

Abstract
The Andes of South America hosts perhaps the highest amphibian species diversity in the world, and a sizable component of that diversity is comprised of direct-developing frogs of the genus Pristimantis (Anura: Craugastoridae). In order to better understand the initial stages of species formation in these frogs, this study quantified local-scale spatial genetic structuring in three species of Pristimantis. DNA sequences of two mitochondrial gene fragments (16S and COI) were obtained from P. brevifrons, P. palmeri and P. jubatus at different locations in the Cordillera Occidental. We found high levels of genetic diversity in the three species, with highly structured populations (as measured by F ST) in P. brevifrons and P. palmeri while P. jubatus showed panmixia. Large effective population sizes, inferred from the high levels of genetic diversity, were found in the three species and two highly divergent lineages were detected within P. jubatus and P. palmeri. Estimated divergence times among populations within P. brevifrons and P. palmeri coincide with the Pleistocene, perhaps due to similar responses to climatic cycling or recent geological history. Such insights have important implications for linking alpha and beta diversity, suggesting regional scale patterns may be associated with local scale processes in promoting differentiation among populations in the Andes.
Introduction
The Andes of South America contain the highest levels of total species richness in the world [1], [2]. The high biodiversity of tropical montane regions may be due to their old age [3], higher rates of adaptive divergence across elevation gradients [4], [5], greater opportunities for vicariant speciation [6], [7], [8] or a mixture of factors [9], [10]. Montane diversity is often spatially partitioned, i.e., beta diversity (species composition between sites) is quite high compared to the lowlands, while alpha diversity (species within a community) is relatively low [11]. The observation that sister species, at least in vertebrates, tend to be in similar habitats [8] suggests that simple vicariance models [12] could potentially account for much montane diversity compared with models invoking ecological gradients [5]. Indeed, mountain ridges and valleys have long been considered as agents of allopatric speciation by acting as effective barriers to dispersal [13].
A given geographic barrier may not have the same effect on all species, however. Organismal biology and natural history interact with landscape features to determine to what extent a potential barrier will affect dispersal [6], [14]. Over evolutionary time the context-specific dispersal capacity of an organism will determine spatial genetic patterns [15], [16]. Even congeneric species may have contrasting ecological requirements that result in distinct phylogeographic histories [17].
Using molecular data it is possible to quantify the magnitude of genetic variation within populations and patterns of gene flow among conspecific populations. Moreover, if shared past events led to the formation of current genetic patterns, then comparing phylogeographic patterns of sympatric species may enhance our understanding of the relative roles performed by the forces of climatic, geological and ecological conditions in structuring within-species genetic variation [18], [19], [20]. Molecular studies of different amphibians in the montane Neotropics have revealed high levels of genetic diversity across small geographic areas [21], [22], [23]. However, in South America and especially in Colombia, there are only a few molecular studies on genetic diversity of widely distributed montane amphibian taxa endemic to particular regions [9]. The mountains of southwestern Colombia are considered hotspots of diversity for their unique assemblages of anurans and other taxa [24]. The number of species of anurans in the Cordillera Occidental of Colombia is steadily increasing [24], with over 150 species currently found at elevations above 1,000 meters [25].
The genus Pristimantis (Anura: Terrarana: Craugastoridae) is the dominant component of the total amphibian fauna of Colombia, with 200 species, i.e., 27% of the country’s 750 species [26]. Nevertheless, little is known about their ecology, phylogenetic relationships [27], population genetic structure, or levels of genetic polymorphism. This study focused on three species of Pristimantis with distributions in the Cordillera Occidental of Colombia. Pristimantis jubatus [28] is found at 2,450–2,750 meters above sea level (masl) in Munchique National Park, with a total range less than 5,000 km2 in the department of Cauca. This species is considered to be locally abundant, inhabits forests with high (>90%) average humidity and dense vegetation cover (>80%) and appears to be intolerant of habitat modifications [28], [29]. In contrast, Pristimantis palmeri [30] and P. brevifrons [31] are sympatric in the Cordillera Occidental, ranging from the departments of Cauca to Risaralda, with elevational ranges of 900 to 2,400 masl and 1,140–3,200 masl, respectively. They are common species with apparently stable populations, most frequently found in disturbed situations, and are considered as generalist species and easily adaptable to modified landscapes.
These species were chosen because the accessibility of the area, availability of samples, their co-distribution in the Cordillera Occidental, and their contrasting ecological requirements. Thus, they should provide a useful opportunity to investigate the local historical demographic processes behind the larger scale biogeographic patterns in an ecoregion with a dynamic geologic history [32]. Information on intraspecific genetic variation is also important to inform conservation plans at the level of communities and biogeographic subregions [33], because data on population sizes and migration rates may be useful for predicting the effects of disease, climate change and isolation by habitat fragmentation on amphibian populations [34], [35], [36].
Determining the processes promoting diversification and the geographic and temporal scales at which they operate is key to understanding how this group of direct-developing frogs has attained such remarkable levels of beta diversity across the Andes mountains [27]. This study investigates the population histories on different spatial scales of three species of Pristimantis and focuses on the following questions: 1. Is genetic diversity randomly distributed in each species with respect to geography? 2. Do species with broad versus narrow distributions show similar patterns in the distribution of genetic variation? 3. Do sympatric species have common phylogeographic patterns because they share similar environmental histories, or do species with contrasting life histories show distinct phylogeographic patterns?
Results
MtDNA Diversity
The 16S alignment contained 536 base pairs (bp) for P. brevifrons (N = 19 individuals), and 537 bp for P. palmeri (N = 32) and P. jubatus (N = 30). This locus showed the number of segregating sites, S = 18 for P. brevifrons, S = 12 for P. palmeri, and S = 26 for P. jubatus. The COI alignment contained 658 bp for P. brevifrons (N = 16), and 618 bp for P. palmeri (N = 35) and P. jubatus (N = 31). Pristimantis brevifrons showed S = 30 polymorphic sites, while P. palmeri and P. jubatus contained S = 110 and S = 50 sites, respectively. The translation of amino acids using the genetic code for vertebrates showed that COI variants were synonymous (silent).
Haplotype relationships were illustrated in a median-joining network (Fig. 1). The two sampling localities for P. brevifrons (Los Paraguas and Peñas Blancas), located 151.8 km apart and about at roughly same altitude, did not share haplotypes (each locality contained unique haplotypes) either in 16S or COI, which is consistent with a long period of isolation. As expected from the geography (Fig. 2), for P. palmeri the Serranía de Los Paraguas had the greatest differentiation as compared to the other two more proximal localities (Peñas Blancas and Chicoral) (Fig. 1). Chicoral and Peñas Blancas sites are separated by 18 km, while the northern population of Los Paraguas is more than 134 km apart from those sites.
Figure 1. Networks of mitochondrial DNA haplotypes for 16S (left) and COI (right) from P. brevifrons, P. palmeri and P. jubatus.

Branches represent inferred mutational steps, where more than three mutations are denoted by the number of substitutions. The size of the circle is proportional to the number of individuals found for each haplotype. The two highly divergent specimens from Peñas Blancas in P. palmeri and one from Munchique National Park in P. jubatus were included in the networks.
Figure 2. Map of western Colombia showing the four sampling localities in the departments of Cauca and Valle del Cauca.

There was a highly divergent COI haplotype found in two specimens (UVC15845 and UVC15857) within Peñas Blancas population of P. palmeri that were 17% (p-distance) diverged from what were presumably conspecific samples found at the same site. We were unable to amplify 16S from these same two specimens, unfortunately. Furthermore, there was one haplotype from the Chicoral population closely related to samples found at Peñas Blancas, indicating recent gene flow between these populations (Fig. 1). Pristimantis jubatus showed relatively low mtDNA divergences among sampling sites, with a total of four haplotypes for both genes. One individual of P. jubatus (UVC15842) from the site Observatorio was highly diverged (4% and 8%, p-distances for 16S and COI, respectively) from its conspecifics, with 22 nucleotide differences at 16S and 46 differences at COI from others at the same locality (Fig. 1). These divergence haplotypes within P. palmeri and P. jubatus may be migrants from an unsampled locality or cryptic species considering the intra-specific threshold values of divergence proposed by Vences et al. [37] and Fouquet et al. [38] of 3–5% at 16S and 10% at COI.
Population Genetic Structure
For two of the three species, mtDNA variation showed patterns of population subdivision. Population genetic structure was high among localities for the widespread, generalist species P. palmeri and P. brevifrons (Table 1). For P. palmeri the minimum estimated pairwise F ST among populations was found between Chicoral and Peñas Blancas (F ST = 0.682, probability of the null hypothesis of panmixa, p<0.01), which are geographically closer. The greatest population structuring was observed between Paraguas and Peñas Blancas (the two most distant sampling sites) in both P. brevifrons (F ST = 0.98, p<0.01) and P. palmeri (F ST = 0.95, p<0.01). However, substantial polymorphism within populations resulted in low but significant proportions of the total variation partitioned among localities in the AMOVA (29.9%, p<0.01 and 21.6%, p<0.01, for P. brevifrons and P. palmeri, respectively). In the more range-restricted and habitat-specialized species P. jubatus, in contrast, we observed low mtDNA sequences divergence among all three localities (F ST = 0.0076–0.2749) and a low percentage of the total variation partitioned among localities (1.34%, p>0.05). These results suggest that the three sampling localities 3 km apart, Observatorio, Santa Ana and Charguayaco (Table 2) within the Munchique National Park (Fig. 2) form a single genetic population.
Table 1. F ST estimated and confidence limits (in parentheses) between pairs of populations of P. brevifrons and P. palmeri using mtDNA sequences from the 16S and COI genes combined.
| Species | Locality | Chicoral | Peñas Blancas |
| P. brevifrons | Paraguas | 0.9840 (0.976–1.000) | |
| P. palmeri | Paraguas | 0.9387 (0.896–0.982) | 0.9506 (0.910–0.982) |
| Chicoral | 0.6821 (0.468–0.894) |
Table 2. Sampling locations.
| Locality | Taxon | N | Altitude (masl) | Transect (km)* | Description of site |
| Los Paraguas | P. brevifrons | 7 | 1950–2050 | 0.8 | Primary forest |
| Peñas Blancas | P. brevifrons | 12 | 2000–2200 | 1.4 | Secondary forest |
| Los Paraguas | P. palmeri | 7 | 1950–2200 | 1.0 | Primary forest |
| Chicoral | P. palmeri | 15 | 1800–1850 | 0.8 | Secondary forest |
| Peñas Blancas | P. palmeri | 13 | 1950–2050 | 1.2 | Secondary forest |
| Observatorio | P. jubatus | 13 | 2550 | 0.2 | Primary forest |
| Santa Ana | P. jubatus | 8 | 2700 | 0.3 | Primary forest |
| Charguayaco | P. jubatus | 10 | 2450 | 0.2 | Primary forest |
Transect refers to the straight distance along which samples were collected at each site. The three sites for P. jubatus are contained within the Munchique National Park (Fig. 2).
Tajima’s D statistic for P. jubatus for the combined Observatorio-Santa Ana-Charguayaco data (treated as one population, based on the above results), showed a significantly site-frequency skew for the mitochondrial dataset (DT ≤ - 2.5 for both genes, Table 3). These values were due to the sample UVC15842 (found in Observatorio, see above), which increased the number of singleton sites and caused a rejection of the standard neutral model. If this sample was excluded, neutrality was not rejected (Table 3). Summary statistics of genetic polymorphisms for all populations of the three species of Pristimantis are provided in Table 3.
Table 3. Genetic polymorphism data for mitochondrial DNA sequences from the Pristimantis populations in this study.
| Loci | Species | Population | N | h | S | θW | π | DT | DT 95% I.C. | Prob. DT≠0 Simul. Coal. |
| 16S | P. brevifrons | Paraguas | 7 | 2 | 1 | 0.00076 | 0.00053 | −1.0062 | −1.0062 to 1.341 | 0.490 |
| Peñas Blancas | 12 | 4 | 3 | 0.00186 | 0.00094 | −1.6292 | −1.629 to 1.972 | 0.543 | ||
| P. jubatus | Observatorio-Santa Ana-Charguayaco | 30 | 4 | 19 | 0.01047 | 0.0029 | −2.5212*/−1.248 | −1.712 to 1.816/−1.509 to 2.014 | 0.000 | |
| P. palmeri | Paraguas | 7 | 1 | 0 | 0 | 0 | N.A. | N.A. | N.A. | |
| Chicoral | 14 | 4 | 4 | 0.00235 | 0.00152 | −1.1644 | −1.797 to 1.886 | 0.1440 | ||
| Peñas Blancas | 11 | 1 | 0 | 0 | 0 | N.A. | N.A. | N.A. | ||
| COI | P. brevifrons | Paraguas | 6 | 2 | 3 | 0.00201 | 0.00153 | −1.2331 | −1.233 to 1.909 | 0.268 |
| Peñas Blancas | 10 | 2 | 1 | 0.00057 | 0.00032 | −1.1117 | −1.111 to 1.463 | 0.412 | ||
| P. jubatus | Observatorio-Santa Ana-Charguayaco | 31 | 4 | 48 | 0.02036 | 0.00604 | −2.6023*/0.2368 | −1.719 to 1.792/−1.507 to 2.027 | 0.000 | |
| P. palmeri | Paraguas | 7 | 3 | 2 | 0.00132 | 0.0017 | 1.1684 | −1.237 to 1.649 | 0.522 | |
| Chicoral | 15 | 7 | 10 | 0.00498 | 0.0045 | −0.3644 | −1.802 to 1.827 | 0.554 | ||
| Peñas Blancas | 13 | 5 | 94 | 0.04901 | 0.04527 | −0.3921 | −1.760 to 1.700 | 0.543 |
N = number of sequences, h = number of haplotypes, S = segregating sites, θW = population mutation rate per site, π = nucleotide diversity per site, DT = Tajima’s D statistic [73]. An asterisk (*) in the DT column indicates a significant deviation from the expected value under neutrality (DT = 0) as calculated from coalescence simulations. DT values in bold obtained for P. jubatus when the divergent sample UVC15842 was excluded. N.A. = Not Applicable. Indels and sites with missing data were not taken into account in pairwise comparisons.
Estimation of Historical Demographic Parameters
Gene flow (Nm) since initial divergence was very low between the two sampling localities of P. brevifrons, and within P. palmeri between the northerly Los Paraguas and the two southern sites. In contrast, significant and asymmetric gene flow was detected between the two adjacent populations of P. palmeri, with migration from Chicoral into Peñas Blancas being lower than in the reverse direction (Table 4, Fig. S1).
Table 4. Estimates obtained from IMa for the population size parameter theta (Ne), gene flow (Nm) and divergence times (t) and 90% highest posterior density (HPD) intervals for populations of P. brevifrons (one pairwise comparison among two localities) and P. palmeri (three pairwise comparisons among three localities), based on the combined COI and 16S data.
| Species | Samples | Parameter | ||||
| Ne 1 | Ne 2 | N 1 m 1 | N 2 m 2 | t, years | ||
| P. brevifrons | Paraguas/Peñas Blancas | 38595 | 68147 | 0.0006 | 0.0009 | 802500 |
| 90% HPD | 6056–149013 | 21238–181175 | 0.0001–1.6 | 0.0002–1.6 | 565227–1362954* | |
| P. palmeri | Paraguas/Peñas Blancas | 8697 | 52638 | 0.0002 | 0.0009 | 584545 |
| 90% HPD | 486–59206 | 16184–145029 | 0.00001–1.1 | 0.0002–1.8 | 406363–1817272* | |
| Paraguas/Chicoral | 12572 | 143393 | 0.0004 | 0.001 | 541818 | |
| 90% HPD | 813–73170 | 69452–280711 | 0.00003–1.5 | 0.0009–2.3 | 344000–1453818* | |
| Chicoral/Peñas Blancas | 102922 | 46075 | 1.2 | 0.004 | 146818 | |
| 90% HPD | 39411–240720 | 11765–133970 | 0.003–13.9 | 0.001–10.0 | 67727–908636* | |
indicates that HPD estimates are unreliable because the tail of the distribution did not reach zero.
Based on estimates of θ for the combined mitochondrial gene sequences and using an inheritance scalar of 0.25 and a mutation rate estimated from closely related frogs (see Methods), we calculated an effective population size (Ne) for P. brevifrons of 38,000 (90% HPD 6,000–150,000) and 68,000 (90% HPD 21,000–181,000) reproductive individuals in Paraguas and Peñas Blancas, respectively. For P. palmeri we estimated Ne at 10,000 (90% HPD 500–73,000) individuals in Paraguas, 50,000 (90% HPD 12,000–145,000) for Peñas Blancas and 100,000 (90% HPD 40,000–280,000) at Chicoral (Table 4, Fig. S2). To be more conservative in our estimates of N e we removed the unusually divergent specimens of P. palmeri (UVC15857 and UVC15845) from Peñas Blancas. Including these samples would inflate the number of segregating sites and bias estimates of N e upwards. Because the three collecting localities for P. jubatus apparently represented a single genetic population, their effective population size was obtained using MIGRATE [39]. As the specimen UVC15842 of P. jubatus caused significant departures from standard neutral expectations (see above), we removed this sample prior to calculating N e, yielding a more conservative estimated effective population size for this species of 94,000 (95% C.I. 15,600–470,000) individuals.
The upper tail of the posterior distribution for divergence times did not reach zero probability so we were unable to estimate a strict 90% HPD value for this parameter for any species. The partial posterior distributions, however, suggested the following tentative approximations of divergence times. The mode of the posterior distribution of divergence time among P. palmeri populations was about 0.58 million years ago (Mya) (90% HPD roughly 0.40–1.8 Mya) between Paraguas and Peñas Blancas, and divergence time between adjacent populations Chicoral and Peñas Blancas was 0.14 Mya (0.06–0.90 Mya) (Table 4, Fig. S3). Divergence times across the same terrain (Paraguas to Peñas Blancas) were slightly older in P. brevifrons (0.80 Mya; 90% HPD 0.56–1.4 Mya) than in P. palmeri (0.58).
Discussion
Studies on the intraspecific genetic differentiation and population structure of anurans in montane regions are sorely needed, considering that work in montane areas lags behind work done in low elevation environments of the Neotropics (e.g. Central America and Amazonia) [40], [41], [42]. We argue here that much interspecific diversity was likely derived from within-species diversity through vicariance processes, and these processes in term were actively promoted by millions of years of remarkable geological dynamism of the Andean Cordillera [21], [40]. Thus, an understanding of interspecific diversity patterns is informed by the study of within-species historical demographic processes [40], [43].
This study characterizes the genetic diversity, population structure and phylogeography of P. brevifrons, P. palmeri and P. jubatus using the mitochondrial markers COI and 16S. Estimates of gene flow, population sizes, and divergence time revealed important aspects of the historical demography of these three Neotropical montane species. Our data highlighted the undifferentiated structure of P. jubatus at a local geographical scale as well as the similar phylogeographic structure in P. brevifrons and P. palmeri, two species with similar life histories.
Comparative Phylogeographic of Pristimantis Frogs
For P. brevifrons and P. palmeri analyses of the two mitochondrial loci revealed high genetic differentiation in space cross the same geographic scale landscape (approx. 150 km.) and a similar inferred age of population differentiation in both species (around 0.80 Mya) (Fig. 1, Table 4). This implies that the two widespread species responded similarly to shared environmental processes producing similar population histories at the same spatial scale. Because these species are similar in terms of their ecological requirements, their parallel phylogeographic patterns suggest an eco-geological explanation of the geographical pattern during the Pleistocene [18], [19], [44].
The northern population of Los Paraguas showed low migration rates with the two southern sites, and a corresponding high pairwise population differentiation. This population appears to have split from the other locations during the Mid-Pleistocene (0.58–0.80 Mya), i.e., following the last episode of uplift of the northern portion of the Andes in the Pliocene (5.0–2.0 Mya) [45], [46]. The Serranía de Los Paraguas is a mountain formation that follows the western flank of the Cordillera Occidental, diverging from the main mountain range. This geological separation may be the cause of the restricted gene flow with the southern localities, thereby promoting the divergence of the Los Paraguas population. In the Cordillera Oriental of Colombia a similar pattern of isolation by ridges is found in unrelated frog species [9], [47].
Despite the dearth of ecological data for species of Pristimantis, these frogs are thought to have small home ranges along with low vagility [29], [48], which is supported by genetic data in a few species [40], [42]. At a smaller spatial scale (∼3 km), our analysis for the narrowly distributed P. jubatus, showed no significant spatial differentiation. We expected that the habitat specialist P. jubatus would show high structure but we likely need more geographically extensive sampling. On the other hand, we might have expected direct-developing frogs to show minimal population structuring as these amphibians do not require a water source for ovipositing or mating, so they may breed anywhere within a continuous forest of high relative humidity [40]. Pristimantis palmeri perhaps fits this prediction an intermediate spatial scale, i.e., the 18 km between Chicoral and Peñas Blancas. We estimated the asymmetric migration rates at 1.2 (90% HPD 0.003–13.9) and 0.004 (0.001–10.0), thus we could not reject the hypothesis of substantial levels of gene flow at this distance.
To date, there is a limited yet growing number of studies on genetic structure at local or regional scales in Neotropical frogs e.g., [40], [42], [49]. In other studies of terraranid frogs, Elmer et al. [42] found genetic structure at 4 km but Crawford [40] found no mitochondrial or nuclear differentiation between populations separated by 10 km. Also, Guarnizo et al. [9] found identical DNA sequences in localities as far away as 26 km in the high-elevation Andean hylid frog, Dendropsophus labialis. These results indicate that Andean frogs may disperse long distances at least in some cases or in some geographic context [27] and their range limits may be perhaps closely associated to altitude due to thermal limitations [50], [51], that may increase opportunities for speciation and high beta diversity in the Andes [52].
Our IMa analyses produced robust historical demographic parameter estimates, with the exception of divergences times. Features of the evolutionary history of our three study species revealed by coalescent analyses are particularly informative because few data on historical demography are currently available for tropical amphibians. Demographic estimates of population size are not available for P. brevifrons, P. palmeri or P. jubatus to corroborate the genetic-based estimates, however, the estimated population sizes for P. brevifrons, P. palmeri and P. jubatus are quite similar compared to those obtained by Crawford [40] for the terraranid species Craugastor bransfordii (3.1×105) and C. stejnegerianus (1.0×105). Thus, the effective population sizes are potentially very large in our three species of Pristimantis. Large population sizes could be positively correlated to higher speciation rates [53], yet a more parsimonious explanation may be that the beta diversity of Pristimantis in montane regions has been promoted by barriers, such as the Peñas Blancas ridge. Our results here inform these biogeographic models by confirming that even localized and isolated populations of these relatively small frogs may hold substantial amounts of genetic diversity, thus contributing to the maintenance of viable populations through subsequent vicariant events during montane diversification [23]. MtDNA lineages within Pristimantis frogs showed signs of demographic changes associated with recent Pleistocene and geological events. It will be interesting to examine similarly widespread species to evaluate the generality of these patterns and conduct more fine-scaled phylogeographical analyses to explore the possible mechanisms leading to speciation within this group of direct-developing frogs.
Materials and Methods
Ethics Statement
This study was carried out in strict accordance with the evaluations of environmental, social, and educational impacts established for research projects under Universidad del Valle Convocatoria Interna number CI-746. Permit for sampling of specimens was obtained from Ministerio de Medio Ambiente, Vivienda y Desarrollo Territorial de Colombia (Resolución 573 del 10 de abril/08). Euthanasia of frogs was performed with Chloretone, and all efforts were made to minimize suffering. Specimens were deposited at Herpetological Collection – Universidad del Valle (Voucher numbers: UVC15812– UVC15954) (Table S1) and DNA sequences data have been submitted to Genbank: accession numbers JN104663– JN104683; JN370956– JN371118.
Sampling
Sampling was carried out at four localities (Fig. 2): La Serranía de Los Paraguas, municipality El Cairo, on the border of the departments of Valle del Cauca and Chocó, 2,000–2,200 masl (4.7333°, −76.3°); Chicoral, municipality La Cumbre, department of Valle del Cauca, 1,800–1,900 masl (3.5833°, −76.6°); Peñas Blancas, in Farallones de Cali National Park, municipality Cali, department of Valle del Cauca, 1,900–2,200 masl (3.4333°, −76.6667°); and Munchique National Park, municipality El Tambo, department of Cauca, 2,400–2,800 masl (2.6667°, −76.9167°).
At each locality we established approximately straight-line transects across which samples were collected. Samples sizes ranged from 7–15 individuals per location (Table 2). Pristimantis jubatus was sampled at three different locations in the Munchique National Park (Observatorio, Santa Ana and Charguayaco), while widespread, generalist species P. brevifrons and P. palmeri were sampled at La Serranía de Los Paraguas and Peñas Blancas, and Serranía de Los Paraguas, Peñas Blancas and Chicoral, respectively. Tissues were collected in the field and kept in a solution of 1 M Tris HCl, 0.5 M EDTA, 5 M NaCl and SDS to 20%. Specimen identification was confirmed by Dr. John D. Lynch, Curator of Amphibians at the Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Bogotá.
Laboratory Techniques
Total DNA was extracted from liver or thigh tissue using the Qiagen Tissue DNeasy kit (Qiagen) following the manufacturer’s instructions. Mitochondrial DNA (mtDNA) fragments were amplified using PCR primers 16 Sbr plus 16 Sar for the ARN region of the 16S ribosomal RNA gene [54] and LCO1490 plus HCO2198 for the 5′ half of the cytochrome oxidase subunit I (COI), also known as the Folmer fragment or Barcode of Life fragment for animals [55], [56]. The DNA amplification reactions contained 2.5 mM MgCl2, 0.3 mM dNTPs, 0.3 µM of each Primer, 0.625 U of Taq polymerase and 1–2 µl total DNA. PCR conditions followed Goebel et al. [57] and Vences et al. [58]. Negative controls were used to monitor potential contamination in PCR. Amplification products were purified with QIAquick PCR clean-up kit (Qiagen). Both H (heavy) and L (light) strands were sequenced using the chemical reaction Big-Dye Terminator v. 3.1 reagents in an automated DNA sequencer (ABI 3730XL, Applied Biosystems Inc.). The sequences were deposited in GenBank (Table S1).
Analyses
DNA sequence analysis
Sequences were edited, assembled and aligned using the program Sequencher 4.6 (Gene Codes Corporation) and verified by eye. COI sequences were translated into amino acids with the program DnaSP 5.0 [59] to confirm the absence of inferred nonsense mutations. We used DnaSP to calculate the following summary statistics for genetic variation within each population: haplotype diversity (Hd), nucleotide diversity per site (π ) [60], the number of segregating sites (S), Watterson’s estimator of the per-site population mutation rate (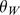 ) [61] and Tajima’s D statistic (D
T). Sites with indels or missing data were eliminated in pairwise sequence comparisons. Coalescence simulations were run in DnaSP to test for significant departures from the expected value D
T = 0 under the standard neutral model using 5,000 replicates conditional on sample size and S.
) [61] and Tajima’s D statistic (D
T). Sites with indels or missing data were eliminated in pairwise sequence comparisons. Coalescence simulations were run in DnaSP to test for significant departures from the expected value D
T = 0 under the standard neutral model using 5,000 replicates conditional on sample size and S.
Analysis of population differentiation
We used the programs Arlequin 3.1 [62] and DnaSP 5.0 to conduct an Analysis of Molecular Variance (AMOVA) and calculate Lynch & Crease’s pairwise F ST [63] to test the null hypothesis of panmixia between pairs of populations [64] using 1000 permutations and significance level of 0.05. F ST was chosen as the metric parameter of the genetic differentiation among populations because it provides a more effective summary of the effects of population structure [65]. To visualize the relationship between the genealogy of haplotypes and geography, haplotype networks were inferred using median joining as implemented in the program Network 4.5.1.0 [66].
Estimation historical demographic parameters
We estimated the effective population sizes (N e), asymmetric pairwise migration rates (Nm) and time since population divergence (t) with the programs IMa [67] and MIGRATE [39]. To convert parameter estimates of IMa into demographic units, an inheritance scalar of 0.25 and generation time of one year were assumed. To obtain N e from θ ( = 4N e μ, under the standard neutral model) we assumed a mutation rate, μ, of 1.9×10−8 per site per year, based on estimated rates of silent site divergence in a genus of frogs closely related to Pristimantis [68]. While the COI gene typically has higher overall rates of substitution relative to the more conserved 16S gene [37], [69], [70], we assume that these two mitochondrial genes experience comparable rates of mutation [71] (as opposed to substitution rates). We used an infinite-site model of substitution [72] for both genes, 16S and COI. Several preliminary MCMC simulations with broad priors were performed to set appropriate bounds for each parameter (θ, m, t). We compared the convergence of the marginal distributions of each parameter among multiple runs and obtained the credibility intervals based on 90% highest posterior density (HPD).
Supporting Information
Posterior probability distributions for gene flow from IMa analysis.
(TIF)
Posterior probability distributions for effective population sizes from IMa analysis.
(TIF)
Posterior probability distributions for divergence times from IMa analysis.
(TIF)
Sample information for taxa analyzed in this study.
(DOC)
Acknowledgments
We thank the Ministerio de Medio Ambiente, Vivienda y Desarrollo Territorial de Colombia for research permission (Resolución 573 del 10 de abril/08) and the Universidad del Valle for funding (Convocatoria Interna CI-746). The Programa de Postgrado de Ciencias-Biología and the Grupo de Estudios Ecogenéticos y Biología Molecular of the Universidad del Valle provided training in molecular techniques, support and academic assistance. Thanks to Ranulfo González and Guillermo Barreto for their advice, Diana López for access to software and literature at the Instituto de Investigaciones Alexander von Humboldt, John D. Lynch from the Instituto de Ciencias Naturales-UNAL for access to specimens, Luz Piedad Aguirre and Juan Esteban Zuluaga from the Vicerrectoria de Investigaciones of the Universidad del Valle for support, and to the rangers and inhabitants of La Romelia -Munchique National Park-, Peñas Blancas, Pichindé, El Cairo, Chicoral, and Loma Alta. Our work in Los Paraguas was facilitated by Serraniaguas. Wilmar Bolivar, Nelsy Pinto, Fernanda Castillo and an anonymous reviewer for insightful and helpful comments on various versions of the manuscript. Isabel Castro, Kyle Morrison and Alexander Wilson helped in the translation of the manuscript. This work partially fulfilled the requirements for the Masters degree of JCG-R at the Universidad del Valle.
Funding Statement
This research was supported by Universidad del Valle (Convocatoria Interna CI-746). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Myers N, Mittermeier RA, Mittermeier CG, da Fonseca GAB, Kent J (2000) Biodiversity hotspots for conservation priorities. Nature 403: 853–858. [DOI] [PubMed] [Google Scholar]
- 2.Kessler M, Kluge J (2008) Diversity and endemism in tropical montane forests - from patterns to processes. In: Gradstein SR, Homeier J, Gansert D, editors. The Tropical mountain forest: Patterns and processes in a biodiversity hotspot: Göttingen Centre for Biodiversity and Ecology. 35–50. [Google Scholar]
- 3. Wiens JJ, Parra-Olea G, García-París M, Wake DB (2007) Phylogenetic history underlies elevational biodiversity patterns in tropical salamanders. Proceedings of the Royal Society B: Biological Sciences 274: 919–928. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Cadena CD (2007) Testing the role of interspecific competition in the evolutionary origin of elevational zonation: An example with Buarremon Brush-Finches (Aves, Emberizidae) in the neotropical mountains. Evolution 61: 1120–1136. [DOI] [PubMed] [Google Scholar]
- 5. Moritz C, Patton JL, Schneider CJ, Smith TB (2000) Diversification of rainforest faunas: an integrated molecular approach. Annual Review of Ecology and Systematics 31: 533–563. [Google Scholar]
- 6. Janzen DH (1967) Why mountain passes are higher in the Tropics. The American Naturalist 101: 233. [Google Scholar]
- 7. Ghalambor CK, Huey RB, Martin PR, Tewksbury JJ, Wang G (2006) Are mountain passes higher in the tropics? Janzen’s hypothesis revisited. Integrative and Comparative Biology 46: 5–17. [DOI] [PubMed] [Google Scholar]
- 8.Cadena CD, Kozak KH, Gómez JP, Parra JL, McCain CM, et al.. (2011) Latitude, elevational climatic zonation and speciation in New World vertebrates. Proceedings of the Royal Society B: Biological Sciences. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Guarnizo CE, Amézquita A, Bermingham E (2009) The relative roles of vicariance versus elevational gradients in the genetic differentiation of the high Andean tree frog, Dendropsophus labialis . Molecular Phylogenetics and Evolution 50: 84–92. [DOI] [PubMed] [Google Scholar]
- 10. Smith SA, Montes de Oca AN, Reeder TW, Wiens JJ (2007) A phylogenetic perspective on elevational species richness patterns in Middle American treefrogs: Why so few species in lowland tropical rainforests? Evolution 61: 1188–1207. [DOI] [PubMed] [Google Scholar]
- 11. Jankowski JE, Ciecka AL, Meyer NY, Rabenold KN (2009) Beta diversity along environmental gradients: implications of habitat specialization in tropical montane landscapes. Journal of Animal Ecology 78: 315–327. [DOI] [PubMed] [Google Scholar]
- 12. Wiens JJ (2004) Speciation and ecology revisited: Phylogenetic niche conservatism and the origin of species. Evolution 58: 193–197. [DOI] [PubMed] [Google Scholar]
- 13. Lynch JD, Duellman WE (1997) Frogs of the genus Eleutherodactylus in western Ecuador: systematics, ecology, and biogeography. The University of Kansas, Natural History Museum Special Publication 23: 1–236. [Google Scholar]
- 14. Burney CW, Brumfield RT (2009) Ecology predicts levels of genetic differentiation in Neotropical birds. The American Naturalist 174: 358–368. [DOI] [PubMed] [Google Scholar]
- 15.Avise JC (2000) Phylogeography: The history and formation of species. Cambridge, Massachusetts: Harvard University Press. [Google Scholar]
- 16. Beavis A, Sunnucks P, Rowell D (2011 ) Microhabitat preferences drive phylogeographic disparities in two Australian funnel web spiders Biological Journal of the Linnean Society. 104: 805–819. [Google Scholar]
- 17. Crawford AJ, Bermingham E, Polanía C (2007) The role of tropical dry forest as a long-term barrier to dispersal: a comparative phylogeographical analysis of dry forest tolerant and intolerant frogs. Molecular Ecology 16: 4789–4807. [DOI] [PubMed] [Google Scholar]
- 18. Soltis D, Morris A, McLachlan J, Manos P, Soltis P (2006 ) Comparative phylogeography of unglaciated eastern North America Molecular Ecology. 15: 4261–4293. [DOI] [PubMed] [Google Scholar]
- 19. Dawson M (2012) Parallel phylogeographic structure in ecologically similar sympatric sister taxa Molecular Ecology. 21: 987–1004. [DOI] [PubMed] [Google Scholar]
- 20. Bermingham E, Moritz C (1998) Comparative phylogeography: concepts and applications. Molecular Ecology 7: 367–369. [Google Scholar]
- 21. Garcia-Paris M, Good DA, Parra-Olea G, Wake DB (2000) Biodiversity of Costa Rican salamanders: Implications of high levels of genetic differentiation and phylogeographic structure for species formation. Proceedings of the National Academy of Sciences 97: 1640–1647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Elmer KR, Davila JA, Lougheed SC (2007) Cryptic diversity and deep divergence in an upper Amazonian leaflitter frog, Eleutherodactylus ockendeni . BMC Evolutionary Biology 7: 1–55. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Streicher JW, Crawford AJ, Edwards CW (2009) Multilocus molecular phylogenetic analysis of the montane Craugastor podiciferus species complex (Anura: Craugastoridae) in Isthmian Central America. Molecular Phylogenetics and Evolution 53: 620–630. [DOI] [PubMed] [Google Scholar]
- 24. Lynch JD (1998) New species of Eleutherodactylus from the Cordillera Occidental of western Colombia with a synopsis of the distributions of species in western Colombia. Revista de la Academia Colombiana de Ciencias Exactas, Fisicas y Naturales 22: 117–148. [Google Scholar]
- 25. Bernal MH, Lynch JD (2008) Review and analysis of altitudinal distribution of the Andean anurans in Colombia. Zootaxa 1826: 1–25. [Google Scholar]
- 26.AmphibiaWeb: Information on amphibian biology and conservation. [web application]. 2012. Berkeley, California: AmphibiaWeb. Available: http://amphibiaweb.org/. (Accessed: Jan 4, 2012).
- 27. Pinto-Sánchez NR, Ibáñez R, Madriñán S, Sanjur OI, Bermingham E, et al. (2012) The great American biotic interchange in frogs: Multiple and early colonization of Central America by the South American genus Pristimantis (Anura: Craugastoridae). Molecular Phylogenetics and Evolution 62: 954–972. [DOI] [PubMed] [Google Scholar]
- 28. García-R JC, Lynch JD (2006) A new species of frog (genus Eleutherodactylus) from a cloud forest in Western Colombia. Zootaxa 1171: 39–45. [Google Scholar]
- 29. García-R JC, Castro F, Cárdenas H (2005) Relación entre la distribución de anuros y variables del hábitat en el sector La Romelia del Parque Nacional Natural Munchique (Cauca, Colombia). Caldasia 27: 299–310. [Google Scholar]
- 30. Boulenger GA (1912) Descriptions of new batrachians from the Andes of South America, preserved in the British Museum. Annals and Magazine of Natural History 8: 185–191. [Google Scholar]
- 31. Lynch JD (1981) Two new species of Eleutherodactylus from western Colombia (Amphibia: Anura: Leptodactylidae). Occasional papers of the museum of Zoology University of Michigan 697: 1–12. [Google Scholar]
- 32. Graham A (2009) The Andes: A geological overview from a biological perspective. Annals of the Missouri Botanical Garden 96: 371–385. [Google Scholar]
- 33. Moritz C (2002) Strategies to protect biological diversity and the evolutionary processes that sustain it. Systematic Biology 51: 238–254. [DOI] [PubMed] [Google Scholar]
- 34. Young BE, Lips KR, Reaser JK, Ibáñez R, Salas AW, et al. (2001) Population declines and priorities for amphibian conservation in Latin America. Conservation Biology 15: 1213–1223. [Google Scholar]
- 35. Velásquez-E BE, Castro F, Bolívar-G W, Herrera MI (2008) Infección por el hongo quitrido Batrachochytrium dendrobatidis en anuros de la Cordillera Occidental de Colombia. Herpetotropicos 4: 65–70. [Google Scholar]
- 36.Crawford AJ, Lips KR, Bermingham E (2010) Epidemic disease decimates amphibian abundance, species diversity, and evolutionary history in the highlands of central Panama. Proceedings of the National Academy of Sciences. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Vences M, Thomas M, van der Meijden A, Chiari Y, Vieites DR (2005) Comparative performance of the 16S rRNA gene in DNA barcoding of amphibians. Frontiers in Zoology 2: 5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Fouquet A, Gilles A, Vences M, Marty C, Blanc M, et al. (2007) Underestimation of species richness in Neotropical frogs revealed by mtDNA analyses. PLoS ONE 2: e1109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Beerli P, Felsenstein J (2001) Maximum likelihood estimation of a migration matrix and effective population sizes in n subpopulations by using a coalescent approach. Proceedings of the National Academy of Sciences 98: 4563–4568. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Crawford AJ (2003) Huge populations and old species of Costa Rican and Panamanian dirt frogs inferred from mitochondrial and nuclear gene sequences. Molecular Ecology 12: 2525–2540. [DOI] [PubMed] [Google Scholar]
- 41. Wang IJ, Crawford AJ, Bermingham E (2008) Phylogeography of the Pygmy Rain Frog (Pristimantis ridens) across the lowland wet forests of isthmian Central America. Molecular Phylogenetics and Evolution 47: 992–1004. [DOI] [PubMed] [Google Scholar]
- 42. Elmer KR, Davila JA, Lougheed SC (2007) Applying new inter-individual approaches to assess fine-scale population genetic diversity in a neotropical frog, Eleutherodactylus ockendeni . Heredity 99: 506–515. [DOI] [PubMed] [Google Scholar]
- 43. Kliman RM, Hey J (1993) DNA sequence variation at the period locus within and among species of the Drosophila melanogaster complex. Genetics 133: 375–387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Van der Hammen T (2000) Aspectos de historia y ecologia de la biodiversidad norandina y amazonica. Revista de la Academia Colombiana de Ciencias Exactas, Fisicas y Naturales 24: 231–245. [Google Scholar]
- 45. Gregory-Wodzicki KM (2000) Uplift history of the Central and Northern Andes: A review. Geological Society of America Bulletin 112: 1091–1105. [Google Scholar]
- 46. de Porta J (2003) La formación del istmo de Panamá. Su incidencia en Colombia. Revista de la Academia Colombiana de Ciencias Exactas, Fisicas y Naturales XXVII: 191–216. [Google Scholar]
- 47. Bernal XE, Guarnizo C, Luddecke H (2005) Geographic variation in advertisement call and genetic structure of Colostethus palmatus (Anura, Dendrobatidae) from the Colombian Andes. Herpetologica 61: 395–408. [Google Scholar]
- 48. Ovaska K (1992) Short- and long-term movements of the frog Eleutherodactylus johnstonei in Barbados, West Indies. Copeia 1992: 569–573. [Google Scholar]
- 49. Dixo M, Metzger J, Morgante J, Zamudio K (2009) Habitat fragmentation reduces genetic diversity and connectivity among toad populations in the Brazilian Atlantic coastal forest. Biological Conservation 12: 1560–1569. [Google Scholar]
- 50. Lynch JD (1999) Ranas pequeñas, la geometría de evolución y la especiación en los Andes colombianos. Revista de la Academia Colombiana de Ciencias Exactas, Fisicas y Naturales 23: 143–159. [Google Scholar]
- 51. Graham CH, Silva N, Velásquez-Tibatá J (2010) Evaluating the potential causes of range limits of birds of the Colombian Andes. Journal of Biogeography 37: 1863–1875. [Google Scholar]
- 52. Cadena CD, Kozak K, Gomez J, Parra J, McCain C, et al. (2011) Latitude, elevational climatic zonation, and speciation in New World vertebrates. Proceedings of the Royal Society B 279: 194–201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Hubbell SP (2001) The unified neutral theory of biodiversity and biogeography. Monographs in Population Biology 32: 1–375. [Google Scholar]
- 54.Kessing B, Croom H, Martin A, McIntosh C, Owen MW, et al.. (2004) The simple fool’s guide to PCR, version 1.0. Special Publication. Honolulu: Dept. of Zoology, University of Hawaii. [Google Scholar]
- 55. Folmer O, Black M, Hoeh W, Lutz R, Vrijenhoek R (1994) DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Molecular Marine Biology and Biotechnology 3: 294–299. [PubMed] [Google Scholar]
- 56. Hebert PDN, Cywinska A, Ball SL, deWaard JR (2003) Biological identifications through DNA barcodes. Proceedings of the Royal Society B 270: 313–321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Goebel AM, Donnelly JM, Atz ME (1999) PCR primers and amplification methods for 12S ribosomal DNA, the control region, cytochrome oxidase I, and cytochrome b in bufonids and other frogs, and an overview of PCR primers which have amplified DNA in amphibians successfully. Molecular Phylogenetics and Evolution 11: 163–199. [DOI] [PubMed] [Google Scholar]
- 58. Vences M, Kosuch J, Glaw F, Böhme W, Veith M (2003) Molecular phylogeny of hyperoliid treefrogs: biogeographic origin of Malagasy and Seychellean taxa and re-analysis of familial paraphyly. Journal of Zoological Systematics and Evolutionary Research 41: 205–215. [Google Scholar]
- 59. Librado P, Rozas J (2009) DnaSP v5: a software for comprehensive analysis of DNA polymorphism data. Bioinformatics 25: 1451–1452. [DOI] [PubMed] [Google Scholar]
- 60. Nei M, Li W-H (1979) Mathematical model for studying variation in terms of restriction endonucleases. Proceedings of the National Academy of Sciences USA 76: 5269–5273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. Watterson GA (1975) On the number of segregating sites in genetic models without recombination. Theoretical Population Biology 7: 256–276. [DOI] [PubMed] [Google Scholar]
- 62. Excoffier L, Laval G, Schneider S (2005) Arlequin (version 3.0): An integrated software package for population genetics data analysis. Evolutionary Bioinformatics Online 1: 47–50. [PMC free article] [PubMed] [Google Scholar]
- 63. Lynch M, Crease TJ (1990) The analysis of population survey data on DNA sequence variation. Molecular Biology and Evolution 7: 377–394. [DOI] [PubMed] [Google Scholar]
- 64. Raymond M, Rousset F (1995) An exact test for population differentiation. Evolution 49: 1280–1283. [DOI] [PubMed] [Google Scholar]
- 65. Whitlock MC (2011) G’ST and D do not replace FST . Molecular Ecology 20: 1083–1091. [DOI] [PubMed] [Google Scholar]
- 66. Bandelt H-J, Forster P, Rohl A (1999) Median-joining networks for inferring intraspecific phylogenies. Molecular Biology and Evolution 16: 37–48. [DOI] [PubMed] [Google Scholar]
- 67. Hey J, Nielsen R (2007) Integration within the Felsenstein equation for improved Markov chain Monte Carlo methods in population genetics. Proceedings of the National Academy of Sciences 104: 2785–2790. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68. Crawford AJ (2003) Relative rates of nucleotide substitution in frogs. Journal of Molecular Evolution 57: 636–641. [DOI] [PubMed] [Google Scholar]
- 69.Vences M, Nagy ZT, Sonet G, Verheyen E (2012) DNA barcoding amphibians and reptiles. In: Kress WJ, Erickson DL, editors: Humana Press. 79–107. [DOI] [PubMed] [Google Scholar]
- 70. Vences M, Thomas M, Bonett RM, Vieites DR (2005) Deciphering amphibian diversity through DNA barcoding: Chances and challenges. Philosophical Transactions of The Royal Society B 360: 1859–1868. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71. Ingman M, Kaessmann H, Paabo S, Gyllensten U (2000) Mitochondrial genome variation and the origin of modern humans. Science 408: 708–713. [DOI] [PubMed] [Google Scholar]
- 72. Kimura M (1969) The number of heterozygous nucleotide sites maintained in a finite population due to a steady flux of mutations. Genetics 61: 893–903. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73. Tajima F (1989) Statistical method for testing the neutral mutation hypothesis by DNA polymorphism. Genetics 123: 585–595. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Posterior probability distributions for gene flow from IMa analysis.
(TIF)
Posterior probability distributions for effective population sizes from IMa analysis.
(TIF)
Posterior probability distributions for divergence times from IMa analysis.
(TIF)
Sample information for taxa analyzed in this study.
(DOC)