

Summary
The Arabidopsis guard cell anion channel SLAC1 is essential for stomatal closure in response to various endogenous and environmental stimuli. Interestingly, here we reveal an unexpected impairment of slac1 alleles on stomatal opening.
We report that mutations in SLAC1 unexpectedly slow stomatal opening induced by light, low CO2 and elevated air humidity in intact plants and that this is caused by the severely reduced activity of inward K+ (K+in) channels in slac1 guard cells.
Expression of channels and transporters involved in stomatal opening showed small, but significant reductions in transcript levels in slac1 guard cells, however this was deemed insufficient to explain the severely impaired K+in channel activity in slac1. We further examined resting cytosolic [Ca2+]cyt levels and K+in channel sensitivity to cytosolic [Ca2+]cyt in slac1. These experiments showed higher resting [Ca2+]cyt in slac1 guard cells and that reducing [Ca2+]cyt to <10 nM rapidly restored the activity of K+in channels in slac1 closer to wild type levels.
These findings demonstrate an unanticipated compensatory feedback control in plant stomatal regulation, that counteracts the impaired stomatal closing response of slac1, by down-regulating stomatal opening mechanisms and implicates enhanced [Ca2+]cyt sensitivity priming as a mechanistic basis for the down-regulated K+in channel activity.
Keywords: abscisic acid (ABA), anion channel, Ca2+ signaling, cytosolic Ca2+ concentration, guard cells, potassium uptake channel, SLAC1, stomatal opening
Introduction
Each stomatal pore in the epidermis of aerial plant tissues is formed by a pair of guard cells. They enable carbon dioxide (CO2) uptake into leaves for photosynthesis and control transpirational water loss to the atmosphere. The aperture of stomatal pores is regulated by guard cell turgor pressure adjusted by modification of guard cell ion and organic solute content (Raschke et al., 1988). Stomata respond to various environmental and endogenous factors, such as light, air humidity, CO2 concentration, abscisic acid (ABA) and cytosolic Ca2+. Elevation of cytosolic Ca2+ functions in ABA-, and CO2-induced stomatal closing (McAinsh et al., 1990; Webb et al., 1996; Grabov & Blatt, 1998; Staxen et al., 1999; MacRobbie, 2000; Allen et al., 2001; Marten et al., 2007a; Young et al., 2006; Siegel et al., 2009; Hubbard et al., 2012).
Stomatal opening is driven by the guard cell plasma membrane H+-ATPase, which pumps protons from the guard cell cytosol to the cell wall and creates an electrochemical gradient across the plasma membrane (Assmann et al., 1985; Shimazaki et al., 1986; Thiel et al., 1992; for review see Shimazaki et al., 2007). Membrane hyperpolarization triggers the activation of voltage-gated potassium uptake channels (K+in) that mediate the uptake of K+ ions (Schroeder et al., 1984). Guard cell K+in channels function as multimers and single channel gene knockouts do not have apparent stomatal and K+in channel activity phenotypes (Szyroki et al., 2001; Very & Sentenac 2003). However, over-expression of a dominant negative K+in channel mutant (Uozumi et al., 1995) resulted in a slowing and decrease in light-induced stomatal opening by c. 45% (Kwak et al., 2001). Furthermore, even stronger repression of K+in channels in kincless, which is a knockout of KAT2 transformed with a dominant negative version of KAT2, resulted in substantially reduced light-induced stomatal opening (Lebaudy et al., 2008). The activity of K+in channels in guard cells is rapidly down-regulated by [Ca2+]cyt elevation in guard cells (Schroeder and Hagiwara, 1989; Kelly et al., 1995; Grabov and Blatt, 1999). Recent research has shown that ABA enhances the [Ca2+]cyt sensitivity of K+in channel down-regulation in guard cells (Siegel et al., 2009), which is in-line with ABA inhibition of stomatal closing.
When plants are subjected to factors causing stomatal closure, slow (S-type) and rapid (R-type) type anion channels are activated (Keller et al., 1989; Schroeder and Hagiwara, 1989; Pei et al., 1997; for review see Barbier-Brygoo et al., 2011; Kollist et al., 2011). Efflux of anions leads to depolarization of the plasma membrane, which in turn activates K+ efflux channels (Schroeder et al., 1987; Blatt and Armstrong, 1993; Roelfsema et al., 2001). A screen for Arabidopsis ozone sensitive mutants (Overmyer et al., 2000; Overmyer et al., 2008, Vahisalu et al., 2008) and CO2-insensitive mutants (Negi et al., 2008) led to the identification of SLAC1, a protein essential for guard cell plasma membrane S-type anion channel function (Vahisalu et al., 2008). SLAC1 is required for stomatal closure induced by ozone, CO2, ABA, calcium, light/dark transitions and reduction in air humidity (Negi et al., 2008; Vahisalu et al., 2008). Mutations in SLAC1 impaired S-type anion channel function, but R-type anion channels and ABA-activated Ca2+-permeable channel activity was not altered (Vahisalu et al., 2008). Furthermore, guard cell protoplasts of slac1 over-accumulated of osmoregulatory ions, such as malate, fumarate, Cl- and K+ (Negi et al., 2008). The plasma membrane localization of SLAC1 and the homology of SLAC1 to anion transporters (Camarasa et al., 2001) suggested that SLAC1 encodes guard cell S-type anion channel (Vahisalu et al., 2008). Heterologous expression of SLAC1 in Xenopus oocytes induced S-type anion currents similar to those observed in guard cells confirming this model (Geiger et al., 2009; Lee et al., 2009).
During stomatal opening, SLAC1 must be inactivated (Schroeder et al., 1993; Schwarz et al., 1995; Marten et al., 2007). This implies that stomatal opening should be faster in slac1 plants. However, in complete contrast to this prediction, we found that slac1 exhibited a reduced rate of stomatal opening in response to three physiological stimuli of stomatal opening: light, low CO2 and high humidity in intact plants. Interestingly, although SLAC1 encodes for S-type anion channels, we demonstrate that slac1 alleles exhibit strongly impaired K+in channel activities, which provides a mechanistic basis for slower stomatal opening in slac1 mutants. Furthermore, the dramatic reduction in K+in channel currents in slac1 guard cells is shown to be rapidly reversed upon lowering the cytosolic free Ca2+ concentration, showing an enhanced [Ca2+]cyt sensitivity of K+in channels in slac1.
These findings imply that eliminating SLAC1 function (Vahisalu et al., 2008; Negi et al., 2008) unexpectedly affects the activities of other membrane proteins essential for mediation of stomatal opening, and they point to compensatory mechanisms that counteract transpirational water loss.
Materials and Methods
Plant growth and gas exchange
Arabidopsis thaliana (L.) Heynh (Col-0 as wild-type) seeds were planted in soil containing 4:3 (v:v) peat:vermiculite and grown through a hole in a glass plate as described (Kollist et al., 2007). Soil moisture was kept at 60-80% of maximum water capacity. Plants were grown in growth chambers (AR-66LX and AR-22L, Percival Scientific, IA, USA) at 12 h : 12 h photoperiod and 150 μmol m-2 s-1 light. Temperature was 23°C in the light and 18°C in the dark, relative humidity of the air was 70-80%. For experiments 23-26 d old plants were used.
Stomatal conductance was measured in intact plants using a rapid-response gas exchange measurement device consisting of eight through-flow whole-rosette cuvettes (Kollist et al., 2007). Inlet air composition was monitored and adjusted to desired values. Air humidity and CO2 concentration were decreased by filtering air through a granular potassium hydroxide. Humidity was increased by a humidifying thermostat system and CO2 was added to inlet air as necessary. Light conditions were changed manually by covering plants from the light source. Stomatal conductance measurements for air humidity and CO2 treatments were carried out during 11:00-15:00 h; light treatment was applied at the onset of regular light regime (07:30 h).
Guard cell protoplast extraction and RT-PCR
Guard cells from plants grown as described above were isolated using the protocol by Leonhardt et al. (2004). Plant RNA was extracted using the RNeasy Plant Mini Kit and converted to cDNA using QuantiTect Reverse Transcription Kit (both from Qiagen GmbH, Germany). 7900HT Fast Real-Time PCR System was used with MicroAmpTM Optical 384-well Reaction Plate (Applied Biosystems, Singapore) and MicroAmpTM Optical Adhesive Film (Applied Biosystems, USA).
For qPCR reactions, the Maxima™ SYBR Green/ROX qPCR Master Mix (2X) (Fermentas) was used. The TIP41-like family protein (At4g34270; (Czechowski et al., 2005) was used as normalization standard. Primers used for RT-PCR are listed in Supporting Information Table S1. The following genes were analysed: Guard cell expressed H+ATPase genes AHA1 (At2g18960), AHA2 (At4g30190), and AHA5 (At2g24520) (Ueno et al., 2005). The inward rectifying K+ channels KAT1 (At5g46240) (Nakamura et al., 1995), KAT2 (At4g18290) (Pilot et al., 2001), AKT2 (At4g22200) (Cao et al., 1995), KC1 (At4g32650) (Reintanz et al., 2002) and K+ outflow channel GORK (At5g37500) (Ache et al., 2000). We also measured expression levels of AtABCB14 (At1g28010) which mediates malate uptake (Lee et al., 2008), aluminum activated malate transporter, ALMT12 (Meyer et al., 2010; Sasaki et al., 2010) and TPC1 (At4g03560), a vacuolar Ca2+ channel required for Ca2+-inhibition of stomatal opening (Peiter et al., 2005).
Patch clamping
Arabidopsis thaliana plants (Columbia ecotype) were grown in soil (Sungro Special blend Professional Growing Mix; Seba Beach, Alberta, Canada) in a growth chamber (Conviron, Manitoba, Canada) under a 16-h-light : 8-h-dark cycle at a photon fluence rate of 75 μmol m-2 s-1 and a temperature of 20-22°C with ≥70% humidity described previously (Mori et al., 2006). Arabidopsis guard cell protoplasts were isolated enzymatically as described previously (Vahisalu et al., 2008). To analyze inward potassium channel currents at physiological [Ca2+]cyt, the bath solution contained 30 mM KCl, 1 mM CaCl2, 2 mM MgCl2, 10 mM Mes-Tris, pH 5.5, pipette solution contained 30 mM KCl, 70 mM K-Glu, 2 mM MgCl2, 6.7 mM EGTA, 10 mM HEPES-Tris, pH 7.1, and 3.35 mM CaCl2 was added to result in 250 nM free Ca2+ as described previously (Pei et al., 1997). Inward potassium currents were recorded within 2 to 3 min after accessing whole-cell configurations and for continued durations >5 min. Thus the pipette solution that dialyzes the cytoplasm (Pusch & Neher, 1988) enabled the cytoplasmic free Ca2+ concentration to reach a stable state. To analyze inward potassium channel currents at low free [Ca2+]cyt, no Ca2+ was added to the pipette solution, other components in the pipette solution remained unchanged, resulting in a calculated free Ca2+ concentration of <10 nM. The same bath solution was used for all conditions. The osmolarities of the bath and pipette solutions for inward potassium current recordings were adjusted with D-sorbitol to 485 mM and 500 mM, respectively (Pei et al., 1997). To analyze outward malate currents, the bath solution contained 30 mmol/L Cs-Malate, 1 mM CaCl2, 2 mM MgCl2, 10 mM Mes-Tris, pH 5.6, and the pipette solution contained 150 mM Cs-Malate 2 mM MgCl2, 10 mM Hepes-Tris, pH 7.05, plus 6.67 mM EGTA and 5.86 mM CaCl2 to result in 2 μM free Ca2+. The osmolarities of bath and pipette solutions were adjusted with D-Sorbitol to 485 mM and 500 mM, respectively. For malate current analyses, guard cell protoplasts were extracellularly pre-incubated for 30 min in the same bath solution with 40 mM CaCl2 added before patch clamping, the CaCl2 concentration in the bath solution was reduced from 40 mM to 1 mM by perfusion before patch clamping, and whole-cell recordings with giga seals (>10 GΩ) were achieved within 30 min after the pre-incubation as described previously (Vahisalu et al., 2008).
Ratiometric quantitation of cytosolic Ca2+ with yellow cameleon 3.6
Cytosolic Ca2+ concentration changes were monitored using the non-invasive FRET Ca2+ reporter yellow cameleon. 3.6 as previously described (Mori et al., 2006). Intact leaf epidermes adhered to a cover slip were incubated 3 h in stomatal opening buffer (50 mM KCl, 10 mM MES-Tris (pH 5.6), 0 mM added CaCl2) for 3 h in 200 μmol m-2s-1 white light and baseline cytosolic Ca2+ concentrations were monitored for 0.5 h in the same stomatal opening buffer in > 30 wild-type and >30 slac1 guard cells, as indicated.
ABA-induced stomatal closure
Stomatal measurements were performed by excising c. 4 wk old rosette leaves. Intact leaves were glued abaxial side down onto cover slips using Hollister medical adhesive and mesophyll cells were removed from the intact epidermis using single edge industrial razor blades (Young et al., 2006). The glued epidermes were submerged in stomatal opening buffer (10 mM KCl, 7.5 mM iminodiacetic acid, 10 mM MES and pH 6.2 adjusted with KOH) for 3 h in light (100 μmol m-2 s-1) and then incubated for an additional 3 h or 1 h in stomatal opening buffer in which the indicated ABA concentrations had been added. Stomatal apertures (30 stomata per treatment per experiment) were then measured on a light microscope (Nikon Diaphot 300) via scion image. Measurements were performed as double blind in which genotype and ABA concentrations were not known to the experimenter.
Results
Mutations in SLAC1 anion channel slows high humidity-, light- and low CO2-induced stomatal opening
In initial experiments, we observed an unexpected slowing of stomatal opening when plants kept in darkness for 2 h were exposed to 130 μmol m-2 s-1 white light (Fig. S1a). As in these experiments there were differences in pre-treatment stomatal conductances between wild-type and slac1 mutant alleles (Fig. S1b), we sought for conditions that produce similar steady-state starting stomatal conductances for further investigations of stimulus-induced stomatal opening. We found that by adapting plants initially to low air humidity (water vapor concentration difference between leaf and air 23.5 ± 0.5 mmol mol-1, relative air humidity in cuvette 17.9 ± 0.7%) for 2-3 h the slac1-1 and slac1-3 and wild-type plants equilibrated at similar stomatal conductances. To analyze humidity-induced stomatal opening air humidity was increased (water vapor concentration difference between leaf and air 14.9 ± 0.3 mmol mol-1, relative air humidity in cuvette 47.3 ± 0.6%) while keeping CO2 and light conditions constant. Such treatments induced first a sudden decline in stomatal conductance (Fig. 1a,b), which has been proposed to be caused by a rapid decrease of water efflux from epidermal cells and concomitant increase of their pressure on guard cells (Ivanoff, 1928). This was followed by a rapid increase in stomatal opening in both wild type and in slac1 mutants. However, the initial rates of stomatal conductance changes were unexpectedly faster in wild type than in slac1 mutants (at 12 min 0.74 ± 0.19, 0.21 ± 0.07 and 0.15 ± 0.05 mmol m-2 s-1 per min for wild type, slac1-1 and slac1-3, respectively, P < 0.01).
Fig. 1.
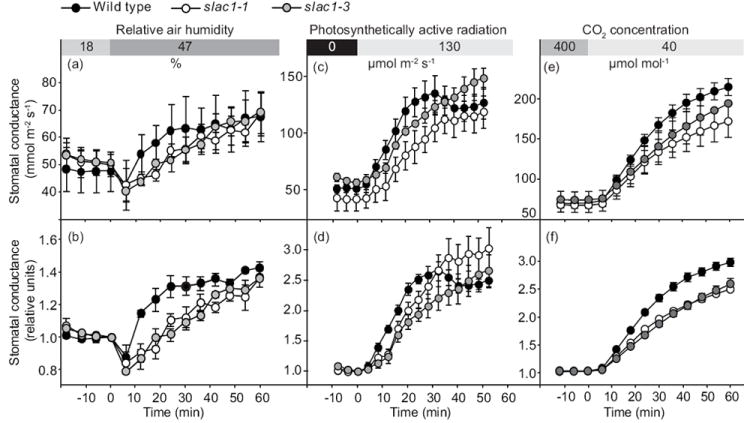
Humidity-, light- and low CO2-induced increase in whole-plant stomatal conductance is reduced in Arabidopsis slac1 mutants. (Wild type, black circles; slac1-1, white circles; slac1-3, grey circles.) (a, b) Wild type, slac1-1, and slac1-3 plants were kept in low relative air humidity (17.9 ± 0.7%) for 120–180 min, thereafter humidity was abruptly increased to 47.3 ± 0.6% at time 0 as indicated in the upper panel. Error bars indicate ±SEM, n = 3. (c, d) Stomatal conductance patterns of wild type, slac1-1, and slac1-3 plants kept in darkness overnight (12 h) and then exposed to 130 μmol m-2 s-1 light as indicated in upper panel are shown. Error bars indicate ±SEM, n = 4. (e, f) To address low CO2-induced stomatal opening of wild type, slac1-1, and slac1-3 plants were kept in 400 μmol mol-1 of CO2 and 20-25% relative air humidity until stomatal conductance values had stabilized (2-3 h), thereafter CO2 concentration was decreased to 40 μmol mol-1 at time 0 (±SEM, n = 4-8). Patterns of absolute (a, c, e) as well as relative stomatal conductance normalized to value at 0 time point (b, d, f) are shown.
To address light–induced stomatal opening, plants were kept in gas-exchange cuvettes over regular night (12 h) period. In the morning slac1 and wild-type plants had similar stomatal conductance. After onset of light, conductance began to increase both in wild-type and in slac1 mutants (Fig. 1c,d). However, the initial rates of increases in stomatal conductance were significantly slower in slac1 mutants than in wild-type (at 12 min 2.9 ± 0.4, 0.9 ± 0.3 and 1.1 ± 0.4 mmol m-2 s-1 per min, in wild-type, slac1-1 and slac1-3, respectively, P < 0.01). After 30 min in white light, the increase in stomatal conductance in wild type plants ceased and even slightly declined until it reached a steady state. In slac1 only a gradual slower rate of stomatal opening was observed without the subsequent slight closure.
To study the stomatal responsiveness of slac1 to low CO2, we first acclimatized plants under reduced humidity for 2-3 h to achieve similar stomatal conductances of slac1 mutants and wild-type plants. Thereafter CO2 was reduced from 400 to 40 μmol mol-1. Such treatment caused a rapid, three-fold increase in stomatal conductance in all genotypes (Fig. 1e,f). Still, the initial rate of stomatal opening was clearly slower in slac1 mutants (at 12 min 1.5 ± 0.1 and 1.5 ± 0.2 mmol m-2 s-1 per min in slac1-1 and slac1-3, compared to 2.4 ± 0.2 in wild-type, P < 0.01).
Earlier experiments have shown that slac1 plants are largely insensitive to high CO2-induced stomatal closure (Negi et al., 2008; Vahisalu et al., 2008). To research whether elevation of CO2 from 100 μmol mol-1 to 400 μmol mol-1 will induce stomatal closure in slac1, plants were acclimatized under low CO2 for 3 h followed by an increase of CO2 to 400 μmol mol-1. This caused clear, though significantly slower, stomatal closure also in slac1 mutants (Fig. S2). Also stepwise reduction of CO2 concentration from 400 to 0 μmol mol-1 followed by increase to 800 μmol mol-1 indicated that slac1 had a clear, though reduced, stomatal response to CO2 changes (Fig. S3). These results are in agreement with the findings that SLAC1 is not the only anion channel in guard cells, as R-type anion channels and SLAH3 remain intact in slac1 (Vahisalu et al., 2008; Geiger et al., 2011).
slac1 guard cells exhibit dramatically reduced K+in channel currents
Inward K+ (K+in) channels provide a pathway contributing to K+ uptake during stomatal opening driven by H+-ATPases in guard cells (Schroeder et al., 1984; Schroeder et al., 1987; Thiel et al., 1992; Kwak et al., 2001; Lebaudy et al., 2008). In parallel independent research, we analyzed K+in channel currents by whole-cell patch clamping. These measurements showed dramatically reduced K+in channel activity in slac1-1 and slac1-3 guard cells compared to wild type (Fig. 2), indicating that the disruption of S-type anion channel function in slac1 unexpectedly also greatly reduced the activity of K+in channels. In slac1-1 and slac1-3 the reduction (by 84% and 79%, respectively) in K+in channel current magnitude of slac1 alleles at -180 mV was similar to low K+in current magnitudes that caused slow stomatal opening when K+in channels were blocked or genetically impaired (Kelly et al., 1995; Kwak et al., 2001; Lebaudy et al., 2008). Next we investigated possible mechanisms causing the dramatic reduction in K+in channel activity in slac1 alleles.
Fig. 2.
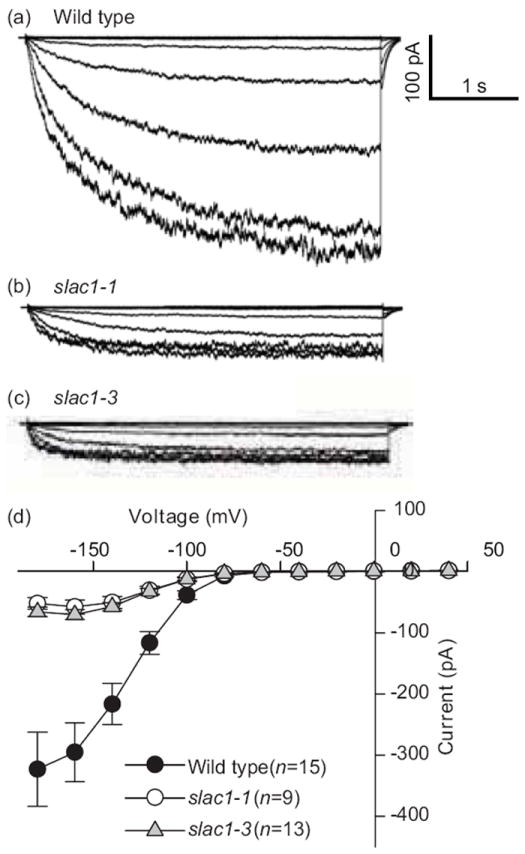
Impaired SLAC1 leads to reduced K+in channel activities in Arabidopsis guard cells. Whole-cell recordings of inward K+ currents in the presence of 30 mM KCl in the bath solution in (a) wild-type, (b) slac1-1, and (c) slac1-3 guard cells. (d) Average current–voltage relationships. K+in channel currents were activated by voltage pulses with +20 mV increment from -180 mV to +40 mV. Error bars indicate ±SEM.
Mutations in SLAC1 cause partly reduced expression levels of ion channels in guard cells
We investigated whether the reduced K+in channel activity in slac1 could be a result of reduced K+in channel subunit mRNA levels, and/or whether impaired SLAC1 has affected the expression of other known transporters expressed in guard cells (Leonhardt et al., 2004; Yang et al., 2008) involved in stomatal opening. We analyzed mRNA levels of several guard cell ion channels and transporters in slac1 and wild type isolated guard cells, including AHA1, AHA2, AHA5, KAT1, KAT2, AKT1, AKT2, AtABCB14, GORK, TPC1, AtALMT12 and AtKC1 (For details see Table S1). Transcript levels of several guard cell-expressed transporter genes were reduced in slac1 guard cells in four independent experiments (Fig. 3). The three guard cell-expressed H+-ATPase transcripts showed slight average reductions in slac1 mutants guard cells, albeit at a 90% statistical confidence level only for AHA1 and AHA2 in slac1-3. Also the transcript levels of guard cell malate importer AtABCB14 which was reported to limit light and CO2-induced stomatal responses (Lee et al., 2009) were reduced in slac1-1. Moreover, KAT2 transcript level was significantly reduced in both slac1 alleles at a 95% statistical confidence level (Fig. 3). However, we concluded that the observed differences in ion channel transcript levels were deemed unlikely to cause the severely reduced K+in channel activity in slac1 mutants.
Fig. 3.
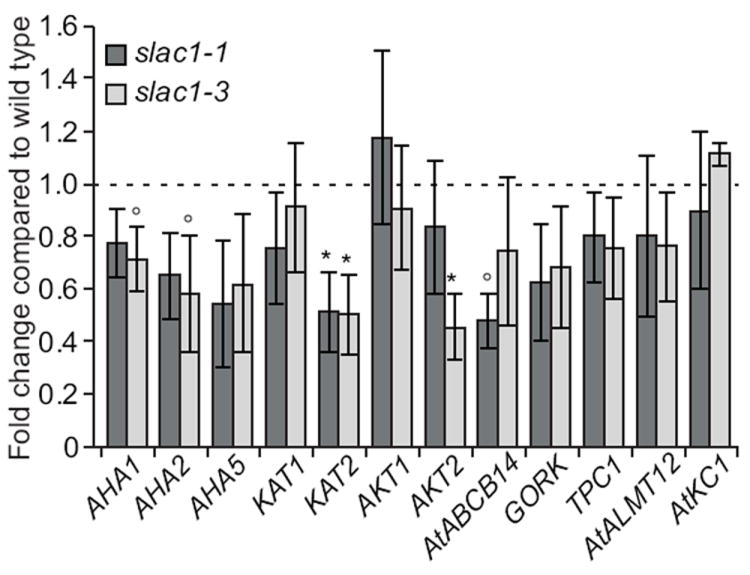
Mutations in SLAC1 suppress expression of several guard cell ion channel and transporter genes. Relative expression of Arabidopsis plasma membrane H+-ATPases AHA1, AHA2, AHA5, transporter AtABCB14 and ion channels KAT1, KAT2, AKT1, AKT2, TPC1, GORK, AtALMT12, and AtKC1 was measured in isolated guard cells of wild type, slac1-1 (dark bars), and slac1-3 (light bars). Error bars indicate ±SEM, n = 4 (*, P < 0.05; °, P < 0.10; One-way ANOVA and LSD test).
Decreasing the cytosolic Ca2+ concentration rapidly restores K+in channel currents in slac1 guard cells
Elevation in the cytosolic Ca2+ is known to activate S-type anion channels in guard cells (Schroeder & Hagiwara, 1989; Mori et al., 2006; Vahisalu et al., 2008; Siegel et al., 2009; Chen et al., 2010). Furthermore, [Ca2+]cyt elevation is also known to down-regulate K+in channel activity, as a mechanism contributing to inhibition of stomatal opening (Schroeder & Hagiwara, 1989; Kelly et al., 1995; Grabov & Blatt, 1999; Siegel et al., 2009). To explore whether cytosolic Ca2+ plays a role in the substantially reduced K+in channel activity of slac1 guard cells, patch clamp experiments were performed in which the free Ca2+ concentration in the patch clamp pipette solution was decreased from 250 nM to <10 nM by adding 6.7 mM EGTA and no Ca2+ to the pipette solution (see Methods). Interestingly, recording of K+in channel currents only 2 to 3 min after reducing [Ca2+]cyt to <10 nM by whole-cell patch clamp access to the cytoplasm of guard cells (Hamill et al., 1981) resulted in markedly increased K+in channel current magnitudes in slac1 mutant guard cells. At <10 nM free [Ca2+]cyt in the pipette solution that dialyzes the cytoplasm of guard cells, K+in channel currents were only c. 20% smaller than in wild type guard cells (Fig. 4a,b). This lay in stark contrast to experiments in which [Ca2+]cyt was buffered to higher levels (250 nM) in the pipette solution (Fig. 2). These results showed that K+in channel currents could be rescued by lowering [Ca2+]cyt to an unphysiologically low concentration of 10 nM, thus revealing an enhanced sensitivity of K+in channels to [Ca2+]cyt in slac1 guard cells. To determine whether resting cytosolic Ca2+ levels are higher in slac1 than in wild-type guard cells, we used plants transformed with the non-invasive yellow cameleon construct 3.6 (YC3.6) (Mori et al., 2006; Siegel et al., 2009; Vahisalu et al., 2008). Analysis of YC3.6 transformed wild-type guard cells, showed that average baseline fluorescence ratio in wild-type guard cells was 1.71 ± 0.10 SD (± 0.02 SEM, n = 35 guard cells). The average baseline fluorescence ratio of slac1 guard cells was 1.79 ± 0.1 SD (± 0.01 SEM, n = 39 slac1 guard cells). Note that we compared baseline FRET ratios of the cameleon Ca2+ reporter, rather than attempt calibration of free [Ca2+]cyt ratios. Calibrations usually do not calibrate each individual cell and therefore introduce errors with any Ca2+ reporter. Therefore comparative [Ca2+]cyt imaging analyses often report unbiased raw [Ca2+]cyt ratios (Siegel et al., 2009), as analyzed here. Thus analyses of a large number of wild-type and slac1 guard cells indicated that slac1 guard cells had slightly but statistically significant (P < 0.001)higher baseline [Ca2+]cyt ratios (Fig. 4c), which suggests that slac1 mutants guard cells have an elevated cytosolic Ca2+ concentration.
Fig. 4.
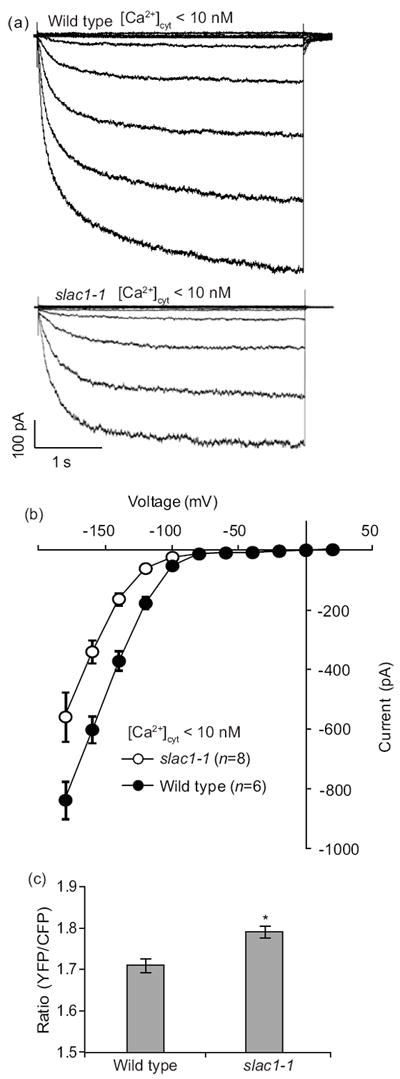
Decrease of intracellular Ca2+ concentration rapidly restores K+in channel activity and cameleon Ca2+ reporter analyses indicates slightly elevated cytosolic Ca2+ in Arabidopsis slac1 mutant guard cells. (a) Whole-cell recordings of K+in currents with <10 nM free [Ca2+]cyt in the pipette solution in slac1-1 and in wild type guard cells. (b) Average steady state current–voltage relationships for guard cells, as recorded in (a). (c) Baseline cytosolic Ca2+-dependent ratiometric fluorescence levels in wild type (n = 35) and slac1-1 (n = 39) guard cells transformed with yellow cameleon 3.6; *, P < 0.001. Error bars indicate ±SEM.
Prolonged treatment with ABA induces partial stomatal closure in slac1 mutants
We have previously shown that ABA-induced stomatal closure was strongly impaired in slac1 when up to 10 μM ABA was applied for 40 min (Vahisalu et al., 2008). Since R-type anion channels remained functional in slac1 guard cells (Vahisalu et al., 2008), we analyzed slac1 stomatal responses to higher ABA concentrations and longer exposure times. This demonstrated that application of 50 μM ABA for 3 h induced a clear but partial stomatal response in slac1 (Fig. 5a). To exclude that this result was due to changes in experimental conditions, we repeated earlier experiments with application of a more physiological lower ABA concentration and analyzed short-term responses (Fig. 5b). In line with previous findings (Vahisalu et al., 2008), these conditions did not induce stomatal closure in slac1 (Fig. 5b). Together, these data reinforce the observation that both S-type and R-type anion channels in guard cells contribute to stomatal closure, and suggest that the S-type anion channel is important for fast responses as also indicated by the lack of rapid ozone/ROS responses in slac1 (Vahisalu et al., 2010).
Fig. 5.
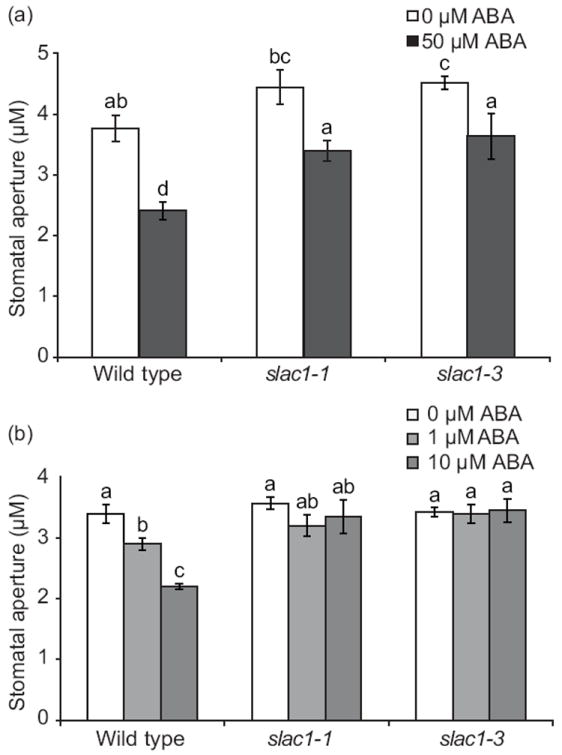
Partial abscisic acid (ABA)-induced stomatal closure in Arabidopsis slac1 mutants depends on ABA concentration and duration of ABA exposure. (a) Leaves of wild type, slac1-1, and slac1-3 were incubated in 0 μM ABA or 50 μM ABA for 3 h (±SEM, n = 20-33). (b) Impaired ABA-induced stomatal response in slac1-1 and slac1-3 mutants, at lower ABA concentrations (1 and 10 μM ABA) and reduced exposure time to ABA (1 h). ABA-induced stomatal closure in wild type, slac1-1, and slac1-3, intact leaf epidermis treated with the indicated ABA concentrations for 1 h (±SEM, n = 3 experiments, 30 stomata per condition per experiment). Experiments were conducted as genotype and [ABA] blind experiments. Letters refer to significant (P < 0.05) differences (ANOVA and LSD test).
Given that, in contrast to wild type, slac1 guard cells retain high intracellular organic anion concentrations, including malate (Negi et al., 2008), we analyzed whether S-type anion channels can mediate malate efflux from Arabidopsis guard cells. Patch clamp studies with malate in the pipette solution that dialyzes the cytoplasm of guard cells did not display clear anion efflux currents (outward currents), suggesting that S-type anion channels in Arabidopsis guard cells are not highly permeable to malate anions (Fig. S4). These results correlate with a report that S-type anion currents are suppressed by malate (Wang & Blatt, 2011). Thus the high malate and fumarate content of slac1 guard cells (Negi et al., 2008) may also result from an influence of impaired SLAC1 function on additional transport processes in guard cells.
Discussion
The guard cell plasma membrane S-type anion channel SLAC1 is required for stomatal closure in responses to multiple environmental and endogenous stimuli (Negi et al., 2008; Vahisalu et al., 2008). slac1 guard cells exhibit greatly reduced S-type anion channel function, whereas R-type anion channels and ABA-activated Ca2+-permeable channels remained functional (Vahisalu et al., 2008). These data illustrate the rate-limiting role of S-type anion channels for the onset of stomatal closure (Schroeder & Hagiwara, 1989; Schroeder et al., 1993; Kim et al., 2010; Kollist et al., 2011). Stomatal opening, on the contrary, requires accumulation of anions and thus S-type anion channels are inactivated (Schwartz et al., 1995; Marten et al., 2007). This implies that the absence of SLAC1 function might result in faster stomatal opening. However, our experiments show the opposite, the impaired SLAC1 activity in two independent slac1 mutant alleles clearly reduced the rate of stomatal opening in response to three distinct stimuli in intact plants: light, low CO2 and an increase in air humidity (Fig. 1). As SLAC1 mediates anion efflux, it is unlikely that impairment in SLAC1 activity would directly be responsible for the slow stomatal opening phenotype observed in slac1. Furthermore, humidity, light and CO2 induce stomatal opening by different signaling pathways (Roelfsema & Hedrich, 2005; Shimazaki et al., 2007), which suggests that a basic property of stomatal opening and not only a single signaling pathway is affected in slac1. The basic property, which is the same for light-, CO2- and humidity-induced stomatal opening, could be the uptake of ions leading to water inflow and swelling of guard cells.
Down-regulation of guard cell potassium uptake channel activity by enhanced [Ca2+]cyt sensitivity in SLAC1 mutants
We found a dramatic reduction of K+in channel activity in slac1 (Fig. 2). Based on previous knock-down and pharmacological experiments, blocking of K+in channel activity by 80% or more in guard cells, causes a slowing in the rate of stomatal opening (Kelly et al., 1995; Kwak et al., 2001; Lebaudy et al., 2008). Therefore, the observed reduction (≈ 80%) in K+in channel activity in slac1 alleles (Fig. 2) provides a functional basis that can cause slowed stomatal opening responses to light, low CO2 and high humidity (Fig. 1).
We also analyzed K+in channel and other ion transporter transcript levels which revealed decreased expression of KAT2, AKT2, AtABCB14, AHA1 and AHA2 (Fig. 3). However, the decreases in transcript levels were modest and are unlikely to fully explain the large reduction of K+in currents. Further exploration of the reduction in K+in channel activity in slac1 guard cells (Fig. 2) revealed that the K+in current activities were rapidly restored in slac1 guard cells by lowering [Ca2+]cyt to below physiological resting levels of <10 nM (Fig. 4). A recent study showed that ABA enhances the [Ca2+]cyt sensitivity of K+in channel down-regulation in Arabidopsis guard cells demonstrating that the [Ca2+]cyt sensitivity of K+in channels is tunable (Siegel et al., 2009). This stimulus-induced enhancement of the [Ca2+]cyt sensitivity of guard cell signaling mechanisms has been termed ‘Ca2+ sensitivity priming’ (Young et al., 2006; Siegel et al., 2009; Xue et al., 2011). The present results suggest that an impaired SLAC1 channel may similarly enhance the Ca2+ sensitivity of K+in channel, such that free [Ca2+]cyt levels of 250 nM dramatically down-regulate K+in channel activity (Fig. 2). This response could provide an adaptive response of guard cells to counteract the tendency of slac1 mutant stomata to open too widely.
Ca2+ imaging analyses showed that slac1 guard cells have a slightly elevated cytosolic [Ca2+]cyt concentration (Fig. 4c). The impairment in S-type anion channel activity in slac1 guard cells (Vahisalu et al., 2008), is predicted to cause more negative membrane potentials. This in turn can increase the activation of hyperpolarization-activated Ca2+ permeable ICa channels in guard cells (Pei et al., 2000; Hamilton et al., 2000), which may provide a basis for the observed elevation in baseline [Ca2+]cyt in slac1 guard cells. Although the increase in [Ca2+]cyt was statistically significant, more research would be needed to determine the relative contribution of this elevation to the slowed stomatal opening, as K+in channels also exhibited an enhanced sensitivity to [Ca2+]cyt per se. Previous studies have shown that exposure of guard cells to elevated extracellular Ca2+ concentrations primes guard cells to respond to cytosolic Ca2+ (Allen et al., 2002; Mori et al., 2006). Thus the elevated baseline [Ca2+]cyt concentrations in slac1 guard cells (Fig. 4c), may contribute to the constitutively primed (enhanced) Ca2+ sensitivity of K+in channels in slac1 guard cells (Figs 2, 4).
Intracellular Ca2+ elevation regulates several key mechanisms and ion channels that function in stomatal closing (Schroeder & Hagiwara, 1989; McAinsh et al., 1990; Hedrich et al., 1990; Webb et al., 1996; MacRobbie, 2000; Siegel et al., 2009). The rapid “Ca2+-reactive” phase of stomatal closing is mediated by [Ca2+]cyt activation of SLAC1 channels (Vahisalu et al., 2008) and has been shown to be independent of the [Ca2+]cyt oscillation frequency, in contrast to the long-term ‘Ca2+-programmed’ phase (inhibition of stomatal re-opening after initial closure) (Allen et al., 2001). The rapid initial Ca2+-reactive phase of stomatal closing (Allen et al., 2001) has been shown to include ABA- and CO2-induced enhancement (priming) of the [Ca2+]cyt sensitivity of S-type anion channel activation (Siegel et al., 2009; Chen et al., 2010; Xue et al., 2011) and enhancement in the [Ca2+]cyt sensitivity of K+in channel down-regulation (Siegel et al., 2009).
Furthermore, knock-out of SLAC1 can be predicted to cause more negative guard cell membrane potentials, based on the lack of the major depolarizing SLAC1 anion efflux activity (Schroeder & Hagiwara, 1989), which would drive enhanced K+ influx and more rapid stomatal opening. To counter these effects, an enhanced Ca2+ sensitivity of K+in channels would provide a mechanism to at least partially counteract the detrimental effects of slac1 mutation. The unexpected strongly reduced K+in channel activity previously found in abi2-1 guard cells also correlates with the much wider stomatal apertures in abi2-1 guard cells under the imposed conditions (Pei et al., 1997). Thus the present study suggests that guard cells adapt to mutations through compensatory feedback Ca2+ signaling mechanisms.
Recent studies indicate a direct link between regulation mechanisms of SLAC1 and the K+ uptake channel KAT1. Phosphorylation of SLAC1 by OST1 and by Ca2+ dependent protein kinases can activate S-type anion currents in guard cells and SLAC1-dependent anion currents in oocytes (Geiger et al., 2009, 2010; Lee et al., 2009; Brandt et al., 2012) add: Mori et al., 2006). At the same time OST1-dependent phosphorylation of the guard cell K+ uptake channel KAT1 leads to inhibition of K+in currents in oocytes (Sato et al., 2009). Activation of OST1 is controlled by PYR/PYL/RCAR receptor proteins and ABA (Ma et al., 2009; Park et al., 2009; Nishimura et al., 2010). The concentration of ABA tends to be increased in seeds of ABA insensitive mutants such as abi2-1 (Koornneef et al., 1984; Ma et al., 2009). Thus it is possible that a feedback mechanism exists which leads to increased ABA biosynthesis in plants when SLAC1 is impaired and which in turn leads to enhanced activation of protein kinases such as OST1 and concomitant inactivation of KAT1 and reduction of K+in currents. However, we did not observe any statistically significant differences in ABA concentration between slac1 mutants and wild type plants in whole plant extracts (Fig. S5), which does not exclude that there may be differences at the guard cell level.
CO2- and ABA-induced stomatal closure in slac1 mutants
Previously it was shown that slac1 mutants are insensitive to increases in CO2 concentration (Negi et al., 2008; Vahisalu et al., 2008). Here we observed that stomatal closure induced by an increase in [CO2] was reduced, but not completely abolished in slac1 plants (Figs S2, S3). In addition, as shown earlier (Vahisalu et al., 2008), SLAC1 is required for rapid 10 μM [ABA]-induced stomatal closure (Fig. 5b). Interestingly, application of a higher concentration of ABA for a longer period of time induced partial stomatal closure in slac1 (Fig. 5a). Stomatal closure induced by darkness or reduction of air humidity was also present in slac1, albeit delayed and with slower kinetics compared to wild type plants (Vahisalu et al., 2008). Both S- and R-type anion channels are activated by ABA and CO2 in Vicia faba (Raschke et al., 2003). Recently it was shown that AtALMT12 encodes a guard cell R-type anion channel (Meyer et al., 2010) and indeed CO2-, darkness- and ABA-induced stomatal closure were partially suppressed in plants lacking AtALMT12 (Meyer et al., 2010; Sasaki et al., 2010). On the basis of this information we suggest that, whereas SLAC1 is required for fast stomatal responses, other channels such as R-type anion channels and possibly also SLAH3 (Geiger et al., 2011) can contribute to stomatal closure when the stimuli are sufficiently strong, depending on growth conditions and/or after extended time courses.
S-type and R-type anion channels have different dynamics (Linder & Raschke, 1992; Schroeder & Keller, 1992). R-type anion channel currents are malate permeable (Keller et al., 1989). By contrast, S-type anion currents did not show large malate efflux currents in Arabidopsis guard cells (Fig. S4) or in oocytes expressing SLAC1 (Geiger et al., 2009). Note that previous research (Vahisalu et al., 2008) analyzed an upper limit for relative malate permeability through extrapolation of chloride carrying currents with malate on the extracellular membrane side and therefore did not analyze malate transport mediated by S-type anion channels.
Conclusions
The present study demonstrates that slac1 mutants exhibit an unexpected pronounced slowing of stomatal opening and a dramatic reduction in K+in channel currents. These findings provide evidence that guard cells have a compensatory machinery that counteracts mutations that would cause enhanced stomatal apertures. Here, mechanisms have been identified that can provide a mechanistic basis for the slowed stomatal opening and the reduced K+in channel activity in slac1 alleles. The sensitivity of guard cell K+in channels to the free cytosolic Ca2+ concentration is enhanced (‘primed’). Large K+in channel activity can be rapidly recovered in slac1 guard cells by lowering [Ca2+]cyt. Furthermore, slac1 guard cells exhibit slightly elevated baseline [Ca2+]cyt levels. In addition, the partially reduced expression of guard cell ion channel and transporter transcripts, may also contribute a small component to the reported compensatory slowing of stomatal closing mechanisms. The compensatory slowing of stomatal opening in response to light, low CO2 and high humidity may provide an important feedback mechanism for gas exchange regulation in plants.
Supplementary Material
Fig. S1 Light-induced increase in whole-plant stomatal conductance is reduced in slac1 mutants.
Fig. S2 CO2-induced stomatal closure in slac1 mutants occurs at below-ambient CO2 concentrations.
Fig. S3 CO2-induced changes in stomatal conductance of wild type and slac1-3 plants.
Fig. S4 Arabidopsis guard cells S-type anion channels do not mediate large malate efflux currents.
Fig. S5 Whole-plant abscisic acid concentrations of wild type, slac1-1 and slac1-3 do not differ.
Table S1 Primers used for real time PCR
Acknowledgments
We thank Amber Ries for assistance with plant growth, Dr Heino Moldau for very valuable technical assistance and scientific comments, and Prof. Michael R. Blatt for preliminary data analyzing KAT1 vesicular trafficking in slac1 and for comments on an early version of the manuscript communicated to M. R. B. in 20.10.2011. This work was supported by the grants from Estonian Science Foundation (7763, 7361 and 9208, theme SF0180071s07) and by European Regional Fund (the Center of Excellence in Environmental Adaptation) to H.K., by grants from the NIH (R01GM060396), NSF (MCB0918220) to J.I.S. A grant from the Chemical Sciences, Geosciences, and Biosciences Division of the Office of Basic Energy Sciences at the US Department of Energy (DE-FG02-03ER15449) to J.I.S. supported initial K+ channel analyses. This research was also supported in part by grants from National Natural Science Foundation of China (31170227) and National Basic Research Program of China (973 program) (2012CB114300) to Y-F.W., and by the Academy of Finland Centre of Excellence program (2006-2011) to J.K.
Footnotes
Author Contributions: K.L. and Y-F.W. contributed equally to this study. K.L. found the slowed stomatal opening phenotypes of slac1 in H.K.’s laboratory, and Y-F.W. independently found the reduced K+in channel activities in slac1 alleles in J.I.S.’ laboratory. This study was conceived and experiments designed by H.K. and J.I.S.
Supporting Information
Additional supporting information may be found in the online version of this article.
Please note: Wiley–Blackwell are not responsible for the content or functionality of any supporting information supplied by the authors. Any queries (other than missing material) should be directed to the New Phytologist Central Office.
References
- Ache P, Becker D, Ivashikina N, Dietrich P, Roelfsema R, Hedrich R. GORK, a delayed outward rectifier expressed in guard cells of Arabidopsis thaliana, is a K+-selective, K+-sensing ion channel. FEBS Letters. 2000;8:93–98. doi: 10.1016/s0014-5793(00)02248-1. [DOI] [PubMed] [Google Scholar]
- Allen GJ, Chu SP, Harrington CL, Schumacher K, Hoffman T, Tang YY, Grill E, Schroeder JI. A defined range of guard cell calcium oscillation parameters encodes stomatal movements. Nature. 2001;411:1053–1057. doi: 10.1038/35082575. [DOI] [PubMed] [Google Scholar]
- Allen GJ, Murata Y, Chu SP, Nafisi M, Schroeder JI. Hypersensitivity of abscisic acid-induced cytosolic calcium increases in the Arabidopsis farnesyltransferase mutant era1-2. Plant Cell. 2002;14:1649–1662. doi: 10.1105/tpc.010448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Assmann SM, Simoncini L, Schroeder JI. Blue light activates electrogenic ion pumping in guard cell protoplasts of Vicia faba L. Nature. 1985;318:285–287. [Google Scholar]
- Barbier-Brygoo H, De Angeli A, Filleur S, Frachisse JM, Gambale F, Thomine S, Wege S. Anion channels/transporters in plants: from molecular bases to regulatory networks. Annual Review of Plant Biology. 2011;62:25–51. doi: 10.1146/annurev-arplant-042110-103741. [DOI] [PubMed] [Google Scholar]
- Blatt MR, Armstrong F. K+ channels of stomatal guard cells: abscisic-acid-evoked control of the outward rectifier mediated by cytoplasmic pH. Planta. 1993;191:330–341. [Google Scholar]
- Brandt B, Brodsky DE, Xue S, Negi J, Iba K, Kangasjärvi J, Ghassemian M, Stephan AB, Hu H, Schroeder JI. Reconstitution of abscisic acid activation of SLAC1 anion channel by CPK6 and OST1 kinases and branched ABI1 PP2C phosphatase action. Proceedings of the National Academy of Sciences, USA. 2012;109:10593–10598. doi: 10.1073/pnas.1116590109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Camarasa C, Bidard F, Bony M, Barre P, Dequin S. Characterization of Schizosaccharomyces pombe malate permease by expression in Saccharomyces cerevisiae. Applied and Environmental Microbiology. 2001;67:4144–4151. doi: 10.1128/AEM.67.9.4144-4151.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cao Y, Ward JM, Kelly WB, Ichida AM, Gaber RF, Anderson JA, Uozumi N, Schroeder JI, Crawford NM. Multiple genes, tissue specificity and expression-dependent modulation contribute to the functional diversity of potassium channels in Arabidopsis thaliana. Plant Physiology. 1995;109:1093–1106. doi: 10.1104/pp.109.3.1093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen ZH, Hills A, Lim CK, Blatt MR. Dynamic regulation of guard cell anion channels by cytosolic free Ca2+ concentration and protein phosphorylation. Plant Journal. 2010;61:816–25. doi: 10.1111/j.1365-313X.2009.04108.x. [DOI] [PubMed] [Google Scholar]
- Czechowski T, Stitt M, Altmann T, Udvardi MK, Scheible WR. Genome-wide identification and testing of superior reference genes for transcript normalization in Arabidopsis. Plant Physiology. 2005;139:5–17. doi: 10.1104/pp.105.063743. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Geiger D, Scherzer S, Mumm P, Stange A, Marten I, Bauer H, Ache P, Matschi S, Liese A, Al-Rasheid KAS, et al. Activity of guard cell anion channel SLAC1 is controlled by drought-stress signaling kinase-phosphatase pair. Proceedings of the National Academy of Sciences, USA. 2009;106:21425–21430. doi: 10.1073/pnas.0912021106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Geiger D, Scherzer S, Mumm P, Marten I, Ache P, Matschi S, Liese A, Wellmann C, Al-Rasheid KAS, Grill E, et al. Guard cell anion channel SLAC1 is regulated by CDPK protein kinases with distinct Ca2+ affinities. Proceedings of the National Academy of Sciences, USA. 2010;107:8023–8028. doi: 10.1073/pnas.0912030107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Geiger D, Maierhofer T, AL-Rasheid KAS, Scherzer S, Mumm P, Liese A, Ache P, Wellmann C, Marten I, Grill E, et al. Stomatal closure by fast abscisic acid signaling is mediated by the guard cell anion channel SLAH3 and the receptor RCAR1. Science Signaling. 2011;4:173, ra32. doi: 10.1126/scisignal.2001346. [DOI] [PubMed] [Google Scholar]
- Grabov A, Blatt MR. Membrane voltage initiates Ca2+ waves and potentiates Ca2+ increases with abscisic acid in stomatal guard cells. Proceedings of the National Academy of Sciences, USA. 1998;95:4778–4783. doi: 10.1073/pnas.95.8.4778. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grabov A, Blatt MR. A steep dependence of inward-rectifying potassium channels on cytosolic free calcium concentration increase evoked by hyperpolarization in guard cells. Plant Physiology. 1999;119:277–288. doi: 10.1104/pp.119.1.277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grefen C, Donald N, Hashimoto K, Kudla J, Schumacher K, Blatt MR. A ubiquitin-10 promoter-based vector set for fluorescent protein tagging facilitates temporal stability and native protein distribution in transient and stable expression studies. Plant Journal. 2010;64:355–365. doi: 10.1111/j.1365-313X.2010.04322.x. [DOI] [PubMed] [Google Scholar]
- Hedrich R, Busch H, Raschke K. Ca2+ and nucleotide dependent regulation of voltage dependent anion channels in the plasma membrane of guard cells. EMBO Journal. 1990;9:3889–3892. doi: 10.1002/j.1460-2075.1990.tb07608.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hamill OP, Marty A, Neher E, Sakmann B, Sigworthm FJ. Improved patch-clamp techniques for high-resolution current recording from cells and cell-free membrane patches. Pflugers Archiv-European Journal of Physiology. 1981;391:85–100. doi: 10.1007/BF00656997. [DOI] [PubMed] [Google Scholar]
- Hamilton DWA, Hills A, Kohler B, Blatt MR. Ca2+ channels at the plasma membrane of stomatal guard cells are activated by hyperpolarization and abscisic acid. Proceedings of the National Academy of Sciences, USA. 2000;97:4967–4972. doi: 10.1073/pnas.080068897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hubbard KE, Siegel RS, Valerio G, Brandt B, Schroeder JI. Abscisic acid and CO2 signalling via calcium sensitivity priming in guard cells, new CDPK mutant phenotypes and a method for improved resolution of stomatal stimulus–response analyses. Annals of Botany. 2012;109:5–17. doi: 10.1093/aob/mcr252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ivanoff L. Zur Methodik der Transpirationsbestimmung am Standort. Berichte der Deutschen Botanischen Gesellschaft. 1928;46:306–310. [Google Scholar]
- Keller BU, Hedrich R, Raschke K. Voltage-dependent anion channels in the plasma membrane of guard cells. Nature. 1989;341:450–453. doi: 10.1002/j.1460-2075.1990.tb07608.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kelly WB, Esser JE, Schroeder JI. Effects of cytosolic calcium and limited, possible dual, effects of G protein modulators on guard cell inward potassium channels. Plant Journal. 1995;8:479–489. [Google Scholar]
- Kim TH, Böhmer M, Hu H, Nishimura N, Schroeder JI. Guard cell signal transduction network: advances in understanding abscisic acid, CO2, and Ca2+ sgnaling. Annual Review of Plant Biology. 2010;61:561–591. doi: 10.1146/annurev-arplant-042809-112226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kollist T, Moldau H, Rasulov B, Oja V, Rämma H, Hüve K, Jaspers P, Kangasjärvi J, Kollist H. A novel device detects a rapid ozone-induced transient stomatal closure in intact Arabidopsis and its absence in abi2 mutant. Physiologia Plantarum. 2007;129:796–803. [Google Scholar]
- Kollist H, Jossier M, Laanemets K, Thominem S. Anion channels in plant cells. FEBS Journal. 2011;278:4277–4292. doi: 10.1111/j.1742-4658.2011.08370.x. [DOI] [PubMed] [Google Scholar]
- Koornneef M, Reuling G, Karssen CM. The isolation and characterization of abscisic acid-insensitive mutants of Arabidopsis thaliana. Physiologia Plantarum. 1984;61:377–383. [Google Scholar]
- Kwak JM, Murata Y, Baizabal-Aguirre VM, Merrill J, Wang M, Kemper A, Hawke SD, Tallman G, Schroeder JI. Dominant negative guard cell K+ channel mutants reduce inward-rectifying K+ currents and light-induced stomatal opening in Arabidopsis. Plant Physiology. 2001;127:473–485. [PMC free article] [PubMed] [Google Scholar]
- Lebaudy A, Vavasseur A, Hosy E, Dreyer I, Leonhardt N, Thibaud JB, Véry AA, Simonneau T, Sentenac H. Plant adaptation to fluctuating environment and biomass production are strongly dependent on guard cell potassium channels. Proceedings of the National Academy of Sciences, USA. 2008;105:5271–5276. doi: 10.1073/pnas.0709732105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee M, Choi Y, Burla B, Kim YY, Jeon B, Maeshima M, Yoo JY, Martinoia E, Lee Y. The ABC transporter AtABCB14 is a malate importer and modulates stomatal response to CO2. Nature Cell Biology. 2008;10:1217–1223. doi: 10.1038/ncb1782. [DOI] [PubMed] [Google Scholar]
- Lee SC, Lan W, Buchanan BB, Luan S. A protein kinase-phosphatase pair interacts with an ion channel to regulate ABA signaling in plant guard cells. Proceedings of the National Academy of Sciences, USA. 2009;106:21419–21424. doi: 10.1073/pnas.0910601106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leonhardt N, Kwak JM, Robert N, Waner D, Leonhardt G, Schroeder JI. Microarray expression analyses of Arabidopsis guard cells and isolation of a recessive abscisic acid hypersensitive protein phosphatase 2C mutant. Plant Cell. 2004;16:596–615. doi: 10.1105/tpc.019000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Linder B, Raschke K. A slow anion channel in guard cells, activating at large hyperpolarization, may be principal for stomatal closing. FEBS Letters. 1992;313:27–30. doi: 10.1016/0014-5793(92)81176-m. [DOI] [PubMed] [Google Scholar]
- Ma Y, Szostkiewicz I, Korte A, Moes D, Yang Y, Christmann A, Grill E. Regulators of PP2C phosphatase activity function as abscisic acid sensors. Science. 2009;324:1064–1068. doi: 10.1126/science.1172408. [DOI] [PubMed] [Google Scholar]
- MacRobbie EAC. ABA activates multiple Ca2+ fluxes in stomatal guard cells, triggering vacuolar K+ (Rb+) release. Proceedings of the National Academy of Sciences, USA. 2000;97:12361–12368. doi: 10.1073/pnas.220417197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marten H, Hedrich R, Roelfsema MRG. Blue light inhibits guard cell plasma membrane anion channels in a phototropin-dependent manner. Plant Journal. 2007a;50:29–39. doi: 10.1111/j.1365-313X.2006.03026.x. [DOI] [PubMed] [Google Scholar]
- Marten H, Konrad KR, Dietrich P, Roelfsema MRG, Hedrich R. Ca2+-dependent and -independent abscisic acid activation of plasma membrane anion channels in guard cells of Nicotiana tabacum. Plant Physiology. 2007b;143:28–37. doi: 10.1104/pp.106.092643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McAinsh MR, Brownlee C, Hetherington AM. Abscisic acid-induced elevation of guard cell cytosolic Ca2+ precedes stomatal closure. Nature. 1990;343:186–188. [Google Scholar]
- Meyer S, Mumm P, Imes D, Endler A, Weder B, Al-Rasheid KAS, Geiger D, Marten I, Martinoia E, Hedrich R. AtALMT12 represents an R-type anion channel required for stomatal movement in Arabidopsis guard cells. Plant Journal. 2010;63:1054–1062. doi: 10.1111/j.1365-313X.2010.04302.x. [DOI] [PubMed] [Google Scholar]
- Mori IC, Murata Y, Yang Y, Munemasa S, Wang YF, Andreoli S, Tiriac H, Alonso JM, Harper JF, Ecker JR, et al. CDPKs CPK6 and CPK3 function in ABA regulation of guard cell S-type anion- and Ca2+-permeable channels and stomatal closure. PLoS Biology. 2006;4:e327. doi: 10.1371/journal.pbio.0040327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakamura RL, McKendree WL, Jr, Hirsch RE, Sedbrook JC, Gaber RF, Sussman MR. Expression of an Arabidopsis potassium channel gene in guard cells. Plant Physiology. 1995;109:371–374. doi: 10.1104/pp.109.2.371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Negi J, Matsuda O, Nagasawa T, Oba Y, Takahashi H, Kawai-Yamada M, Uchimiya H, Hashimoto M, Iba K. CO2 regulator SLAC1 and its homologues are essential for anion homeostasis in plant cells. Nature. 2008;452:483–486. doi: 10.1038/nature06720. [DOI] [PubMed] [Google Scholar]
- Nishimura N, Sarkeshik A, Nito K, Park SY, Wang A, Carvalho PC, Lee S, Caddell DF, Cutler SR, Chory J, et al. PYR/PYL/RCAR family members are major in-vivo ABI1 protein phosphatase 2C-interacting proteins in Arabidopsis. Plant Journal. 2010;61:290–299. doi: 10.1111/j.1365-313X.2009.04054.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Overmyer K, Kollist H, Tuominen H, Betz C, Langebartels C, Wingsle G, Kangasjärvi S, Brader G, Munnineaux P, Kangasjärvi J. Complex phenotypic profiles leading to ozone sensitivity in Arabidopsis thaliana mutants. Plant, Cell & Environment. 2008;31:1237–1249. doi: 10.1111/j.1365-3040.2008.01837.x. [DOI] [PubMed] [Google Scholar]
- Overmyer K, Tuominen H, Kettunen R, Betz C, Langebartels C, Sandermann H, Kangasjärvi J. Ozone-sensitive Arabidopsis rcd1 mutant reveals opposite roles for ethylene and jasmonate signaling pathways in regulating superoxide-dependent cell death. Plant Cell. 2000;12:1849–1862. doi: 10.1105/tpc.12.10.1849. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Park SY, Fung P, Nishimura N, Jensen DR, Fujii H, Zhao Y, Lumba S, Santiago J, Rodrigues A, Chow TF, et al. Abscisic acid inhibits type 2C protein phosphatases via the PYR/PYL family of START proteins. Science. 2009;324:1068–1071. doi: 10.1126/science.1173041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pei ZM, Kuchitsu K, Ward JM, Schwarz M, Schroeder JI. Differential abscisic acid regulation of guard cell slow anion channels in Arabidopsis wild-type and abi1 and abi2 mutants. Plant Cell. 1997;9:409–423. doi: 10.1105/tpc.9.3.409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pei Z-M, Murata Y, Benning G, Thomine S, Klusener B, Allen GJ, Grill E, Schroeder JI. Calcium channel activated by hydrogen peroxide mediate abscisic acid signalling in guard cells. Nature. 2000;406:731–734. doi: 10.1038/35021067. [DOI] [PubMed] [Google Scholar]
- Peiter E, Maathuis FJ, Mills LN, Knight H, Pelloux J, Hetherington AM, Sanders D. The vacuolar Ca2+-activated channel TPC1 regulates germination and stomatal movement. Nature. 2005;434:404–408. doi: 10.1038/nature03381. [DOI] [PubMed] [Google Scholar]
- Pilot G, Lacombe B, Gaymard F, Chérel I, Boucherez J, Thibaud JB, Sentenac H. Guard cell inward K+ channel activity in Arabidopsis involves expression of the twin channel subunits KAT1 and KAT2. Journal of Biological Chemistry. 2001;276:3215–3221. doi: 10.1074/jbc.M007303200. [DOI] [PubMed] [Google Scholar]
- Pusch M, Neher E. Rates of diffusional exchange between small cells and a measuring pipette. Pflugers Archiv-European Journal of Physiology. 1988;411:204–211. doi: 10.1007/BF00582316. [DOI] [PubMed] [Google Scholar]
- Raschke K, Hedrich R, Reckmann U, Schroeder JI. Exploring biophysical and biochemical components of the osmotic motor that drives stomatal movement. Botanica Acta. 1988;101:283–94. [Google Scholar]
- Raschke K, Shabahang M, Wolf R. The slow and the quick anion conductance in whole guard cells: their voltage-dependent alternation, and the modulation of their activities by abscisic acid and CO2. Planta. 2003;217:639–650. doi: 10.1007/s00425-003-1033-4. [DOI] [PubMed] [Google Scholar]
- Reintanz B, Szyroki A, Ivashikina N, Ache P, Godde M, Becker D, Palme K, Hedrich R. AtKC1, a silent Arabidopsis potassium channel α-subunit modulates root hair K+ influx. Proceedings of the National Academy of Sciences, USA. 2002;99:4079–4084. doi: 10.1073/pnas.052677799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roelfsema MRG, Steinmeyer R, Staal M, Hedrich R. Single guard cell recordings in intact plants: light-induced hyperpolarization of the plasma membrane. Plant Journal. 2001;26:1–13. doi: 10.1046/j.1365-313x.2001.01000.x. [DOI] [PubMed] [Google Scholar]
- Roelfsema MRG, Hedrich R. In the light of stomatal opening: new insights into ‘the Watergate’. New Phytologist. 2005;167:665–691. doi: 10.1111/j.1469-8137.2005.01460.x. [DOI] [PubMed] [Google Scholar]
- Sasaki T, Mori IC, Furuichi T, Munemasa S, Toyooka K, Matsuoka K, Murata Y, Yamamoto Y. Closing plant stomata requires a homolog of an aluminum-activated malate transporter. Plant Cell Physiology. 2010;51:354–365. doi: 10.1093/pcp/pcq016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sato A, Sato Y, Fukao Y, Fujiwara M, Umezawa T, Shinozaki K, Hibi T, Taniguchi M, Miyake H, Goto D, et al. Threonine at position 306 of the KAT1 potassium channel is essential for channel activity and is a target site for ABA-activated SnRK2/OST1/SnRK2.6 protein kinase. Biochemical Journal. 2009;424:439–448. doi: 10.1042/BJ20091221. [DOI] [PubMed] [Google Scholar]
- Schroeder JI, Hagiwara S. Cytosolic calcium regulates ion channels in the plasma membrane of Vicia faba guard cells. Nature. 1989;338:427–430. [Google Scholar]
- Schroeder JI, Hedrich R, Fernandez JM. Potassium-selective single channels in guard cell protoplasts of Vicia faba. Nature. 1984;312:361–362. [Google Scholar]
- Schroeder JI, Keller BU. Two types of anion channel currents in guard cells with distinct voltage regulation. Proceedings of the National Academy of Sciences, USA. 1992;89:5025–5029. doi: 10.1073/pnas.89.11.5025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schroeder JI, Raschke K, Neher E. Voltage dependence of K+ channels in guard-cell protoplasts. Proceedings of the National Academy of Sciences, USA. 1987;84:4108–4112. doi: 10.1073/pnas.84.12.4108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schroeder JI, Schmidt C, Sheaffer J. Identification of high-affinity slow anion channel blockers and evidence for stomatal regulation by slow anion channels in guard cells. Plant Cell. 1993;5:1831–1841. doi: 10.1105/tpc.5.12.1831. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schwartz A, Ilan N, Schwarz M, Scheaffer J, Assmann SM, Schroeder JI. Anion channel blockers inhibit S-type anion channels and abscisic acid responses in guard cells. Plant Physiology. 1995;109:651–658. doi: 10.1104/pp.109.2.651. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shimazaki K, Iino M, Zeiger E. Blue light-dependent proton extrusion by guard-cell protoplasts of Vicia faba. Nature. 1986;319:324–326. [Google Scholar]
- Shimazaki K, Doi M, Assmann SM, Kinoshita T. Light regulation of stomatal movement. Annual Review of Plant Biology. 2007;58:219–247. doi: 10.1146/annurev.arplant.57.032905.105434. [DOI] [PubMed] [Google Scholar]
- Siegel RS, Xue SW, Murata Y, Yang YZ, Nishimura N, Wang A, Schroeder JI. Calcium elevation-dependent and attenuated resting calcium-dependent abscisic acid induction of stomatal closure and abscisic acid-induced enhancement of calcium sensitivities of S-type anion and inward-rectifying K+ channels in Arabidopsis guard cells. Plant Journal. 2009;59:207–220. doi: 10.1111/j.1365-313X.2009.03872.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Szyroki A, Ivashikina N, Dietrich P, Roelfsema MR, Ache P, Reintanz B, Deeken R, Godde M, Felle H, Steinmeyer R, et al. KAT1 is not essential for stomatal opening. Proceedings of the National Academy of Sciences, USA. 2001;98:2917–2921. doi: 10.1073/pnas.051616698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Staxen I, Pical C, Montgomery LT, Gray JE, Hetherington AM, McAinsh MR. Abscisic acid induces oscillations in guard-cell cytosolic free calcium that involve phosphoinositide-specific phospholipase C. Proceedings of the National Academy of Sciences, USA. 1999;96:1779–1784. doi: 10.1073/pnas.96.4.1779. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thiel G, MacRobbie EAC, Blatt MR. Membrane transport in stomatal guard cells: the importance of voltage control. Journal of Membrane Biology. 1992;126:1–18. doi: 10.1007/BF00233456. [DOI] [PubMed] [Google Scholar]
- Ueno K, Kinoshita T, Inoue S, Emi T, Shimazaki K. Biochemical characterization of plasma membrane H+-ATPase activation in guard cell protoplasts of Arabidopsis thaliana in response to blue light. Plant Cell Physiology. 2005;46:955–961. doi: 10.1093/pcp/pci104. [DOI] [PubMed] [Google Scholar]
- Uozumi N, Gassmann W, Cao Y, Schroeder JI. Identification of strong modifications in cation selectivity in an Arabidopsis inward rectifying potassium channel by mutant selection in yeast. Journal of Biological Chemistry. 1995;270:24276–24281. doi: 10.1074/jbc.270.41.24276. [DOI] [PubMed] [Google Scholar]
- Vahisalu T, Kollist H, Wang YF, Nishimura N, Chan WY, Valerio G, Lamminmäki A, Brosché M, Moldau H, Desikan R, Schroeder JI, et al. SLAC1 is required for plant guard cell S-type anion channel function in stomatal signalling. Nature. 2008;452:487–491. doi: 10.1038/nature06608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vahisalu T, Puzõrjova I, Brosché M, Valk E, Lepiku M, Moldau H, Pechter P, Wang YS, Lindgren O, Salojärvi J, et al. Ozone-triggered rapid stomatal response involves the production of reactive oxygen species, and is controlled by SLAC1 and OST1. Plant Journal. 2010;62:442–453. doi: 10.1111/j.1365-313X.2010.04159.x. [DOI] [PubMed] [Google Scholar]
- Very A-A, Sentenac H. Molecular mechanisms and regulation of K+ transport in higher plants. Annual Review of Plant Biology. 2003;54:575–603. doi: 10.1146/annurev.arplant.54.031902.134831. [DOI] [PubMed] [Google Scholar]
- Wang Y, Blatt MR. Anion channel sensitivity to cytosolic organic acids implicates a central role for oxaloacetate in integrating ion flux with metabolism in stomatal guard cells. Biochemica Journal. 2011;439:161–70. doi: 10.1042/BJ20110845. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Webb AAR, McAinsh MR, Mansfield TA, Hetherington AM. Carbon dioxide induces increases in guard cell cytosolic free calcium. Plant Journal. 1996;9:297–304. [Google Scholar]
- Yang Y, Costa A, Leonhardt N, Siegel RS, Schroeder JI. Isolation of a strong Arabidopsis guard cell promoter and its potential as a research tool. Plant Methods. 2008;4:6. doi: 10.1186/1746-4811-4-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Young JJ, Mehta S, Israelsson M, Godoski J, Grill E, Schroeder JI. CO2 signaling in guard cells: calcium sensitivity response modulation, a Ca2+ -independent phase, and CO2 insensitivity of the gca2 mutant. Proceedings of the National Academy of Sciences, USA. 2006;103:7506–7511. doi: 10.1073/pnas.0602225103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xue S, Hu H, Ries A, Merilo E, Kollist H, Schroeder JI. Central functions of bicarbonate in S-type anion channel activation and OST1 protein kinase in CO2 signal transduction in guard cell. EMBO Journal. 2011;30:1645–1658. doi: 10.1038/emboj.2011.68. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Fig. S1 Light-induced increase in whole-plant stomatal conductance is reduced in slac1 mutants.
Fig. S2 CO2-induced stomatal closure in slac1 mutants occurs at below-ambient CO2 concentrations.
Fig. S3 CO2-induced changes in stomatal conductance of wild type and slac1-3 plants.
Fig. S4 Arabidopsis guard cells S-type anion channels do not mediate large malate efflux currents.
Fig. S5 Whole-plant abscisic acid concentrations of wild type, slac1-1 and slac1-3 do not differ.
Table S1 Primers used for real time PCR