

1. Introduction
Enzymes are very efficient catalysts that are essential for the functioning of living organisms. The low efficiency of biocatalysts produced de novo relative to those that have evolved naturally demonstrates that our understanding of enzymatic catalysis is still incomplete.1−4 The dynamic motion of enzymes during catalytic events is one of the many aspects of protein chemistry that are currently insufficiently well understood.5−9 On one hand, proteins need to have well-defined and organized structures in order to maintain stable functionality in the intracellular environment. On the other hand, some degree of flexibility is often required for catalytic activity. Molecular dynamics simulations have provided key insights into the importance of protein dynamics in catalysis, such as the observation of substrate access and product exit pathways that cannot be identified by inspecting crystal structures.10 Csermely et al. recently reported that mutations in regions that affect protein dynamics, such as hinge regions that are important in substrate binding, can have dramatic effects on catalytic activity.11 In this review, we highlight the role of protein gates as another class of highly dynamic structures that play key roles in protein function.
Given the importance of gates for enzymatic catalysis, the number of studies that have examined them systematically is surprisingly small. Conformational gating in proteins was first described by McCammon and co-workers in 1981, but there have been relatively few systematic studies in this area since then.12−14 Moreover, much of the available data on gates in macromolecular systems is hidden or otherwise dispersed within the scientific literature, partly because there is currently no consensus regarding what defines a gate. Some authors describe all residues that affect the ligand’s access to a target area as gating residues, whereas others apply the term exclusively to structural features that undergo large movements during the gating event. In this review, we define a gate as a dynamic system consisting of individual residues, loops, secondary structure elements, or domains that can reversibly switch between open and closed conformations and thereby control the passage of small molecules—substrates, products, ions, and solvent molecules—into and out of the protein structure. Under this definition, the anchoring residues that stabilize the open or closed conformations of a gate are not themselves gating residues. However, because of their various interactions with the gating residues, they can control the size and properties of the ligands that pass through the gate as well as the frequency of the exchange events.
Gates can be found in various systems, including enzymes, ion channels, protein–protein complexes, and protein–nucleic acid complexes.14,15 In this work, we focus specifically on gates in enzymes. We attempt to answer three basic questions—why, how, where—by describing the molecular function, structural basis, and location of gates within protein structures. We discuss 71 illustrative examples of enzymes that together contain 129 different molecular gates and propose a system for their classification. Reviewed enzymes were chosen based on a literature search with a set of keywords corresponding to gates and conformational changes in enzymes. A preliminary set of protein structures was filtered out, leaving only those entries for which both open and closed conformations were described. The final set represents different classes of enzymes (Figure 1A) and different protein folds (Figure 1C) and spans structurally and functionally the entire enzyme world. Among 71 chosen enzymes, only 17 have higher than 20% sequence identity with other set members (Figure 1E). The proposed classification system provides a useful framework for comparing gates of different enzymes and drawing general conclusions about gate function, structure, and position. Moreover, the classification scheme is easily extendable to describe the new gate types that will almost certainly be revealed by structural and functional analyses of newly isolated enzymes in the future.
Figure 1.

Distribution of (A) reviewed proteins and (B) proteins from the PDB database according to EC classes, (C) reviewed proteins and (D) proteins from the PDB database according to SCOP classification, and (E) identity matrix of reviewed enzymes (only the enzymes with sequence identities above 20% are shown for clarity).
2. Molecular Function of Gates
Analyses of protein dynamics have identified a number of enzymes with gates, suggesting that these structures are rather common. What is the molecular function of the gates? It seems that in enzymes they facilitate precise control over processes that are directly linked to catalysis. Enzyme gates can (i) contribute to enzyme selectivity by controlling substrate access to the active site, (ii) prevent solvent access to specific regions of the protein, and (iii) synchronize processes occurring in distant parts of the protein (Figure 2). The proper function of even the simplest gates can potentially be essential for catalysis, and the gating event may even represent the rate-limiting step of the catalytic cycle. Interestingly, different gating residues within a single protein molecule may be responsible for restricting the access of specific substrates. High variability of the gating residues within an enzyme scaffold can lead to the evolution of enzyme families whose members are selective for specific substrate types. The best known example of such specialization within a single enzyme family is provided by the cytochromes P450.16
Figure 2.
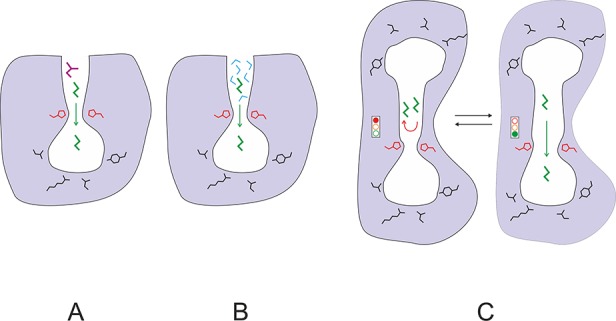
Schematic illustration of the molecular functions of protein gates: (A) control of substrate access, (B) control of solvent access, (C) control and synchronization of reactions. Protein is represented by the area colored in gray, active site cavity by the area in white, gating residues by red lines, substrate molecules by green or violet lines, and water molecules by blue lines.
2.1. Control of Substrate Access
Enzyme selectivity has been traditionally explained by the “lock and key” model,17 which was subsequently complemented by the “induced fit” or “hand in glove”,18 “selected fit”,19 and “keyhole, lock, and key”20 models. In many cases, these models provide an adequate description of enzyme selectivity based on adjustable complementarity between the active site and the cognate substrates. However, research conducted over the past decade has shown that regions located further from the active site can also affect enzyme selectivity. Substrate access pathways, which often incorporate molecular gates, impose additional constraints on ligand binding to the active site.20 The ability of ligands to traverse these access pathways can be controlled by (i) size discrimination at the narrowest point along the pathway forming a bottleneck, (ii) geometrical constraints, e.g., the curvature of the pathway, and (iii) specific molecular interactions such as hydrogen bonds, electrostatic interactions, and hydrophobic interactions with the residues comprising the access pathway. Protein gates can be regarded as molecular filters that discriminate between molecules as similar as molecular oxygen and carbon monoxide in NiFe hydrogenases21,22 or water and hydroxyperoxide in catalases.23,24 Gates act as filters in a wide range of enzymes, controlling the range of substrates that can be accepted by broad-specificity cytochromes P450,25 the stereospecificity of epoxide hydrolases,26 and product length in undecaprenyl-pyrophosphate synthases.27
One of the first systematic descriptions of the influence of the gating process on substrate binding was reported by Szabo et al., who assumed that the switching between the open and the closed conformations of the gate was a stochastic process.28−30 This model was successfully used to demonstrate that despite conformation gating13 acetylcholinesterase can bind acetylcholine with a rate constant of 109 M–1 s–1 and predict the rate of formation of the enzyme–substrate complex in choline oxidase.31 Since gates create a barrier on the substrate access pathway, the kinetic rate constant for passage over the barrier can be obtained using Kramers’ reaction rate theory or its later modifications.32−34 This methodology was used to compare the results of computational and experimental studies on the passage of the tetramethylammonium cation through acetylcholinesterase35 and migration of ammonia through carbamoyl phosphate synthetase.36
2.2. Control of Solvent Access
Spatial localization of the hydrophobic and hydrophilic regions within the structure of a protein is important in maintaining its proper fold and can also be crucial for catalytic function. The various steps of an enzymatic reaction may require different environments. These distinct environments can be generated by having the individual steps occur in spatially separate regions of the protein, but this does not eliminate the problem of transporting the substrate between these sites. There are important problems to be addressed, including transporting polar molecules from a polar environment to a nonpolar one and separating hydrophilic compartments from hydrophobic ones within the structure of a single protein.
In some proteins, these problems are addressed by the presence of selective barriers that permit passage of solutes but not water molecules. Crystallographic and NMR data can be used to identify cavities within a protein structure accessible to water molecules. Exclusion of water from some parts of the cavity, such as the active site or a specific tunnel, is essential for functioning of numerous enzymes. In simple cases, the gates may prevent the entrance of water molecules into the cavity when a substrate or a cofactor is not present, as occurs in rabbit 20a-hydroxysteroid dehydrogenase.37 In more complex cases, the gates may permit access only to a specific part of the cavity, as occurs in carbamoyl phosphate synthetase36 and imidazole glycerol phosphate synthase.38 In the cytochromes P450, a “water channel” controls hydration of the substrate in the active site, which is extremely important for cytochrome activity.16 The potential importance of gates that act as solvent barriers is further illustrated by the example of enzymes with ammonia tunnels. In these proteins, gates prevent water from entering the channel and protonating the ammonia, which is essential for maintaining its nucleophilic character.39
Control of water access can be seen as a special case of the function described in section 2.1. However, when discussing water exclusion, the main emphasis is on the water permeability of the gates and their ability to distinguish water molecules from other ligands. The gate can simultaneously act as a barrier to passage of water molecules while acting as a selective filter for other molecules, allowing them to access the active site. Gates of this type resemble semiselective membranes that can distinguish between species such as water and ammonia, allowing only the second to pass. It is worth noting that passage of ‘permitted’ species can be facilitated by rearrangement of an individual gating residue, such as K99 in imidazole glycerol phosphate synthase. Conversely, passage of water molecules through gates of this type would often require significant conformational changes in all of the residues that comprise the gate.38 This makes gates in enzymes far more sophisticated than semipermeable membranes.
2.3. Control and Synchronization of Reactions
Another function of gates becomes apparent when considering enzymes with two or more active sites. Many protein structures contain tunnels to facilitate efficient migration of intermediates and gates to synchronize chemical reactions. Such arrangements can be compared to a pair of workers on an assembly line. The second worker has to be ready before he can receive a product from the first one. Moreover, the products generated by the first worker must satisfy certain standards. The control gates located between the workers regulate the exchange of products over a well-defined period of time. Gates of this kind are common in ammonia-transferring enzymes, suggesting that they are old in evolutionary terms and functionally important.40−43 The need to efficiently transport ammonia within the interior of the protein may be related to its high cellular toxicity. We speculate that gates of this kind may be present in many enzymes that have multiple active sites connected by internal tunnels for the transport of intermediates. Many such enzymes have been studied in some detail, including carbamoyl phosphate synthetase, which has tunnels for ammonia and carbamate transportation;36 asparagine synthetase,44 glucosamine 6-phosphate synthase,45 and glutamate synthase,46 all of which have tunnels for ammonia transportation; tryptophan synthase for indole47 and carbon monoxide dehydrogenase/acetyl coenzyme A synthase for carbon monoxide transportation.48
Systematic analysis of the functions of the known gates in the 71 proteins discussed in this article revealed the following distribution of gate types: 40% of the studied gates control substrate access, 19% control solvent access, 15% control and synchronize catalytic events, and 26% have other function (Figure 3).
Figure 3.
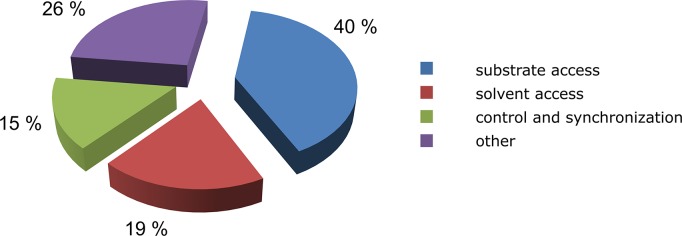
Distribution of gate functions; 71 proteins with 129 different gates were analyzed. Percentages shown in the figure are based on all of the identified functions of each gate; individual gate may perform multiple functions. Detailed description of the analyzed proteins is provided in Table 3.
3. Structural Basis of Gates
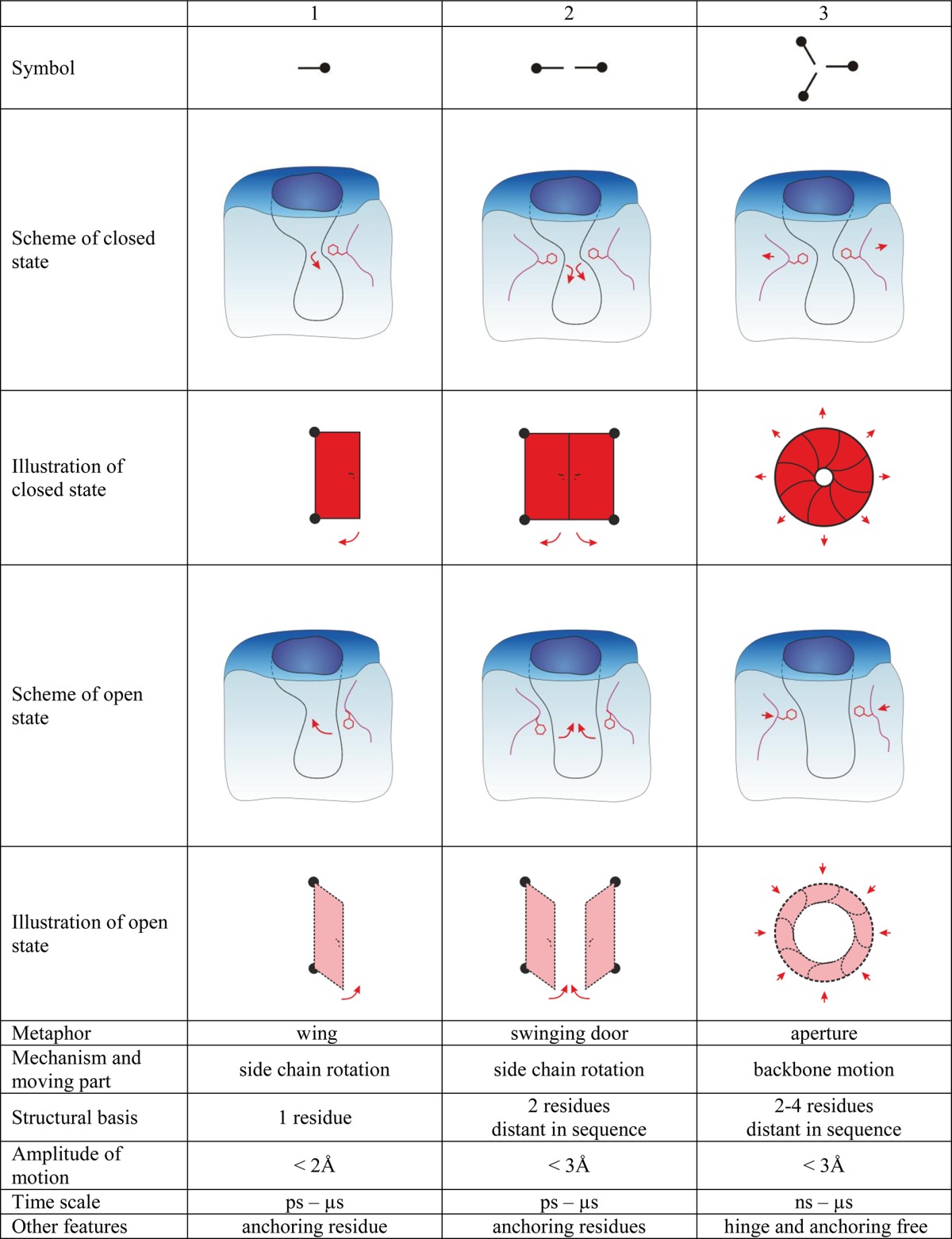
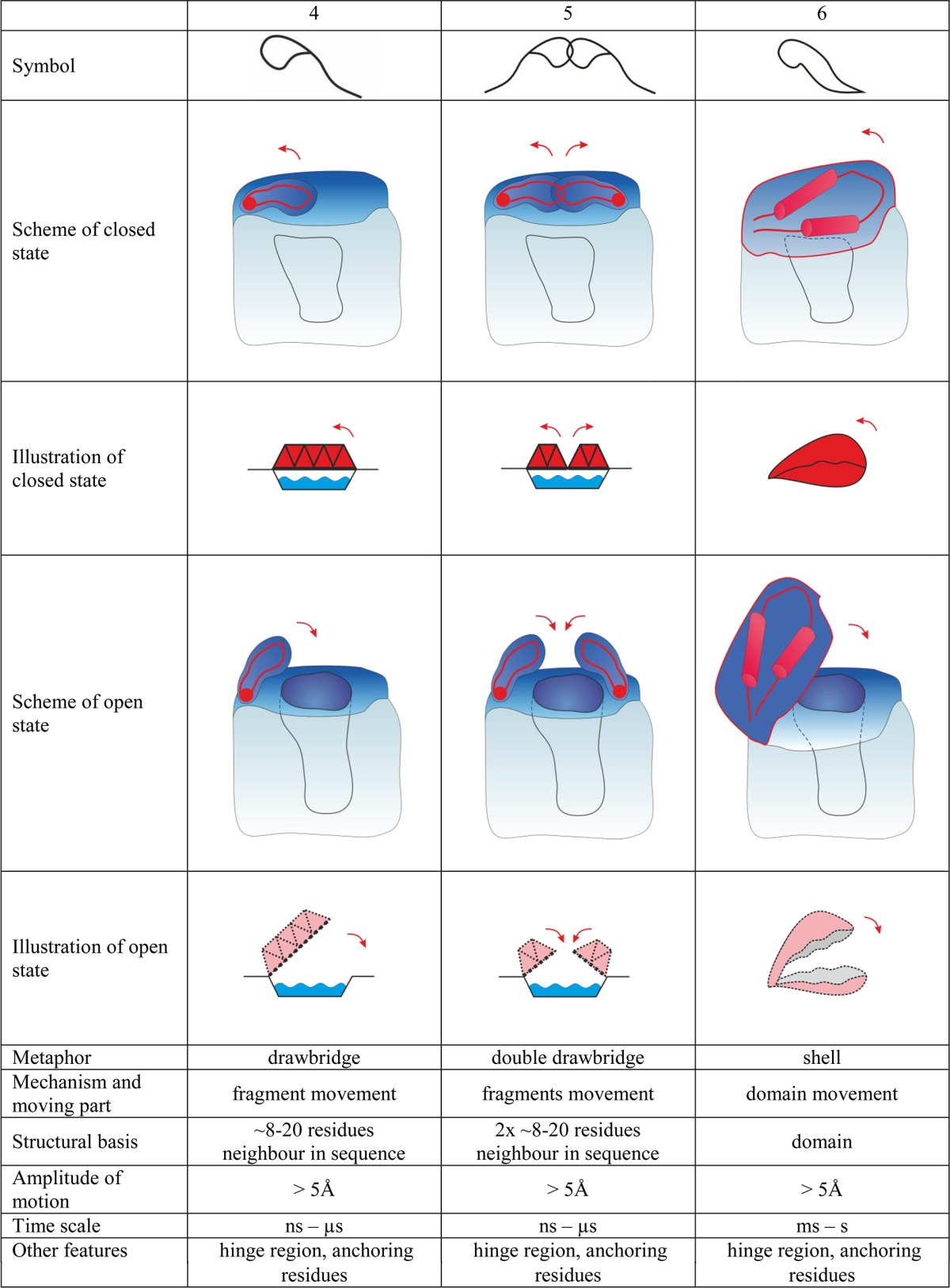
Gates are dynamic systems that can make reversible transitions between open and closed states. They vary in size and complexity, from individual amino acid residues to loops, secondary structure elements, and even domains. The simplest gates consist of only one amino acid side chain that can close or open an access pathway by rotating. Opening and closing of more complicated systems can involve the synchronized movement of two or more residues, and the largest systems involve rearrangements of secondary elements or even entire domains (Table 1). For larger systems, movement of the gate may cause formation of a tunnel or enclosed cavity in addition to permitting or denying access to selected species.49,50 The following parameters can be useful for describing and discriminating between gates: (i) their constituent residues, (ii) their anchoring residues, (iii) the hinge region, i.e., the amino acids that make the structure flexible and allow it to move, (iv) the gate’s position, (v) the gate’s bottleneck diameter in the open and closed states, (vi) changes in the bottleneck’s size over time, (vii) the energy required to switch the gate from one state to the other, and (viii) the energy required for passage of specific molecules through the gate.
Table 1. Classifying Enzyme Gates According to Their Structural Basis.
3.1. Residue Motion: Wings
The energetic barriers for residue rotation are quite small, 1–16 kcal/mol.51 While generally low, such barriers can nevertheless be large enough to significantly affect the probability that a given species will be able to pass through the gate or the rate at which they do so. Depending on the particular amino acid and its surroundings, one or both states of the gate may be stabilized by interactions with anchoring residues, e.g., hydrophobic interactions, H bonds, ionic interactions, salt bridges, and π–π interactions. The strongest effect on the control of the passage is achieved when a large gating residue is located in the bottleneck of the pathway. The most common residues in this role are those whose side chains contain aromatic rings, i.e., W, F, and Y (Figure 4). Wing-type gates are common and can be found in enzymes such as imidazole glycerol phosphate synthase,38 cytidine triphosphate synthetase,52 methane monooxygenase hydroxylase,53 FabZ β-hydroxyacyl-acyl carrier protein dehydratase,54 and cytochrome P450.25,55 Even small gates of this type may require an activating agent to open. For example, the gate in the water channel of human monooxygenase CYP3A4 is created by the interaction of the conserved residue R375 with the heme, which opens upon cytochrome P450 reductase binding to the enzyme.56
Figure 4.

Relative occurrences of specific amino acid residues in wing and swinging door gates; 71 proteins with 129 gates were analyzed, and 154 residues that form wings or swinging doors were identified. Detailed description of the analyzed proteins is provided in Table 3. Values were normalized against the frequency with which each amino acid appears in all of the protein structures of the UniProtKB/Swiss-Prot database (2012_07).
3.2. Residue Motion: Swinging Doors
A more complex type of gate consists of two residues that can rotate but are stabilized in the closed conformation by a mutual interaction. Lario et al. introduced the phrase “swinging door” to describe gates of this type that were identified in cholesterol oxidase type I.57 Some swinging door gates open by having both residues rotate in the same direction, while in others the two residues rotate in opposite directions. Common stabilizing interactions in swinging door gates include π stacking as occurs in the F–F pair of cytochrome P4503A458,59 and acetylcholinesterase,60 ionic interactions as in toluene-4-monooxygenase61 and cytochrome P450cam, P450BM3, and P450eryF,25,55 aliphatic hydrophobic interactions such as those between the F–I, the F–V, and the F–L pairs of cytochrome P4503A4,58,59 aliphatic interactions such as those between the R–L and the L–I pairs of cytochrome P4503A4,58,59 and hydrogen bonds such as that between the R–S pair in cytochrome P4503A4.58 The open conformations of one or both of the gate residues may also be anchored, depending on the amino acids surrounding the gate. In comparison to wing gates, gates consisting of two residues can control wider tunnels and channels. It is worth mentioning that the individual residues that comprise a swinging door gate may simultaneously be components of another gate, as occurs in cytochrome P4503A4.58 Literature data indicate that most gates of this type consist of F–F pairs, and one way to screen for potential gates is to search for phenylalanine sandwiches.
3.3. Residue Motion: Apertures
Proteins undergo low-frequency breathing motions that may involve synchronized movements of bottleneck residues. In contrast to the previously described gates, the residues that form aperture type gates do not need to rotate and can maintain a rigid conformation. Their movements occur as a result of the synchronized relocalization of the enzyme backbone during its breathing motions. The ability of a given species to pass through gates of this type depends on the length of time the gate remains in the open state, which is determined by the enzyme’s rigidity (especially in terms of the compartments housing the gating amino acids) and the strength of the interactions between the gating residues. Gates of this kind can therefore switch between states at different frequencies, which can be adjusted by mutating the gating residues. Aperture-type gates have been identified in several enzymes including carbamoyl phosphate synthetase,36 choline oxidase,31 glutamate synthases,62 extradiol dioxygenases-homoprotocatechuate 2,3-dioxygenase,63 cytochrome P450eryF,25 and acetylcholinesterase.64
3.4. Motions of Loops and Secondary Structure Elements: Drawbridges and Double Drawbridges
The movements of loops and secondary structure elements can provide an energetically favorable method of controlling access for larger ligands. The gates described above consist of individual residues and would not provide sufficient control for enzymes that have large substrates and correspondingly large active site cavities. In many cases, the loops involved in access control also contribute to formation of substrate/cofactor binding cavities. Alternatively, in enzymes with complex systems of internal tunnels such as the members of the cytochrome P450 family, the dynamic motion of the protein structure, especially the flexible B–C and F–G loops in the cytochromes P450, plays a vital role in the opening and closing of the tunnels.16 Protein motions of this type can also merge different tunnels, creating a wider opening. Here, gating elements control the access of large substrates by merging and dividing the space shared by the tunnels.16 However, in such cases the movements of the loops can cause formation of smaller and more selective gates such as the swinging doors described in the preceding sections.56,58
Movements of loops and secondary structure elements can change the solvation of a cavity or the gate itself. The equilibrium between the open and the closed conformations depends on the anchoring residues and the flexibility of the hinge region. All of these elements play important roles in the movements of large gates. The conserved GxG motif found in most cytochrome P450 family members provides a good example.65 Depending on cytochrome isoform, the motif flanks either one or both ends of the B–C loop. It increases the flexibility of the loops, lowers the energy required for their motion, and facilitates tunnel opening and closing. The loops’ variable lengths and levels of flexibility mean that each member of the P450 family has a gate with unique properties.16
3.5. Domain Motion: Shell
Large domain motions in enzymes are not generally regarded as gating systems. “Gates” of this scale are common in ion channels, which are beyond the scope of this review.14 However, one might expect that such large gates could be present in enzymes that catalyze reactions of very large substrates. Indeed, a gate of this kind has been observed in RNA polymerase, whose clamp domain opens to permit entry of promoter DNA during initiation, closes to establish a tight grip on the DNA during elongation, and then opens again to release the DNA during termination.66 Interestingly, movement of large domains may protect enzymes from small molecule leakage and control their transport through long tunnel networks. This has been observed in carbon monoxide dehydrogenase/acetyl coenzyme A synthase, which operates as a tetrameric complex of distinct subunits.67 The cap domain movement of epoxide hydrolase from Mycobacterium tuberculosis is another enzyme with a domain-scale gate that controls substrate access to the active site cavity.68 Monomers of phospholipase A2 control access to their interface and the active site by adopting a different conformation during dimer aggregation.69 Large domain movements often require an additional source of energy. For example, in the ATP-dependent protease HsIVU,70 ATP hydrolysis is required to initiate conformational changes and propagate them to the residues that form the gate.
Structures of the gates found in 71 different proteins were analyzed systematically, yielding the results presented in Tables 2 and 3. The most common gate types are wings, swinging doors, and drawbridges, while apertures, double drawbridges, and shells are less common (Figure 5). However, these numbers may be distorted by the difficulty of identifying different gate classes by experimental methods or molecular dynamics simulations. Movements of only a few residues are more easily captured than those of secondary structure elements or domains due to the different time scales involved and the sizes of the moving structures. Moreover, gates may be controlled by overlapping processes that occur on different time scales and affect different structural regions.50
Table 2. List of Enzymes Possessing Gates Described in the Scientific Literature with Indication of Their Function, Structural Basis, and Location.


Table 3. Detailed Description of Enzymes Possessing Gates Presented in the Scientific Literature.
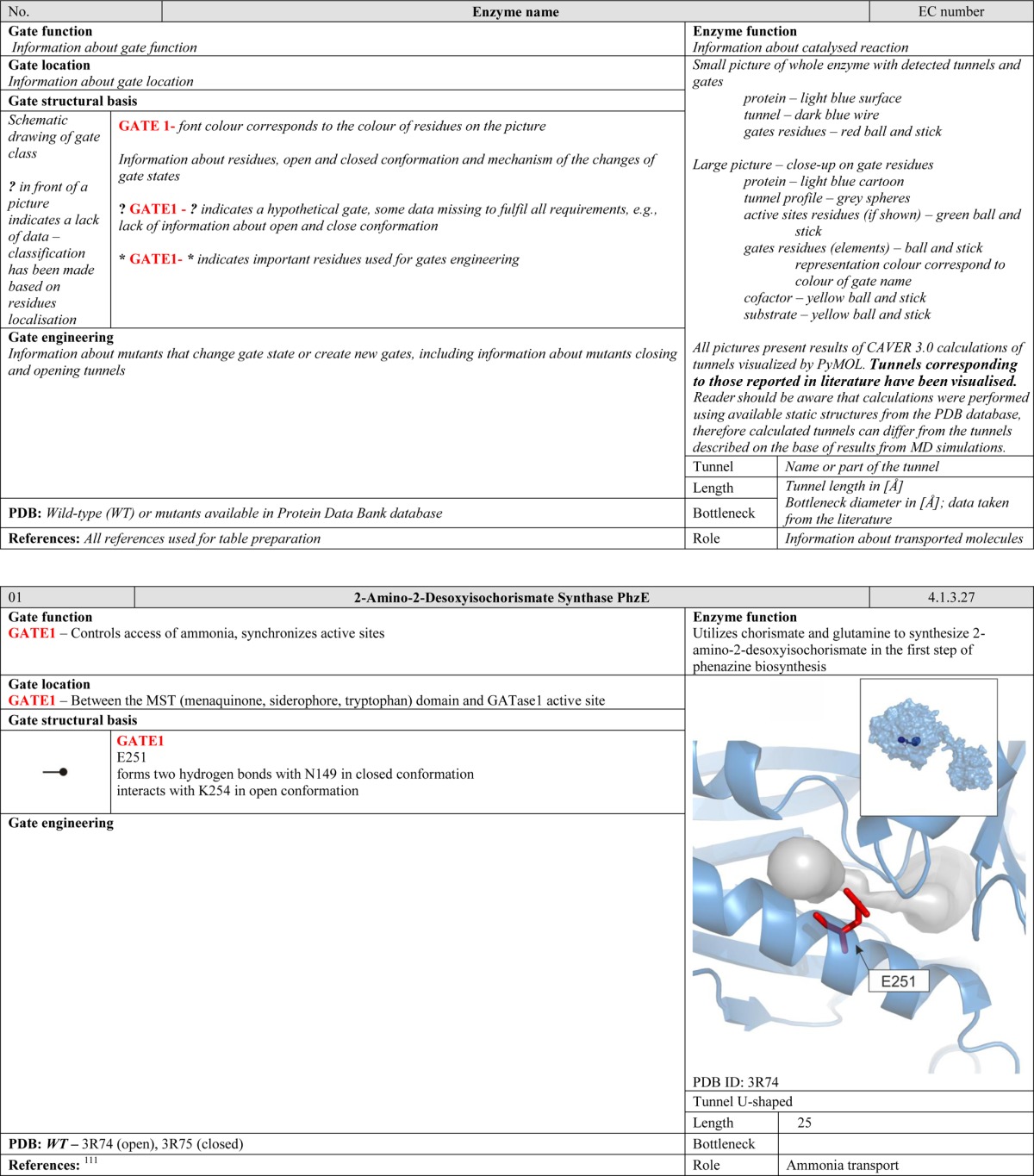

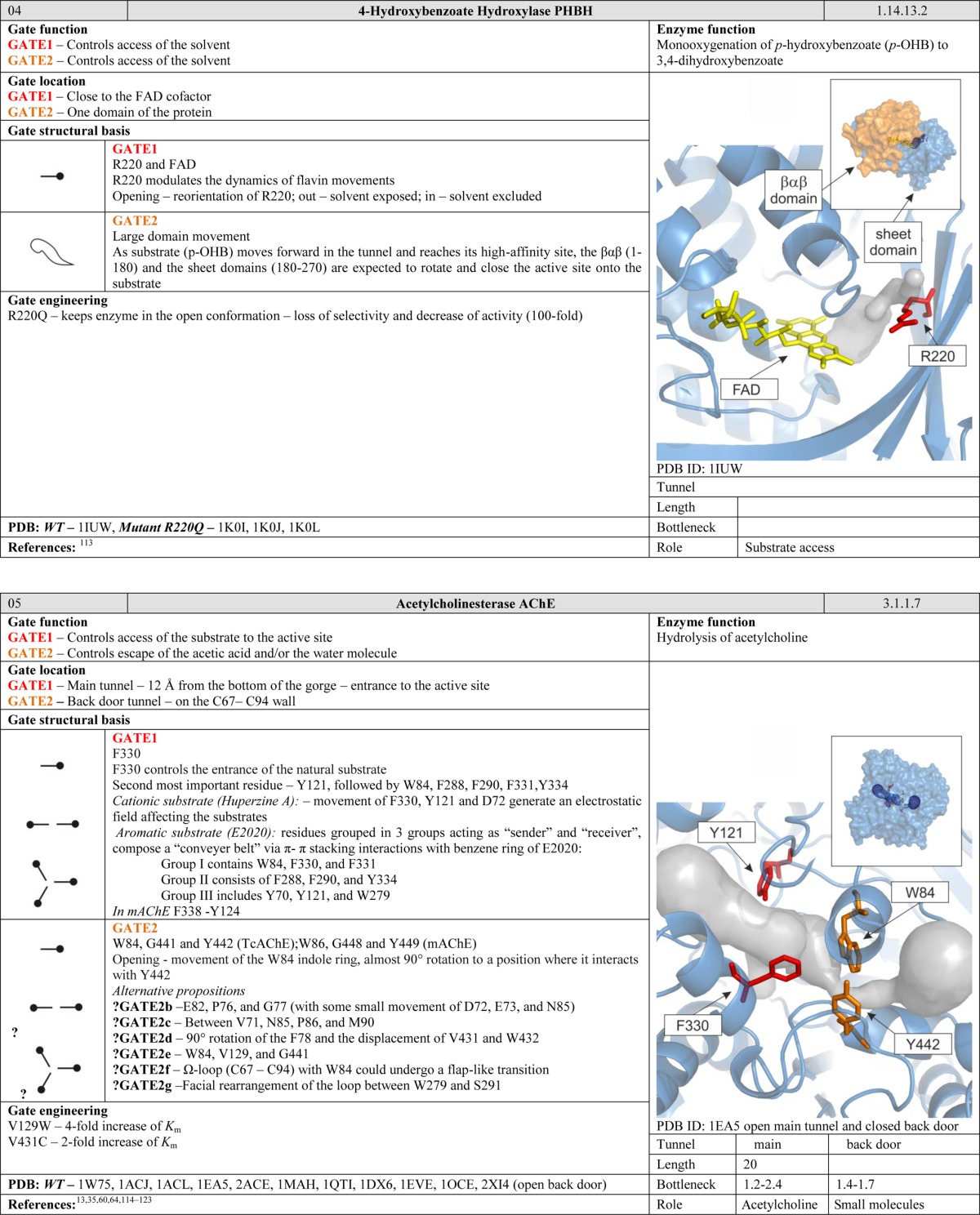


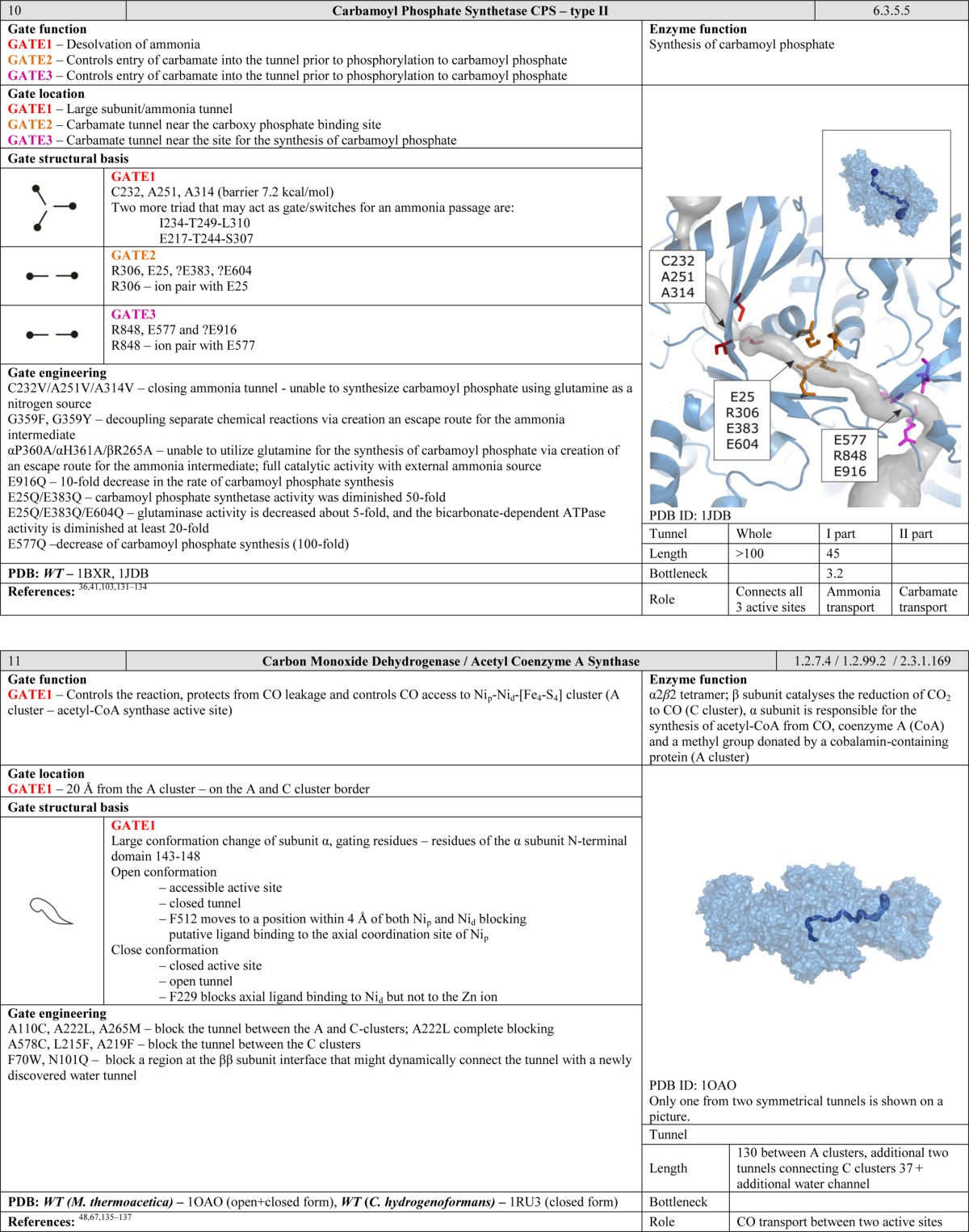

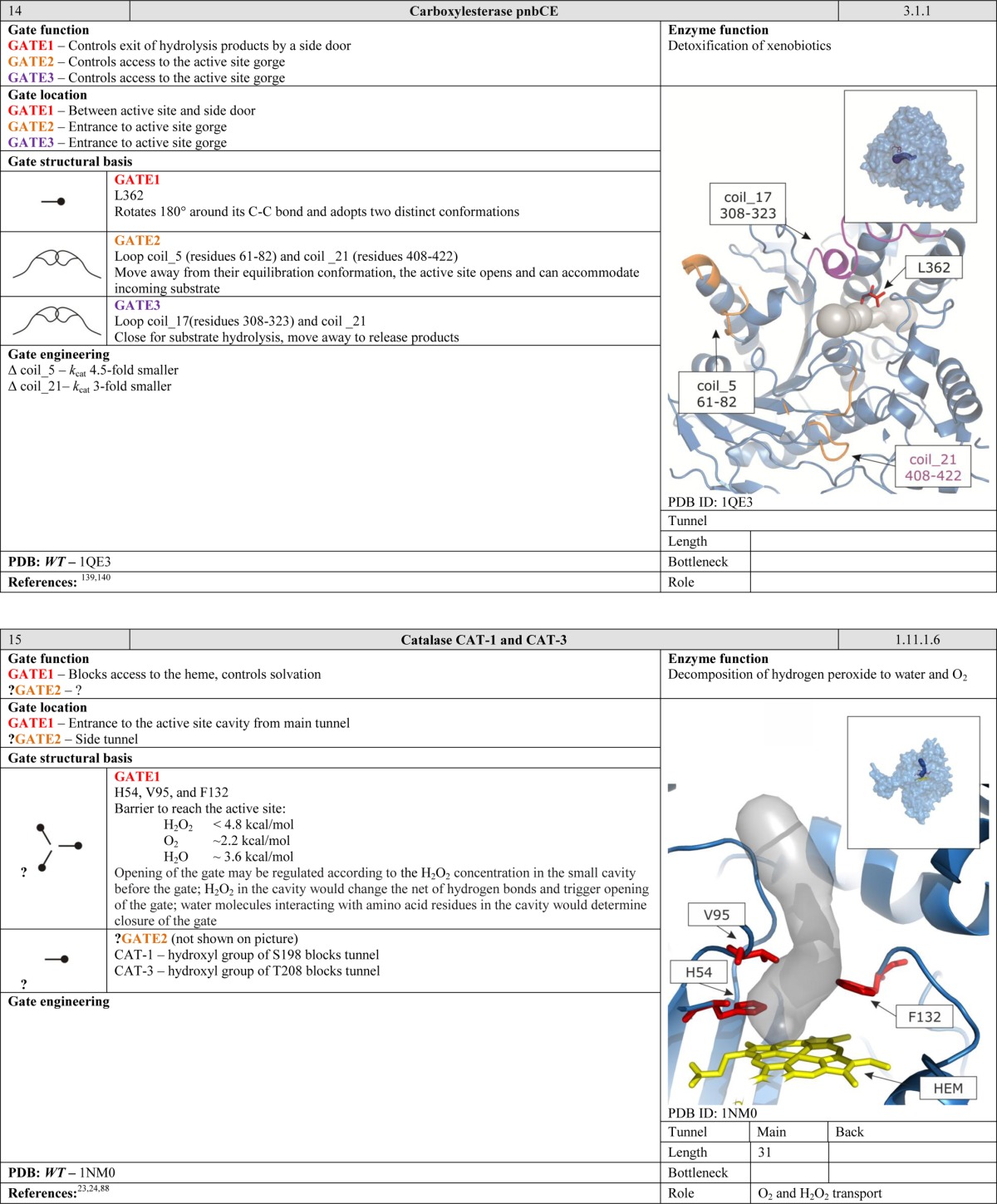
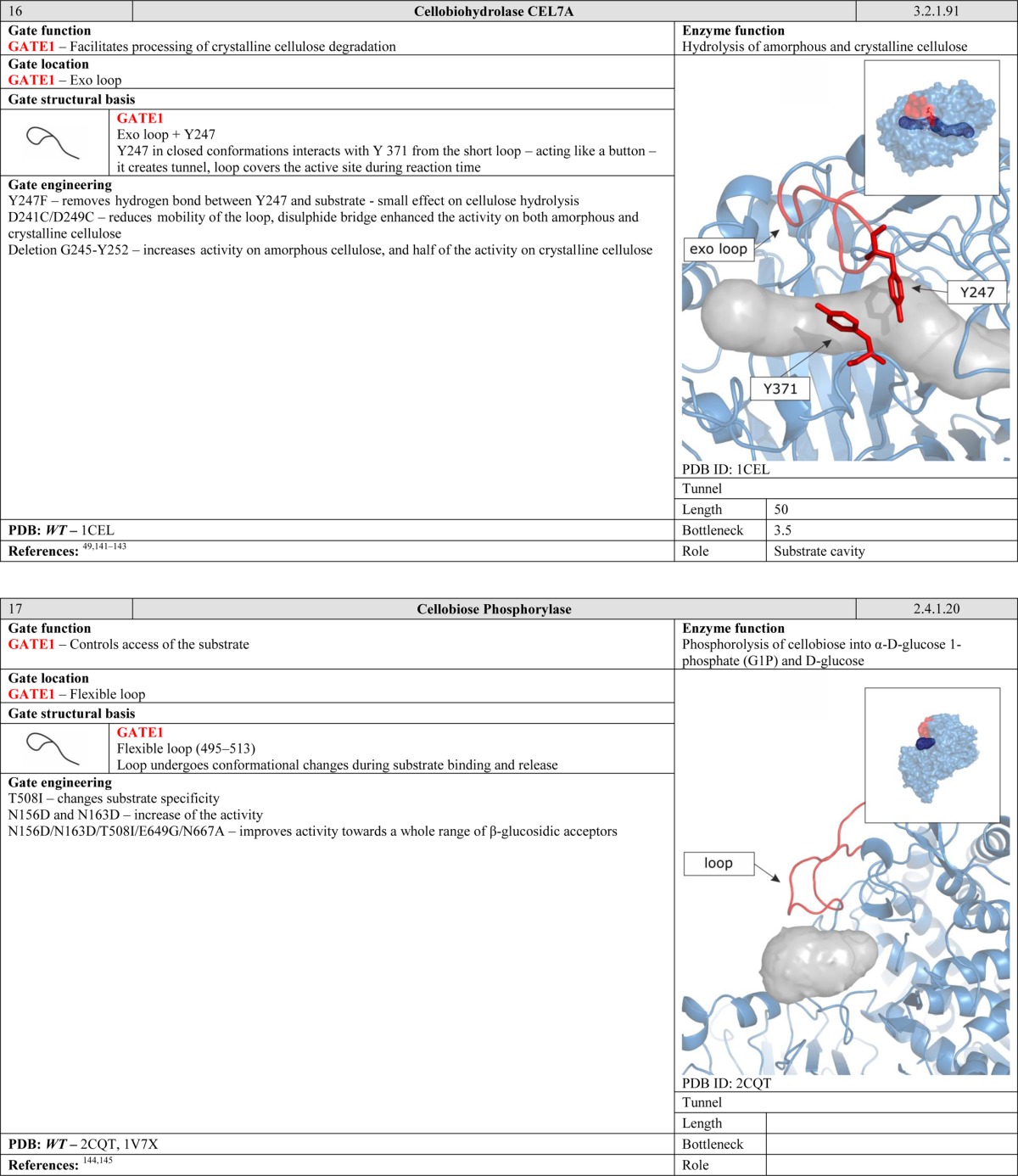

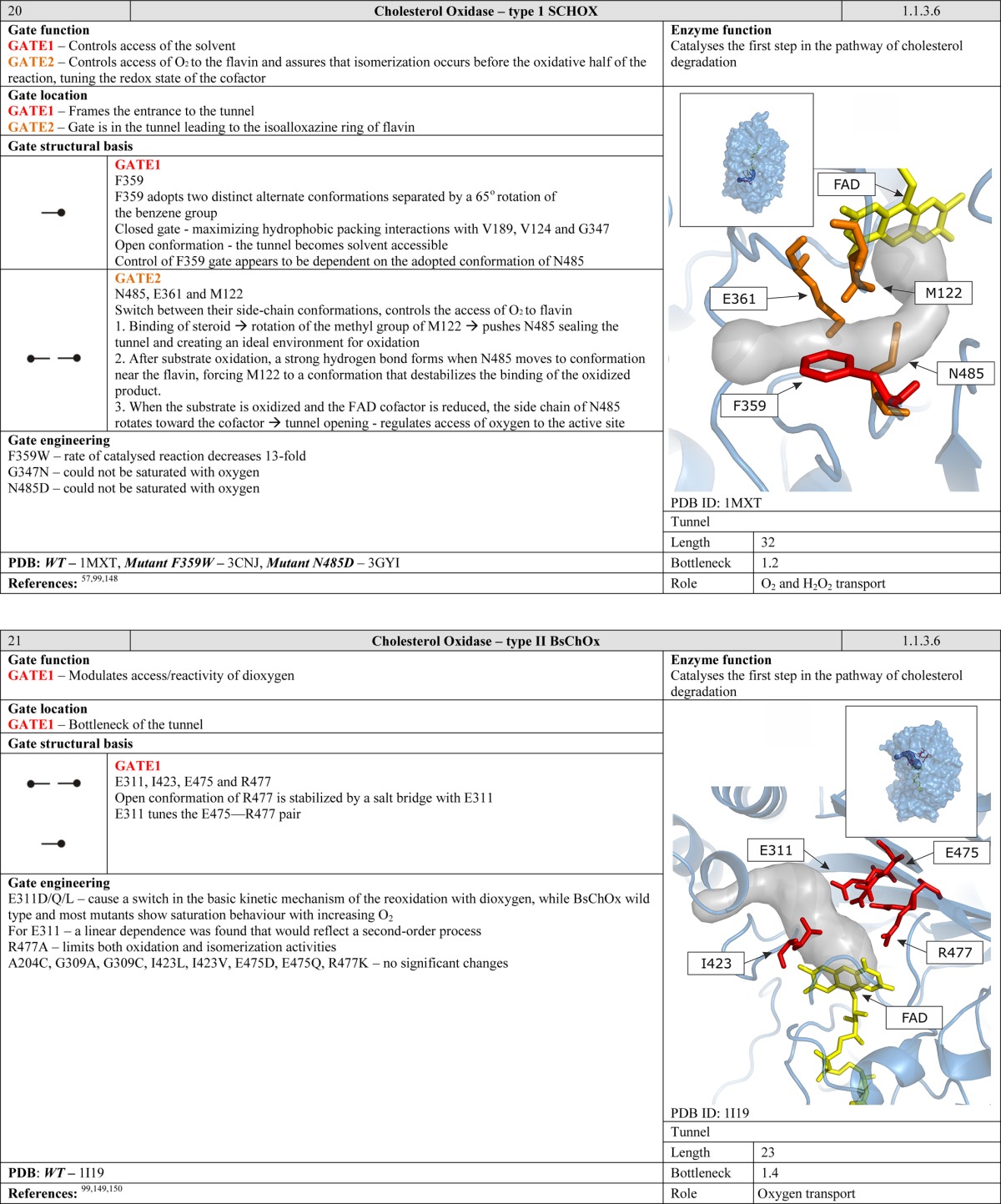



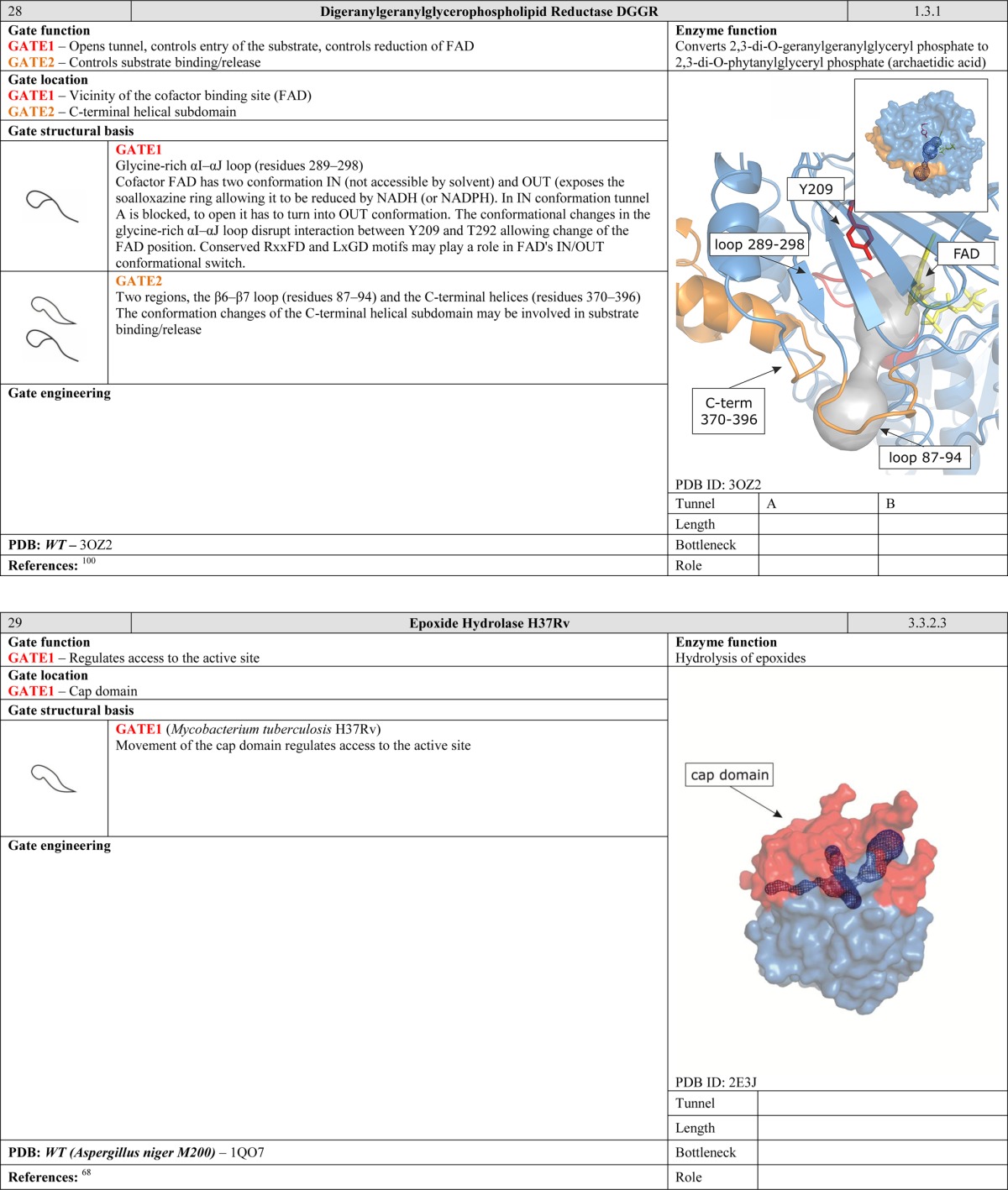
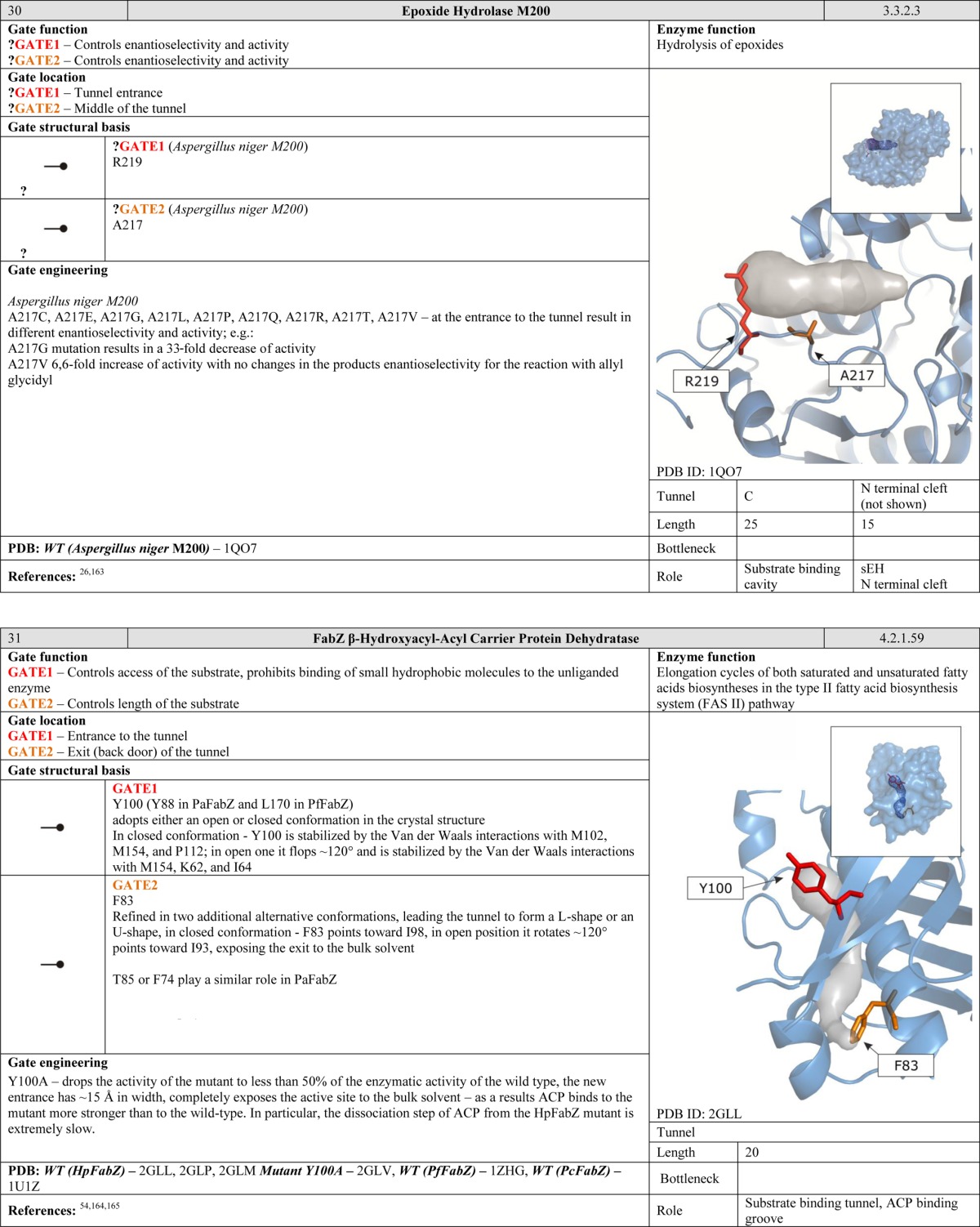
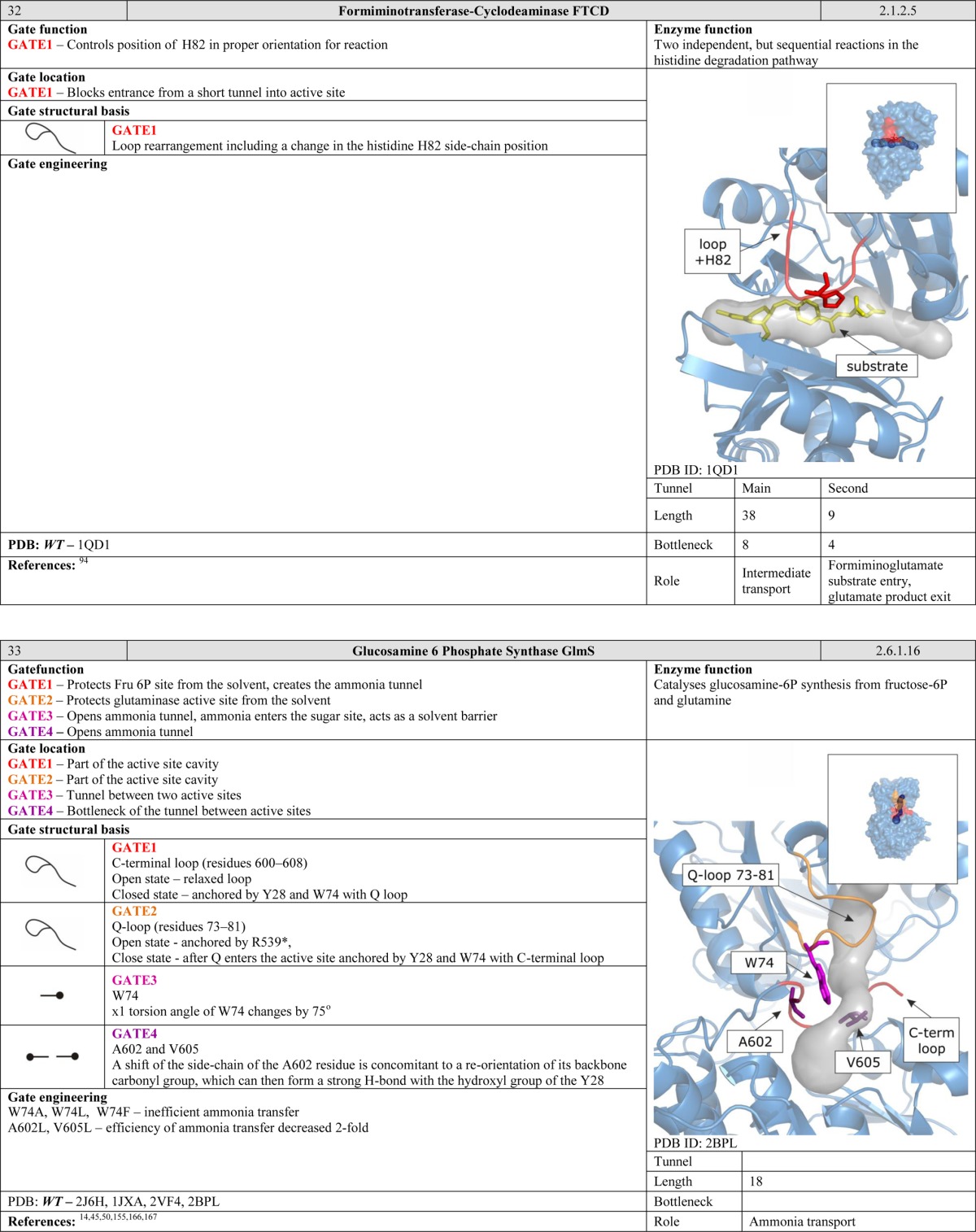
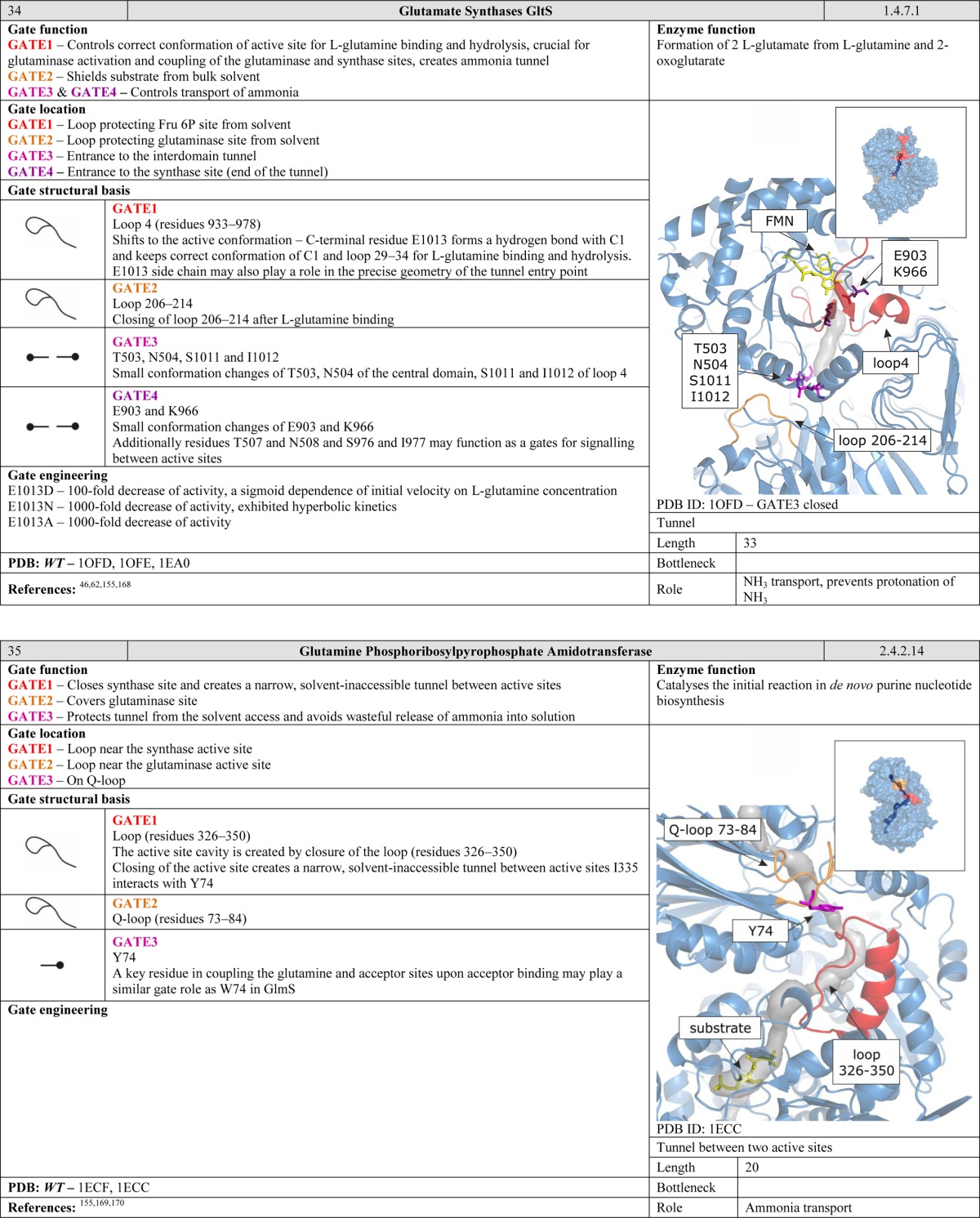
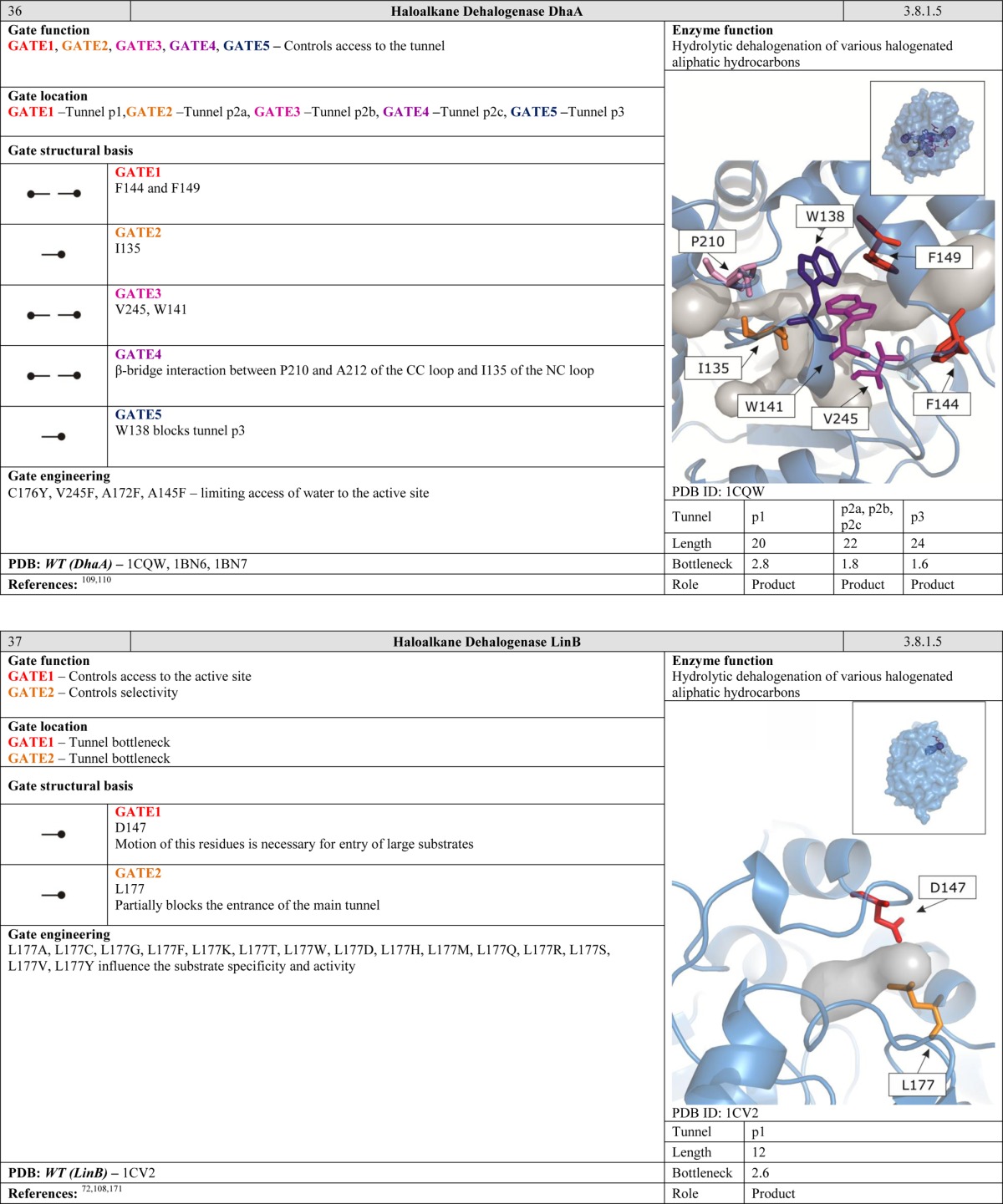
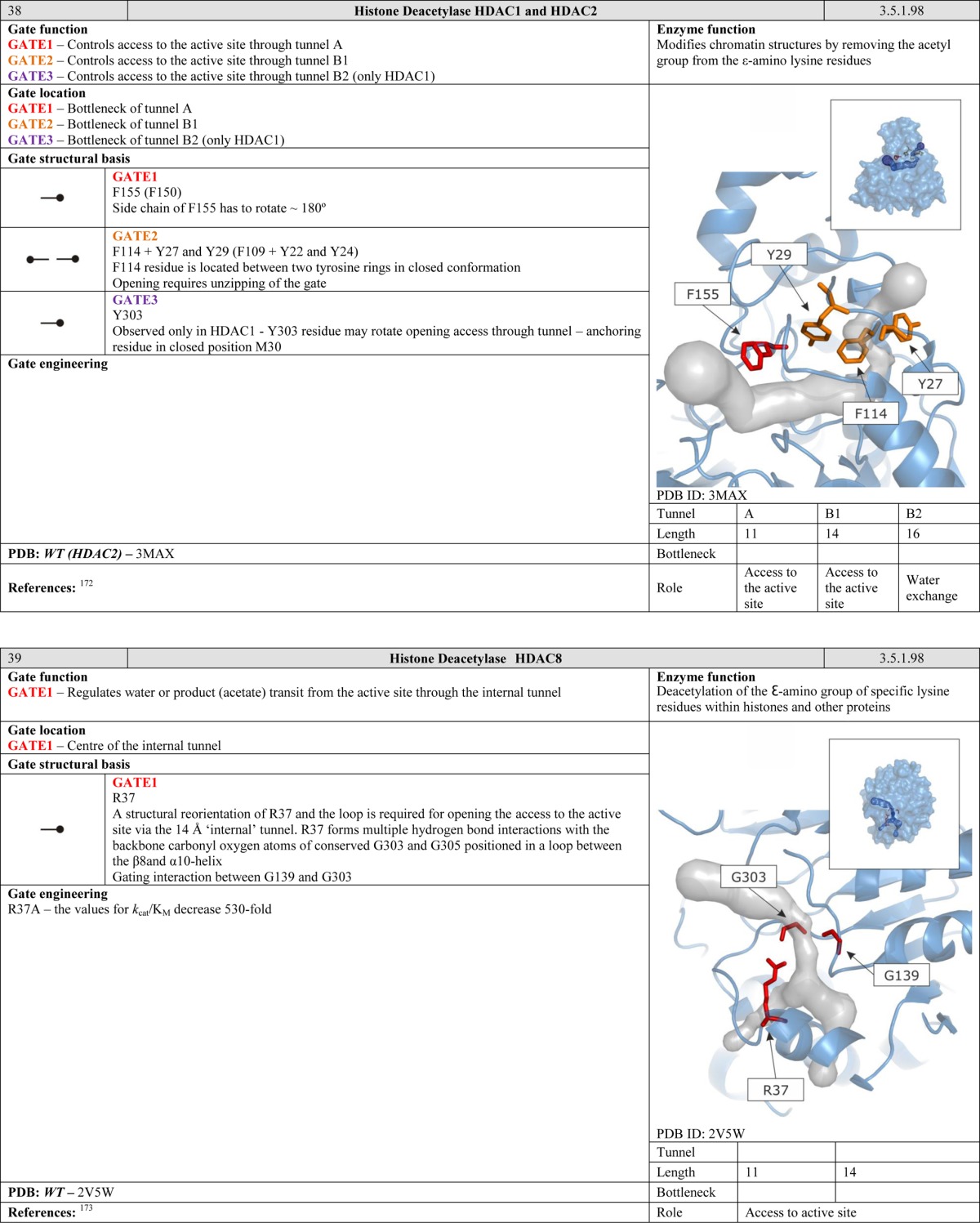











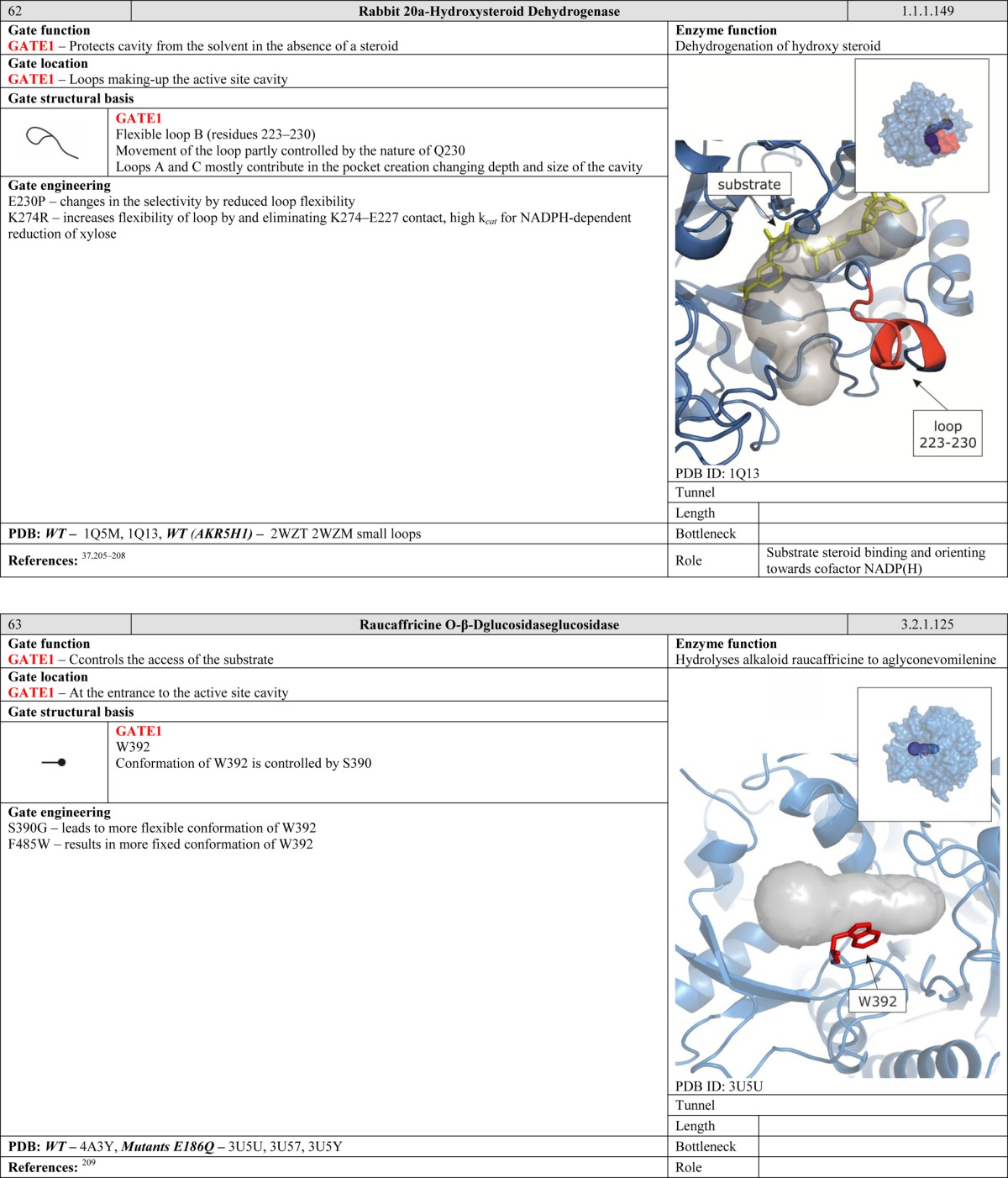



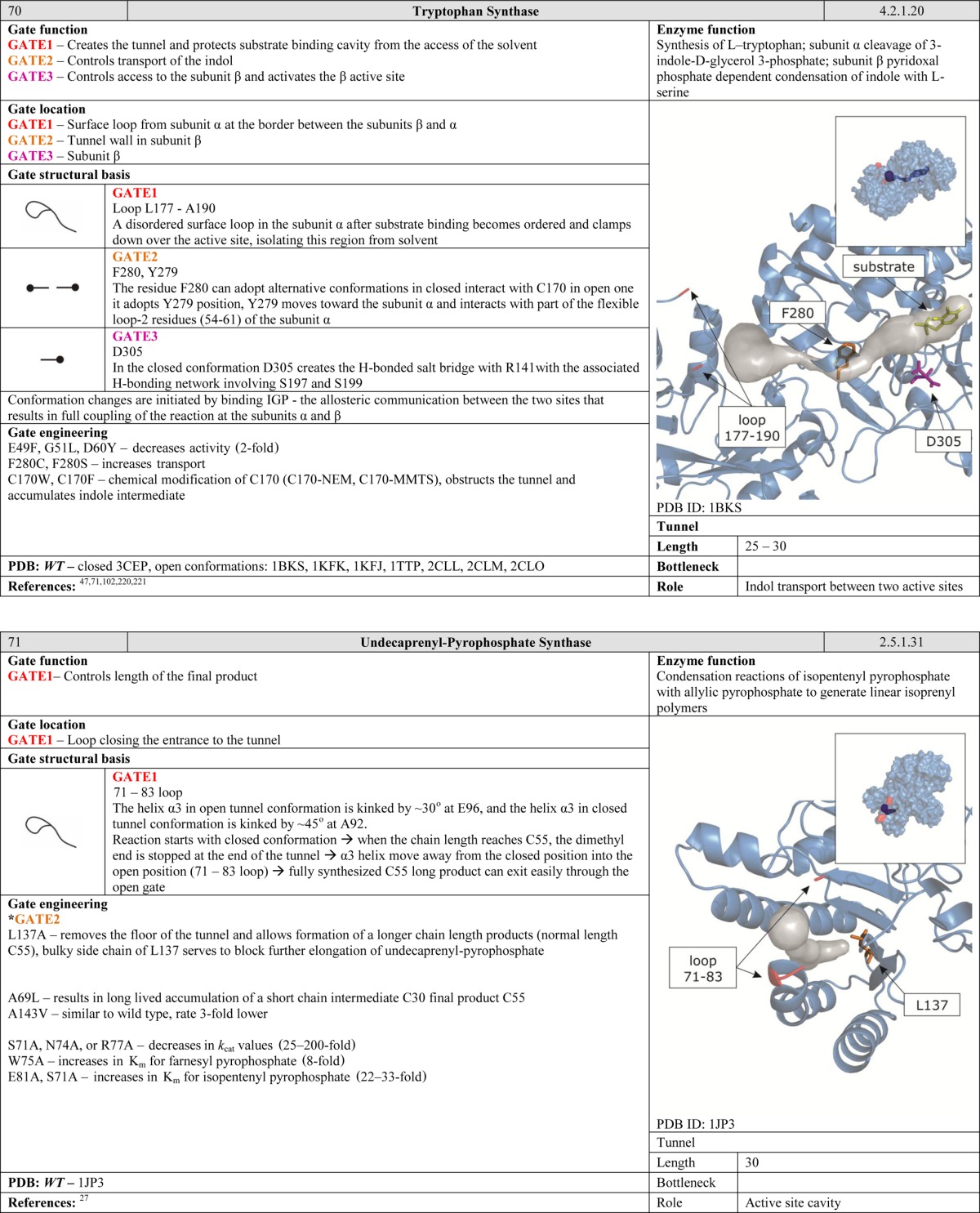
Figure 5.
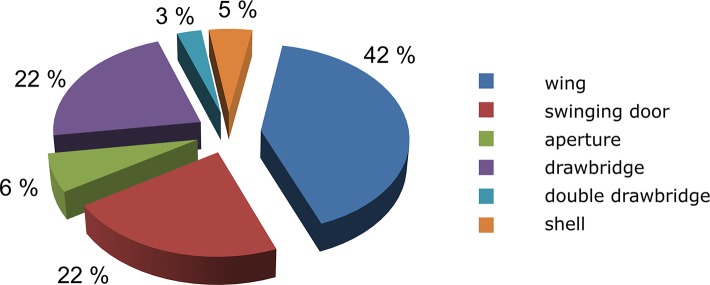
Frequencies of different gate types based on analysis of 71 proteins with 129 gates. Detailed description of the analyzed proteins is provided in Table 3.
4. Locations of Gates
The roles of gates in the enzymatic catalysis discussed above suggest that these structures are natural hot spots for modifying enzyme properties. Identification of structural components of natural gates would therefore be very useful to protein designers. This raises a question: how and where should one look for the gates? Gates in proteins can be identified experimentally by protein crystallography and NMR spectroscopy and computationally by molecular dynamics simulations and normal-mode analysis.
Crystallographic analyses can provide information on alternative configurations of particular amino acids in a protein structure. The presence of residues that can adopt both open- and closed-type configurations along the access or release pathways of ligands, ions, and solvents may suggest the presence of a gate. However, in order for a gate to be detected by crystallography, it is necessary for both the open and the closed conformations to be sufficiently represented. Despite this restriction, crystallographic analyses have identified gates in tryptophan synthase,71 haloalkane dehalogenase LinB,72l-amino acid oxidase,73 and toluene-o-xylene monooxygenase.74 In some cases, only one conformation will be present in the solved crystal structure, which may create a somewhat distorted picture, suggesting the absence of a gate when the open conformation is stabilized or the absence of a pathway when the closed conformation is stabilized.31,61,75
The limitations of crystallographic analysis can be overcome by advanced NMR spectroscopy, which makes it possible to study multiple protein conformations simultaneously, over time scales ranging from picoseconds to milliseconds.7,76 Such analyses provide information on both the open and the closed states as well as the population of each state and rate of their interconversion. NMR techniques have been used to measure the rate of exchange between the open and the closed conformations of triosephophate isomerase,77,78 HIV-1 protease,79 and dihydrofolatereductase.80,81 Overall, the utility of NMR for studying distant effects of mutations on protein dynamics suggests that it has great potential for investigating gating mechanisms.9
Some of the most useful tools for identifying gates are the computer programs developed for detecting tunnels, channels, and cavities in protein structures.82 The outputs of CAVER,83 MOLE,84 and MOLAXIS85 can be analyzed to detect bottleneck residues that form a potential gate or identify the best position for introduction of a new gate. Mutations at these “hot spots” can provide enzymes with new selectivities or activities.86 Zawaira et al.59 used the CAVER software together with the Protein Interaction Calculator87 for identifying gating residues within the cytochrome P450 family.
MD simulations are well suited for identification and analysis of gates and their behavior over time. Detailed descriptions of MD methods and their applications in simulating ligand migration can be found in recent reviews.9,88 Movements of large protein fragments on microsecond time scales can be investigated using Brownian dynamics,31,89 while Random Expulsion Molecular Dynamics and Steered Molecular Dynamics can be used to study pathways dedicated to transport of specific ligands.58,90 Some proteins have multiple pathways, each of which accommodates a different ligand or ligand class. This may in fact be a lot more common than is currently realized and can dramatically increase the complexity of gating systems arising from protein movement and the difficulty of identifying the true gating residues. For example, different residues control the ability of inhibitors E2020 and Huperzine A to access the active site of Torpedo californica acetylcholinesterase.60 Similarly, in cytochrome P4503A4, different residues in the same tunnel control access of temazepan and testosterone-6OH.58 The importance of a gating residue identified by computational methods can be confirmed experimentally by site-directed mutagenesis and kinetic experiments.
Studies using the experimental and theoretical approaches for gate identification discussed above have demonstrated that their locations within the protein can vary widely. Gates have been observed (i) at the entry to the active site or even directly inside the active site, (ii) at the entry or in the bottleneck of the protein tunnel connecting the buried active site to the protein surface or connecting two active site cavities, and (iii) at the interface of the cofactor and active site cavities (Figure 6).
Figure 6.
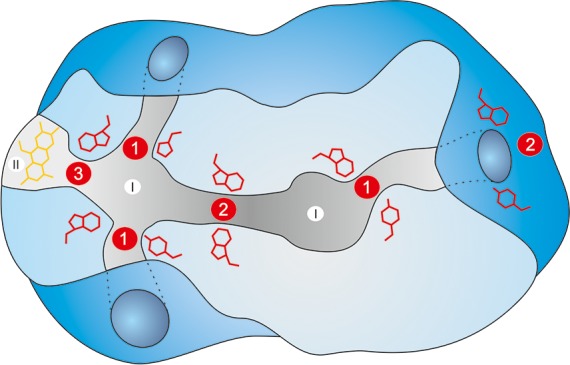
Locations of gates within a protein structure. Schematic representation of an enzyme with two active sites connected by a tunnel (I), a cofactor cavity (II), and multiple access tunnels. Gating residues in red may be located at the entrance to the active site (1), at the entrance or the bottleneck of the tunnel (2), and between the active site cavity and the cofactor cavity (3).
4.1. Active Site Entrance and Active Site
The entrance to the active site cavity is a suitable location for a gate, and gates situated here can have strong effects on enzyme activity. In some cases, the gating residues may even be a part of the active site.91 The simplest gates serve as filters that discriminate between potential substrates and thus play an important role in controlling enzyme selectivity. More advanced systems can prevent substrate entry when the active site residues are not properly oriented, e.g., in enzymes that require conformational changes before substrate binding. Many enzymes have gates at the entrance to their active sites, including acetylcholinesterase,60 imidazole glycerol phosphate synthase,38 glutamate synthase,46 toluene-o-xylene monooxygenase,91 monooxygenase,92 choline oxidase,31 NiFe hydrogenases,21 carbonic anhydrases,93 formiminotransferase-cyclodeaminase,94 type III polyketide synthases,95 and FabZ β-hydroxyacyl–acyl carrier protein dehydratase.54
4.2. Tunnel Entrance and Tunnel Bottleneck
The ability of ligands and solvent molecules to move from the media surrounding the protein to the active site can be controlled by gates located at any point along the tunnel. Gating residues may be situated at the tunnel entrance. However, it is more common to find them at the tunnel bottleneck. The tunnel entrance refers to the first shell of residues that define the tunnel and have contact with the bulk solvent. The tunnel bottleneck refers to the narrowest part that can be positioned anywhere along the tunnel (Figure 6). Even a single large residue whose side chain can project into the interior of the tunnel can exert efficient control over the access pathway. One might speculate that it might be favorable to have gates located inside tunnels because this allows their position to be more tightly controlled; their movements are restricted by the surrounding residues, and both the open and the closed conformations can be stabilized via interactions with neighboring amino acids. In contrast, residues located on the surface of the protein possess more degrees of freedom, and it is rare for both the open and the closed conformations to be stabilized. Examples of such gates inside the tunnels can be found in cholesterol oxidase type I,57 toluene-4-monooxygenase,61 undecaprenyl-pyrophosphate synthase,27 homoprotocatechuate 2,3-dioxygenase,63 4-hydroxy-2-ketovalerate aldolase/acylating acetaldehyde dehydrogenase,96 epoxide hydrolase from Aspergillus niger M200,26 and FabZ β-hydroxyacyl-acyl carrier protein dehydratase.54 Similarly, gates can be situated in the bottlenecks of tunnels connecting two active sites. Gates in such positions are essential for enzymes that catalyze two reactions requiring different environments, such as glucosamine 6 phosphate synthase,45 imidazole glycerol phosphate synthase,38 cytidine triphosphate synthetase,52 carbamoyl phosphate synthetase,36 and glutamate synthases.46
4.3. Cofactor Cavity
Gates can be positioned at the interface of the active site and the cofactor cavity, allowing for more fine-grained control during the reaction. In NADH oxidase, the W47 residue acts as a gate that controls the accessibility of the FAD flavin ring and thus plays a crucial role during the catalytic cycle. The closed conformation is stabilized by hydrogen bonds between the cofactor and the peptide backbone, whereas stabilization of the open form may be advantageous during the initial steps of substrate binding since it is believed to slow down product dissociation.22,97 Other gates of this type have been reported in 3-hydroxybenzoate hydroxylase,98 4-hydroxy-2-ketovalerate aldolase/acylating acetaldehyde dehydrogenase,96 and cholesterol oxidase type I57 and type II.99 Moreover, cofactors themselves can also function as gates. The FAD cofactor of digeranylgeranylglycerophospholipid reductase has two different conformations, referred to as the “in” and “out” conformations. In the “in” conformation, the tunnel is blocked by FAD. To open the tunnel, FAD has to adopt the “out” conformation.100
Gates are most commonly located at the tunnel entry and the tunnel bottleneck (51%). This is to be expected because the bottleneck represents the narrowest point of the tunnel, and its diameter often dictates the tunnel’s permeability. Another common location is the entrance to the active site cavity (28%). Gates at the entrance of the cofactor cavity are less common (5%), which is not surprising since not all enzymes have a cofactor cavity. In 16% of the cases studied, the gate was not located within any of these functional regions (Figure 7).
Figure 7.
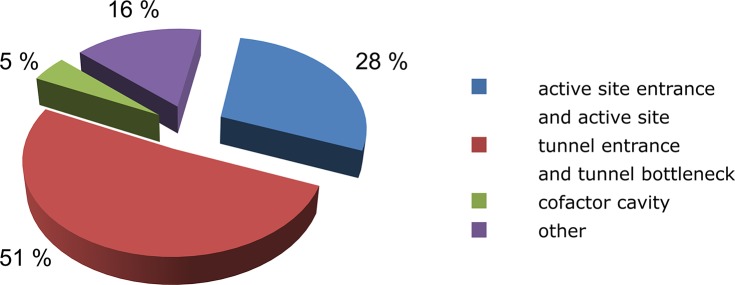
Distribution of gate locations within protein structures based on analysis of 71 proteins with 129 gates. Detailed description of the analyzed proteins is provided in Table 3.
5. Engineering of Gates
The average rate of evolution of the gating residues in the cytochrome P450 family is significantly greater than that for the protein sequence as a whole.59 The high rate of evolution at the gating residues suggests that gate engineering may be an attractive alternative to other rational enzyme design strategies. This idea is supported by a few observations: (i) the gates are often spatially separated from the active site, and so mutations at the gating residues should not be deleterious to protein function, (ii) the opening and closing of the access pathways can affect ligand exchange and thus enzyme activity and selectivity, and (iii) gate modification can modulate the solvent’s ability to access the active site, which in turn affects solvation and stabilization of the transition state and also product release. In the following section, we describe selected cases in which an enzyme’s catalytic properties have been successfully altered by modifying its gates.
5.1. Gate Modification
Gates can be modified by substitution of the gating residues, hinge residues, or anchoring residues depending on the nature of the gate in question. Modification of the gating residues has been shown to change the selectivity and activity of toluene-o-xylene monooxygenase, with the E214G mutation improving oxidation of p-nitrophenol by a factor of 15.101 A similar improvement in overall activity was achieved in a lipase from Burkholderia cepacia by the mutations L17S + L287I. This double mutant also exhibited a 10-fold increase in enantioselectivity compared to the wild-type enzyme.86 The T78F or P76F mutations in imidazole glycerol phosphate synthase override some of the control exerted by the wild-type gate and block the passage of ammonia through the tunnel.38 In NiFe hydrogenases, mutations of the V74 and V74 + L122 residues changes the rates of transport for H2, CO, and O2, thereby modulating the overall rate of reaction.21
Gates that incorporate secondary structure elements are dependent on hinge and anchoring regions. It has been demonstrated that modification of the hinge region can change an enzyme’s activity and selectivity. Notably, the Q230P mutation in rabbit 20A-hydroxysteroid dehydrogenase decreases the flexibility of a key loop and thereby changes its selectivity.37 Similarly, access to the active site in the HIV-1 protease is controlled by two β-turn flaps.89 Results from Brownian dynamics simulations suggest that the G48V + V82A or I84V + L90M mutations in this enzyme reduce the likelihood that the active site will be exposed at any given point in time from 14% in the wild type to 2% in the mutants.89 The importance of the anchoring residues in the HIV-1 protease was demonstrated by the F53L mutation, which generates a semiopen conformation due to removal of the stabilizing F53–I50 interaction.89
5.2. Gate Removal
Removing gates typically increases substrate and product exchange rates but also allows more extensive access of water molecules to protein tunnels and cavities. The overall effect of gate removal is therefore equal to the combined effects of these two processes. Gate deletion has been shown to increase the rate of substrate binding to tryptophan synthase.102 The F280C and F280S mutations both increased the rate of indole binding by a factor of 2. Similarly, the T78A mutation allowed ammonia to rapidly pass through the tunnel in imidazole glycerol phosphate synthase.38 The R239A mutation in the cyclase caused a 1000-fold decrease in the enzyme’s kcat/Km value and decoupling of the reaction.38 This dramatic change in enzyme catalytic efficiency was attributed to creation of a new route for ammonia release. A similar leakage of ammonia was caused by the G359F and G359Y mutations in carbamoyl phosphate synthetase.103,104 Negative consequences of gate removal were also observed for the FabZ-β-hydroxyacyl-acyl carrier protein dehydratase (HpFabZ),54 in which the Y100A mutation leaves the active site completely exposed to the bulk solvent. As a result, the acyl carrier protein binds to the HpFabZ Y100A mutant much more strongly than to the wild-type HpFabZ, decreasing the mutant enzyme’s activity by more than 50% due to the very slow dissociation of the acyl carrier protein.
In some cases, gate removal enables bulky substrates to access the active site cavity. Mutations D285I and D285Q in toluene-4-monooxygenase improved its ability to oxidize the large and bulky substrates 2-phenylethanol and methyl p-tolyl sulfide by factors of 8 and 11, respectively, while the D285S mutation improved the rate of styrene oxidation 1.7-fold.105 The L137A mutation in undecaprenyl-pyrophosphate synthase removed the bottom of the tunnel in this enzyme, allowing formation of products with longer chain lengths.27 A similar situation was encountered in type III polyketide synthases from Aloe arborescens PCS, in which the M207 residue controls the number of condensations of malonyl-CoA.106 The M207G mutation opened a connection between the tunnel and two hidden pockets located behind the active site, resulting in formation of extended products. Further product elongation was achieved with the triple mutant F80A + Y82A + M207G.
5.3. Gate Insertion
To best of our knowledge, there have been no reports of an intentional introduction of a new gate into an enzyme structure. However, there have been studies in which an access tunnel was systematically modified with multiple substitutions, and it is reasonable to expect that some of these mutations might have created new gates. More research will clearly be needed to confirm this expectation. To verify successful intentional insertion of a new gate into a protein structure, it would be necessary to confirm the existence of both the open and the closed conformations at a position where previously only a single conformation could be adopted.
Site-directed mutagenesis targeting specific residues at various positions along the access tunnel of Candida rugosa lipase has been used to alter the acceptable substrate chain length for this enzyme. The mutants for which this was observed were P246F, L413F, L410W, L410F + S300E, and L410F + S365L.107 We note that the aromatic residues F and W, which are common in wing and swinging door gates, were introduced in each of these variants.
In another study, the residue L177 that is located near the entrance to the access tunnel of the haloalkane dehalogenase LinB from Sphingobium japonicum UT26 was substituted with all of the natural amino acids, yielding 19 mutants with significantly altered substrate specificity and activity.108 Preliminary computational analyses of these variants using molecular dynamics revealed that the two residues possessing a single aromatic ring (F and Y) exhibited large fluctuations, as might be expected for gating.
Residue A217 is located at the entrance to the tunnel in the epoxide hydrolase EH from Aspergillus niger M200. This residue was substituted with C, E, G, L, P, Q, R, T, and V, and the effect of each mutation on the enzyme’s activity and enantioselectivity was studied.26 The mutants exhibited different enantioselectivity and activity relative to the wild type. For instance, the activity of the A217G mutation toward allyl glycidyl ether was lower than that of the wild type by a factor of 33, whereas the A217V mutation increased activity toward this substrate 6.6-fold.
Residues I135, W141, C176, V245, L246, and Y273 are positioned close to the entrance to the main and side tunnels of the haloalkane dehalogenase DhaA from Rhodococcus rhodochrous NCIMB 13064. These residues were simultaneously permuted in an attempt to improve this enzyme’s activity against 1,2,3-trichloropropane. The most successful mutant, which featured the I135F, C176Y, V245F, L246I, and Y273F substitutions, showed 26-fold greater activity toward the target substrate than did the wild type.109 In this mutant, three aromatic residues were introduced in place of aliphatic ones in the vicinity of the tunnels. Computational analysis of product release from the mutant suggests that substitutions introduced a transient rather than permanent structural feature and gating residues prevented access of water to the active site.110 Crystallographic analysis of the mutant revealed two distinct conformations for the Y176 side chain.110
6. Conclusions
This review highlights the importance of gates in enzymes. Gates play vital roles in controlling the catalytic activity and selectivity of enzymes and are more common in protein structures than is generally thought. In particular, gates control substrate access to the active site and product release, prevent or restrict solvent access to specific regions of the protein, and can synchronize processes occurring in distinct parts of the enzyme. Our literature survey of 129 gates in 71 enzymes revealed a large variety of systems with sophisticated structures. We presented a rigorous definition of gates and established a new scheme for their classification. The large number of inspected cases allowed us to build a catalogue of gates assigned to six distinct classes—wings, swinging doors, apertures, drawbridges, double drawbridges, and shells—with three different functions and three distinguishable locations. We also presented summary statistics that give a preliminary overview of the propensity of specific amino acid residues to occur in particular gate classes. The proposed classification scheme can be easily extended and updated but even in its present form can provide guidance for analysis and engineering of gates in biomolecular systems.
The biochemical relevance and specific location of gates within protein structures make them attractive targets for protein engineering. Attempts to rationally redesign gates typically involve computer-assisted gate identification followed by modification using focused directed evolution. This approach is compatible with a recent trend in protein engineering that stresses construction of small and smart libraries. Gate modification and deletion have been demonstrated in numerous cases, but the intentional insertion of new gates remains a challenge. Convenient methods for identifying gates in protein structures are essential prerequisites for their engineering. In silico, this can be achieved by coupling the software tools developed for describing pathways to tools developed for study of protein dynamics. Of the available experimental techniques, NMR spectroscopy is particularly suitable for analysis of highly dynamic protein structures and can be expected to play an indispensable role in the study of gate dynamics at the atomic level. The field would also benefit from development of new experimental techniques for monitoring the passage of ligands through the protein pathways. One day it will be possible to control the catalytic properties of enzymes by rational engineering of their gates. To achieve this goal, we have to learn how gates evolved, how they interact with the other parts of the protein structure as well as with the ligand and solvent molecules, and how they fulfill their biological functions.
Acknowledgments
This work was supported by the Grant Agency of the Czech Republic (P207/12/0775 and P503/12/0572), the Grant Agency of the Czech Academy of Sciences (IAA401630901) and the European Regional Development Fund (CZ.1.05/2.1.00/01.0001). The work of A.G. was supported by SoMoPro programme No. SIGA762 and has received a financial contribution from the European Union within seventh FP (FP/2007-2013) under grant agreement No. 229603 and is cofinanced by the South Moravian Region. MetaCentrum is acknowledged for providing access to computing facilities, supported by the Czech Ministry of Education of the Czech Republic (LM2010005).
Biographies

Artur Gora received his M.Sc. degree in Environmental Protection in 1998 and in Molecular Biology in 1999 and Ph.D. degree in Chemistry in 2002 from the Jagiellonian University in Poland. From 2002 to 2009 he carried out postdoctoral research at the Institute of Catalysis and Surface Chemistry of the Polish Academy of Sciences. In 2004 he was awarded a JSPS fellowship, and from 2004 to 2006 he was working at the National Institute of Advanced Industrial Science and Technology in Japan. Since 2010, he has been researcher at the Loschmidt Laboratories of the Masaryk University in Brno, Czech Republic. His recent research is focused on rational redesign of proteins.

Jan Brezovsky received his M.Sc. degree in Biophysics at Masaryk University, Brno, Czech Republic, in 2006. In 2011 he received his Ph.D. degree in Environmental Chemistry with his thesis “Molecular Modeling of Enzymes” at the same university. Since then, he has been working as junior researcher at Loschmidt Laboratories of Masaryk University focusing on computational protein engineering and molecular modeling. His main research topics are ration redesign of dynamics and selectivity of enzymes.

Jiri Damborsky is the Loschmidt Chair Professor at the Department of Experimental Biology and the Research Centre for Toxic Compounds in the Environment, Masaryk University, Brno, Czech Republic. He received his MSc. degree in 1993 and Ph.D. degree in Microbiology in 1997 at Masaryk University. His research interests lie in the field of protein engineering and synthetic biology. His research group develops new concepts and software tools for rational design of enzymes and bacteria. He is holder of the EMBO/HHMI Scientist award of the European Molecular Biology Organisation and the Howard Hughes Medical Institute.
The authors declare no competing financial interest.
References
- Jiang L.; Althoff E. A.; Clemente F. R.; Doyle L.; Röthlisberger D.; Zanghellini A.; Gallaher J. L.; Betker J. L.; Tanaka F.; Barbas C. F.; Hilvert D.; Houk K. N.; Stoddard B. L.; Baker D. Science 2008, 319, 1387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Röthlisberger D.; Khersonsky O.; Wollacott A. M.; Jiang L.; DeChancie J.; Betker J.; Gallaher J. L.; Althoff E. A.; Zanghellini A.; Dym O.; Albeck S.; Houk K. N.; Tawfik D. S.; Baker D. Nature 2008, 453, 190. [DOI] [PubMed] [Google Scholar]
- Nanda V.; Koder R. L. Nat. Chem. 2010, 2, 15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Richter F.; Leaver-Fay A.; Khare S. D.; Bjelic S.; Baker D. PLoS One 2011, 6, e19230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kamerlin S. C. L.; Warshel A. Proteins 2010, 78, 1339. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schwartz S. D.; Schramm V. L. Nat. Chem. Biol. 2009, 5, 551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Henzler-Wildman K.; Kern D. Nature 2007, 450, 964. [DOI] [PubMed] [Google Scholar]
- Henzler-Wildman K. A.; Thai V.; Lei M.; Ott M.; Wolf-Watz M.; Fenn T.; Pozharski E.; Wilson M. A.; Petsko G. A.; Karplus M.; Hübner C. G.; Kern D. Nature 2007, 450, 838. [DOI] [PubMed] [Google Scholar]
- Lee J.; Goodey N. M. Chem. Rev. 2011, 111, 7595. [DOI] [PubMed] [Google Scholar]
- Lüdemann S. K.; Lounnas V.; Wade R. C. J. Chem. Biol. 2000, 303, 797. [DOI] [PubMed] [Google Scholar]
- Csermely P.; Palotai R.; Nussinov R. Trends Biochem. Sci. 2010, 35, 539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McCammon J. A.; Northrup S. H. Nature 1981, 293, 316. [DOI] [PubMed] [Google Scholar]
- Zhou H.-X.; Wlodek S. T.; McCammon J. A. Proc. Natl. Acad. Sci. U.S.A. 1998, 95, 9280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou H. X.; McCammon J. A. Trends Biochem. Sci. 2010, 35, 179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McCammon J. A. BMC Biophys. 2011, 4, 4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cojocaru V.; Winn P. J.; Wade R. C. Biochim. Biophys. Acta 2007, 1770, 390. [DOI] [PubMed] [Google Scholar]
- Fischer E. Ber. Dtsch. Chem. Ges. 1894, 27, 2985. [Google Scholar]
- Koshland D. E. Proc. Nat. Acad. Sci. U.S.A. 1958, 44, 98. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsai C. J.; Kumar S.; Ma B.; Nussinov R. Protein Sci. 1999, 8, 1181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prokop Z.; Gora A.; Brezovsky J.; Chaloupkova R.; Stepankova V.; Damborsky J. Engineering of protein tunnels: Keyhole-lock-key model for catalysis by the enzymes with buried active sites. In Protein Engineering Handbook; Lutz S., Bornscheuer U. T., Eds.; Wiley-VCH: Weinheim, 2012; p 421. [Google Scholar]
- Liebgott P. P.; Leroux F.; Burlat B.; Dementin S.; Baffert C.; Lautier T.; Fourmond V.; Ceccaldi P.; Cavazza C.; Meynial-Salles I.; Soucaille P.; Fontecilla-Camps J. C.; Guigliarelli B.; Bertrand P.; Rousset M.; Leger C. Nat. Chem. Biol. 2010, 6, 63. [DOI] [PubMed] [Google Scholar]
- Volbeda A.; Martin L.; Cavazza C.; Matho M.; Faber B. W.; Roseboom W.; Albracht S. P.; Garcin E.; Rousset M.; Fontecilla-Camps J. C. J. Biol. Inorg. Chem. 2005, 10, 239. [DOI] [PubMed] [Google Scholar]
- Amara P.; Andreoletti P.; Jouve H. M.; Field M. J. Protein Sci. 2001, 10, 1927. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kalko S. G.; Gelpí J. L.; Fita I.; Orozco M. J. Am. Chem. Soc. 2001, 123, 9665. [DOI] [PubMed] [Google Scholar]
- Winn P. J.; Ludemann S. K.; Gauges R.; Lounnas V.; Wade R. C. Proc. Natl. Acad. Sci. U.S.A. 2002, 99, 5361. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kotik M.; Stepanek V.; Kyslik P.; Maresova H. J. Biotechnol. 2007, 132, 8. [DOI] [PubMed] [Google Scholar]
- Ko T. P.; Chen Y. K.; Robinson H.; Tsai P. C.; Gao Y. G.; Chen A. P. C.; Liang P. H.; Wang A. H. J. J. Biol. Chem. 2001, 276, 47474. [DOI] [PubMed] [Google Scholar]
- Szabo A.; Shoup D.; Northrup S. H.; McCammon J. A. J. Chem. Phys. 1982, 77, 4484. [Google Scholar]
- Zhou H. X.; Szabo A. Biophys. J. 1996, 71, 2440. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou H.-X. J. Chem. Phys. 1998, 108, 8146. [Google Scholar]
- Xin Y.; Gadda G.; Hamelberg D. Biochemistry 2009, 48, 9599. [DOI] [PubMed] [Google Scholar]
- Kramers H. A. Physica 1940, 7, 284. [Google Scholar]
- Case D. Computer 1993, 26, 47. [Google Scholar]
- Smith B. B. Biochem. Educ. 1989, 17, 220. [Google Scholar]
- Bui J. M.; Henchman R. H.; McCammon J. A. Biophys. J. 2003, 85, 2267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fan Y.; Lund L.; Shao Q.; Gao Y. Q.; Raushel F. M. J. Am. Chem. Soc. 2009, 131, 10211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Couture J.-F.; Legrand P.; Cantin L.; Labrie F.; Luu-The V.; Breton R. J. Mol. Biol. 2004, 339, 89. [DOI] [PubMed] [Google Scholar]
- Amaro R. E.; Myers R. S.; Davisson V. J.; Luthey-Schulten Z. A. Biophys. J. 2005, 89, 475. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Badet B.; Vermoote P.; Haumont P. Y.; Lederer F.; LeGoffic F. Biochemistry 1987, 26, 1940. [DOI] [PubMed] [Google Scholar]
- Lund L.; Fan Y.; Shao Q.; Gao Y. Q.; Raushel F. M. J. Am. Chem. Soc. 2010, 132, 3870. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miles B. W.; Raushel F. M. Biochemistry 2000, 39, 5051. [DOI] [PubMed] [Google Scholar]
- Li K. K.; Beeson; Ghiviriga I.; Richards N. G. J. Biochemistry 2007, 46, 4840. [DOI] [PubMed] [Google Scholar]
- Myers R. S.; Amaro R. E.; Luthey-Schulten Z. A.; Davisson V. J. Biochemistry 2005, 44, 11974. [DOI] [PubMed] [Google Scholar]
- Meyer M. E.; Gutierrez J. A.; Raushel F. M.; Richards N. G. J. Biochemistry 2010, 49, 9391. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Floquet N.; Mouilleron S.; Daher R.; Maigret B.; Badet B.; Badet-Denisot M. A. FEBS Lett. 2007, 581, 2981. [DOI] [PubMed] [Google Scholar]
- Van den Heuvel R. H. .; Svergun D. I.; Petoukhov M. V.; Coda A.; Curti B.; Ravasio S.; Vanoni M. A.; Mattevi A. J. Chem. Biol. 2003, 330, 113. [DOI] [PubMed] [Google Scholar]
- Brzovic P. S.; Sawa Y.; Hyde C. C.; Miles E. W.; Dunn M. F. J. Biol. Chem. 1992, 267, 13028. [PubMed] [Google Scholar]
- Darnault C.; Volbeda A.; Kim E. J.; Legrand P.; Vernède X.; Lindahl P. A.; Fontecilla-Camps J. C. Nat. Struct. Mol. Biol. 2003, 10, 271. [DOI] [PubMed] [Google Scholar]
- Von Ossowski I.; Ståhlberg J.; Koivula A.; Piens K.; Becker D.; Boer H.; Harle R.; Harris M.; Divne C.; Mahdi S.; Zhao Y.; Driguez H.; Claeyssens M.; Sinnott M. L.; Teeri T. T. J. Mol. Biol. 2003, 333, 817. [DOI] [PubMed] [Google Scholar]
- Mouilleron S.; Badet-Denisot M.-A.; Badet B.; Golinelli-Pimpaneau B. Arch. Biochem. Biophys. 2011, 505, 1. [DOI] [PubMed] [Google Scholar]
- Doig A. J. Biophys. Chem. 1996, 61, 131. [DOI] [PubMed] [Google Scholar]
- Endrizzi J. A.; Kim H.; Anderson P. M.; Baldwin E. P. Biochemistry 2005, 44, 13491. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sazinsky M. H.; Lippard S. J. J. Am. Chem. Soc. 2005, 127, 5814. [DOI] [PubMed] [Google Scholar]
- Zhang L.; Liu W.; Hu T.; Du L.; Luo C.; Chen K.; Shen X.; Jiang H. J. Biol. Chem. 2008, 283, 5370. [DOI] [PubMed] [Google Scholar]
- Oprea T. I.; Hummer G.; García A. E. Proc. Natl. Acad. Sci. U.S.A. 1997, 94, 2133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fishelovitch D.; Shaik S.; Wolfson H. J.; Nussinov R. J. Phys. Chem. B 2010, 114, 5964. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lario P. I.; Sampson N.; Vrielink A. J. Mol. Biol. 2003, 326, 1635. [DOI] [PubMed] [Google Scholar]
- Fishelovitch D.; Shaik S.; Wolfson H. J.; Nussinov R. J. Phys. Chem. B 2009, 113, 13018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zawaira A.; Coulson L.; Gallotta M.; Karimanzira O.; Blackburn J. J. Struct. Biol. 2011, 173, 282. [DOI] [PubMed] [Google Scholar]
- Niu C.; Xu Y.; Xu Y.; Luo X.; Duan W.; Silman I.; Sussman J. L.; Zhu W.; Chen K.; Shen J.; Jiang H. J. Phys. Chem. B 2005, 109, 23730. [DOI] [PubMed] [Google Scholar]
- Lountos G. T.; Mitchell K. H.; Studts J. M.; Fox B. G.; Orville A. M. Biochemistry 2005, 44, 7131. [DOI] [PubMed] [Google Scholar]
- Vanoni M. A.; Curti B. Arch. Biochem. Biophys. 2005, 433, 193. [DOI] [PubMed] [Google Scholar]
- Xu L.; Zhao W.; Wang X. Eur. Biophys. J. 2010, 39, 327. [DOI] [PubMed] [Google Scholar]
- Xu Y.; Colletier J.-P.; Weik M.; Jiang H.; Moult J.; Silman I.; Sussman J. L. Biophys. J. 2008, 95, 2500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wester M. R.; Johnson E. F.; Marques-Soares C.; Dansette P. M.; Mansuy D.; Stout C. D. Biochemistry 2003, 42, 6370. [DOI] [PubMed] [Google Scholar]
- Sevostyanova A.; Belogurov G. A.; Mooney R. A.; Landick R.; Artsimovitch I. Mol. Cell. 2011, 43, 253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tan X.; Loke H.-K.; Fitch S.; Lindahl P. A. J. Am. Chem. Soc. 2005, 127, 5833. [DOI] [PubMed] [Google Scholar]
- Biswal B. K.; Morisseau C.; Garen G.; Cherney M. M.; Garen C.; Niu C.; Hammock B. D.; James M. N. G. J. Mol. Biol. 2008, 381, 897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Da Silva Giotto M. T.; Garratt R. C.; Oliva G.; Mascarenhas Y. P.; Giglio J. R.; Cintra A. C. O.; De Azevedo W. F.; Arni R. K.; Ward R. J. Proteins: Struct., Funct., Bioinf. 1998, 30, 442. [DOI] [PubMed] [Google Scholar]
- Park E.; Rho Y. M.; Koh O.-J.; Ahn S. W.; Seong I. S.; Song J.-J.; Bang O.; Seol J. H.; Wang J.; Eom S. H.; Chung C. H. J. Biol. Chem. 2005, 280, 22892. [DOI] [PubMed] [Google Scholar]
- Rhee S.; Parris K. D.; Ahmed S. A.; Miles E. W.; Davies D. R. Biochemistry 1996, 35, 4211. [DOI] [PubMed] [Google Scholar]
- Oakley A. J.; Klvana M.; Otyepka M.; Nagata Y.; Wilce M. C. J.; Damborsky J. Biochemistry 2004, 43, 870. [DOI] [PubMed] [Google Scholar]
- Pawelek P. D.; Cheah J.; Coulombe R.; Macheroux P.; Ghisla S.; Vrielink A. EMBO J. 2000, 19, 4204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sazinsky M. H.; Bard J.; Di Donato A.; Lippard S. J. J. Biol. Chem. 2004, 279, 30600. [DOI] [PubMed] [Google Scholar]
- Fiorucci S.; Golebiowski J.; Cabrol-Bass D.; Antonczak S. Proteins 2006, 64, 845. [DOI] [PubMed] [Google Scholar]
- Eisenmesser E. Z.; Bosco D. A.; Akke M.; Kern D. Science 2002, 295, 1520. [DOI] [PubMed] [Google Scholar]
- Rozovsky S.; Jogl G.; Tong L.; McDermott A. E. J. Mol. Biol. 2001, 310, 271. [DOI] [PubMed] [Google Scholar]
- Massi F.; Wang C.; Palmer A. G. III Biochemistry 2006, 45, 10787. [DOI] [PubMed] [Google Scholar]
- Katoh E.; Louis J. M.; Yamazaki T.; Gronenborn A. M.; Torchia D. A.; Ishima R. Protein Sci. 2003, 12, 1376. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Osborne M. J.; Schnell J.; Benkovic S. J.; Dyson H. J.; Wright P. E. Biochemistry 2001, 40, 9846. [DOI] [PubMed] [Google Scholar]
- Boroujerdi A. F. B.; Young J. K. Biopolymers 2009, 91, 140. [DOI] [PubMed] [Google Scholar]
- Brezovsky J.; Chovancova E.; Gora A.; Pavelka A.; Biedermannova L.; Damborsky J. Biotechnol. Adv. 2013, 31, 38. [DOI] [PubMed] [Google Scholar]
- Chovancova E.; Pavelka A.; Benes P.; Strnad O.; Brezovsky J.; Kozilkova B.; Gora A.; Sustr V.; Klvana M.; Medek P.; Biedermannova L.; Sochor J.; Damborsky J. PLoS Comput. Biol. 2012, 8, e1002708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Petrek M.; Kosinova P.; Koca J.; Otyepka M. Structure 2007, 15, 1357. [DOI] [PubMed] [Google Scholar]
- Yaffe E.; Fishelovitch D.; Wolfson H. J.; Halperin D.; Nussinov R. Proteins 2008, 73, 72. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lafaquiere V.; Barbe S.; Puech-Guenot S.; Guieysse D.; Cortes J.; Monsan P.; Simeon T.; Andre I.; Remaud-Simeon M. ChemBioChem 2009, 10, 2760. [DOI] [PubMed] [Google Scholar]
- Tina K. G.; Bhadra R.; Srinivasan N. Nucleic Acids Res. 2007, 35, W473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arroyo-Mañez P.; Bikiel D. E.; Boechi L.; Capece L.; Di Lella S.; Estrin D. A.; Martí M. A.; Moreno D. M.; Nadra A. D.; Petruk A. A. Biochim. Biophys. Acta: Proteins Proteomics 2011, 1814, 1054. [DOI] [PubMed] [Google Scholar]
- Chang C.-E.; Shen T.; Trylska J.; Tozzini V.; McCammon J. A. Biophys. J. 2006, 90, 3880. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schleinkofer K.; Sudarko; Winn P. J.; Ludemann S. K.; Wade R. C. EMBO Rep. 2005, 6, 584. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murray L. J.; García-Serres R.; McCormick M. S.; Davydov R.; Naik S. G.; Kim S.-H.; Hoffman B. M.; Huynh B. H.; Lippard S. J. Biochemistry 2007, 46, 14795. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sciara G.; Kendrew S. G.; Miele A. E.; Marsh N. G.; Federici L.; Malatesta F.; Schimperna G.; Savino C.; Vallone B. EMBO J. 2003, 22, 205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rowlett R. S. Biochim. Biophys. Acta 2010, 1804, 362. [DOI] [PubMed] [Google Scholar]
- Kohls D.; Sulea T.; Purisima E. O.; MacKenzie R. E.; Vrielink A. Structure 2000, 8, 35. [DOI] [PubMed] [Google Scholar]
- Abe I.; Morita H. Nat. Prod. Rep. 2010, 27, 809. [DOI] [PubMed] [Google Scholar]
- Manjasetty B. A.; Powlowski J.; Vrielink A. Proc. Natl. Acad. Sci. U.S.A. 2003, 100, 6992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Toth K.; Sedlak E.; Sprinzl M.; Zoldak G. Biochim. Biophys. Acta 2008, 1784, 789. [DOI] [PubMed] [Google Scholar]
- Hiromoto T.; Fujiwara S.; Hosokawa K.; Yamaguchi H. J. Chem. Biol. 2006, 364, 878. [DOI] [PubMed] [Google Scholar]
- Vrielink A.; Ghisla S. FEBS J. 2009, 276, 6826. [DOI] [PubMed] [Google Scholar]
- Xu Q.; Eguchi T.; Mathews I. I.; Rife C. L.; Chiu H.-J.; Farr C. L.; Feuerhelm J.; Jaroszewski L.; Klock H. E.; Knuth M. W.; Miller M. D.; Weekes D.; Elsliger M.-A.; Deacon A. M.; Godzik A.; Lesley S. A.; Wilson I. A. J. Mol. Biol. 2010, 404, 403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vardar G.; Wood T. K. J. Bacteriol. 2005, 187, 1511. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ruvinov S. B.; Yang X. J.; Parris K. D.; Banik U.; Ahmed S. A.; Miles E. W.; Sackett D. L. J. Biol. Chem. 1995, 270, 6357. [DOI] [PubMed] [Google Scholar]
- Thoden J. B.; Huang X.; Raushel F. M.; Holden H. M. J. Biol. Chem. 2002, 277, 39722. [DOI] [PubMed] [Google Scholar]
- Huang X.; Raushel F. M. Biochemistry 2000, 39, 3240. [DOI] [PubMed] [Google Scholar]
- Brouk M.; Derry N. L.; Shainsky J.; Zelas Z. B. B.; Boyko Y.; Dabush K.; Fishman A. J. Mol. Catal. B: Enzym. 2010, 66, 72. [Google Scholar]
- Morita H.; Kondo S.; Oguro S.; Noguchi H.; Sugio S.; Abe I.; Kohno T. Chem. Biol. 2007, 14, 359. [DOI] [PubMed] [Google Scholar]
- Schmitt J.; Brocca S.; Schmid R. D.; Pleiss J. Protein Eng. 2002, 15, 595. [DOI] [PubMed] [Google Scholar]
- Chaloupkova R.; Sykorova J.; Prokop Z.; Jesenska A.; Monincova M.; Pavlova M.; Tsuda M.; Nagata Y.; Damborsky J. J. Biol. Chem. 2003, 278, 52622. [DOI] [PubMed] [Google Scholar]
- Pavlova M.; Klvana M.; Prokop Z.; Chaloupkova R.; Banas P.; Otyepka M.; Wade R. C.; Tsuda M.; Nagata Y.; Damborsky J. Nat. Chem. Biol. 2009, 5, 727. [DOI] [PubMed] [Google Scholar]
- Klvana M.; Pavlova M.; Koudelakova T.; Chaloupkova R.; Dvorak P.; Prokop Z.; Stsiapanava A.; Kuty M.; Kuta-Smatanova I.; Dohnalek J.; Kulhanek P.; Wade R. C.; Damborsky J. J. Mol. Biol. 2009, 392, 1339. [DOI] [PubMed] [Google Scholar]
- Li Q.-A.; Mavrodi D. V.; Thomashow L. S.; Roessle M.; Blankenfeldt W. J. Biol. Chem. 2011, 286, 18213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carere J.; Baker P.; Seah S. Y. K. Biochemistry 2011, 50, 8407. [DOI] [PubMed] [Google Scholar]
- Wang J.; Ortiz-Maldonado M.; Entsch B.; Massey V.; Ballou D.; Gatti D. L. Proc. Natl. Acad. Sci. U.S.A. 2002, 99, 608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Faerman C.; Ripoll D.; Bon S.; Le Feuvre Y.; Morel N.; Massoulie J.; Sussman J. L.; Silman I. FEBS Lett. 1996, 386, 65. [DOI] [PubMed] [Google Scholar]
- Alisaraie L.; Fels G. J. Mol. Model. 2006, 12, 348. [DOI] [PubMed] [Google Scholar]
- Wlodek S. T.; Clark T. W.; Scott L. R.; McCammon J. A. J. Am. Chem. Soc. 1997, 119, 9513. [Google Scholar]
- Xu Y.; Shen J.; Luo X.; Silman I.; Sussman J. L.; Chen K.; Jiang H. J. Am. Chem. Soc. 2003, 125, 11340. [DOI] [PubMed] [Google Scholar]
- Rydberg E. H.; Brumshtein B.; Greenblatt H. M.; Wong D. M.; Shaya D.; Williams L. D.; Carlier P. R.; Pang Y.-P.; Silman I.; Sussman J. L. J. Med. Chem. 2006, 49, 5491. [DOI] [PubMed] [Google Scholar]
- Tai K.; Shen T.; Börjesson U.; Philippopoulos M.; McCammon J. A. Biophys. J. 2001, 81, 715. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanson B.; Colletier J.-P.; Xu Y.; Lang P. T.; Jiang H.; Silman I.; Sussman J. L.; Weik M. Protein Sci. 2011, 20, 1114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Greenblatt H. M.; Guillou C.; Guénard D.; Argaman A.; Botti S.; Badet B.; Thal C.; Silman I.; Sussman J. L. J. Am. Chem. Soc. 2004, 126, 15405. [DOI] [PubMed] [Google Scholar]
- Harel M.; Schalk I.; Ehret-Sabatier L.; Bouet F.; Goeldner M.; Hirth C.; Axelsen P. H.; Silman I.; Sussman J. L. Proc. Natl. Acad. Sci. U.S.A. 1993, 90, 9031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dvir H.; Jiang H. L.; Wong D. M.; Harel M.; Chetrit M.; He X. C.; Jin G. Y.; Yu G. L.; Tang X. C.; Silman I.; Bai D. L.; Sussman J. L. Biochemistry 2002, 41, 10810. [DOI] [PubMed] [Google Scholar]
- Harmat V.; Domokos K.; Menyhard D. K.; Palló A.; Szeltner Z.; Szamosi I.; Beke-Somfai T.; Náray-Szabó G.; Polgár L. J. Biol. Chem. 2011, 286, 1987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Papaleo E.; Renzetti G.; Tiberti M. PLoS One 2012, 7, e35686. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Santos C. R.; Tonoli C. C. C.; Trindade D. M.; Betzel C.; Takata H.; Kuriki T.; Kanai T.; Imanaka T.; Arni R. K.; Murakami M. T. Proteins: Struct., Funct., Bioinf. 2011, 79, 547. [DOI] [PubMed] [Google Scholar]
- Tesson A. R.; Soper T. S.; Ciustea M.; Richards N. G. Arch. Biochem. Biophys. 2003, 413, 23. [DOI] [PubMed] [Google Scholar]
- Larsen T. M.; Boehlein S. K.; Schuster S. M.; Richards N. G. J.; Thoden J. B.; Holden H. M.; Rayment I. Biochemistry 1999, 38, 16146. [DOI] [PubMed] [Google Scholar]
- Wang J.; Song J. J.; Franklin M. C.; Kamtekar S.; Im Y. J.; Rho S. H.; Seong I. S.; Lee C. S.; Chung C. H.; Eom S. H. Structure 2001, 9, 177. [DOI] [PubMed] [Google Scholar]
- Wang J.; Song J. J.; Seong I. S.; Franklin M. C.; Kamtekar S.; Eom S. H.; Chung C. H. Structure 2001, 9, 1107. [DOI] [PubMed] [Google Scholar]
- Kim J.; Raushel F. M. Biochemistry 2004, 43, 5334. [DOI] [PubMed] [Google Scholar]
- Kim J.; Raushel F. M. Arch. Biochem. Biophys. 2004, 425, 33. [DOI] [PubMed] [Google Scholar]
- Huang X.; Raushel F. M. Biochemistry 2000, 39, 3240. [DOI] [PubMed] [Google Scholar]
- Holden H. M.; Thoden J. B.; Raushel F. M. Curr. Opin. Struct. Biol. 1998, 8, 679. [DOI] [PubMed] [Google Scholar]
- Tan X.; Volbeda A.; Fontecilla-Camps J. C.; Lindahl P. A. J. Biol. Inorg. Chem. 2006, 11, 371. [DOI] [PubMed] [Google Scholar]
- Tan X.; Lindahl P. A. J. Biol. Inorg. Chem. 2008, 13, 771. [DOI] [PubMed] [Google Scholar]
- Volbeda A.; Fontecilla-Camps J. C. J. Biol. Inorg. Chem. 2004, 9, 525. [DOI] [PubMed] [Google Scholar]
- Teng Y.-B.; Jiang Y.-L.; He Y.-X.; He W.-W.; Lian F.-M.; Chen Y.; Zhou C.-Z. BMC Struct. Biol. 2009, 9, 67. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wierdl M.; Morton C. L.; Nguyen N. K.; Redinbo M. R.; Potter P. M. Biochemistry 2004, 43, 1874. [DOI] [PubMed] [Google Scholar]
- Yu X.; Sigler S. C.; Hossain D.; Wierdl M.; Gwaltney S. R.; Potter P. M.; Wadkins R. M. J. Mol. Model. 2011, 18, 2869. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Varrot A.; Hastrup S.; Schülein M.; Davies G. J. Biochem. J. 1999, 337, 297. [PMC free article] [PubMed] [Google Scholar]
- Schülein M. Biochim. Biophys. Acta 2000, 1543, 239. [DOI] [PubMed] [Google Scholar]
- Koivula A.; Kinnari T.; Harjunpää V.; Ruohonen L.; Teleman A.; Drakenberg T.; Rouvinen J.; Jones T. A.; Teeri T. T. FEBS Lett. 1998, 429, 341. [DOI] [PubMed] [Google Scholar]
- De Groeve M. R. M.; Remmery L.; Van Hoorebeke A.; Stout J.; Desmet T.; Savvides S. N.; Soetaert W. Biotechnol. Bioeng. 2010, 107, 413. [DOI] [PubMed] [Google Scholar]
- Fushinobu S.; Mertz B.; Hill A. D.; Hidaka M.; Kitaoka M.; Reilly P. J. Carbohydr. Res. 2008, 343, 1023. [DOI] [PubMed] [Google Scholar]
- Morita H.; Wanibuchi K.; Nii H.; Kato R.; Sugio S.; Abe I. Proc. Natl. Acad. Sci. U.S.A. 2010, 107, 19778. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Podzelinska K.; Latimer R.; Bhattacharya A.; Vining L. C.; Zechel D. L.; Jia Z. J. Mol. Biol. 2010, 397, 316. [DOI] [PubMed] [Google Scholar]
- Chen L.; Lyubimov A. Y.; Brammer L.; Vrielink A.; Sampson N. S. Biochemistry 2008, 47, 5368. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coulombe R.; Yue K. Q.; Ghisla S.; Vrielink A. J. Biol. Chem. 2001, 276, 30435. [DOI] [PubMed] [Google Scholar]
- Piubelli L.; Pedotti M.; Molla G.; Feindler-Boeckh S.; Ghisla S.; Pilone M. S.; Pollegioni L. J. Biol. Chem. 2008, 283, 24738. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang W.; Boju L.; Tkalec L.; Su H.; Yang H. O.; Gunay N. S.; Linhardt R. J.; Kim Y. S.; Matte A.; Cygler M. Biochemistry 2001, 40, 2359. [DOI] [PubMed] [Google Scholar]
- Johnson B. J.; Cohen J.; Welford R. W.; Pearson A. R.; Schulten K.; Klinman J. P.; Wilmot C. M. J. Biol. Chem. 2007, 282, 17767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilmot C. M.; Saysell C. G.; Blessington A.; Conn D. A.; Kurtis C. R.; McPherson M. J.; Knowles P. F.; Phillips S. E. V. FEBS Lett. 2004, 576, 301. [DOI] [PubMed] [Google Scholar]
- Wilmot C. M.; Murray J. M.; Alton G.; Parsons M. R.; Convery M. A.; Blakeley V.; Corner A. S.; Palcic M. M.; Knowles P. F.; McPherson M. J.; Phillips S. E. Biochemistry 1997, 36, 1608. [DOI] [PubMed] [Google Scholar]
- Mouilleron S.; Golinelli-Pimpaneau B. Curr. Opin. Struct. Biol. 2007, 17, 653. [DOI] [PubMed] [Google Scholar]
- Endrizzi J. A.; Kim H.; Anderson P. M.; Baldwin E. P. Biochemistry 2004, 43, 6447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goto M.; Omi R.; Nakagawa N.; Miyahara I.; Hirotsu K. Structure 2004, 12, 1413. [DOI] [PubMed] [Google Scholar]
- Khan K. K.; He Y. Q.; Domanski T. L.; Halpert J. R. Mol. Pharmacol. 2002, 61, 495. [DOI] [PubMed] [Google Scholar]
- Wade R. C.; Winn P. J.; Schlichting I.; Sudarko J. Inorg. Biochem. 2004, 98, 1175. [DOI] [PubMed] [Google Scholar]
- Bhabha G.; Lee J.; Ekiert D. C.; Gam J.; Wilson I. A.; Dyson H. J.; Benkovic S. J.; Wright P. E. Science 2011, 332, 234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luo J.; Bruice T. C. J. Biomol. Struct. Dyn. 2009, 27, 245. [DOI] [PubMed] [Google Scholar]
- Swanwick R. S.; Shrimpton P. J.; Allemann R. K. Biochemistry 2004, 43, 4119. [DOI] [PubMed] [Google Scholar]
- Zou J.; Hallberg B. M.; Bergfors T.; Oesch F.; Arand M.; Mowbray S. L.; Jones T. A. Structure 2000, 8, 111. [DOI] [PubMed] [Google Scholar]
- Kimber M. S.; Martin F.; Lu Y.; Houston S.; Vedadi M.; Dharamsi A.; Fiebig K. M.; Schmid M.; Rock C. O. J. Biol. Chem. 2004, 279, 52593. [DOI] [PubMed] [Google Scholar]
- Swarnamukhi P. L.; Sharma S. K.; Bajaj P.; Surolia N.; Surolia A.; Suguna K. FEBS Lett. 2006, 580, 2653. [DOI] [PubMed] [Google Scholar]
- Mouilleron S.; Badet-Denisot M.-A.; Golinelli-Pimpaneau B. J. Biol. Chem. 2006, 281, 4404. [DOI] [PubMed] [Google Scholar]
- Teplyakov A.; Obmolova G.; Badet B.; Badet-Denisot M. A. J. Mol. Biol. 2001, 313, 1093. [DOI] [PubMed] [Google Scholar]
- Dossena L.; Curti B.; Vanoni M. A. Biochemistry 2007, 46, 4473. [DOI] [PubMed] [Google Scholar]
- Kim J. H.; Krahn J. M.; Tomchick D. R.; Smith J. L.; Zalkin H. J. Biol. Chem. 1996, 271, 15549. [DOI] [PubMed] [Google Scholar]
- Krahn J. M.; Kim J. H.; Burns M. R.; Parry R. J.; Zalkin H.; Smith J. L. Biochemistry 1997, 36, 11061. [DOI] [PubMed] [Google Scholar]
- Biedermannova L.; Prokop Z.; Gora A.; Chovancova E.; Kovacs M.; Damborsky J.; Wade R. C. J. Biol. Chem. 2012, 287, 29062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kalyaanamoorthy S.; Chen Y.-P. P. J. Chem. Inf. Model. 2012, 52, 589. [DOI] [PubMed] [Google Scholar]
- Haider S.; Joseph C. G.; Neidle S.; Fierke C. A.; Fuchter M. J. Bioorg. Med. Chem. Lett. 2011, 21, 2129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hornak V.; Okur A.; Rizzo R. C.; Simmerling C. Proc. Natl. Acad. Sci. U.S.A. 2006, 103, 915. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hornak V.; Okur A.; Rizzo R. C.; Simmerling C. J. Am. Chem. Soc. 2006, 128, 2812. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu F.; Kovalevsky A. Y.; Louis J. M.; Boross P. I.; Wang Y.-F.; Harrison R. W.; Weber I. T. J. Chem. Biol. 2006, 358, 1191. [DOI] [PubMed] [Google Scholar]
- Torbeev V. Y.; Raghuraman H.; Mandal K.; Senapati S.; Perozo E.; Kent S. B. H. J. Am. Chem. Soc. 2008, 131, 884. [DOI] [PubMed] [Google Scholar]
- Nicolet Y.; Cavazza C.; Fontecilla-Camps J. C. J. Inorg. Biochem. 2002, 91, 1. [DOI] [PubMed] [Google Scholar]
- Cohen J.; Kim K.; King P.; Seibert M.; Schulten K. Structure 2005, 13, 1321. [DOI] [PubMed] [Google Scholar]
- Cohen J.; Kim K.; Posewitz M.; Ghirardi M. L.; Schulten K.; Seibert M.; King P. Biochem. Soc. Trans. 2005, 33, 80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Omi R.; Mizuguchi H.; Goto M.; Miyahara I.; Hayashi H.; Kagamiyama H.; Hirotsu K. J. Biochem. 2002, 132, 759. [DOI] [PubMed] [Google Scholar]
- Chaudhuri B. N.; Lange S. C.; Myers R. S.; Chittur S. V.; Davisson V. J.; Smith J. L. Structure 2001, 9, 987. [PubMed] [Google Scholar]
- Amaro R.; Luthey-Schulten Z. Chem. Phys. 2004, 307, 147. [Google Scholar]
- Myers R. S.; Jensen J. R.; Deras I. L.; Smith J. L.; Davisson V. J. Biochemistry 2003, 42, 7013. [DOI] [PubMed] [Google Scholar]
- Douangamath A.; Walker M.; Beismann-Driemeyer S.; Vega-Fernandez M. C.; Sterner R.; Wilmanns M. Structure 2002, 10, 185. [DOI] [PubMed] [Google Scholar]
- Chaudhuri B. N.; Lange S. C.; Myers R. S.; Davisson V. J.; Smith J. L. Biochemistry 2003, 42, 7003. [DOI] [PubMed] [Google Scholar]
- Josephine H. R.; Ravichandran K. R.; Hedstrom L. Biochemistry 2010, 49, 10674. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johansson P.; Wiltschi B.; Kumari P.; Kessler B.; Vonrhein C.; Vonck J.; Oesterhelt D.; Grininger M. Proc. Natl. Acad. Sci. U.S.A. 2008, 105, 12803. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moustafa I. M.; Foster S.; Lyubimov A. Y.; Vrielink A. J. Mol. Biol. 2006, 364, 991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qian Z.; Horton J. R.; Cheng X.; Lutz S. J. Mol. Biol. 2009, 393, 191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Uppenberg J.; Ohrner N.; Norin M.; Hult K.; Kleywegt G. J.; Patkar S.; Waagen V.; Anthonsen T.; Jones T. A. Biochemistry 1995, 34, 16838. [DOI] [PubMed] [Google Scholar]
- Magnusson A. O.; Rotticci-Mulder J. C.; Santagostino A.; Hult K. ChemBioChem 2005, 6, 1051. [DOI] [PubMed] [Google Scholar]
- Cha S.-S.; An Y. J.; Lee C. R.; Lee H. S.; Kim Y.-G.; Kim S. J.; Kwon K. K.; De Donatis G. M.; Lee J.-H.; Maurizi M. R.; Kang S. G. EMBO J. 2010, 29, 3520. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klimacek M.; Brunsteiner M.; Nidetzky B. J. Biol. Chem. 2012, 287, 6655. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murray L. J.; Lippard S. J. Acc. Chem. Res. 2007, 40, 466. [DOI] [PubMed] [Google Scholar]
- Son S.-Y.; Ma J.; Kondou Y.; Yoshimura M.; Yamashita E.; Tsukihara T. Proc. Natl. Acad. Sci. U.S.A. 2008, 105, 5739. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang J.; Harris J.; Mousseau D. D.; Edmondson D. E. FEBS J. 2009, 276, 4569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Milczek E. M.; Binda C.; Rovida S.; Mattevi A.; Edmondson D. E. FEBS J. 2011, 278, 4860. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hritz J.; Zoldak G.; Sedlak E. Proteins 2006, 64, 465. [DOI] [PubMed] [Google Scholar]
- Feldman-Salit A.; Wirtz M.; Lenherr E. D.; Throm C.; Hothorn M.; Scheffzek K.; Hell R.; Wade R. C. Structure 2012, 20, 292. [DOI] [PubMed] [Google Scholar]
- Oliaro-Bosso S.; Caron G.; Taramino S.; Ermondi G.; Viola F.; Balliano G. PLoS One 2011, 6, e22134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arni R. K.; Ward R. J.; Gutierrez J. M.; Tulinsky A. Acta Crystallogr., Sect. D: Biol. Crystallogr. 1995, 51, 311. [DOI] [PubMed] [Google Scholar]
- Ullah A.; Souza T. A. C. B.; Betzel C.; Murakami M. T.; Arni R. K. Int. J. Biol. Macromol. 2012, 51, 209. [DOI] [PubMed] [Google Scholar]
- Goldstein R.; Cheng J.; Stec B.; Roberts M. F. Biochemistry 2012, 51, 2579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scoble J.; McAlister A. D.; Fulton Z.; Troy S.; Byres E.; Vivian J. P.; Brammananth R.; Wilce M. C. J.; Le Nours J.; Zaker-Tabrizi L.; Coppel R. L.; Crellin P. K.; Rossjohn J.; Beddoe T. J. Mol. Biol. 2010, 398, 26. [DOI] [PubMed] [Google Scholar]
- Jeudy S.; Monchois V.; Maza C.; Claverie J.-M.; Abergel C. Proteins 2006, 62, 302. [DOI] [PubMed] [Google Scholar]
- Leitgeb S.; Petschacher B.; Wilson D. K.; Nidetzky B. FEBS Lett. 2005, 579, 763. [DOI] [PubMed] [Google Scholar]
- Kozma E.; Brown E.; Ellis E. M.; Lapthorn A. J. J. Biol. Chem. 2002, 277, 16285. [DOI] [PubMed] [Google Scholar]
- Xia L.; Ruppert M.; Wang M.; Panjikar S.; Lin H.; Rajendran C.; Barleben L.; Stöckigt J. ACS Chem. Biol. 2012, 7, 226. [DOI] [PubMed] [Google Scholar]
- Labonté P.; Axelrod V.; Agarwal A.; Aulabaugh A.; Amin A.; Mak P. J. Biol. Chem. 2002, 277, 38838. [DOI] [PubMed] [Google Scholar]
- Tagami S.; Sekine S.; Kumarevel T.; Hino N.; Murayama Y.; Kamegamori S.; Yamamoto M.; Sakamoto K.; Yokoyama S. Nature 2010, 468, 978. [DOI] [PubMed] [Google Scholar]
- McCormick M. S.; Sazinsky M. H.; Condon K. L.; Lippard S. J. J. Am. Chem. Soc. 2006, 128, 15108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Notomista E.; Cafaro V.; Bozza G.; Di Donato A. Appl. Environ. Microbiol. 2009, 75, 823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang Y.; Berlow R. B.; Loria J. P. Biochemistry 2009, 48, 4548. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Malabanan M. M.; Amyes T. L.; Richard J. P. Curr. Opin. Struct. Biol. 2010, 20, 702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wierenga R. K.; Kapetaniou E. G.; Venkatesan R. Cell. Mol. Life Sci. 2010, 67, 3961. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schmitt E.; Panvert M.; Blanquet S.; Mechulam Y. Structure 2005, 13, 1421. [DOI] [PubMed] [Google Scholar]
- Wu J.; Bu W.; Sheppard K.; Kitabatake M.; Kwon S.-T.; Söll D.; Smith J. L. J. Mol. Biol. 2009, 391, 703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oshikane H.; Sheppard K.; Fukai S.; Nakamura Y.; Ishitani R.; Numata T.; Sherrer R. L.; Feng L.; Schmitt E.; Panvert M.; Blanquet S.; Mechulam Y.; Söll D.; Nureki O. Science 2006, 312, 1950. [DOI] [PubMed] [Google Scholar]
- Pompliano D. L.; Peyman A.; Knowles J. R. Biochemistry 1990, 29, 3186. [DOI] [PubMed] [Google Scholar]
- Dierkers A. T.; Niks D.; Schlichting I.; Dunn M. F. Biochemistry 2009, 48, 10997. [DOI] [PMC free article] [PubMed] [Google Scholar]