

Abstract
Ischemic stroke is a devastating brain injury and an important cause of neurologic disability worldwide and across the lifespan. Despite the physical, social, and economic burdens of this disease there is only a single approved medicine for the treatment of acute stroke, and its use is unfortunately limited to the small fraction of patients presenting within the narrow therapeutic window. Following stroke, there is a period of plasticity involving cell genesis, axon growth, and synaptic modulation that is essential to spontaneous recovery. Treatments focusing on neuroprotection and enhancing recovery have been the focus of intense preclinical studies, but translation of these treatments into clinical use has been disappointing thus far. The important role of epigenetic mechanisms in disease states is becoming increasingly apparent, including in ischemic stroke. These regulators of gene expression are poised to be critical mediators of recovery following stroke. In this review we discuss evidence for the role of epigenetics in neuroplasticity and the implications for stroke recovery.
Keywords: DNA methylation, Histone, MiRNA, Ischemia, Neurogenesis, Axon growth, Synaptogenesis
Introduction
Stroke is one of the leading causes of neurologic morbidity and mortality worldwide. United States data estimate the annual incidence of stroke in adults at nearly 800,000, with a corresponding economic burden surpassing $35 billion (Go et al., 2014). While more common in the elderly, stroke afflicts people across the entire age span including infants and children and thus represents an important cause of neurologic disability in the pediatric population as well. The past two decades have witnessed the advent of dedicated stroke centers, along with the widespread use of thrombolytic treatment. These advances have substantially improved our management of acute stroke, but little success has been realized in developing therapies that provide true neuroprotection and enhanced recovery. Recognizing the burden of stroke-related disability in our population and limitations of our ability to provide hyperacute therapies such as t-PA due to narrow therapeutic time windows, the development of neurorestorative therapies without such restrictive uses is imperative.
Epigenetics refers to changes in gene expression that are not based on mutation of the underlying DNA sequence (Ma et al., 2010). Epigenetic changes are generally considered to be long lasting and heritable through successive cell generations, but recent evidence also suggests the potential for previously unappreciated dynamic changes under certain conditions (Felling et al., 2012). Although the field of epigenetics is now well established, interest in the epigenetic mechanisms involved in stroke pathophysiology has only recently gained traction. Our understanding of the epigenetics of neural plasticity has been substantially informed by the study of learning and memory (Levenson and Sweatt, 2005). Using this knowledge as a basis to better understand the structural and functional changes that occur following stroke could provide innovative approaches to stroke recovery and rehabilitation because motor learning is a critical component of this process (Krakauer, 2006). The primary epigenetic mechanisms often considered involve DNA methylation, histone modifications including methylation and acetylation, and posttranscriptional mechanisms of regulation through small, noncoding RNAs. Recent reviews support the emerging interest in the relevance of this field to stroke pathophysiology, but these largely focus on the injury process (Qureshi and Mehler, 2010a,b, 2011). In this review we discuss the available evidence supporting epigenetic mechanisms of neuroplasticity, with emphasis on implications for stroke recovery. This is an emerging domain with the potential to offer important insight into the biology of regeneration and recovery after stroke.
A critical period of injury-induced plasticity
Stroke recovery is an incredibly complex process and therefore any discussion of underlying mechanisms requires a good framework. Most clinical measures of recovery focus on the ability to accomplish various tasks essential to everyday life. In this sense, recovery can be achieved a number of ways, the most efficient of which is arguably compensatory adaptation, or learning to accomplish the task in a different way. For instance, if I have suffered a left middle cerebral artery stroke and can no longer reach for an object with my right hand, the easiest way to obtain the desired object is to reach instead with my left hand. Much of today's clinical focus concentrates on such means of compensatory adaptation. This does not reflect any degree of real neurologic recovery, and the holy grail of brain recovery research is the true restoration of function to the injured brain. Stroke patients do exhibit a spectrum of true recovery, but this is frequently far too limited. Understanding the mechanisms underlying this spontaneous recovery is an essential prerequisite to augmenting it.
There is tremendous evidence that the majority of spontaneous recovery occurs within a defined period of time after stroke. A large study of the natural history of stroke demonstrated that patients reached their maximal improvement by 3 months regardless of the initial severity of their symptoms (Jorgensen et al., 1999). Additionally, animal models indicate that early initiation of rehabilitative therapies within the first days after stroke leads to better functional outcomes (Krakauer et al., 2012). Despite significant challenges in studying similar effects in human stroke patients, clinical studies have also demonstrated trends toward beneficial effects of early rehabilitation (Cifu and Stewart, 1999). Some have drawn comparisons between this early recovery phase after stroke and the critical periods of plasticity that occur during development (Nahmani and Turrigiano, 2014). This leads to 2 important concepts: 1. Interventions designed to truly target reduced neurologic impairment after stroke need to be implemented within this critical period; and 2. Understanding the molecular and cellular characteristics that define critical period may allow a similar window of opportunity to be recreated long after a stroke occurs.
What characteristics of the early post-stroke time period are so critical to the recovery process? The immediate post-stroke epoch can be conceptualized as a period of enhanced plasticity, in many ways resembling the time of neurodevelopment (Cramer and Chopp, 2000). This enhanced plasticity includes the generation of new cells and blood vessels, sprouting and growth of new axons, and modulation of new and existing synapses (Carmichael, 2006). How the mature brain can suddenly launch into a period of renewed growth and development remains largely mysterious. The possibility that key components in epigenetic regulation stand poised to mediate this process in response to injury is a promising concept. In this review we highlight epigenetic mechanisms that altered in the aftermath of stroke and are known to have important functions in neuroplasticity (Table 1).
Table 1.
Selected epigenetic mechanisms influencing neuroplasticity. The mechanisms listed in this table have plausible roles in stroke recovery based on alterations of the key molecular players following stroke.
| Neurogenesis, gliogenesis, angiogenesis | Axon growth | Synaptic plasticity | |
|---|---|---|---|
| DNA methylation |
|
|
|
| Histone Modification |
|
|
|
| MicroRNA |
|
|
|
Global epigenetic changes following stroke
Before discussing the roles of epigenetics specific to recovery, summarized in Table 1, we would like to introduce the global epigenetic changes induced by stroke and briefly mention the evidence that these may generally be involved in stroke physiology. These include some roles in neuroprotection and preconditioning, two processes that are certainly important to outcomes after stroke although not directly related to repair processes which by definition require injury to have occurred first. While the general mechanisms of epigenetic regulation are beyond the scope of this review, we do provide a brief introduction to each and refer the reader to excellent reviews of these topics for further detailed discussions.
DNA methylation
The methylation of cytosine residues was first observed by Johnson and Coghill (1925) but not implicated in the regulation of gene expression until posited by Holliday and Pugh (1975). The methylation of cytosine-guanine (CpG) dinucleotides by a family of methyltransferase enzymes (DNMTs 1-4) has since been well characterized (Goll and Bestor, 2005). The role of this DNA modification within CpG-rich islands near 5′ promoter sites has long been appreciated as an effective means of gene silencing (Bird, 1986), but more recently scientists have expanded the classical view of DNA methylation. The role of intragenic methylation, which in fact comprises most of the methylated residues under homeostatic conditions, has garnered significant attention (Maunakea et al., 2010). Furthermore CpG dinucleotides may not be the exclusive site of methylation in the mammalian genome as previously thought (Ramsahoye et al., 2000), at least in neurons (Xie et al., 2012; Lister et al., 2013; Guo et al., 2014). Evidence of active demethylation of DNA has also called into question the stability of this epigenetic mark (Ma et al., 2009a; Guo et al., 2011a). These recent advances demonstrate the evolving nature of our understanding of DNA methylation.
Following stroke the level of global DNA methylation increases significantly in the infarcted tissue compared to the contralateral hemisphere (Endres et al., 2000). Interestingly, this occurred without measurable changes in DNMT protein or enzymatic activity. While the authors suggest that this may have been due to the technical limitations of the assays used, it may also reflect the complexity and regional specificity of events occurring throughout the brain following stroke. This is highlighted by the fact that DNMT1 reduction protects against stroke, but complete absence of the enzyme does not (Endres et al., 2001). The roles of DNA methylation following stroke are likely varied, and may play a part in both the injury process as well as recovery.
Chromatin modifications
DNA associates with histone proteins in subunits called nucleosomes that form chromatin. The chromatin organization dictates, in part, the access of the genetic code to a cell's transcriptional machinery. Histones can undergo a number of modifications that can allow or prevent transcription (Kouzarides, 2007). These modifications are catalyzed by a variety of enzymes often with reciprocal functions making the modifications largely reversible and allowing dynamic changes to gene expression in response to the cellular environment. Modulation of these enzymes and thus the underlying chromatin structure has produced interesting results in animal models of stroke, and the role of histone modifications in recovery after stroke has also been reviewed recently (J. Elder et al., 2013).
The polycomb group (PcG) and trithorax group (thxG) are families of proteins with reciprocal capabilities of repressing and activating gene transcription, respectively, by coordinating the posttranslational modification of histones (Schuettengruber et al., 2007). Bmi-1 is a PcG protein important in protecting neurons from oxidative stress (Chatoo et al., 2009). This prompted investigation of its role in ischemic preconditioning, a model of potent neuroprotection from ischemic stroke. In this study, levels of Bmi1 increased in animals subjected to both preconditioning and subsequent injurious ischemia, but not in animals subjected to either stimulus alone (Stapels et al., 2010). In vivo knockdown of Bmi1 or another PcG protein, Scmh1, using siRNA essentially abolished the protection afforded by preconditioning, and overexpression of either protected cells in an in vitro model of oxygen glucose deprivation (Stapels et al., 2010).
Acetylation and deacetylation are histone modifications that are made by histone acetyltransferases (HATs) and histone deacetylases (HDACs), respectively. Ischemia alters the expression of multiple HDAC proteins (Baltan et al., 2011), and these have become popular targets for preclinical neuroprotection studies in stroke. Several studies now have demonstrated protective effects of HDAC inhibition in animal models of stroke with various compounds including valproic acid, trichostatin A, and sodium butyrate (Kim et al., 2007; Wang et al., 2012; George et al., 2013). The action of HDAC enzymes is complex, with some subtypes exhibiting protective effects while others may promote cell death (Langley et al., 2008; Chuang et al., 2009). Furthermore, certain classes of HDAC enzymes are expressed outside of the nucleus where they influence the function of diverse proteins in a nonepigenetic manner (Cho and Cavalli, 2014). While many compounds used are often considered to be general inhibitors of HDACs, important work has demonstrated substantial class-specificity of many compounds (Bradner et al., 2010). This is a critical point of future investigation because some studies suggest that selective inhibition of specific HDAC isoforms, such as HDAC6, can offer neuroprotection without associated cell toxicity (Rivieccio et al., 2009). Further study of individual HDACs will help to refine our understanding of the complex role of these proteins in stroke pathogenesis and subsequent recovery.
MicroRNAs
Small noncoding RNAs are a relatively novel class of epigenetic regulators that exert their influence on the genome in complex and as yet incompletely understood ways. They function largely, although not exclusively, through posttranscriptional repression of gene expression (Hobert, 2008). The miRNAs interact with the Argonaute family of proteins to form the RNA induced silencing complexes (RISCs) which then bind to and silence specific mRNA transcripts (Carthew and Sontheimer, 2009). MiRNAs have emerged as important regulators during CNS development, but they are also modulated following a variety of CNS injuries which has gotten them significant attention as potential therapeutic targets (Bhalala et al., 2013).
Several groups have investigated the expression profiles of miRNAs following stroke in animal models (Dharap et al., 2009; X.S. Liu et al., 2011; Gubern et al., 2013) and humans (Tan et al., 2009). These exhibit variable changes in expression during the acute and recovery phases of stroke. Unfortunately, there is little overlap between these studies in terms of consistency of findings, which muddles the picture of which miRNAs are truly important in the evolution of stroke and recovery. The time periods examined in reference to injury and recovery are different. In fact there is likely significant overlap between these phases, and dissecting which miRNAs are involved in the regulation of each will be important work moving forward.
Determining regional changes in miRNA expression may be important in this effort. MiRNA 181a for instance increases in the ischemic core, but decreases in the penumbra (Ouyang et al., 2012). Inhibition of miRNA-181a through antagomir silencing (complementary miRNAs that bind to target miRNAs) reduces cell death in models of both focal and global ischemia with Grp78 and Bcl-2 being potential target transcripts of importance (Ouyang et al., 2012; Moon et al., 2013). Inhibition of let7f or miRNA-1 appears to provide neuroprotection via IGF1 pathways, substantially reducing infarct volume even when done 4 h after stroke, whereas inhibition of miR-124 does not (Selvamani et al., 2012). Conversely, viral mediated overexpression of miR-124 protects against both stroke in vivo and oxygen-glucose deprivation in vitro (Doeppner et al., 2013; Sun et al., 2013), but some have demonstrated conflicting results showing that miR-124 actually promotes cell death by suppressing the expression of apoptosis inhibitors (X. Liu et al., 2013b; Zhu et al., 2014). Many miRNAs can target multiple different genes which may explain some conflicting findings, and trying to dissect the multiple functions that these regulators can serve in the pathophysiology of stroke will require sophisticated investigation.
Neurogenesis, gliogenesis, and angiogenesis
In multiple species including humans, neurogenesis continues to occur postnatally and throughout adulthood in distinct brain regions, specifically the subgranular zone (SGZ) of the hippocampus and the subventricular zone (SVZ) (Palmer et al., 1995; Eriksson et al., 1998; Gould et al., 1999). These neurogenic regions harbor populations of neural stem and progenitor cells (NSPs), and changes within this niche can modulate the process of neurogenesis. Following ischemic stroke, more cells are produced from these regions and exhibit altered paths of migration toward the region of injury (Felling et al., 2006; Arvidsson et al., 2002; Thored et al., 2006). While some of these cells do appear to incorporate into local circuitry (Hou et al., 2008), the vast majority of the cells die, and the extent to which this process contributes meaningfully to regeneration remains unclear. The concept of a “neurovascular unit” illustrates the coupling of neural cells with blood vessels, and many growth factors that influence neurogenesis also contribute to angiogenesis. Angiogenesis also occurs following stroke and should be considered an important component of the NSP niche (Ohab et al., 2006). Epigenetic changes provide an enticing mechanism by which stroke can influence changes within the NSP niche and thus influence the behavior of these cells.
DNA methylation
One way by which methylation silences gene expression is through recruitment of specific binding proteins to the promoter element. The methyl-CpG binding domain (MBD) family of proteins includes MBD1-4 and Mecp2 (Hendrich and Bird, 1998). Following transient ischemia, many of these proteins exhibit altered expression within subregions of the hippocampus, one of the important niches for postnatal neurogenesis. MBD1 and Mecp2 both remained unchanged initially but were elevated by 24 hours after ischemia, while MBD2 expression increased by 6 h after ischemia. In contrast, MBD3 was significantly reduced as early as 3 h after ischemia (Jung et al., 2002). These proteins can be important regulators in the process of neurogenesis. MBD1 is expressed in neural stem and progenitor cells. Mice deficient in this protein exhibit impaired neurogenesis as assessed by BrdU incorporation, partly explained by decreased survival of newly generated cells (Zhao et al., 2003). FGF-2 is an important growth factor for neurogenesis, and further study of Mbd1 knockdown demonstrated increased expression of FGF-2 along with specific hypomethylation of its promoter (Li et al., 2008).
The interrelationship between the expression of MBD1 and promoter methylation is intriguing and may represent a role for these proteins in either establishing or possibly protecting the methylation state. DNA methylation has classically been considered a very stable means of gene silencing, but recent evidence suggests that methylation states may be more dynamic than previously appreciated. The growth arrest and DNA-damage inducible 45 (Gadd45) family of proteins are important components to a process of active demethylation of cytosine residues (Barreto et al., 2007; Ma et al., 2009a,b). This process appears to be mediated through DNA repair pathways, and Gadd45 may function by coupling the necessary enzymatic machinery (Ma et al., 2009a,b; Schafer, 2013). Importantly, Gadd45-dependent demethylation appears to occur in specific genes, leaving global DNA methylation relatively unchanged (Jin et al., 2008; Engel et al., 2009). Gadd45b is essential to neurogenesis that occurs in the DG following electroconvulsive treatment, possibly via Tet1-mediated demethylation of important growth factors including FGF-1 and BDNF (Ma et al., 2009b; Guo et al., 2011b). Neurogenesis is similarly seen following various animal models of stroke, and ischemia substantially increases the expression of Gadd45 proteins in both the adult and perinatal rat brain after stroke (Chen et al., 1998; Schmidt-Kastner et al., 1998; Charriaut-Marlangue et al., 1999). Fig. 1 provides a mechanistic model by which Gadd45 mediated demethylation could lead to the increased expression specific genes important to neuroplasticity.
Figure 1.
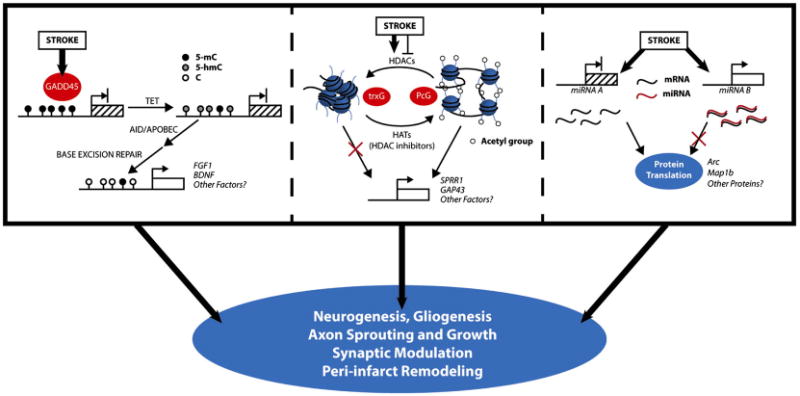
Proposed mechanistic models by which epigenetic changes can influence neuroplasticity following stroke. Active demethylation of DNA leads to increases in gene expression which promotes plasticity. Shifts in histone acetylation may increase or decrease the transcription of specific growth factors. Altered expression of specific non-coding miRNAs can result in increased or decreased translation of proteins. The specific mechanisms reviewed here have strong evidence in neuroplasticity and have been observed to be altered by stroke. The importance in modulating plasticity after stroke requires further specific investigation.
Histone modifications
As discussed previously, members of the polycomb group of proteins can influence the outcome of stroke. Their involvement in neurogenesis in the normal brain is well documented. Bmi-1 knockout reduces the self-renewal of neural stem cells in the developing brain without affecting the fate of lineage-restricted progenitors (Molofsky et al., 2003). Conversely overexpression of Bmi-1 enhances the self-renewal of NSCs in both the developing and adult brain through mechanisms that require the transcription factor Foxg1 (Fasano et al., 2009). These results, coupled with its role in histone modification, provide an appealing mechanism by which Bmi-1 can confer an “epigenetic memory” of differentiation state through subsequent cell divisions (Ringrose and Paro, 2007; Ng and Gurdon, 2008). Given that overexpression of Bmi-1 can substantially reduce infarction in stroke models (Stapels et al., 2010), the role of this protein in regulating neurogenesis following stroke is a worthwhile target of further investigation. EZH2 is a component of the polycomb repressive complex 2 (PRC2) that promotes epigenetic gene silencing by catalyzing histone H3 trimethylation (Cao et al., 2002). Downregulation of EZH2 can promote neuronal differentiation in human mesenchymal stem cells (hMSCs), and transplantation of hMSCs with knocked down EZH2 provides improved functional recovery after stroke compared to transplanted hMSCs with normal EZH2 function (Yu et al., 2013).
Oligodendrogenesis is also altered following stroke, and new evidence supports an important role for histone deacetylase activity in this process. In the early period following stroke, oligodendrocyte progenitors (OPCs) in the peri-infarct white matter demonstrate increased expression of HDAC1 and HDAC2, concurrent with increased proliferation. Mature oligodendrocytes, on the other hand, showed decreased HDAC1 and increased HDAC2 (Kassis et al., 2014). In addition to implying an important role for HDACs in oligodendrogenesis following stroke, this work demonstrates that even individual HDAC isoforms within the same class may have differential effects on cell maturation.
Additional indirect evidence supporting a role for histone modification in post-stroke neurogenesis stems from the effects of HDAC inhibitors on stroke recovery. Valproic acid (VPA) is a potent HDAC inhibitor and improves functional recovery after stroke (Wang et al., 2012). One possible mechanism is through the promotion of angiogenesis, as VPA treatment increased microvessel density and improved cerebral blood flow to the ischemic hemisphere 14 days after stroke (Wang et al., 2012). VPA is also known to promote neuronal differentiation in hippocampal progenitor cells (Hsieh et al., 2004) and thus may also mediate recovery through neurogenesis.
MicroRNAs
The recent explosion of miRNA profiling studies has yielded important results demonstrating the importance of these regulators in neurogenesis. MiR-124 is one molecule that has importance in the acute phase of stroke as discussed above, but also may influence repair after stroke by regulating the behavior of progenitor cells given its role as a neuronal fate determinant in the normal SVZ through targeting of the Notch ligand Jagged1 (JAG1) (Cheng et al., 2009). In the ischemic brain, miR-124a is reduced in the SVZ 7 days after stroke, corresponding to a time of significant neurogenesis (X. S. Liu et al., 2011). Introduction of exogenous miR-124a substantially reduces signaling through the Notch pathway, reduces progenitor proliferation, and induces neuronal differentiation, suggesting that the decreased expression of miR-124a observed after ischemia may be an important mediator of the progenitor cell response (X.S. Liu et al., 2011). MiR-9 is another microRNA transcript of potential importance to post-stroke repair. Loss of this transcript suppresses proliferation in cultured human neural progenitor cells and enhances the migration of these cells when transplanted into the ischemic brain (Delaloy et al., 2010). A potential target of miR-9 important in the ischemic brain is serum response factor, a transcription factor capable of promoting oligodendrocyte differentiation (Buller et al., 2012).
Axon sprouting and growth
The immediate border of an infarct is characterized by the formation of a glial scar in which upregulation of growth inhibitors such as myelin-associated proteins, extracellular matrix proteins, and other growth inhibitors prevent effective axon regeneration (Silver and Miller, 2004). Immediately adjacent to the glial scar however, is a region of periinfarct cortex characterized by the expression of multiple growth-promoting factors that is actually permissive to axon sprouting (Carmichael et al., 2005). Stroke induces extensive changes in the organization of affected networks (Napieralski et al., 1996; Carmichael et al., 2001; Dancause et al., 2005). Axon sprouting both in peri-infarct cortex as well as more remote areas of connected cortex is an important contributor to this remodeling process (Carmichael et al., 2005). Understanding the epigenetic mechanisms regulating the establishment of permissive and inhibitory environments for axon growth will help to further illuminate the means by which network remodeling occurs after stroke, and possible ways to facilitate it.
DNA methylation
Little is known about the importance of DNA methylation in regulating axon sprouting after stroke, but an important role can be inferred from studies in other models of injury. Small proline-rich repeat protein 1 (SPRR1) is expressed following axotomy and can promote axon outgrowth in this scenario (Bonilla et al., 2002). SPRR1 is also expressed at high levels in the peri-infarct cortex early after stroke (Carmichael et al., 2005). Keratinocytes exhibit robust expression of SPRR1 when exposed to the hypomethylating agent 5-azacytidine, suggesting that its expression is influenced by methylation state (J. T. Elder and Zhao, 2002). Whether altered methylation of SPRR1 following stroke is important in its expression still needs to be demonstrated.
Histone Modifications
SPRR1 can be induced by hypomethylating agents as discussed above, but its expression can also be modulated through histone modification. Similar to the effects of 5-azacytidine on keratinocytes, SPRR1 expression is increased in these cells following treatment with the HDAC inhibitor sodium butyrate (J.T. Elder and Zhao, 2002), but this has yet to be examined in the brain. GAP43 is a growth cone-associated protein that promotes neurite outgrowth by regulate cytoskeletal organization via protein kinase C signaling (Benowitz and Routtenberg, 1997). Gap43 expression decreases following neurodevelopment, but is highly induced in peri-infarct cortex after stroke (Skene et al., 1986; Carmichael et al., 2005). VPA is another commonly used drug that can inhibit HDACs, and VPA administration can induce the expression of GAP43 as well as other growth and survival proteins while promoting neurite outgrowth (Yuan et al., 2001). Supporting these findings in an optic nerve crush model of axonal injury, over expression of the HAT, p300, increases GAP43 and SPRR1 expression coincident with increased histone H3 acetylation and promotes axon regeneration (Gaub et al., 2011).
As mentioned in the introduction of this section, the glial scar surrounding and infarct likely limits axonal regeneration due to the presence specific inhibitory factors such as chondroitin sulfate, myelin associated proteins, and proteoglycans. Interestingly, Gaub and colleagues used the HDAC inhibitor trichostatin A to examine effects on neurite outgrowth. They found that HDAC inhibition increased levels of the histone acetyltransferases CREB-binding protein/p300 (CBP/p300) and the p300-CBP-associated factor (P/CAF) as well as hyperacetylation of histone H3. This was accompanied by a reduction of growth cone collapse not only on permissive substrates, but also on inhibitory substrates containing myelin and chondroitin sulfate proteoglycans (Gaub et al., 2010). Recent data also demonstrate the importance of histone 4 acetylation levels in establishing a permissive environment for axon growth. H4 hypoacetylation is associated with diminished axon growth potential, but following a conditioning lesion paradigm of spinal cord injury H4 acetylation levels are restored, triggering the expression of multiple regeneration-associated genes through the activity of a transcriptional complex involving Smad1 (Finelli et al., 2013). It remains to be seen whether similar effects can be demonstrated in the brain after stroke.
MiRNA
Studies have demonstrated an important role for several miRNAs in the processes of axon outgrowth, guidance, and branching (Chiu et al., 2014). In the studies of miRNA expression profiles following ischemia, there are few reports of differential expression of any of these in either the acute or recovery phase of stroke. One exception is miR-9 which is downregulated in ischemic white matter (Buller et al., 2012). MiR-9 is expressed in the axons of primary cortical neurons in the developing brain where it represses microtubule associated protein 1b (Map1b) translation. Inhibition of miR-9 by RNA interference resulted in significantly increased axon length but reduced branching patterns, effects that were dependent on the regulation of Map1b translation (Dajas-Bailador et al., 2012). Another observation of these experiments is that brain derived neurotrophic factor (BDNF), a signaling molecule important in axon development, influences miR-9 in a biphasic manor. Short pulses of BDNF decreased miR-9 expression and resulted in axon growth; whereas, longer pulses of BDNF actually increased the expression of miR-9 and promoted multiple branch points in axons (Dajas-Bailador et al., 2012). Given these modes of action, a role for miR-9 in regulating axon regeneration in the setting of stroke is very plausible but requires specific investigation.
Synaptic plasticity
Synaptic plasticity is central to the establishment and maintenance of neural networks. On one hand, Hebbian plasticity dictates that the strength of a synapse is dependent on its activity, manifested in the processes of long term potentiation and long term depression. Homeostatic mechanisms of plasticity function to maintain average neuronal excitability, thereby balancing activity-dependent synaptic changes that would have otherwise proceeded unchecked and providing stability to the system (Nelson and Turrigiano, 2008). Both of these mechanisms are important considerations in spontaneous recovery from stroke (Murphy and Corbett, 2009). Stroke results in marked functional cortical remapping both in animal models as well as human patients (Chollet et al., 1991; Clarkson et al., 2013). These changes are accompanied by increased synaptic protein expression in peri-infarct tissue as well as more remote contralateral brain several weeks after stroke (Stroemer et al., 1995). Epigenetic regulation is critical to synaptic plasticity and warrants specific investigation in the setting of stroke recovery (Guzman-Karlsson et al., 2014).
DNA methylation
We have previously discussed the role of DNMT activity in protecting against ischemia, but little is known regarding the importance of DNMT's in the recovery phase. There is evidence, however, that DNMTs are important for synaptic plasticity. Inhibition of DNMT activity blocks long term potentiation in the hippocampus and results in decreased methylation of the reelin and BDNF promoters, genes known to be involved in the induction of synaptic plasticity (Levenson and Sweatt, 2005). Furthermore mice deficient in DNMT1 and DNMT3a exhibit impaired learning and memory along with abnormal hippocampal plasticity (Feng et al., 2010). Given this data, it would be interesting to examine the effects of DNMT deficiency on synaptic remodeling after stroke.
Following stroke, neurons in the peri-infarct cortex exhibit intrinsic hyperexcitability (Schiene et al., 1996; Brown et al., 2009). This increased excitability is intriguing because neuronal activity has recently been linked to the dynamic regulation of DNA methylation states (Guo et al., 2011a; Felling et al., 2012). Neuronal activity has been shown to promote the specific demethylation of BDNF through the activity of Gadd45b and TET1 (Ma et al., 2009a,b; Guo et al., 2011b). In Gadd45b knockout mice, changes in dendritic morphology typically induced by electroconvulsive treatment do not occur (Ma et al., 2009a,b). It is possible that these mechanisms of demethylation are important in the recovery of stroke, but this has yet to be studied.
Histone modifications
We have already discussed the potential role for HDAC inhibitors in protecting against stroke, but abundant evidence demonstrates that histone acetylation is important in synaptic plasticity and may therefore be important in the recovery phase as well. Use of HDAC inhibitors enhances LTP and memory formation in the hippocampus (Vecsey et al., 2007). Conversely, another group was able to perform essentially the reverse experiment by genetically reducing CBP, thereby lowering histone acetylation, and demonstrated impaired LTP (Chatterjee et al., 2013). Anatomically, HDAC inhibition significantly increases dendritic spine formation in hippocampal neurons, an effect that was demonstrated to be specifically related to HDAC2 using genetic knockout or overexpression (Guan et al., 2009). Investigating the effects of these types of modulation on synaptic plasticity in relation to stroke recovery will provide important mechanistic insight.
MiRNA
The immediate early gene Arc is important regulator of synaptic plasticity (Shepherd and Bear, 2011). Arc expression is decreased in the ischemic core, but significantly increased in the peri-infarct cortex soon after stroke likely due to glutamate release and neuronal activation (Berger et al., 2003). Evidence now implicates multiple miRNAs in the regulation of Arc, at least one of which is differentially expressed after stroke. MiR-324 was among a group of 16 miRNAs studied that could inhibit the expression of a reporter containing the Arc 3′ untranslated region (Wibrand et al., 2012). A member of the miR-324 family also exhibits decreased expression in the brain during the recovery phase after ischemia (F.J. Liu et al., 2013). This presents a plausible mechanism by which Arc expression could be increased to regulate synaptic plasticity after stroke.
MiR-124 is highly specific to neurons and has been implicated in various aspects of stroke pathophysiology. It is also a key component of a pathway involved in long term potentiation. This pathway involves the exchange protein directly activated by cAMP (EPAC) and Zi268, and disruption of the pathway through knockout of EPAC results in impaired LTP and overt learning deficits that can be reversed with knockdown of miR-124 (Yang et al., 2012). MiR-181 has been shown to influence the degree of infarction, and its expression decreases in the penumbra following ischemia (Ouyang et al., 2012). Other studies have shown it to be an inhibitor of dendritic spine formation in the nucleus accumbens (Saba et al., 2012), therefore, decreased expression may contribute to synaptic plasticity after stroke.
Concluding remarks
Epigenetic mechanisms have established roles in neuroplasticity within the normal brain. These have been demonstrated through studies of neurodevelopment, learning and memory. The role of such mechanisms in neuroplasticity following injury have not been clearly defined. Epigenetic changes have been shown following stroke, but mostly in the context of injury evolution. In this review we have examined the intersection of these evolving fields of investigation, and proposed mechanisms by which epigenetic players may influence post-stroke remodeling and plasticity (Fig. 1). These roles need to be examined independently in the context of the injured brain to provide a more comprehensive understanding of how repair processes are initiated in the acute phase of stroke, and how these contribute to later recovery.
While we have focused on positive aspects of plasticity, an important point to note is that the emerging concept of “maladaptive plasticity” suggests that some aspects may actually inhibit spontaneous recovery or otherwise lead to negative consequences such as epileptogenesis (Takeuchi and Izumi, 2012; Jang, 2013). Additionally while certain types of plasticity are enhanced following stroke, other types such as visual plasticity may actually be impaired (Jablonka et al., 2007; Greifzu et al., 2011, 2014). The role of epigenetic regulation in these facets of stroke recovery must also be explored.
We have only begun to scratch the surface in understanding the complex epigenetic changes in the ischemic brain. In contrast to the injured brain, there is an abundance of literature describing the epigenetic mechanisms responsible for the normal functions of plasticity during neurodevelopment and learning. In this review we have broken down the cellular aspects of stroke recovery into parallel physiological processes and identified potential mechanisms with evidence in both stroke and plasticity. This framework helps to identify plausible candidates important to stroke-induced plasticity, but in many cases the roles have yet to be directly investigated. Furthermore, as discussed in the introduction, stroke is a disease with devastating impact across the lifespan. Aging has profound influence on the epigenome (Issa, 2014), and the mechanisms underlying stroke induced plasticity in the aged brain are unlikely to be the same as those in the young and developing brain, thus necessitating consideration of effects of developmental maturation on recovery from stroke.
Epigenetic mechanisms of regulation are potent mediators of changes in gene expression that can be influenced by both intrinsic and extrinsic factors. This places them in a prime position to facilitate increased plasticity following stroke. As discussed, clinical and scientific observations suggest a critical period of plasticity following stroke during which true neurologic repair is a possibility. Further investigation of histone modifications, DNA methylation and demethylation, and miRNA regulation following stroke will provide important insights into the basic mechanisms of such plasticity and further enhance our understanding of the brain's inherent regenerative capacity. Clinical epigenetics is a rapidly advancing field, spurred largely by advances in cancer medicine. Combining these tools with a sophisticated understanding of the underlying biology of spontaneous neurologic remodeling could dramatically improve our clinical approach to stroke patients.
Acknowledgments
The research in the authors' laboratory was supported by grants from National Institutes of Health (R37NS047344, R21ES021957, and K12NS0001696) and the Simons Foundation (SFARI240011).
References
- Arvidsson A, Collin T, Kirik D, Kokaia Z, Lindvall O. Neuronal replacement from endogenous precursors in the adult brain after stroke. Nat Med. 2002;8(9):963–970. doi: 10.1038/nm747. http://dx.doi.org/10.1038/nm747. [DOI] [PubMed] [Google Scholar]
- Baltan S, Bachleda A, Morrison RS, Murphy SP. Expression of histone deacetylases in cellular compartments of the mouse brain and the effects of ischemia. Transl Stroke Res. 2011;2(3):11–423. doi: 10.1007/s12975-011-0087-z. http://dx.doi.org/10.1007/s12975-011-0087-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barreto G, Schafer A, Marhold J, Stach D, Swaminathan SK, Handa V, Niehrs C. Gadd45a promotes epigenetic gene activation by repair-mediated DNA demethylation. Nature. 2007;445(7128):671–675. doi: 10.1038/nature05515. http://dx.doi.org/10.1038/nature05515. [DOI] [PubMed] [Google Scholar]
- Benowitz LI, Routtenberg A. GAP-43: an intrinsic determinant of neuronal development and plasticity. Trends Neurosci. 1997;20(2):84–91. doi: 10.1016/s0166-2236(96)10072-2. [DOI] [PubMed] [Google Scholar]
- Berger UV, Lu XC, Liu W, Tang Z, Slusher BS, Hediger MA. Effect of middle cerebral artery occlusion on mRNA expression for the sodium-coupled vitamin C transporter SVCT2 in rat brain. J Neurochem. 2003;86(4):896–906. doi: 10.1046/j.1471-4159.2003.01891.x. [DOI] [PubMed] [Google Scholar]
- Bhalala OG, Srikanth M, Kessler JA. The emerging roles of microRNAs in CNS injuries. Nat Rev Neurol. 2013;9(6):328–339. doi: 10.1038/nrneurol.2013.67. http://dx.doi.org/10.1038/nrneurol.2013.67. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bird AP. CpG-rich islands and the function of DNA methylation. Nature. 1986;321(6067):209–213. doi: 10.1038/321209a0. http://dx.doi.org/10.1038/321209a0. [DOI] [PubMed] [Google Scholar]
- Bonilla IE, Tanabe K, Strittmatter SM. Small proline-rich repeat protein 1A is expressed by axotomized neurons and promotes axonal outgrowth. J Neurosci Off J Soc Neurosci. 2002;22(4):1303–1315. doi: 10.1523/JNEUROSCI.22-04-01303.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bradner JE, West N, Grachan ML, Greenberg EF, Haggarty SJ, Warnow T, Mazitschek R. Chemical phylogenetics of histone deacetylases. Nat Chem Biol. 2010;6(3):238–243. doi: 10.1038/nchembio.313. doi:nchembio.313 [pii] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brown CE, Aminoltejari K, Erb H, Winship IR, Murphy TH. In vivo voltage-sensitive dye imaging in adult mice reveals that somatosensory maps lost to stroke are replaced over weeks by new structural and functional circuits with prolonged modes of activation within both the peri-infarct zone and distant sites. J Neurosci Off J Soc Neurosci. 2009;29(6):1719–1734. doi: 10.1523/JNEUROSCI.4249-08.2009. http://dx.doi.org/10.1523/JNEUROSCI.4249-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buller B, Chopp M, Ueno Y, Zhang L, Zhang RL, Morris D, Zhang ZG. Regulation of serum response factor by miRNA-200 and miRNA-9 modulates oligodendrocyte progenitor cell differentiation. Glia. 2012;60(12):1906–1914. doi: 10.1002/glia.22406. http://dx.doi.org/10.1002/glia.22406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cao R, Wang L, Wang H, Xia L, Erdjument-Bromage H, Tempst P, Zhang Y. Science. 5595. Vol. 298. New York, N.Y.: 2002. Role of histone H3 lysine 27 methylation in polycomb-group silencing; pp. 1039–1043. http://dx.doi.org/10.1126/science.1076997. [DOI] [PubMed] [Google Scholar]
- Carmichael ST. Cellular and molecular mechanisms of neural repair after stroke: making waves. Ann Neurol. 2006;59(5):735–742. doi: 10.1002/ana.20845. http://dx.doi.org/10.1002/ana.20845. [DOI] [PubMed] [Google Scholar]
- Carmichael ST, Wei L, Rovainen CM, Woolsey TA. New patterns of intracortical projections after focal cortical stroke. Neurobiol Dis. 2001;8(5):910–922. doi: 10.1006/nbdi.2001.0425. http://dx.doi.org/10.1006/nbdi.2001.0425. [DOI] [PubMed] [Google Scholar]
- Carmichael ST, Archibeque I, Luke L, Nolan T, Momiy J, Li S. Growth-associated gene expression after stroke: Evidence for a growth-promoting region in peri-infarct cortex. Exp Neurol. 2005;193(2):291–311. doi: 10.1016/j.expneurol.2005.01.004. http://dx.doi.org/10.1016/j.expneurol.2005.01.004. [DOI] [PubMed] [Google Scholar]
- Carthew RW, Sontheimer EJ. Origins and mechanisms of miRNAs and siRNAs. Cell. 2009;136(4):642–655. doi: 10.1016/j.cell.2009.01.035. http://dx.doi.org/10.1016/j.cell.2009.01.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Charriaut-Marlangue C, Richard E, Ben-Ari Y. DNA damage and DNA damage-inducible protein Gadd45 following ischemia in the P7 neonatal rat. Brain Res Dev Brain Res. 1999;116(2):133–140. doi: 10.1016/s0165-3806(99)00084-x. [DOI] [PubMed] [Google Scholar]
- Chatoo W, Abdouh M, David J, Champagne MP, Ferreira J, Rodier F, Bernier G. The polycomb group gene Bmi1 regulates antioxidant defenses in neurons by repressing p53 pro-oxidant activity. J Neurosci Off J Soc Neurosci. 2009;29(2):529–542. doi: 10.1523/JNEUROSCI.5303-08.2009. http://dx.doi.org/10.1523/JNEUROSCI.5303-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chatterjee S, Mizar P, Cassel R, Neidl R, Selvi BR, Mohankrishna DV, Boutillier AL. A novel activator of CBP/p300 acetyltransferases promotes neurogenesis and extends memory duration in adult mice. J Neurosci Off J Soc Neurosci. 2013;33(26):10698–10712. doi: 10.1523/JNEUROSCI.5772-12.2013. http://dx.doi.org/10.1523/JNEUROSCI.5772-12.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen J, Uchimura K, Stetler RA, Zhu RL, Nakayama M, Jin K, Simon RP. Transient global ischemia triggers expression of the DNA damage-inducible gene GADD45 in the rat brain. J Cereb Blood Flow Metab Off J Int Soc Cereb Blood Flow Metab. 1998;18(6):646–657. doi: 10.1097/00004647-199806000-00007. http://dx.doi.org/10.1097/00004647-199806000-00007. [DOI] [PubMed] [Google Scholar]
- Cheng LC, Pastrana E, Tavazoie M, Doetsch F. miR-124 regulates adult neurogenesis in the subventricular zone stem cell niche. Nat Neurosci. 2009;12(4):399–408. doi: 10.1038/nn.2294. http://dx.doi.org/10.1038/nn.2294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chiu H, Alqadah A, Chang C. The role of microRNAs in regulating neuronal connectivity. Front Cell Neurosci. 2014;7:283. doi: 10.3389/fncel.2013.00283. http://dx.doi.org/10.3389/fncel.2013.00283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cho Y, Cavalli V. HDAC signaling in neuronal development and axon regeneration. Curr Opin Neurobiol. 2014;27C:118–126. doi: 10.1016/j.conb.2014.03.008. doi:S0959-4388(14)00058-0 [pii] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chollet F, DiPiero V, Wise RJ, Brooks DJ, Dolan RJ, Frackowiak RS. The functional anatomy of motor recovery after stroke in humans: A study with positron emission tomography. Ann Neurol. 1991;29(1):63–71. doi: 10.1002/ana.410290112. http://dx.doi.org/10.1002/ana.410290112. [DOI] [PubMed] [Google Scholar]
- Chuang DM, Leng Y, Marinova Z, Kim HJ, Chiu CT. Multiple roles of HDAC inhibition in neurodegenerative conditions. Trends Neurosci. 2009;32(11):591–601. doi: 10.1016/j.tins.2009.06.002. http://dx.doi.org/10.1016/j.tins.2009.06.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cifu DX, Stewart DG. Factors affecting functional outcome after stroke: A critical review of rehabilitation interventions. Arch Phys Med Rehabil. 1999;80(5 Suppl. 1):S35–S39. doi: 10.1016/s0003-9993(99)90101-6. doi:S000399939900163X [pii] [DOI] [PubMed] [Google Scholar]
- Clarkson AN, Lopez-Valdes HE, Overman JJ, Charles AC, Brennan KC, Thomas Carmichael S. Multimodal examination of structural and functional remapping in the mouse photothrombotic stroke model. J Cereb Blood Flow Metab Off J Int Soc Cereb Blood Flow Metab. 2013;33(5):716–723. doi: 10.1038/jcbfm.2013.7. http://dx.doi.org/10.1038/jcbfm.2013.7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cramer SC, Chopp M. Recovery recapitulates ontogeny. Trends Neurosci. 2000;23(6):265–271. doi: 10.1016/s0166-2236(00)01562-9. [DOI] [PubMed] [Google Scholar]
- Dajas-Bailador F, Bonev B, Garcez P, Stanley P, Guillemot F, Papalopulu N. microRNA-9 regulates axon extension and branching by targeting Map1b in mouse cortical neurons. Nat Neurosci. 2012 doi: 10.1038/nn.3082. http://dx.doi.org/10.1038/nn.3082. [DOI] [PubMed]
- Dancause N, Barbay S, Frost SB, Plautz EJ, Chen D, Zoubina EV, Nudo RJ. Extensive cortical rewiring after brain injury. J Neurosci Off J Soc Neurosci. 2005;25(44):10167–10179. doi: 10.1523/JNEUROSCI.3256-05.2005. http://dx.doi.org/10.1523/JNEUROSCI.3256-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Delaloy C, Liu L, Lee JA, Su H, Shen F, Yang GY, Gao FB. MicroRNA-9 coordinates proliferation and migration of human embryonic stem cell-derived neural progenitors. Cell Stem Cell. 2010;6(4):323–335. doi: 10.1016/j.stem.2010.02.015. http://dx.doi.org/10.1016/j.stem.2010.02.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dharap A, Bowen K, Place R, Li LC, Vemuganti R. Transient focal ischemia induces extensive temporal changes in rat cerebral microRNAome. J Cereb Blood Flow Metab Off J Int Soc Cereb Blood Flow Metab. 2009;29(4):675–687. doi: 10.1038/jcbfm.2008.157. http://dx.doi.org/10.1038/jcbfm.2008.157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Doeppner TR, Doehring M, Bretschneider E, Zechariah A, Kaltwasser B, Muller B, Michel U. MicroRNA-124 protects against focal cerebral ischemia via mechanisms involving Usp14-dependent REST degradation. Acta Neuropathol. 2013;126(2):251–265. doi: 10.1007/s00401-013-1142-5. http://dx.doi.org/10.1007/s00401-013-1142-5. [DOI] [PubMed] [Google Scholar]
- Elder JT, Zhao X. Evidence for local control of gene expression in the epidermal differentiation complex. Exp Dermatol. 2002;11(5):406–412. doi: 10.1034/j.1600-0625.2002.110503.x. [DOI] [PubMed] [Google Scholar]
- Elder J, Cortes M, Rykman A, Hill J, Karuppagounder S, Edwards D, Ratan RR. The epigenetics of stroke recovery and rehabilitation: from polycomb to histone deacetylases. Neurother J Am Soc Exp Neurothe. 2013;10(4):808–816. doi: 10.1007/s13311-013-0224-3. http://dx.doi.org/10.1007/s13311-013-0224-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Endres M, Meisel A, Biniszkiewicz D, Namura S, Prass K, Ruscher K, Dirnagl U. DNA methyltransferase contributes to delayed ischemic brain injury. J Neurosci Off J Soc Neurosci. 2000;20(9):3175–3256. doi: 10.1523/JNEUROSCI.20-09-03175.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Endres M, Fan G, Meisel A, Dirnagl U, Jaenisch R. Effects of cerebral ischemia in mice lacking DNA methyltransferase 1 in post-mitotic neurons. Neuroreport. 2001;12(17):3763–3766. doi: 10.1097/00001756-200112040-00032. [DOI] [PubMed] [Google Scholar]
- Engel N, Tront JS, Erinle T, Nguyen N, Latham KE, Sapienza C, Liebermann DA. Conserved DNA methylation in Gadd45a(-/-) mice. Epigenet Off J DNA Methylat Soc. 2009;4(2):98–99. doi: 10.4161/epi.4.2.7858. doi:7858 [pii] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eriksson PS, Perfilieva E, Bjork-Eriksson T, Alborn AM, Nordborg C, Peterson DA, Gage FH. Neurogenesis in the adult human hippocampus. Nat Med. 1998;4(11):1313–1317. doi: 10.1038/3305. http://dx.doi.org/10.1038/3305. [DOI] [PubMed] [Google Scholar]
- Fasano CA, Phoenix TN, Kokovay E, Lowry N, Elkabetz Y, Dimos JT, Temple S. Bmi-1 cooperates with Foxg1 to maintain neural stem cell self-renewal in the forebrain. Genes Dev. 2009;23(5):561–574. doi: 10.1101/gad.1743709. http://dx.doi.org/10.1101/gad.1743709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Felling RJ, Snyder MJ, Romanko MJ, Rothstein RP, Ziegler AN, Yang Z, Givogri MI, Bongarzone ER, Levison SW. Neural stem/progenitor cells participate in the regenerative response to perinatal hypoxia/ischemia. J Neurosci. 2006;26(16):4359–4369. doi: 10.1523/JNEUROSCI.1898-05.2006. http://dx.doi.org/10.1523/jneurosci.1898-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Felling RJ, Guo JU, Song H. Neuronal activation and insight into the plasticity of DNA methylation. Epigenomics. 2012;4(2):125–127. doi: 10.2217/epi.12.2. http://dx.doi.org/10.2217/epi.12.2. [DOI] [PubMed] [Google Scholar]
- Finelli MJ, Wong JK, Zou H. Epigenetic regulation of sensory axon regeneration after spinal cord injury. J Neurosci Off J Soc Neurosci. 2013;33(50):19664–19676. doi: 10.1523/JNEUROSCI.0589-13.2013. http://dx.doi.org/10.1523/JNEUROSCI.0589-13.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gaub P, Tedeschi A, Puttagunta R, Nguyen T, Schmandke A, Di Giovanni S. HDAC inhibition promotes neuronal outgrowth and counteracts growth cone collapse through CBP/p300 and P/CAF-dependent p53 acetylation. Cell Death Differ. 2010;17:1392–1408. doi: 10.1038/cdd.2009.216. http://dx.doi.org/10.1038/cdd.2009.216. [DOI] [PubMed] [Google Scholar]
- Gaub P, Joshi Y, Wuttke A, Naumann U, Schnichels S, Heiduschka P, Di Giovanni S. The histone acetyltransferase p300 promotes intrinsic axonal regeneration. Brain. 2011;134(Pt 7):2134–2148. doi: 10.1093/brain/awr142. http://dx.doi.org/10.1093/brain/awr142. [DOI] [PubMed] [Google Scholar]
- George S, Kadam SD, Irving ND, Markowitz GJ, Raja S, Kwan A, Comi AM. Impact of trichostatin A and sodium valproate treatment on post-stroke neurogenesis and behavioral outcomes in immature mice. Front Cell Neurosci. 2013;7:123. doi: 10.3389/fncel.2013.00123. http://dx.doi.org/10.3389/fncel.2013.00123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Go AS, Mozaffarian D, Roger VL, Benjamin EJ, Berry JD, Blaha MJ American Heart Association Statistics Committee and Stroke Statistics Subcommittee. Heart disease and stroke statistics—2014 update: A report from the american heart association. Circulation. 2014;129(3):e28–e292. doi: 10.1161/01.cir.0000441139.02102.80. http://dx.doi.org/10.1161/01.cir.0000441139.02102.80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goll MG, Bestor TH. Eukaryotic cytosine methyltransferases. Annu Rev Biochem. 2005;74:481–514. doi: 10.1146/annurev.biochem.74.010904.153721. http://dx.doi.org/10.1146/annurev.biochem.74.010904.153721. [DOI] [PubMed] [Google Scholar]
- Gould E, Reeves AJ, Graziano MS, Gross CG. Science. 5439. Vol. 286. New York, N.Y.: 1999. Neurogenesis in the neocortex of adult primates; pp. 548–552. [DOI] [PubMed] [Google Scholar]
- Greifzu F, Schmidt S, Schmidt KF, Kreikemeier K, Witte OW, Lowel S. Global impairment and therapeutic restoration of visual plasticity mechanisms after a localized cortical stroke. Proc Natl Acad Sci U S A. 2011;108(37):15450–15455. doi: 10.1073/pnas.1016458108. http://dx.doi.org/10.1073/pnas.1016458108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Greifzu F, Pielecka-Fortuna J, Kalogeraki E, Krempler K, Favaro PD, Schluter OM, Lowel S. Environmental enrichment extends ocular dominance plasticity into adulthood and protects from stroke-induced impairments of plasticity. Proc Natl Acad Sci U S A. 2014;111(3):1150–1155. doi: 10.1073/pnas.1313385111. http://dx.doi.org/10.1073/pnas.1313385111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guan JS, Haggarty SJ, Giacometti E, Dannenberg JH, Joseph N, Gao J, Tsai LH. HDAC2 negatively regulates memory formation and synaptic plasticity. Nature. 2009;459(7243):55–60. doi: 10.1038/nature07925. http://dx.doi.org/10.1038/nature07925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gubern C, Camos S, Ballesteros I, Rodriguez R, Romera VG, Canadas R, Castellanos M. miRNA expression is modulated over time after focal ischaemia: Up-regulation of miR-347 promotes neuronal apoptosis. FEBS J. 2013;280(23):6233–6246. doi: 10.1111/febs.12546. http://dx.doi.org/10.1111/febs.12546. [DOI] [PubMed] [Google Scholar]
- Guo JU, Su Y, Zhong C, Ming GL, Song H. Hydroxylation of 5-methylcytosine by TET1 promotes active DNA demethylation in the adult brain. Cell. 2011a;145(3):423–434. doi: 10.1016/j.cell.2011.03.022. http://dx.doi.org/10.1016/j.cell.2011.03.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guo JU, Su Y, Zhong C, Ming GL, Song H. Cell Cycle. 16. Vol. 10. Georgetown, Tex.: 2011b. Emerging roles of TET proteins and 5-hydroxymethylcytosines in active DNA demethylation and beyond; pp. 2662–2668. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guo JU, Su Y, Shin JH, Shin J, Li H, Xie B, Song H. Distribution, recognition and regulation of non-CpG methylation in the adult mammalian brain. Nat Neurosci. 2014;17(2):215–222. doi: 10.1038/nn.3607. http://dx.doi.org/10.1038/nn.3607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guzman-Karlsson MC, Meadows JP, Gavin CF, Hablitz JJ, Sweatt JD. Transcriptional and epigenetic regulation of hebbian and non-hebbian plasticity. Neuropharmacology. 2014 doi: 10.1016/j.neuropharm.2014.01.001. http://dx.doi.org/10.1016/j.neuropharm.2014.01.001. [DOI] [PMC free article] [PubMed]
- Hendrich B, Bird A. Identification and characterization of a family of mammalian methyl-CpG binding proteins. Mol Cell Biol. 1998;18(11):6538–6547. doi: 10.1128/mcb.18.11.6538. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hobert O. Science. 5871. Vol. 319. New York, N.Y.: 2008. Gene regulation by transcription factors and microRNAs; pp. 1785–1786. http://dx.doi.org/10.1126/science.1151651. [DOI] [PubMed] [Google Scholar]
- Holliday R, Pugh JE. Science. 4173. Vol. 187. New York, N.Y.: 1975. DNA modification mechanisms and gene activity during development; pp. 226–232. [PubMed] [Google Scholar]
- Hou SW, Wang YQ, Xu M, Shen DH, Wang JJ, Huang F, Sun FY. Functional integration of newly generated neurons into striatum after cerebral ischemia in the adult rat brain. Stroke J Cereb Circ. 2008;39(10):2837–2844. doi: 10.1161/STROKEAHA.107.510982. http://dx.doi.org/10.1161/STROKEAHA.107.510982. [DOI] [PubMed] [Google Scholar]
- Hsieh J, Nakashima K, Kuwabara T, Mejia E, Gage FH. Histone deacetylase inhibition-mediated neuronal differentiation of multipotent adult neural progenitor cells. Proc Natl Acad Sci U S A. 2004;101(47):16659–16664. doi: 10.1073/pnas.0407643101. http://dx.doi.org/10.1073/pnas.0407643101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Issa JP. Aging and epigenetic drift: a vicious cycle. J Clin Invest. 2014;124(1):24–29. doi: 10.1172/JCI69735. http://dx.doi.org/10.1172/JCI69735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jablonka JA, Witte OW, Kossut M. Photothrombotic infarct impairs experience-dependent plasticity in neighboring cortex. Neuroreport. 2007;18(2):165–169. doi: 10.1097/WNR.0b013e328010feff. http://dx.doi.org/10.1097/WNR.0b013e328010feff. [DOI] [PubMed] [Google Scholar]
- Jang SH. Motor function-related maladaptive plasticity in stroke: a review. NeuroRehabilitation. 2013;32(2):311–316. doi: 10.3233/NRE-130849. http://dx.doi.org/10.3233/NRE-130849. [DOI] [PubMed] [Google Scholar]
- Jin SG, Guo C, Pfeifer GP. GADD45A does not promote DNA demethylation. PLoS Genet. 2008;4(3):e1000013. doi: 10.1371/journal.pgen.1000013. http://dx.doi.org/10.1371/journal.pgen.1000013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnson TB, Coghill RD. Researches on pyrimidines. C111. the discovery of 5-methyl-cytosine in tuberculinic acid, the nucleic acid of the tubercle bacillus. J Am Chem Soc. 1925;47(11):2838–2844. [Google Scholar]
- Jorgensen HS, Nakayama H, Raaschou HO, Olsen TS. Stroke. neurologic and functional recovery the Copenhagen stroke study. Phys Med Rehabil Clin N Am. 1999;10(4):887–906. [PubMed] [Google Scholar]
- Jung BP, Zhang G, Ho W, Francis J, Eubanks JH. Transient forebrain ischemia alters the mRNA expression of methyl DNA-binding factors in the adult rat hippocampus. Neuroscience. 2002;115(2):515–524. doi: 10.1016/s0306-4522(02)00383-4. [DOI] [PubMed] [Google Scholar]
- Kassis H, Chopp M, Liu XS, Shehadah A, Roberts C, Zhang ZG. Histone deacetylase expression in white matter oligodendrocytes after stroke. Neurochem Int. 2014 doi: 10.1016/j.neuint.2014.03.006. http://dx.doi.org/10.1016/j.neuint.2014.03.006. [DOI] [PMC free article] [PubMed]
- Kim HJ, Rowe M, Ren M, Hong JS, Chen PS, Chuang DM. Histone deacetylase inhibitors exhibit anti-inflammatory and neuroprotective effects in a rat permanent ischemic model of stroke: multiple mechanisms of action. J Pharmacol Exp Ther. 2007;321(3):892–901. doi: 10.1124/jpet.107.120188. http://dx.doi.org/10.1124/jpet.107.120188. [DOI] [PubMed] [Google Scholar]
- Kouzarides T. SnapShot: histone-modifying enzymes. Cell. 2007;131(4):822. doi: 10.1016/j.cell.2007.11.005. http://dx.doi.org/10.1016/j.cell.2007.11.005. [DOI] [PubMed] [Google Scholar]
- Krakauer JW. Motor learning: its relevance to stroke recovery and neurorehabilitation. Curr Opin Neurol. 2006;19(1):84–90. doi: 10.1097/01.wco.0000200544.29915.cc. [DOI] [PubMed] [Google Scholar]
- Krakauer JW, Carmichael ST, Corbett D, Wittenberg GF. Getting neurorehabilitation right: what can be learned from animal models? Neurorehabil Neural Repair. 2012;26(8):923–931. doi: 10.1177/1545968312440745. http://dx.doi.org/10.1177/1545968312440745. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Langley B, D'Annibale MA, Suh K, Ayoub I, Tolhurst A, Bastan B, Ratan RR. Pulse inhibition of histone deacetylases induces complete resistance to oxidative death in cortical neurons without toxicity and reveals a role for cytoplasmic p21(waf1/cip1) in cell cycle-independent neuroprotection. J Neurosci Off J Soc Neurosci. 2008;28(1):163–176. doi: 10.1523/JNEUROSCI.3200-07.2008. http://dx.doi.org/10.1523/JNEUROSCI.3200-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Levenson JM, Sweatt JD. Epigenetic mechanisms in memory formation. Nat Rev Neurosci. 2005;6(2):108–118. doi: 10.1038/nrn1604. http://dx.doi.org/10.1038/nrn1604. [DOI] [PubMed] [Google Scholar]
- Li X, Barkho BZ, Luo Y, Smrt RD, Santistevan NJ, Liu C, Zhao X. Epigenetic regulation of the stem cell mitogen fgf-2 by Mbd1 in adult neural stem/progenitor cells. J Biol Chem. 2008;283(41):27644–27652. doi: 10.1074/jbc.M804899200. http://dx.doi.org/10.1074/jbc.M804899200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lister R, Mukamel EA, Nery JR, Urich M, Puddifoot CA, Johnson ND, Ecker JR. Science. 6146. Vol. 341. New York, N.Y.: 2013. Global epigenomic reconfiguration during mammalian brain development; p. 1237905. http://dx.doi.org/10.1126/science.1237905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu XS, Chopp M, Zhang RL, Tao T, Wang XL, Kassis H, Zhang ZG. MicroRNA profiling in subventricular zone after stroke: MiR-124a regulates proliferation of neural progenitor cells through notch signaling pathway. PloS One. 2011;6(8):e23461. doi: 10.1371/journal.pone.0023461. http://dx.doi.org/10.1371/journal.pone.0023461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu FJ, Lim KY, Kaur P, Sepramaniam S, Armugam A, Wong PT, Jeyaseelan K. microRNAs involved in regulating spontaneous recovery in embolic stroke model. PloS One. 2013a;8(6):e66393. doi: 10.1371/journal.pone.0066393. http://dx.doi.org/10.1371/journal.pone.0066393. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu X, Li F, Zhao S, Luo Y, Kang J, Zhao H, Ji X. MicroRNA-124-mediated regulation of inhibitory member of apoptosis-stimulating protein of p53 family in experimental stroke. Stroke J Cereb Circ. 2013b;44(7):1973–1980. doi: 10.1161/STROKEAHA.111.000613. http://dx.doi.org/10.1161/STROKEAHA.111.000613. [DOI] [PubMed] [Google Scholar]
- Ma DK, Guo JU, Ming GL, Song H. Cell Cycle. 10. Vol. 8. Georgetown, Tex.: 2009a. DNA excision repair proteins and Gadd45 as molecular players for active DNA demethylation; pp. 1526–1531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ma DK, Jang MH, Guo JU, Kitabatake Y, Chang ML, Pow-Anpongkul N, Song H. Science. 5917. Vol. 323. New York, N.Y.: 2009b. Neuronal activity-induced Gadd45b promotes epigenetic DNA demethylation and adult neurogenesis; pp. 1074–1077. http://dx.doi.org/10.1126/science.1166859. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ma DK, Marchetto MC, Guo JU, Ming GL, Gage FH, Song H. Epigenetic choreographers of neurogenesis in the adult mammalian brain. Nat Neurosci. 2010;13(11):1338–1344. doi: 10.1038/nn.2672. http://dx.doi.org/10.1038/nn.2672;10.1038/nn.2672. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maunakea AK, Nagarajan RP, Bilenky M, Ballinger TJ, D'Souza C, Fouse SD, Costello JF. Conserved role of intragenic DNA methylation in regulating alternative promoters. Nature. 2010;466(7303):253–257. doi: 10.1038/nature09165. http://dx.doi.org/10.1038/nature09165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Molofsky AV, Pardal R, Iwashita T, Park IK, Clarke MF, Morrison SJ. Bmi-1 dependence distinguishes neural stem cell self-renewal from progenitor proliferation. Nature. 2003;425(6961):962–967. doi: 10.1038/nature02060. http://dx.doi.org/10.1038/nature02060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moon JM, Xu L, Giffard RG. Inhibition of microRNA-181 reduces forebrain ischemia-induced neuronal loss. J Cereb Blood Flow Metab Off J Int Soc Cereb Blood Flow Metab. 2013;33(12):1976–1982. doi: 10.1038/jcbfm.2013.157. http://dx.doi.org/10.1038/jcbfm.2013.157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murphy TH, Corbett D. Plasticity during stroke recovery: From synapse to behaviour. Nat Rev Neurosci. 2009;10(12):861–872. doi: 10.1038/nrn2735. http://dx.doi.org/10.1038/nrn2735. [DOI] [PubMed] [Google Scholar]
- Nahmani M, Turrigiano GG. Adult cortical plasticity following injury: Recapitulation of critical period mechanisms? Neuroscience. 2014 doi: 10.1016/j.neuroscience.2014.04.029. http://dx.doi.org/10.1016/j.neuroscience.2014.04.029. in press. [DOI] [PMC free article] [PubMed]
- Napieralski JA, Butler AK, Chesselet MF. Anatomical and functional evidence for lesion-specific sprouting of corticostriatal input in the adult rat. J Comp Neurol. 1996;373(4):484–497. doi: 10.1002/(SICI)1096-9861(19960930)373:4<484::AID-CNE2>3.0.CO;2-Y. doi:2-Y. [DOI] [PubMed] [Google Scholar]
- Nelson SB, Turrigiano GG. Strength through diversity. Neuron. 2008;60(3):477–482. doi: 10.1016/j.neuron.2008.10.020. http://dx.doi.org/10.1016/j.neuron.2008.10.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ng RK, Gurdon JB. Cell Cycle. 9. Vol. 7. Georgetown, Tex.: 2008. Epigenetic inheritance of cell differentiation status; pp. 1173–1177. [DOI] [PubMed] [Google Scholar]
- Ohab JJ, Fleming S, Blesch A, Carmichael ST. A neurovascular niche for neurogenesis after stroke. J Neurosci Off J Soc Neurosci. 2006;26(50):13007–13016. doi: 10.1523/JNEUROSCI.4323-06.2006. http://dx.doi.org/10.1523/JNEUROSCI.4323-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ouyang YB, Lu Y, Yue S, Xu LJ, Xiong XX, White RE, Giffard RG. miR-181 regulates GRP78 and influences outcome from cerebral ischemia in vitro and in vivo. Neurobiol Dis. 2012;45(1):555–563. doi: 10.1016/j.nbd.2011.09.012. http://dx.doi.org/10.1016/j.nbd.2011.09.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Palmer T, Ray J, Gage FH. FGF-2-responsive neuronal progenitors reside in proliferative and quiescent regions of the adult rodent brain. Molecular and Cellular Neuroscience; FGF-2-Responsive Neuronal Progenitors Reside in Proliferative and Quiescent Regions of the Adult Rodent Brain. 1995 doi: 10.1006/mcne.1995.1035. http://dx.doi.org/10.1006/mcne.1995.1035. [DOI] [PubMed]
- Qureshi IA, Mehler MF. The emerging role of epigenetics in stroke: II. RNA regulatory circuitry. Arch Neurol. 2010a;67(12):1435–1441. doi: 10.1001/archneurol.2010.300. http://dx.doi.org/10.1001/archneurol.2010.300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qureshi IA, Mehler MF. Emerging role of epigenetics in stroke: Part 1: DNA methylation and chromatin modifications. Arch Neurol. 2010b;67(11):1316–1322. doi: 10.1001/archneurol.2010.275. http://dx.doi.org/10.1001/archneurol.2010.275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qureshi IA, Mehler MF. The emerging role of epigenetics in stroke: III. Neural stem cell biology and regenerative medicine. Arch Neurol. 2011;68(3):294–302. doi: 10.1001/archneurol.2011.6. http://dx.doi.org/10.1001/archneurol.2011.6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ramsahoye BH, Biniszkiewicz D, Lyko F, Clark V, Bird AP, Jaenisch R. Non-CpG methylation is prevalent in embryonic stem cells and may be mediated by DNA methyltransferase 3a. Proc Natl Acad Sci U S A. 2000;97(10):5237–5242. doi: 10.1073/pnas.97.10.5237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ringrose L, Paro R. Dev. 2. Vol. 134. Cambridge; England: 2007. Polycomb/trithorax response elements and epigenetic memory of cell identity; pp. 223–232. http://dx.doi.org/10.1242/dev.02723. [DOI] [PubMed] [Google Scholar]
- Rivieccio MA, Brochier C, Willis DE, Walker BA, D'Annibale MA, McLaughlin K, Langley B. HDAC6 is a target for protection and regeneration following injury in the nervous system. Proc Natl Acad Sci U S A. 2009;106(46):19599–19604. doi: 10.1073/pnas.0907935106. http://dx.doi.org/10.1073/pnas.0907935106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saba R, Storchel PH, Aksoy-Aksel A, Kepura F, Lippi G, Plant TD, Schratt GM. Dopamine-regulated microRNA MiR-181a controls GluA2 surface expression in hippocampal neurons. Mol Cell Biol. 2012;32(3):619–632. doi: 10.1128/MCB.05896-11. http://dx.doi.org/10.1128/MCB.05896-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schafer A. Gadd45 proteins: Key players of repair-mediated DNA demethylation. Adv Exp Med Biol. 2013;793:35–50. doi: 10.1007/978-1-4614-8289-5_3. http://dx.doi.org/10.1007/978-1-4614-8289-5_3. [DOI] [PubMed] [Google Scholar]
- Schiene K, Bruehl C, Zilles K, Qu M, Hagemann G, Kraemer M, Witte OW. Neuronal hyperexcitability and reduction of GABAA-receptor expression in the surround of cerebral photothrombosis. J Cereb Blood Flow Metab Off J Int Soc Cereb Blood Flow Metab. 1996;16(5):906–914. doi: 10.1097/00004647-199609000-00014. http://dx.doi.org/10.1097/00004647-199609000-00014. [DOI] [PubMed] [Google Scholar]
- Schmidt-Kastner R, Zhao W, Truettner J, Belayev L, Busto R, Ginsberg MD. Pixel-based image analysis of HSP70, GADD45 and MAP2 mRNA expression after focal cerebral ischemia: hemodynamic and histological correlates. Brain Res Mol Brain Res. 1998;63(1):79–97. doi: 10.1016/s0169-328x(98)00263-0. [DOI] [PubMed] [Google Scholar]
- Schuettengruber B, Chourrout D, Vervoort M, Leblanc B, Cavalli G. Genome regulation by polycomb and trithorax proteins. Cell. 2007;128(4):735–745. doi: 10.1016/j.cell.2007.02.009. http://dx.doi.org/10.1016/j.cell.2007.02.009. [DOI] [PubMed] [Google Scholar]
- Selvamani A, Sathyan P, Miranda RC, Sohrabji F. An antagomir to microRNA Let7f promotes neuroprotection in an ischemic stroke model. PloS One. 2012;7(2):e32662. doi: 10.1371/journal.pone.0032662. http://dx.doi.org/10.1371/journal.pone.0032662. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shepherd JD, Bear MF. New views of arc, a master regulator of synaptic plasticity. Nat Neurosci. 2011;14(3):279–284. doi: 10.1038/nn.2708. http://dx.doi.org/10.1038/nn.2708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Silver J, Miller JH. Regeneration beyond the glial scar. Nat Rev Neurosci. 2004;5(2):146–156. doi: 10.1038/nrn1326. http://dx.doi.org/10.1038/nrn1326. [DOI] [PubMed] [Google Scholar]
- Skene JH, Jacobson RD, Snipes GJ, McGuire CB, Norden JJ, Freeman JA. Science. 4765. Vol. 233. New York, N.Y.: 1986. A protein induced during nerve growth (GAP-43) is a major component of growth-cone membranes; pp. 783–786. [DOI] [PubMed] [Google Scholar]
- Stapels M, Piper C, Yang T, Li M, Stowell C, Xiong ZG, Zhou A. Polycomb group proteins as epigenetic mediators of neuroprotection in ischemic tolerance. Sci Signal. 2010;3(111):ra15. doi: 10.1126/scisignal.2000502. http://dx.doi.org/10.1126/scisignal.2000502. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stroemer RP, Kent TA, Hulsebosch CE. Neocortical neural sprouting, synaptogenesis, and behavioral recovery after neocortical infarction in rats. Stroke J Cereb Circ. 1995;26(11):2135–2144. doi: 10.1161/01.str.26.11.2135. [DOI] [PubMed] [Google Scholar]
- Sun Y, Gui H, Li Q, Luo ZM, Zheng MJ, Duan JL, Liu X. MicroRNA-124 protects neurons against apoptosis in cerebral ischemic stroke. CNS Neurosci Ther. 2013;19(10):813–819. doi: 10.1111/cns.12142. http://dx.doi.org/10.1111/cns.12142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takeuchi N, Izumi S. Maladaptive plasticity for motor recovery after stroke: Mechanisms and approaches. Neural Plast. 2012:359728. doi: 10.1155/2012/359728. http://dx.doi.org/10.1155/2012/359728. [DOI] [PMC free article] [PubMed]
- Tan KS, Armugam A, Sepramaniam S, Lim KY, Setyowati KD, Wang CW, Jeyaseelan K. Expression profile of MicroRNAs in young stroke patients. PloS One. 2009;4(11):e7689. doi: 10.1371/journal.pone.0007689. http://dx.doi.org/10.1371/journal.pone.0007689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thored P, Arvidsson A, Cacci E, Ahlenius H, Kallur T, Darsalia V, Lindvall O. Stem Cells. 3. Vol. 24. Dayton, Ohio: 2006. Persistent production of neurons from adult brain stem cells during recovery after stroke; pp. 739–747. http://dx.doi.org/10.1634/stemcells.2005-0281. [DOI] [PubMed] [Google Scholar]
- Vecsey CG, Hawk JD, Lattal KM, Stein JM, Fabian SA, Attner MA, Wood MA. Histone deacetylase inhibitors enhance memory and synaptic plasticity via CREB:CBP-dependent transcriptional activation. J Neurosci Off J Soc Neurosci. 2007;27(23):6128–6140. doi: 10.1523/JNEUROSCI.0296-07.2007. http://dx.doi.org/10.1523/JNEUROSCI.0296-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang B, Zhu X, Kim Y, Li J, Huang S, Saleem S, Cao W. Histone deacetylase inhibition activates transcription factor Nrf2 and protects against cerebral ischemic damage. Free Radic Biol Med. 2012;52(5):928–936. doi: 10.1016/j.freeradbiomed.2011.12.006. http://dx.doi.org/10.1016/j.freeradbiomed.2011.12.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wibrand K, Pai B, Siripornmongcolchai T, Bittins M, Berentsen B, Ofte ML, Bramham CR. MicroRNA regulation of the synaptic plasticity-related gene arc. PloS One. 2012;7(7):e41688. doi: 10.1371/journal.pone.0041688. http://dx.doi.org/10.1371/journal.pone.0041688. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xie W, Barr CL, Kim A, Yue F, Lee AY, Eubanks J, Ren B. Base-resolution analyses of sequence and parent-of-origin dependent DNA methylation in the mouse genome. Cell. 2012;148(4):816–831. doi: 10.1016/j.cell.2011.12.035. http://dx.doi.org/10.1016/j.cell.2011.12.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang Y, Shu X, Liu D, Shang Y, Wu Y, Pei L, Lu Y. EPAC null mutation impairs learning and social interactions via aberrant regulation of miR-124 and Zif268 translation. Neuron. 2012;73(4):774–788. doi: 10.1016/j.neuron.2012.02.003. http://dx.doi.org/10.1016/j.neuron.2012.02.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu YL, Chou RH, Shyu WC, Hsieh SC, Wu CS, Chiang SY, Hung MC. Smurf2-mediated degradation of EZH2 enhances neuron differentiation and improves functional recovery after ischaemic stroke. EMBO Mol Med. 2013;5(4):531–547. doi: 10.1002/emmm.201201783. http://dx.doi.org/10.1002/emmm.201201783. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yuan PX, Huang LD, Jiang YM, Gutkind JS, Manji HK, Chen G. The mood stabilizer valproic acid activates mitogen-activated protein kinases and promotes neurite growth. J Biol Chem. 2001;276(34):31674–31683. doi: 10.1074/jbc.M104309200. http://dx.doi.org/10.1074/jbc.M104309200. [DOI] [PubMed] [Google Scholar]
- Zhao X, Ueba T, Christie BR, Barkho B, McConnell MJ, Nakashima K, Gage FH. Mice lacking methyl-CpG binding protein 1 have deficits in adult neurogenesis and hippocampal function. Proc Natl Acad Sci U S A. 2003;100(11):6777–6782. doi: 10.1073/pnas.1131928100. http://dx.doi.org/10.1073/pnas.1131928100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhu F, Liu JL, Li JP, Xiao F, Zhang ZX, Zhang L. J Mol Neurosci. 1. Vol. 52. MN: 2014. MicroRNA-124 (miR-124) regulates Ku70 expression and is correlated with neuronal death induced by ischemia/reperfusion; pp. 148–155. http://dx.doi.org/10.1007/s12031-013-0155-9. [DOI] [PubMed] [Google Scholar]