

Abstract
Biodiesel, as one type of renewable energy, is an ideal substitute for petroleum-based diesel fuel and is usually made from triacylglycerides by transesterification with alcohols. Biodiesel production based on microbial fermentation aiming to establish more efficient, less-cost and sustainable biodiesel production strategies is under current investigation by various start-up biotechnology companies and research centers. Genetic engineering plays a key role in the transformation of microbes into the desired cell factories with high efficiency of biodiesel production. Here, we present an overview of principal microorganisms used in the microbial biodiesel production and recent advances in metabolic engineering for the modification required. Overexpression or deletion of the related enzymes for de novo synthesis of biodiesel is highlighted with relevant examples.
Keywords: genetic engineering, microorganisms, biodiesel, fatty acids, de novo synthesis
Introduction
Energy is essential for life while the industry development and the global economy actually run on energy. The application of fossil fuels as energy sources is unsustainable due to depleting limited resources and also due to the accumulation of greenhouse gases in the environment.1 The current consumption of crude oil is about 11.6 million tons per day, from which we can concluded that the entire resources will suffice for a rather short time period only.2,3 Without appropriate alternatives to crude oil, the global economy will suffer a dramatic collapse by reason of exploding oil prices as the demand will continuously rise.4 It is inevitable to resolve the dependence to crude oil and the increasing impairment of the environment by establishing a sustainable and competitive alternative which is based on renewable and abundant feedstock like biomass5,6 or on other regenerative sources. Environmental concerns and the depletion of oil reserves have resulted in research promotion on environmentally friendly and sustainable biofuels.7
Among all the biofuels, biodiesel has been receiving perhaps the most attention, due to the similarity with conventional diesel in terms of chemical structure and energy content. Additionally, no modification of the diesel engine is required, as biodiesel is compatible with existing engine models and has been commercially blended with diesel as a transportation fuel in a number of countries including Germany, Italy and Malaysia.8 Biodiesel is mainly obtained by the transesterification of fat and vegetable oils in the presence of a catalyst by an alcohol leading to a fatty acid methyl esters (FAMEs) or a fatty acid ethyl ester (FAEEs).9 Unlike petroleum diesel, biodiesel is biodegradable and non-toxic, and it significantly reduces toxicant and other emissions when burned as a fuel.10
The traditional methods of biodiesel production include chemical catalysis and enzymatic catalysis. Chemical catalysis is of prime importance for the current industrial production of biodiesel,11 while enzymatic catalysis is also an attractive approach for its mild reaction conditions, easy product separation and so on.12,13 However, the current biodiesel production faces several problems: deficiency of a stable, sufficient feedstock supply system, inconsistent performance, and challenging economics.14 The microbial production has the potential to overcome these challenges due to some advantages such as short producing period, little labor required, easy to scale up, and regardless of venue, season, climate change and other factors.15 Making full use of microorganism for biodiesel production has a very large potential. Here, we present a simple overview of principle microorganisms used in the biodiesel production, important metabolism pathways related with biodiesel synthesis and potential routes for construction efficient and less-cost biodiesel production systems.
Principal Microorganisms for Biodiesel Production
Microorganisms, usually one-celled organisms, could not only provide the biodiesel synthesis with the substrates such as fatty acids sources and alcohols, but also be used as a catalyst in this process.
Grease microorganisms to supply fatty acid sources
Grease microorganisms, also called oleaginous microorganisms, is frequently applied and studied in the current biodiesel production to supply fatty acids source for transesterification. Grease microorganisms, coming from a rich microorganism resource, can utilize or convert various agro-industrial materials (e.g., plant biomass) to cellular lipids.16 The fatty acid composition of the lipids extracted from grease microorganisms is generally similar to vegetable oils, which can be converted into biodiesel.17 Grease strains could be found in the species of bacteria, yeasts, molds and algae.17 The key characteristics could be summarized as follows: (1) more than 50% of lipids should be accumulated in the cells; (2) the strains can be used in industrialization culture with simple apparatus; (3) the strains should have fast growth speed and anti-pollution ability; and (4) the oil is easy to be extracted.
In recent years, microalgae are attracting considerable interest as an economically viable oil feedstock due to its obvious superiority of easily culture, big group, diverse metabolic capabilities and high fatty acid content.18 Many autotrophic microalgaes capable of oil accumulation have been found, such as Chlorella vulgaris, Botryococcus braunii, Navicula pelliculosa, Scenedsmus acutus, Crypthecodinium cohnii, Dunaliella primolecta, Monallanthus salina, Neochloris oleoabundans, Phaeodactylum tricornutum and Tetraselmis sueica.19 The average oil content of microalgae varies from 1 to 70% and would significantly affected by the cultivation conditions and algae species.20 Khozin-Goldberg and Cohen21 observed that TAG content increased in Monodus subterraneus under phosphate limitation conditions. Xin et al.22 showed that microalga Scenedesmus sp LX1 yielded the highest amount of lipid with a content of 35.7% when only the cultivation temperature was 20ºC. Microalgae have been considered to be the inevitable development trend of future biodiesel. However, several barriers, such as low growth rate, strict breeding condition and large upfront investment, need to be overcome before microalgae can be used as an economically viable biofuel feedstock.18
Ethanol producers to supply alcohol sources
Alcohol is another substrate in the transesterification reaction to generate the corresponding fatty acid ester. Methanol or ethanol is the most frequently used acyl acceptor, while FAMEs is cheaper, more reactive and volatile than FAEEs. However, ethanol is less toxic and is considered more renewable. Ethanol can be easily produced from renewable sources by microbial fermentation,10 which has been demonstrated by many reports and reviews.23,24 Owning to so much effort from many researchers, the yield of ethanol can even reach the theoretical value.25,26 The most commonly used microbe for ethanol production is yeast, among which, Saccharomyces cerevisiae is the preferred one producing ethanol to give concentration as high as 18% of the fermentation broth.23 S. cerevisiae has been widely used as a key cell factory platform for future biorefineries.27 Zymomonas mobilis is another well studied ethanol-producing strain with published genome sequence.28 Z. mobilis has higher ethanol yields and faster specific rates compared with yeast, although it has a limited carbon substrate range of simple C6 sugars glucose, fructose and sucrose. The genetic manipulation systems in Z. mobilis have been well established, while many researches have been performed on its metabolic engineering focused on extending its substrate range for ethanol production.29 In the study of Hideshi Yanase et al.25 Z. mobilis was engineered to convert the wood hydrolysate containing glucose, mannose and xylose as major sugar components to ethanol by introducing genes encoding mannose and xylose catabolic enzymes. The yield of ethanol reached 89.8% of the theoretical yield within 72 h. Ethanol production could also be achieved by genetic engineering of E. coli strains, which was one of the early successful applications of metabolic engineering.30
Recombinant microorganisms capable of de novo biosynthesis of biodiesel
Pathway manipulation by genetic engineering allows the transformation of microbes into “cell factories”.16 De novo biosynthesis of biodiesel has been realized in some reports by metabolic engineering.31-34 Figure 1 illustrates a common de novo biosynthetic pathway of biodiesel like FAEEs in the microbes. Several specific pathways such as ethanol production, fatty acid metabolism and transesterification are involved in this pathway.

Figure 1. Representative pathways in microorganisms could be engineered for de novo biosynthetic process of biodiesel (e.g., fatty acid ethyl esters).
E. coli is an excellent host for genetic engineering to produce high-value chemicals given its plethora of sophisticated genetic tools as well as the wealth of information available on its metabolism.35,36 The world’s first report of de novo biosynthetic pathway of biodiesel is showed in E. coli by the simultaneous overexpression of the ethanol production genes from Z. mobilis and the wax ester synthase/acyl-CoA-diacylglycerol acyltransferase (WS/DGAT) gene from the Acinetobacter baylyi strain ADP1.37 By this approach, ethanol production in the cells was combined with subsequent esterification when they were cultivated under aerobic conditions in the presence of glucose and oleic acid. It is notable that biosynthesis of FAEEs in Kalscheuer’s work37 relied on the supplementation of exogenous fatty acids. The uptake of exogenous fatty acids from the medium and their activation to the corresponding acyl-CoA thioesters becomes a factor limiting FAEEs accumulation in the report of Kalscheuer. Yangkai Duan and his group members32 further modified the fatty acids pathway of E. coli to increase the content of endogenous fatty acids, which supplied as acyl moieties of coenzyme A thioesters of fatty acids for transesterification. To decrease the process cost, Steen et al.33 engineered the substrate metabolism pathway of the E. coli with FAEEs producing capability. This newly engineered strain was able to accumulate FAEEs utilizing hemicellulose as raw materials. Figure 2 illustrated the detailed genetic manipulations sucessfully used in E. coli to produce FAEEs directly from lignocellulose biomass without the addition of expensive enzymes.

Figure 2. The successfully engineered pathways in E. coli for de novo biosynthesis of biodiesel. An endoxylanase catalytic domain (Xyn10B) from Clostridium stercorarium and a xylanase (Xsa) from Bacteroides ovatus were introduced to hydrolyse the hemicellulose to xylose. Ethanol-forming pathway was introduced by overexpression of pdc gene and adhB gene, coding pyruvate decarboxylase (PDC) and alcohol dehydrogenase (ADH), respectively. Flux through the E. coli fatty acid pathway was increased to improve production of free fatty acids and acyl-CoAs by knockout of the fadE gene coding acyl-CoA dehydrogenase (FadE), by overexpression of acetyl-CoA carboxylase (ACCase), thioesterases (TE) and fatty acyl-CoA synthase (FadD). The atfA gene from A. baylyi coding the wax ester synthase/acyl-coenzyme A: diacylglycerol acyltransferase (WS/DGAT) were introduced for fatty acid ethyl esters formation.
In addition to E. coli, S. cerevisiae may also be a potential candidate for genetic manipulation. S. cerevisiae is a good ethanol producer and is able to accumulate fatty acids with a chain length of mainly 16 or 18 carbon atoms.38 In S. cerevisiae, ethanol is accumulated at a high concentration and hence biodiesel production is not limited by ethanol supply.39,40 The FAEEs synthesis in the S. cerevisiae would initiated only when acyltransferase is introduced, which synthesize fatty acid esters from alcohols and fatty acyl coenzyme A thioesters. On the other hand, comprehensive modifications in more and more functional microorganisms to produce less-cost biofuel will also be possible with advances in genetic transformation methods and increased knowledge regarding expression systems. For example, genes for microbial biodiesel production could be expressed in some of the oleaginous microorganisms independent from fatty acid or lipid feeding, since the fatty acyl-CoA supply in such microorganisms is sufficient.34 The more and more fundamental understanding on the metabolism pathways and genetic manipulation technologies provided in oleaginous microorganisms, such as Rhodococcus opacus41 and Yarrowia lipolytica,42 suggests their potential application in microbial biodiesel production. Another promising functional microorganisms as the candidate for engineering is cellulosic microorganisms, which have multiple cellulase enzyme systems for complete biomass digest. Establishing the plasmid for FAEEs production in cellulosic microorganisms used as an expression host, such as cellulose-degrading or hemicellulose degrading strains, may help to decrease the cost from both raw materials and production process. In addition, microalgal is also recognized as a good model for biodiesel production though manipulation, although the application of genetic engineering in eukaryotic microalgae is still in its infancy.43
Genetic Engineering in Fatty Acids Pathway
Understanding microbial fatty acid metabolism is of great interest for biodiesel production. Both the quantity and the quality of biodiesel precursors from a specific strain are closely linked to how fatty acid metabolism is controlled.
Overview of fatty acids pathway
Cellular fatty acids are derived from three different sources: external supply, endogenous lipid (protein) turnover, and de novo synthesis and elongation.38 The major supply of fatty acids during cellular growth depends on de novo synthesis. Figure 3 illustrates the biochemical reactions involved in fatty acid metabolism. In essence, the fatty acids biosynthesis of microbes is similar with that of animals and plants, both of which are performed by the fatty acid synthetase (FAS) and requires the constant supply of acetyl-CoA and malonyl-CoA.44 In non-oleaginous microbes, acetyl-CoA is usually produced from the glycolysis pathway by breaking down pyruvate and from the cytosolic pyruvate-acetaldehyde-acetate pathway.45 Additional acetyl-CoA could be provided from the cleavage of citrate by ATP citrate lyase (ACL) in oleaginous microorganisms.46 Acetyl-CoA is then catalyzed to form malonyl-CoA by acetyl-CoA carboxylase (ACC), which is the important committing step of the fatty acid synthetic pathway.47 In E. coli, ACC is a protein containing four subunits with a biotin carboxyl carrier protein, a biotin-carboxylase subunit and a carboxyl-transferase subunit joined together into a heterotrimeric complex.38 In contrast, the ACC in the eukaryotic cells is a multi-domain single polypeptide, having all the functions of the ACC.38,47 Once malonyl-CoA is synthesized, it is condensed with acetyl-CoA catalyzed by the fatty acid synthetase (FAS) for fatty acid elongation. This resulting compound is transformed via three successive reactions, i.e., reduction, dehydration and reduction, and then condensed with another malonyl-CoA. This cycle is repeated until the saturated chain of a palmitic (16:0) or a stearic acid (18:0) is formed. Type II FAS, a multi-subunit protein in which each individual peptide is dissociable and can catalyze an enzymatic transacetylation reaction, is the major enzyme responsible for this process in bacteria and plant.48,49 In yeast, this enzyme is found to be the type I FAS, a multifunctional protein.45 To obtain longer or unsaturated chains, elongases and desaturases are required, which are usually located in endoplasmic reticulum membrane and mitochondria. During the fatty acids biosynthesis, NADPH is one of the important substrates required for two reduction steps in the fatty acid elongation cycle, which is mainly produced by malic enzyme (ME) and the pentose phosphate pathway.50 Fatty acid produced in the cell will then be transformed to various fatty acid esters or be degraded. β-oxidation is the principal metabolic pathway responsible for the degradation of fatty acids, which is taking place in the peroxisome,51 although the transport mechanism remains unclear.45
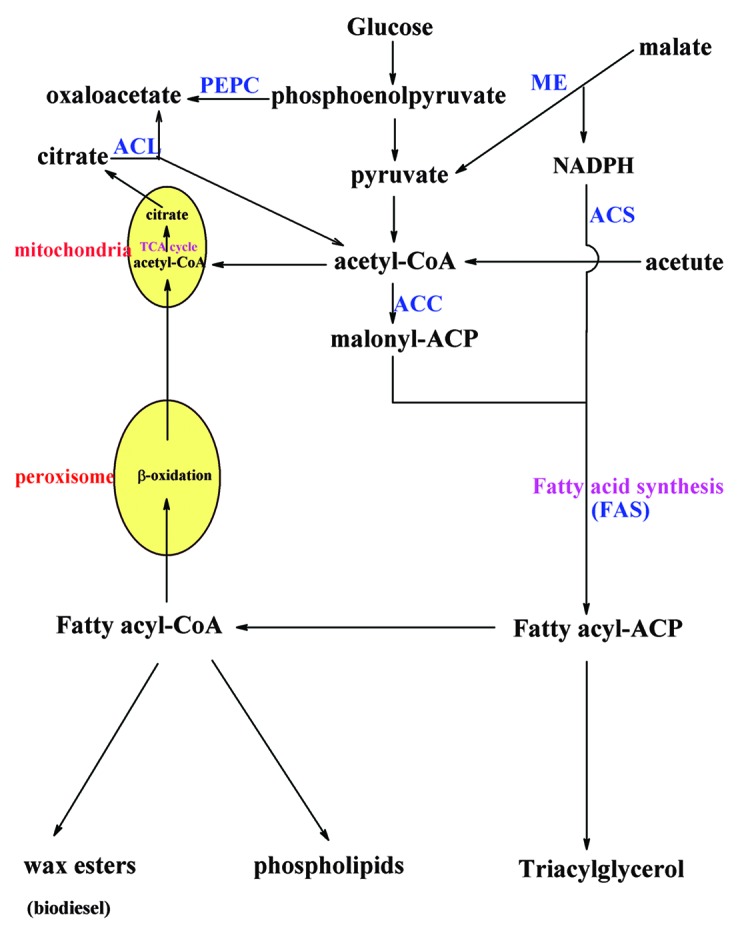
Figure 3. Simplified overview of fatty acid metabolism in microbes. ACL, ATP citrate lyase; ACS, acyl-CoA synthetases; ACC, acetyl-CoA carboxylase; ME, malic enzymes; PEPC, phosphoenolpyruvate carboxylase; FAS, fatty acid synthetase
Improving the fatty acids biosynthesis
The biochemical approach refers to the strategy of enhancing fatty acids accumulation by controlling the nutritional or cultivation conditions to channel metabolic flux into lipid biosynthesis.43 There is apparently a dilemma in the biochemical strategy, i.e., the conditions that stimulates fatty acids esters accumulation in cells may result in severely impeded cell growth and hence lowered overall lipid productivity.52 It is likely that the dilemma could be solved by employing metabolic engineering approaches aiming at enhancing the metabolic flux into fatty acids biosynthesis. Numerous studies have been carried using the genetic strategy to enhance the lipid accumulation in different species.
As an important enzyme catalyzing the first committed step in fatty acid biosynthesis in many organisms, ACC has traditionally been thought to exert a strong control on the metabolic flux of fatty acid synthesis. Overexpression of ACC did enhance the fatty acid synthesis in various organisms, including plant, bacteria, yeast, and so on.53-55 Ruenwai et al.55 isolated a multi-domain ACC from an oleaginous fungus Mucor rouxii. Overexpressing the M. rouxii ACC in a non-oleaginous yeast Hansenula polymorpha resulted in a 40% increase in the total fatty acid content. By the overproduction of ACC in E. coli, 100-fold increase in the malonyl-CoA pool was obtained accompanying by a 6-fold increase in fatty acid production.53 Similarly, Klaus et al.56 achieved a 30% increase in total fatty acid content in Solanum tuberosum (potato) by overexpressing the ACC from Arabidopsis. Nevertheless, the low increase ratios in the above-mentioned reports indicate that overexpression of ACC alone may not be adequate to affect the whole fatty acids biosynthesis significantly. Actually, ACC was also successfully overexpressed in the diatoms C. cryptica and N. saprophila. However, no significant increase of lipid accumulation was observed in the transgenic diatoms.57,58
Subsequent steps in fatty acid biosynthesis can also limit fatty acid flux. There are also other enzymes presenting a strong correlation with the fatty acid biosynthesis.50,59 The ATP: citrate lyase (ACL) catalyzes the conversion of citrate into acetyl-CoA and oxaloacetate, and represents a source of acetyl-CoA for fatty acid biosynthesis. An increased amount of fatty acids by 16% was obtained in tobacco by overexpression of a rat ACL gene leading to a 4-fold increase in the total ACL activity.60 Similar enhancement in the fatty acid synthesis could also be achieved by overexpression of acyl-CoA synthetases (ACSs), a family of enzymes that catalyze the thioesterification of fatty acids with coenzymeA to form activated intermediates.61 It was observed by Lin et al.62 that overexpressing the acs gene in E. coli resulted in a significant increase of the assimilation of acetate from the medium and contributed to an increase in the rate of fatty acid synthesis. ME, an enzyme catalyzes the conversion of malate into pyruvate and simultaneously reduces a NADP+ molecule into NADPH, is suggested to be a reducing power supplier for lipogenic enzymes such as ACC, FAS and ACL.63 Overexpressing ME has been demonstrated as an effective method to enhance lipid accumulation in some strains, such as Mucor circinelloides59 and S. cerevisiae.50 Thioesterases is responsible for the release of fatty acid chains from the acyl carrier protein and the production of free fatty acids. Overexpression of thioesterases is also able to increase fatty acid production substantially in many strains such as E. coli and cyanobacteria.64-67
Blocking the lipid degradation pathway and other metabolic pathways would also lead to the accumulation of fatty acid sources. For example, starchless mutant of Chlorella pyrenoidosa has been shown to have elevated polyunsaturated fatty acid content.68 Besides, β-oxidation is one of the major pathways responsible for metabolizing fatty acids, which consists of a set of four reactions operating at the carbons 2 or 3 of acyl-CoA esters and shortening of the acyl-chain.69 It is found that knocking out genes involved in β-oxidation not only can lead to increased amounts of intracellular free fatty acids but also result in extracellular fatty acid secretion in some cases.70-72 For example, deletion of the fadD gene (encoding the fatty acyl-CoA synthetase) in E. coli could result in an enhanced fatty acids accumulation.32 The same positive effect on the fatty acids accumulation achieved by knocking out of the fadE gene coding acyl-CoA dehydrogenase in E. coli.32 However, cells would rely on the β-oxidation of fatty acids for cellular energy under certain physiological conditions. As a result, knocking out lipid catabolism genes may have deleterious effects on cellular growth and proliferation.73 Cao et al.74 demonstrated that using an indirect method, i.e., inhibiting the acetyl-CoA transportation system required for coupling the β-oxidization in peroxisome and the TCA cycle in mitochondria but not any enzyme of the β-oxidation, is capable of inhibiting the β-oxidization of Candida tropicali. The third competitive pathway is the conversion of phosphoenolpyruvate to oxaloacetate, which is catalyzed by the phosphoenolpyruvate carboxylase (PEPC). Significantly enhanced lipid contents could be obtained by breaking the PEPC activity.75,76 By expressing antisense PEPC in Brassica napus, Chen et al.77 achieved a 6.4–18% increase in oil content, which suggests that reduced PEPC activity enhanced the fatty acid esters accumulation.
Recently, more and more researchers present a comprehensive modification suggestion of engineering more than one enzyme relevant to the fatty acids accumulation.43 One example of the comprehensive modification is shown in E. coli, which resulted in a 20-fold increase in free fatty acid production.66 Genetic engineering of the transcription factor, which affects up or downregulation of genes responsible for the global fatty acid pathways, would also be a potentially promising strategy among various comprehensive modifications.78-80 Reik et al.80 overexpressed a zinc-finger protein transcription factors that binds a DNA sequence within the promoter in microalgae and led to an enhanced lipid synthesis.
Modifying the fatty acids profiles
In addition to engineering microorganisms for the increased production of fatty acid esters, it is also reasonable to attempt to improve the quality of the fatty acid esters, with regard to suitability as a diesel fuel feedstock. The carbon chain length and degree of unsaturation of the fatty acids can affect the cold flow and oxidative stability properties of a biodiesel fuel which is derived from this feedstock.18
The chain lengths of fatty acids are mainly determined by acyl-ACP thioesterases, which can release the fatty acid chain from the FAS. There are several acyl-ACP thioesterases from a variety of organisms that are specific for certain fatty acid chain lengths. Transgenic overexpression of thioesterases are able to change fatty acid chain length.81,82 As in most other organisms, the major end products of the plant and E. coli FAS are usually 16- or 18-carbon fatty acids.83,84 However, certain plant species synthesize in developing seeds large amounts of predominantly medium-chain (C8 to C14) fatty acids, which are deposited in TAGs for long-term carbon storage.85 It is proposed that transgenically overexpression of shorter chain length specific acyl-ACP thioesterases would lead to the enhancement of shorter chain fatty acids. Actually, transformation and expression of a 12:0-biased thioesterase from Umbellularia californica in E. coli changed its lipid profiles drastically, with a great increase in the production of lauric acid.65 Besides, the production of myristic acid was found to be greatly increased by the expression of a 14:0-biased thioesterase from Cinnamomum camphorum.86 The similar phenomenon has also been shown in microalgae87 that the heterologous expression of two thioesterases, biased toward the production of lauric (C12:0) and myristic acid (C14:0), led to a increased accumulation of shorter chain length fatty acids in the eukaryotic microalgae Phaeodactylum tricornutum.
There are other enzymes having significant effect on the fatty acid modification reaction. Overexpression of the FabH protein, which is one of three β-ketoacyl-ACP synthases (KAS) in E. coli that catalyze the elongations necessary for fatty acid biosynthesis, influenced the cell by producing more shorter-chain fatty acids.88 Blocking the degradation of fatty acyl-CoA in E. coli through the deletion of the fadE gene encoding acyl-CoA dehydrogenase caused significant changes in the FAEEs composition and major products were ethyl palmitate (16:0) and ethyl oleate (18:1) rather than ethyl myristate (14:0) after its deletion.32
Genetic Engineering for Extracellular Secretion of Lipophilic Compounds
Prior to the use in the biodiesel industry, fatty acid esters or TAGs have to be separated from cells through a series of energy-intensive steps such as cell harvest, drying and solvent extraction. The cost of the energy-intensive processes usually accounts for more than 80% of total cost of biofuel production.89 Secretion of such lipophilic products into the culture medium rather than intracellular accumulation can significantly reduce the costs of product recovery by skipping these separation processes. Besides, extracellular production of lipophilic compounds would not be directly limited by cell density or cell volume. Indeed, the localization of neutral lipids in organisms is not restricted to the cell cytoplasm, as extracellular lipid deposition has been shown in various strains. There are many strains of bacteria presenting the ability of extracellular TAGs production, like Alcanivorax,90 Acinetobacter,91 Alcaligenes and Pseudomonas.92 It was also reported that similar secretion of fatty acids or TAGs can be achieved by metabolic engineering and mutagenesis, not only in bacterium E. coli, but also in some yeast strains and microalgae.71,93,94
Diffusion by concentration gradient and transportation by carrier proteins are the two ways for lipophilic compounds exporting.95-97 It is suggested that manipulation that allows yeast cells to accumulate high levels of intracellular fatty acids or that allows yeast to change the fatty acids length may be favorable for extracellular secretion of free fatty acids.18,98 Inactivation of genes involved in β-oxidation is one of the important manipulations that performed for enhancing fatty acid secretion. ACSs has been recognized as an intensively used gene in such cases.93 As a family of enzymes that catalyze the thioesterification of fatty acids with coenzyme A to form activated intermediates, ACSs plays a fundamental role in lipid metabolism, which are also necessary for fatty acid import into cells through the process of vectorial acylation.61 ACSs increases the concentration gradient between extracellular and intracellular fatty acid by converting fatty acid into acyl-CoA and contributes to the uptake of fatty acid.95 Thus, the deletion of ACSs gene might increase the concentration of free fatty acid in the cytosol and enhance fatty acid export out of cells. Scharnewski et al.93 demonstrated that in a mutant strain of S. cerevisiae deficient in ACSs, which could secrete fatty acid out of cells with a maximum titer of 200 μmol L−1. There is also ample evidence in E. coli proving fatty acid export out of cells is relevant with ACSs.95,98 Nevertheless, it was recently suggested that the deletion of ACSs genes is unable to improve the extracellular fatty acid production when the cell associated fatty acid titer was high. It is clear that fatty acid secretion was not only determined by the concentration gradient, but also related with other carrier proteins or some undetermined mechanisms.65,98 Thioesterase is also known to influence the fatty acids secretion. For instance, expression of acyl-ACP thioesterase from Umbellularia californica in the fatty acid-degradation mutant E. coli led to the secretion of free fatty acid into medium.65 In the study of Liu et al.,98 an engineered E. coli was obtained with enhanced extracellular fatty acids production through the cytosolic expression of an engineered E. coli thioesterase (a ‘leadless’ version of thioesterase without the NH2-terminal 26 amino acid residues). The similar phenomenon was also found in the cyanobacterium Synechocystis sp PCC 6803. By introducing acyl-acyl carrier protein thioesterases, the cyanobacterium Synechocystis sp PCC 6803 were able to overproduce fatty acids (C10-C18) and secrete them into the medium at an efficiency of up to 133 ± 12 mg L−1 of culture per day at a cell density of 1.5 × 108 cells mL−1 (0.23 g of dry weight per liter). Transportation with the help of carrier proteins is proposed to be another type for fatty acid export out of cells. The process for fatty acid transport is inducible and commensurate with the expression of specific sets of proteins thought to participate in the secretion.99,100 FadL is outer membrane protein in E. coli, which has been found to be specifically involved in the binding and translocation of the long-chain fatty acids across the outer membrane of the cell. A tight linkage exists between FadL and fatty acyl CoA synthetase, indicating that these two proteins must act in concert in the facilitated transport and activation of these hydrophobic compounds.96,98,101,102 As another recently discovered lipid transporter, MsbA is the ATP-binding cassette (ABC) transporter and is implicated in the export of lipopolysaccharides and phospholipids in E. coli.103,104 The fatty acid transport apparatus in yeast is quite distinct from that defined in E. coli but may emulate that being defined in higher eukaryotic cells. In S. cerevisiae, a protein Fat1p (encoded within the fat1 gene) was identified as a plasma membrane-bound long-chain fatty acid transporter,105 which acted with other two proteins (i.e., Faa1p and Faa4p) to facilitate the transport and activation of exogenous long-chain fatty acids.93,105,106 Besides, the secretion yields of fatty acid or lipid could also be increased by weakening the S-layer and peptidoglycan layers to facilitate fatty acids secretion.98,107
Genetic Engineering for Intracellular Transesterification
Enzymatic transesterification by lipases is a method extensively used for FAMEs or FAEEs production.9 Fatty acids sources and short chain alcohols are essential substrates in the transesterification processes with lipases as biocatalyst, while most of the catalyzed processes are performed in organic solvents in the absence of water.12 Over the past few years, much effort has been devoted to constructing intracellular transesterification systems for microbial biodiesel production. Acyltransferases, a class of enzymes which are responsible for wax ester or TAG biosynthesis in vivo (Fig. 4), present the potentiality to be used as an alternative for lipase-catalyzed acylation in the biodiesel production.108-111 Long-chain fatty alcohols or diacylglycerols and fatty acid coenzyme A thioesters (acyl-CoA) is used as substrates for the transesterification of acyltransferases for wax ester or TAG biosynthesis.112 The biotechnologically relevant synthesis reactions are performed in aqueous systems, while the enzymes are only active with CoA-activated fatty acids.113 The dependence of acyltransferases on CoA esters suggests that the acyltransferase-catalyzed acylation is the native reaction under physiological conditions. Thus, the acyltransferanse mediated biodiesel synthesis would preferably be performed in vivo. In vitro applications using acyltransferanse as biocatalyst in biotechnological processes will not be economic in analogy to lipase catalyzed esterifications. However, the acyltransferanse mediated biodiesel production is the possible development of processes that can utilize bulk materials, such as sugars, or better starch, cellulose, and hemicellulose as substrates, which are not dependant on oilseed crops. This will not only significantly extend the substrate range that can be used for fuel production, but will also use resources that do not compete with food and feed production.113

Figure 4. In vivo reaction catalyzed by acyltransferases for the synthesis of wax ester and triacylglycerol (TAGs). WS, Wax ester synthase; DGAT, diacylglycerol acyltransferase
There are three unrelated families of acyltransferases found in higher plants, mammals and bacteria, which might be suitable for biodiesel synthesis. The first identified acyltransferase of plants is jojoba embryo wax ester synthase, which did not show activity when heterologously expressed in S. cerevisiae.114 The second group of identified acyltransferases was from bacteria, which is a bifunctional enzyme functioning as wax ester acyltransferases (WS) and as acyl-CoA:diacylglycerol acyltransferase (DGAT). The third group of acyltransferases is from mammals such as the acyltransferase from Mus musculus C57BL/6.115 The WS/DGAT from bacterium A. baylyi ADP1 is an acyltransferase extensively used for microbial biodiesel production.33,108-111,116 However, the reaction rate of the WS/DGAT enzyme is low with ethanol as substrate,110,113 since the WS/DGAT from A. baylyi ADP1 displayed a general preference for long-chain alcohols and a lower activity for short-chain alcohols. Luckily, WS/DGAT homologs are frequently found in many organism genomes.112 All the acyltransferase from A. baylyi ADP1, Marinobacter hydrocarbonoclasticus DSM 8798, Rhodococcus opacus PD630, M. musculus C57BL/6 and Psychrobacter arcticus 273–4 can responsible for the formation of biodiesel.39 It is very likely that more effective candidates for biodiesel synthesis can be found among the acyltransferases from different organisms with various substrate and product chain-length preferences. Indeed, an acyltransferase from M. hydrocarbonoclasticus DSM 8798 without DGAT activity was found to be more suitable for producing biodiesel since it avoids direction of fatty acyl-CoAs toward TAG biosynthesis.117 Additionally, directed evolution of acyltransferases may result in selection of enzymes with higher specificity toward ethanol. Furthermore, the substrate specificities of WSs vary with the different expression hosts,109,114 indicating the importance of exploring suitable hosts for the biodiesel synthesis. The WS/DGAT from A. baylyi strain ADP1 is active in different microbial hosts, such as E. coli, Pseudomonas citronellolis and S. cerevisiae.109,118,119 Active expression of the WS/DGAT homolog from M. smegmatis could also be achieved in E. coli and Rhodococcus opacus.118,120
Genetic Engineering in Substrates Metabolism
Fermentation economics depend on a variety of factors, including investment costs for the fermentation equipment, raw materials and maintenance plus costs for upstream and downstream processes as well as the process yield and throughput.121 In particular, it may be highly desirable and perhaps necessary to incorporate the feature of naturally utilization of agro-industrial wastes into these cell factories to decrease the investment costs when developing recombinant biodiesel producers. Utilization of agro-industrial residues to make biofuel has become one of the hottest points in current researches.122-125
Lignocellulosic biomass
De novo biosynthesis of FAEEs from lignocellulosic biomass has been achieved by genetic manipulation in the study of Duan et al.32As inexpensive materials for biotechnology production, lignocellulosic plant biomass represents the largest source of renewable carbon and consists of 40–55% cellulose, 25–50% hemicellulose and 10–40% lignin. The social and economic benefits of producing biofuel from lignocellulosic biomass instead of crops are widely appreciated.126 To do so, lignocellulosic plant biomass must be broken down and hydrolyzed into fermentable sugars.127 Large quantities of glycoside hydrolase (GH) enzymes are demand for the efficiently conversion of lignocellulose into fermentable sugars. Actually, the substantial capital and material expense occurred in the enzymes preparation has become the second highest contribution to raw material cost after the feedstock itself.127 Combination of lignocellulose-utilization and product formation properties in one single microorganism, also called consolidated bioprocess (CBP), might be a potential breakthrough. Hydrolyze the cellulose and hemicelluloses in biomass and produce a valuable product currently accomplished in different reactors or different organisms, are combined in CBP.128 The key point of CBP for biodiesel production is the engineering of a microorganism that can efficiently de-polymerize biomass polysaccharides to fermentable sugars and efficiently convert this mixed-sugar hydrolysate into FAEEs (Fig. 5).

Figure 5. Graphic illustration of a lignocellulose conversion to biodiesel in a single microbe by consolidated bioprocess (CBP). The enzymatic hydrolysis of the cellulose and hemicellulose fractions to fermentable hexoses and pentoses requires the production of both cellulases and hemicellulases (dashed lines). The subsequent conversion of the hexoses and pentoses to biodiesel initiated by the modifying fatty acids pathways, introducing ethanol producing pathway and overexpressing acyltransferase. FFA, fatty acid synthesis; WS/DGAT, acyltransferase for fatty acid esters synthesis; EtOH, ethanol producing pathway.
Two strategies can be used to develop CBP organisms: (1) engineering naturally cellulolytic microorganisms to improve product-related properties; and (2) engineering non-cellulolytic organisms with high product yields to become cellulolytic.129 Selection of suitable and efficient cellulolytic enzymes is a key factor in the process of engineering non-cellulolytic organisms with high product yields. Full enzymatic hydrolysis of crystalline cellulose usually requires synergistic action of three major types of enzymatic activity: (1) endoglucanases (EGs) (1,4-β-D-glucan 4-glucanohydrolases; EC 3.2.1.4); (2) exoglucanases, including cellodextrinases (1,4-β-D-glucan glucanohydrolases; EC 3.2.1.74), and cellobiohydrolases (CBHs) (1,4-β-D-glucan cellobiohydrolases; EC 3.2.1.91); and (3) β-glucosidases (BGLs) (β-glucoside glucohydrolases; EC 3.2.1.21).130 Endoglucanases are active on the amorphous regions of cellulose and yield cellobiose and cello-oligosaccharides as hydrolysis products, while cellobiohydrolases are active on the crystalline regions of cellulose and yield almost exclusively cellobiose as their main hydrolysis product. Furthermore, β-glucosidases convert cellobiose and some cellooligosaccharides to glucose and then taken up by organisms. Likewise, the synergistic action of a plethora of enzymes is required for the hydrolysis of hemicellulose, another fraction contained in the lignocellulosic biomass with high content. Over the past decade, a lot of efficient lignocellulose degrading enzymes have been exploited and functionally active expressed in non-cellulolytic or weak-cellulolytic microorganisms (Table 1). Directly into ethanol or other fuels from some model cellulosic and hemicellulosic substrates has been achieved in several non-cellulolytic microorganisms after the introduction of lignocellulose degrading enzyme genes. For example, introduction of cellulase genes enabled the yeast S. cerevisiae123 and the bacterium Klebsiella oxytoca131 directly convert phosphoric acid swollen cellulose (PASC) to ethanol without the addition of exogenous cellulase and hemicellulase. E. coli is a popular host for engineering in the biofuel production. Construction of recombinant E. coli with CBP capability is of great practical significance. However, the inferior capacity in E. coli for protein export has rendered it unable to extracellular produce cellulase or hemicellulase in quantities required for industrial-scale lignocellulose hydrolysis. Various techniques, developed over decades of research, can be applied to generate secreted yields from E. coli,132 although these concentrations are still too low for an industrial process. In 2010, Steen et al.33 constructed an FAEEs-producing E. coli with extracellular hemicellulase activity to use the xylan in the growth medium. Then, Bokinsky et al. engineered E. coli extracellular expressing cellulase, xylanase, β-glucosidase, and xylobiosidase enzymes under control of native E. coli promoters by the fusion with export related protein (e.g., protein OsmY). The engineered E. coli grew well and produced fuel substitutes using either the cellulose or hemicellulose components of ionic liquid-pretreated switchgrass or on both components when combined as a coculture.133 Removal of soluble inhibitors in the hydrolysates may reduce the enzyme loading required for the industrial process in the biomass hydrolysis.134,135 Therefore, even though the yields of secreted protein (<0.1 mg enzymes/g solids) in the study of Bokinsky et al.133 was low, the E. coli is capable of growing on the cellulose and hemicellulose fractions of the ionic liquid-pretreated plant biomass.
Table 1. Common lignocellulose degrading enzymes expressed in non-cellulolytic or weak-cellulolytic microorganisms.
| Enzyme or gene |
Source-species |
Receiver-species |
|---|---|---|
| Cellulose degradation: endoglucanase (EG) | ||
| EGI |
Trichoderma reesei |
Saccharomyces cerevisiae,143Yarrowia lipolytica,144 Ashbya gossypii145 |
| EGII |
T. reesei |
S. cerevisiae,124Y. lipolytica,146Pichia pastoris,147Kluyveromyces marxianus148 |
| EG(CelE) |
Clostridium thermocellum |
S. cerevisiae,149Escherichia coli150 |
| EG (CelA) |
C. thermocellum |
Bacillus subtilis,151Lactobacillus gasseri,152Lactobacillus johnsoni152 |
| EG (Cel9A) |
Thermobifida fusca |
S. cerevisiae
153
|
| EGIII | Trichoderma harzianum | P. pastoris 154 |
| Cellobiohydrolase (CBH) | ||
|---|---|---|
| CBHI |
T. reesei |
S.cerevisiae,153P. pastoris155 |
| CBHII |
T. reesei |
S. cerevisiae156,Y. lipolytica,146P.pastoris,146Aspergillus oryzae156 |
| CBHII |
Magnaporthe oryzae |
A. oryzae
156
|
| CBHI |
Talaromyces emersonii |
S. cerevisiae
157
|
| CBH (Cel48) | Clostridium phytofermentans | E. col,158B. subtilis158 |
| β-Glucosidase (BGL) | ||
|---|---|---|
| BGL1 |
Saccharomycopsis fibuligera |
S.cerevisiae
149
|
| BGL1 |
Aspergillus aculeatus |
S. cerevisiae,148K. marxianus,147T. reesei,159Issatchenkia orientalis160 |
| BGL1 |
A. oryzae |
S.cerevisiae
125
|
| BGL | Penicillium brasilianum | A. oryzae 161 |
| Hemicellulose degradation: | ||
|---|---|---|
| β-Xylanase(Xyn2) |
T. reesei |
E. coli,162P. pastoris,163S. cerevisiae164 |
| β-Xylanase (xynA) |
Penicillium citrinum |
P. pastoris
165
|
| β-Xylosidase |
T. reesei |
S. cerevisiae
166
|
| β-Mannanase |
A. aculeatus |
Y. lipolytica,167S. cerevisiae,168Aspergillus niger169 |
| endo-β-1,4-Glucanase | A. aculeatus | A. oryzae 170 |
Crude glycerol
The upsurge in biodiesel production has resulted in a market surplus of glycerol, a byproduct of the chemical transesterification process.14 Glycerol can be used as inexpensive and readily available raw material to produce fuels and chemicals in some industrial fermentation processes.136 There have been intensive attempts to identify glycerol assimilation pathway in bacterial or yeast strains.137-140 The possibility to recycle glycerol, which is the byproduct of FAEEs, for FAEEs production was demonstrated in the report of Yu et al.40 and microbial biodiesel production from glycerol was successfully achieved in S. cerevisiae. The application of glycerol for biodiesel production was also demonstrated by Elbahloul and Steinbuchel34 by the pilot-scale cultivation of E. coli harboring a plasmid with ethanol producing enzymes and acyltransferases. Besides, crude glycerol contains water, salts and other organic materials including residual methanol and free fatty acids and each component varies widely in contents.141 Therefore, exploration of microbial strains tolerant to undesirable inhibitory components such as salts and organic solvents that present in crude glycerol becomes a challenge in low-grade crude glycerol fermentation.
Restaurant oil wastes
Restaurant oil wastes, rich in free fatty acids, have be utilized to provide fatty acid sources for FAEEs-based biodiesel production in the report of Wang et al.31 The exploration of strategies to utilize restaurant oil wastes for biodiesel production offers significant advantages, such as enhancing the economic viability of biodiesel, not competing with the food market and maintaining the health of the environment.142 However, it should be noted that the FAEEs yielded from the restaurant oil wastes is still extremely lower than that from the standard medium containing sodium oleate, which indicates that much more further investigations are needed before its industrial application.
Conclusion
Biodiesel is produced by transesterification of fatty acid sources (e.g., TAGs) with short-chain alcohols yielding monoalkyl esters of long-chain fatty acids such as FAMEs and FAEEs. To realize this process, microorganisms must be developed to have the ability to produce fatty acids and short-chain alcohols available for transesterfication and to possess acyltransferases with a higher activity for short-chain alcohols. Although all of these capabilities are involved by known microorganisms, de novo synthesis of fatty acid ester based biodiesel (FAEEs) has not been found in nature microorganisms to date. Experimental evidence and fundamental principles of the related pathways support the feasibility of de novo biosynthesis of biodiesel by one single microorganism after reasonable modification of potential host organisms. Strains of E. coli are found to be the most popular host for this engineering. However, it is clear that biodiesel production in the current engineered strains is still far from industrial application. Developing microbes with both higher efficiency and capability of costless substrates is urgent in order to compete with the fossil fuel. It could be noted many pathways would influence the biodiesel synthesis. Overexpression of one or several specific enzyme genes may not cause dramatic facilitation. Systems metabolic engineering might be more suitable for the development of such microbial strains having potential commercial applications. Furthermore, how robust the microorganism for a large-scale industrial process is another problem should be given adequate consideration.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Acknowledgments
This study was supported by the International Cooperation Project in Science and Technology of Zhejiang Province (2008C14038), the Science and Technology Project of Zhejiang Province (2011C13016), the National Natural Science Foundation of China (31070079), and the National Key Technology Research and Development Program of the Ministry of Science and Technology of China (2012BAC17B04).
Submitted
10/30/12
Revised
12/02/12
Accepted
12/04/12
Footnotes
Previously published online: www.landesbioscience.com/journals/bioe/article/23114
References
- 1.Khan SA. Rashmi, Hussain MZ, Prasad S, Banerjee UC. Prospects of biodiesel production from microalgae in India. Renew Sustain Energy Rev. 2009;13:2361–72. doi: 10.1016/j.rser.2009.04.005. [DOI] [Google Scholar]
- 2.Shafiee S, Topal E. When will fossil fuel reserves be diminished? Energy Policy. 2009;37:181–9. doi: 10.1016/j.enpol.2008.08.016. [DOI] [Google Scholar]
- 3.Vasudevan PT, Briggs M. Biodiesel production--current state of the art and challenges. J Ind Microbiol Biotechnol. 2008;35:421–30. doi: 10.1007/s10295-008-0312-2. [DOI] [PubMed] [Google Scholar]
- 4.Sorrell S, Speirs J, Bentley R, Brandt A, Miller R.Global oil depletion: an assessment of the evidence for a near-term peak in global oil production. 2009.
- 5.Ali Y, Hanna MA. Alternative diesel fuels from vegetable oils. Bioresour Technol. 1994;50:153–63. doi: 10.1016/0960-8524(94)90068-X. [DOI] [Google Scholar]
- 6.Kaushik N, Kumar K, Kumar S. Potential of Jatropha curcas for Biofuels. Journal of Biobased Materials and Bioenergy. 2007;1:301–14. doi: 10.1166/jbmb.2007.002. [DOI] [Google Scholar]
- 7.Wen D, Jiang H, Zhang K. Supercritical fluids technology for clean biofuel production. Prog Nat Sci. 2009;19:273–84. doi: 10.1016/j.pnsc.2008.09.001. [DOI] [Google Scholar]
- 8.Lam MK, Tan KT, Lee KT, Mohamed AR. Malaysian palm oil: Surviving the food versus fuel dispute for a sustainable future. Renew Sustain Energy Rev. 2009;13:1456–64. doi: 10.1016/j.rser.2008.09.009. [DOI] [Google Scholar]
- 9.Parawira W. Biotechnological production of biodiesel fuel using biocatalysed transesterification: A review. Crit Rev Biotechnol. 2009;29:82–93. doi: 10.1080/07388550902823674. [DOI] [PubMed] [Google Scholar]
- 10.Yusuf NNAN, Kamarudin SK, Yaakub Z. Overview on the current trends in biodiesel production. Energy Convers Manage. 2011;52:2741–51. doi: 10.1016/j.enconman.2010.12.004. [DOI] [Google Scholar]
- 11.Lotero E, Liu YJ, Lopez DE, Suwannakarn K, Bruce DA, Goodwin JG. Synthesis of biodiesel via acid catalysis. Ind Eng Chem Res. 2005;44:5353–63. doi: 10.1021/ie049157g. [DOI] [Google Scholar]
- 12.Du W, Li W, Sun T, Chen X, Liu DH. Perspectives for biotechnological production of biodiesel and impacts. Appl Microbiol Biotechnol. 2008;79:331–7. doi: 10.1007/s00253-008-1448-8. [DOI] [PubMed] [Google Scholar]
- 13.Nielsen PM, Brask J, Fjerbaek L. Enzymatic biodiesel production: technical and economical considerations. Eur J Lipid Sci Technol. 2008;110:692–700. doi: 10.1002/ejlt.200800064. [DOI] [Google Scholar]
- 14.Ma FR, Hanna MA. Biodiesel production: a review. Bioresour Technol. 1999;70:1–15. doi: 10.1016/S0960-8524(99)00025-5. [DOI] [Google Scholar]
- 15.Yan-yan ZZHUA, Bo L. How to Secure Triacylglycerol Supply for Chinese Biodiesel Industry. Progress In Biotechnology. 2005;11 [Google Scholar]
- 16.Peralta-Yahya PP, Keasling JD. Advanced biofuel production in microbes. Biotechnol J. 2010;5:147–62. doi: 10.1002/biot.200900220. [DOI] [PubMed] [Google Scholar]
- 17.Kosa M, Ragauskas AJ. Lipids from heterotrophic microbes: advances in metabolism research. Trends Biotechnol. 2011;29:53–61. doi: 10.1016/j.tibtech.2010.11.002. [DOI] [PubMed] [Google Scholar]
- 18.Radakovits R, Jinkerson RE, Darzins A, Posewitz MC. Genetic engineering of algae for enhanced biofuel production. Eukaryot Cell. 2010;9:486–501. doi: 10.1128/EC.00364-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Li Q, Du W, Liu D. Perspectives of microbial oils for biodiesel production. Appl Microbiol Biotechnol. 2008;80:749–56. doi: 10.1007/s00253-008-1625-9. [DOI] [PubMed] [Google Scholar]
- 20.Mata TM, Martins AA, Caetano NS. Microalgae for biodiesel production and other applications: A review. Renew Sustain Energy Rev. 2010;14:217–32. doi: 10.1016/j.rser.2009.07.020. [DOI] [Google Scholar]
- 21.Khozin-Goldberg I, Cohen Z. The effect of phosphate starvation on the lipid and fatty acid composition of the fresh water eustigmatophyte Monodus subterraneus. Phytochemistry. 2006;67:696–701. doi: 10.1016/j.phytochem.2006.01.010. [DOI] [PubMed] [Google Scholar]
- 22.Li X, Hu HY, Zhang YP. Growth and lipid accumulation properties of a freshwater microalga Scenedesmus sp. under different cultivation temperature. Bioresour Technol. 2011;102:3098–102. doi: 10.1016/j.biortech.2010.10.055. [DOI] [PubMed] [Google Scholar]
- 23.Lin Y, Tanaka S. Ethanol fermentation from biomass resources: current state and prospects. Appl Microbiol Biotechnol. 2006;69:627–42. doi: 10.1007/s00253-005-0229-x. [DOI] [PubMed] [Google Scholar]
- 24.Weber C, Farwick A, Benisch F, Brat D, Dietz H, Subtil T, et al. Trends and challenges in the microbial production of lignocellulosic bioalcohol fuels. Appl Microbiol Biotechnol. 2010;87:1303–15. doi: 10.1007/s00253-010-2707-z. [DOI] [PubMed] [Google Scholar]
- 25.Yanase H, Miyawaki H, Sakurai M, Kawakami A, Matsumoto M, Haga K, et al. Ethanol production from wood hydrolysate using genetically engineered Zymomonas mobilis. Appl Microbiol Biotechnol. 2012;94:1667–78. doi: 10.1007/s00253-012-4094-0. [DOI] [PubMed] [Google Scholar]
- 26.Wang YZ, Manow R, Finan C, Wang JH, Garza E, Zhou SD. Adaptive evolution of nontransgenic Escherichia coli KC01 for improved ethanol tolerance and homoethanol fermentation from xylose. J Ind Microbiol Biotechnol. 2011;38:1371–7. doi: 10.1007/s10295-010-0920-5. [DOI] [PubMed] [Google Scholar]
- 27.Hong KK, Nielsen J. Metabolic engineering of Saccharomyces cerevisiae: a key cell factory platform for future biorefineries. Cell Mol Life Sci. 2012;69:2671–90. doi: 10.1007/s00018-012-0945-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Seo JS, Chong H, Park HS, Yoon KO, Jung C, Kim JJ, et al. The genome sequence of the ethanologenic bacterium Zymomonas mobilis ZM4. Nat Biotechnol. 2005;23:63–8. doi: 10.1038/nbt1045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Rogers PL, Jeon YJ, Lee KJ, Lawford HG. Zymomonas mobilis for fuel ethanol and higher value products. Adv Biochem Eng Biotechnol. 2007;108:263–88. doi: 10.1007/10_2007_060. [DOI] [PubMed] [Google Scholar]
- 30.Ingram LO, Conway T, Clark DP, Sewell GW, Preston JF. Genetic engineering of ethanol production in Escherichia coli. Appl Environ Microbiol. 1987;53:2420–5. doi: 10.1128/aem.53.10.2420-2425.1987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Wang B, Lin H, Zhan J, Yang Y, Zhou Q, Zhao Y. Biodiesel synthesis by a one-step method in a genetically engineered Escherichia coli using rice straw hydrolysate and restaurant oil wastes as raw materials. J Appl Microbiol. 2012;113:531–40. doi: 10.1111/j.1365-2672.2012.05357.x. [DOI] [PubMed] [Google Scholar]
- 32.Duan YK, Zhu Z, Cai K, Tan XM, Lu XF. De novo biosynthesis of biodiesel by Escherichia coli in optimized fed-batch cultivation. PLoS One. 2011;6:e20265. doi: 10.1371/journal.pone.0020265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Steen EJ, Kang Y, Bokinsky G, Hu Z, Schirmer A, McClure A, et al. Microbial production of fatty-acid-derived fuels and chemicals from plant biomass. Nature. 2010;463:559–62. doi: 10.1038/nature08721. [DOI] [PubMed] [Google Scholar]
- 34.Elbahloul Y, Steinbüchel A. Pilot-scale production of fatty acid ethyl esters by an engineered Escherichia coli strain harboring the p(Microdiesel) plasmid. Appl Environ Microbiol. 2010;76:4560–5. doi: 10.1128/AEM.00515-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Handke P, Lynch SA, Gill RT. Application and engineering of fatty acid biosynthesis in Escherichia coli for advanced fuels and chemicals. Metab Eng. 2011;13:28–37. doi: 10.1016/j.ymben.2010.10.007. [DOI] [PubMed] [Google Scholar]
- 36.Huffer S, Roche CM, Blanch HW, Clark DS. Escherichia coli for biofuel production: bridging the gap from promise to practice. Trends Biotechnol. 2012;30:538–45. doi: 10.1016/j.tibtech.2012.07.002. [DOI] [PubMed] [Google Scholar]
- 37.Kalscheuer R, Stölting T, Steinbüchel A. Microdiesel: Escherichia coli engineered for fuel production. Microbiology. 2006;152:2529–36. doi: 10.1099/mic.0.29028-0. [DOI] [PubMed] [Google Scholar]
- 38.Tehlivets O, Scheuringer K, Kohlwein SD. Fatty acid synthesis and elongation in yeast. Biochim Biophys Acta. 2007;1771:255–70. doi: 10.1016/j.bbalip.2006.07.004. [DOI] [PubMed] [Google Scholar]
- 39.Shi SB, Valle-Rodriguez JO, Khoomrung S, Siewers V, Nielsen J. Functional expression and characterization of five wax ester synthases in Saccharomyces cerevisiae and their utility for biodiesel production. Biotechnology for Biofuels 2012; 5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Yu KO, Jung J, Kim SW, Park CH, Han SO. Synthesis of FAEEs from glycerol in engineered Saccharomyces cerevisiae using endogenously produced ethanol by heterologous expression of an unspecific bacterial acyltransferase. Biotechnol Bioeng. 2012;109:110–5. doi: 10.1002/bit.23311. [DOI] [PubMed] [Google Scholar]
- 41.Holder JW, Ulrich JC, DeBono AC, Godfrey PA, Desjardins CA, Zucker J, et al. Comparative and functional genomics of Rhodococcus opacus PD630 for biofuels development. PLoS Genet. 2011;7:e1002219. doi: 10.1371/journal.pgen.1002219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Loira N, Dulermo T, Nicaud JM, Sherman DJ. A genome-scale metabolic model of the lipid-accumulating yeast Yarrowia lipolytica. BMC Syst Biol. 2012;6:35. doi: 10.1186/1752-0509-6-35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Courchesne NMD, Parisien A, Wang B, Lan CQ. Enhancement of lipid production using biochemical, genetic and transcription factor engineering approaches. J Biotechnol. 2009;141:31–41. doi: 10.1016/j.jbiotec.2009.02.018. [DOI] [PubMed] [Google Scholar]
- 44.Jian-jun XGLIU, Yuan-sen ZXLI. Application of microbial oil and research on oleaginous microorganisms. Cereals & Oils. 2008;6:004. [Google Scholar]
- 45.Beopoulos A, Nicaud JM, Gaillardin C. An overview of lipid metabolism in yeasts and its impact on biotechnological processes. Appl Microbiol Biotechnol. 2011;90:1193–206. doi: 10.1007/s00253-011-3212-8. [DOI] [PubMed] [Google Scholar]
- 46.Ratledge C. Fatty acid biosynthesis in microorganisms being used for Single Cell Oil production. Biochimie. 2004;86:807–15. doi: 10.1016/j.biochi.2004.09.017. [DOI] [PubMed] [Google Scholar]
- 47.Sasaki Y, Nagano Y. Plant acetyl-CoA carboxylase: structure, biosynthesis, regulation, and gene manipulation for plant breeding. Biosci Biotechnol Biochem. 2004;68:1175–84. doi: 10.1271/bbb.68.1175. [DOI] [PubMed] [Google Scholar]
- 48.Subrahmanyam S, Cronan JE., Jr. Overproduction of a functional fatty acid biosynthetic enzyme blocks fatty acid synthesis in Escherichia coli. J Bacteriol. 1998;180:4596–602. doi: 10.1128/jb.180.17.4596-4602.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Rock CO, Jackowski S. Forty years of bacterial fatty acid synthesis. Biochem Biophys Res Commun. 2002;292:1155–66. doi: 10.1006/bbrc.2001.2022. [DOI] [PubMed] [Google Scholar]
- 50.Moreira dos Santos M, Raghevendran V, Kötter P, Olsson L, Nielsen J. Manipulation of malic enzyme in Saccharomyces cerevisiae for increasing NADPH production capacity aerobically in different cellular compartments. Metab Eng. 2004;6:352–63. doi: 10.1016/j.ymben.2004.06.002. [DOI] [PubMed] [Google Scholar]
- 51.Shen YQ, Burger G. Plasticity of a key metabolic pathway in fungi. Funct Integr Genomics. 2009;9:145–51. doi: 10.1007/s10142-008-0095-6. [DOI] [PubMed] [Google Scholar]
- 52.Ratledge C. Regulation of lipid accumulation in oleaginous micro-organisms. Biochem Soc Trans. 2002;30:1047–50. doi: 10.1042/BST0301047. [DOI] [PubMed] [Google Scholar]
- 53.Davis MS, Solbiati J, Cronan JE., Jr. Overproduction of acetyl-CoA carboxylase activity increases the rate of fatty acid biosynthesis in Escherichia coli. J Biol Chem. 2000;275:28593–8. doi: 10.1074/jbc.M004756200. [DOI] [PubMed] [Google Scholar]
- 54.Klaus D, Ohlrogge JB, Neuhaus HE, Dörmann P. Increased fatty acid production in potato by engineering of acetyl-CoA carboxylase. Planta. 2004;219:389–96. doi: 10.1007/s00425-004-1236-3. [DOI] [PubMed] [Google Scholar]
- 55.Ruenwai R, Cheevadhanarak S, Laoteng K. Overexpression of acetyl-CoA carboxylase gene of Mucor rouxii enhanced fatty acid content in Hansenula polymorpha. Mol Biotechnol. 2009;42:327–32. doi: 10.1007/s12033-009-9155-y. [DOI] [PubMed] [Google Scholar]
- 56.Klaus D, Ohlrogge JB, Neuhaus HE, Dörmann P. Increased fatty acid production in potato by engineering of acetyl-CoA carboxylase. Planta. 2004;219:389–96. doi: 10.1007/s00425-004-1236-3. [DOI] [PubMed] [Google Scholar]
- 57.Dunahay TG, Jarvis EE, Roessler PG. Genetic transformation of the diatoms Cyclotella cryptica and Navicula saprophila. J Phycol. 1995;31:1004–12. doi: 10.1111/j.0022-3646.1995.01004.x. [DOI] [Google Scholar]
- 58.Dunahay TG, Jarvis EE, Dais SS, Roessler PG. Manipulation of microalgal lipid production using genetic engineering. Appl Biochem Biotechnol. 1996;57-8:223–31. doi: 10.1007/BF02941703. [DOI] [Google Scholar]
- 59.Zhang Y, Adams IP, Ratledge C. Malic enzyme: the controlling activity for lipid production? Overexpression of malic enzyme in Mucor circinelloides leads to a 2.5-fold increase in lipid accumulation. Microbiology. 2007;153:2013–25. doi: 10.1099/mic.0.2006/002683-0. [DOI] [PubMed] [Google Scholar]
- 60.Rangasamy D, Ratledge C. Genetic enhancement of fatty acid synthesis by targeting rat liver ATP:citrate lyase into plastids of tobacco. Plant Physiol. 2000;122:1231–8. doi: 10.1104/pp.122.4.1231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Black PN, DiRusso CC. Yeast acyl-CoA synthetases at the crossroads of fatty acid metabolism and regulation. Biochim Biophys Acta. 2007;1771:286–98. doi: 10.1016/j.bbalip.2006.05.003. [DOI] [PubMed] [Google Scholar]
- 62.Lin H, Castro NM, Bennett GN, San KY. Acetyl-CoA synthetase overexpression in Escherichia coli demonstrates more efficient acetate assimilation and lower acetate accumulation: a potential tool in metabolic engineering. Appl Microbiol Biotechnol. 2006;71:870–4. doi: 10.1007/s00253-005-0230-4. [DOI] [PubMed] [Google Scholar]
- 63.Wynn JP, bin Abdul Hamid A, Ratledge C. The role of malic enzyme in the regulation of lipid accumulation in filamentous fungi. Microbiology. 1999;145:1911–7. doi: 10.1099/13500872-145-8-1911. [DOI] [PubMed] [Google Scholar]
- 64.Lennen RM, Braden DJ, West RA, Dumesic JA, Pfleger BF. A process for microbial hydrocarbon synthesis: Overproduction of fatty acids in Escherichia coli and catalytic conversion to alkanes. Biotechnol Bioeng. 2010;106:193–202. doi: 10.1002/bit.22660. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Voelker TA, Davies HM. Alteration of the specificity and regulation of fatty acid synthesis of Escherichia coli by expression of a plant medium-chain acyl-acyl carrier protein thioesterase. J Bacteriol. 1994;176:7320–7. doi: 10.1128/jb.176.23.7320-7327.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Lu XF, Vora H, Khosla C. Overproduction of free fatty acids in E. coli: implications for biodiesel production. Metab Eng. 2008;10:333–9. doi: 10.1016/j.ymben.2008.08.006. [DOI] [PubMed] [Google Scholar]
- 67.Jiang P, Cronan JE., Jr. Inhibition of fatty acid synthesis in Escherichia coli in the absence of phospholipid synthesis and release of inhibition by thioesterase action. J Bacteriol. 1994;176:2814–21. doi: 10.1128/jb.176.10.2814-2821.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Ramazanov A. Ramazanov. Z. Isolation and characterization of a starchless mutant of Chlorella pyrenoidosa STL-PI with a high growth rate, and high protein and polyunsaturated fatty acid content. Phycol Res 2006. [Google Scholar]
- 69.Hiltunen JK, Qin Y. beta-oxidation - strategies for the metabolism of a wide variety of acyl-CoA esters. Biochim Biophys Acta. 2000;1484:117–28. doi: 10.1016/S1388-1981(00)00013-5. [DOI] [PubMed] [Google Scholar]
- 70.Michinaka Y, Shimauchi T, Aki T, Nakajima T, Kawamoto S, Shigeta S, et al. Extracellular secretion of free fatty acids by disruption of a fatty acyl-CoA synthetase gene in Saccharomyces cerevisiae. J Biosci Bioeng. 2003;95:435–40. doi: 10.1016/s1389-1723(03)80041-5. [DOI] [PubMed] [Google Scholar]
- 71.Nojima Y, Kibayashi A, Matsuzaki H, Hatano T, Fukui S. Isolation and characterization of triacylglycerol-secreting mutant strain from yeast, Saccharomyces cerevisiae. J Gen Appl Microbiol. 1999;45:1–6. doi: 10.2323/jgam.45.1. [DOI] [PubMed] [Google Scholar]
- 72.Scharnewski M, Pongdontri P, Mora G, Hoppert M, Fulda M. Mutants of Saccharomyces cerevisiae deficient in acyl-CoA synthetases secrete fatty acids due to interrupted fatty acid recycling. FEBS J. 2008;275:2765–78. doi: 10.1111/j.1742-4658.2008.06417.x. [DOI] [PubMed] [Google Scholar]
- 73.Germain V, Rylott EL, Larson TR, Sherson SM, Bechtold N, Carde JP, et al. Requirement for 3-ketoacyl-CoA thiolase-2 in peroxisome development, fatty acid beta-oxidation and breakdown of triacylglycerol in lipid bodies of Arabidopsis seedlings. Plant J. 2001;28:1–12. doi: 10.1046/j.1365-313X.2001.01095.x. [DOI] [PubMed] [Google Scholar]
- 74.Cao Z, Gao H, Liu M, Jiao P. Engineering the acetyl-CoA transportation system of candida tropicalis enhances the production of dicarboxylic acid. Biotechnol J. 2006;1:68–74. doi: 10.1002/biot.200500008. [DOI] [PubMed] [Google Scholar]
- 75.Song D, Fu J, Shi D. Exploitation of Oil-bearingMicroalgae for Biodiesel. Chinese Journal of Biotechnology. 2008;24:341–8. doi: 10.1016/s1872-2075(08)60016-3. [DOI] [PubMed] [Google Scholar]
- 76.Sugimoto T, Tanaka K, Monma M, Kawamura Y, Saio K. Phosphoenolpyruvate Carboxylase Level in Soybean Seed Highly Correlates to Its Contents of Protein and Lipid. Agr Biol Chem Tokyo. 1989;53:885–7. doi: 10.1271/bbb1961.53.885. [DOI] [Google Scholar]
- 77.Chen J, Lang C, Hu Z, Liu Z, Huang R. Antisense PEP gene regulates to ratio of protein and lipid content in Brassica napus. J Agric Biotechnol. 1999;7:316–20. [Google Scholar]
- 78.Chisti Y. Biodiesel from microalgae. Biotechnol Adv. 2007;25:294–306. doi: 10.1016/j.biotechadv.2007.02.001. [DOI] [PubMed] [Google Scholar]
- 79.Grotewold E. Transcription factors for predictive plant metabolic engineering: are we there yet? Curr Opin Biotechnol. 2008;19:138–44. doi: 10.1016/j.copbio.2008.02.002. [DOI] [PubMed] [Google Scholar]
- 80.Reik A, Zhou Y, Collingwood TN, Warfe L, Bartsevich V, Kong Y, et al. Enhanced protein production by engineered zinc finger proteins. Biotechnol Bioeng. 2007;97:1180–9. doi: 10.1002/bit.21304. [DOI] [PubMed] [Google Scholar]
- 81.Voelker TA, Worrell AC, Anderson L, Bleibaum J, Fan C, Hawkins DJ, et al. Fatty acid biosynthesis redirected to medium chains in transgenic oilseed plants. Science. 1992;257:72–4. doi: 10.1126/science.1621095. [DOI] [PubMed] [Google Scholar]
- 82.Voelker TA, Hayes TR, Cranmer AM, Turner JC, Davies HM. Genetic engineering of a quantitative trait: metabolic and genetic parameters influencing the accumulation of laurate in rapeseed. Plant J. 2002;9:229–41. doi: 10.1046/j.1365-313X.1996.09020229.x. [DOI] [Google Scholar]
- 83.Neidhardt FC, Ingraham JL, Low KB, Magasanik B, Schaechter M, Umbarger H. Escherichia coli and Salmonella typhimurium. Cellular and Molecular Biology, Vol. 1 and II. American Society for Microbiology, 1987. [Google Scholar]
- 84.Harwood JL. Fatty acid metabolism. Annu Rev Plant Physiol Plant Mol Biol. 1988;39:101–38. doi: 10.1146/annurev.pp.39.060188.000533. [DOI] [Google Scholar]
- 85.Gunstone F, Harwood J, Padley F. The Lipid HandbookChapman and Hall. London, New York 1986. [Google Scholar]
- 86.Yuan L, Voelker TA, Hawkins DJ. Modification of the substrate specificity of an acyl-acyl carrier protein thioesterase by protein engineering. Proc Natl Acad Sci U S A. 1995;92:10639–43. doi: 10.1073/pnas.92.23.10639. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Radakovits R, Eduafo PM, Posewitz MC. Genetic engineering of fatty acid chain length in Phaeodactylum tricornutum. Metab Eng. 2011;13:89–95. doi: 10.1016/j.ymben.2010.10.003. [DOI] [PubMed] [Google Scholar]
- 88.Tsay JT, Oh W, Larson TJ, Jackowski S, Rock CO. Isolation and characterization of the beta-ketoacyl-acyl carrier protein synthase III gene (fabH) from Escherichia coli K-12. J Biol Chem. 1992;267:6807–14. [PubMed] [Google Scholar]
- 89.Molina Grima E, Belarbi EH, Acién Fernández FG, Robles Medina A, Chisti Y. Recovery of microalgal biomass and metabolites: process options and economics. Biotechnol Adv. 2003;20:491–515. doi: 10.1016/S0734-9750(02)00050-2. [DOI] [PubMed] [Google Scholar]
- 90.Manilla-Pérez E, Reers C, Baumgart M, Hetzler S, Reichelt R, Malkus U, et al. Analysis of lipid export in hydrocarbonoclastic bacteria of the genus Alcanivorax: identification of lipid export-negative mutants of Alcanivorax borkumensis SK2 and Alcanivorax jadensis T9. J Bacteriol. 2010;192:643–56. doi: 10.1128/JB.00700-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Singer ME, Tyler SM, Finnerty WR. Growth of Acinetobacter sp. strain HO1-N on n-hexadecanol: physiological and ultrastructural characteristics. J Bacteriol. 1985;162:162–9. doi: 10.1128/jb.162.1.162-169.1985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Goutx M, Acquaviva M, Bertrand JC. Cellular and Extracellular Carbohydrates and Lipids from Marine-Bacteria during Growth on Soluble Substrates and Hydrocarbons. Mar Ecol Prog Ser. 1990;61:291–6. doi: 10.3354/meps061291. [DOI] [Google Scholar]
- 93.Scharnewski M, Pongdontri P, Mora G, Hoppert M, Fulda M. Mutants of Saccharomyces cerevisiae deficient in acyl-CoA synthetases secrete fatty acids due to interrupted fatty acid recycling. FEBS J. 2008;275:2765–78. doi: 10.1111/j.1742-4658.2008.06417.x. [DOI] [PubMed] [Google Scholar]
- 94.Nojima Y, Yagi T, Miyakawa T, Matsuzaki H, Hatano T, Fukui S. Extracellular Formation of Triglycerides from Glucose by a Mutant Strain of Trichosporon. J Ferment Bioeng. 1995;80:88–90. doi: 10.1016/0922-338X(95)98182-K. [DOI] [Google Scholar]
- 95.Stahl A, Gimeno RE, Tartaglia LA, Lodish HF. Fatty acid transport proteins: a current view of a growing family. Trends Endocrinol Metab. 2001;12:266–73. doi: 10.1016/S1043-2760(01)00427-1. [DOI] [PubMed] [Google Scholar]
- 96.Nunn WD, Colburn RW, Black PN. Transport of long-chain fatty acids in Escherichia coli. Evidence for role of fadL gene product as long-chain fatty acid receptor. J Biol Chem. 1986;261:167–71. [PubMed] [Google Scholar]
- 97.Wang X, Karbarz MJ, McGrath SC, Cotter RJ, Raetz CR. MsbA transporter-dependent lipid A 1-dephosphorylation on the periplasmic surface of the inner membrane: topography of francisella novicida LpxE expressed in Escherichia coli. J Biol Chem. 2004;279:49470–8. doi: 10.1074/jbc.M409078200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Liu H, Yu C, Feng DX, Cheng T, Meng X, Liu W, et al. Production of extracellular fatty acid using engineered Escherichia coli. Microb Cell Fact. 2012;•••:11. doi: 10.1186/1475-2859-11-41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Berk PD, Stump DD. Mechanisms of cellular uptake of long chain free fatty acids. Mol Cell Biochem. 1999;192:17–31. doi: 10.1023/A:1006832001033. [DOI] [PubMed] [Google Scholar]
- 100.DiRusso CC, Black PN. Long-chain fatty acid transport in bacteria and yeast. Paradigms for defining the mechanism underlying this protein-mediated process. Mol Cell Biochem. 1999;192:41–52. doi: 10.1023/A:1006823831984. [DOI] [PubMed] [Google Scholar]
- 101.Black PN. Primary sequence of the Escherichia coli fadL gene encoding an outer membrane protein required for long-chain fatty acid transport. J Bacteriol. 1991;173:435–42. doi: 10.1128/jb.173.2.435-442.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Kumar GB, Black PN. Bacterial long-chain fatty acid transport. Identification of amino acid residues within the outer membrane protein FadL required for activity. J Biol Chem. 1993;268:15469–76. [PubMed] [Google Scholar]
- 103.Zhou Z, White KA, Polissi A, Georgopoulos C, Raetz CRH. Function of Escherichia coli MsbA, an essential ABC family transporter, in lipid A and phospholipid biosynthesis. Faseb J. 1998;12:A1284–A. doi: 10.1074/jbc.273.20.12466. [DOI] [PubMed] [Google Scholar]
- 104.Doerrler WT, Reedy MC, Raetz CRH. An Escherichia coli mutant defective in lipid export. J Biol Chem. 2001;276:11461–4. doi: 10.1074/jbc.C100091200. [DOI] [PubMed] [Google Scholar]
- 105.Watkins PA, Lu JF, Steinberg SJ, Gould SJ, Smith KD, Braiterman LT. Disruption of the Saccharomyces cerevisiae FAT1 gene decreases very long-chain fatty acyl-CoA synthetase activity and elevates intracellular very long-chain fatty acid concentrations. J Biol Chem. 1998;273:18210–9. doi: 10.1074/jbc.273.29.18210. [DOI] [PubMed] [Google Scholar]
- 106.Faergeman NJ, DiRusso CC, Elberger A, Knudsen J, Black PN. Disruption of the Saccharomyces cerevisiae homologue to the murine fatty acid transport protein impairs uptake and growth on long-chain fatty acids. J Biol Chem. 1997;272:8531–8. doi: 10.1074/jbc.272.13.8531. [DOI] [PubMed] [Google Scholar]
- 107.Liu X, Brune D, Vermaas W, Curtiss R., 3rd Production and secretion of fatty acids in genetically engineered cyanobacteria. Proc Natl Acad Sci U S A. 2010 doi: 10.1073/pnas.1001946107. [DOI] [PubMed] [Google Scholar]
- 108.Kalscheuer R, Uthoff S, Luftmann H, Steinbuchel A. In vitro and in vivo biosynthesis of wax diesters by an unspecific bifunctional wax ester synthase/acyl-CoA: diacylglycerol acyltransferase from Acinetobacter calcoaceticus ADP1. Eur J Lipid Sci Technol. 2003;105:578–84. doi: 10.1002/ejlt.200300840. [DOI] [Google Scholar]
- 109.Kalscheuer R, Luftmann H, Steinbüchel A. Synthesis of novel lipids in Saccharomyces cerevisiae by heterologous expression of an unspecific bacterial acyltransferase. Appl Environ Microbiol. 2004;70:7119–25. doi: 10.1128/AEM.70.12.7119-7125.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Stöveken T, Kalscheuer R, Malkus U, Reichelt R, Steinbüchel A. The wax ester synthase/acyl coenzyme A:diacylglycerol acyltransferase from Acinetobacter sp. strain ADP1: characterization of a novel type of acyltransferase. J Bacteriol. 2005;187:1369–76. doi: 10.1128/JB.187.4.1369-1376.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Uthoff S, Stöveken T, Weber N, Vosmann K, Klein E, Kalscheuer R, et al. Thio wax ester biosynthesis utilizing the unspecific bifunctional wax ester synthase/acyl coenzyme A:diacylglycerol acyltransferase of Acinetobacter sp. strain ADP1. Appl Environ Microbiol. 2005;71:790–6. doi: 10.1128/AEM.71.2.790-796.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Wältermann M, Stöveken T, Steinbüchel A. Key enzymes for biosynthesis of neutral lipid storage compounds in prokaryotes: properties, function and occurrence of wax ester synthases/acyl-CoA: diacylglycerol acyltransferases. Biochimie. 2007;89:230–42. doi: 10.1016/j.biochi.2006.07.013. [DOI] [PubMed] [Google Scholar]
- 113.Stöveken T, Steinbüchel A. Bacterial acyltransferases as an alternative for lipase-catalyzed acylation for the production of oleochemicals and fuels. Angew Chem Int Ed Engl. 2008;47:3688–94. doi: 10.1002/anie.200705265. [DOI] [PubMed] [Google Scholar]
- 114.Lardizabal KD, Metz JG, Sakamoto T, Hutton WC, Pollard MR, Lassner MW. Purification of a jojoba embryo wax synthase, cloning of its cDNA, and production of high levels of wax in seeds of transgenic arabidopsis. Plant Physiol. 2000;122:645–55. doi: 10.1104/pp.122.3.645. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Cheng JB, Russell DW. Mammalian wax biosynthesis. II. Expression cloning of wax synthase cDNAs encoding a member of the acyltransferase enzyme family. J Biol Chem. 2004;279:37798–807. doi: 10.1074/jbc.M406226200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Kalscheuer R, Stölting T, Steinbüchel A. Microdiesel: Escherichia coli engineered for fuel production. Microbiology. 2006;152:2529–36. doi: 10.1099/mic.0.29028-0. [DOI] [PubMed] [Google Scholar]
- 117.Holtzapple E, Schmidt-Dannert C. Biosynthesis of isoprenoid wax ester in Marinobacter hydrocarbonoclasticus DSM 8798: identification and characterization of isoprenoid coenzyme A synthetase and wax ester synthases. J Bacteriol. 2007;189:3804–12. doi: 10.1128/JB.01932-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Kalscheuer R, Steinbüchel A. A novel bifunctional wax ester synthase/acyl-CoA:diacylglycerol acyltransferase mediates wax ester and triacylglycerol biosynthesis in Acinetobacter calcoaceticus ADP1. J Biol Chem. 2003;278:8075–82. doi: 10.1074/jbc.M210533200. [DOI] [PubMed] [Google Scholar]
- 119.Kalscheuer R, Stöveken T, Luftmann H, Malkus U, Reichelt R, Steinbüchel A. Neutral lipid biosynthesis in engineered Escherichia coli: jojoba oil-like wax esters and fatty acid butyl esters. Appl Environ Microbiol. 2006;72:1373–9. doi: 10.1128/AEM.72.2.1373-1379.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Wang L, Takayama K, Goldman DS, Schnoes HK. Synthesis of alcohol and wax ester by a cell-free system in Mycobacterium tuberculosis. Biochim Biophys Acta. 1972;260:41–8. doi: 10.1016/0005-2760(72)90072-0. [DOI] [PubMed] [Google Scholar]
- 121.Peters D. Carbohydrates for fermentation. Biotechnol J. 2006;1:806–14. doi: 10.1002/biot.200600041. [DOI] [PubMed] [Google Scholar]
- 122.Alvira P, Tomás-Pejó E, Ballesteros M, Negro MJ. Pretreatment technologies for an efficient bioethanol production process based on enzymatic hydrolysis: A review. Bioresour Technol. 2010;101:4851–61. doi: 10.1016/j.biortech.2009.11.093. [DOI] [PubMed] [Google Scholar]
- 123.Den Haan R, Rose SH, Lynd LR, van Zyl WH. Hydrolysis and fermentation of amorphous cellulose by recombinant Saccharomyces cerevisiae. Metab Eng. 2007;9:87–94. doi: 10.1016/j.ymben.2006.08.005. [DOI] [PubMed] [Google Scholar]
- 124.Fujita Y, Takahashi S, Ueda M, Tanaka A, Okada H, Morikawa Y, et al. Direct and efficient production of ethanol from cellulosic material with a yeast strain displaying cellulolytic enzymes. Appl Environ Microbiol. 2002;68:5136–41. doi: 10.1128/AEM.68.10.5136-5141.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Kotaka A, Bando H, Kaya M, Kato-Murai M, Kuroda K, Sahara H, et al. Direct ethanol production from barley beta-glucan by sake yeast displaying Aspergillus oryzae beta-glucosidase and endoglucanase. J Biosci Bioeng. 2008;105:622–7. doi: 10.1263/jbb.105.622. [DOI] [PubMed] [Google Scholar]
- 126.Nigam PS, Singh A. Production of liquid biofuels from renewable resources. Prog Energ Combust. 2011;37:52–68. doi: 10.1016/j.pecs.2010.01.003. [DOI] [Google Scholar]
- 127.Himmel ME, Ding SY, Johnson DK, Adney WS, Nimlos MR, Brady JW, et al. Biomass recalcitrance: engineering plants and enzymes for biofuels production. Science. 2007;315:804–7. doi: 10.1126/science.1137016. [DOI] [PubMed] [Google Scholar]
- 128.Lynd LR, van Zyl WH, McBride JE, Laser M. Consolidated bioprocessing of cellulosic biomass: an update. Curr Opin Biotechnol. 2005;16:577–83. doi: 10.1016/j.copbio.2005.08.009. [DOI] [PubMed] [Google Scholar]
- 129.la Grange DC, den Haan R, van Zyl WH. Engineering cellulolytic ability into bioprocessing organisms. Appl Microbiol Biotechnol. 2010;87:1195–208. doi: 10.1007/s00253-010-2660-x. [DOI] [PubMed] [Google Scholar]
- 130.Zhang YHP, Lynd LR. Toward an aggregated understanding of enzymatic hydrolysis of cellulose: noncomplexed cellulase systems. Biotechnol Bioeng. 2004;88:797–824. doi: 10.1002/bit.20282. [DOI] [PubMed] [Google Scholar]
- 131.Zhou SD, Ingram LO. Simultaneous saccharification and fermentation of amorphous cellulose to ethanol by recombinant Klebsiella oxytoca SZ21 without supplemental cellulase. Biotechnol Lett. 2001;23:1455–62. doi: 10.1023/A:1011623509335. [DOI] [Google Scholar]
- 132.Georgiou G, Segatori L. Preparative expression of secreted proteins in bacteria: status report and future prospects. Curr Opin Biotechnol. 2005;16:538–45. doi: 10.1016/j.copbio.2005.07.008. [DOI] [PubMed] [Google Scholar]
- 133.Bokinsky G, Peralta-Yahya PP, George A, Holmes BM, Steen EJ, Dietrich J, et al. Synthesis of three advanced biofuels from ionic liquid-pretreated switchgrass using engineered Escherichia coli. Proc Natl Acad Sci U S A. 2011;108:19949–54. doi: 10.1073/pnas.1106958108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Kim Y, Ximenes E, Mosier NS, Ladisch MR. Soluble inhibitors/deactivators of cellulase enzymes from lignocellulosic biomass. Enzyme Microb Technol. 2011;48:408–15. doi: 10.1016/j.enzmictec.2011.01.007. [DOI] [PubMed] [Google Scholar]
- 135.Li CL, Knierim B, Manisseri C, Arora R, Scheller HV, Auer M, et al. Comparison of dilute acid and ionic liquid pretreatment of switchgrass: Biomass recalcitrance, delignification and enzymatic saccharification. Bioresour Technol. 2010;101:4900–6. doi: 10.1016/j.biortech.2009.10.066. [DOI] [PubMed] [Google Scholar]
- 136.Choi WJ. Glycerol-based biorefinery for fuels and chemicals. Recent Pat Biotechnol. 2008;2:173–80. doi: 10.2174/187220808786241006. [DOI] [PubMed] [Google Scholar]
- 137.Papanikolaou S, Fakas S, Fick M, Chevalot I, Galiotou-Panayotou M, Komaitis M, et al. Biotechnological valorisation of raw glycerol discharged after bio-diesel (fatty acid methyl esters) manufacturing process: Production of 1,3-propanediol, citric acid and single cell oil. Biomass Bioenergy. 2008;32:60–71. doi: 10.1016/j.biombioe.2007.06.007. [DOI] [Google Scholar]
- 138.Lee PC, Lee WG, Lee SY, Chang HN. Succinic acid production with reduced by-product formation in the fermentation of Anaerobiospirillum succiniciproducens using glycerol as a carbon source. Biotechnol Bioeng. 2001;72:41–8. doi: 10.1002/1097-0290(20010105)72:1<41::AID-BIT6>3.0.CO;2-N. [DOI] [PubMed] [Google Scholar]
- 139.Johnson DT, Taconi KA. The glycerin glut: Options for the value-added conversion of crude glycerol resulting from biodiesel production. Environ Prog. 2007;26:338–48. doi: 10.1002/ep.10225. [DOI] [Google Scholar]
- 140.Murarka A, Dharmadi Y, Yazdani SS, Gonzalez R. Fermentative utilization of glycerol by Escherichia coli and its implications for the production of fuels and chemicals. Appl Environ Microbiol. 2008;74:1124–35. doi: 10.1128/AEM.02192-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Thompson JC, He BB. Characterization of crude glycerol from biodiesel production from multiple feedstocks. Appl Eng Agric. 2006;22:261–5. [Google Scholar]
- 142.Chhetri AB, Watts KC, Islam MR. Waste Cooking Oil as an Alternate Feedstock for Biodiesel Production. Energies. 2008;1:3–18. doi: 10.3390/en1010003. [DOI] [Google Scholar]
- 143.Den Haan R, Rose SH, Lynd LR, van Zyl WH. Hydrolysis and fermentation of amorphous cellulose by recombinant Saccharomyces cerevisiae. Metab Eng. 2007;9:87–94. doi: 10.1016/j.ymben.2006.08.005. [DOI] [PubMed] [Google Scholar]
- 144.Park CS, Chang CC, Ryu DD. Expression and high-level secretion of Trichoderma reesei endoglucanase I in Yarrowia lipolytica. Appl Biochem Biotechnol. 2000;87:1–15. doi: 10.1385/ABAB:87:1:1. [DOI] [PubMed] [Google Scholar]
- 145.Ribeiro O, Wiebe M, Ilmén M, Domingues L, Penttilä M. Expression of Trichoderma reesei cellulases CBHI and EGI in Ashbya gossypii. Appl Microbiol Biotechnol. 2010;87:1437–46. doi: 10.1007/s00253-010-2610-7. [DOI] [PubMed] [Google Scholar]
- 146.Boonvitthya N, Bozonnet S, Burapatana V, O’Donohue MJ, Chulalaksananukul W. Comparison of the Heterologous Expression of Trichoderma reesei Endoglucanase II and Cellobiohydrolase II in the Yeasts Pichia pastoris and Yarrowia lipolytica. Mol Biotechnol. 2012 doi: 10.1007/s12033-012-9557-0. [DOI] [PubMed] [Google Scholar]
- 147.Yanase S, Hasunuma T, Yamada R, Tanaka T, Ogino C, Fukuda H, et al. Direct ethanol production from cellulosic materials at high temperature using the thermotolerant yeast Kluyveromyces marxianus displaying cellulolytic enzymes. Appl Microbiol Biotechnol. 2010;88:381–8. doi: 10.1007/s00253-010-2784-z. [DOI] [PubMed] [Google Scholar]
- 148.Fujita Y, Ito J, Ueda M, Fukuda H, Kondo A. Synergistic saccharification, and direct fermentation to ethanol, of amorphous cellulose by use of an engineered yeast strain codisplaying three types of cellulolytic enzyme. Appl Environ Microbiol. 2004;70:1207–12. doi: 10.1128/AEM.70.2.1207-1212.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Jeon E, Hyeon JE, Suh DJ, Suh YW, Kim SW, Song KH, et al. Production of cellulosic ethanol in Saccharomyces cerevisiae heterologous expressing Clostridium thermocellum endoglucanase and Saccharomycopsis fibuligera beta-glucosidase genes. Mol Cells. 2009;28:369–73. doi: 10.1007/s10059-009-0131-y. [DOI] [PubMed] [Google Scholar]
- 150.Hazlewood GP, Davidson K, Clarke JH, Durrant AJ, Hall J, Gilbert HJ. Endoglucanase E, produced at high level in Escherichia coli as a lacZ’ fusion protein, is part of the Clostridium thermocellum cellulosome. Enzyme Microb Technol. 1990;12:656–62. doi: 10.1016/0141-0229(90)90004-A. [DOI] [PubMed] [Google Scholar]
- 151.Liu JM, Xin XJ, Li CX, Xu JH, Bao J. Cloning of thermostable cellulase genes of Clostridium thermocellum and their secretive expression in Bacillus subtilis. Appl Biochem Biotechnol. 2012;166:652–62. doi: 10.1007/s12010-011-9456-z. [DOI] [PubMed] [Google Scholar]
- 152.Cho JS, Choi YJ, Chung DK. Expression of Clostridium thermocellum endoglucanase gene in Lactobacillus gasseri and Lactobacillus johnsonii and characterization of the genetically modified probiotic lactobacilli. Curr Microbiol. 2000;40:257–63. doi: 10.1007/s002849910051. [DOI] [PubMed] [Google Scholar]
- 153.van Wyk N, den Haan R, van Zyl WH. Heterologous co-production of Thermobifida fusca Cel9A with other cellulases in Saccharomyces cerevisiae. Appl Microbiol Biotechnol. 2010;87:1813–20. doi: 10.1007/s00253-010-2618-z. [DOI] [PubMed] [Google Scholar]
- 154.Generoso WC, Malagó-Jr W, Pereira N, Jr., Henrique-Silva F. Recombinant expression and characterization of an endoglucanase III (cel12a) from Trichoderma harzianum (Hypocreaceae) in the yeast Pichia pastoris. Genet Mol Res. 2012;11:1544–57. doi: 10.4238/2012.May.21.11. [DOI] [PubMed] [Google Scholar]
- 155.Godbole S, Decker SR, Nieves RA, Adney WS, Vinzant TB, Baker JO, et al. Cloning and expression of Trichoderma reesei cellobiohydrolase I in Pichia pastoris. Biotechnol Prog. 1999;15:828–33. doi: 10.1021/bp9901116. [DOI] [PubMed] [Google Scholar]
- 156.Takahashi M, Takahashi H, Nakano Y, Konishi T, Terauchi R, Takeda T. Characterization of a cellobiohydrolase (MoCel6A) produced by Magnaporthe oryzae. Appl Environ Microbiol. 2010;76:6583–90. doi: 10.1128/AEM.00618-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Voutilainen SP, Murray PG, Tuohy MG, Koivula A. Expression of Talaromyces emersonii cellobiohydrolase Cel7A in Saccharomyces cerevisiae and rational mutagenesis to improve its thermostability and activity. Protein Eng Des Sel. 2010;23:69–79. doi: 10.1093/protein/gzp072. [DOI] [PubMed] [Google Scholar]
- 158.Zhang XZ, Zhang Z, Zhu Z, Sathitsuksanoh N, Yang Y, Zhang YH. The noncellulosomal family 48 cellobiohydrolase from Clostridium phytofermentans ISDg: heterologous expression, characterization, and processivity. Appl Microbiol Biotechnol. 2010;86:525–33. doi: 10.1007/s00253-009-2231-1. [DOI] [PubMed] [Google Scholar]
- 159.Nakazawa H, Kawai T, Ida N, Shida Y, Kobayashi Y, Okada H, et al. Construction of a recombinant Trichoderma reesei strain expressing Aspergillus aculeatus β-glucosidase 1 for efficient biomass conversion. Biotechnol Bioeng. 2012;109:92–9. doi: 10.1002/bit.23296. [DOI] [PubMed] [Google Scholar]
- 160.Kitagawa T, Tokuhiro K, Sugiyama H, Kohda K, Isono N, Hisamatsu M, et al. Construction of a beta-glucosidase expression system using the multistress-tolerant yeast Issatchenkia orientalis. Appl Microbiol Biotechnol. 2010;87:1841–53. doi: 10.1007/s00253-010-2629-9. [DOI] [PubMed] [Google Scholar]
- 161.Krogh KB, Harris PV, Olsen CL, Johansen KS, Hojer-Pedersen J, Borjesson J, et al. Characterization and kinetic analysis of a thermostable GH3 beta-glucosidase from Penicillium brasilianum. Appl Microbiol Biotechnol. 2010;86:143–54. doi: 10.1007/s00253-009-2181-7. [DOI] [PubMed] [Google Scholar]
- 162.Jun H, Bing Y, Keying Z, Xuemei D, Daiwen C. Expression of a Trichoderma reesei beta-xylanase gene in Escherichia coli and activity of the enzyme on fiber-bound substrates. Protein Expr Purif. 2009;67:1–6. doi: 10.1016/j.pep.2008.07.015. [DOI] [PubMed] [Google Scholar]
- 163.He J, Yu B, Zhang KY, Ding XM, Chen DW. Expression of endo-1, 4-beta-xylanase from Trichoderma reesei in Pichia pastoris and functional characterization of the produced enzyme. BMC Biotechnol. 2009;•••:9. doi: 10.1186/1472-6750-9-56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.La Grange DC, Claeyssens M, Pretorius IS, Van Zyl WH. Coexpression of the Bacillus pumilus beta-xylosidase (xynB) gene with the Trichoderma reesei beta xylanase 2 (xyn2) gene in the yeast Saccharomyces cerevisiae. Appl Microbiol Biotechnol. 2000;54:195–200. doi: 10.1007/s002530000372. [DOI] [PubMed] [Google Scholar]
- 165.Tanaka H, Nakamura T, Hayashi S, Ohta K. Purification and properties of an extracellular endo-1,4-beta-xylanase from Penicillium citrinum and characterization of the encoding gene. J Biosci Bioeng. 2005;100:623–30. doi: 10.1263/jbb.100.623. [DOI] [PubMed] [Google Scholar]
- 166.Fujii T, Yu G, Matsushika A, Kurita A, Yano S, Murakami K, et al. Ethanol production from xylo-oligosaccharides by xylose-fermenting Saccharomyces cerevisiae expressing β-xylosidase. Biosci Biotechnol Biochem. 2011;75:1140–6. doi: 10.1271/bbb.110043. [DOI] [PubMed] [Google Scholar]
- 167.Roth R, Moodley V, van Zyl P. Heterologous expression and optimized production of an Aspergillus aculeatus endo-1,4-beta-mannanase in Yarrowia lipolytica. Mol Biotechnol. 2009;43:112–20. doi: 10.1007/s12033-009-9187-3. [DOI] [PubMed] [Google Scholar]
- 168.Setati ME, Ademark P, van Zyl WH, Hahn-Hägerdal B, Stålbrand H. Expression of the Aspergillus aculeatus endo-beta-1,4-mannanase encoding gene (man1) in Saccharomyces cerevisiae and characterization of the recombinant enzyme. Protein Expr Purif. 2001;21:105–14. doi: 10.1006/prep.2000.1371. [DOI] [PubMed] [Google Scholar]
- 169.van Zyl PJ, Moodley V, Rose SH, Roth RL, van Zyl WH. Production of the Aspergillus aculeatus endo-1,4-beta-mannanase in A. niger. J Ind Microbiol Biotechnol. 2009;36:611–7. doi: 10.1007/s10295-009-0551-x. [DOI] [PubMed] [Google Scholar]
- 170.Pauly M, Andersen LN, Kauppinen S, Kofod LV, York WS, Albersheim P, et al. A xyloglucan-specific endo-beta-1,4-glucanase from Aspergillus aculeatus: expression cloning in yeast, purification and characterization of the recombinant enzyme. Glycobiology. 1999;9:93–100. doi: 10.1093/glycob/9.1.93. [DOI] [PubMed] [Google Scholar]