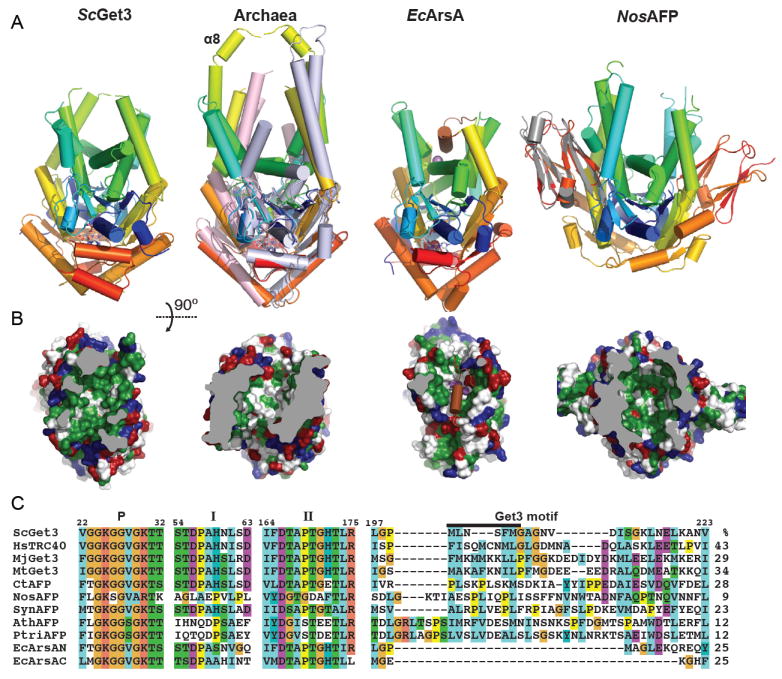
A. Cartoon representations of dimers of members of the ArsA fold viewed down the putative TA binding groove. The structures are the transition state S. cerevisiae Get3 (PDB ID: 2WOJ), the archaeal M. jannaschii (PDB ID: 3UG6, monomers color ramped) and M. thermoautotropicum Get3s (PDB ID: 3ZQ6, monomers light blue/pink), E. coli ArsA (PDB ID: 1F48, each pseudo-dimer color ramped with the helix filling the groove in brown), and the cyanobacterial homolog Nostoc sp. all4481 (PDB ID: 3IGF, monomers color ramped). The human α-crystallin structure (PDB ID: 2WJ7) has been aligned in gray to the equivalent cyanobacterial domain. Metals are shown as spheres and nucleotides as sticks. B. Surface representations of the structures in A rotated 90° with residues colored as in Fig. 2D. Some foreground residues have been removed for clarity and the truncation is colored grey. MjGet3 is shown for the archaeal homologs and the loop filling the groove has been removed in the ArsA surface calculation to show the comparable groove. C. An alignment of notable regions of the ArsA family proteins from ClustalW. Numbering and region names are based on the S. cerevisiae protein similar to Fig. 2C. A black bar identifies residues corresponding to α8. The sequences are ScGet3 (Saccharomyces cerevisiae UniProt ID: Q12154), HsTRC40 (Homo sapiens UniProt ID: O43681), MjGet3 (Methanocaldococcus jannaschii UniProt ID: Q58542), MtGet3 (Methanobacter thermoautotropicum UniProt ID: O27555) CtAFP (Chlorobium tepidum UniProt ID: Q46366), NosAFP (Nostoc sp PCC 7120 UniProt ID: Q8YNT0), SynAFP (Synechocystis sp. PCC 6803 UniProt ID: F7UTP7), AthAFP (Arabidopsis thaliana UniProt ID: Q6DYE4), PtriAFP (Populus trichocarpa or California poplar UniProt ID: B9HWM7) and EcArsA (Escherichia coli plasmid R773 N- and C-terminal domains UniProt ID: P08690). Numbers to the right are percent identity to ScGet3 excluding crystallin domains.
Fungal Get3 has approximately 25% identity to each half of ArsA; therefore, based on homology alone, distinguishing an ArsA from a Get3 is difficult. In most genomic annotations members of this fold family are identified as ArsA homologs. Several key differences to distinguish Get3 from ArsA have been identified. The simplest is that Get3 is a homo-dimer. Instead of containing the ArsA metal ligand residues [20,21], Get3 homologs contain a unique “Get3 motif” insertion [9] (Fig. 3C). Moreover, Get3 homologs typically contain a pair of cysteines, a CXXC motif, at their dimer interface that coordinate zinc presumably to stabilize the dimer, analogous to the ArsA linker [21,22].
Archaea contain both ArsA and Get3 homologs [23]. Only about half of the sequenced genomes contain the Get3 homolog and half of these lack the CXXC motif [10]. While the role of the ArsA homolog is likely similar to that in eubacteria, the presence of a Get3 homolog suggests membrane protein targeting occurs. Indeed, in heterologous systems archaeal Get3 can form complexes with a variety of TA-proteins and in one case facilitates insertion [10,24]. This implies a unique pathway, as other GET pathway members have not been identified. Recent structural studies of the homologs from Methanobacter thermoautotripicum (MtGet3) and Methanocaldococcus jannaschii (MjGet3) confirm the structural homology of archaeal Get3 to fungal Get3 (Fig. 3A & B) [10,24]. Both archaeal structures are in the closed form; however, unlike fungal Get3 homologs the substrate binding loops are ordered and extended. In MjGet3 this results in a tetramer, a dimer of dimers, with a closed hydrophobic chamber stabilized by α8 (Fig. 2E & S1) [10]. Moreover, of the archaeal homologs where oligomeric state was analyzed, all are capable of forming stable tetramers in solution including the Thermococcus kodakaerensis homolog, which lacks the CXXC motif and is competent for TA binding [10].
Looking more broadly at sequence homology a third class of this fold family becomes apparent. Here, the fold is found in photosynthetic organisms. For clarity, we will refer to members of this protein class as the ‘ArsA Family fold associated with Photosynthesis’ or AFP. They can be found in green sulfur bacteria, cyanobacteria and are nuclear encoded with chloroplast-targeting signals in plants. Each of these organisms is characterized by photosynthetic machinery in specialized membranes, either the chlorosome or thylakoid [25]. A recent structure from this class, all4481 from the cyanobacteria Nostoc sp PCC 7120 (NosAFP), was deposited by the Northeast Structural Genomics Consortium. The structure reveals a closed Get3-like fold containing the hydrophobic groove but lacking the nucleotide-binding pocket (Fig. 3A & B). A small heat shock protein (HSP)/crystallin domain is appended to the C-terminus that contacts the Get3 surface used in yeast for partner recognition (see below). The remarkable structural similarity of NosAFP is not evident from the low sequence identity (9%) to ScGet3; however, similar homologs such as that from the green sulfur bacteria Chlorobium tepidum (CtAFP) have higher identity (28%) and appear to include the nucleotide-binding pocket (Fig. 3C). The conservation of the AFP class from ancient photosynthetic organisms to modern plants suggests a critical role in biosynthesis of the photosynthetic membranes. It will be exciting to see if there is a role for this protein in protein targeting to membranes.
Get1 and Get2 regulate membrane insertion steps
The integral membrane proteins Get1 and Get2 form the ER membrane bound complex required for TA-insertion by Get3 [26]; however, only a homolog for Get2 is found in higher eukaryotes [27]. Both proteins have single cytoplasmic domains that can bind Get3 in the absence of the membrane components. The Get1 cytoplasmic domain, which connects the first and second transmembrane helices, is a coiled-coil motif that extends between the subunits of a Get3 dimer [28,29] (Fig. 4A). A portion of Get1 binds at a groove on the surface of Get3 formed by α10 and α11. At the other Get3 subunit, Get1 extends into the nucleotide-binding pocket, preventing ATP or ADP from binding (Fig. 4A, C & D). Get3 is in an open or semi-open conformation, with Get1 acting as a wedge that would block a complete transition to the closed state. Nearly the entire cytoplasmic loop is modeled in these structures; therefore, bound Get3 must be in close proximity to the ER membrane.
The cytoplasmic domain of Get2 consists of the N-terminus of the protein. Overall sequence conservation is poor, with the exception of the first 35 amino acids that are sufficient to bind Get3 [28,29]. This region forms two helices that wrap along the outer surface of Get3, including the groove where Get1 interacts (Fig. 4B-D). Unlike Get1, the ordered portions of Get2 do not contact both subunits of Get3. With bound Mg2+ADP·AlF4−, Get3 is nearly identical to previous transition state structures [5,9,28]. A second structure containing Mg2+ADP also is a closed Get3 dimer [29]. Despite this, helices that line the TA-protein-binding site are in conformations similar to open structures although resolution limited the extent of modeling and refinement (Fig. S1). Intriguingly, in both structures the surface of Get3 that interacts with Get2 is unperturbed relative to Get3 alone. Since Get2 does not contact the TA-protein binding loops or the nucleotide-binding pocket, it is unclear why a closed conformation of Get3 would be selected. It is also noteworthy that in the Mg2+ADP bound structure a crystallographic axis relates a second Get3 dimer into an arrangement similar to the M. jannaschii tetramer [10] (Fig. S1).
The structures of Get1 and Get2 and other biochemical data led to a general model for events at the membrane [28-30] (Fig. 1). The flexible cytoplasmic domain of Get2 initially captures the Get3/TA-protein complex. As Get3 approaches the membrane, Get1 first displaces Get2 and then facilitates a transition from closed to open Get3. This corresponds with integration of the TA-protein into the membrane and release of bound nucleotide. ATP then displaces Get1, releasing Get3 from the membrane complex. The exact nature, however, of the Get1/Get2/Get3 complex at the membrane remains to be determined and is crucial for understanding the mechanism of insertion. Without a clear structural explanation for state selectivity for Get2, it is not obvious how the Get3/TA-protein complex is favored from free Get3 or how immediate rebinding after an insertion cycle is prevented. Uncertainty about the stoichiometry of Get1 and Get2 results in alternative models of Get1 and Get2 binding at different subunits of dimeric or tetrameric Get3. Finally, the biophysical mechanism of TA-protein insertion into the membrane and the requirements of Get3 or full-length Get1 and Get2, in this process are largely unknown.
Sgt2, Get4 and Get5 load Get3 with TA-protein
The HSP co-chaperone Sgt2 mediates the committed step in TA-protein targeting. It recruits a variety of HSP families via an internal tetratricopeptide repeat (TPR) domain [30,31]. The Sgt2 C-terminal domain binds the transmembrane helix of ER-destined TA-proteins, which are then handed to Get3 [30,32]. In contrast, mitochondrial TA-proteins remain associated with bound HSPs. This domain, rich in glutamine, methionine and asparagine, contains only a short conserved sequence and is weakly predicted as helical. The mechanism of TA-protein selection and handoff to Get3 are, therefore, intriguing problems. Sgt2 is a homodimer mediated by its small N-terminal domain, and the TPR and C-terminal domains of the two subunits extend away from each other [31] (Fig 4E). The N-terminal domain also mediates the association with Get5, where a dimer of Sgt2 binds a single copy of Get5 [31,33].
Get4 and Get5 act as an adaptor complex linking Sgt2 to Get3. Get4 is an alpha helical repeat protein that tightly binds to the N-terminal domain of Get5 [33-35]. The Get5 sequence is followed by a ubiquitin-like domain (Ubl) and a C-terminal homodimerization domain, resulting in an extended heterotetrameric complex [35,36] (Fig. 4F). The N-terminal face of Get4 is a conserved basic surface that mediates interaction with Get3 [33,35,37]. Although a high-resolution structure of this complex remains to be determined, mutagenesis and molecular modeling indicate that, like Get1 and Get2, Get4 binds Get3 at α10 and α11 (Fig. 4D). The in vivo stoichiometry, however, has not been established. In order for a single Get4/Get5 heterotetramer to bind both subunits of a Get3 dimer, as it has been proposed [35,37], the Ubl and C-terminal domains of Get5 would either thread through the TA binding groove of Get3 or wrap around the NHDs, possibly positioning Sgt2 away from the TA-protein binding groove. If the two Get4 copies bind independent Get3 dimers, the Sgt2/TA-protein complex could be more accessible to Get3. This model also provides a framework for Get3 to tetramerize upon TA-protein capture (Fig. 1)[36].
A complex analogous to Get4/Get5/Sgt2 appears to exist in vertebrates although with some differences in architecture and function. The homologs of Get4 and Get5, named TRC35 and Ubl4a, bind to Bag6 (alternatively named Bat-3 or Scythe) forming the Bag6 complex [38]. Bag6 is an approximately 1000-residue protein with an N-terminal Ubl domain and a C-terminal Bag domain, with internal proline rich regions. SGTA, the homolog of Sgt2, uses its N-terminal domain to associate with Bag6, and although it has not yet been demonstrated experimentally, Ubl4A is expected to bridge these proteins [2,39]. The Bag6 complex not only has the ability to load TA-proteins onto the Get3 homolog, TRC40 [38,40], but also mediates protein degradation pathways for mislocalized membrane proteins, retrotranslocated ER proteins and other defective proteins [32,41,42]. Determining how the interplay between these functions makes or breaks membrane proteins will provide fundamental insight into how TM segments are manipulated by the cell.
Concluding remarks
We are rapidly acquiring structural information for all of the components of the TA-protein-targeting pathway. Careful analysis of analogous systems and components of the large and dynamic complexes generate testable models. These are tied together by low-resolution and biochemical methodologies. Despite recent leaps in understanding, many crucial questions remain to be answered. How are TA-proteins initially directed towards the Get4/Get5/Sgt2 sorting complex? How does this complex recruit Get3 and what is the mechanism for handoff? What is the structure of the physiological Get3/TA-protein complex? How is the TA-protein integrated into the membrane? Further afield are questions about the functional roles of structurally homologous proteins in other domains of life. Clearly, understanding the detailed biosynthesis of this important class of proteins will continue to excite for some time to come.
Supplementary Material
Structures of Get3 tetramers. A. Tetrameric MjGet3 (PDB ID: 3UG6) shown in a cartoon representation with one subunit color ramped from N to C-terminus and additional subunits colored gray, blue and green. B. A crystallographic tetramer of ScGet3 (PDB ID: 3SJD, shown as in A) in complex with the soluble domain of Get2 (cyan).
Conformations of the NHDs of NifH and ParA in different nucleotide states. A. A structural alignment of NifH NHDs represented as in Fig. 2C. Apo (PDB ID: 2AFH), ADP bound (PDB ID: 2AFI), ADP·AlF4− bound (PDB ID: 1M34) and AMPPCP bound (PDB ID: 2AFK) forms are shown. B. As in A for ParA structures. Apo (PDB ID: 3EZ7, PDB ID: 3EZ9, PDB ID: 3EZF), ADP bound (PDB ID: 3CWQ, PDB ID: 3EZ2) and ATP/ATPγS bound (PDB ID: 3EA0, PDB ID: 2OZE) forms are shown.
Highlights.
Get3 is a central nucleotide hydrolase in the GET pathway, and models for interaction of tail-anchored (TA) proteins with Get3 include a binding groove formed by a Get3 dimer and a tetramer with a central hydrophobic chamber.
ArsA is a representative member of a family of ATPases that includes Get3.
Archaeal Get3 homologs are similar in structure and function to fungal Get3s suggesting a unique TA-targeting pathway in this domain.
Some photosynthetic organisms have a conserved protein with structural similarity to Get3 that may be involved in biosynthesis of photosynthetic membranes.
Get1 and Get2 form a membrane receptor for Get3/TA-protein complexes, and complexes of Get1 and Get2 soluble domains with reveal a mechanism for release at the membrane.
Acknowledgments
We are grateful to Harry Gristick, Meera Rao, Doug Rees and Michael Rome for commenting on the manuscript. Research in the Clemons lab is funded by an R01 from the National Institutes of Health grant GM097572.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Shao S, Hegde RS. Membrane protein insertion at the endoplasmic reticulum. Annual Rev Cell Dev Biology. 2011;27:25–56. doi: 10.1146/annurev-cellbio-092910-154125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- **2.Hegde RS, Keenan RJ. Tail-anchored membrane protein insertion into the endoplasmic reticulum. Nature Rev Mol Cell Biol. 2011;12:787–798. doi: 10.1038/nrm3226. This review provides an excellent overview of the TA protein-targeting problem and outlines the elucidation of components of the GET/TRC pathways in a framework for the current model. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Simpson PJ, Schwappach B, Dohlman HG, Isaacson RL. Structures of Get3, Get4, and Get5 Provide New Models for TA Membrane Protein Targeting. Structure. 2010;18:897–902. doi: 10.1016/j.str.2010.07.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Leipe D, Wolf Y, Koonin E, Aravind L. Classification and evolution of P-loop GTPases and related ATPases. J Mol Biol. 2002;317:41–72. doi: 10.1006/jmbi.2001.5378. [DOI] [PubMed] [Google Scholar]
- 5.Bozkurt G, Stjepanovic G, Vilardi F, Amlacher S, Wild K, Bange G, Favaloro V, Rippe K, Hurt E, Dobberstein B, et al. Structural insights into tail-anchored protein binding and membrane insertion by Get3. Proc Natl Acad Sci USA. 2009;106:21131–21136. doi: 10.1073/pnas.0910223106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Hu J, Li J, Qian X, Denic V, Sha B. The crystal structures of yeast get3 suggest a mechanism for tail-anchored protein membrane insertion. PLoS ONE. 2009;4:e8061. doi: 10.1371/journal.pone.0008061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Suloway CJM, Chartron JW, Zaslaver M, Clemons WM., Jr Model for eukaryotic tail-anchored protein binding based on the structure of Get3. Proc Natl Acad Sci USA. 2009;106:14849–14854. doi: 10.1073/pnas.0907522106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Yamagata A, Mimura H, Sato Y, Yamashita M, Yoshikawa A, Fukai S. Structural insight into the membrane insertion of tail-anchored proteins by Get3. Genes to Cells. 2010;15:29–41. doi: 10.1111/j.1365-2443.2009.01362.x. [DOI] [PubMed] [Google Scholar]
- 9.Mateja A, Szlachcic A, Downing ME, Dobosz M, Mariappan M, Hegde RS, Keenan RJ. The structural basis of tail-anchored membrane protein recognition by Get3. Nature. 2009;461:361–366. doi: 10.1038/nature08319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- **10.Suloway CJ, Rome ME, Clemons WM., Jr Tail-anchor targeting by a Get3 tetramer: the structure of an archaeal homologue. EMBO J. 2012;31:707–719. doi: 10.1038/emboj.2011.433. This paper shows that heterologously expressed Get3/TA protein complexes from both eukaryotes and archaea contain tetramer of Get3. Evidence is presented that supports a novel tetramer model for targeting. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Leznicki P, Warwicker J, High S. A biochemical analysis of the constraints of tail-anchored protein biogenesis. Biochem J. 2011;436:719–727. doi: 10.1042/BJ20101737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Favaloro V, Vilardi F, Schlecht R, Mayer MP, Dobberstein B. Asna1/TRC40-mediated membrane insertion of tail-anchored proteins. J Cell Sci. 2010;123:1522–1530. doi: 10.1242/jcs.055970. [DOI] [PubMed] [Google Scholar]
- 13.Lutkenhaus J, Sundaramoorthy M. MinD and role of the deviant Walker A motif, dimerization and membrane binding in oscillation. Mol Microbiol. 2003;48:295–303. doi: 10.1046/j.1365-2958.2003.03427.x. [DOI] [PubMed] [Google Scholar]
- 14.Schindelin H, Kisker C, Schlessman JL, Howard JB, Rees DC. Structure of ADP x AIF4(-)-stabilized nitrogenase complex and its implications for signal transduction. Nature. 1997;387:370–376. doi: 10.1038/387370a0. [DOI] [PubMed] [Google Scholar]
- 15.Ajees AA, Yang J, Rosen BP. The ArsD As(III) metallochaperone. Biometals. 2011;24:391–399. doi: 10.1007/s10534-010-9398-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Lin Y, Yang J, Rosen B. ArsD: an As(III) metallochaperone for the ArsAB As(III)-translocating ATPase. J Bioenerg Biomembr. 2007;39:453–458. doi: 10.1007/s10863-007-9113-y. [DOI] [PubMed] [Google Scholar]
- 17.Burgess BK, Lowe DJ. Mechanism of Molybdenum Nitrogenase. Chemical Rev. 1996;96:2983–3012. doi: 10.1021/cr950055x. [DOI] [PubMed] [Google Scholar]
- 18.Zhou T, Radaev S, Rosen BP, Gatti DL. Structure of the ArsA ATPase: the catalytic subunit of a heavy metal resistance pump. EMBO J. 2000;19:4838–4845. doi: 10.1093/emboj/19.17.4838. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Fu H-L, Ajees AA, Rosen BP, Bhattacharjee H. Role of signature lysines in the deviant walker a motifs of the ArsA ATPase. Biochemistry. 2010;49:356–364. doi: 10.1021/bi901681v. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Boskovic J, Soler-Mira A, García-Cantalejo JM, Ballesta JP, Jiménez A, Remacha M. The sequence of a 16,691 bp segment of Saccharomyces cerevisiae chromosome IV identifies the DUN1, PMT1, PMT5, SRP14 and DPR1 genes, and five new open reading frames. Yeast. 1996;12:1377–1384. doi: 10.1002/(SICI)1097-0061(199610)12:13%3C1377::AID-YEA35%3E3.0.CO;2-R. [DOI] [PubMed] [Google Scholar]
- 21.Stefanovic S, Hegde RS. Identification of a targeting factor for posttranslational membrane protein insertion into the ER. Cell. 2007;128:1147–1159. doi: 10.1016/j.cell.2007.01.036. [DOI] [PubMed] [Google Scholar]
- 22.Metz J, Wächter A, Schmidt B, Bujnicki JM, Schwappach B. The yeast Arr4p ATPase binds the chloride transporter Gef1p when copper is available in the cytosol. J Biol Chem. 2006;281:410–417. doi: 10.1074/jbc.M507481200. [DOI] [PubMed] [Google Scholar]
- 23.Borgese N, Righi M. Remote Origins of Tail-Anchored Proteins. Traffic. 2010;11:877–885. doi: 10.1111/j.1600-0854.2010.01068.x. [DOI] [PubMed] [Google Scholar]
- 24.Sherrill J, Mariappan M, Dominik P, Hegde RS, Keenan RJ. A conserved archaeal pathway for tail-anchored membrane protein insertion. Traffic. 2011;12:1119–1123. doi: 10.1111/j.1600-0854.2011.01229.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Hohmann-Marriott MF, Blankenship RE. Evolution of photosynthesis. Annual Rev of Plant Biol. 2011;62:515–548. doi: 10.1146/annurev-arplant-042110-103811. [DOI] [PubMed] [Google Scholar]
- 26.Schuldiner M, Metz J, Schmid V, Denic V, Rakwalska M, Schmitt HD, Schwappach B, Weissman JS. The GET Complex Mediates Insertion of Tail-Anchored Proteins into the ER Membrane. Cell. 2008;134:634–645. doi: 10.1016/j.cell.2008.06.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Vilardi F, Lorenz H, Dobberstein B. WRB is the receptor for TRC40/Asna1-mediated insertion of tail-anchored proteins into the ER membrane. Journal of Cell Science. 2011;124:1301–1307. doi: 10.1242/jcs.084277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- *28.Mariappan M, Mateja A, Dobosz M, Bove E, Hegde RS, Keenan RJ. The mechanism of membrane-associated steps in tail-anchored protein insertion. Nature. 2011;477:61–66. doi: 10.1038/nature10362. Get1 and Get2 are shown to be the minimum membrane components needed for TA protein integration by Get3. Structures of Get1/Get3 and Get2/Get3 complexes revealed a shared binding site. The effect of nucleotide on the Get1/Get3 interaction provides a mechanism for pathway regeneration. [DOI] [PMC free article] [PubMed] [Google Scholar]
- *29.Stefer S, Reitz S, Wang F, Wild K, Pang YY, Schwarz D, Bomke J, Hein C, Lohr F, Bernhard F, et al. Structural basis for tail-anchored membrane protein biogenesis by the Get3-receptor complex. Science. 2011;333:758–762. doi: 10.1126/science.1207125. This is an independent report presenting the crystal structures of the Get1/Get3 and Get2/Get3 complexes. As with Mariappan et al., nucleotide is shown to affect the Get1 and Get3 interaction. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Wang F, Brown EC, Mak G, Zhuang J, Denic V. A Chaperone Cascade Sorts Proteins for Posttranslational Membrane Insertion into the Endoplasmic Reticulum. Mol Cell. 2010;40:159–171. doi: 10.1016/j.molcel.2010.08.038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Chartron JW, Gonzalez GM, Clemons WM., Jr A structural model of the Sgt2 protein and its interactions with chaperones and the Get4/Get5 complex. J Biol Chem. 2011;286:34325–34334. doi: 10.1074/jbc.M111.277798. [DOI] [PMC free article] [PubMed] [Google Scholar]
- *32.Wang Q, Liu Y, Soetandyo N, Baek K, Hegde R, Ye Y. A ubiquitin ligase-associated chaperone holdase maintains polypeptides in soluble states for proteasome degradation. Mol Cell. 2011;42:758–770. doi: 10.1016/j.molcel.2011.05.010. This report shows that the Bag-6/Ubl4A/TRC35 complex is not only involved in TA protein targeting, but also binds to the transmembrane segments of integral membrane proteins prior to degradation by the proteasome. [DOI] [PMC free article] [PubMed] [Google Scholar]
- *33.Chang Y-W, Chuang Y-C, Ho Y-C, Cheng M-Y, Sun Y-J, Hsiao C-D, Wang C. Crystal structure of Get4/Get5 complex and its interactions with Sgt2, Get3 and Ydj1. J Biol Chem. 2010;286:9962–9970. doi: 10.1074/jbc.M109.087098. These report the first crystal structures of Get4 in complex with a portion of Get5 and map the interaction surface with Get3. Chartron et al further shows a nucleotide dependence for Get3 recognition. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Bozkurt G, Wild K, Amlacher S, Hurt E, Dobberstein B, Sinning I. The structure of Get4 reveals an alpha-solenoid fold adapted for multiple interactions in tail-anchored protein biogenesis. FEBS Lett. 2010 doi: 10.1016/j.febslet.2010.02.070. [DOI] [PubMed] [Google Scholar]
- *35.Chartron JW, Suloway CJM, Zaslaver M, Clemons WM., Jr Structural characterization of the Get4/Get5 complex and its interaction with Get3. Proc Natl Acad Sci USA. 2010;107:12127–12132. doi: 10.1073/pnas.1006036107. These report the first crystal structures of Get4 in complex with a portion of Get5 and map the interaction surface with Get3. Chartron et al further shows a nucleotide dependence for Get3 recognition. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Chartron JW, Vandervelde DG, Rao M, Clemons WM., Jr The Get5 carboxyl terminal domain is a novel dimerization motif that tethers an extended Get4/Get5 complex. J Biol Chem. 2012 doi: 10.1074/jbc.M111.333252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Chang YW, Lin TW, Li YC, Huang YS, Sun YJ, Hsiao CD. Interaction Surface and Topology of Get3-Get4-Get5 Protein Complex, Involved in Targeting Tail-anchored Proteins to Endoplasmic Reticulum. J Biol Chem. 2012;287:4783–4789. doi: 10.1074/jbc.M111.318329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Mariappan M, Li X, Stefanovic S, Sharma A, Mateja A, Keenan RJ, Hegde RS. A ribosome-associating factor chaperones tail-anchored membrane proteins. Nature. 2010;466:1120–1124. doi: 10.1038/nature09296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Winnefeld M, Grewenig A, Schnölzer M, Spring H, Knoch TA, Gan EC, Rommelaere J, Cziepluch C. Human SGT interacts with Bag-6/Bat-3/Scythe and cells with reduced levels of either protein display persistence of few misaligned chromosomes and mitotic arrest. Exp Cell Res. 2006;312:2500–2514. doi: 10.1016/j.yexcr.2006.04.020. [DOI] [PubMed] [Google Scholar]
- 40.Leznicki P, Clancy A, Schwappach B, High S. Bat3 promotes the membrane integration of tail-anchored proteins. J Cell Sci. 2010 doi: 10.1242/jcs.066738. [DOI] [PMC free article] [PubMed] [Google Scholar]
- *41.Hessa T, Sharma A, Mariappan M, Eshleman HD, Gutierrez E, Hegde RS. Protein targeting and degradation are coupled for elimination of mislocalized proteins. Nature. 2011;475:394–397. doi: 10.1038/nature10181. This report shows that the Bag6 complex plays a general triaging role for membrane proteins in the cytoplasm selecting for degradation or the TA pathway. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Minami R, Hayakawa A, Kagawa H, Yanagi Y, Yokosawa H, Kawahara H. BAG-6 is essential for selective elimination of defective proteasomal substrates. J Cell Biol. 2010;190:637–650. doi: 10.1083/jcb.200908092. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Structures of Get3 tetramers. A. Tetrameric MjGet3 (PDB ID: 3UG6) shown in a cartoon representation with one subunit color ramped from N to C-terminus and additional subunits colored gray, blue and green. B. A crystallographic tetramer of ScGet3 (PDB ID: 3SJD, shown as in A) in complex with the soluble domain of Get2 (cyan).
Conformations of the NHDs of NifH and ParA in different nucleotide states. A. A structural alignment of NifH NHDs represented as in Fig. 2C. Apo (PDB ID: 2AFH), ADP bound (PDB ID: 2AFI), ADP·AlF4− bound (PDB ID: 1M34) and AMPPCP bound (PDB ID: 2AFK) forms are shown. B. As in A for ParA structures. Apo (PDB ID: 3EZ7, PDB ID: 3EZ9, PDB ID: 3EZF), ADP bound (PDB ID: 3CWQ, PDB ID: 3EZ2) and ATP/ATPγS bound (PDB ID: 3EA0, PDB ID: 2OZE) forms are shown.