

Summary
Only two decades ago antibodies to fungi were thought to have little or no role in protection against fungal diseases. However, subsequent research has provided convincing evidence that certain antibodies can modify the course of fungal infection to the benefit or detriment of the host. Hybridoma technology was the breakthrough that enabled the characterization of antibodies to fungi, illuminating some of the requirements for antibody efficacy. As discussed in this review, fungal specific antibodies mediate protection through direct actions on fungal cells and through classical mechanisms such as phagocytosis and complement activation. Although mechanisms of antibody-mediated protection are often species-specific, numerous fungal antigens can be targeted to generate vaccines and therapeutic immunoglobulins. Furthermore, the study of antibody function against medically important fungi has provided fresh immunological insights into the complexity of humoral immunity that are likely to apply to other pathogens.
The past two decades have been momentous in advancing our understanding of the role of antibody-mediated immunity (AMI) in host defense against fungi and they have brought about a paradigm shift in our thinking on this question. Prior to the 1990s AMI was considered to be irrelevant in host defense against fungi (for review see (Casadevall, 1995)) as the experimental methods that were in use at the time were not able to consistently establish a role for AMI. These methods, including passive transfer of immune sera to naive hosts and correlating the presence of serum antibody with immunity to fungal disease often yielded negative results and there was a lack of association between invasive fungal diseases and known antibody defects in humans. By contrast, ample evidence that cell-mediated immunity (CMI) was essential for resistance to fungal diseases resulted in CMI being viewed as the arm of the immune system responsible for host defense against fungi. In a prior essay we described the practice of characterizing microbes by whether host defense against them was dependent on AMI or CMI as the ‘great immunological catastrophe of the 20th century’ because this subdivision limited research on microbial pathogenesis and immunity to a single arm of the immune system, ignoring the other/s (Casadevall and Pirofski, 2011). However, for fungi the situation changed rapidly after Dromer and her colleagues showed that a monoclonal antibody (mAb) to Cryptococcus neoformans was protective against lethal cryptococcal infection in mice (Dromer et al., 1987b). At about the same time a protective mAb was reported against the fungus Pneumocistis carinii although this organism was thought to be a protozoan at the time (Gigliotti and Hughes, 1988). Subsequently, protective mAbs have been successfully generated against five medically important fungi (Table 1). The fact that certain antigens recognized by some of the aforementioned mAbs are expressed by different fungi has raised optimism that universal anti-fungal vaccines that protect via AMI could be generated.
Table 1.
Fungal antigens that have been shown to elicit protective antibodies
| Fungus | Antigen | Reference |
|---|---|---|
| Aspergillus fumigatus | Beta-1,3 glucan | (Torosantucci et al., 2009) |
| Cell wall glycoprotein | (Chaturvedi et al., 2005) | |
| Candida albicans | Secreted aspartyl proteinase2 | (Sandini et al., 2011) |
| Beta-1,3 glucan | (Torosantucci et al., 2009; Torosantucci et al., 2005) | |
| Mp58 mannoprotein | (Viudes et al., 2004) | |
| Beta-(1–2)-linked mannotriose | (Han et al., 1997) | |
| Heat-shock protein 90 | (Matthews et al., 1991) | |
| Als3 | (Brena et al., 2007) | |
| Phosphoglycerate kinase | (Calcedo et al., 2011) | |
| Fructose bisphophate aldolase | (Calcedo et al., 2011) | |
| Cryptococcus neoformans | Beta-1,3 glucan | (Torosantucci et al., 2009) |
| Glucuronoxylomannan (GXM) | (Dromer et al., 1987a; Sanford et al., 1990; Mukherjee et al., 1992; Fleuridor et al., 1998; Beenhouwer et al., 2007) | |
| glucosylceramide | (Rodrigues et al., 2000) | |
| Melanin | (Rosas et al., 2001) | |
| Histoplasma capsulatum | Heat shock protein60 | (Guimaraes et al., 2009b) |
| Histone-like protein | (Nosanchuk et al., 2003) | |
| 70 KD cell surface protein | (Lopes et al., 2010a) | |
| Paracoccidioides brasiliensis | gp43 | (Buissa-Filho et al., 2008) |
| Melanin | (da Silva et al., 2006) | |
| Pneumocystis spp | Glycoprotein A | (Gigliotti et al., 1998) |
| Major surface glycoprotein | (Smulian et al., 2000) |
The breakthrough that made the identification of protective antibodies to fungi possible was the mAb technology. In contrast to polyclonal sera, mAbs provided defined reagents that recognized a single antigenic determinant and yielded consistent and reproducible results. Furthermore, and importantly, studies with mAbs led to the discovery that depending on their specificity and isotype, mAbs to fungi can mediate three different effects in being protective, non-protective (indifferent) or disease-enhancing. The observed disease-enhancing properties of mAbs provided an explanation for historical difficulties in establishing a role for AMI with polyclonal preparations, as these intrinsically heterogeneous reagents were likely to contain a variety of antibodies in varying proportions with each of the foregoing activities. Hence, studies with mAbs established definitively that protective immunoglobulins to fungi can be produced and that the historical inability to establish a role for AMI in protection against fungi was most likely a function of the heterogeneous preparations used rather than a fundamental limitation of AMI.
Given the rising tide of mycotic diseases, understanding the role of AMI in host defense against fungi is particularly important. Mycotic diseases have increased significantly as a result of use of antibacterial agents, which alter the host associate microbiota and immunosuppressive therapies, which induce impaired immunity. Most invasive fungal infections occur in patients with impaired immunity due to one of the interventions noted above or acquired immunodeficiency, such as HIV/AIDS. Given that immunosuppression enhances fungal pathogenesis, it is not surprising that despite ample in vitro activity, antifungal drugs often cannot eradicate the fungal burden in patients with impaired immunity. As such, fungal diseases are difficult to treat, have high morbidity and mortality, and often result in latency that can reactivate in the setting of immune suppression. Consequently, there is a great need to develop new approaches to treat and prevent fungal diseases. Immunoglobulins offer a very attractive approach. Vaccines that elicit AMI have a long track record of efficacy against many infectious diseases, and having served as inaugural antimicrobial agents in the preantibiotic era, immunoglobulins are increasingly being considered as potential therapies for a variety of microbes (Casadevall et al., 2004). Consequently, there is now great interest in the development of vaccines and therapeutic antibodies for fungi but progress in this area must contend with the complexity of AMI for this group of pathogens.
In this review, we discuss the tremendous progress that has been made in the field of AMI to fungal pathogens with emphasis on how information learned with the fungi has informed immunology on basic mechanisms of antibody action.
Challenges posed by fungal pathogens
In containing fungi the immune system faces unique challenges unlike those posed by bacteria and viruses. Many fungal pathogens exhibit dimorphism, a characteristic that enables them to exist as hyphae or yeast cells, with each form having a distinct surface and antigenic composition. Fungal cells are surrounded and protected by a cell wall, which is a structure composed of crosslinked polysaccharides including chitin and other components. An important functional effect of the fungal cell wall is to inhibit the ability of complement to damage fungal cell membranes, even though complement activation can elicit an antifungal inflammatory response. Hence, antibody-dependent complement activation does not result in direct damage to fungal cells. However, there are now several examples of mAbs that bind to cell wall components and inhibit replication (Table 2). Although the mechanism by which mAbs bind and affect the fungal cell wall is not understood, it is possible that the complex nature of the cell wall requires a tightly regulated metabolism that is disrupted by the attachment of immunoglobulin molecules. Fungal cells are notorious for producing secondary metabolites with deleterious effects. For example, mannitol production can interfere with host oxidative defenses and mycotoxins such gliotoxin are toxic to immune effector cells. Adding to the difficulties encountered by the immune system fungal pathogens can exhibit antigenic variation. The phenomenon of fungal dimorphism means that many pathogenic fungi can exist in two different forms in the host, each of which is antigenically distinct. For C. neoformans, antigenic diversity stems from the capsular polysaccharide and it is estimated that each cell is antigenically unique owing to combinatorial diversity of the polysaccharide capsule (McFadden et al., 2007). In addition, the C. neoformans capsule is antiphagocytic and capable of quenching free radicals produced by host immune cell oxidative burst (Zaragoza et al., 2008). Capsular polysaccharide released by C. neoformans cells in tissue interferes with the inflammatory response and inhibits recruitment and the function of effector cells that are empowered by specific antibodies. Fungi are also capable of forming very large structures in tissue. Aspergillus spp. form dense collections of hyphae in tissue that induce surrounding tissue necrosis and C. neoformans and Coccidioides immitis can each generate enormous structures in tissue know as giant or titan cells and spherules, respectively. C. neoformans giant cells are far larger than macrophages, precluding phagocytosis and posing a formidable problem for host defenses (Zaragoza et al., 2010; Okagaki et al., 2010). Although fungi exert a multitude of effects that thwart host defense, these very effects provide rational targets for AMI as antibodies to fungal cell wall determinants, metabolites and capsular polysaccharides can serve to inhibit their deleterious effects.
Table 2.
Mechanisms of antibody-mediated protection against fungi
| Mode | Mechanism | Comment | References |
|---|---|---|---|
| Indirect | Phagocytosis | This mechanism may be particularly important for fungi that actively resist phagocytosis such as C. neoformans. However, phagocytosis may not be sufficient as a host protective mechanism since many fungi can survive in the intracellularly. | (Schlagetter and Kozel, 1990) |
| Complement activation | Appears to be critical for IgM but may be dispensable for IgG | (Shapiro et al., 2002; Han et al., 2001) | |
| Antibody-directed cellular cytotoxicity | Best example is the requirement for specific antibody in NK cell-mediated antifungal effects against C. neoformans | (Nabavi and Murphy, 1986) | |
| Phagosome activation | mAbs to H. capsulatum promote phagosome maturation and thus interfere with fungal mechanisms of intracellular survival | (Shi et al., 2008) | |
| Modulation of the inflammatory response | mAbs to C. neoformans mediate changes in inflammatory responses that are associated with better control of infection in tissue | (Feldmesser et al., 2002; Feldmesser and Casadevall, 1997) | |
| Direct | Inhibition of biofilm formation | An in vitro observation for C. neoformans that could reflect the ability of specific antibody to the capsule to interfere with polysaccharide release | (Martinez and Casadevall, 2005) |
| Inhibition of polysaccharide release | Given the many deleterious effects associate with polysaccharide in tissue the ability of antibody to reduce capsule shedding from C. neoformans could translate into more effective immune responses | (Martinez et al., 2004) | |
| Inhibition of replication and cytotoxicity | The mechanism is unknown but antibodies that bind cell wall components can inhibit replication and/or trigger cell death. | (Brena et al., 2007; Moragues et al., 2003; Magliani et al., 2005; Rosas et al., 2001) | |
| Interference with dimorphic changes | Some mAbs to C. albicans interfere with germ tube formation | (Moragues et al., 2003) | |
| Alterations in gene expression | Antibody binding to the C. neoformans capsule triggers changes in gene expression that can make the fungus more vulnerable to antifungal agents | (McClelland et al., 2010) | |
| Iron starvation | Iron starvation appears to be the mechanism by which a mAb is cytotoxic and interferes with morphogenesis | (Brena et al., 2011) | |
| Mimicking killer toxin | Anti-idiotypic antibodies to yeast killer toxin mimic the structure of killer toxin and are active against many fungal species | (Magliani et al., 2005) |
Mechanisms of AMI against fungi
Most current knowledge about mechanisms of AMI against fungi comes from studies with C. neoformans and C. albicans, which are among the most common and most studied human pathogenic fungi. Historically, mechanisms of AMI, or classical mechanisms, were held to include opsonization, complement activation, viral and toxin neutralization and antibody-dependent cellular cytotoxicity (ADCC) (Casadevall and Pirofski, 2011). Among these mechanisms, viral neutralization is not immediately relevant, although some of the mechanisms by which viral neutralization is mediated could have parallels in fungal-antibody interactions. Each of the other classical functions could have a role in host defense against fungi. For example, opsonization is very important for host defense against C. neoformans as the polysaccharide capsule abrogates phagocytosis and complement is not an effective opsonin for some strains (Zaragoza et al., 2003). Whether ingestion is an effective mechanism for controlling C. neoformans depends on the activation state of the cell and the immunological state of the host as this fungus has the capacity for intracellular replication and can even escape the phagocytic cell by non-lytic exocytosis. In fact, intracellular replication can be faster than extracellular growth. In animal models of cryptococcosis, complement activation appears to be essential for the function of specific IgM, but may be dispensable for IgG (Shapiro et al., 2002). In theory, toxin neutralization could contribute to host defense against fungi that produce mycotoxins, but this has not been demonstrated to date. Fc receptor-dependent ADCC can mediate protection against C. albicans (van Spriel et al., 2001) and antibody-dependent NK cell-mediated activity against C. neoformans has been demonstrated (Miller et al., 1990; Nabavi and Murphy, 1986). Another antibody-mediated antifungal mechanism is exemplified by the ability of anti-idiotypic antibodies mimicking killer toxin to directly damage fungal cells by interacting with the killer toxin receptor (Magliani et al., 2005).
In addition to the aforementioned classical mechanisms of AMI, new mechanisms of antibody action against fungi have been identified (Table 2). One such mechanism is that antibodies to certain fungal determinants are able to mediate direct antifungal effects without the need for host cells. For example, a mAb to Candida albicans mannoprotein has been shown to mediate three different activities: inhibition of adherence, inhibition of germination, and direct candidacidal activity (Moragues et al., 2003), with the the direct effects involving antibody-mediated iron starvation (Brena et al., 2011). In addition, antibody-mediated inhibition of an aspartyl proteinase has been associated with protection against vaginal candidiasis (De Bernardis et al., 1997), and antibodies to 1,2 β mannotriose (Xin et al., 2008) and cell wall protein-derived peptides protect against disseminated candidiasis in mice (Xin and Cutler, 2011). Antibodies to cell wall β-glucans have been shown to interfere with adherence and cell wall remodeling (Torosantucci et al., 2005; Torosantucci et al., 2009) and have demonstrated the capacity for direct antifungal activity via growth inhibition of C. albicans as well as Cryptococcus neoformans (Torosantucci et al., 2009). As exemplified by the direct anti-fungal and growth-inhibitory capacity of β-glucan antibodies, and the ability of AMI elicited by a cell wall β-glucan-based vaccine to protect against multiple fungi (Torosantucci et al., 2005), studies with mAbs have provided proof of principle that the development of universal fungal therapies and vaccines that provide AMI upon binding conserved fungal β-glucans could be possible.
For Cryptococcus neoformans, studies with mAbs have demonstrated several direct antibody-mediated effects including interference with capsular polysaccharide release and biofilm formation. However, there is also considerable evidence that specific antibody to the C. neoformans capsule can mediate protection indirectly. These indirect mechanisms of protection alter the inflammatory response and depend on collaboration with effector cells and/or CMI. In fact, the primary mechanism of antibody-mediated protection against C. neoformans is most likely via modification of the inflammatory response in a manner that that enhances fungal clearance and minimizes inflammatory damage to tissue (reviewed in (Casadevall and Pirofski, 2003)). Specific antibody can also alter the nature of C. neoformans-macrophage interaction as exemplified by data showing that antibody-opsonized C. neoformans exits macrophages as biofilm-like microcolonies, whereas planktonic cells exit after complement opsonization (Alvarez et al., 2008). A similar mechanism may underlie antibody-mediated protection against murine histoplasmosis, as antibody administration was associated with reduced inflammatory damage (Guimaraes et al., 2009a). The mechanism for these effects may be a result of antibody-mediated changes in the H. capsulatum-macrophage interaction as antibody-dependent opsonization enhanced phagosome activation, acidification and maturation, which can result in more efficient T cell activation and more effective CMI (Shi et al., 2008).
AMI in natural resistance to fungi
Cryptococcus neoformans
Despite overwhelming evidence that individual mAbs can modify the course of fungal infection to the benefit of the host (see Table 1), a definitive role for natural AMI in host defense against fungi has been difficult to establish. Historically, the lack of 1) a clear association between known B cell defects and a predisposition to fungal disease and 2) a clear relationship between serum antibody to fungal antigens and resistance to fungi led most to conclude that AMI has no role in host defense against fungi in patients. However, this conclusion was flawed as it was based on negative (a lack) of association. Part of the difficulty in establishing a relationship between antibody and resistance to fungal disease is that most immunologically intact individuals produce many antibodies that bind fungal antigens, entities to which they are constantly exposed. In fact, immunologically intact individuals are at very low risk of ever developing disease. Since antibodies to major fungal pathogens, such as Cryptococcus neoformans and C. albicans are commonly present in individuals who have no history of disease as well as many who do, the presence of serum antibody does not discriminate those who are immune from those who are susceptible (Casadevall and Pirofski, 2007). However, the same information could be interpreted differently. For example, one could argue that the high prevalence of serum antibodies to fungal determinants in normal individuals together with the very low incidence of fungal disease in this group is consistent with a protective role for AMI against fungi. Further, some of these observations might be reconciled if AMI is beneficial when present but not necessarily required. Nonetheless, serum antibody can identify individuals who are susceptible to reactivation-associated disease. For example, as serological evidence against C. neoformans is detectable early in childhood (Goldman et al., 2001; Abadi and Pirofski, 1999), the presence of specific antibody has been cited to suggest that solid organ transplantation-associated cryptococcosis stems from reactivation of latent infection in patients with prior infection (Saha et al., 2007). However, such data also imply that preexisting antibody did not prevent reactivation. This is not surprising in view of the critical importance of CMI in resistance to C. neoformans and ample evidence that natural resistance to disease requires both AMI and CMI (Rivera et al., 2005; Yuan et al., 1997; Beenhouwer et al., 2001). Nonetheless, cryptococcal disease has been reported in patients with B cell defects, including X-linked hyperIgM (Winkelstein et al., 2003; de et al., 2005), a disorder with B, in addition to T cell defects (Agematsu et al., 1998), and hypogammaglobulinemia (Gupta et al., 1987; Wahab et al., 1995; Neto et al., 2000). In addition, IgG2 deficiency was identified in an otherwise normal individual who developed meningitis with C. gattii (Marr et al., 2012) and compared to those without HIV/AIDS, adults and children with HIV/AIDS have lower titers of specific IgG2 the cryptococcal capsular polysaccharide glucuronoxylomannan (GXM) (Abadi and Pirofski, 1999; Abadi and Pirofski, 1999; DeShaw and Pirofski, 1995). IgG2 is the predominant IgG subclass to polysaccharide antigens in humans (Pirofski, 2001).
The most common immune deficiency among patients with cryptococcal disease is HIV and HIV-associated B cell defects have been linked to cryptococcal disease susceptibility. HIV-infected participants in the Multicenter AIDS Cohort Study (MACS) who developed cryptococcosis had lower levels of serum antibody expressing VH3 genes and IgM to GXM than those who did not. Similar observations have been reported with HIV-infected individuals in other cohorts (DeShaw and Pirofski, 1995; Fleuridor et al., 1999; Subramaniam et al., 2009). The loss of B cells and serum antibody expressing VH3 depletion is a well-documented HIV-associated defect (Berberian et al., 1994; Berberian et al., 1993). Hence, as human antibodies to GXM use VH3 genes (Fleuridor et al., 1998; Fleuridor et al., 1999; Pirofski et al., 1995), HIV-associated depletion of VH3-expressing B cells could translate into a hole in the GXM-reactive repertoire. Another HIV-associated deficit that has been linked to cryptococcal disease susceptibility is IgM memory B cell depletion. Identified as one of the original HIV-associated B cell deficiencies (Lane et al., 1983), IgM memory B cell depletion remains a complication of HIV/AIDS even in the HAART era (Moir and Fauci, 2009) and has been linked to HIV-associated cryptococcal disease susceptibility (Subramaniam et al., 2009). IgM memory B cells are the source of natural IgM that has the capacity to bind conserved microbial carbohydrates (Baumgarth et al., 2005; Griffin et al., 2011). Hence, the lower levels of IgM to GXM that have been reported in the sera of patients with HIV/AIDS (DeShaw and Pirofski, 1995; Fleuridor et al., 1999; Subramaniam et al., 2009) as well as pre-transplant sera of patients who developed cryptococcal disease after solid organ transplantation (Jalali et al., 2006) could contribute to the development of cryptococcal disease in addition to being a possible sentinel for IgM memory B cell deficiency.
In contrast to the uncertain relationship between AMI and resistance to fungi that has arisen from serological studies in humans, investigations in experimental animals provide strong evidence for a contribution of natural AMI to host defense against fungi. An early study demonstrated increased susceptibility to C. neoformans in mice with X-linked immunodeficiency (Marquis et al., 1985). More recently, B cell deficient (Rivera et al., 2005) and IgM-deficient (sIgM−/−) (Subramaniam et al., 2010b) mice have each been shown to be more susceptible to pulmonary infection with C. neoformans. Interestingly, when sIgM−/− mice were infected intraperitoneally, they were protected against C. neoformans (Subramaniam et al., 2010a). Seemingly paradoxical, this finding underscores the complexity of AMI to fungi and highlights that it is often tissue specific. Hence, while secreted IgM appears to be crucial for granuloma formation and preventing dissemination of C. neoformans from the lungs (Subramaniam et al., 2010b), an excess of B-1 B cells and IgG2 (which is characteristic of sIgM−/− mice) appears to mediate protection in the peritoneal cavity despite the absence of IgM, most likely by enhancing the fungicidal activity of macrophages (Subramaniam et al., 2010a). Hence, IgM and B cells exhibit different, tissue specific roles in resistance to cryptococccal dissemination and death in mice.
Candida albicans
Observational studies in patients with candidiasis led to the identification of antibodies to an antigen, subsequently recognized as hsp90, that were associated with survival (Matthews et al., 1991). This observation spawned work that led to the development of an antibody-based fragment for treatment of fungal disease, Mycograb (Pachi et al., 2006). Although Mycograb enhanced the activity of antifungal agents against C. albicans in mice (Matthews et al., 2003), and improved outcome in invasive candidiasis in combination with liposomal amphotericin in a randomized, multicenter clinical trial (Pachi et al., 2006), its clinical development has been discontinued (see below). The complex states of human C. albicans infection, from commensalism to disease, present a unique challenge in efforts to characterize the role of AMI in resistance to disease and harness the potential of antibody-based therapy in patients. Studies in mice have been more straightforward, but have yielded different results depending on the fungal strain, route of infection and animal model. B cell deficient mice succumbed more rapidly than wild type mice when infected with filamentous forms of C. albicans (Saville et al., 2008), and mice depleted of B cells were more susceptible (Sinha et al., 1987). In another model, B cell deficient mice survived primary infection with C. albicans and Aspergillus fumigatus (Montagnoli et al., 2003), but C. albicans-infected mice did not survive a secondary challenge due to a failure of immunoregulation, rather than fungal clearance. Normal mouse serum reversed the effects and reconstituted a survival phenotype (Montagnoli et al., 2003). This study reiterated findings from the aforementioned C. neoformans models in which AMI conferred protection via immunoregulation and control of inflammation.
Other fungal pathogens
In contrast to C. albicans and C. neoformans, B cell depletion did not alter mortality in mice with primary H. capsulatum infection (Allendoerfer et al., 1999), however, B cells protected against fungal reactivation in immune mice (Allen and Deepe, Jr., 2006).
B cells have also been shown to have an immunoregulatory role in immunity to Pneumocystis in mice (Lund et al., 2006). B-1 B cells and natural IgM to conserved β-glucan determinants secreted by B-1 B cells were shown to enhance innate immunity to Pneumocystis via dendritic cell priming (Rapaka et al., 2010), providing another instance of IgM-mediated anti-fungal immunity as demonstrated for C. neoformans (Subramaniam et al., 2010b). Further evidence for AMI-mediated resistance to Pneumocystis is found in a report that simian immunodeficiency virus-infected macaques with IgG to a Pneumocystis protein (KEX-1) were more resistant to Pneumocystis colonization and lung disease than macaques without IgG (Rapaka et al., 2010). In contrast to their protective role in Pneumocystis and intraperitoneal infection with C. neoformans (as IgM deficient mice have elevated levels of B-1 B cells (Subramaniam et al., 2010a)), B-1 B cells had a detrimental effect in pulmonary P. brasilienses infection in mice, albeit in a different mouse model in which disease enhancement was attributed to B-1 B cell production of IL-10 and impairment of phagocytosis (Popi et al., 2008).
AMI has been shown to be important for the generation of long lasting immunity to C. albicans and A. fumigatus (Montagnoli et al., 2003), and as noted above for maintenance of immunity to H. capsulatum (Allen and Deepe, Jr., 2006). In addition to IgM, IgA could also play a role in natural resistance to C. albicans as tetraspanin protein (CD37) deficient mice, which have 15-fold higher levels of IgA, were less susceptible than wild-type mice and sera from CD37 deficient mice enhanced resistance of wild-type mice (van Spriel et al., 2009).
Contribution of AMI in pathogenesis and susceptibility to fungi
Given that AMI is a critical arm of the immune response, and fungi cause disease in the setting of immunodeficiency and excessive immune responses, i.e. at both ends of the damage-response spectrum (Casadevall and Pirofski, 2011), it is likely that some antibody responses can be deleterious. For example, IgE to fungal antigens has been implicated in the development of allergic pulmonary aspergillosis, fungal sinusitis and forms of asthma that are caused by C. neoformans (Goldman et al., 2006). However, evidence that AMI contributes to the pathogenesis of and susceptibility to invasive fungal disease is indirect at best, with most data coming from experimental animal models. The capacity of AMI to enhance fungal disease was discovered with the identification of disease-enhancing mAbs that shortened survival when they were administered to C. neoformans-infected mice (Nussbaum et al., 1996). However, enhancement was limited to certain mouse strains and the same mAb that was enhancing in one strain was protective in another (Rivera and Casadevall, 2005). Recently, disease-enhancing monoclonal antibodies have also been reported for Scedosporium apiospermum, a soil fungus that causes invasive and non-invasive infection in susceptible individuals (Lopes et al., 2010b). A comparison of Balb/c and C.B-17 mice, which differ in their capacity to clear pulmonary C. neoformans infection revealed that human immunoglobulin locus genes were associated with slower clearance in Balb/c mice (Lovchik et al., 1999). The enhanced susceptibility of Balb/c mice to cryptococcosis has been associated with significantly higher levels of C. neoformans-specific IgM, IgG1, IgG2a and IgG3 and Th2 polarization of the pulmonary inflammatory response (Lovchik et al., 1999). In contrast, susceptible C57Bl/6 mice had higher levels of IgG2b after intraperitoneal infection with C. neoformans (Subramaniam et al., 2010a). IgG2b has been associated with inferior opsonic activity in vitro (Sanford et al., 1990) for some mAbs, but not others (Yuan et al., 1998). Methamphetamine administration increased susceptibility to H. capsulatum in mice, a phenomenon associated with pleitropic effects on the immune system including alterations in antibody responses (Martinez et al., 2009). For C. albicans, a comparison of genetically resistant and susceptible mouse strains revealed qualitative and quantitative differences in the antibody response, but such results do not infer causality (Costantino et al., 1995). Similarly, differences in the susceptibility of mice to Paracoccidioides brasilensis correlated with differences in the isotype composition such that resistant and susceptible strains produced IgG2a and IgG2b isotypes, respectively (Kashino et al., 2000). However, we note that these differences occurred in a background of many immunological differences, making the degree to which AMI contributed to disease pathogenesis unclear. On the other hand, specific IgG was associated with resistance to colonization and lung disease after exposure to Pneumocystis in macaques (Kling et al., 2010) and IgM deficient mice that exhibited resistance to intraperitoneal infection with C. neoformans had higher levels of IgG2a than susceptible mice (Subramaniam et al., 2010a). However, it should be noted that a given antibody type can be the byproduct of a Th2-polarized response, thus being only a small part of a complex B and T cell response. Thus, links between the presence of antibody and an outcome of fungal infection are associations that do not imply or establish a causal relationship between AMI and an outcome of fungal infection. Nonetheless, immune responses to fungi are considered notorious for eliciting antibodies that are not considered to be protective as exemplified for antibodies to Candida (Farah and Ashman, 2005) and Coccidiodes spp. (Cole et al., 2004) and the phenomenon may be more widespread.
Efficacy of induced AMI against fungi in prophylaxis and therapy
Given the high mortality, chronicity, and refractoriness of fungal diseases to therapy, there is currently considerable interest in the use of antibodies to prevent disease and as adjuncts to standard antifungal therapy. AMI can be induced actively and passively. The efficacy of passive antibody administration is convincingly established against various fungi (Table 1) and several vaccine formulations have also been shown to mediate protection by eliciting protective antibody responses in mice (Table 3). One concern in inducing AMI is that prozone-like effects have been demonstrated against fungi, in which loss of protective efficacy is apparent at high relative to lower antibody concentrations (Taborda et al., 2003). Hence, it is conceivable that highly immunogenic vaccines could fail if they elicit exuberant antibody responses with high antibody titers. Nevertheless, the experience with many effective vaccines against bacterial and viral pathogens provides strong encouragement that effective antifungal vaccines can be developed. A vaccine against recurrent vulvovaginal candidiasis known as PEV7 which expresses recombinant aspartyl protease on virosomes produced by Pevion Biotech AG (Bern, Switzerland) is currently in clinical evaluation. A murine IgG1 mAb for C. neoformans that was studied in a clinical trial was found to be safe and to reduce serum GXM when used at high doses (Larsen et al., 2005). Further clinical development of this mAb has been hampered by economic issues resulting from the high costs of drug development for a disease of relatively low prevalence. A recombinant antibody fragment to C. albicans HSP90 (efungumab, Mycograb) showed promising efficacy in clinical trials (Pachi et al., 2006) but subsequent development of this immunoglobulin derivative has been hampered by production difficulties and was abandoned by Novartis (http://www.novartis.com/newsroom/media-releases/en/2010/1449020.shtml). Despite the absence of licensed products that prevent or treat fungal diseases based on AMI, the fact that antibodies and vaccines have been tested clinically is a measure of the interest in and acceptance of the concept that the pursuit of AMI for fungal diseases warrants consideration in clinical medicine. Furthermore, newer types of antibody preparations are on the horizon. For example, the efficacy of antibody can be enhanced by linking it to a radioactive isotope since this converts the antibody in a fungicidal molecule (Dadachova et al., 2003) irrespective of its natural biological function. In fact, by targeting antigens shared across fungal species it may be possible to develop broad spectrum radioimmunotherapy for fungal diseases (Bryan et al., 2011). This could be an attractive option for developing new antifungal strategies against the devastating mold infections that occur in severely immunocomprised individuals.
Table 3.
Experimental vaccines that protect through AMI
| Vaccine | Target | Mechanism | Reference |
|---|---|---|---|
| Glucuronoxylomannan-tetanus toxoid conjugate | C. neoformans | Vaccine elicits antibody responses to the polysaccharide capsule | (Devi, 1996) |
| Peptide mimotope | C. neoformans | Peptide elicits antibody responses that react with the capsule | (Fleuridor et al., 2001) |
| Synthetic glycopeptide | C. albicans | Antibodies to cell wall beta mannans | (Xin et al., 2008) |
| Mannan-protein conjungate | C. albicans | Antibodies to cell wall beta mannans | (Han et al., 1999) |
| Beta-glucan conjugate | C. albicans, A. fumigatus, C. neoformans | Antibodies to cell wall beta glucans | (Torosantucci et al., 2005; Torosantucci et al., 2009) |
| Recombinant and truncated aspartic protease | C. albicans | Protease neutralization | (Sandini et al., 2011) |
| Glycan-peptide conjugates | C. albicans | Antibodies to cell wall | (Xin and Cutler, 2011) |
Summary
In a little more than two decades the field of AMI to medically important fungi has gone from being a remote backwater of scientific study to a vibrant field that is now generating promising vaccines and therapeutic immunoglobulins while making major contributions to basic immunology and microbial pathogenesis. In fact, we have recently proposed a new synthesis for AMI based largely on discoveries made with antibodies to fungi (Casadevall and Pirofski, 2011). Regardless of this progress and despite the work of many investigators it is clear that we have barely scratched the surface in this complex topic. We anticipate that in the coming years many new targets of AMI will be identified and hopefully the promising antibody-based reagents and vaccines that are currently in various stages of pre-clinical and clinical development will lead to licensed products that will transform our approach to fungal diseases. However, in our view, the greatest challenge facing this field, and all of immunology, is how to integrate AMI with the other components of the immune system to produce a coherent picture of effective and non-effective immune responses that would be predictive and useful in guiding the design of new vaccines and immune therapies.
Figure 1.
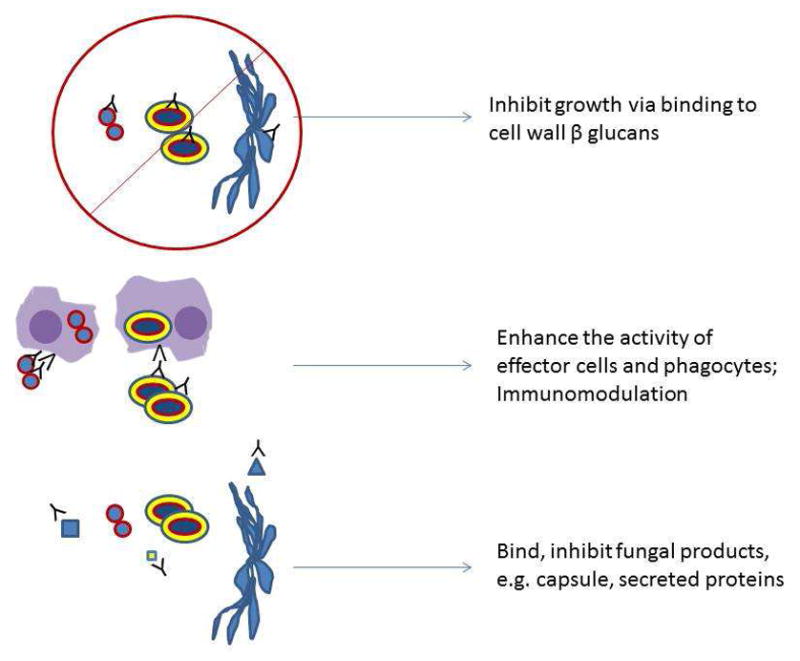
The proposed mechanisms of antibody action against fungi can be considered under three general categories involving A) direct inhibition of growth, B) immunomodulation and potentiation of innate immune mechanisms and C) neutralization of the untoward effects of fungal products on host tissues.
Acknowledgments
AC is supported by NIH grants NIH awards HL059842, AI033774, AI033142, AI052733 and Center for AIDS Research at Albert Einstein College of Medicine. LP received support from NIH grants R01-AI045459 and R01-AI044374.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Reference List
- Abadi J, Pirofski L. Antibodies reactive with the cryptococcal capsular polysaccharide glucuronoxylomannan are present in sera from children with and without HIV infection. J Infect Dis. 1999;180:915–919. doi: 10.1086/314953. [DOI] [PubMed] [Google Scholar]
- Agematsu K, Nagumo H, Shinozaki K, Hokibara S, Yasui K, Terada K, Kawamura N, Toba T, Nonoyama S, Ochs HD, Komiyama A. Absence of IgD-CD27(+) memory B cell population in X-linked hyper-IgM syndrome. J Clin Invest. 1998;102:853–860. doi: 10.1172/JCI3409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Allen HL, Deepe GS., Jr B cells and CD4-CD8− T cells are key regulators of the severity of reactivation histoplasmosis. J Immunol. 2006;177:1763–1771. doi: 10.4049/jimmunol.177.3.1763. [DOI] [PubMed] [Google Scholar]
- Allendoerfer R, Brunner GD, Deepe GS., Jr Complex requirements for nascent and memory immunity in pulmonary histoplasmosis. J Immunol. 1999;162:7389–7396. [PubMed] [Google Scholar]
- Alvarez M, Saylor C, Casadevall A. Antibody action after phagocytosis promotes Cryptococcus neoformans and Cryptococcus gattii macrophage exocytosis with biofilm-like microcolony formation. Cell Microbiol. 2008;10:1622–1633. doi: 10.1111/j.1462-5822.2008.01152.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baumgarth N, Tung JW, Herzenberg LA. Inherent specificities in natural antibodies: a key to immune defense against pathogen invasion. Springer Semin Immunopathol. 2005;26:347–362. doi: 10.1007/s00281-004-0182-2. [DOI] [PubMed] [Google Scholar]
- Beenhouwer DO, Shapiro S, Feldmesser M, Casadevall A, Scharff MD. Both Th1 and Th2 cytokines affect the ability of monoclonal antibodies to protect mice against Cryptococcus neoformans. Infect Immun. 2001;69:6445–6455. doi: 10.1128/IAI.69.10.6445-6455.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beenhouwer DO, Yoo EM, Lai CW, Rocha MA, Morrison SL. Human immunoglobulin G2 (IgG2) and IgG4, but not IgG1 or IgG3, protect mice against Cryptococcus neoformans infection. Infect Immun. 2007;75:1424–1435. doi: 10.1128/IAI.01161-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berberian L, Goodglick L, Kipps TJ, Braun J. Immunoglobulin VH3 gene products: natural ligands for HIV gp120. Science. 1993;261:1588–1591. doi: 10.1126/science.7690497. [DOI] [PubMed] [Google Scholar]
- Berberian L, Shukla J, Jefferis R, Braun J. Effects of HIV infection on VH3 (D12 idiotope) B cells in vivo. J Acquir Immune Defic Syndr. 1994;7:641–646. [PubMed] [Google Scholar]
- Brena S, Cabezas-Olcoz J, Moragues MD, Fernandez dLI, Dominguez A, Quindos G, Ponton J. Fungicidal monoclonal antibody C7 interferes with iron acquisition in Candida albicans. Antimicrob Agents Chemother. 2011;55:3156–3163. doi: 10.1128/AAC.00892-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brena S, Omaetxebarria MJ, Elguezabal N, Cabezas J, Moragues MD, Ponton J. Fungicidal monoclonal antibody C7 binds to Candida albicans Als3. Infect Immun. 2007;75:3680–3682. doi: 10.1128/IAI.01840-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bryan RA, Guimaraes AJ, Hopcraft S, Jiang Z, Bonilla K, Morgenstern A, Bruchertseifer F, Del PM, Torosantucci A, Cassone A, Nosanchuk JD, Casadevall A, Dadachova E. Toward Developing a Universal Treatment for Fungal Disease Using Radioimmunotherapy Targeting Common Fungal Antigens. Mycopathologia. 2011 doi: 10.1007/s11046-011-9476-9. (in press) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buissa-Filho R, Puccia R, Marques AF, Pinto FA, Munoz JE, Nosanchuk JD, Travassos LR, Taborda CP. The monoclonal antibody against the major diagnostic antigen of Paracoccidioides brasiliensis mediates immune protection in infected BALB/c mice challenged intratracheally with the fungus. Infect Immun. 2008;76:3321–3328. doi: 10.1128/IAI.00349-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Calcedo R, Garcia AR, Abad A, Rementeria A, Ponton J, Hernando FL. Phosphoglycerate kinase and fructose bisphosphate aldolase of Candida albicans as new antigens recognized by human salivary IgA. Rev Iberoam Micol. 2011 doi: 10.1016/j.riam.2011.07.004. (in press) [DOI] [PubMed] [Google Scholar]
- Casadevall A. Antibody immunity and invasive fungal infections. Infect Immun. 1995;63:4211–4218. doi: 10.1128/iai.63.11.4211-4218.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Casadevall A, Dadachova E, Pirofski L. Passive antibody therapy for infectious diseases. Nature Microbiol Rev. 2004;2:695–703. doi: 10.1038/nrmicro974. [DOI] [PubMed] [Google Scholar]
- Casadevall A, Pirofski LA. Antibody-mediated regulation of cellular immunity and the inflammatory response. Trends Immunol. 2003;24:474–478. doi: 10.1016/s1471-4906(03)00228-x. [DOI] [PubMed] [Google Scholar]
- Casadevall A, Pirofski LA. Accidental virulence, cryptic pathogenesis, martians, lost hosts, and the pathogenicity of environmental microbes. Eukaryot Cell. 2007;6:2169–2174. doi: 10.1128/EC.00308-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Casadevall A, Pirofski LA. A new synthesis for antibody-mediated immunity. Nat Immunol. 2011;13:21–28. doi: 10.1038/ni.2184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chaturvedi AK, Kavishwar A, Shiva Keshava GB, Shukla PK. Monoclonal immunoglobulin G1 directed against Aspergillus fumigatus cell wall glycoprotein protects against experimental murine aspergillosis. Clin Diagn Lab Immunol. 2005;12:1063–1068. doi: 10.1128/CDLI.12.9.1063-1068.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cole GT, Xue JM, Okeke CN, Tarcha EJ, Basrur V, Schaller RA, Herr RA, Yu JJ, Hung CY. A vaccine against coccidioidomycosis is justified and attainable. Med Mycol. 2004;42:189–216. doi: 10.1080/13693780410001687349. [DOI] [PubMed] [Google Scholar]
- Costantino PJ, Gare NF, Warmington JR. Humoral immune responses to systemic Candida albicans infection in inbred mouse strains. Immunol Cell Biol. 1995;73:125–133. doi: 10.1038/icb.1995.20. [DOI] [PubMed] [Google Scholar]
- da Silva MB, Marques AF, Nosanchuk JD, Casadevall A, Travassos LR, Taborda CP. Melanin in the dimorphic fungal pathogen Paracoccidioides brasiliensis: effects on phagocytosis, intracellular resistance and drug susceptibility. Microbes Infect. 2006;8:197–205. doi: 10.1016/j.micinf.2005.06.018. [DOI] [PubMed] [Google Scholar]
- Dadachova E, Nakouzi A, Bryan RA, Casadevall A. Ionizing radiation delivered by specific antibody is therapeutic against a fungal infection. Proc Natl Acad Sci U S A. 2003;100:10942–10947. doi: 10.1073/pnas.1731272100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Bernardis F, Boccanera M, Adriani D, Spreghini E, Santoni G, Cassone A. Protective role of antimannan and anti-aspartyl proteinase antibodies in an experimental model of Candida albicans vaginitis in rats. Infect Immun. 1997;65:3399–3405. doi: 10.1128/iai.65.8.3399-3405.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de GM, Erice A, Gil A, Gutierrez J, Rivas P, Hernando C, Rodriguez MC. Cryptococcal meningitis in a patient with X-linked hyper-IgM1 syndrome. Scand J Infect Dis. 2005;37:526–528. doi: 10.1080/00365540510036570. [DOI] [PubMed] [Google Scholar]
- DeShaw M, Pirofski LA. Antibodies to the Cryptococcus neoformans capsular glucuronoxylomannan are ubiquitous in serum from HIV+ and HIV− individuals. Clin Exp Immunol. 1995;99:425–432. doi: 10.1111/j.1365-2249.1995.tb05568.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Devi SJN. Preclinical efficacy of a glucuronoxylomannan-tetanus toxoid conjugate vaccine of Cryptococcus neoformans in a murine model. Vaccine. 1996;14:841–842. doi: 10.1016/0264-410x(95)00256-z. [DOI] [PubMed] [Google Scholar]
- Dromer F, Charreire J, Contrepois A, Carbon C, Yeni P. Protection of mice against experimental cryptococcosis by anti-Cryptococcus neoformans monoclonal antibody. Infect Immun. 1987a;55:749–752. doi: 10.1128/iai.55.3.749-752.1987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dromer F, Salamero J, Contrepois A, Carbon C, Yeni P. Production, characterization, and antibody specificity of a mouse monoclonal antibody reactive with Cryptococcus neoformans capsular polysaccharide. Infect Immun. 1987b;55:742–748. doi: 10.1128/iai.55.3.742-748.1987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Farah CS, Ashman RB. Active and passive immunization against oral Candida albicans infection in a murine model. Oral Microbiol Immunol. 2005;20:376–381. doi: 10.1111/j.1399-302X.2005.00240.x. [DOI] [PubMed] [Google Scholar]
- Feldmesser M, Casadevall A. Effect of serum IgG1 against murine pulmonary infection with Cryptococcus neoformans. J Immunol. 1997;158:790–799. [PubMed] [Google Scholar]
- Feldmesser M, Mednick A, Casadevall A. Antibody-mediated protection in murine Cryptococcus neoformans infection is associated with subtle pleotrophic effects on the cytokine and leukocyte response. Infect Immun. 2002;70:1571–1580. doi: 10.1128/IAI.70.3.1571-1580.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fleuridor R, Lees A, Pirofski L. A cryptococcal capsular polysaccharide mimotope prolongs the survival of mice with Cryptococcus neoformans infection. J Immunol. 2001;166:1087–1096. doi: 10.4049/jimmunol.166.2.1087. [DOI] [PubMed] [Google Scholar]
- Fleuridor R, Lyles RH, Pirofski L. Quantitative and qualitative differences in the serum antibody profiles of human immunodeficiency virus-infected persons with and without Cryptococcus neoformans meningitis. J Infect Dis. 1999;180:1526–1535. doi: 10.1086/315102. [DOI] [PubMed] [Google Scholar]
- Fleuridor R, Zhong Z, Pirofski L. A human IgM monoclonal antibody prolongs survival of mice with lethal cryptococcosis. J Infect Dis. 1998;178:1213–1216. doi: 10.1086/515688. [DOI] [PubMed] [Google Scholar]
- Gigliotti F, Garvy BA, Harmsen AG. Antibody-mediated shift in the profile of glycoprotein A phenotypes observed in a mouse model of Pneumocystis carinii pneumonia. Infect Immun. 1998;64:1892–1899. doi: 10.1128/iai.64.6.1892-1899.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gigliotti F, Hughes WT. Passive immunoprophylaxis with specific monoclonal antibodies confers partial protection against Pneumocystis carinii pneumonitis in animal models. J Clin Invest. 1988;81:1666–1668. doi: 10.1172/JCI113503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldman DL, Davis J, Bommartio F, Shao X, Casadevall A. Enhanced allergic inflammation and airway responsiveness in rats with chronic Cryptococcus neoformans infection suggests a potential role for fungal pulmonary infection in the pathogenesis of asthma. J Infect Dis. 2006;193:1178–1186. doi: 10.1086/501363. [DOI] [PubMed] [Google Scholar]
- Goldman DL, Khine H, Abadi J, Lindenberg DJ, Pirofski L, Niang R, Casadevall A. Serologic evidence for Cryptococcus infection in early childhood. Pediatrics. 2001;107:E66. doi: 10.1542/peds.107.5.e66. [DOI] [PubMed] [Google Scholar]
- Griffin DO, Holodick NE, Rothstein TL. Human B1 cells in umbilical cord and adult peripheral blood express the novel phenotype CD20+ CD27+ CD43+ CD70−. J Exp Med. 2011;208:67–80. doi: 10.1084/jem.20101499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guimaraes AJ, Frases S, Gomez FJ, Zancope-Oliveira RM, Nosanchuk JD. Monoclonal antibodies to heat shock protein 60 alter the pathogenesis of Histoplasma capsulatum. Infect Immun. 2009a;77:1357–1367. doi: 10.1128/IAI.01443-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guimaraes AJ, Frases S, Gomez FJ, Zancope-Oliveira RM, Nosanchuk JD. Monoclonal antibodies to heat shock protein 60 alter the pathogenesis of Histoplasma capsulatum. Infect Immun. 2009b;77:1357–1367. doi: 10.1128/IAI.01443-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gupta S, Ellis M, Cesario T, Ruhling M, Vayuvegula B. Disseminated cryptococcal infection in a patient with hypogammaglobulinemia and normal T-cell function. Am J Med. 1987;82:129–131. doi: 10.1016/0002-9343(87)90388-3. [DOI] [PubMed] [Google Scholar]
- Han Y, Kanbe T, Cherniak R, Cutler JE. Biochemical characterization of Candida albicans epitopes that can elicit protective and nonprotective antibodies. Infect Immun. 1997;65:4100–4107. doi: 10.1128/iai.65.10.4100-4107.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Han Y, Kozel TR, Zhang MX, MacGill RS, Carroll MC, Cutler JE. Complement is essential for protection by an IgM and an IgG3 monoclonal antibody against experimental, hematogenously disseminated candidiasis. J Immunol. 2001;167:1550–1557. doi: 10.4049/jimmunol.167.3.1550. [DOI] [PubMed] [Google Scholar]
- Han Y, Ulrich MA, Cutler JE. Candida albicans mannan extract-protein conjugates induce a protective immune response against experimental candidiasis. J Infect Dis. 1999;179:1477–1484. doi: 10.1086/314779. [DOI] [PubMed] [Google Scholar]
- Jalali Z, Ng L, Singh N, Pirofski LA. Antibody response to Cryptococcus neoformans capsular polysaccharide glucuronoxylomannan in patients after solid-organ transplantation. Clin Vaccine Immunol. 2006;13:740–746. doi: 10.1128/CVI.00139-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kashino SS, Fazioli RA, Cafalli-Favati C, Meloni-Bruneri LH, Vaz CA, Burger E, Singer LM, Calich VL. Resistance to Paracoccidioides brasiliensis infection is linked to a preferential Th1 immune response, whereas susceptibility is associated with absence of IFN-gamma production. J Interferon Cytokine Res. 2000;20:89–97. doi: 10.1089/107999000312766. [DOI] [PubMed] [Google Scholar]
- Kling HM, Shipley TW, Patil SP, Kristoff J, Bryan M, Montelaro RC, Morris A, Norris KA. Relationship of Pneumocystis jiroveci humoral immunity to prevention of colonization and chronic obstructive pulmonary disease in a primate model of HIV infection. Infect Immun. 2010;78:4320–4330. doi: 10.1128/IAI.00507-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lane HC, Masur H, Edgar L, Whalen G, Rook AH, Fauci AS. Abnormalities of B-cell activation and immunoregulation in patients with the acquired immunodeficiency syndrome. N Engl J Med. 1983;309:453–458. doi: 10.1056/NEJM198308253090803. [DOI] [PubMed] [Google Scholar]
- Larsen RA, Pappas PG, Perfect JR, Aberg JA, Casadevall A, Cloud GA, James R, Filler S, Dismukes WE. A phase I evaluation of the safety and pharmacodynamic activity of a murine-derived monoclonal antibody 18B7 in subjects with treated cryptococcal meningitis. Antimicrob Agents Chemother. 2005;49:952–958. doi: 10.1128/AAC.49.3.952-958.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lopes LC, Guimaraes AJ, de Cerqueira MD, Gomez BL, Nosanchuk JD. A histoplasma capsulatum-specific IgG1 isotype monoclonal antibody, H1C, to a 70-kilodalton cell surface protein is not protective in murine histoplasmosis. Clin Vaccine Immunol. 2010a;17:1155–1158. doi: 10.1128/CVI.00033-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lopes LC, Rollin-Pinheiro R, Guimaraes AJ, Bittencourt VC, Martinez LR, Koba W, Farias SE, Nosanchuk JD, Barreto-Bergter E. Monoclonal antibodies against peptidorhamnomannans of Scedosporium apiospermum enhance the pathogenicity of the fungus. PLoS Negl Trop Dis. 2010b;4:e853. doi: 10.1371/journal.pntd.0000853. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lovchik JA, Wilder JA, Huffnagle GB, Riblet R, Lyons CR, Lipscomb MF. Ig heavy chain complex-linked genes influence the immune response in a murine cryptococcal infection. J Immunol. 1999;163:3907–3913. [PubMed] [Google Scholar]
- Lund FE, Hollifield M, Schuer K, Lines JL, Randall TD, Garvy BA. B cells are required for generation of protective effector and memory CD4 cells in response to Pneumocystis lung infection. J Immunol. 2006;176:6147–6154. doi: 10.4049/jimmunol.176.10.6147. [DOI] [PubMed] [Google Scholar]
- Magliani W, Conti S, Frazzi R, Ravanetti L, Maffei DL, Polonelli L. Protective antifungal yeast killer toxin-like antibodies. Curr Mol Med. 2005;5:443–452. doi: 10.2174/1566524054022558. [DOI] [PubMed] [Google Scholar]
- Marquis G, Montplaisir S, Pelletier M, Mousseau S, Auger P. Genetic resistance to murine cryptococcosis: increased susceptibility in the CBA/N XID mutant strain of mice. Infect Immun. 1985;47:282–287. doi: 10.1128/iai.47.1.282-287.1985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marr KA, Datta K, Pirofski LA, Barnes R. Cryptococcus gattii Infection in Healthy Hosts: A Sentinel for Subclinical Immunodeficiency? Clin Infect Dis. 2012;54:153–154. doi: 10.1093/cid/cir756. [DOI] [PubMed] [Google Scholar]
- Martinez LR, Casadevall A. Specific antibody can prevent fungal biofilm formation and this effect correlates with protective efficacy. Infect Immun. 2005;73:6350–6362. doi: 10.1128/IAI.73.10.6350-6362.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martinez LR, Mihu MR, Gacser A, Santambrogio L, Nosanchuk JD. Methamphetamine enhances histoplasmosis by immunosuppression of the host. J Infect Dis. 2009;200:131–141. doi: 10.1086/599328. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martinez LR, Moussai D, Casadevall A. Antibody to Cryptococcus neoformans glucuronoxylomannan inhibits the release of capsular antigen. Infect Immun. 2004;72:3674–3679. doi: 10.1128/IAI.72.6.3674-3679.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matthews RC, Burnie JP, Howat D, Rowland T, Walton F. Autoantibody to heat shock protein 90 can mediate protection against systemic candidosis. Immunology. 1991;74:20–24. [PMC free article] [PubMed] [Google Scholar]
- Matthews RC, Rigg G, Hodgetts S, Carter T, Chapman C, Gregory C, Illidge C, Burnie J. Preclinical assessment of the efficacy of mycograb, a human recombinant antibody against fungal HSP90. Antimicrob Agents Chemother. 2003;47:2208–2216. doi: 10.1128/AAC.47.7.2208-2216.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McClelland EE, Nicola AM, Prados-Rosales R, Casadevall A. Ab binding alters gene expression in Cryptococcus neoformans and directly modulates fungal metabolism. J Clin Invest. 2010;120:1355–1361. doi: 10.1172/JCI38322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McFadden DC, Fries BC, Wang F, Casadevall A. Capsule structural heterogeneity and antigenic variation in Cryptococcus neoformans. Eukaryot Cell. 2007;6:1464–1473. doi: 10.1128/EC.00162-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miller MF, Mitchell TG, Storkus WJ, Dawson JR. Human natural killer cells do not inhibit growth of Cryptococcus neoformans in the absence of antibody. Infect Immun. 1990;58:639–645. doi: 10.1128/iai.58.3.639-645.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moir S, Fauci AS. B cells in HIV infection and disease. Nat Rev Immunol. 2009;9:235–245. doi: 10.1038/nri2524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Montagnoli C, Bozza S, Bacci A, Gaziano R, Mosci P, Morschhauser J, Pitzurra L, Kopf M, Cutler J, Romani L. A role for antibodies in the generation of memory antifungal immunity. Eur J Immunol. 2003;33:1193–1204. doi: 10.1002/eji.200323790. [DOI] [PubMed] [Google Scholar]
- Moragues MD, Omaetxebarria MJ, Elguezabal N, Sevilla MJ, Conti S, Polonelli L, Ponton J. A monoclonal antibody directed against a Candida albicans cell wall mannoprotein exerts three anti-C. albicans activities. Infect Immun. 2003;71:5273–5279. doi: 10.1128/IAI.71.9.5273-5279.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mukherjee J, Scharff MD, Casadevall A. Protective murine monoclonal antibodies to Cryptococcus neoformans. Infect Immun. 1992;60:4534–4541. doi: 10.1128/iai.60.11.4534-4541.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nabavi N, Murphy JW. Antibody-dependent natural killer cell-mediated growth inhibition of Cryptococcus neoformans. Infect Immun. 1986;51:556–562. doi: 10.1128/iai.51.2.556-562.1986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Neto RJ, Guimaraes MC, Moya MJ, Oliveira FR, Louzada PL, Jr, Martinez R. Hypogammaglobulinemia as risk factor for Cryptococcus neoformans infection: report of 2 cases. Rev Soc Bras Med Trop. 2000;33:603–608. [PubMed] [Google Scholar]
- Nosanchuk JD, Steenbergen JN, Shi L, Deepe GS, Jr, Casadevall A. Antibodies to a cell surface histone-like protein protect against Histoplasma capsulatum. J Clin Invest. 2003;112:1164–1175. doi: 10.1172/JCI19361. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nussbaum G, Yuan R, Casadevall A, Scharff MD. Immunoglobulin G3 blocking antibodies to Cryptococcus neoformans. J Exp Med. 1996;183:1905–1909. doi: 10.1084/jem.183.4.1905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Okagaki LH, Strain AK, Nielsen JN, Charlier C, Baltes NJ, Chretien F, Heitman J, Dromer F, Nielsen K. Cryptococcal cell morphology affects host cell interactions and pathogenicity. PLoS Pathog. 2010;6:e1000953. doi: 10.1371/journal.ppat.1000953. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pachi J, Svoboda P, Jacobs F, van der Hoven B, Spronk P, Masterson G, Malbrain M, Aoun M, Garbino J, Takala J, Drgona L, Burnie J, Matthews R. A randomized, blinded, multicenter trial in patients with invasive candidiasis comparing lipid-associated amphotericin B alone and in combination with Mycograb, an antibody based inhibitor of Hsp90. Clin Infect Dis. 2006 doi: 10.1086/503428. (in press) [DOI] [PubMed] [Google Scholar]
- Pirofski L. Polysaccharides, mimotopes and vaccines for fungal and encapsulated pathogens. Trends Microbiol. 2001;9:445–451. doi: 10.1016/s0966-842x(01)02134-5. [DOI] [PubMed] [Google Scholar]
- Pirofski L, Lui R, DeShaw M, Kressel AB, Zhong Z. Analysis of human monoclonal antibodies elicited by vaccination with a Cryptococcus neoformans glucuronoxylomannan capsular polysaccharide vaccine. Infect Immun. 1995;63:3005–3014. doi: 10.1128/iai.63.8.3005-3014.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Popi AF, Godoy LC, Xander P, Lopes JD, Mariano M. B-1 cells facilitate Paracoccidioides brasiliensis infection in mice via IL-10 secretion. Microbes Infect. 2008;10:817–824. doi: 10.1016/j.micinf.2008.04.012. [DOI] [PubMed] [Google Scholar]
- Rapaka RR, Ricks DM, Alcorn JF, Chen K, Khader SA, Zheng M, Plevy S, Bengten E, Kolls JK. Conserved natural IgM antibodies mediate innate and adaptive immunity against the opportunistic fungus Pneumocystis murina. J Exp Med. 2010;207:2907–2919. doi: 10.1084/jem.20100034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rivera J, Casadevall A. Mouse genetic background is a major determinant of isotype-related differences for antibody-mediated protective efficacy against Cryptococcus neoformans. J Immunol. 2005;174:8017–8026. doi: 10.4049/jimmunol.174.12.8017. [DOI] [PubMed] [Google Scholar]
- Rivera J, Zaragoza O, Casadevall A. Antibody-mediated protection against Cryptococcus neoformans is dependent on B cells. Infect Immun. 2005;73:1141–1150. doi: 10.1128/IAI.73.2.1141-1150.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rodrigues ML, Travassos LR, Miranda KR, Franzen AJ, Rozental S, De Souza W, Alviano CS, Barreto-Bergter E. Human antibodies against a purified glucosylceramide from Cryptococcus neoformans inhibit cell budding and fungal growth. Infect Immun. 2000;68:7049–7060. doi: 10.1128/iai.68.12.7049-7060.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosas AL, Nosanchuk JD, Casadevall A. Passive immunization with melanin-binding monoclonal antibodies prolongs survival in mice with lethal Cryptococcus neoformans infection. Infect Immun. 2001;69:3410–3412. doi: 10.1128/IAI.69.5.3410-3412.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saha DC, Goldman DL, Shao X, Casadevall A, Husain S, Limaye AP, Lyon M, Somani J, Pursell K, Pruett TL, Singh N. Serologic evidence for reactivation of cryptococcosis in solid-organ transplant recipients. Clin Vaccine Immunol. 2007;14:1550–1554. doi: 10.1128/CVI.00242-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sandini S, La VR, Deaglio S, Malavasi F, Cassone A, De BF. A highly immunogenic recombinant and truncated protein of the secreted aspartic proteases family (rSap2t) of Candida albicans as a mucosal anticandidal vaccine. FEMS Immunol Med Microbiol. 2011;62:215–224. doi: 10.1111/j.1574-695X.2011.00802.x. [DOI] [PubMed] [Google Scholar]
- Sanford JE, Lupan DM, Schlagetter AM, Kozel TR. Passive immunization against Cryptococcus neoformans with an isotype-switch family of monoclonal antibodies reactive with cryptococcal polysaccharide. Infect Immun. 1990;58:1919–1923. doi: 10.1128/iai.58.6.1919-1923.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saville SP, Lazzell AL, Chaturvedi AK, Monteagudo C, Lopez-Ribot JL. Use of a genetically engineered strain to evaluate the pathogenic potential of yeast cell and filamentous forms during Candida albicans systemic infection in immunodeficient mice. Infect Immun. 2008;76:97–102. doi: 10.1128/IAI.00982-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schlagetter AM, Kozel TR. Opsonization of Cryptococcus neoformans by a family of isotype-switch variant antibodies specific for the capsular polysaccharide. Infect Immun. 1990;58:1914–1918. doi: 10.1128/iai.58.6.1914-1918.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shapiro S, Beenhouwer DO, Feldmesser M, Taborda C, Carroll MC, Casadevall A, Scharff MD. Immunoglobulin G monoclonal antibodies to Cryptococcus neoformans protect mice deficient in complement component C3. Infect Immun. 2002;70:2598–2604. doi: 10.1128/IAI.70.5.2598-2604.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shi L, Albuquerque PC, Lazar-Molnar E, Wang X, Santambrogio L, Gacser A, Nosanchuk JD. A monoclonal antibody to Histoplasma capsulatum alters the intracellular fate of the fungus in murine macrophages. Eukaryot Cell. 2008;7:1109–1117. doi: 10.1128/EC.00036-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sinha BK, Prasad S, Monga DP. Studies of the role of B-cells in the resistance of mice to experimental candidiasis. Zentralbl Bakteriol Mikrobiol Hyg A. 1987;266:316–322. doi: 10.1016/s0176-6724(87)80045-7. [DOI] [PubMed] [Google Scholar]
- Smulian AG, Sullivan DW, Theus SA. Immunization with recombinant Pneumocystis carinii p55 antigen provides partial protection against infection: characterization of epitope recognition associated with immunization. Microbes Infect. 2000;2:127–136. doi: 10.1016/s1286-4579(00)00275-6. [DOI] [PubMed] [Google Scholar]
- Subramaniam K, Metzger B, Hanau LH, Guh A, Rucker L, Badri S, Pirofski LA. IgM(+) memory B cell expression predicts HIV-associated cryptococcosis status. J Infect Dis. 2009;200:244–251. doi: 10.1086/599318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Subramaniam KS, Datta K, Marks MS, Pirofski LA. Improved survival of mice deficient in secretory immunoglobulin M following systemic infection with Cryptococcus neoformans. Infect Immun. 2010a;78:441–452. doi: 10.1128/IAI.00506-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Subramaniam KS, Datta K, Quintero E, Manix C, Marks MS, Pirofski LA. The absence of serum IgM enhances the susceptibility of mice to pulmonary challenge with Cryptococcus neoformans. J Immunol. 2010b;184:5755–5767. doi: 10.4049/jimmunol.0901638. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taborda CP, Rivera J, Zaragoza O, Casadevall A. More is not necessarily better: ‘Prozone-like’ effects in passive immunization with Immunoglobulin G. J Immunol. 2003;140:3621–3630. doi: 10.4049/jimmunol.170.7.3621. [DOI] [PubMed] [Google Scholar]
- Torosantucci A, Bromuro C, Chiani P, De Bernardis F, Berti F, Galli C, Norelli F, Bellucci C, Polonelli L, Costantino P, Rappuoli R, Cassone A. A novel glyco-conjugate vaccine against fungal pathogens. J Exp Med. 2005;202:597–606. doi: 10.1084/jem.20050749. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Torosantucci A, Chiani P, Bromuro C, De BF, Palma AS, Liu Y, Mignogna G, Maras B, Colone M, Stringaro A, Zamboni S, Feizi T, Cassone A. Protection by anti-beta-glucan antibodies is associated with restricted beta-1,3 glucan binding specificity and inhibition of fungal growth and adherence. PLoS ONE. 2009;4:e5392. doi: 10.1371/journal.pone.0005392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Spriel AB, Leusen JH, van EM, Dijkman HB, Assmann KJ, Mayadas TN, van de Winkel JG. Mac-1 (CD11b/CD18) is essential for Fc receptor-mediated neutrophil cytotoxicity and immunologic synapse formation. Blood. 2001;97:2478–2486. doi: 10.1182/blood.v97.8.2478. [DOI] [PubMed] [Google Scholar]
- van Spriel AB, Sofi M, Gartlan KH, van der Schaaf A, Verschueren I, Torensma R, Raymakers RA, Loveland BE, Netea MG, Adema GJ, Wright MD, Figdor CG. The tetraspanin protein CD37 regulates IgA responses and anti-fungal immunity. PLoS Pathog. 2009;5:e1000338. doi: 10.1371/journal.ppat.1000338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Viudes A, Lazzell A, Perea S, Kirkpatrick WR, Peman J, Patterson TF, Martinez JP, Lopez-Ribot JL. The C-terminal antibody binding domain of Candida albicans mp58 represents a protective epitope during candidiasis. FEMS Microbiol Lett. 2004;232:133–138. doi: 10.1016/S0378-1097(04)00042-4. [DOI] [PubMed] [Google Scholar]
- Wahab JA, Hanifah MJ, Choo KE. Bruton’s agammaglobulinaemia in a child presenting with cryptococcal empyema thoracis and periauricular pyogenic abscess. Singapore Med J. 1995;36:686–689. [PubMed] [Google Scholar]
- Winkelstein JA, Marino MC, Ochs H, Fuleihan R, Scholl PR, Geha R, Stiehm ER, Conley ME. The X-linked hyper-IgM syndrome: clinical and immunologic features of 79 patients. Medicine (Baltimore) 2003;82:373–384. doi: 10.1097/01.md.0000100046.06009.b0. [DOI] [PubMed] [Google Scholar]
- Xin H, Cutler JE. Vaccine and monoclonal antibody that enhance mouse resistance to candidiasis. Clin Vaccine Immunol. 2011;18:1656–1667. doi: 10.1128/CVI.05215-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xin H, Dziadek S, Bundle DR, Cutler JE. Synthetic glycopeptide vaccines combining beta-mannan and peptide epitopes induce protection against candidiasis. Proc Natl Acad Sci U S A. 2008;105:13526–13531. doi: 10.1073/pnas.0803195105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yuan R, Casadevall A, Oh J, Scharff MD. T cells cooperate with passive antibody to modify Cryptococcus neoformans infection in mice. Proc Natl Acad Sci. 1997;94:2483–2488. doi: 10.1073/pnas.94.6.2483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yuan R, Spira G, Oh J, Paizi M, Casadevall A, Scharff MD. Isotype switching increases antibody protective efficacy to Cryptococcus neoformans infection in mice. Infect Immun. 1998;66:1057–1062. doi: 10.1128/iai.66.3.1057-1062.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zaragoza O, Chrisman CJ, Castelli MV, Frases S, Cuenca-Estrella M, Rodriguez-Tudela JL, Casadevall A. Capsule enlargement in Cryptococcus neoformans confers resistance to oxidative stress suggesting a mechanism for intracellular survival. Cell Microbiol. 2008;10:2043–2057. doi: 10.1111/j.1462-5822.2008.01186.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zaragoza O, Garcia-Rodas R, Nosanchuk JD, Cuenca-Estrella M, Rodriguez-Tudela JL, Casadevall A. Fungal cell gigantism during mammalian infection. PLoS Pathog. 2010;6:e1000945. doi: 10.1371/journal.ppat.1000945. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zaragoza O, Taborda CP, Casadevall A. The efficacy of complement-mediated phagocytosis of Cryptococcus neoformans is dependent on the location of C3 in the polysaccharide capsule and involves both direct and indirect C3-mediated interactions. Eur J Immunol. 2003;33:1957–1967. doi: 10.1002/eji.200323848. [DOI] [PubMed] [Google Scholar]