

Abstract
Aims/hypothesis
We have previously documented a high heritability of insulin clearance in a Hispanic cohort. Here, our goal was to confirm the high heritability in a second cohort and search for genetic loci contributing to insulin clearance.
Methods
Hyperinsulinaemic–euglycaemic clamps were performed in 513 participants from 140 Hispanic families. Heritability was estimated for clamp-derived insulin clearance and a two-phase genome-wide linkage scan was conducted using a variance components approach. Linkage peaks were further investigated by candidate gene association analysis in two cohorts.
Results
The covariate-adjusted heritability of insulin clearance was 73%, indicating that the majority of the phenotypic variance is due to genetic factors. In the Phase 1 linkage scan, no signals with a logarithm of odds (LOD) score >2 were detected. In the Phase 2 scan, two linkage peaks with an LOD >2 for insulin clearance were identified on chromosomes 15 (LOD 3.62) and 20 (LOD 2.43). These loci harbour several promising candidate genes for insulin clearance, with 12 single nucleotide polymorphisms (SNPs) on chromosome 15 and six SNPs on chromosome 20 being associated with insulin clearance in both Hispanic cohorts.
Conclusions/interpretation
In a second Hispanic cohort, we confirmed that insulin clearance is a highly heritable trait and identified chromosomal loci that harbour genes regulating insulin clearance. The identification of such genes may improve our understanding of how the body clears insulin, thus leading to improved risk assessment, diagnosis, prevention and therapy of diabetes, as well as of other hyper-insulinaemic disorders, such as the metabolic syndrome and polycystic ovary syndrome.
Keywords: Candidate gene, Heritability, Insulin clearance, Linkage, Type 2 diabetes mellitus
Introduction
The incidence of type 2 diabetes mellitus has reached epidemic proportions and continues to grow; it has been estimated that one in three Americans (one in two in some minority groups) born in 2000 will develop diabetes in their lifetime [1]. The prevalence of diagnosed diabetes is twice as high in non-Hispanic blacks and Mexican–Americans compared with non-Hispanic whites [2, 3]. Therefore, an improved understanding of the pathophysiology of diabetes is imperative in minority populations.
The investigation of the pathophysiology of type 2 diabetes has focused on insulin resistance and defective insulin secretion, both of which must be present for type 2 diabetes to develop [4]. Intact insulin secretion maintains normoglycaemia in the presence of insulin resistance via compensatory hyperinsulinaemia. Consistent with this notion, most type 2 diabetes susceptibility genes appear to affect insulin secretion [5]. To avoid type 2 diabetes, insulin levels must increase to overcome tissue unresponsiveness. The resultant hyperinsulinaemia, while maintaining normal glucose levels, contributes to cardiometabolic risk [6].
A much overlooked factor in studies of the pathophysiology of diabetes is the role of insulin clearance, i.e. removal of insulin from the circulation. A few physiological studies have demonstrated that insulin resistance elicits a reduction in insulin clearance, suggesting that altering insulin clearance is an additional means of achieving compensatory hyperinsulinaemia [7–10]. We were the first to report that whole-body insulin clearance, as measured by hyperinsulinaemic–euglycaemic clamps performed in Mexican–Americans, had a heritability (h2) of 0.58, which was greater than other insulin-related traits examined (fasting insulin h2 0.38, insulin resistance h2 0.44) [11]. The high heritability of insulin clearance raises the possibility that inherited variability in insulin clearance may affect an individual’s risk of developing diabetes by influencing the ability to achieve compensatory hyperinsulinaemia in response to insulin resistance. Whether this is the case is at present unknown because insulin clearance has not been measured in any large candidate gene or genome-wide association studies (GWAS) probing type 2 diabetes susceptibility.
In the current study, our goal was to confirm the high heritability of insulin clearance in an independent cohort of Mexican–Americans and to conduct the first genome-wide linkage scan to search for regions possibly harbouring genes for insulin clearance. Linkage peaks were further investigated by association studies to search for loci involved in insulin clearance.
Methods
Participants and phenotyping
The participants in the heritability, linkage and association studies were drawn from the Hypertension–Insulin Resistance (HTN–IR) cohort, which consists of Los Angeles Hispanic–American families ascertained via a proband with essential hypertension. The recruitment and phenotyping of this cohort have been described previously [12]. The linkage study in that cohort used prior microsatellite genotyping conducted on 754 participants from 145 families. Subsequently, additional participants were recruited (bringing the total to 922 participants in 163 families), all of whom were genotyped in the present genetic association study. Approximately 75% of the HTN–IR participants had undergone the euglycaemic clamp with steady-state plasma insulin (SSPI) levels and were therefore ultimately analysed in this study. Flowcharts in electronic supplementary material [ESM] Fig. 1 provide details on the number of participants used in the current linkage and association studies.
Fig. 1.

Plots of the maximum linkage signal for insulin clearance on chromosome 15 (a) and chromosome 20 (b). Microsatellite markers are listed at top. Solid line, linkage for the entire cohort; dashed line, linkage for Phase 1; dotted line, linkage for Phase 2
Participants from the Mexican–American Coronary Artery Disease (MACAD) study [11] were used in the association analyses conducted in conjunction with the HTN–IR cohort. In the present report, after data cleaning as described below, 179 families were included, comprising 676 participants from the offspring generation (adult offspring of probands with coronary artery disease and the spouses of those offspring) who underwent phenotyping for insulin clearance.
By design, participants undergoing detailed phenotyping in both cohorts were free of known diabetes and clinically manifest cardiovascular disease, thus avoiding secondary changes in phenotype caused by overt disease. A small percentage of participants in both cohorts were newly diagnosed with diabetes as a result of their participation in the studies. These participants were not taking any glucose-lowering medications when they were phenotyped.
Participants in the HTN–IR and MACAD cohorts underwent the hyperinsulinaemic–euglycaemic clamp procedure, performed with the same techniques and assays (described in the ESM Methods). The metabolic clearance rate of insulin (ml m−2 min−1) is the insulin infusion rate divided by the final SSPI level (average of insulin levels at 100, 110 and 120 min) of the euglycaemic clamp. Because all of our participants received the same insulin infusion rate (60 mU m−2 min−1), the SSPI is a direct measure of insulin clearance at steady state and was the trait used in genetic analysis in this study. Insulin sensitivity was quantified using the marker of insulin sensitivity (M value) from the euglycaemic clamp, as previously described [11]. Insulin secretion was estimated using the HOMA of beta cell function (HOMA-%B) using fasting glucose and insulin levels; HOMA of insulin resistance (HOMA-IR) was also calculated [13].
All studies were approved by the Institutional Review Boards at the participating institutions. All participants gave informed consent before participation.
Microsatellite marker genotyping
A 10 cM genome scan using microsatellite markers was conducted on 754 participants (570 with SSPI data) from 145 families of the HTN–IR cohort in two separate batches. According to the original study design, a Phase 1 linkage scan was conducted, followed by recruitment of additional participants who comprised a Phase 2 linkage scan. Phase 1 consisted of 71 families with 354 individuals; Phase 2 comprised 74 families with 400 individuals (includes non-phenotyped parents and phenotyped offspring). For the Phase 1 sample, 378 fluorescently tagged microsatellite markers (Marshfield Version 9; http://research.marshfieldclinic.org/genetics) were successfully genotyped at the University of California at Los Angeles (UCLA); for the Phase 2 sample, 377 micro-satellite markers (Screening Set 13, see http://research.marshfieldclinic.org/genetics and Ghebranious et al. [14]) were successfully genotyped at PreventionGenetics (Marshfield, WI, USA). The number of overlapping markers between Phase 1 and Phase 2 was 246; we therefore report the individual as well as the combined linkage analyses.
After data cleaning, described in detail in the ESM Methods, 513 individuals from 140 families (227 individuals from 67 families in Phase 1, 286 individuals from 73 families in Phase 2) had SSPI data and were used in the heritability and linkage analyses (ESM Fig. 1).
Linkage analysis
Heritability estimates and a genome-wide linkage scan were conducted using a variance components approach implemented in SOLAR (Sequential Oligogenic Linkage Analysis Routines; available from http://solar.txbiomedgenetics.org/download.html) [15], in which the total variation in a phenotype ( ) is partitioned into components of variance due to a major gene ( ), additive polygenes ( ) and individual specific environment ( ):
| (1) |
Heritability (h2) was estimated by the proportion of total variation in the phenotype due to genetic effects. Similarly, the expected genetic covariances between arbitrary relatives i and j at a given marker locus can be specified as a function of identity by descent (IBD) and relationships quantified by the kinship coefficient:
| (2) |
where IBDij is the predicted probability that individuals i and j share alleles that are identical by IBD at the major locus, φij is the corresponding kinship coefficient and δij = 1 if i = j and 0 otherwise.
Linkage was tested by evaluating the null hypothesis that the additive genetic variance due to a quantitative trait locus (QTL) equals zero. The linkage analyses were adjusted for age, sex, BMI and diabetes status. To calculate empirical p values, we simulated a fully informative marker and applied gene dropping for offspring using the LODADJ option in SOLAR. The data were simulated 10,000 times and subsequent linkage analyses were performed to obtain the distribution of the test statistic under the null hypothesis of no linkage. The empirical p values were obtained as the proportion of the 10,000 replicates that had a logarithm of odds (LOD) score greater than or equal to the nominal LOD score observed for the original linked locus. p values were converted to LOD scores by: LOD = χ2/[2* ln(10)].
Positional candidate single nucleotide polymorphisms
We sought candidate loci under the −1 LOD interval of any linkage peaks with an LOD score greater than 2. We took advantage of the fact that the HTN–IR and MACAD cohorts have recently been genotyped on the Cardio-Metabochip as part of a separate project (unpublished data, M.O. Goodarzi, J.I. Rotter). The Cardio-Metabochip was designed to provide high-throughput genotyping for replication and fine-mapping of GWAS results for cardiac (e.g. myocardial infarction, QT interval), metabolic (e.g. diabetes, fasting glucose, fasting insulin), anthropometric (e.g. BMI, WHR) and lipid traits (detailed description of Cardio-Metabochip design, see the ESM Methods). Besides providing a relatively dense set of markers under the −1 LOD intervals, this chip provides single nucleotide polymorphism (SNPs) of particular interest to the study of insulin clearance, given the close relationship between insulin levels and most of the traits represented on the chip. Only the SNPs under the −1 LOD intervals of the two linkage peaks with an LOD >2 were examined here. Details of Cardio-Metabochip genotyping, quality control and population stratification analysis in the HTN–IR and MACAD studies are given in the ESM Methods. After quality control (ESM Fig. 1), 638 HTN–IR and 676 MACAD genotyped participants with SSPI were available for association analysis (clinical characteristics, see ESM Table 1).
Table 1.
Characteristics of the participants in the heritability and linkage studies
| Trait | All | Phase 1 | Phase 2 | p value |
|---|---|---|---|---|
| Families (n) | 140 | 67 | 73 | |
| Participants (n) | 513 | 227 | 286 | |
| Male sex (%) | 41.9 | 41.4 | 42.4 | 0.85 |
| Age (years) | 32.7±10.9 | 29.7±8.7 | 35.1±11.9 | <0.0001 |
| BMI (kg/m2) | 28.5±5.9 | 28.2±5.9 | 28.8±6.0 | 0.27 |
| Diabetes (%) | 7.7 | 6.8 | 8.9 | 0.36 |
| Fasting insulin (pmol/l) | 90.6±59.4 | 94.8±57.1 | 87.0±61.2 | 0.16 |
| Fasting glucose (mmol/l) | 5.31±0.46 | 5.33±0.47 | 5.30±0.44 | 0.41 |
| HOMA-IR | 3.9±3.4 | 4.2±3.6 | 3.6±3.3 | 0.03 |
| HOMA-%B | 184.6±134.2 | 212.1±150.4 | 160.8±113.4 | <0.0001 |
| M value (μmol m−2 min−1) | 1,308.8±575.0 | 1,306.6±593.9 | 1,310.5±561.2 | 0.94 |
| SSPI (pmol/l) | 832.2±197.4 | 814.8±201.6 | 830.2±193.8 | 0.41 |
Unless otherwise stated, values are mean ± SD
Association analysis
Association between individual candidate SNPs and insulin clearance measures was evaluated using the general estimating equations (GEE1) method implemented in the Genome-Wide Association Analyses with Family Data (GWAF) program [16]. GWAF uses functions in existing R packages to properly model the residual correlations within families in the test of genotype–phenotype association. The additive genetic model was used in both cohorts. Potential confounding factors, including age, sex, BMI and diabetes status were included as covariates in all analyses. We sought loci associated with SSPI in HTN–IR and MACAD. The power of this two-cohort study to detect association is displayed in ESM Table 2. With the two cohorts studied, we had 81% power to detect SNPs that explain 1.6% of SSPI variance. Even for a SNP that explains as little as 1.2% of variance, we had a power of 65% to detect association.
Table 2.
Loci with evidence of association with SSPI in two Hispanic cohorts
| Gene locus
|
HTN–IR
|
MACAD
|
Meta-analysis p value | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Chr | Base P | SNPa | SNP in gene | Nearest gene(s)b | MAF | Beta | p value | MAF | Beta | p value | |
| 15 | 64118445 | rs12899971 | MEGF11 | 0.022 | −18.317 | 9.13×10−3 | 0.030 | −14.430 | 1.10×10−2 | 2.90×10−4 | |
| 15 | 72839988 | rs11072505 | CYP1A2 (4 kb), CSK (21.5 kb) | 0.295 | 4.892 | 2.53×10−2 | 0.270 | 5.115 | 1.68×10−2 | 1.07×10−3 | |
| 15 | 72850248 | rs60001770 | CYP1A2 (14.3 kb), CSK (11.2 kb) | 0.084 | 9.761 | 5.98×10−3 | 0.087 | 10.126 | 3.41×10−3 | 6.04×10−5 | |
| 15 | 72879437 | rs2301249 | CSK | 0.515 | −4.451 | 2.88×10−2 | 0.501 | −4.060 | 3.92×10−2 | 2.82×10−3 | |
| 15 | 72882536 | rs7085 | CSK | 0.518 | −4.367 | 3.15×10−2 | 0.501 | −4.060 | 3.92×10−2 | 3.04×10−3 | |
| 15 | 72894933 | rs7176022 | LMAN1L | 0.517 | −4.043 | 4.74×10−2 | 0.501 | −3.840 | 4.97×10−2 | 5.45×10−3 | |
| 15 | 76785585 | chr15:76785585 | CHRNB4 (45.7 kb), ADAMTS7 (53 kb) | 0.191 | 5.520 | 3.17×10−2 | 0.173 | 6.976 | 2.47×10−3 | 2.17×10−4 | |
| 15 | 76785802 | chr15:76785802 | CHRNB4 (45.9 kb), ADAMTS7 (52.8 kb) | 0.191 | 5.520 | 3.17×10−2 | 0.176 | 7.128 | 1.92×10−3 | 1.73×10−4 | |
| 15 | 76793880 | chr15:76793880 | CHRNB4 (54 kb), ADAMTS7 (44.7 kb) | 0.194 | 5.726 | 2.59×10−2 | 0.173 | 6.868 | 3.71×10−3 | 2.58×10−4 | |
| 15 | 76795005 | rs11638490 | CHRNB4 (55.1 kb), ADAMTS7 (43.6 kb) | 0.429 | 5.736 | 5.98×10−3 | 0.379 | 3.540 | 4.80×10−2 | 1.02×10−3 | |
| 15 | 76806665 | rs3813565 | CHRNB4 (66.7 kb), ADAMTS7 (31.9 kb) | 0.427 | 5.383 | 9.68×10−3 | 0.379 | 3.759 | 3.48×10−2 | 1.03×10−3 | |
| 15 | 76807207 | rs35583595 | CHRNB4 (67.3 kb), ADAMTS7 (31.4 kb) | 0.429 | 5.736 | 5.98×10−3 | 0.379 | 3.759 | 3.48×10−2 | 7.06×10−4 | |
| 20 | 39222875 | rs34887308 | PLCG1 | 0.053 | 9.688 | 3.77×10−2 | 0.040 | 10.124 | 4.92×10−2 | 4.46×10−3 | |
| 20 | 39225155 | rs34454281 | PLCG1 | 0.054 | 9.496 | 3.89×10−2 | 0.041 | 12.869 | 1.55×10−2 | 1.47×10−3 | |
| 20 | 39284214 | chr20:39284214 | ZHX3 | 0.053 | 9.688 | 3.77×10−2 | 0.038 | 11.245 | 3.24×10−2 | 2.93×10−3 | |
| 20 | 39323693 | chr20:39323693 | ZHX3 | 0.053 | 9.688 | 3.77×10−2 | 0.039 | 10.827 | 3.69×10−2 | 3.33×10−3 | |
| 20 | 39338518 | chr20:39338518 | ZHX3 | 0.053 | 9.670 | 3.83×10−2 | 0.039 | 10.827 | 3.69×10−2 | 3.38×10−3 | |
| 20 | 39338917 | chr20:39338917 | ZHX3 | 0.053 | 9.688 | 3.77×10−2 | 0.039 | 10.827 | 3.69×10−2 | 3.33×10−3 | |
SNPs that do not have rs numbers are indicated by genomic coordinates (NCBI Genome Build 36.3, www.ncbi.nlm.nih.gov/genome/guide/human/release_notes.html)
If the SNP is not located in a gene, the nearest upstream and the nearest downstream genes are listed, with distances from the SNP
Base P, base position; Chr, chromosome; MAF, minor allele frequency
For those SNPs associated with SSPI in HTN–IR and MACAD, the proportion of variance explained by each associated SNP was estimated using variance components analysis methods. For these SNPs, we also combined the results of HTN–IR and MACAD using fixed-effects, inverse-variance weighting meta-analysis as implemented in METAL [17].
Results
Characteristics of the heritability and linkage study cohort are given in Table 1. Phase 1 and Phase 2 samples were comparable for BMI, sex distribution, fasting glucose and insulin, as well as M value and SSPI. The Phase 2 sample was older on average and had lower HOMA values. Age-, sex-, BMI- and diabetes status-adjusted heritability values were calculated for the insulin-related traits available in this cohort. The heritability of fasting insulin (0.41, SD 2.33, SEM 0.11, p< 0.0001), HOMA of insulin resistance (HOMA-IR) (0.42, SD 1.96, SEM 0.09, p<0.0001) and HOMA-%B (0.38, SD 1.95, SEM 0.09, p<0.0001) was similar. The clamp-derived M value displayed a higher heritability of 0.63 (SD 2.19, SEM 0.10, p<0.0001), while the most heritable insulin-related trait was SSPI at 0.73 (SD 2.21, SEM 0.10, p<0.0001).
The most significant linkage peaks for SSPI were observed in the Phase 2 scan at chromosome 15 (75 cM, empirical LOD 3.62, p = 0.0001) and chromosome 20 (54 cM, empirical LOD 2.43, p = 0.0004), while no linkage signals with a LOD score greater than 2 were observed in the Phase 1 scan. When combining Phase 1 and Phase 2 samples, the empirical LOD score decreased to 2.53 for chromosome 15 (p = 0.0004), but increased to 2.71 for chromosome 20 (p = 0.0001). The maximum LOD scores for SSPI on chromosomes 15 and 20 are displayed in Fig. 1 for Phase 1, Phase 2 and the combined data. While the linkage signal on chromosome 15 was mainly from the Phase 2 sample (Fig. 1a), the linkage signal on chromosome 20 was more consistent between the Phase 1 and Phase 2 samples, although with a higher magnitude for the Phase 2 sample (Fig. 1b).
We further explored the linkage peaks by conducting association analyses of Cardio-Metabochip variants located under the −1 LOD interval of each peak. To identify the most robust loci, we sought only SNPs associated with SSPI in both (i.e. HTN–IR and MACAD) cohorts. A total of 2,759 SNPs was located under the −1 LOD interval of the chromosome 15 peak. Of these, 12 had a p value of p<0.05 for association with SSPI in both cohorts, with the same direction of effect (Table 2). Three discrete regions are delineated by these 12 SNPs (Fig. 2). The first SNP (rs12899971) resides in the MEGF11 gene. The next five SNPs form a 54 kb haplotype block near or inclusive of the CYP1A2, CSK and LMAN1L genes. The final six SNPs form a 21 kb haplotype block flanked by the CHRNB4 and ADAMTS7 genes.
Fig. 2.

Gene and linkage disequilibrium plot for the chromosome 15 SNPs associated with insulin clearance. Genes are indicated by arrows (arrow size not to scale). The locations of the genotyped SNPs relative to the genes are indicated. The linkage disequilibrium plot beneath the genes displays D′ values (%) for each pair of SNPs in the box at the intersection of the diagonals from each SNP. The bright red blocks indicate D′ = 1 (100%) for the corresponding pair of variants with an LOD score ≥2. The boxes with lighter shades of red indicate D′ <1, with an LOD score ≥2. The SNPs consist of a single SNP at MEGF11, plus two distinct blocks of SNPs. The region of the first block of SNPs is expanded to provide details. SNPs rs12899971 and rs2301249 are intronic; rs7085 resides in the 3′ untranslated region of CSK. SNPs that do not have rs numbers are indicated by genomic coordinates (NCBI Genome Build 36.3, www.ncbi.nlm.nih.gov/genome/guide/human/release_notes.html)
The association study of the chromosome 20 peak, also conducted using Cardio-Metabochip variants under the −1 LOD interval, encompassed 1,002 SNPs, six of which had a p value of p<0.05 for association with SSPI in both (i.e. the HTN–IR and MACAD) samples, with the same direction of effect (Table 2). These six SNPs, which are in linkage disequilibrium, span a 116 kb region across the terminal portions of the PLCG1 and ZHX3 genes (Fig. 3).
Fig. 3.
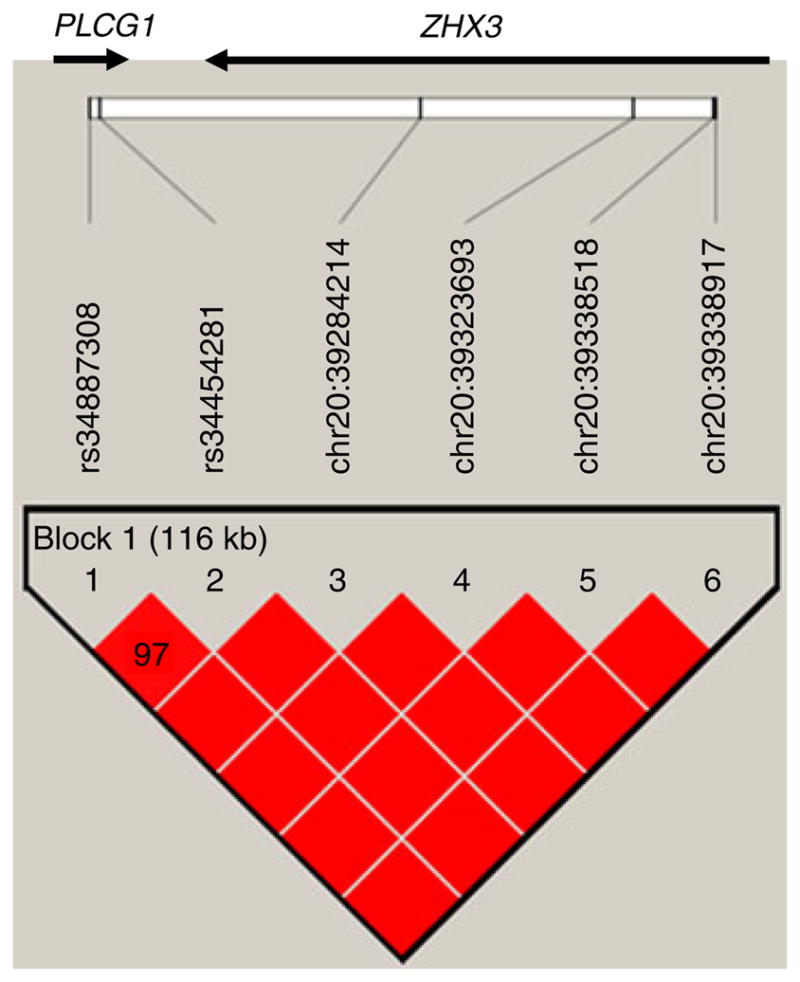
Gene and linkage disequilibrium plot for the chromosome 20 SNPs associated with insulin clearance. Genes are indicated by arrows (arrow size not to scale). The locations of the genotyped SNPs relative to the genes are indicated. The linkage disequilibrium plot beneath the genes displays D′ values (%) for each pair of SNPs in the box at the intersection of the diagonals from each SNP. The bright red blocks indicate D′ = 1 (100%) for the corresponding pair of variants with an LOD score of ≥2. All intragenic SNPs are intronic. SNPs that do not have rs numbers are indicated by genomic coordinates (NCBI Genome Build 36.3, www.ncbi.nlm.nih.gov/genome/guide/human/release_notes.html)
For the 18 SNPs identified above (12 on chromosome 15, six on chromosome 20) and found to be associated with SSPI in the HTN–IR and MACAD cohorts, we conducted a meta-analysis, the results of which are given in Table 2. Meta-analysis p values ranged from 5×10−3 to 6×10−5. We also estimated the percentage of variance explained by the SNP(s). The 18 SNPs individually accounted for 0.32% to 2.0% of the variance in SSPI in the HTN–IR and for 0.33% to 1.5% in the MACAD cohorts. Considered jointly, the 18 SNPs accounted for 5.5% and 6.1% of the variance in SSPI in HTN–IR and MACAD, respectively.
Discussion
This study confirmed the high heritability of insulin clearance in Mexican–Americans. We also identified, via linkage and association, regions of chromosomes 15 and 20 that may harbour quantitative trait loci that influence insulin clearance.
While many studies have implicated insulin resistance and defective insulin secretion in the pathophysiology of type 2 diabetes, a few studies suggest that reductions of insulin clearance may be an adaptive response to insulin resistance, aimed at maintaining normoglycaemia [7, 10]. Insulin resistance has been found to have an inverse correlation with insulin clearance [8, 9], while weight loss resulted in an increase in clearance [9]. Studies of rats [18] and dogs [19] with diet-induced obesity have also found that reduced insulin clearance accompanies insulin resistance. Treatment with the insulin-sensitising agent rosiglitazone resulted in reduced insulin secretion and increased clearance [20]. Thus, a reduction of insulin clearance accompanies increasing insulin secretion in response to insulin resistance. An improved understanding of this trait and its genetic determinants may therefore provide insight into why some insulin-resistant individuals develop diabetes.
In pursuit of this goal, we examined Mexican–Americans, an ethnic group with a high prevalence of the metabolic syndrome [21] and diabetes [2, 3]. Our logic was that study of a group with a high prevalence of abnormalities in the insulin system could increase the chances of discovering the responsible genetic variants. The high heritability of insulin clearance may be unique to Mexican–Americans and perhaps related to their high rates of diabetes. Studies in other ethnic groups will be needed to determine whether insulin clearance is similarly heritable in other populations.
We used SNPs from the Cardio-Metabochip in our association study of the −1 LOD intervals under the two linkage peaks. While not representing all SNPs contained in the intervals of interest, these SNPs are of particular relevance because they have been previously implicated in traits relevant to diabetes, obesity, coronary artery disease, dyslipidaemia and hypertension. Given that hyperinsulinaemia is a common feature of these conditions, Cardio-Metabochip SNPs are particularly pertinent to our study of insulin clearance, a major factor determining circulating insulin levels [22].
SNPs in or near the MEGF11, CYP1A2, CSK, LMAN1L, CHRNB4 and ADAMTSL7 genes located under the chromosome 15 linkage peak were associated with insulin clearance in two independent Hispanic cohorts. As commonly occurs in systematic association studies (e.g. GWAS), the identified genes do not have an immediately clear relevance to insulin clearance. These genes were localised to three regions (Fig. 2). The first region contains MEGF11, which has no known relationship to insulin but is expressed in the kidney [23], which clears insulin. The MEGF11 locus was not extensively covered on the Cardio-Metabochip; further genotyping will be necessary to elucidate its association with insulin clearance.
The second region, comprising CYP1A2, CSK and LMAN1L, was fine-mapped on the Cardio-Metabochip to investigate a strong GWAS signal for blood pressure (rs1378942) [24]. Insulin binding to its receptor triggers internalisation of the insulin–insulin receptor complex by endocytosis. Within the resulting endosome, insulin is cleaved by insulin-degrading enzyme, protein disulfide isomerase and other enzymes. c-Src tyrosine kinase (CSK) participates in actin cytoskeleton remodelling [25], which might affect insulin clearance via vesicular trafficking along the actin cytoskeleton. Insulin has been found to promote the association of CSK with phosphorylated caveolin-1, which is located at points of contact of the actin cytoskeleton with the plasma membrane [26]. Given that ligand-bound insulin receptors localise to caveolae [27], the interaction of CSK with caveolin might play a role in the internalisation of the insulin receptor–insulin complex. A GWAS meta-analysis associated a variant in CYP1A2 with caffeine consumption [28]. Cytochrome P450, family 1, subfamily A, polypeptide 2 (CYP1A2) metabolises xenobiotics such as caffeine; it is not known whether CYP1A2 participates in insulin metabolism. Thus, while further studies are needed, CSK appears the most likely of the three candidate genes mentioned above to be involved in insulin clearance.
The third region on chromosome 15, flanked by CHRNB4 and ADAMTS7, was fine-mapped on the Cardio-Metabochip in follow-up of a signal for coronary artery disease. A missense variant (rs3825807) in ADAMTS7 was associated with coronary artery disease [29], but none of the SNPs associated with SSPI was in linkage disequilibrium with this variant. ADAMTS7, which codes for a disintegrin/metalloproteinase, is expressed in tissues that clear insulin, including liver, kidney and skeletal muscle [30]; however, a role in insulin clearance has yet to be discovered. CHRNB4, which codes for a neuronal nicotinic receptor, has no known role in insulin metabolism, but appears to have a role in central regulation of food intake [31].
The −1 LOD interval of the chromosome 20 peak overlapped portions of the PLCG1 and ZHX3 genes. Interestingly, several previous studies have identified linkage with type 2 diabetes on chromosome 20q, with LOD scores >2 found at 30 to 70 cM [32], overlapping one of our insulin clearance linkage peaks. However, a subsequent meta-analysis of 23 type 2 diabetes linkage scans found evidence, at 62 cM, of linkage with diabetes in European whites, but not in Mexican–American samples [33]. A type 2 diabetes linkage study encompassing the entire long arm of chromosome 20 in Europeans identified not only the above linkage signal at ~55 cM, but also a second linkage peak at ~15 cM; this latter signal contains the PLCG1 gene [34]. Phospholipase C, γ1 (PLCG1) catalyses the conversion of phosphatidylinositol 4,5-bisphosphate to inositol 1,4,5-trisphosphate and diacylglycerol, playing a role in the signal transduction of receptor tyrosine kinases. Its relationship to insulin receptor function is unknown. PLCG1 also plays a role in membrane trafficking and endocytosis [35], and therefore might participate in insulin clearance. Zinc fingers and homeoboxes 3 (ZHX3), a ubiquitous transcriptional repressor [36], has no known relationship to insulin clearance.
One challenge in the future investigation of the genetics of insulin clearance is that it is not a trait routinely available in large epidemiological studies. Thus in the recent GWAS that have dramatically advanced our knowledge of type 2 diabetes genes, most novel genes discovered have been implicated as factors affecting insulin secretion, largely based on association with OGTT-derived measures of insulin secretion (e.g. insulinogenic index) [5]. The impact of insulin clearance on insulin levels during the OGTT is unclear; moreover, insulin clearance is not readily derived from standard OGTT data. As has been recognised for insulin resistance [37], the lack of an accurate measure of insulin clearance in diabetes cohorts will limit the ability to investigate whether insulin clearance is altered by any of the type 2 diabetes genes. Therefore focused studies specifically attempting to associate diabetes genes with insulin clearance will be needed. So far, variants in only two type 2 diabetes loci, HHEX/IDE and PPARG, have been associated with insulin clearance [38, 39]. HHEX/IDE encompasses insulin-degrading enzyme (IDE), the main enzyme responsible for insulin breakdown.
An understanding of the genetics of insulin clearance will be relevant not only to diabetes, but to a number of other common genetic conditions characterised by hyperinsulinaemia, such as obesity, the metabolic syndrome, polycystic ovary syndrome, non-alcoholic fatty liver disease, hypertension and coronary artery disease. These metabolic conditions are highly prevalent in our society and all appear to be, in part, genetically determined. Thus, inherited factors regulating insulin clearance may influence these conditions by modulating circulating insulin levels.
One limitation of our study is the accuracy of SSPI as a measurement of the metabolic clearance rate of insulin. Because we did not measure C-peptide levels during the euglycaemic clamps to document suppression of endogenous insulin secretion, our estimates of insulin clearance may underestimate the true values. However, because the proportion of SSPI represented by residual insulin secretion is expected to be small during hyperinsulinaemic infusion, we are confident that this had a minimal effect on our linkage and association results.
In our association study, we considered a value of p<0.05 to be significant. In general, correction for multiple testing is critical in genetic association studies, particularly when a large number of SNPs is being tested. However, in this study, the 3,761 SNPs analysed in the association study were not randomly selected, but chosen on the basis of their location under two linkage peaks. In this case, while it is not certain how multiple testing should be accounted for, it is clear that a standard approach based on the number of independent SNPs would be overly conservative. We observed 18 SNPs associated with SSPI in two cohorts, a marked excess over the nine expected by chance (3,761× 0.05×0.05). Association in two independent cohorts is one solution to the multiple testing issue; moreover, the observation that meta-analysis of the two studies resulted in greater statistical significance (Table 2) adds further credence to these associations. Our study benefited from being able to use two cohorts of the same ethnicity, originating from the same geographical region and with phenotyping performed using the same techniques. Nevertheless, additional independent confirmation will be needed to unequivocally establish these loci as factors influencing insulin clearance.
Future efforts will involve additional fine-mapping of the linkage signals for insulin clearance, as well as investigation of additional positional candidate genes under these peaks. Under the two linkage peaks we identified (chromosomes 15 and 20), 18 SNPs were associated with SSPI in two cohorts. These SNPs account for only small to moderate proportions of the total variance, indicating that other genetic variants may also contribute to variation in SSPI. Further large-scale studies are necessary to confirm these observations. This report represents an early step in the much-needed genetic characterisation of insulin clearance, a trait with relevance to type 2 diabetes and several other common metabolic disorders.
Supplementary Material
Acknowledgments
Funding This study was supported by National Institutes of Health (NIH) grants R01-DK079888, R01-HL067974, P50-HL055005 and P30-DK063491, and by General Clinical Research Center grants M01-RR000425 and M01-RR000043. The study was also supported by the National Center for Research Resources, Grant UL1-RR033176, now at the National Center for Advancing Translational Sciences, Grant UL1-TR000124. Further support came from the Cedars-Sinai Winnick Clinical Scholars Award (to M.O. Goodarzi) and the Cedars-Sinai Board of Governors’ Chair in Medical Genetics (to J.I. Rotter).
Abbreviations
- CYP1A2
Cytochrome P450, family 1, subfamily A, polypeptide 2
- CSK
c-Src tyrosine kinase
- GWAS
Genome-wide association study
- HOMA-%B
HOMA of beta cell function
- HOMA-IR
HOMA of insulin resistance
- HTN–IR
Hypertension–Insulin Resistance cohort
- IBD
Identity by descent
- LOD
Logarithm of odds
- M value
Marker of insulin sensitivity
- MACAD
Mexican–American Coronary Artery Disease cohort
- PLCG1
Phospholipase C, γ1
- SNP
Single nucleotide polymorphism
- SSPI
Steady-state plasma insulin
- ZHX3
Zinc fingers and homeoboxes 3
Footnotes
Electronic supplementary material The online version of this article (doi:10.1007/s00125-012-2577-2) contains peer-reviewed but unedited supplementary material, which is available to authorised users.
Duality of interest The authors declare that there is no duality of interest associated with this manuscript.
Contribution statement All authors made substantial contributions to conception and design, the acquisition of data or analysis and interpretation of data, as well as to the drafting of the article or its critical revision for important intellectual content. All authors gave final approval of the version to be published.
Contributor Information
X. Guo, Medical Genetics Institute, Cedars-Sinai Medical Center, Los Angeles, CA, USA. Department of Biomedical Sciences, Cedars-Sinai Medical Center, Los Angeles, CA, USA
J. Cui, Medical Genetics Institute, Cedars-Sinai Medical Center, Los Angeles, CA, USA
M. R. Jones, Department of Biomedical Sciences, Cedars-Sinai Medical Center, Los Angeles, CA, USA. Division of Endocrinology, Diabetes and Metabolism, Cedars-Sinai Medical Center, 8700 Beverly Blvd., Room B-131, Los Angeles, CA 90048, USA
T. Haritunians, Medical Genetics Institute, Cedars-Sinai Medical Center, Los Angeles, CA, USA. Department of Biomedical Sciences, Cedars-Sinai Medical Center, Los Angeles, CA, USA
A. H. Xiang, Department of Research and Evaluation, Kaiser Permanente Southern California Medical Group, Pasadena, CA, USA
Y.-D. I. Chen, Medical Genetics Institute, Cedars-Sinai Medical Center, Los Angeles, CA, USA. Department of Biomedical Sciences, Cedars-Sinai Medical Center, Los Angeles, CA, USA
K. D. Taylor, Medical Genetics Institute, Cedars-Sinai Medical Center, Los Angeles, CA, USA. Department of Biomedical Sciences, Cedars-Sinai Medical Center, Los Angeles, CA, USA
T. A. Buchanan, Department of Medicine, Keck School of Medicine, University of Southern California, Los Angeles, CA, USA
R. C. Davis, Department of Medicine, David Geffen School of Medicine at UCLA, Los Angeles, CA, USA
W. A. Hsueh, Diabetes Research Center, Division of Diabetes, Obesity and Lipids, Methodist Hospital Research Institute, Houston, TX, USA
L. J. Raffel, Medical Genetics Institute, Cedars-Sinai Medical Center, Los Angeles, CA, USA. Department of Biomedical Sciences, Cedars-Sinai Medical Center, Los Angeles, CA, USA
J. I. Rotter, Medical Genetics Institute, Cedars-Sinai Medical Center, Los Angeles, CA, USA. Department of Biomedical Sciences, Cedars-Sinai Medical Center, Los Angeles, CA, USA
M. O. Goodarzi, Email: mark.goodarzi@cshs.org, Medical Genetics Institute, Cedars-Sinai Medical Center, Los Angeles, CA, USA. Department of Biomedical Sciences, Cedars-Sinai Medical Center, Los Angeles, CA, USA. Division of Endocrinology, Diabetes and Metabolism, Cedars-Sinai Medical Center, 8700 Beverly Blvd., Room B-131, Los Angeles, CA 90048, USA
References
- 1.Narayan KM, Boyle JP, Thompson TJ, Sorensen SW, Williamson DF. Lifetime risk for diabetes mellitus in the United States. JAMA. 2003;290:1884–1890. doi: 10.1001/jama.290.14.1884. [DOI] [PubMed] [Google Scholar]
- 2.Cowie CC, Rust KF, Byrd-Holt DD, et al. Prevalence of diabetes and impaired fasting glucose in adults in the U.S. population: National Health and Nutrition Examination Survey 1999–2002. Diabetes Care. 2006;29:1263–1268. doi: 10.2337/dc06-0062. [DOI] [PubMed] [Google Scholar]
- 3.Cowie CC, Rust KF, Ford ES, et al. Full accounting of diabetes and pre-diabetes in the U.S. population in 1988–1994 and 2005–2006. Diabetes Care. 2009;32:287–294. doi: 10.2337/dc08-1296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Defronzo RA. Banting Lecture. From the triumvirate to the ominous octet: a new paradigm for the treatment of type 2 diabetes mellitus. Diabetes. 2009;58:773–795. doi: 10.2337/db09-9028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Perry JR, Frayling TM. New gene variants alter type 2 diabetes risk predominantly through reduced beta-cell function. Curr Opin Clin Nutr Metab Care. 2008;11:371–377. doi: 10.1097/MCO.0b013e32830349a1. [DOI] [PubMed] [Google Scholar]
- 6.Fontbonne A, Charles MA, Thibult N, et al. Hyperinsulinaemia as a predictor of coronary heart disease mortality in a healthy population: the Paris Prospective Study, 15-year follow-up. Diabetologia. 1991;34:356–361. doi: 10.1007/BF00405009. [DOI] [PubMed] [Google Scholar]
- 7.Flier JS, Minaker KL, Landsberg L, Young JB, Pallotta J, Rowe JW. Impaired in vivo insulin clearance in patients with severe target-cell resistance to insulin. Diabetes. 1982;31:132–135. doi: 10.2337/diab.31.2.132. [DOI] [PubMed] [Google Scholar]
- 8.Haffner SM, Stern MP, Watanabe RM, Bergman RN. Relationship of insulin clearance and secretion to insulin sensitivity in non-diabetic Mexican Americans. Eur J Clin Invest. 1992;22:147–153. doi: 10.1111/j.1365-2362.1992.tb01819.x. [DOI] [PubMed] [Google Scholar]
- 9.Jones CN, Abbasi F, Carantoni M, Polonsky KS, Reaven GM. Roles of insulin resistance and obesity in regulation of plasma insulin concentrations. Am J Physiol Endocrinol Metab. 2000;278:E501–E508. doi: 10.1152/ajpendo.2000.278.3.E501. [DOI] [PubMed] [Google Scholar]
- 10.Scheen AJ, Paquot N, Letiexhe MR, Paolisso G, Castillo MJ, Lefebvre PJ. Glucose metabolism in obese subjects: lessons from OGTT, IVGTT and clamp studies. Int J Obes Relat Metab Disord. 1995;19(Suppl 3):S14–S20. [PubMed] [Google Scholar]
- 11.Goodarzi MO, Taylor KD, Guo X, et al. Variation in the gene for muscle-specific AMP deaminase is associated with insulin clearance, a highly heritable trait. Diabetes. 2005;54:1222–1227. doi: 10.2337/diabetes.54.4.1222. [DOI] [PubMed] [Google Scholar]
- 12.Xiang AH, Azen SP, Raffel LJ, et al. Evidence for joint genetic control of insulin sensitivity and systolic blood pressure in Hispanic families with a hypertensive proband. Circulation. 2001;103:78–83. doi: 10.1161/01.cir.103.1.78. [DOI] [PubMed] [Google Scholar]
- 13.Matthews DR, Hosker JP, Rudenski AS, Naylor BA, Treacher DF, Turner RC. Homeostasis model assessment: insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia. 1985;28:412–419. doi: 10.1007/BF00280883. [DOI] [PubMed] [Google Scholar]
- 14.Ghebranious N, Vaske D, Yu A, Zhao C, Marth G, Weber JL. STRP screening sets for the human genome at 5 cM density. BMC Genomics. 2003;4:6. doi: 10.1186/1471-2164-4-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Almasy L, Blangero J. Multipoint quantitative-trait linkage analysis in general pedigrees. Am J Hum Genet. 1998;62:1198–1211. doi: 10.1086/301844. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Chen MH, Yang Q. GWAF: an R package for genome-wide association analyses with family data. Bioinformatics. 2010;26:580–581. doi: 10.1093/bioinformatics/btp710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Willer CJ, Li Y, Abecasis GR. METAL: fast and efficient meta-analysis of genomewide association scans. Bioinformatics. 2010;26:2190–2191. doi: 10.1093/bioinformatics/btq340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Stromblad G, Bjorntorp P. Reduced hepatic insulin clearance in rats with dietary-induced obesity. Metabolism. 1986;35:323–327. doi: 10.1016/0026-0495(86)90148-4. [DOI] [PubMed] [Google Scholar]
- 19.Mittelman SD, van Citters GW, Kim SP, et al. Longitudinal compensation for fat-induced insulin resistance includes reduced insulin clearance and enhanced beta-cell response. Diabetes. 2000;49:2116–2125. doi: 10.2337/diabetes.49.12.2116. [DOI] [PubMed] [Google Scholar]
- 20.Kim SH, Abbasi F, Chu JW, et al. Rosiglitazone reduces glucose-stimulated insulin secretion rate and increases insulin clearance in nondiabetic, insulin-resistant individuals. Diabetes. 2005;54:2447–2452. doi: 10.2337/diabetes.54.8.2447. [DOI] [PubMed] [Google Scholar]
- 21.Ford ES. Prevalence of the metabolic syndrome defined by the International Diabetes Federation among adults in the U.S. Diabetes Care. 2005;28:2745–2749. doi: 10.2337/diacare.28.11.2745. [DOI] [PubMed] [Google Scholar]
- 22.Goodarzi MO, Cui J, Chen YD, Hsueh WA, Guo X, Rotter JI. Fasting insulin reflects heterogeneous physiological processes: role of insulin clearance. Am J Physiol Endocrinol Metab. 2011;301:E402–E408. doi: 10.1152/ajpendo.00013.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Nagase T, Nakayama M, Nakajima D, Kikuno R, Ohara O. Prediction of the coding sequences of unidentified human genes. XX. The complete sequences of 100 new cDNA clones from brain which code for large proteins in vitro. DNA Res. 2001;8:85–95. doi: 10.1093/dnares/8.2.85. [DOI] [PubMed] [Google Scholar]
- 24.Newton-Cheh C, Johnson T, Gateva V, et al. Genome-wide association study identifies eight loci associated with blood pressure. Nat Genet. 2009;41:666–676. doi: 10.1038/ng.361. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Lowry WE, Huang J, Ma YC, et al. Csk, a critical link of g protein signals to actin cytoskeletal reorganization. Dev Cell. 2002;2:733–744. doi: 10.1016/s1534-5807(02)00175-2. [DOI] [PubMed] [Google Scholar]
- 26.Cao H, Sanguinetti AR, Mastick CC. Oxidative stress activates both Src-kinases and their negative regulator Csk and induces phosphorylation of two targeting proteins for Csk: caveolin-1 and paxillin. Exp Cell Res. 2004;294:159–171. doi: 10.1016/j.yexcr.2003.11.010. [DOI] [PubMed] [Google Scholar]
- 27.Engelman JA, Zhang X, Galbiati F, et al. Molecular genetics of the caveolin gene family: implications for human cancers, diabetes, Alzheimer disease, and muscular dystrophy. Am J Hum Genet. 1998;63:1578–1587. doi: 10.1086/302172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Cornelis MC, Monda KL, Yu K, et al. Genome-wide meta-analysis identifies regions on 7p21 (AHR) and 15q24 (CYP1A2) as determinants of habitual caffeine consumption. PLoS Genet. 2011;7:e1002033. doi: 10.1371/journal.pgen.1002033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Schunkert H, Konig IR, Kathiresan S, et al. Large-scale association analysis identifies 13 new susceptibility loci for coronary artery disease. Nat Genet. 2011;43:333–338. doi: 10.1038/ng.784. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Hurskainen TL, Hirohata S, Seldin MF, Apte SS. ADAM-TS5, ADAM-TS6, and ADAM-TS7, novel members of a new family of zinc metalloproteases. General features and genomic distribution of the ADAM-TS family. J Biol Chem. 1999;274:25555–25563. doi: 10.1074/jbc.274.36.25555. [DOI] [PubMed] [Google Scholar]
- 31.Mineur YS, Abizaid A, Rao Y, et al. Nicotine decreases food intake through activation of POMC neurons. Science. 2011;332:1330–1332. doi: 10.1126/science.1201889. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Permutt MA, Wasson J, Love-Gregory L, et al. Searching for type 2 diabetes genes on chromosome 20. Diabetes. 2002;51(Suppl 3):S308–S315. doi: 10.2337/diabetes.51.2007.s308. [DOI] [PubMed] [Google Scholar]
- 33.Guan W, Pluzhnikov A, Cox NJ, Boehnke M. Meta-analysis of 23 type 2 diabetes linkage studies from the International Type 2 Diabetes Linkage Analysis Consortium. Hum Hered. 2008;66:35–49. doi: 10.1159/000114164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Zouali H, Hani EH, Philippi A, et al. A susceptibility locus for early-onset non-insulin dependent (type 2) diabetes mellitus maps to chromosome 20q, proximal to the phosphoenolpyruvate carboxykinase gene. Hum Mol Genet. 1997;6:1401–1408. doi: 10.1093/hmg/6.9.1401. [DOI] [PubMed] [Google Scholar]
- 35.Heath CM, Stahl PD, Barbieri MA. Lipid kinases play crucial and multiple roles in membrane trafficking and signaling. Histol Histopathol. 2003;18:989–998. doi: 10.14670/HH-18.989. [DOI] [PubMed] [Google Scholar]
- 36.Yamada K, Kawata H, Shou Z, et al. Analysis of zinc-fingers and homeoboxes (ZHX)-1-interacting proteins: molecular cloning and characterization of a member of the ZHX family, ZHX3. Biochem J. 2003;373:167–178. doi: 10.1042/BJ20021866. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Florez JC. Newly identified loci highlight beta cell dysfunction as a key cause of type 2 diabetes: where are the insulin resistance genes? Diabetologia. 2008;51:1100–1110. doi: 10.1007/s00125-008-1025-9. [DOI] [PubMed] [Google Scholar]
- 38.Pivovarova O, Nikiforova VJ, Pfeiffer AF, Rudovich N. The influence of genetic variations in HHEX gene on insulin metabolism in the German MESYBEPO cohort. Diabetes Metab Res Rev. 2009;25:156–162. doi: 10.1002/dmrr.926. [DOI] [PubMed] [Google Scholar]
- 39.Tschritter O, Fritsche A, Stefan N, et al. Increased insulin clearance in peroxisome proliferator-activated receptor gamma2 Pro12Ala. Metabolism. 2003;52:778–783. doi: 10.1016/s0026-0495(03)00071-4. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.