

Abstract
It is quite important to understand the overall metabolic regulation mechanism of bacterial cells such as Escherichia coli from both science (such as biochemistry) and engineering (such as metabolic engineering) points of view. Here, an attempt was made to clarify the overall metabolic regulation mechanism by focusing on the roles of global regulators which detect the culture or growth condition and manipulate a set of metabolic pathways by modulating the related gene expressions. For this, it was considered how the cell responds to a variety of culture environments such as carbon (catabolite regulation), nitrogen, and phosphate limitations, as well as the effects of oxygen level, pH (acid shock), temperature (heat shock), and nutrient starvation.
1. Introduction
Although living organisms may have been created somehow with the formation of a compartmentalized autocatalytic cycles with the appearance of ribonucleic acid-based or protein-based enzymes gaining complexity, evolved by adapting to the environment on earth, and improved in their effectiveness and robustness [1], it is still not certain that evolution can solve all the mystery of highly efficient, robust, and well-organized cell systems. It might be true that evolution has played some important roles for the improvement of cell's function and robustness to the changes in the environment, but this may not be all that can explain the cell's complexity with efficient function. In the living organisms, metabolic network, defined as the set and topology of metabolic biochemical reactions within a cell, plays an essential role for the cell to survive, where it is under organized control. In living organisms or cells, thousands of different biochemical reactions as well as transport processes are linked together to break down organic compounds to generate energy and to synthesize macromolecular compounds for cell synthesis. Note that the set of enzyme reactions is not static as illustrated in the biochemistry text book, but the set itself changes dynamically in response to the changes in the growth environment and the cell's state. Similarly, complex signaling networks interconvert signals or stimuli that are important for cellular function and interactions with the environment. This implies the transfer of information in signal transduction pathways and cascades designed to maximize efficiency and cellular responses. It may be of importance to understand the evolution of metabolism and signalings for understanding the adaptation processes of cellular life and the emergence of higher levels of organization [1]. However, here the time scale evolution will not be included unless otherwise stated, and a rather in-depth review is made on the metabolic regulation of a bacterial cell system.
The living organism must survive in response to the variety of environmental perturbations by maintaining the cell system by sensing external and/or internal state [2]. The main part of these functional responses concern the metabolic regulation. The enzymes which form the metabolic pathways are subject to multiple levels of regulation, where the transcriptional regulation may play the important role for metabolic regulation [3]. Although its relevance might have been overemphasized [4], it is important to understand the regulatory processes that govern the cellular metabolism. This is the key focus in the recent microbial systems biology, where genome-scale models for metabolic stoichiometry becomes popular [5, 6], together with the detailed topology of transcriptional regulatory networks that describe all known interactions between transcriptional factors and their target genes [3]. It should be noted that among possible topological network, only a subset is active at any given point in time and condition [7–10]. Although the transcriptional regulation is typically analyzed by measurements of mRNA abundance, the intracellular reaction rates or metabolic fluxes must be quantified in particular by 13C-based metabolic flux analysis to assess metabolic function in the network context [11–17]. It has been demonstrated the strong condition dependence of metabolic control with typically sparse networks of active transcriptional control that affect the flux distribution between different pathways in yeast [18].
Van Rijsewijk et al. [12] considered 81 transcription factors, including all known to directly or indirectly control central metabolic enzymes and 10 sigma- and anti-sigma factors in E. coli. Of the 81 transcription factors, 41 have one or more direct target gene that is involved in the central carbon metabolism [12]. In order to clarify the overall picture of the regulation, it is important to integrate different levels of omics data such as transcriptomics, proteomics, metabolomics, and fluxes, where advanced high throughput data are useful. Perhaps the most comprehensive data set containing those for E. coli grown at different growth rates in glucose-limited continuous cultures and upon deletion of 24 glycolysis and pentose phosphate pathway genes [19] may provide an unprecedented opportunity for computational analyses to extract biological insights [20]. The viability of living organism depends on the correct regulation of gene expressions, so that the appropriate proteins and enzymes must be produced in response to the environmental perturbation. Network analysis has been paid recent attention for investigating the transcriptional regulatory systems [21]. Babu et al. [22] considered to relate network topology to the systems' function in directing gene expression changes. In order to provide a comprehensive view of transcription factors (TFs) and their regulatory functions, RegulonDB is an invaluable resource in analyzing E. coli metabolic regulation [23]. Living organisms sense the changes in the environmental condition by detecting extracellular signals such as the concentrations of nutrients such as carbon, nitrogen, and phosphate sources, and the growth condition such as pH level, oxygen availability. These signals eventually feed into the transcriptional regulatory systems, which affect the physiological and morphological changes that enable organisms to adapt effectively for survival [24]. It has been highlighted that three quarters of E. coli TFs respond directly to external stimuli [25]. The 120 E. coli TFs may be classified into three categories depending on the original input signals such as external (58 TFs), internal (29 TFs), and hybrid (33 TFs).
The typical TFs in E. coli contains “two-headed molecules” which constitutes of a DNA-binding domain and an allosteric site to which metabolites bind noncovalently or to which enzymes covalently modify in order to modulate the regulatory activity of TF [24]. As will be mentioned later, two-component signal transduction system is considered to be the important means of detecting extracellular signals and transducing the signals into cytosol for metabolic regulation. These typically involve a phospho-relay from a transmembrane histidine protein kinase sensor to a target TF response regulator (Figure 1(a)). In the case of E. coli, 29 TFs show such regulation with 28 histidine protein kinase [26], where the genes encoding the two components are usually located within the same operon, enabling their coordinated expression, while some of the kinases and regulators are not adjacent on the chromosome, and it is not straightforward to link the partner in such a case [24]. Note that there might exist a cross-talk between noncognate sensors and regulators [27], which complicates the analysis of metabolic regulation.
Figure 1.

Transcription factors and their function.
As typically seen in Lac repressor, another method for sensing exogenous signals is by TF binding of transported small molecules, and the TFs regulate the enzymatic pathways that process these molecules [24]. In addition to exogenous signals, cell can recognize the cell's state by detecting the intracellular metabolites synthesized internally by cytosolic enzymes. The typical example is Cra (FruR) which binds a key intermediate such as F1, 6BP (FDP) and regulates the carbon flow (Figure 1(b)) as will be explained later in more detail. There exist, yet hybrid type of TFs, where they sense the metabolites that are transported from the culture environment or synthesized endogenously (Figure 1(b)). This can be typically seen in regulating amino acid synthetic pathways, possibly because it is preferable for the cell to import essential metabolites where they are freely available rather than expend energy on their production [24].
The central metabolic pathways of a cell are controlled by a number of global regulators or transcription factors, depending on the culture condition as illustrated by Figure 2. Biological systems are known to be robust and adaptable to the culture environment. It became apparent that such robustness is inherent in the biochemical and genetic networks. Several genes that are necessary to respond to various environmental or nutritional changes require specific recognition by RNA polymerase associated with the alternative sigma factors. Here, we consider how the culture environment affects the global regulators as transcription factors, and how the metabolic pathway genes are regulated by the corresponding global regulators.
Figure 2.

Overall metabolic regulation scheme.
2. Variety of Regulation Mechanisms
Living organisms such as bacterial cells have complex but efficient mechanisms to respond to the change in culture environment. This is achieved by the so-called global regulators, where they generally act at transcriptional level. The global transcriptional regulators are themselves regulated by posttranscriptional regulators. Thus, global regulation forms a cascade of regulations [28]. In relation to global regulators, sigma factors play also important roles, where they allow RNA polymerase to be recruited at specific DNA sequences in the promoter regions at which they initiate transcription. In E. coli, seven sigma factors have been found so far, and those play roles depending on the environmental stimuli (Table 1). In Bacillus subtilis, it has been known that multiple sigma factors control sporulation [29].
Table 1.
Sigma factors and their functions.
| σ 19 | Ion transport |
| σ 24 | Extreme temperature |
| σ 28 | Flagella genes |
| σ 32 | Heat shock |
| σ 38 | Stationary phase or carbon starvation, and so forth |
| σ 54 | Nitrogen regulation |
| σ 70 | House keeping |
H-NS (histone-like nucleotide structuring protein) is another type of global transcriptional regulator, found in enterobacteria. It is a small DNA-associated protein that binds preferentially to a curved AT-rich DNA without showing sequence preferences [28]. H-NS regulates a variety of physiological pathways such as metabolism, fimbriae expression, virulence flagella synthesis, and proper function [28].
Other types of global regulators are signaling molecules such as cyclic-AMP (cAMP) and cyclic-di-GMP [30, 31]. As will be explained later in this paper, cAMP is synthesized from ATP by Cya (adenylate cyclase) at low glucose concentration with an increase in phosphorylated EIIAGlc (EIIAGlc-P) involved in PTS, where EIIAGlc-P activates Cya activity. Note that cAMP binds to Crp (cAMP receptor protein), also known as CAP (catabolite activation protein), and cAMP-Crp complex becomes an activated transcription factor in relation to catabolite regulation. Note also that cAMP regulates not only catabolite regulation, but also flagellum synthesis, biofilm formation, quorum sensing, and nitrogen regulation [31–34].
As another level of regulation, small noncoding RNAs (sRNAs) play important roles in the posttranscriptional regulation [35]. The sRNAs are mainly involved in stress response regulation, pathogenesis, and virulence. A single sRNA can affect multiple targets, where sRNAs modify the translation or stability of the targets and chaperone. One such example is SgrS in E. coli, where it binds to the mRNA of ptsG gene, which encodes EIIBCGlc for glucose uptake [36]. SgrS encodes a small protein SgrT, where SgrT is also involved in the inhibition of glucose uptake, and thus regulate ptsG activity [37]. Another sRNAs regulate other regulators, where such example is DsrA sRNA in E. coli, which regulates σ 38 expression [38]. Another group of sRNAs bind to proteins, where such example is CsrB in E. coli [39], and these sRNAs regulate the activity of CsrA global regulator as will be explained later for carbon storage regulation.
Another level of posttranscriptional regulation is the control of protein stability and folding carried out by ATP-dependent proteases and chaperones [28]. Such examples are the E. coli Lon ATP-dependent proteases that regulate flagella expression by degrading σ 38 as well as acid shock tolerance regulon by regulating the amount of GadE, where gadE is under σ 38 transcriptional control as will be explained later in this paper. As will be also explained, posttranslational control is mediated by the C1PXP ATP-dependent protease, which degrades σ 38.
3. Porin Proteins in the Outer Membrane and Their Regulation
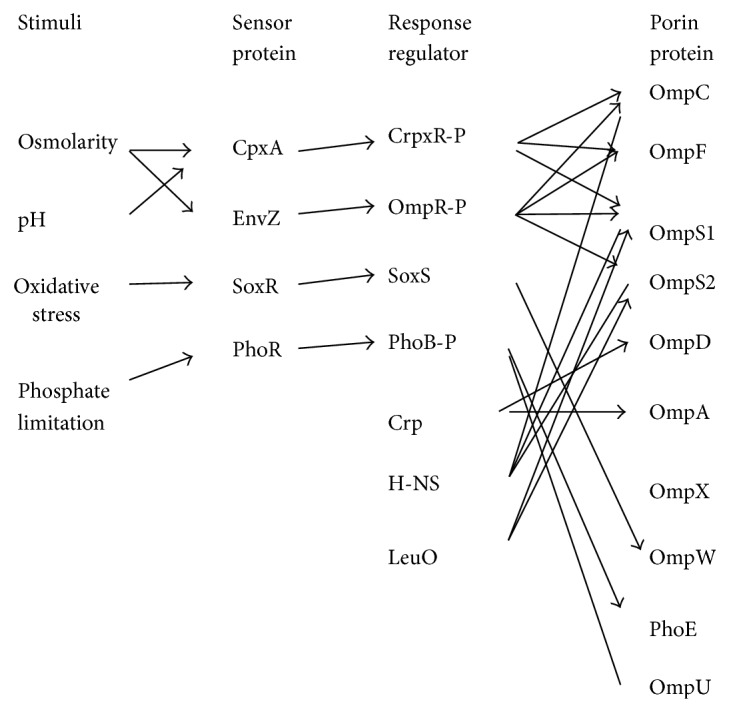
The gram-negative bacteria such as E. coli have two concentric membranes surrounding the cytoplasm, where the space between these two membranes is called periplasm. The outer membrane and cytoplasmic membrane constitute a hydrophobic barrier against polar compounds. The outer membrane contains channel proteins, where the specific molecules can only move across these channels. In the outer membrane of E. coli, 108 channels are formed by the porin proteins [40]. Porins are the outer membrane proteins that produce large, open but regulated water-filled pores that form substrate-specific, ion-selective, or nonspecific channels that allow the influx of small hydrophilic nutrient molecules and the efflux of waste products [41]. They also exclude many antibiotics and inhibitors that are large and lipophilic [42]. Porins including OmpC and OmpF of E. coli form stable trimers with a slight preference for cations over anions [43, 44]. The OmpC and OmpF are the most abundant porins present under typical growth condition representing up to 2% of the total cellular protein [45]. OmpF seems to have slightly larger channel than OmpC. OmpC and OmpF are the constitutive porins. Their relative abundance changes depending on such factors as osmolarity, temperature, and growth phase [46–48]. These porins serve for glucose to enter into the periplasm when glucose is present at a higher concentration than about 0.2 mM [49, 50]. It has been shown that the diffusion rate for glucose is found to be about twofold higher through OmpF than through OmpC [51]. Under glucose limitation, the outer membrane glycoporin LamB is induced [50], where this protein permeates several carbohydrates such as maltose, maltodextrins, and glucose [52]. It has been reported that about 70% of the total glucose import capacity of the cell is contributed by LamB [50]. Glucose transport by diffusion through porins of the outer membrane is a passive process [53].
The porin genes are under control of two-component system such as EnvZ-OmpR system, where EnvZ is an inner membrane sensor kinase and OmpR is the cytoplasmic response regulator (Figure 3). In response to the environmental signals such as osmolarity, pH, temperature, nutrients, and toxins, EnvZ phosphorylates OmpR to form phosphorylated OmpR (OmpR-P), where OmpR-P increases its binding affinity for the promoter regions of porin genes such as ompC and ompF [41]. Note that acetyl phosphate can function as a phosphate donor for OmpR under certain condition. Note also that OmpR controls cellular processes such as chemotaxis and virulence as well [54]. In terms of virulence, abolition of porin production diminishes pathogenesis [41]. There are several other porins than OmpC and OmpF, such as OmpU and OmpT (V. cholerae), OmpH and OmpL (Photobacterium), OmpD (S. typhimurium), OmpS1 and OmpS2 (S. enterica), and OmpW (S. enterica, E. coli, and V. cholerae). Porin genes are also under control of other regulators other than EnvZ-OmpR such as CpxR (under extracytoplasmic stress) [54], PhoB (under phosphate limitation) [55], Lrp (under starvation) [56] Rob (for cationic peptides), MarA (under weak acids), SoxS (under oxidative stress) [57], CadC (at low pH) [58], Crp (under catabolite repression), Fnr (for anaerobiosis) [59], ToxR (for virulence) [60], H-NS, StpA, Ihf, Hu (for nucleotide proteins) [61], and LeuO (for stringent response) [41, 62].
Figure 3.
Outer membrane porin proteins and their regulations.
CpxA and CpxR form a two-component system, where CpxA is the transmembrane sensor kinase, while CpxR is the response regulator. CpxA can be induced by a variety of stimuli such as higher pH (alkali), misfolded proteins and alterations in the membrane composition [63, 64]. Upon the activation of the kinase activity of CpxA, the phosphorylated CpxR (CpxR-P) plays roles as a transcriptional regulator and controls the expressions of ompC and ompF genes, and so forth [41].
PhoR and PhoB also form a two-component system, where phoR is the sensor kinase and detect a low concentration of phosphate or phosphate starvation and activate and phosphorylate PhoB [65] as will be explained later for phosphate regulation. The phosphorylated PhoB (PhoB-P) activates the transcription of phoE gene, where PhoE porin is induced under phosphate limitation [41]. Moreover, PhoB negatively regulates the OmpT, OmpU, and OmpA porins in Vibrio cholera [55].
Lrp is a global regulator which regulates mainly amino acid metabolism. Lrp activity is stimulated in minimal medium (which means low nutrient availability), while it is repressed in rich medium such as LB medium [66]. In minimal medium, Lrp negatively regulates ompC gene, while it positively regulates ompF gene.
MarA, SoxS, and Rob are the members of the AraC/XylS family of transcriptional regulators [67]. These three regulators diminish ompF expression [57]. SoxR and SoxS form a two-component system, where SoxR is a cytoplasmic sensor protein activated by oxidative stress and activates the SoxS regulator as will be explained later for oxidative stress regulation. MarA responds to weak acids like salicylic acid, and so forth [68] and certain antibiotics [69]. Rob may be a general regulator and might be stimulated by cationic peptides [70].
CadC is an inner membrane transcriptional activator that acts both as a signal sensor and as a transcriptional regulator, where it positively regulates the production of OmpC and OmpF at low pH [71, 72].
Crp plays an essential role for catabolite regulation as will be explained later, where Crp regulates ompR-envZ operon by binding directly to the promoter region [73]. The ompA gene in E. coli is positively regulated by Crp [74], while ompX is negatively regulated by Crp by means of CyaR, a small RNA (sRNA) [75]. In Typhimurium, ompD porin gene is activated by cAMP-Crp [59].
Fnr is a DNA-binding protein that senses oxygen level and regulates the metabolism under anaerobic condition together with ArcA/B regulator system as will be explained later for the metabolic regulation under oxygen-limited condition. Fnr positively regulates ompD gene expression under anaerobiosis by the posttranscriptional regulation [59].
ToxR is a transmembrane DNA-binding protein, and it is an important regulator of virulence gene expression in V. cholera. ToxR positively regulates ompT porin gene expression. Note that increased osmolarity enhances OmpT production and diminishes OmpU production, which is similar to that of OmpR on ompC and ompF in E. coli. Moreover, TorX represses ompW gene expression at high osmolarity in V. cholerae in the presence of glucose [76].
Bacteria possess small nucleotide proteins such as H-NS, StpA, Ihf, and Hu with functional similarity to eukaryotic histones, which affect several porin genes [41]. H-NS is a master global regulator, which controls the expressions of several porin genes such as ompC, ompF, ompS1, and ompS2. H-NS represses ompC gene expression and diminishes the production of OmpF. StpA is a paralogue of H-NS and is an RNA chaperone. H-NS and StpA repress ompS1 and ompS2 gene expressions in E. coli and S. Typhimurium. [77, 78]. On the other hand, H-NS and StpA stimulate the production of the outer membrane maltoporin LamB through posttranscriptional control of the maltose regulon activator MalT [79].
Ihf is one of the most abundant sequence-specific DNA-binding proteins and is a global regulator. The Ihf protein negatively regulates ompC expression, and it is necessary for the negative osmoregulation of ompF. Ihf affects ompC and ompF in two distinct ways: directly by binding upstream to the promoter regions and indirectly by influencing the expression of EnvZ-OmpR [80].
LeuO is a LysR-type regulator that controls the expression of several genes in response to stress, virulence, and biofilm accumulation. The OmpS1 and OmpS2 quiescent porins are silenced by H-NS [78, 81], while LeuO acts as an antagonist of H-NS, thereby derepressing ompS1 and ompS2 gene expressions [41, 62].
Small untranslated regulatory RNAs, often referred to as noncoding RNA, also affect porin regulation. MicF is one such example. In general, they inhibit translation of the transcripts by direct RNA-RNA interaction [41]. The sRNAs have been found to play diverse physiological roles in response to stress, metabolic regulation, control of bacterial envelope composition, and bacterial virulence [38, 82–84]. It has been shown that enterobacteria use many sRNAs such as MicC, MicA, InvR, RybB, CyaR, IpeX, and RseX to fine-tune the outer membrane composition at the posttranscriptional level [84].
4. Transport of Carbohydrates and Carbon Catabolite Regulation
4.1. Transport of Substrate Molecules and PTS
The first step in the metabolism of carbohydrates is the transport of these molecules into the cell (Figure 4). In bacteria, various carbohydrates can be taken up by several mechanisms [85]. Primary transport of sugars is driven by ATP, while secondary transport is driven by the electrochemical gradients of the translocated molecules across the membrane [86], where the secondary transport systems contain the symporters which cotransport two or more molecules, uniporters that transport single molecule, and antiporters that countertransport two or more molecules. Sugar symporters usually couple the uphill movement of the sugar to the downhill movement of proton (or sodium ion). Namely, the electrochemical proton (or sodium ion) gradient drives the accumulation of glucose [85]. Sugar uptake by group translocation is unique for bacteria and is involved in the phosphotransferase system (PTS) (Figure 4).
Figure 4.

Outer and inner membrane and periplasm and glucose transport by PTS and non-PTS.
Once glucose was transported inside periplasm, it can be internalized into the cytoplasm by the phosphotransferase system (PTS). It may be considered that the glucose concentration in the periplasm is low due to active transport systems in the cytoplasmic membrane [53]. Once inside the periplasm, glucose can be transported into cytosol by PTS, where PTS is widespread in bacteria and absent in archaea and eukaryotic organisms [87]. PTS is composed of the soluble and nonsugar-specific components Enzyme I (EI) encoded by ptsI and the phosphohistidine carrier protein (HPr) encoded by ptsH, where they transfer phosphoryl group from PEP to the sugar-specific enzyme IIA and IIB. Another component of PTS, is the enzyme IIC (in some cases also IID) which is an integral membrane protein permease that recognizes and transports the sugar molecules, where it is phosphorylated by EIIB. There have been reported to exist 21 different enzyme II complexes in E. coli, that are involved in the transport of about 20 different carbohydrates [88]. In E. coli, EIIGlc and EIIMan are involved in the transport of glucose. The EIIGlc is composed of the soluble EIIAGlc encoded by crr and of the integral membrane permease EIICBGlc encoded by ptsG. The EIIMan complex is composed of the EIIABMan homodimer enzyme and the integral membrane permease EIICDMan (Figure 4), where these proteins are encoded in the manXYZ operon [53]. In addition to mannose, these proteins can also transport glucose, fructose, N-acetylglucosamines, and glucosamine with similar efficiency [89]. In a wild-type strain growing on glucose, ptsG is induced, while manXYZ operon is repressed. In ptsG mutant, the glucose can be transported by EIIMan complex, and the cell can grow with less growth rate than the wild-type strain [90]. When the extracellular glucose concentration is less than about 1 μM, or it is more than about 2 g/L for pts mutants, this can be also utilized [91]. The induction of these genes is caused by the intracellular galactose that functions as an autoinducer of the system [92]. One of the genes induced under glucose limitation is galP, that codes for the low-affinity galactose: H+ symporter GalP (Figure 4).
The genes in the mglABC operon encode an ATP-binding protein, a galactose/glucose periplasmic binding protein, and an integral membrane transporter protein, respectively, forming Mgl system for the galactose/glucose (methyl galactoside) import [53]. This high-affinity porter belongs to the ATP-binding cassette (ABC) superfamily of the primary active class of transporters [53]. When extracellular glucose concentration is very low, the Mgl system together with LamB attains high-affinity glucose transport [53]. The glucose molecule transported either by GalP or Mgl systems must be phosphorylated by Glk encoded by glk from ATP to become G6P (Figure 4) [93].
Note that PTS seems to be quite efficient as it consumes one mole of PEP for each internalized and phosphorylated glucose, where one mole of PEP is equivalent to one mole of ATP, since the conversion of PEP to PYR by Pyk would yield one mole of ATP by substrate-level phosphorylation. The high-affinity Mgl-glucokinase system is energetically the most expensive, as it consumes two moles of ATP per glucose. The GalP-glucokinase system requires one mol of H+ that is internalized into the cytoplasm and one mol of ATP (Figure 4).
4.2. Carbon Catabolite Regulation
Among the culture environment, carbon sources are by far important for the cell from the point of view of energy generation and biosynthesis. Most living organisms including bacteria can use various compounds as carbon sources, where these can be either cometabolized or selectively used with preference for the specific carbon sources among available carbon sources. One typical example of selective carbon-source usage is the diauxie phenomenon observed in Escherichia coli and others when a mixture of glucose and other carbon sources such as lactose was used as a carbon source, where this phenomenon was first observed by Monod [94]. Subsequent investigation on this phenomenon has revealed that selective-carbon source utilization is common and that glucose is the preferred carbon source in many organisms. Moreover, the presence of glucose often prevents the use of other carbon sources. This preference of glucose over other carbon sources has been named as glucose repression, or more generally carbon catabolite repression (CCR) [95]. CCR is observed in most heterotrophic bacteria which include facultatively autotrophic bacteria that repress the genes for CO2 fixation in the presence of organic carbon source [96]. Some pathogenic bacteria such as Chlamydia trachomatis and Mycoplasma pneumonia seem to lack CCR, where these are adapted to nutrient-rich host environments [97, 98]. Another phenomenon can be seen in Corynebacterium glutamicum, where coassimilation of glucose and other carbon sources is made, but it is highly regulated [99, 100]. It is of interest that for some bacteria such as Streptococcus thermophilus, Bifidobacterium longum, and Pseudomonas aeruginosa, glucose is not a primary carbon source, and the genes for glucose utilization are repressed when preferred carbon sources are available, where this phenomenon is called as reverse CCR [101–103]. CCR is one of the most important regulatory phenomena in many bacteria [104–106]. CCR is important for the cells to compete with other organisms in nature, where it is crucial to select a preferred carbon source in order to improve the growth rate, which then results in survival as compared to other competing organisms. Moreover, CCR has a crucial role in the expression of virulence genes, which often enable the organism to access new sources of nutrients. The ability to select the appropriate carbon source that allows fastest growth may be the driving force for the evolution of CCR [107].
The E. coli lac operon is only expressed if allolactose (a lactose isomer formed by β-galactosidase) binds and inactivates the lac repressor. Lactose cannot be transported into the cell in the presence of glucose, because the lactose permease, LacY is inactive in the presence of glucose [108]. As shown in Figure 5, phosphorylated EIIAGlc is dominant when glucose is absent and does not interact with LacY, whereas unphosphorylated EIIAGlc can bind and inactivates LacY when glucose is present [109, 110]. Note that this only occurs if lactose is present [111]. The same mechanism may be seen for the transport of other secondary carbon sources such as maltose, melibiose, raffinose, and galactose [112, 113].
Figure 5.

Inducer exclusion and the activation of adenylate cyclase in the glucose-lactose system.
Inducer exclusion has also been reported for Gram positive bacteria, and HPr is the major player in these organisms. In Lactobacillus brevis, HPr(Ser-P) is formed when glucose is present and binds and inactivate permease [114]. By contrast, the lactose permease of S. thermophilus is controlled by HPr-(His-P-) dependent phosphorylation. In the absence of glucose, HPr (His-P) can phosphorylate PTS-like domain, thereby activating the permease for lactose transport [115]. When glucose is present, HPr becomes phosphorylated on Ser46 and can no longer activate the lactose permease [116].
The central players in carbon catabolite regulation in E. coli are the transcriptional activator Crp (cyclic AMP (cAMP) receptor protein; also called as catabolite gene-activator protein (CAP)), the signal metabolite cAMP, adenylate cyclase (Cya), and the PTSs, where these systems are involved in both transport and phosphorylation of carbohydrates. The PTS in E. coli consists of two common cytoplasmic proteins, EI (enzyme I) encoded by ptsI and HPr (histidine-phosphorylatable protein) encoded by ptsH, as well as carbohydrate-specific EII (enzyme II) complexes. The glucose-specific PTS in E. coli consists of the cytoplasmic protein EIIAGlc encoded by crr and the membrane-bound protein EIICBGlc encoded by ptsG, which transport and concomitantly phosphorylate glucose as explained before. The phosphoryl groups are transferred from PEP via successive phosphorelay reactions in turn by EI, HPr, EIIAGlc and EIICBGlc to glucose. The cAMP-Crp complex and the repressor Mlc are involved in the regulation of ptsG gene and pts operon expressions. It has been demonstrated that unphosphorylated EIICBGlc can relieve the expression of ptsG gene expression by sequestering Mlc from its binding sites through a direct protein-protein interaction in response to glucose concentration. In contrast to Mlc, where it represses the expressions of ptsG, ptsHI, and crr [117], cAMP-Crp complex activates ptsG gene expression [118] (Figure 6). Since intracellular cAMP levels are low during growth on glucose, these two antagonistic regulatory mechanisms guarantee a precise adjustments of ptsG expression levels under various conditions [119] (Figure 6). It should be noted that unphosphorylated EIIAGlc inhibits the uptake of other non-PTS carbohydrates by the so-called inducer exclusion [120], while phosphorylated EIIAGlc (EIIAGlc-P) activates adenylate cyclase (Cya), which generates cAMP from ATP and leads to an increase in the intracellular cAMP level [121] (Figure 6). In the absence of glucose, Mlc binds to the upstream of ptsG gene and prevents its transcription. If glucose is present in the medium, the amount of unphosphorylated EIICBGlc increases due to the phosphate transfer to glucose. In this situation, Mlc binds to EIICBGlc, and thus it does not bind to the operator region of pts genes [119, 122, 123]. Note that if the concentration ratio between PEP and PYR (PEP/PYR) is high, EIIAGlc is predominantly phosphorylated, whereas if this ratio is low, then EIIAGlc is predominantly dephosphorylated [124, 125]. EIIAGlc is preferentially dephosphorylated when E. coli cells grow rapidly with glucose as a carbon source [124, 125]. Note also that cAMP levels are low during growth with non-PTS carbohydrates such as lactose, where PEP/PYR ratio is the key factor that controls phosphorylation of EIIAGlc, which explains dephosphorylation of EIIAGlc, resulting in low cAMP pool [124, 125]. As stated above, inducer exclusion is the dominant factor for the glucose-lactose diauxie [109, 126]. The roles of cAMP-Crp is then to express lac operon, and it is involved in CCR by activating the expression of ptsG and EIICB domain of the glucose-specific PTS, and therefore the transport of glucose [127].
Figure 6.
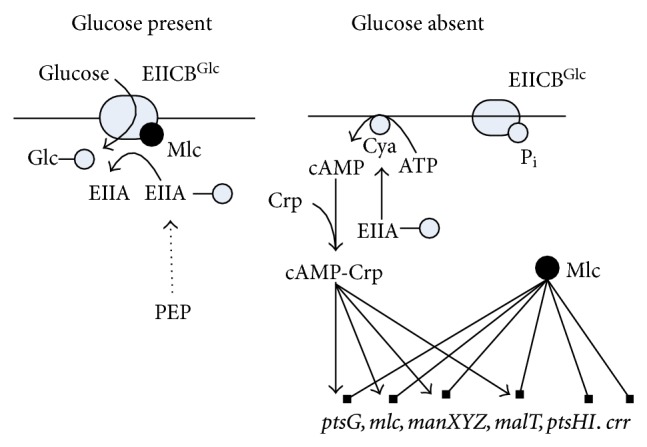
The multiple regulations by Mlc and cAMP-Crp.
4.3. Carbohydrate Uptake by Various PTS and without PTS in E. coli
As stated above, PTS is a transport system that catalyzes the uptake of a variety of carbohydrates and their conversion into their respective phosphoesters during transport [128]. PTS is composed of EI, HPr, and E II, where those accept phosphoryl group from a donor and transfer to an acceptor, thus cycling between the phosphorylated and unphosphorylated states [128]. EI and HPr are common to all PTS carbohydrates, while EII is carbohydrate specific. Thus, bacteria usually contain many different E IIs. Each E II complex consists of one or two hydrophobic integral membrane domains (C and D) and two hydrophilic domains (A and B), which together are responsible for the transport of the carbohydrate across the membrane as well as its phosphorylation. E. coli contains different EII complexes, where EII complexes are formed either by distinct proteins that contain EI and/or HPr domains exist [128]. A prominent example for the latter is FPr, which consists of HPr and EIIA domain and mediates the phosphotransfer in the uptake of fructose by E. coli. As shown in Figure 7, fructose can be transferred and phosphorylated by the fructose PTS (EIIBCFru) or ATP-dependent mannofruct kinase Mak [129]. The EIIBCFru encoded by fruA phosphorylate fructose concomitantly with transport to fructose 1-phosphate, which is further converted to FDP by an ATP-dependent Pfk [130].
Figure 7.
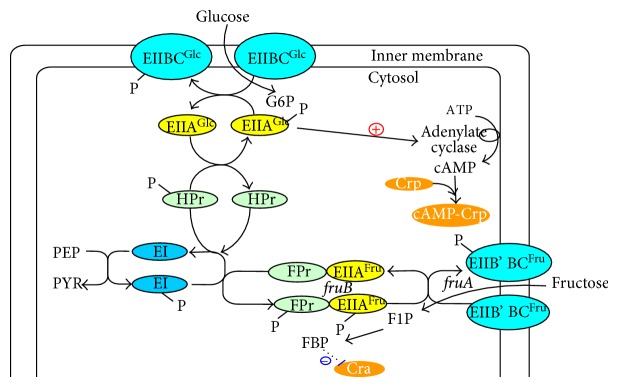
Glucose PTS and fructose PTS.
There are three pathways for the utilization of fructose as shown in Figure 8 [131]. In the primary pathway, fructose (Fru) is transported via the membrane-spanning protein FruA and concomitantly phosphorylated by a PEP: D-fructose 1-phosphotransferase (fructose PTS) system (ATP: D-fructose 1-phosphotransferase, EC 2.7.1.3), which is induced by D-fructose and enter the cell as D-fructose 1-phosphate (F1P), where this process is affected by the transfer of a phosphoryl moiety from PEP to the hexose by the concerted action of two cytoplasmic proteins: EI of PTS and a membrane-associated diphosphoryl protein (DTP). F1P is then converted to fructose 1,6-diphosphate (FDP) by ATP and by the inducible enzyme D-fructose-1-phosphate kinase (F1PK) (ATP: D-fructose-1-1phosphate 6-phosphotransferase).
Figure 8.
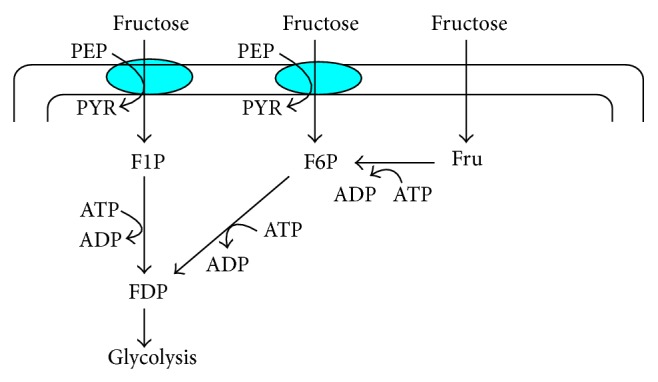
Fructose uptake pathways.
In the second pathway, fructose enters the cell via a membrane-spanning proteins that have a general ability to recognize sugars possessing the 3,4,5-D-arabino-hexose configuration which include the permeases for mannose (ManXYZ), glucitol (SrlA), and mannitol (MtlA) [131] D-fructose is converted to F6P by a specific sucrose-induced D-fructokinase (ATP: D-fructose 6-phosphotransferase, EC 2.7.1.4), and then converted to FDP by Pfk of the EMP pathway.
In the 3rd pathway, fructose enters the cell by diffusion, using an isoform of the glucose transporter PtsG. Since this mode of entry does not involve the PTS, the free fructose has to be phosphorylated by ATP to become F6P.
D-xylose is converted to D-xylulose by xylose isomerase (D-xylose ketoisomerase, EC 5.3.1.5) (Figure 9). D-Xylulose is subsequently phosphorylated by xylulokinase (ATP: D-xylulose 5-phosphotransferase, EC 2.7.1.17) to form D-xylulose 5-phosphate (X5P). Under anaerobic condition, xylulose reductase (XR) is induced, and xylitol and xylitol 5-phosphate are produced, where they may inhibit the cell growth.
Figure 9.
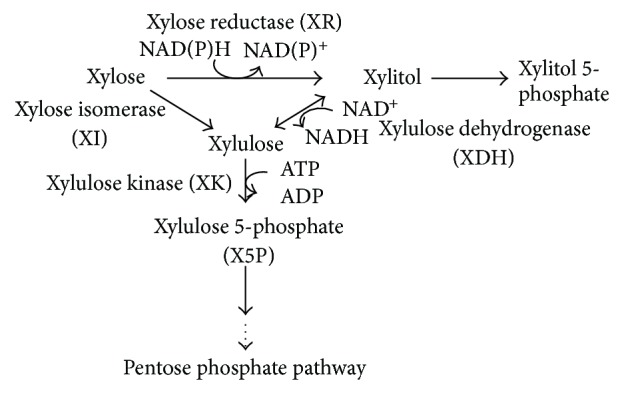
Xylose uptake pathways.
Glycerol is oxidized to dihydroxyacetone by a glycerol dehydrogenase (glycerol: NAD oxidoreductase, EC 1.1.1.6). Dihydroxyacetone is then phosphorylated by a kinase using ATP (Figure 10). Another pathway for glycerol utilization is that glycerol is phosphorylated by glycerol kinase (ATP: glycerol phosphotransferase, EC 2.7.1.30) to form L-glycerol 3-phosphate, which then is converted to GAP.
Figure 10.
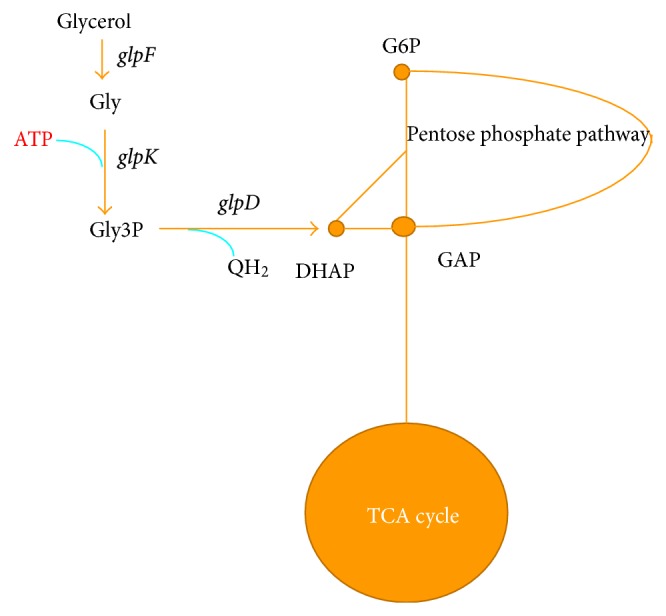
Glycerol uptake pathways.
4.4. CCR in Other Bacteria Than E. coli
The key players in CCR in Bacillus subtilis are the pleiotropic transcription factor CcpA (catabolite control protein A), the Hpr protein of the PTS, the bifunctional HPr kinase/phosphorylase (HPrK) and the glycolytic intermediates such as FDP and G6P [132–134]. Unlike E. coli, HPr phosphorylation plays an important role, where phosphorylated HPr serves as the effector for the dimeric CcpA, which controls the expressions of CCR genes [107]. The phosphorylation of HPr is catalysed by HPrK, that binds ATP, and its activity is triggered by the availability of FDP as an indicator of high glycolytic activity [135–137]. By contrast, phosphorylase activity prevails under nutrient limitation, and the activation is stimulated by the inorganic phosphate in the cell [137, 138]. Under nutrient rich condition, HPrK acts as a kinase and phosphorylates HPr, and the cofactor for CcpA is formed. The interaction between CcpA and the phosphorylated HPr is enhanced by FDP and G6P [139, 140].
With the exception of the mycoplasmas, a Firmicutes also use HPr, HPrK, and CcpA for CCR [132]. CcpA in lactic acid bacteria such as Lactococcus lactis represses not only genes of carbon metabolism, but also controls metabolic pathway genes such as glycolysis and lactic acid formation pathway genes [107].
In streptomyces coelicolor and the related species, glucose kinase is the key player of CCR, where it is independent of the PTS [141]. Corynebacterium glutamicum is important in the industrial production of amino acids, where it prefers to use multiple carbon sources simultaneously. Diauxic growth is observed for the case of using a mixture of glutamate or ethanol and glucose, where the repressor protein RamB is activated when glucose is present and binds to the promoter regions of the genes involved in acetate and ethanol catabolism [142, 143]. The ramB expression is regulated by the feedback of RamB and RamA, where RamA is activated when acetate is present. The 13 two-component systems have been found so far, and the role of five has been elucidated recently [144].
Pseudomona putida can assimilate various aromatic and aliphatic hydrocarbons, where it has been reported that the use of hydrocarbons is represses by succinate, and this seems to be a general feature of CCR in this organism [103, 145]. Under CCR, the translation of operon-specific regulators is inhibited by the binding of an RNA-binding protein Crc to mRNAs of the regulator transcript, and thus CCR seems to be governed by an RNA-binding protein at the level of posttranscriptional control rather than by a DNA-binding transcriptional regulator [146, 147].
CCR is crucial for the expression of virulence genes and for pathogenicity in many pathogenic bacteria. Note that the primary aim of pathogenic bacteria is to gain access to nutrients rather than to cause damage to the host and that the expression of virulent genes is linked to the nutrient supply of the bacteria [107].
In many Firmicutes, the mutants devoid of the HPr kinase grow significantly slower than wild-type cells. It is, therefore, suggested that HPr kinase, which generates the cofactor for CcpA, might be a suitable drug target, where the compound that inhibits the kinase activity of HPr has been identified, and this compound inhibits the growth of B. subtilis but not of E. coli, where E. coli does not contain HPr kinase [107].
Crp and cAMP are essential for the expression of virulence genes in enteric bacteria, and therefore, the corresponding crp and cya mutant strains of S. enterica and Y. enterocolitica can be used as live vaccines in mice and pigs [148–150].
4.5. Carbon Storage Regulation
The carbon storage regulator (Csr) system influences a variety of physiological processes such as central carbon metabolism, biofilm formation, motility, peptide uptake, virulence and pathogenesis, quorum sensing, and oxidative stress response [28, 151–153] (Figure 11). Csr is controlled by the RNA-binding protein CsrA, a posttranscriptional global regulator that regulates mRNA stability and translation [154–156]. CsrA binds to the 5′ untranslated region of its target mRNAs, often in the region spanning the Shine-Dalgarno (SD) site [157]. CsrA is regulated by two sRNAs called CsrB and CsrC in E. coli [158–160]. These sRNAs are composed of multiple CsrA-binding sites that bind and sequester CsrA [39].
Figure 11.
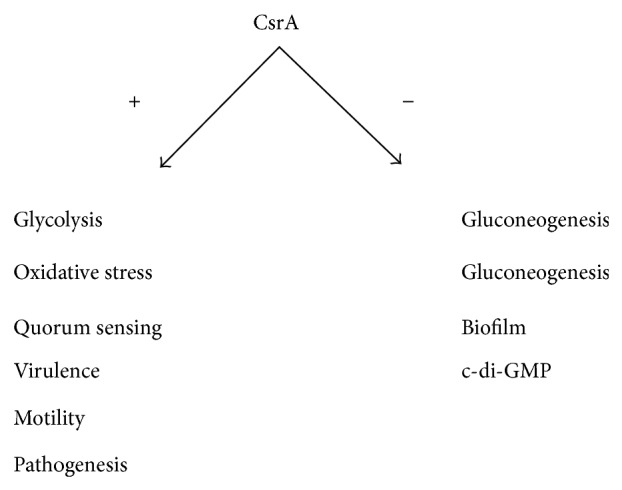
Regulation of Csr.
CsrA is a global regulator and regulates a variety of pathways as stated above, where the central carbon metabolism is of practical interest among them. CsrA negatively regulates glycogen accumulation by regulating the expressions of glgCAP operon and glgB of glgBX operon [28, 161]. As illustrated in Figure 12, CsrA regulates central carbon metabolism and glycogenesis such that glycogen synthesis pathway genes such as pgm, glgC, glgA, and glgB, as well as gluconeogenic genes such as fbp, ppsA, and pckA genes are repressed, while glycolysis genes such as pgi, pfkA (but not pfkB), tpiA, eno, and pykF genes are activated [28, 162]. It has been shown that phenylalanine production could be enhanced by manipulation of Csr [163]. More recently, it was shown that biofuel production could be enhanced by manipulating (enhancing) CsrB in E. coli [162].
Figure 12.

Pathway regulation by CsrA.
4.6. Carbon Flow Control in E. coli
In addition to cAMP-Crp, which acts depending on the level of glucose concentration, the catabolite repressor/activator protein (Cra) originally characterized as the fructose repressor (FruR) plays an important role in the control of carbon flow in E. coli [164–166]. The carbon uptake and glycolysis genes such as ptsHI, pfkA, pykF, zwf, and edd-eda are reported to be repressed, while ppsA, fbp, pckA, icdA, aceA, and aceB are activated by the Cra protein [164, 166] (Figure 13). It has been shown that the genes such as pfkA, pykF, and edd-eda have Cra binding sites that overlap or follow the RNAP-binding site [167–170]. It is known that a mutant defective in the cra gene is unable to grow on gluconeogenic substrates such as pyruvate, acetate, and lactate [171]. This appears to be due to deficiency in the gluconeogenic enzymes such as Pps, Pck, some TCA cycle enzymes, the two glyoxylate-shunt enzymes, and certain electron transport carriers [171]. Molecular level research on cra gene expression has been made by several researchers using lacZ-transcriptional fusion [170, 172–175]. The gluconeogenic pathway is deactivated by the knockout of cra gene, and the carbon flow toward glycolysis and the glucose consumption rate are expected to increase since glycolysis pathway genes such as ptsHI, pfkA, and pykF are activated by the cra gene knockout. It has been shown that cra gene knockout enables the increase of the glucose consumption rate and thus improve the rate of metabolite production under certain culture conditions [176]. However, the regulation mechanism is complex, and it must be careful since icdA, aceA, B, and cydB genes are repressed, while zwf and edd gene expressions are activated, and thus ED pathway is activated by cra gene knockout [177]. Phue et al. [178] also studied the role of the cra gene in relation to high density cell cultures of E. coliB and E. coliK.
Figure 13.
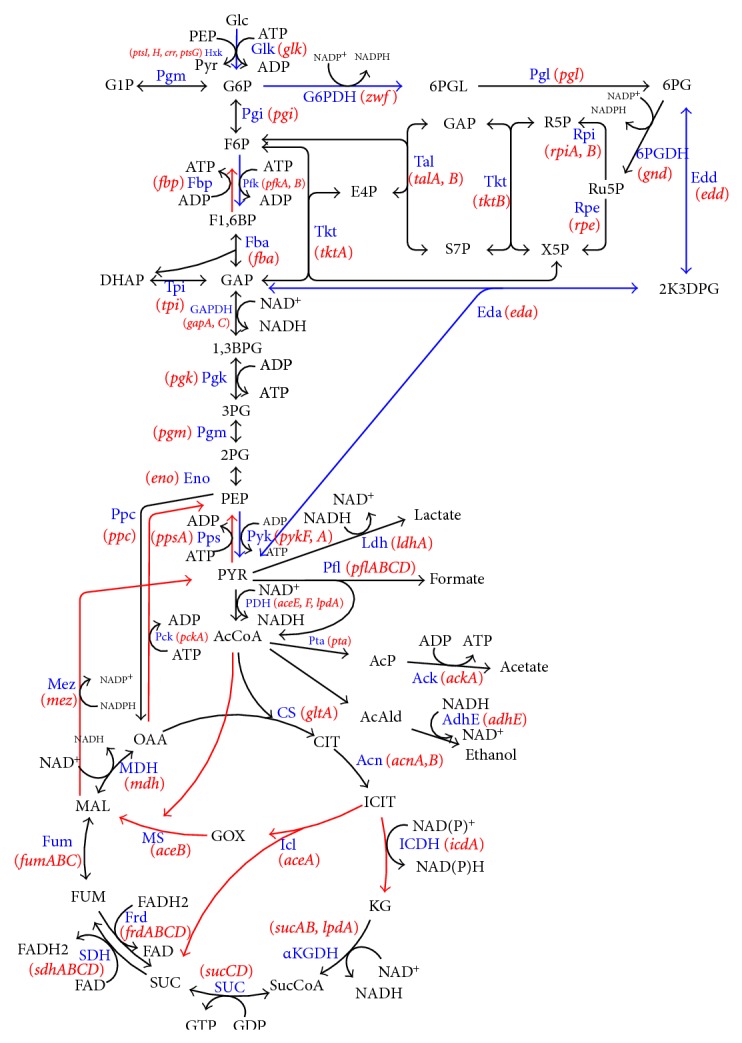
Pathway gene regulation of Cra.
4.7. Effect of Glucose Concentration on Gene Expressions in E. coli
Let us consider how the culture condition such as glucose concentration affects the global regulators and metabolic pathway genes of wild type E. coli (BW25113) [179]. Table 1 shows the fermentation characteristics of the wild type E. coli for the continuous culture at different dilution rates, where it indicates that the specific glucose uptake rate, the specific acetate production rate, and the specific CO2 evolution rate (CER) increase as the dilution rate was increased [179]. Figure 14 shows the effect of the dilution rate (the specific growth rate) on gene transcript levels, where it indicates that in accordance with the increased specific glucose consumption rate, the transcript levels of ptsG, ptsH, and pfkA are increased as the dilution rate increased, where cra transcript level decreased and crp as well as mlc decreased accordingly. The decrease in crp is also coincident with the decrease in cyaA which encodes Cya. The transcript levels of zwf, gnd, edd, and eda increased as the dilution rate increased in accordance with the decrease in cra. The transcript level of ppc increased while pckA decreased as the dilution rate was increased. Moreover, the transcript levels of fadR and iclR increased, and aceA and aceB decreased as the dilution rate increased. In accordance with the increase in the specific acetate production rate, the transcript levels of pta and ackA increased. Further observation indicates that in accordance with the decrease in rpoS transcript level, tktB, acnA, and fumC decreased as the dilution rate increased, where this phenomenon will be discussed later for nutrient starvation.
Figure 14.

The effect of dilution rate on the gene transcript levels. (a) Global regulator genes; (b) PTS, glycolysis, anaplerotic pathway, cyaA, and yfiD genes; (c) TCA and glyoxylate pathway genes; (d) PP pathway genes; (e) Respiratory chain genes. White bar: 0.2; Light grey bar: 0.4; Dark grey bar: 0.6; Black bar: 0.7.
5. Nitrogen Regulation
Next to carbon (C) source metabolism, nitrogen (N) metabolism is also important in understanding the metabolic regulation. In E. coli, assimilation of N-source such as ammonia/ammonium (NH4 +) using α-KG results in the synthesis of glutamate and glutamine (Figure 15). Glutamine synthetase (GS, encoded by glnA) catalyzes the only pathway for glutamine biosynthesis. Glutamate can be synthesized by two pathways through the combined actions of GS and glutamate synthase (GOGAT, encoded by gltBD) forming GS/GOGAT cycle, or by glutamate dehydrogenase (GDH encoded by gdhA) [180]. The GS/GOGAT cycle has a high-affinity for NH4 + (K m < 0.2 mM for GS), and therefore it is dominant when nitrogen is scarce in the medium, whereas GDH has a low-affinity for NH4 + (K m < 1 mM) and is utilized when sufficient nitrogen source is available in the medium. When extracellular NH4 + concentration is low around 5 μM or less, ammonium enters into the cell via AmtB and is converted to glutamine by GS, and UTase uridylylates both GlnK and GlnB [181] (Figure 16). When extracellular NH4 + concentration is more than 50 μM, the metabolic demand for glutamine pool rises, and UTase deuridylylates GlnK and GlnB. GlnK complexes with AmtB, thereby inhibiting the transport via AmtB, where GlnB interacts with NtrB and activates its phosphatase activity leading to dephosphorylation of NtrC and NtrC-dependent gene expression ceases [181] (Figure 16). The nitrogen intermediates such as glutamate and glutamine provide nitrogen for the synthesis of all the other N-containing components. About 88% of cellular nitrogen comes from glutamate, and the rest from glutamine [182]. The ATP required for the nitrogen assimilation using GS/GOGAT cycle under N-limiting condition accounts for 15% of the total requirement in E. coli. A significant amount of NADPH is also required for nitrogen assimilation [180, 182]. The other pathways involved in maintaining cellular nitrogen balance under specific conditions include aspartate-oxaloacetate and alanine-pyruvate shunts [183, 184].
Figure 15.

Central metabolic pathways and NH3-assimilation pathways.
Figure 16.
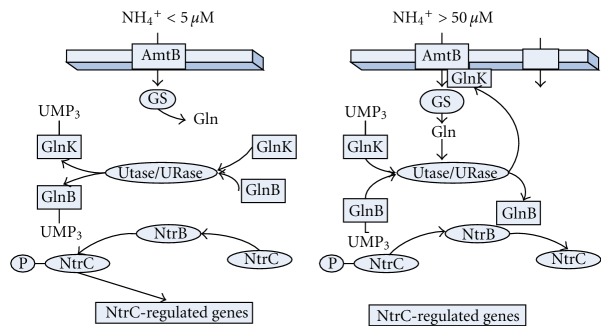
Ammonia assimilation under different NH4 + concentration.
It should be noted that carbon metabolism is not only controlled by carbon-derived signals, but also by the availability of nitrogen and other nutrient [185]. From the studies on interdependence of different metabolic routes, two of the major signal transduction systems of nitrogen and carbon metabolisms have been identified as PII, a small nitrogen regulatory protein and PTS as explained before. Because of the important roles in the regulatory functions, PII and PTS can be regarded as the central processing units of N and C metabolisms, respectively. The PII protein senses αKG and ATP, thus link the state of central carbon and energy metabolism for the control of N assimilation [185]. The glucose catabolism is modulated by the global regulators such as Cra, Crp, Cya, and Mlc while N assimilation is regulated by PII-Ntr system together with global regulators such as Crp, providing a novel regulatory network between C and N assimilation in E. coli [186]. The effects of C and N limitations on E. coli metabolism have been investigated for the continuous culture [187–191]. The C and N metabolisms may be linked by energy metabolism. It has been reported that the PII protein controls N assimilation by acting as a sensor of adenylate energy charge, which is the measure of energy available for the metabolism. The signal transduction requires ATP binding to PII, which is synergistic with the binding of αKG. Moreover, αKG serves as a cellular signal of C and N status and strongly regulates PII functions [192]. The studies on the C and N pathway interdependence have so far focused on the conversion of αKG to glutamate [193]. It is evident that the regulatory mechanism of this conversion is critical for the interdependence of C and N assimilation.
Figure 17 shows the effect of C/N ratio on the fermentation characteristics during aerobic continuous culture at the dilution rate of 0.2 h−1, where the C/N ratio is the value of the feed substrate concentrations [193]. Figure 17 indicates that the glucose concentration increases, whereas the cell concentration decreases as C/N ratio increases. Figure 17 also shows that the glucose concentration is very low at 100% and 60% of N concentrations (C-limitation), whereas its concentration is high at 20% and 10% of N concentrations (N-limitation). Note that the specific glucose consumption rate as well as the specific acetate and CO2 production rates tended to increase as C/N ratio increases.
Figure 17.
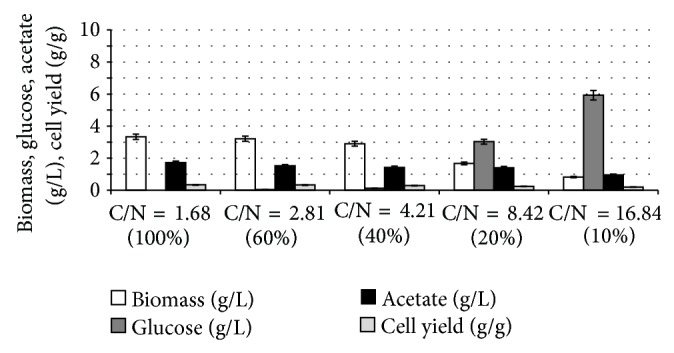
Effect of C/N ratio on the fermentation characteristics for the continuous culture at the dilution rate of 0.2 h−1.
In order to interpret the fermentation characteristics as shown in Figure 17, the relative mRNA levels are shown for different C/N ratios in Figure 18, where it shows that crp transcript level became lower as C/N ratio increases, which corresponds to the fact that cAMP-Crp level decreases as glucose concentration increases. In accordance with the change in crp transcript level, mlc level changed in a similar fashion [194]. Figure 18(a) also shows that the transcript levels of such genes as soxR/S and rpoS became higher as C/N ratio increases, which may be due to oxygen stress caused by higher respiratory activity for the former [188], along with nutrient stress for the latter [195].
Figure 18.

Comparison of the transcriptional mRNA levels of the wild-type E. coli genes cultivated at 100% (C/N = 1.68), 40% (C/N = 4.21), 20% (C/N = 8.42), and 10% (C/N = 16.8) N concentration.
The transcript level of rpoN, which encodes σ 54, increased as C/N ratio increases (Figure 18(b)). Figure 18(b) also shows that the expressions of glnA, glnL, glnG, and gltD genes changed in a similar fashion as rpoN, indicating the activation of GS-GOGAT pathway under N-limitation. The glnB gene which codes for PII also changed in a similar fashion, while glnD which controls the uridylylation and deuridylylation shows somewhat different, but the trend seems to be similar (Figure 18(b)). PII paralogue-encoding gene, glnK shows very high expressions at 20% and 10% of N-limitation (Figure 18(b)). The expression pattern of nac is similar to that of rpoN, whereas gdhA shows a reverse pattern, implying that gdhA is repressed by Nac (Figure 18(b)). The GDH pathway is favored when the organism is stressed for energy because GDH does not use ATP as does GS pathway [196]. Figure 18(b) shows the decreased expression of gdhA as C/N ratio increases. Liang and Houghton [197] investigated the effect of NH4Cl concentration on GDH and GS activities and showed the upregulations of GDH and transhydrogenase activities at lower NH4Cl concentration.
The availability of nitrogen is sensed by PII protein at the level of intracellular glutamine, where glutamine is synthesized by glutamine synthetase (GS) encoded by glnA and is transported mainly by GlnHPQ. The glnHPQ operon is under the control of tandem promoters such as glnHp1 and glnHp2, where the former is σ 70 dependent, and the latter is σ 54 dependent and NtrC-P dependent [198, 199]. It has been shown that as the major transcriptional effector of the glucose effect, Crp affects nitrogen regulation [186]. Namely, glnAp1 is activated by Crp with glutamine as N-source (Figure 19). Through glnHPQ-dependent signaling, Crp acts to decrease the amount of the phosphorylated NtrC activator, which in turn causes the decrease in glnAp2 expression [186]. However, this regulation is more complex. It has been suggested that σ 54-dependent Ntr genes of E. coli form a gene cascade in response to N-limitation [200]. The central participants of Ntr response are NRI (or NtrC) and NRII (or NtrB), and RNA polymerase complexed to σ 54. NRI is the transcriptional activator of σ 54-dependent promoters, while NRII is a bifunctional protein that can either transfer phosphate to NRI or control the dephosphorylation of NRI-phosphate. N-limitation results in the phosphorylation of NRI, which in turn stimulates the expression of glnALG operon. The expression of the glnALG operon is controlled by tandem promoters such as glnAp1 and glnAp2, where glnAp1 is a σ 70-dependent weak promoter, and its transcription can be activated by Crp and blocked by Ntr-P. On the other hand, glnAp2 is transcribed by RNA polymerase (Eσ 54) and is activated by Ntr-P. Therefore, glnAp2 is responsible for activating glnA transcription under N-limitation [201].
Figure 19.
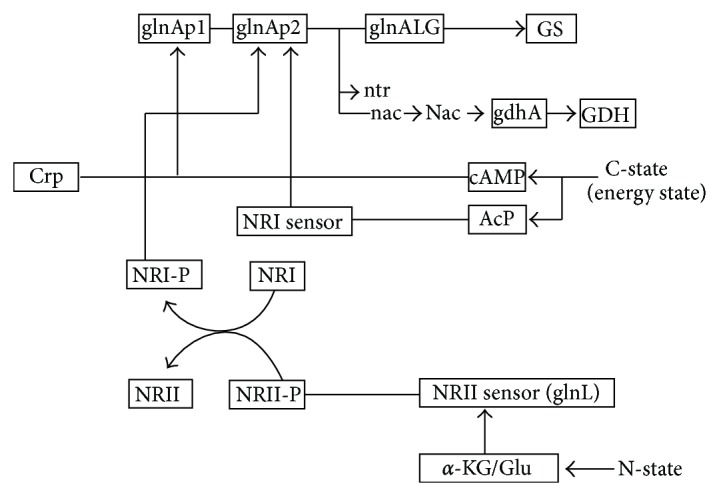
The interaction between nitrogen regulation and catabolite regulation.
It has been reported that there is no NRI-P binding sites in the gdhA regulatory region [202], and it is unlikely for NRI to directly repress gdhA promoter [203]. As it has been shown that Nac is involved in the transcriptional repression of gdhA gene under N-limitation [203], Nac seems to repress gdhA gene as shown in Figure 18, where it shows that the transcript level of gdhA gene is lower, while gltB and D genes are higher under N-limitation as compared to C-limitation. NADPH is an important cofactor in GDH and (GS)-GOGAT activities and it has been reported that transhydrogenase plays some role in the regulation of these pathways [197]. Under N-limitation, the glutamate and glutamine synthetic pathways are expected to be repressed due to shortage of NH3 for those reactions, and thus NADPH is less utilized, resulting in overproduction of NADPH. Part of this may be converted to NADH by transhydrogenase, and the converted NADH together with other NADH formed may be utilized for ATP production through respiratory chain.
E. coli possesses two closely related PII paralogues such as GlnB and GlnK, where GlnB is produced constitutively, and it regulates the NtrB (NRII)/NtrC (NRI) two-component system [181]. It has been shown that the intracellular concentrations of NRI and NRII increase upon N-limitation [204–206]. The phosphorylated NtrC is an activator of various nitrogen-controlled genes such as glnA which codes for GS [200] and glnK encoding the second PII paralogues [206]. The increased NRI, presumably in the phosphorylated form such as NRI-P activates the expression of glnK and nac promoters under N-limitation [207, 208]. Figure 18 shows that the transcript levels of glnK and nac genes increased as C/N ratio increases, where it has been reported that glnK and nac promoters are sharply activated when ammonia is used up [208]. The gltBDF operon which has been found to have binding affinity with global regulators such as Fnr and Crp in the promoter region [209].
The Ntr system is composed of four enzymes (Figure 20): a uridylyltransferase/uridylyl-removing enzyme (UTase/UR) encoded by glnD gene, a small trimeric protein, PII encoded by glnB, and the two-component system composed of NtrB and NtrC. GlnD controls the activity of GS by adenylylation/deadenylation through a bifunctional enzyme adenylyltransferase (ATase), the glnE gene product [210–212]. The activity of GlnK becomes high under N-limitation and contributes to the regulation of NtrC-dependent genes [213]. It has been shown that on GS adenylation, ATase activity is regulated by UTase/UR and PII such that upon nitrogen limitation, UTase covalently modifies PII by addition of a UMP group at a specific residue and the resultant uridylylated form of PII promotes deadenylylation of GS by ATase (Figure 20). Conversely, under N-rich condition, the uridylyl-removing activity of GlnD predominates, and the deuridylylated PII promotes adenylation of GS by ATase. Adenylylation by ATase is promoted by deuridylylated PII which is produced by UR action on PII (UMP)3 under higher N-concentration (low C/N ratio) (Figure 20). These indicate that UTase/UR and PII acting together sense the intracellular nitrogen status [214]. The PII signal transduction proteins such as GlnB and GlnK are uridylylated/deuridylylated in response to intracellular glutamine level, where low intracellular glutamine level, signalling N-limitation, leads to uridylylation of GlnB [214]. GlnB is shown to be allosterically regulated by α-KG, and thus GlnB may play a role in integrating signals of C/N status. The NtrB/NtrC two-component system and GlnE which adenylylates/deadenylylates GS are the receptors of GlnB signal transduction [213]. It has been suggested that the carbon/cAMP effect is mediated through GlnB uridylylation [213].
Figure 20.

Overall mechanism of nitrogen assimilation in E. coli under C-limited (N-rich) and N-limited conditions.
The phosphorylated NRI/NtrC (NRI/NtrC-P) activates transcription from N-regulated σ 54-dependent promoters by binding to the enhancers [214–217]. PII and the related GlnK protein control the phosphorylation state of NRII/NtrB by stimulating the phosphatase activity of NRII. The ability of GlnK and PII to regulate the activities of NRII is in turn regulated by the intracellular signals of C and N availability via allosteric control [215].
6. Phosphate Regulation
The phosphate (P) metabolism is also quite important from the energy generation and phosphorelay regulation points of view. The phosphorous compounds serve as major building blocks of many biomolecules and have important roles in signal transduction [65]. The phosphorus compounds serve as major building blocks of many biomolecules and have important roles in signal transduction [65]. The phosphate is contained in lipids, nucleic acids, proteins, and sugars and is involved in many biochemical reactions by the transfer of phosphoryl groups [218]. Moreover, phosphate metabolism is closely related to the diverse metabolisms such as energy and central carbon metabolisms [219]. All living cells sophisticatedly regulate the phosphate uptake, and survive even under phosphate-limiting condition [220, 221]. Escherichia coli contains about 15 mg of phosphate (P) per g (dry cell weight) [222]. Depending on the concentration of environmental phosphate, E. coli controls phosphate metabolism through Pho regulon, which forms a global regulatory circuit involved in a bacterial phosphate management [65, 223]. The PhoR-PhoB two-component system plays an important role in detecting and responding to the changes of the environmental phosphate concentration [224–226]. It has been known that PhoR is an inner-membrane histidine kinase sensor protein that appears to respond to variations in periplasmic orthophosphate (Pi) concentration through interaction with a phosphate transport system, and that PhoB is a response regulator that acts as a DNA-binding protein to activate or inhibit specific gene transcription [65, 227–229]. The activation signal, a phosphate concentration below 4 μM, is transmitted by a phosphorelay from PhoR to PhoB. Phospho-PhoB in turn controls Pho regulon gene expressions. PhoB is phosphorylated by PhoR under phosphate starvation or by PhoM (or CreC) in the absence of functional PhoR [230–236].
The E. coli Pho regulon includes 31 (or more) genes arranged in eight separate operons such as eda, phnCDEFGHIJKLMNOP, phoA, phoBR, phoE, phoH, psiE, pstSCAB-phoU, and ugpBAECQ [237]. When Pi is in excess, PhoR, Pst, and PhoU together turn off the Pho regulon by dephosphorylating PhoB. In addition, two Pi-independent controls that may be formed of cross-regulation turn on the Pho regulon in the absence of PhoR. The sensor CreC, formerly called PhoM, phosphorylates PhoB in response to some (unknown) catabolite, while acetyl phosphate may directly phosphorylate PhoB [223]. When Pi is in excess, Pi is taken up by the low-affinity Pi transporter, Pit. Four proteins such as PstS, PstC, PstA, and PstB form an ABC transporter important for the high-affinity capture of periplasmic inorganic phosphate (Pi) and its low-velocity transport into the cytosol [238]. These proteins are encoded together with PhoU as the pstSCAB-phoU operon. PstS is a periplasmic protein that binds Pi with high-affinity. PstC and PstA are inner membrane channel proteins for Pi entry, while PstB is an ATP-dependent permease that provides the energy necessary for Pi transport from periplasm to cytosol (Figure 21). When phosphate is in excess, the Pst system forms a repression complex with PhoR, and prevents activation of PhoB. PhoU and PstB are also required for dephosphorylation of phospho-PhoB under P-rich condition [239]. Indeed, PhoU is essential for the repression of the Pho regulon under high phosphate condition [64]. It may be considered that PhoU acts by binding to PhoR, PhoB or PhoR/PhoB, complex to promote dephosphorylation of phosphorylated PhoB or by inhibiting formation of the PhoR-PhoB complex [240].
Figure 21.
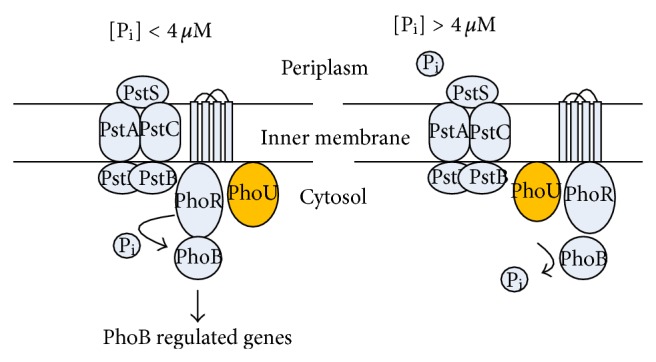
Molecular mechanism of phosphate regulation.
It has been shown that phoB mutant does not synthesize alkaline phosphatase (phoA gene product) [241–246] and phosphate-binding protein (pstS gene product) [242, 245, 246]. It was observed that phoU expression changed depending on phosphate concentration of the phoB mutant [247]. Since the phoA gene mutation leads to the decreased content of membrane proteins or completely lacks them, mutations in the phoB gene result in the loss of alkaline phosphate and two membrane proteins [248]. Nesmeianova et al. [240] found that phoB mutation leads to the loss of polyphosphate kinase activity which catalyzes the synthesis of polyP in E. coli. Ault-Riché et al. [248] also found that the strains with deletion of phoB failed to accumulate polyP in response to osmotic stress or nitrogen limitation. Mutations in the phoB gene had no effect on pepN [249] and lky (tolB) expressions [250].
The expressions of the genes under the control of the PhoR-PhoB two-component system were found to be affected by the duration of P-limitation in response to phosphate starvation in E. coli. This means that the roles of the PhoR-PhoB two-component regulatory system seem to be more complex [226]. Since phosphate starvation is a relatively inexpensive means of gene induction in practice, the phoA promoter has been used for overexpression of heterologous genes [251]. A better understanding of the Pho regulon would allow for the optimization of such processes [238].
Figure 22 shows the effect of P concentration on the transcript levels, where it indicates that phoB transcript level increased as P concentration decreases, and phoB regulated genes such as phoA, phoE, phoH, phnC, pstS, and ugpB were all increased in a similar fashion [252]. Note that phoU and phoM changed in a similar fashion as phoR, and also that the transcript level of rpoD, which encodes the RNA polymerase holoenzyme containing σ 70, increased in a similar fashion as PhoB regulatory genes [253].
Figure 22.
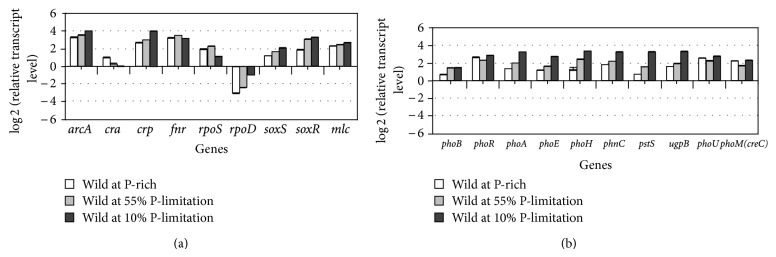
Comparison of the transcript levels of the wild-type E. coli cultivated with different P concentrations of the feed (100%, 55%, and 10%): (a) global regulatory genes and (b) PhoB regulatory genes.
The effect of phoR gene knockout on the selected gene transcript levels is also shown in Figure 23 where it indicates that the phoB-regulated genes such as phoA, phoE, phoH, phnC, pstS, and ugpB were more downregulated for the phoR mutant as compared to phoB mutant, whereas phoU and phoM (creC) were less affected by phoR gene knockout [252].
Figure 23.

Comparison of the transcript levels of Pho regulon genes for the wild-type, phoB, and phoR mutants cultivated at 10% P-concentration.
When cells enter into Pi-starvation phase in the batch culture, the Pho regulon is activated and σ S starts to accumulate in the cytosol [65, 254, 255]. The promoters of the Pho genes are recognized by σ D-associated RNA polymerase. A mutation in rpoS, significantly increases the level of AP (alkaline phosphatase) activity, and the overexpression of σ S inhibits it [256]. It has been reported that in rpoS mutant, the expression of AP was considerably higher than that in wild-type strain, implying that σ S is involved in the regulation of AP. Other Pho genes such as phoE and ugpB are likewise affected by σ S. The rpoS may inhibit the transcriptions of phoA, phoB, phoE, and ugpB, but not that of pstS [256]. The pst may be transcribed by both σ S and σ D. The Pho regulon is thus evolved to maintain a tradeoff between cell nutrition and cell survival during Pi-starvation [256]. The previous reports suggest that the Pho regulon and the stress response are interrelated [255–260].
The presence of glucose or mutations in cya or cAMP receptor protein gene (crp) leads to the induction of phoA gene in phoR mutatnt. This induction requires the sensor PhoM (CreC) and the regulator PhoB [261]. However, PhoM (CreC) may not detect glucose per se, where it may detect an intermediate in the central metabolism. Therefore, cya or crp mutation may indirectly affect PhoM-(CreC-) dependent control. In addition to Pi control, two Pi-independent controls may lead to the activation of PhoB. These two may be connected to control pathways in carbon and energy metabolisms, in which intracellular Pi is incorporated into ATP. One Pi-independent control is the regulation by the synthesis of acetyl phosphate (AcP), where Pi is incorporated into ATP at Ack (acetate kinase) pathway. AcP may act indirectly on PhoB.
7. Oxygen Level Regulation
7.1. Effect of Oxygen Limitation on the Metabolism
In addition to nutrient sources, oxygen level is also quite important from the metabolic regulation point of view. Global regulators such as Fnr and ArcAB are mainly responsible for the regulation of the availability of oxygen and other electron acceptors in the culture environment, where Fnr regulates the expressions of metabolic pathway genes under anaerobic condition [262], while ArcAB regulates under both anaerobic and microaerobic conditions [263, 264]. It has been shown that ArcA/B system exerts more significant effect on the cell metabolism under microaerobic condition than under aerobic or anaerobic condition. The effect of ArcAB system on the flux distribution at pyruvate node has been investigated based on the extracellular metabolite concentrations [265, 266]. It was shown that lactate can be overproduced by arcA/fnr double mutant [265] in a similar way as pfl gene knockout [267, 268].
Reoxidation of the reducing equivalents such as NADH generated by the oxidation of the energy source occurs in the respiratory chain under aerobic or microaerobic condition. In E. coli, NADH is oxidized in the respiratory chain via a coupled NADH dehydrogenase NDH-1 encoded by nuo or an uncoupled dehydrogenase NDH-2 encoded by ndh, and the electron flows into quinone and quinol pool. Quinol is then oxidized by either the cytochrome bo or the cytochrome bd terminal oxidase complex, which in turn passes the electrons to oxygen with concomitant production of water. The two terminal oxidases differ in their affinities for oxygen as well as in their H+/e-stoichiometries, where cytochrome bo oxidase has a low-affinity for oxygen and translocates two H+s per e−, while cytochrome bd has a high-affinity to oxygen and translocates one H+ per e−. The cyoABCDE operon is represses by both ArcA and Fnr, while cydAB operon is activated by ArcA and repressed by Fnr [269].
The microbial cells such as E. coli can generate energy as ATP under wide ranges of redox condition. The reducing equivalents such as NADH are reoxidized in the respiratory chain, where oxygen, nitrate, fumarate, and dimethyl sulfoxide, and so forth, are the electron acceptors. This process is coupled to the formation of a proton motive force (PMF), which is utilized for ATP generation from ADP and Pi. In the absence of oxygen, or other electron accepters, ATP is generated via substrate level phosphorylation through the process of degradation of carbon source in the metabolic pathways. Under such fermentation condition, the cell such as E. coli excretes such metabolites as lactate, ethanol, succinate, and formate (CO2 and H2 as well) as well as acetate, where the relative production rates for these metabolites are governed by the demand for redox neutrality. The succinate is formed from PEP via Ppc. Pyruvate serves as a common substrates for pyruvate formate-lyase (Pfl) and the pyruvate dehydrogenase complex (PDHc), and this branch point involves the cleavage of PYR. The expressions of pfl genes which encode Pfl is activated by ArcA and Fnr, and it becomes higher at lower oxygen concentrations, whereas aceE,F which encode α and β subunits of PDHc is repressed by ArcA under oxygen limited condition. At the branch point of AcCoA, the product of both Pfl and PDHc reactions, is converted to either acetate and ethanol or subsequently undergo further oxidation in the TCA cycle. The interconversion of Pfl between inactive and active glycyl radical-bearing species occurs at low oxygen concentration and is controlled by the activities of the iron-sulfur protein, Pfl activase, and the product of the adhE gene, Pfl deactivase [270, 271]. The active glycyl radical form of Pfl is irreversibly destroyed by molecular oxygen and hence must be either protected from oxygen damage or converted to the inactive, oxygen-stable species during the transition between anaerobiosis and aerobiosis [272, 273].
7.2. Regulation by ArcA/B System
The Arc (anoxic respiration control) system, composed of ArcA, the cytosolic response regulator, and ArcB, the membrane bound sensor kinase, regulates the TCA cycle genes depending on the oxygen level or redox state. The ArcB protein has three cytoplasmic domains: a primary transmitter domain (H1) containing a conserved His292, a receiver domain (D1) containing a conserved Asp576, and a secondary transmitter domain (H2) containing a conserved His717 [272, 274–276]. The primary transmitter domain of ArcB is autophosphorylated at His292 at the expense of ATP [277, 278]. The phosphoryl group is then sequentially transferred to Asp576 and His717 and from there to Asp54 of ArcA. However, the phosphoryl group on His292 could also be directly transferred to ArcA at a very low rate [277]. On the other hand, the phosphoryl group from His717 could also be transferred to ArcA, but this transfer is regulated by redox conditions [279]. Namely, upon stimulation by the redox state, ArcB undergoes autophosphorylation, and the phosphoryl group is transferred to ArcA by the His→Asp→His→Asp phosphorelay [280]. Consequently, the phosphorylated ArcA binds to the promoter regions of the TCA cycle and other genes. It has been reported that ArcA, when phosphorylated, represses the expressions of the genes involved in the TCA cycle and the glyoxylate shunt genes such as gltA, acnAB, icdA, sucABCD, sdhCDAB, fumA, mdh, and aceA, B [281–286]. Moreover, the genes which encode the primary dehydrogenases such as glpD, lctPRD, aceE,F and lpdA are also repressed by ArcA [287–289]. Escherichia coli possesses two terminal quinol oxidases in the respiratory chain. The genes cyoABCDE, which encode cytochrome o oxidase that has a low oxygen affinity and mainly functions under aerobic condition, are repressed by ArcA [290]. On the other hand, the cydAB genes which encode cytochrome d oxidase that has high oxygen affinity are activated by ArcA [287, 291, 292].
Alexeeva et al. [263] investigated the effects of different oxygen supply rates on the catabolism in arcA mutant. It was shown that ArcAB system exerts more significant effect on the cell's catabolism under microaerobic condition than under aerobic or anaerobic condition. A strong link is demonstrated between redox ratio (NADH/NAD+) and acetate overflow in E. coli [293]. It was shown that the commencement of acetate overflow occurred above the critical NADH/NAD+ ratio of 0.06 [293]. Moreover, acetate overflow is delayed by the expression of heterologous NADH oxidase (NOX), an enzyme that serves to reduce the NADH/NAD+ ratio [293]. The redox state has been reported to trigger the Arc regulon [294, 295].
Since phosphorylated ArcA represses TCA cycle genes, the arcA gene deletion activates the TCA cycle, resulting in the reduction in the acetate formation [293]. The NADH oxidation by the expression of NOX in the arcA gene knockout mutant further reduced the acetate formation, resulting in the increased recombinant protein production [293]. Since TCA cycle is the main source of energy generation and provides important precursors for amino acids such as glutamate, and lysine, it is of practical interest to enhance the TCA cycle activity. As stated above, the arcA/B genes knockout in E. coli transcriptionally activates the TCA cycle and overproduces NADH, which may in turn repress the TCA cycle by its allosteric regulation. Moreover, it has been reported that ArcAB does not control the TCA cycle under aerobic condition due to the fact that oxidized quinone electron carriers inhibit autophosphorylation of ArcB, and it can not transphosphorylate ArcA [294] (Figure 24). As expected from the above mentioned regulation, the TCA cycle is activated by arcA/B gene knockout, which then causes higher NADH/NAD ratio, which in turn represses TCA cycle activity [296]. Vemuri et al. [296, 297] considered to express heterologous nox gene to oxidize NADH, and in turn activate TCA cycle, while nicotinic acid and Na nitrate may also activate TCA cycle [298].
Figure 24.

Fnr and ArcA/B and respiratory chain regulation.
Since the TCA cycle is the source of energy generation and provide some of the precursors for the cell synthesis, the activation of the TCA cycle may lead to the improvement of ATP production for the cell growth and/or the TCA cycle-related metabolite productions in practice. It should be noted that the activation of TCA cycle reduced the acetate production rate, which is the common obstacle for the metabolite production using E. coli. It should, however, also be noted that the activation of the TCA cycle caused the decrease in the cell yield due to higher production of CO2 in the TCA cycle. This may be overcome by activating the glyoxylate pathway by fadR gene knockout [299], and so forth. It is controversial whether the cell metabolism is controlled to maximize ATP generation or cell synthesis, and so forth [300].
7.3. Fnr and Nar Systems
Respiration is a fundamental cellular process utilizing different terminal electron acceptors such as oxygen and nitrate. The ability to sense these electron acceptors is a key for the cells to survive. Escherichia coli is a metabolically versatile chemoheterotroph grown on a variety of substrate under various oxygen concentrations with fumarate or nitrate, replacing oxygen as terminal electron acceptor under anaerobic condition [301]. Many bacteria utilize oxygen as the terminal electron acceptor, but they can switch to other acceptors such as nitrate under oxygen limitation. In E. coli, this switch from aerobic to anaerobic respiration is controlled by Fnr (fumarate and nitrate reduction), where it was identified by Spiro and Guest [302]. Under oxygen limitation, Fnr binds a [4Fe-4S]2+ cluster and becomes a transcriptionally active dimeric form.
Escherichia coli possesses sensing/regulation systems for the rapid response to the availability of oxygen, redox state as represented by NADH/NAD+ ratio, and the presence of other electron acceptors. Those regulation systems channel electrons from donor to terminal acceptors. The pyridine nucleotides such as NADH and NAD+ function as the important redox carriers involved in the metabolism. These coenzymes not only serve as electron acceptors in the breakdown of substrates, but also provide the reducing power for the redox reactions in the anaerobic and aerobic respirations. A balance for oxidation and reduction of these nucleotides is regulated for catabolism and anabolism, since the turnover of the nucleotides is very high compared to their concentrations [303]. Under anaerobic condition, the reoxidation of NADH and the formation of reduced compounds occur, whereas NADH oxidation is coupled to the respiration by electron transfer under aerobic or nitrate respiration.
The metabolic regulation is made by the binding of dimeric Fnr to the promoter regions of the relevant genes with affinities depending on the redox state [304]. The ability of Fnr to bind DNA is regulated by the change in equilibrium between monomeric apo Fnr (inactive) and dimeric Fnr (active) in vivo. The active form of Fnr binds to DNA to regulate the corresponding genes under anaerobic condition. Molecular oxygen can oxidize the ion-sulfer cluster of the corresponding region, resulting in monomerization of the protein and subsequent loss of its ability to bind DNA [305]. Nar (nitrate reduction) plays a role when nitrate is present, and belongs to the two-component redox regulation systems, where it comprises a membrane sensor (NarX) that may act as a kinase causing phosphorylation of the regulator (NarL) under certain conditions. The Nar system activates such genes as nitrate reduction encoding nitrate and nitrite reductases and represses such genes as fumarate reductase genes.
The sequence of the fnr gene revealed that it encodes a protein which shows a significant homology to CAP/Crp (for catabolic activator protein). However, a number of significant differences between the two proteins have been investigated. Fnr is a monomeric protein, and it does not have the conserved group of surface residues that interact with cyclic AMP. It contains an oxygen labile iron-sulfur center as a sensor element for anaerobiosis [282, 306–308]. Several studies have been conducted on the structure and gene sequence for Fnr and Crp proteins. From these studies, it was found that both Fnr and Crp proteins possess almost similar structure and gene sequence. The genes that are controlled by these two global regulators have similar binding sites [308–310]. Even if some mutation changed the structure of proteins, the mutation in Fnr protein could convert to Crp protein, and similarly Crp protein could convert to Fnr protein [302]. It may be also considered that both Crp and Fnr proteins can form heterodimer, which might not allow both of them to function properly [306–308]. Then, the absence of Fnr protein or gene allows Crp protein to bind more effectively to the target gene sequence.
Since Fnr is known to activate frd and pfl genes, the fnr mutant produced less succinate and formate [311]. Although arcA is known to be activated by Fnr, the regulation mechanism is somewhat complicated (Figure 24). Namely, cyo and cyd genes are repressed, by Fnr, while cyo is repressed, and cyd is activated by ArcA. The fnr mutant shows a decreased gene expression of arcA, and an increased gene expressions of both cyoA and cydB. This implies that the activated cytochrome oxydase increased quinone pool, which inhibits ArcB phosphorylation and in turn decreases phosphorylation of ArcA, where arcA gene expression also decreases due to fnr gene knockout.
7.4. Effect of Excess Oxygen on the Metabolism
On the other hand, the microbial cell responds to oxidative stress by inducing antioxidant proteins such as superoxide dismutase and catalase [312]. The well characterized pleiotropic regulators of the antioxidant responses are the OxyR and SoxR [313]. The activation of both proteins results in the transcriptional enhancement of sets of genes whose products relieve the stress by eliminating oxidants and preventing or repairing oxidative damage [313]. SoxR is a member of the MerR family of metal-binding transcription factors, and it exists in solution as a homodimer with each subunit containing a [2Fe-2S] cluster. These clusters are in the reduced state in inactivated SoxR, and their oxidation activates SoxR as a powerful transcription factor [314]. The active form of SoxR activates transcription of sox gene. The sox gene product, SoxS, belongs to the AraC/XylS family of DNA-binding transcription factors [315]. SoxS regulates the expressions of more than 17 genes or operons [316–320].
Oxygen derivatives such as superoxide (O2 •−), hydrogen peroxide (H2O2), and the hydroxyl radical (•OH) are usually generated as toxic by-products of aerobic metabolism in a cascade of monovalent reductions from molecular oxygen. Although these are not so reactive per se, O•− and H2O2 have been reported to cause severe cell damage. H2O2 along with Fe2+ via the Fenton reaction produces •OH, which can react with any macromolecule such as protein, membrane constituents, and DNA [321, 322]. O2 •− exacerbates the Fenton reaction by increasing the intracellular pool of “free iron,” for instance, by releasing iron from O2 •−-oxidized [4Fe-4S] clusters. O2 •− may also react with nitric oxide (•NO), producing highly reactive peroxynitrite (ONOO−), which can generate •OH. Despite their potential toxicity, reactive oxygen species (ROS) at low concentration have been shown to be actively involved in the cell's life and, therefore, should not be entirely eliminated. Potent basic defense systems maintain ROS at harmless levels but cannot deal with sudden increases in ROS production. This creates an imbalance between production and elimination, referred to as oxidative stress.
Early studies using two-dimensional gel electrophoresis to analyze variations in protein expressions have shown that the synthesis of more than 80 proteins was activated in response to oxidative stress [312]. Some of these induced proteins are identified as possessing fundamental antioxidant functions, for example, superoxide dismutase and catalase. The search for mutants with altered antioxidant defenses led to the isolation and characterization of pleiotropic regulators that operate as redox-regulated genetic switches [315, 323, 324]. The best characterized pleiotropic regulators of the antioxidant responses are the OxyR and SoxR proteins [313]. Both proteins have the remarkable ability of directly transducing oxidative signals to genetic regulation. Both proteins are expressed constitutively in an inactive state and are transiently activated in cells under specific types of oxidative stress. The activation of the OxyR and SoxR proteins results in the transcriptional enhancement of sets of genes (regulons) whose products relieve the stress by eliminating oxidants and preventing or repairing oxidative damage [313].
The two enzymes involved in the oxidative PP pathway, G6PDH and 6PGDH that provide NADPH for biosynthesis, are significantly affected in both soxR and soxS mutants [325]. The activities of G6PDH and 6PGDH decreased in both soxR and soxS mutants, compared to the parent stain. The downregulations of these two enzymes agreed with the slower growth rates in both mutants, since these enzymes are known to be under growth rate-dependent regulation [326]. The downregulation of zwf gene in both mutants is also due to the effects of soxS and soxR genes deletion, since zwf is a member of soxRS and multiple antibiotic resistance (mar) regulons. Thus, unlike gnd, zwf expression is transcriptionally activated by SoxS during episodes of oxidative stress [323, 327]. The pntA (membrane bound transhydrogenase) transcripts which is involved in NADPH generation [328], are upregulated in both soxR and soxS mutants. This may be due to the down-regulation of NADPH-generating enzymes such as G6PDH and 6PGDH in PP pathway, and ICDH in TCA cycle, since NADPH plays a significant role to reduce oxidative stress [323].
8. Acid Schock or the Effect of pH
The acid barrier of the stomach represents a strong challenge for many pathogenic enterobacteria. Enteric bacteria that cause disease in the human intestine endure a transient but extreme acid condition in the stomach. The normal stomach shows an acid environment at around pH 2 with an emptying time of about 2 h [329]. Unlike acid sensitive Vibrio cholerae, Escherichia coli, and Shigella have a potent acid resistant systems that are able to withstand at low pH at around 2 for at least 2 h [330, 331]. E. coli possesses a level of acid resistance rivaling that of gastric pathogen Helicobacter Pylori [332]. As such, it is quite important to understand the cell metabolism in relation to acidic condition from both medical and fermentation points of view. The molecular and physiological response to acid stress has been thus the subject of intense investigation (Table 3) [333, 334].
Table 3.
Regulators involved in regulating glutamate-dependent acid resistance.
| Protein | Descriptor | Function in acid resistance |
|---|---|---|
| RpoD | σ 70 | Transcription of gadA/BC |
| RpoS | σ 38 | Transcription of gadX |
| EvgAS | Two-component signal transduction | Activates ydeO and gadE transcription |
| YdeO | AraC-like regulator | Activates gadE transcription |
| GadE | LuxR-related activator | Required for acid resistance, binds to Gad box, activates transcription of gadA/BC, autoactivates gadE, and represses ydeO |
| GadX | AraC-like regulator | Activates gadE, coactivates gadA/BC, and represses gadW |
| GadW | AraC-like regulator | Inhibits RpoS production, activates gadE, can coactivate gadA/BC at pH 8 |
| Crp | cAMP receptor protein | Inhibits RpoS production |
| TrmE | Era-like GTPase | Activates gadE mRNA production and stimulates translation of gadA and gadB mRNA |
| HNS | Histone-like protein | Negative regulator |
| TorR | Response regulator of TMAO reductase | Negative regulator of gadE |
[333].
Several acid stress response systems that can protect E. coli from acidic condition have been investigated [333–336]. Some of these depend on the available extracellular amino acids such as glutamate, arginine, and lysine, where the intracellular proton is consumed by the reductive decarboxylation of the amino acid followed by the excretion of the product such as γ-amino butyric acid (GABA) from cytoplasm to the periplasm by a dedicated antiporter that also imports the original amino acid [333] (Figure 25). E. coli cells have been demonstrated to exhibit acid resistance by such genes as gadAB which encodes glutamate decarboxylase and gadC which encodes glutamate/GABA antiporter. Glutamate decarboxylase production has been shown to increase in response to acid, osmotic, and stationary phase signals. The gadA and gadB genes for glutamate decarboxylase isozymes form a glutamate-dependent acid response system, where the process of decarboxylation consumes an intracellular proton and helps maintain pH homeostasis. It has also been known that there exists similar acid resistant systems for the case of using arginine instead of glutamate by arginine decarboxylase, where the antiporter is AdiC in this case [330, 331, 337, 338], and for the case of using lysine by lysine decarboxylase [338]. Note that the cells grown in media rich with amino acids such as LB are acid resistant [333].
Figure 25.
The role of glutamate decarboxylase for acid resistance.
In the typical batch culture, organic acids are most accumulated at the late growth phase or the stationary phase, and it has been known that GadA and GadB proteins increase in response to stationary phase and low pH [339]. The sigma factor σ S or RpoS which increases its amount at the late growth phase and the stationary phase as well as Crp are involved in acid resistance [330, 333]. As would be implied by the involvement of Crp, the resistance system is repressed when glucose is present. Moreover, it has been known that FoF1 proton-translocating ATPase is involved in this system [336]. The FoF1 ATPase is utilized for the protons in the periplasm move into the cytosol across the cell membrane producing ATP from ADP and Pi by the negative proton motive force (PMF) [340]. Since the basic problem of acid stress is the accumulated proton in the cytosol, this proton can be pumped out through FoF1 ATPase by hydrolyzing ATP and reversed proton move due to positive PMF at low pH such as pH 2 or 3 [336]. Without amino acid available in the media, this acid response system is activated by utilizing FoF1 ATPase [335, 341], where the positive proton motive force (PMF) pumps extra protons (H+) from the cytoplasm with consumption of ATP [336]. Namely, PMF is operated in the reverse direction as compared to the case of producing ATP.
Table 2 shows 11 regulators involved in regulating glutamate-dependent acid resistance. In the typical batch culture, the gadA/BC loci can be induced during growth in acidic minimal media (pH 5.5) or in the stationary phase regardless of pH [333]. However, in a complex media such as LB, locus is not induced until the culture enter into stationary phase.
Table 2.
Effect of dilution rate on fermentation characteristics of wild type E. coli.
| Dilution rate (h−1) | Biomass conc. (g/L) | Specific glucose uptake rate (mmol/g/h) | Specific acetate formation rate (mmol/g/h) | Biomass yield (g/g) | Specific CER (mmol/g/h) |
|---|---|---|---|---|---|
| 0.2 | 1.45 ± 0.06 | 3.07 ± 0.13 | ND∗ | 0.37 ± 0.015 | 9.15 |
| 0.4 | 1.87 ± 0.09 | 4.75 ± 0.23 | 0.01 | 0.47 ± 0.023 | 11.61 |
| 0.6 | 2.0 ± 0.09 | 6.67 ± 0.3 | 0.88 ± 0.04 | 0.5 ± 0.023 | 13.17 |
| 0.7 | 1.93 ± 0.08 | 8.05 ± 0.34 | 1.33 ± 0.06 | 0.48 ± 0.02 | 15.83 |
∗ND: not detectable, where glucose detectable limit is 0.038 g/L.
The expressions of gadA/BC genes are under control of GadE and the response regulator RcsB [342], where RcsB is part of the RcsCDB phosphorelay, a signal transduction system conserved in the members of Enterobacteriaceae. The RcsB can be also activated independently of the phosphorelay, by binding of different coregulators such as RcsA (main one), RmpA, TviA, and PhoP [342]. As shown in Figure 26 [343], EvgS is a sensor kinase and phosphorylates EvgA, where the phosphorylated EvgA activates gadE gene as well as ydeO, where YdeO also regulates gadE gene. It has also been shown that small membrane protein B1500 connects the signal transduction cascade between EvgS/EvgA and PhoQ/PhoP, where b1500 is located upstream of ydeO and under control of EvgA [344]. It has been known that Mg2+ turns off the PhoQ/PhoP system [345]. Moreover, the phosphorylated PhoP activates gadW, where GadW activates hdeA and gadA, where hdeA is under the control of GadW, PhoP-P, and GadW. EvgA regulates at least eight genes related to acid resistance such as ydeP, b1500, and ydeO [346].
Figure 26.

Acid resistance mechanism under acidic condition.
It has been shown that an acid pH lowers cAMP levels in exponentially growing cells in the minimal glucose medium. This may elevate RpoS that would drive increased expression of gadX. However, GadW represses RpoS synthesis at acidic condition, and in turn GadX synthesis. GadX, when not repressed by GadW, is acid induced due to changes in cAMP. GadW is also acid induced when it is not repressed by GadX. GadX directly binds to the gadW promoter region. GadX and GadW collaborate to repress gadA and gadBC expressions under alkaline conditions [347]. The GadX-GadW regulon has also been investigated by DNA microarray [348].
As explained before, the two-component system of EnvZ (sensor) and OmpR (regulator) regulates porin expression, where OmpR may be a key regulator for acid adaptation, and thus opmR mutant is sensitive to acid exposure [334]. It has been known that the level of OmpC increases with the increased osmolarity when cells are growing in neutral or alkaline media, whereas the level of OmpF decreases at high osmolarity [349]. It has also been known that these porin proteins play important roles at acidic condition.
The acid-inducible asr gene is reported to be regulated by the two-component systems PhoR/B which controls pho regulon in response to phosphate starvation, and thus PhoB-PhoR deletion mutant fails to induce asr gene expression [350]. It has also been suggested that H+ might activate a sensor protein PhoR in the periplasm [350] directly or via its acceptor.
Under anaerobic condition, additional genes like ackA, lpdA, and so forth, as well as hdeA and ompT, and so forth, are induced. In order to avoid deleterious concentration in the cell caused by the production at low pH, ldhA is induced by acid in order to produce lactate instead of more harmful acetate plus formate [351].
9. Heat Shock Response
The organisms respond to a sudden temperature upshift by increasing the synthesis of a set of proteins. This phenomenon is called the heat shock response. The research on heat shock response of a microorganism contributes to the variety of practical applications such as temperature-induced heterologous protein production [352, 353] and simultaneous saccharification and fermentation (SSF) [354]. The heat shock proteins play roles in the assembly and disassembly of macromolecular complex such as GroE [355], the intracellular transport such as Hsp70 [356], transcription such as σ 70 [357], proteolysis such as Lon [358], and translation such as lysyl-tRNA synthetase [359]. The heat shock response in E. coli is mediated by Eσ 32 [360], and it has been known that at least 26 genes are induced by heat shock [361], where E denotes the RNA polymerase holoenzyme. Among them, groEL, dnaK, and htpG are the genes which code for the major chaperones such as Hsp 60, Hsp 70, and Hsp 90. The ClpP, Lon, and HtrC are involved in the proteolysis. DnaK, DnaJ, GrpE, and RpoH are involved in the autoregulation of heat shock response [362–365]. It has been known that DnaK prevents the formation of inclusion bodies by reducing aggregation and promotion of proteolysis of misfolded proteins [366]. A bichaperone system involving DnaK and ClpB mediates the solubilization or disaggregation of proteins [367]. GroEL operates protein transit between soluble and insoluble protein fractions and participates positively in disaggregation and inclusion body formation. Small heat shock proteins such as IbpA and IbpB protect heat-denatured proteins from irreversible aggregation and have been found to be associated with inclusion bodies [368, 369].
It has been reported on the molecular mechanisms of heat shock proteins [370, 371]. Hoffmann et al. [353] investigated the metabolic adaptation of E. coli during temperature-induced recombinant protein production and showed that cAMP/Crp-controlled LpdA of the pyruvate dehydrogenase complex (PDHc) and SdhA in the TCA cycle are induced four times, reaching a maximum at 1 h after temperature upshift. It is also shown that the TCA cycle enzymes such as ICDH and MDH are initially less produced but regained to their respective preshift values about 30 min after the temperature upshift. Gadgil et al. [372] investigated the effect of temperature downshift from 37°C to 33 and 28°C on gene expressions in E. coli. This kind of investigation is useful in analyzing the metabolic changes and investigating the effects of gene modification for strain improvement [373].
Upon temperature upshift from 37°C to 42°C, the expression of rpoH is upregulated, and the expressions of dnaK, groL, groS, htpG, and ibpB were upregulated [374], which are known to be under control of sigma factor (σ 32). It was also shown that arcA gene expression was upregulated, and that the expression of crp gene was upregulated, and the expression of lpdA, which is under control of Crp, was also upregulated [353, 374].
To survive, cells have to control gene expressions precisely in response to the changes in the growth environment. The microorganism such as E. coli attains this primarily at the transcription level. To control the initiation of the specific transcription, the cell uses diverse mechanisms including various sigma factors. The classical heat shock regulon has been shown to be under the control of σ 32 transcription factor, the product of rpoH gene [375]. The regulation of the sigma factor (σ 32) is complex and depends on the feedback control loops involving the dnaK chaperone and temperature-induced changes in mRNA secondary structure [376]. The relative levels of the major heat shock genes such as dnaK, groS, groL, ibpB, lonA, and htpG were upregulated after the temperature upshift. The expressions of heat shock genes such as dnaK, groL, and ibpB increased in the early induction phase (first 10–20 minutes) and then declined. In E. coli, heat shock protein synthesis rates peak at about 5~10 min after the temperature upshift and then declined to a new steady-state levels [377]. The heat shock response is made transcriptionally, where it has been known that the RNA polymerase core (E) binds to new initiation subunit σ 32 [378], and the resulting holoenzyme Eσ 32 transcribes only heat shock genes [379], which have promoter sequences that differ from those transcribed by E plus σ 70, the normal vegetative initiation factor [380]. The transcription factor σ 70 is itself a heat shock protein, and the increase in its concentration after heat shock may contribute to its decline in heat shock protein synthesis. Moreover, other heat shock proteins, in particular the dnaK gene product contributes to the shutoff, since the mutations in their genes prolong the high level synthesis of heat shock proteins [381]. The heat shock response must be tightly regulated in order to allow rapid changes in heat shock protein synthesis rates. Although the level of mRNA transcribed from the rpoH gene increases after heat shock, their increases may be insufficient and too slow to be the sole explanation of the rapid effect of the heat shock. It has been shown that the concentration of active σ 32 limits the expression of heat shock genes, and that the stability of σ 32 is modulated [377].
Because of the rapid turnover (half-life of less than 1 min), the cellular concentration of σ 32 is very low at normal temperature and is limited for the transcription of the heat shock gene. Upon temperature upshift, σ 32 becomes transiently stabilized until the beginning of the shutoff phase of the heat shock response. The heat shock response is induced as a consequence of declining σ 32 levels and inhibition of σ 32 activity. Stress-dependent changes in heat shock gene are mediated by the antagonistic action of σ 32 and negative modulators which act upon σ 32. These modulators are the DnaK chaperone system which inactivates σ 32 by direct association and mediates its degradation by proteases [382]. Degradation of σ 32 is mediated mainly by FtsH and ATP dependent metalloprotease within the inner membrane. The heat shock proteins increased immediately after the temperature upshift, reached a maximum 5–15 min later and decreased to preshift values largely within 1 h, while the maximum induction of many heat-shock proteins including DnaK and HtpG reached at least 30 min later.
The cyclic AMP (cAMP) receptor protein Crp activates transcription for more than 100 promoters. When bound to its allosteric effector cAMP, the Crp homodimer binds to the specific DNA sites near target promoters, enhancing the binding of RNA polymerase holoenzyme (RNAP) and facilitating the initiation of the transcription. It has been shown that crp gene expression increased, and lpdA gene expression followed the similar pattern upon heat shock. It was shown that mlc gene expression followed the same pattern as that of rpoH upon heat shock, which confirms that Eσ 32 is involved in the expression of mlc gene. It has been shown that Eσ 32 plays an important role in balancing the relative concentration of Mlc and EIICB in response to the availability of glucose in order to maintain inducibility of Mlc regulon at higher temperature [383]. When Mlc is overproduced it has been known to reduce acetate accumulation [384] and causes slow growth but gives better performance for recombinant protein production [385]. Mlc is a global regulator of carbohydrate metabolism, and regulates the expression of pts operon. Mlc represses manXYZ encoding enzyme II of the mannose PTS [386], malT encoding the activator of maltose operon, and mlc itself negatively [387]. Moreover, ptsG encoding enzyme IICB of the glucose PTS (EIICBglc) and the pts operon encoding general PTS proteins are also known to be repressed by Mlc [388, 389]. The mlc promoter is very weak because nucleotide sequence of −10 region of the promoter differs from the consensus sequence of the strong promoter of E. coli. In addition, Mlc expression is autoregulated by Mlc itself. Therefore, the intracellular concentration of Mlc is limited in E. coli [390]. The mlc gene has been known to be transcribed by two promoters, P1 and P2, and has a binding site of its own gene product. It has been shown by in vitro transcription assays of mlc gene that P2 promoter could be recognized by RNA polymerase containing the heat shock sigma factor σ 32 (Eσ 32) as well as Eσ 70, while P1 promoter is only recognized by Eσ 70. The overall regulation mechanism against heat shock may be expressed as Figure 27 [374].
Figure 27.
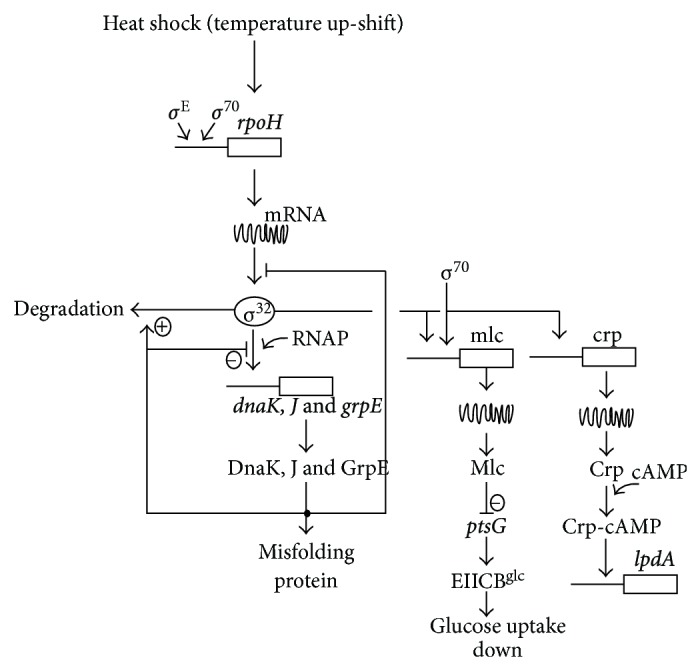
Effect of heat shock on gene and protein expressions and the fermentation characteristics.
Let us consider the production mechanism of acetate at higher temperature. In the typical batch cultivation, the cells must switch efficiently from the rapid growth on a favored carbon source such as glucose to a much slower growth on the excreted by-products such as acetate. Acetate excretion occurs through the Pta-Ack pathway, or may possibly by Pox pathway. Acetate utilization occurs through AcCoA synthetase (Acs). This high-affinity acetate-scavenging enzyme converts acetate to AcCoA, where cells introduce it into the TCA cycle to generate energy and/or the glyoxylate pathway to build cell constituents. The higher expression of acs accelerates acetate assimilation in the presence of acetate [391, 392], which leads to the activation of glyoxylate pathway. Transcription occurs from two σ 70-dependent promoters such as the distal promoter acs P1 and proximal promoter acs P2 [391, 393]. While multiple factors influence transcription, Crp appears to function directly as the critical transcription factor. Cells control this acetate switch primarily by controlling the initiation of acs transcription from the major promoter acs P2 [391, 394]. Activation of acs transcription depends on cAMP-Crp. The cAMP-Crpbinds two sites within the acs regulatory region. However, it has been shown that Fis and Ihf independently modulate Crp-dependent activation of acs P2 transcription [395], and the mechanism is not so simple. As such, the activation of crp may cause acs to be upregulated. The acs gene is also under control of rpoS. It has been shown that acs is expressed in an rpoS-dependent manner during different phases of the growth [396].
Although cellular ATP may increase for short period after the temperature upshift in E. coli [397], it eventually decreases at higher temperature [353, 397]. It has also been reported that the specific CO2 production rate as well as O2 consumption rate increased upon temperature upshift [353, 397, 398]. As a result, the cell yield decreased and the cell maintenance increased [353, 399]. Although it has been reported that the TCA cycle flux increased upon temperature upshift at the specific growth rate of 0.08 h−1 [397], another investigation based on 13C-labeled experiment indicates that the TCA cycle flux became low at the dilution rates of 0.45 and 0.32 h−1 [400].
It has been reported that the respiration is activated during the temperature upshift [353]. It may be due to the activation of cydB gene expression. The arcA showed increased expression after the temperature upshift (especially first 30 mins) and modulated the expressions of such genes as cydB, cyoA, and icdA. The up-regulation of arcA gene may not be the direct effect of heat shock but indirectly due to lower dissolved oxygen concentration caused by the lower solubility at higher temperature [397].
It has been reported that superoxide dismutase gene (sod) is induced in response to the oxidative stress imposed by dioxygen or by the redox active compounds such as viologens or quinones caused by the temperature upshift [401]. It has also been reported that the exposure of sodA/B null mutant E. coli to aerobic heat stress caused a profound loss of viability [399, 402]. Moreover, the sod gene is under control of SoxRS, where it becomes significant under dual osmotic and heat stresses [403].
10. Fatty Acid Metabolic Regulation
The acetate or fatty acid metabolism is also of practical interest. Here, consider this by looking at the effect of fadR gene knockout on the physiology of E. coli, where the parent and its fadR mutant are grown on glucose in a minimal medium under aerobic condition [299]. Compared to the parent strain, acetate production was reduced, whereas the biomass yield was enhanced in the fadR mutant. This cultivation phenomenon is similar to the result of Farmer and Liao [404]. The level of AceK, the bifunctional protein catalyzing phosphorylation/inactivation protein, was higher in the fadR mutant. Acetate-induced periplasmic transporter OppA showed lower expression level in the fadR mutant as compared to wild type [299]. Moreover, it was shown that Icl and MS, the two enzymes of the glyoxylate shunt, are significantly induced in the fadR mutant. CS, Acn, Fum, and MDH were coordinately upregulated to some extent. Moreover, NADP+-dependent malic enzyme (Mez) was upregulated whereas NAD+-dependent malic enzyme (Sfc) down-regulated in the fadR mutant as compared to wild type. In addition, Ppc showed lower activity in the fadR strain, whereas Pck activity increased. In the fermentative pathways, Ack activity is reduced in the fadR mutant as compared to the parent strain, which is in agreement with the decrease of acetate production in the fadR mutant. These trends indicate that the fadR mutant utilized the glyoxylate shunt for the replenishment of OAA for biosynthesis. The activation of the glyoxylate shunt bypassed the TCA cycle and thus prevented the loss of carbons as CO2 in ICDH and KGDH-catalyzed reactions.
Moreover, it was shown that PYR and AcCoA concentrations deceased, whereas the concentrations of isocitrate (ICIT), αKG, MAL, OAA, and aspartate (ASP) increased in the fadR mutant as compared to the parent strain [299]. These results are in agreement with those reported by van de Walle and Shiloach [405] who found that the operation of the glyoxylate shunt in E. coli BL21 resulted in accumulation of TCA cycle intermediates and higher biosynthesis fluxes. Similar to PYR, PEP concentration was also reduced in the fadR mutant. These variations reflected the action of the glyoxylate shunt in the fadR mutant. It is also observed that the intracellular concentrations of intermediates in the glycolysis and the PP pathway such as G6P and 6PG are reduced in the fadR mutant. Apart from these changes, the ratio of NADPH/NADP+ was lower, while that of NADH/NAD+ was higher in the fadR murtant as compared to wild-type strain.
It has been reported that FadR is a transcriptional regulator with a Helix-turn-Helix motif, regulating metabolic pathways, such as the fatty acid biosynthesis and degradation, and glyoxylate shunt and playing a possible role in the regulation of amino acid biosynthesis directly or indirectly [406–408]. The overall effect of the fadR mutant is illustrated in Figure 28 [299].
Figure 28.

Metabolic pathways showing levels of enzymes (or proteins) and intracellular metabolite concentrations in the fadR mutant E. coli relative to those in the parent at the exponential phase grown in glucose minimal medium under aerobic condition. The numbers beside the protein names and the metabolites represent the ratios. Shaded and boxed metabolites mean increased and decreased intracellular concentrations in fadR mutant E. coli.
Induction of the glyoxylate shunt led to the better utilization of AcCoA by increasing the carbon flow through this anaplerotic pathway, which is inferred from the significantly reduced intracellular concentration of AcCoA. The decrease of the intracellular AcCoA pool is therefore suggested to be responsible for the reduced acetate excretion in the fadR mutant. The pools of PEP and PYR are conjointly reduced in the fadR mutant due to the draining of carbon into the TCA cycle and the glyoxylate shunt. PEP is known to be a critical metabolite in E. coli. It involves not only in the PTS as a phosphoryl donor, but also in the regulation of many enzymes as an effecter [409]. It is, therefore, considered that the upregulations of the glycolytic enzymes, such as Pgi and Pfk in the fadR mutant are associated with the release from the inhibition due to the lower PEP concentration, since PEP is an inhibitor of both enzymes. Decrease of G6P concentration is responsible for the faster glucose uptake in the fadR mutant by the upregulation of PTS proteins, since G6P degrades the mRNA of PTS proteins by activating RNaseP enzyme [410]. Other glycolytic enzymes such as Fba, Tpi, GAPDH, and Pgk were concurrently upregulated in the fadR mutant to some extent to form more PEP and PYR, which are consistent with D'Alessio and Josse's results [411] that these enzymes are always regulated proportionally in E. coli.
Concomitant with the induction of the glyoxylate shunt, some of the TCA cycle enzymes, such as CS, Acn, Fum, and MDH, were coordinately upregulated. Besides, SucA, a component of KGDH, and FumA showed higher expression levels. These upregulations are expected to fulfill the role in driving the increased carbon flux due to the action of the glyoxylate shunt. It is reported that CS and Acn, but not ICDH, are regulated in a coordinate mode, which may be due to the fact that citrate is an activator of Acn [412]. However, ICDH activity is subject to phosphorylation/inactivation control at the branch point of isocitrate to force the carbon flux towards the glyoxylate shunt, whose activity was slightly lower in the fadR mutant as compared to the parent strain. The phosphorylation/inactivation of ICDH is exerted by AceK, which is induced in the fadR mutant. Decreased carbon flux via ICDH, therefore, restricted the production of NADPH in the TCA cycle, as shown by the fact that the NADPH concentration is much lower in the mutant than in the parent strain. NADPH is an important cofactor for biosynthesis and mainly formed in the TCA cycle. Up to 60% of the total NADPH is produced in the TCA cycle in the parent strain under aerobic condition [189]. To meet the need for biosynthesis, NADPH has to be generated by other NADPH-producing pathways. One way is through NADPH-dependent malic enzyme, Mez, which is upregulated in the fadR mutant. The up-regulation of Mez is probably related to the reduced intracellular AcCoA concentration as this enzyme is repressed by glucose and AcCoA [413]. This up-regulation also played a role in supplying AcCoA from MAL via PYR in the fadR mutant. On the other hand, the activity of Sfc, which consumes NADH and produces NAD+ by coupling the conversion of PYR to MAL, reduced due to the smaller pool of PYR in the fadR mutant.
Proteome analysis demonstrated that the protein expressions in amino acids biosynthesis, such as AsnB, TrpD, and AroG F1F2 proton-translocating ATPase such as AtpA, AtpC, and AtpD and the ribosomal protein RplI as well as the transcription elongation factor GreA were upregulated in the fadR mutant. These genes are clustered and show growth rate dependent expression [414]. On the contrary, the levels of AccB and fabD involved in fatty acids biosynthetic pathways are negatively affected by fadR mutant, which is consistent with previous studies that the E. coli FadR functions as a repressor of the fatty acid degradative (fad) pathways and can also act as an activator of unsaturated fatty acid synthesis (fab) [405, 415, 416]. In addition, DnaK, the heat shock protein and UspA, the universal stress protein, are induced in the fadR mutant. These proteins are known to protect cells from stressful conditions such as heat shock, starvation, and stress stimulons; thus fadR mutation seems to be a stress to the E. coli cell. The uspA is a member of the fadR regulon, and the transcription of uspA is derepressed during exponential growth in fadR null mutants [416]. Previous studies revealed that RpoS-regulated genes, periplasmic transporters for amino acids and peptides, and metabolic enzymes are induced either by acetate or at low pH [417, 418]. Of these, it is considered that the downregulation of OppA is related to the less accumulation of acetate in the fadR mutant compared to the parent strain.
11. Response to Nutrient Starvation and the Metabolic Regulation by RpoS
Although many industrial fermentations are conducted in the batch mode, most of the studies have focused on the cell growth phase, and little attention has been paid to the late growth and stationary phases. Since the important metabolites are produced at the early stationary or stationary phases, it is quite important to clarify the metabolic regulation that occurs during these phases. During batch fermentation, the cultural conditions change from glucose rich to acetate rich condition, and change further to the carbon-starved condition in E. coli. Several global regulators such as RpoD, SoxRS, Cra, FadR, and IclR may help E. coli to cope with different kinds of metabolic stresses. Apart from these regulators, RpoS, the master regulator of the stationary phase or stress-induced genes in E. coli regulates such genes as those for the carbohydrate PTS, crr, glycolytic pathway genes such as fbaB and pfkB, the acetate-forming gene poxB, the nonoxidative PP pathway genes such as talA and tktB, and TCAcycle genes such as acnA and fumC. In addition, some of the amino acid and fatty acid metabolic pathway genes such as argH, aroM, and yhgY and energy metabolism genes such as narY, appB, and ldcC have also been reported to be regulated in an rpoS-dependent manner [419–425].
E. coli cells are exposed to different stress conditions such as oxidative stress, acid stress or stresses from particular ion or carbon limitation at different phases of growth. Fortunately, E. coli cells possess several regulatory proteins, which through the regulation of a large number of genes help bacteria to cope with continuously changing environment under different stress conditions, including acid stress, or other stresses mentioned above [164]. Of these stress conditions, acid stress, particularly stress from acetate gives a problem for the growth in E. coli. In addition, acetic acid has been recognized to be a problem in recombinant protein production as it easily passes through the thin lipid layer of the bacterial cell wall and cause damage to the protein production [419, 426]. It has been reported that the stress regulatory protein RpoS regulates the expression of approximately 78 genes in E. coli during acid stress [419, 427].
In general, the bacterial culture medium is considered to be rich in carbohydrate such as glucose as the sole carbon source during the exponential growth phase. As the cell such as E. coli utilizes the glucose, acetate is produced as the major fermentative product under aerobic condition, and the cell exhibits a diauxic shift which causes a termination of the exponential growth phase and transition towards the stationary phase. Then, E. coli utilizes acetate as a carbon source during early stationary phase of growth. When acetate is used up, E. coli starts to utilize amino acids as carbon and nitrogen sources during stationary phase. The complex changes occurring among the major metabolic pathway enzymes, their respective genes and the intermediary metabolites, during a shift from carbon rich to carbon-limited conditions, have been a major topic of interest in the metabolic regulation analyses. RpoS is a global regulator that regulates the expressions of many genes at the onset of stationary phase or carbon-starved conditions and other stress conditions in E. coli [420, 423, 424].
RpoS is a sigma factor of RNA polymerase. It is known that the core RNA polymerase consists of four subunits such as two α, one β and one β′. Part of the RNA polymerase that recognizes the promoter-binding site is generally known as sigma factor (σ). Without the sigma factor, RNA polymerase remains inactive [428]. E. coli possesses seven different σ factors [428] as mentioned before. Depending on the environmental condition, different sigma factors bind with the RNA polymerase so that particular gene expressions are initiated. Of these seven different sigma factors, rpoS or σ 38 is important in bacterial metabolism as this transcription factor has been shown to be associated with different kinds of stresses in E. coli [420, 421, 428]. Unlike other regulators, expressions of which are stimulated by certain effector molecules, and these regulators then function by binding to the promoter sites of particular genes, where RpoS itself is a transcription factor, and regulates the expressions of genes at the transcriptional levels. However, once the transcription starts, the sigma factor dissociates from the RNA polymerase.
RpoS has been shown to stimulate the expressions of several oxidative stress response genes such as katE, katG, sodC, and dps and osmotic stress response genes such as osmE, and osmY. Strains lacking a functional rpoS gene also failed to express the genes for acid resistance such as gadA and gadB, near-UV resistance gene nuv, acid phosphatase genes appAR, and so forth [420, 421]. The intracellular level of RpoS itself is regulated by various mechanisms depending on the stress type and growth conditions. For example, rpoS transcription is stimulated by a reduction in growth rate, whereas translation is stimulated by osmotic shock, low temperature, or pH downshifts [420, 421, 427]. The third mechanism that controls the RpoS level is through proteolysis. While under normal situation, RpoS is rapidly degraded by ClpXP proteases, and the proteolytic activity of this enzyme is considerably reduced under stress conditions [420, 429, 430].
Although the roles of RpoS are originally described for various types of stress response, it has been demonstrated that the regulatory roles of rpoS are not restricted to the stress response genes only. In E. coli, RpoS-dependent genes are found all over the chromosome, whose function ranges from DNA repair and protein synthesis to the transport, biosynthesis and metabolism of sugars, amino acids, and fatty acids. Notably, RpoS is found to regulate the expression of DNA repair enzymes such as the exonuclease encoded by xthA and the methyl transferase encoded by ada, the gene that determines the cell morphology such as bolA, the genes encoding transport, and binding proteins such as gabP, and ugpEC. In addition, a considerably large number of unknown proteins are invariably affected by rpoS mutation [422, 427]. Considering the wide range of activities of RpoS, it seems obvious that RpoS could have significant contribution in the maintenance of E. coli metabolic pathways at the stationary phase under carbon-starved conditions.
The complexity of the metabolic system of E. coli is exemplified by the fact that many metabolic pathway genes are found to be regulated by more than one global regulator. For example, icdA of the TCA cycle is regulated by RpoD and Cra, acnA and fumC of the TCA cycle are regulated by SoxRS and RpoS, aceA and aceB of the glyoxylate pathways are regulated by Cra and IclR, and so forth [319, 431–435]. Moreover, the metabolic pathway of E. coli consists of several genes that possess isogenes. These iso-genes are known to encode backup enzymes in response to certain environmental stimuli, and the expressions of these enzymes are often regulated by one or more of the global regulators. It appears that the stress inducible metabolic pathway genes constitute the functional units through which different global regulators coordinate metabolic activities in the face of changing culture environment.
In E. coli, transketolase is encoded by tktA and tktB genes, and transaldolase is encoded by talA and talB genes, respectively. TktA and TalB are reported to be the major enzymes to catalyze transketolase and transaldolase reactions, and TktB and TalA are the minor enzymes of the nonoxidative PP pathway [436, 437]. The non-oxidative PP pathway is important as E4P, the important precursor of the aromatic amino acids, and as S7P, which is the cell wall components of E. coli, are synthesized only through the non-oxidative branch. E4P and S7P are produced through the consecutive reactions catalyzed by Tkt and Tal [436, 438]. While the physiological roles of the major and minor enzymes have been elucidated, the reports on the positive regulation of the minor transketolase (encoded by tktB) and transaldolase (encoded by talA) of the non-oxidative PP pathway by the stress regulator RpoS indicates that these genes might play significant role at the carbon-starved conditions [422, 427, 439].
On the other hand, the TCA cycle genes such as fumC and acnA encode FumC and AcnA enzymes of the TCA cycle, respectively. While fumarase catalyzes the conversion of FUM to MAL by the hydration reaction, Acn catalyzes the conversion of CIT to cis-aconitate to ICIT through dehydration and hydration reactions [440]. However, both FumC and AcnA have iso-enzymes such as FumA and AcnB, which are encoded by the genes fumA and acnB, respectively. It has been reported that FumA and AcnB play the major roles of fumarase and aconitase when the cell grows under optimum growth condition. Both FumA and AcnB possess Fe-S clusters that make these enzymes vulnerable to oxidative stress, and under such condition, FumC and AcnA play as back-up enzymes [432, 434, 435, 441]. It has been reported that RpoS regulates the expressions of fumC and acnA at the stationary phase of growth. Note that it is also reported that the expressions of both fumC and acnA genes are also regulated by the oxidative stress regulators SoxRS as well [285, 442]. In summary, RpoS plays important roles at the late growth phase and the stationary phase [195, 396].
In order to determine how the cell cope with the absence of a vital gene like rpoS, it may be identified such genes that were upregulated at the early stationary phase from the microarray data [425]. A total of 208 genes were upregulated in the mutant excluding the genes for hypothetical proteins. Among these genes, 25 genes (~12%) were upregulated in both phases of growth. Microarray data revealed that, of the central metabolic pathways, significant reduction of the expression was observed for several genes during early stationary phase, with the exception of fumC, which was upregulated. The down-regulation of several genes such as glgA and glgS involved in glycogen synthesis is also an indicator of the bacterial adaptive response to carbon-limited conditions in the absence of rpoS background.
As mentioned earlier, apart from the TCA cycle activity, accumulation of acetate throughout the cultivation period was another notable feature of the rpoS mutant [425]. While two genes, ackA and poxB, involved in acetate production are down-regulated in the mutant at the early stationary phase, microarray data [425] indicate that acetate production could be stimulated by the up-regulation of L-cysteine biosynthesis genes such as cysD, cysE, and cysK, catalyzing the reactions that generate acetate as a by-product [164]. Moreover, enzymes involved in the acetate catabolism such as AcCoA synthetase encoded by acs, the glyoxylate shunt enzymes encoded by the aceBAK operon, and the TCA cycle genes such as gltA, mdh, and sdhC were down-regulated during the stationary phase [424, 443, 444].
Down-regulation of acs during the early stationary phase resulted in a decrease in the intracellular pool of AcCoA in the mutant compared to the wild-type strain. While the major route for AcCoA formation was less expressed, the other pathways for AcCoA formation rely on fatty acid degradation pathway [164]. Several genes that participate in β-oxidation of fatty acids, particularly fadA and fadB are significantly upregulated, and the fatty acid biosynthesis genes such as accB, accC, accD, and fabF were also upregulated in the mutant [425]. The expression of the fatty acid degradation regulator, fadR, was significantly high at the early stationary phase of growth. It is known that FadR regulates fatty acid metabolism by binding to the DNA that contains fadB promoter binding sites, and in this way fadR controls fatty acid metabolism [445, 446]. Down-regulations of aceA and aceB genes correspond to the higher expression of fadR, where FadR represses the glyoxylate shunt encoded by aceBAK by directly regulating the activation of the glyoxylate shunt repressor, iclR [405, 447]. The higher expressions of fadR and iclR also caused acetate accumulation.
12. Conclusions
It was shown in the present paper that a variety of regulation mechanisms are present in response to the changes in culture environment, where the appropriate global regulators or transcription factors together with sigma factors play important roles. Although the findings on the metabolic regulation mechanisms so far are only a tip of ice verg, it is a grand challenge to uncover the overall regulation mechanism in particular by systems biology approach to understand the mystery of how and why the living organisms are so well and efficiently organized for survival.
References
- 1.Caetano-Anollés G., Yafremava L. S., Gee H., Caetano-Anollés D., Kim H. S., Mittenthal J. E. The origin and evolution of modern metabolism. International Journal of Biochemistry and Cell Biology. 2009;41(2):285–297. doi: 10.1016/j.biocel.2008.08.022. [DOI] [PubMed] [Google Scholar]
- 2.Janga S. C., Salgado H., Martínez-Antonio A., Collado-Vides J. Coordination logic of the sensing machinery in the transcriptional regulatory network of Escherichia coli . Nucleic Acids Research. 2007;35(20):6963–6972. doi: 10.1093/nar/gkm743. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Gama-Castro S., Jiménez-Jacinto V., Peralta-Gil M., et al. RegulonDB (version 6.0): gene regulation model of Escherichia coli K-12 beyond transcription, active (experimental) annotated promoters and Textpresso navigation. Nucleic Acids Research. 2008;36(1):D120–D124. doi: 10.1093/nar/gkm994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Heinemann M., Sauer U. Systems biology of microbial metabolism. Current Opinion in Microbiology. 2010;13(3):337–343. doi: 10.1016/j.mib.2010.02.005. [DOI] [PubMed] [Google Scholar]
- 5.Feist A. M., Palsson B. Ø. The growing scope of applications of genome-scale metabolic reconstructions using Escherichia coli . Nature Biotechnology. 2008;26(6):659–667. doi: 10.1038/nbt1401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Palsson B. Metabolic systems biology. FEBS Letters. 2009;583(24):3900–3904. doi: 10.1016/j.febslet.2009.09.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Harbison C. T., Gordon D. B., Lee T. I., et al. Transcriptional regulatory code of a eukaryotic genome. Nature. 2004;430(7004):99–104. doi: 10.1038/nature02800. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Luscombe N. M., Babu M. M., Yu H., Snyder M., Teichmann S. A., Gerstein M. Genomic analysis of regulatory network dynamics reveals large topological changes. Nature. 2004;431(7006):308–312. doi: 10.1038/nature02782. [DOI] [PubMed] [Google Scholar]
- 9.Balázsi G., Barabási A. L., Oltvai Z. N. Topological units of environmental signal processing in the transcriptional regulatory network of Escherichia coli . Proceedings of the National Academy of Sciences of the United States of America. 2005;102(22):7841–7846. doi: 10.1073/pnas.0500365102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Moxley J. F., Jewett M. C., Antoniewicz M. R., et al. Linking high-resolution metabolic flux phenotypes and transcriptional regulation in yeast modulated by the global regulator Gcn4p. Proceedings of the National Academy of Sciences of the United States of America. 2009;106(16):6477–6482. doi: 10.1073/pnas.0811091106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Sauer U. Metabolic networks in motion: 13C-based flux analysis. Molecular Systems Biology. 2006;2, article 62 doi: 10.1038/msb4100109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.van Rijsewijk B. R. B. H., Nanchen A., Nallet S., Kleijn R. J., Sauer U. Large-scale 13C-flux analysis reveals distinct transcriptional control of respiratory and fermentative metabolism in Escherichia coli . Molecular Systems Biology. 2011;7, article 477 doi: 10.1038/msb.2011.9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Zamboni N., Fendt S.-M., Ruhl M., Sauer U. 13C-based metabolic flux analysis. Nature Protocols. 2009;4(6):878–892. doi: 10.1038/nprot.2009.58. [DOI] [PubMed] [Google Scholar]
- 14.Wittman C. Fluome analysis using GC-MS. Microbial Cell Factories. 2007;6:1–17. doi: 10.1186/1475-2859-6-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Shimizu K. Metabolic flux analysis based on 13C-labeling experiments and integration of the information with gene and protein expression patterns. Advances in Biochemical Engineering/Biotechnology. 2004;91:1–49. doi: 10.1007/b94204. [DOI] [PubMed] [Google Scholar]
- 16.Shimizu K. Toward systematic metabolic engineering based on the analysis of metabolic regulation by the integration of different levels of information. Biochemical Engineering Journal. 2009;46(3):235–251. doi: 10.1016/j.bej.2009.06.006. [DOI] [Google Scholar]
- 17.Shimizu K. Bacterial Cellular Metabolic Systems. Oxford, UK: Woodhead Publishing; 2012. [Google Scholar]
- 18.Fendt S. M., Buescher J. M., Rudroff F., Picotti P., Zamboni N., Sauer U. Tradeoff between enzyme and metabolite efficiency maintains metabolic homeostasis upon perturbations in enzyme capacity. Molecular Systems Biology. 2010;6, article 356 doi: 10.1038/msb.2010.11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Ishii N., Nakahigashi K., Baba T., et al. Multiple high-throughput analyses monitor the response of E. coli to perturbations. Science. 2007;316(5824):593–597. doi: 10.1126/science.1132067. [DOI] [PubMed] [Google Scholar]
- 20.Zamboni N., Heinemann M., Sauer U. Getting closer to the whole picture. Science. 2007;316(5824):550–551. doi: 10.1126/science.1142502. [DOI] [PubMed] [Google Scholar]
- 21.Barabasi A. L., Oltvai Z. N. Network topology: understanding the cell’s functional organization. Nature Reviews Genetics. 2004;5:101–113. doi: 10.1038/nrg1272. [DOI] [PubMed] [Google Scholar]
- 22.Babu M. M., Luscombe N. M., Aravind L., Gerstein M., Teichmann S. A. Structure and evolution of transcriptional regulatory networks. Current Opinion in Structural Biology. 2004;14(3):283–291. doi: 10.1016/j.sbi.2004.05.004. [DOI] [PubMed] [Google Scholar]
- 23.Salgado H., Gama-Castro S., Peralta-Gil M., et al. RegulonDB (version 5.0): Escherichia coli K-12 transcriptional regulatory network, operon organization, and growth conditions. Nucleic Acids Research. 2006;34:D394–D397. doi: 10.1093/nar/gkj156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Seshasayee A. S., Bertone P., Fraser G. M., Luscombe N. M. Transcriptional regulatory networks in bacteria: from input signals to output responses. Current Opinion in Microbiology. 2006;9(5):511–519. doi: 10.1016/j.mib.2006.08.007. [DOI] [PubMed] [Google Scholar]
- 25.Martínez-Antonio A., Janga S. C., Salgado H., Collado-Vides J. Internal-sensing machinery directs the activity of the regulatory network in Escherichia coli . Trends in Microbiology. 2006;14(1):22–27. doi: 10.1016/j.tim.2005.11.002. [DOI] [PubMed] [Google Scholar]
- 26.Kanehisa M., Goto S., Hattori M., et al. From genomics to chemical genomics: new developments in KEGG. Nucleic Acids Research. 2006;34:D354–D357. doi: 10.1093/nar/gkj102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Yamamoto K., Hirao K., Oshima T., Aiba H., Utsumi R., Ishihama A. Functional characterization in vitro of all two-component signal transduction systems from Escherichia coli . The Journal of Biological Chemistry. 2005;280(2):1448–1456. doi: 10.1074/jbc.M410104200. [DOI] [PubMed] [Google Scholar]
- 28.Timmermans J., van Melderen L. Post-transcriptional global regulation by CsrA in bacteria. Cellular and Molecular Life Sciences. 2010;67(17):2897–2908. doi: 10.1007/s00018-010-0381-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Kroos L. The Bacillus and Myxococcus developmental networks and their transcriptional regulators. Annual Review of Genetics. 2007;41:13–39. doi: 10.1146/annurev.genet.41.110306.130400. [DOI] [PubMed] [Google Scholar]
- 30.Brückner R., Titgemeyer F. Carbon catabolite repression in bacteria: choice of the carbon source and autoregulatory limitation of sugar utilization. FEMS Microbiology Letters. 2002;209(2):141–148. doi: 10.1016/S0378-1097(02)00559-1. [DOI] [PubMed] [Google Scholar]
- 31.Gutierrez-Ríos R. M., Freyre-Gonzalez J. A., Resendis O., Collado-Vides J., Saier M., Gosset G. Identification of regulatory network topological units coordinating the genome-wide transcriptional response to glucose in Escherichia coli . BMC Microbiology. 2007;7, article 53 doi: 10.1186/1471-2180-7-53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.de Lay N., Gottesman S. The crp-activated small noncoding regulatory RNA CyaR (RyeE) links nutritional status to group behavior. Journal of Bacteriology. 2009;191(2):461–476. doi: 10.1128/JB.01157-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Müller C. M., Åberg A., Straseviçiene J., Emody L., Uhlin B. E., Balsalobre C. Type 1 fimbriae, a colonization factor of uropathogenic Escherichia coli, are controlled by the metabolic sensor CRP-cAMP. PLoS Pathogens. 2009;5(2) doi: 10.1371/journal.ppat.1000303.e1000303 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Zheng D., Constantinidou C., Hobman J. L., Minchin S. D. Identification of the CRP regulon using in vitro and in vivo transcriptional profiling. Nucleic Acids Research. 2004;32(19):5874–5893. doi: 10.1093/nar/gkh908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Gottesman S. Micros for microbes: non-coding regulatory RNAs in bacteria. Trends in Genetics. 2005;21(7):399–404. doi: 10.1016/j.tig.2005.05.008. [DOI] [PubMed] [Google Scholar]
- 36.Vanderpool C. K., Gottesman S. Involvement of a novel transcriptional activator and small RNA in post-transcriptional regulation of the glucose phosphoenolpyruvate phosphotransferase system. Molecular Microbiology. 2004;54(4):1076–1089. doi: 10.1111/j.1365-2958.2004.04348.x. [DOI] [PubMed] [Google Scholar]
- 37.Wadler C. S., Vanderpool C. K. A dual function for a bacterial small RNA: SgrS performs base pairing-dependent regulation and encodes a functional polypeptide. Proceedings of the National Academy of Sciences of the United States of America. 2007;104(51):20454–20459. doi: 10.1073/pnas.0708102104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Majdalani N., Vanderpool C. K., Gottesman S. Bacterial small RNA regulators. Critical Reviews in Biochemistry and Molecular Biology. 2005;40(2):93–113. doi: 10.1080/10409230590918702. [DOI] [PubMed] [Google Scholar]
- 39.Babitzke P., Romeo T. CsrB sRNA family: sequestration of RNA-binding regulatory proteins. Current Opinion in Microbiology. 2007;10(2):156–163. doi: 10.1016/j.mib.2007.03.007. [DOI] [PubMed] [Google Scholar]
- 40.Nikaido H., Nakae T. The outer membrane of gram-negative bacteria. Advances in Microbial Physiology. 1980;20:163–250. doi: 10.1016/S0065-2911(08)60208-8. [DOI] [PubMed] [Google Scholar]
- 41.de la Cruz M. A., Fernández-Mora M., Guadarrama C., et al. LeuO antagonizes H-NS and StpA-dependent repression in Salmonella enterica ompS1 . Molecular Microbiology. 2007;66(3):727–743. doi: 10.1111/j.1365-2958.2007.05958.x. [DOI] [PubMed] [Google Scholar]
- 42.Nikaido H. Molecular basis of bacterial outer membrane permeability revisited. Microbiology and Molecular Biology Reviews. 2003;67(4):593–656. doi: 10.1128/MMBR.67.4.593-656.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Saier M. H., Tran C. V., Barabote R. D. TCDB: the transporter classification database for membrane transport protein analyses and information. Nucleic Acids Research. 2006;34:D181–D186. doi: 10.1093/nar/gkj001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Saier M. H., Yen M. R., Noto K., Tamang D. G., Elkan C. The transporter classification database: recent advances. Nucleic Acids Research. 2009;37(1):D274–D278. doi: 10.1093/nar/gkn862. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Nikaido H. Outer Membrane. Washington, DC, USA: ASM Press; 1996. edited by F. C. Neidhardt. [Google Scholar]
- 46.Hall M. N., Silhavy T. J. The ompB locus and the regulation of the major outer membrane porin proteins of Escherichia coli K12. Journal of Molecular Biology. 1981;146(1):23–43. doi: 10.1016/0022-2836(81)90364-8. [DOI] [PubMed] [Google Scholar]
- 47.Lugtenberg B., Peters R., Bernheimer H., Berendsen W. Influence of cultural conditions and mutations on the composition of the outer membrane proteins of Escherichia coli . Molecular and General Genetics. 1976;147(3):251–262. doi: 10.1007/BF00582876. [DOI] [PubMed] [Google Scholar]
- 48.Pratt L. A., Silhavy T. J. The response regulator SprE controls the stability of RpoS. Proceedings of the National Academy of Sciences of the United States of America. 1996;93(6):2488–2492. doi: 10.1073/pnas.93.6.2488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Nikaido H., Vaara M. Molecular basis of bacterial outer membrane permeability. Microbiological Reviews. 1985;49(1):1–32. doi: 10.1128/mr.49.1.1-32.1985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Death A., Ferenci T. Between feast and famine: endogenous inducer synthesis in the adaptation of Escherichia coli to growth with limiting carbohydrates. Journal of Bacteriology. 1994;176(16):5101–5107. doi: 10.1128/jb.176.16.5101-5107.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Nikaido H., Rosenberg E. Y. Porin channels in Escherichia coli: studies with liposomes reconstituted from purified proteins. Journal of Bacteriology. 1983;153(1):241–252. doi: 10.1128/jb.153.1.241-252.1983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.von Meyenburg K., Nikaido H. Outer membrane of gram-negative bacteria. XVII. Specificity of transport process catalyzed by the λ-receptor protein in Escherichia coli . Biochemical and Biophysical Research Communications. 1977;78(3):1100–1107. doi: 10.1016/0006-291x(77)90534-4. [DOI] [PubMed] [Google Scholar]
- 53.Gosset G. Improvement of Escherichia coli production strains by modification of the phosphoenolpyruvate:sugar phosphotransferase system. Microbial Cell Factories. 2005;4, article 14 doi: 10.1186/1475-2859-4-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Brzostek K., Brzóstkowska M., Bukowska I., Karwicka E., Raczkowska A. OmpR negatively regulates expression of invasin in Yersinia enterocolitica . Microbiology. 2007;153(8):2416–2425. doi: 10.1099/mic.0.2006/003202-0. [DOI] [PubMed] [Google Scholar]
- 55.Kruger W. M. Von, Lery L. M., Soares M. R., et al. The phosphate-starvation response in Vibrio cholerae O1 and phoB mutant under proteomic analysis: disclosing functions involved in adaptation, survival and virulence. Proteomics. 2006;6:1495–1511. doi: 10.1002/pmic.200500238. [DOI] [PubMed] [Google Scholar]
- 56.Ferrario M., Ernsting B. R., Borst D. W., Wiese D. E., II, Blumenthal R. M., Matthews R. G. The leucine-responsive regulatory protein of Escherichia coli negatively regulates transcription of ompC and micF and positively regulates translation of ompF . Journal of Bacteriology. 1995;177(1):103–113. doi: 10.1128/jb.177.1.103-113.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Delihas N., Forst S. MicF: an antisense RNA gene involved in response of Escherichia coli to global stress factors. Journal of Molecular Biology. 2001;313(1):1–12. doi: 10.1006/jmbi.2001.5029. [DOI] [PubMed] [Google Scholar]
- 58.Lee Y. H., Kim B. H., Kim J. H., Yoon W. S., Bang S. H., Park Y. K. CadC has a global translational effect during acid adaptation in Salmonella enterica serovar Typhimurium . Journal of Bacteriology. 2007;189:2417–2425. doi: 10.1128/JB.01277-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Santiviago C. A., Toro C. S., Hidalgo A. A., Youderian P., Mora G. C. Global regulation of the Salmonella enterica serovar Typhimurium major porin, OmpD. Journal of Bacteriology. 2003;185(19):5901–5905. doi: 10.1128/JB.185.19.5901-5905.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Miller V. L., Mekalanos J. J. A novel suicide vector and its use in construction of insertion mutations: osmoregulation of outer membrane proteins and virulence determinants in Vibrio cholerae requires toxR. Journal of Bacteriology. 1988;170(6):2575–2583. doi: 10.1128/jb.170.6.2575-2583.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Deighan P., Free A., Dorman C. J. A role for the Escherichia coli H-NS-like protein StpA in OmpF porin expression through modulation of micF RNA stability. Molecular Microbiology. 2000;38(1):126–139. doi: 10.1046/j.1365-2958.2000.02120.x. [DOI] [PubMed] [Google Scholar]
- 62.Fernández-Mora M., Puente J. L., Calva E. OmpR and LeuO Positively Regulate the Salmonella enterica Serovar Typhi ompS2 porin gene. Journal of Bacteriology. 2004;186(10):2909–2920. doi: 10.1128/JB.186.10.2909-2920.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Dorel C., Lejeune P., Rodrigue A. The Cpx system of Escherichia coli, a strategic signaling pathway for confronting adverse conditions and for settling biofilm communities? Research in Microbiology. 2006;157(4):306–314. doi: 10.1016/j.resmic.2005.12.003. [DOI] [PubMed] [Google Scholar]
- 64.Raivio T. L. Envelope stress responses and gram-negative bacterial pathogenesis. Molecular Microbiology. 2005;56(5):1119–1128. doi: 10.1111/j.1365-2958.2005.04625.x. [DOI] [PubMed] [Google Scholar]
- 65.Wanner B. L. Phosphorus assimilation and control of the phosphate regulon. In: Neidhardt F. C., Curtiss R I. I. I., Ingraham J. L., et al., editors. Escherichia coli and Salmonella: Cellular and Molecular Biology. Washington, DC, USA: ASM Press; 1996. pp. 1357–1381. [Google Scholar]
- 66.Calvo J. M., Matthews R. G. The leucine-responsive regulatory protein, a global regulator of metabolism in Escherichia coli . Microbiological Reviews. 1994;58(3):466–490. doi: 10.1128/mr.58.3.466-490.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Gallegos M. T., Schleif R., Bairoch A., Hofmann K., Ramos J. L. AraC/Xyls family of transcriptional regulators. Microbiology and Molecular Biology Reviews. 1997;61(4):393–410. doi: 10.1128/mmbr.61.4.393-410.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Balagué C., Véscovi E. G. Activation of multiple antibiotic resistance in uropathogenic Escherichia coli strains by aryloxoalcanoic acid compounds. Antimicrobial Agents and Chemotherapy. 2001;45(6):1815–1822. doi: 10.1128/AAC.45.6.1815-1822.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Hachler H., Cohen S. P., Levy S. B. marA, a regulated locus which controls expression of chromosomal multiple antibiotic resistance in Escherichia coli . Journal of Bacteriology. 1991;173(17):5532–5538. doi: 10.1128/jb.173.17.5532-5538.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Oh J. T., Cajal Y., Skowronska E. M., et al. Cationic peptide antimicrobials induce selective transcription of micF and osmY in Escherichia coli . Biochimica et Biophysica Acta. 2000;1463(1):43–54. doi: 10.1016/S0005-2736(99)00177-7. [DOI] [PubMed] [Google Scholar]
- 71.Küper C., Jung K. CadC-mediated activation of the cadBA promoter in Escherichia coli . Journal of Molecular Microbiology and Biotechnology. 2006;10(1):26–39. doi: 10.1159/000090346. [DOI] [PubMed] [Google Scholar]
- 72.Rhee J. E., Kim K. S., Choi S. H. CadC activates pH-dependent expression of the Vibrio vulnificus cadBA operon at a distance through direct binding to an upstream region. Journal of Bacteriology. 2005;187(22):7870–7875. doi: 10.1128/JB.187.22.7870-7875.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Huang L., Tsui P., Freundlich M. Positive and negative control of ompB transcription in Escherichia coli by cyclic AMP and the cyclic AMP receptor protein. Journal of Bacteriology. 1992;174(3):664–670. doi: 10.1128/jb.174.3.664-670.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Gibert I., Barbe J. Cyclic AMP stimulates transcription of the structural gene of the outer-membrane protein ompA of Escherichia coli . FEMS Microbiology Letters. 1990;68(3):307–311. doi: 10.1111/j.1574-6968.1988.tb03197.x. [DOI] [PubMed] [Google Scholar]
- 75.Papenfort K., Pfeiffer V., Lucchini S., Sonawane A., Hinton J. C. D., Vogel J. Systematic deletion of Salmonella small RNA genes identifies CyaR, a conserved CRP-dependent riboregulator of OmpX synthesis. Molecular Microbiology. 2008;68(4):890–906. doi: 10.1111/j.1365-2958.2008.06189.x. [DOI] [PubMed] [Google Scholar]
- 76.Nandi B., Nandy R. K., Sarkar A., Ghose A. C. Structural features, properties and regulation of the outer-membrane protein W (OmpW) of Vibrio cholerae . Microbiology. 2005;151(9):2975–2986. doi: 10.1099/mic.0.27995-0. [DOI] [PubMed] [Google Scholar]
- 77.Hommais F., Krin E., Laurent-Winter C., et al. Large-scale monitoring of pleiotropic regulation of gene expression by the prokaryotic nucleoid-associated protein, H-NS. Molecular Microbiology. 2001;40(1):20–36. doi: 10.1046/j.1365-2958.2001.02358.x. [DOI] [PubMed] [Google Scholar]
- 78.Navarre W. W., Porwollik S., Wang Y., et al. Selective silencing of foreign DNA with low GC content by the H-NS protein in Salmonella . Science. 2006;313(5784):236–238. doi: 10.1126/science.1128794. [DOI] [PubMed] [Google Scholar]
- 79.Johansson J., Dagberg B., Richet E., Uhlin B. E. H-NS and StpA proteins stimulate expression of the maltose regulon in Escherichia coli . Journal of Bacteriology. 1998;180(23):6117–6125. doi: 10.1128/jb.180.23.6117-6125.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Huang L., Tsui P., Freundlich M. Integration host factor is a negative effector of in vivo and in vitro expression of ompC in Escherichia coli . Journal of Bacteriology. 1990;172(9):5293–5298. doi: 10.1128/jb.172.9.5293-5298.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Flores-Valdez M. A., Puente J. L., Calva E. Negative osmoregulation of the Salmonella ompS1 porin gene independently of OmpR in an hns background. Journal of Bacteriology. 2003;185(22):6497–6506. doi: 10.1128/JB.185.22.6497-6506.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Romby P., Vandenesch F., Wagner E. G. H. The role of RNAs in the regulation of virulence-gene expression. Current Opinion in Microbiology. 2006;9(2):229–236. doi: 10.1016/j.mib.2006.02.005. [DOI] [PubMed] [Google Scholar]
- 83.Storz G., Altuvia S., Wassarman K. M. An abundance of RNA regulators. Annual Review of Biochemistry. 2005;74:199–217. doi: 10.1146/annurev.biochem.74.082803.133136. [DOI] [PubMed] [Google Scholar]
- 84.Vogel J., Papenfort K. Small non-coding RNAs and the bacterial outer membrane. Current Opinion in Microbiology. 2006;9(6):605–611. doi: 10.1016/j.mib.2006.10.006. [DOI] [PubMed] [Google Scholar]
- 85.Gunnewijk M. G. W., van den Bogaard P. T. C., Veenhoff L. M., et al. Hierarchical control versus autoregulation of carbohydrate utilization in bacteria. Journal of Molecular Microbiology and Biotechnology. 2001;3(3):401–413. [PubMed] [Google Scholar]
- 86.Poolman B., Konings W. N. Secondary solute transport in bacteria. Biochimica et Biophysica Acta. 1993;1183(1):5–39. doi: 10.1016/0005-2728(93)90003-X. [DOI] [PubMed] [Google Scholar]
- 87.Postma P. W., Lengeler J. W., Jacobson G. R. Phosphoenolpyruvate: carbohydrate phosphotransferase systems. In: Neidhardt F. C., editor. Escherichia coli and Salmonella: Cellular and Molecular Biology. Washington, DC, USA: ASM Press; 1996. pp. 1149–1174. [Google Scholar]
- 88.Tchieu J. H., Norris V., Edwards J. S., Saier M. H., Jr. The complete phosphotransferase system in Escherichia coli . Journal of Molecular Microbiology and Biotechnology. 2001;3(3):329–346. [PubMed] [Google Scholar]
- 89.Curtis S. J., Epstein W. Phosphorylation of D glucose in Escherichia coli mutants defective in glucosephosphotransferase, mannosephosphotransferase, and glucokinase. Journal of Bacteriology. 1975;122(3):1189–1199. doi: 10.1128/jb.122.3.1189-1199.1975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Chou C. H., Bennett G. N., San K. Y. Effect of modulated glucose uptake on high-level recombinant protein production in a dense Escherichia coli culture. Biotechnology Progress. 1994;10(6):644–647. doi: 10.1021/bp00030a009. [DOI] [PubMed] [Google Scholar]
- 91.Flores N., Flores S., Escalante A., et al. Adaptation for fast growth on glucose by differential expression of central carbon metabolism and gal regulon genes in an Escherichia coli strain lacking the phosphoenolpyruvate:carbohydrate phosphotransferase system. Metabolic Engineering. 2005;7(2):70–87. doi: 10.1016/j.ymben.2004.10.002. [DOI] [PubMed] [Google Scholar]
- 92.Death A., Ferenci T. Between feast and famine: Endogenous inducer synthesis in the adaptation of Escherichia coli to growth with limiting carbohydrates. Journal of Bacteriology. 1994;176:5101–5107. doi: 10.1128/jb.176.16.5101-5107.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Lunin V. V., Li Y., Schrag J. D., Iannuzzi P., Cygler M., Matte A. Crystal structures of Escherichia coli ATP-dependent glucokinase and its complex with glucose. Journal of Bacteriology. 2004;186(20):6915–6927. doi: 10.1128/JB.186.20.6915-6927.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Monod J. Recherches sur la Croissance de cultures Bacteriennes [thesis] Paris, France: Hermann et Cie; 1942. [Google Scholar]
- 95.Magasanik B. Catabolite repression. Cold Spring Harbor Symposia on Quantitative Biology. 1961;26:249–256. doi: 10.1101/sqb.1961.026.01.031. [DOI] [PubMed] [Google Scholar]
- 96.Bowien B., Kusian B. Genetics and control of CO2 assimilation in the chemoautotroph Ralstonia eutropha . Archives of Microbiology. 2002;178(2):85–93. doi: 10.1007/s00203-002-0441-3. [DOI] [PubMed] [Google Scholar]
- 97.Nicholson T. L., Chiu K., Stephens R. S. Chlamydia trachomatis lacks an adaptive response to changes in carbon source availability. Infection and Immunity. 2004;72(7):4286–4289. doi: 10.1128/IAI.72.7.4286-4289.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Halbedel S., Eilers H., Jonas B., et al. Transcription in Mycoplasma pneumoniae: analysis of the promoters of the ackA and ldh genes. Journal of Molecular Biology. 2007;371(3):596–607. doi: 10.1016/j.jmb.2007.05.098. [DOI] [PubMed] [Google Scholar]
- 99.Frunzke J., Engels V., Hasenbein S., Gätgens C., Bott M. Co-ordinated regulation of gluconate catabolism and glucose uptake in Corynebacterium glutamicum by two functionally equivalent transcriptional regulators, GntR1 and GntR2. Molecular Microbiology. 2008;67(2):305–322. doi: 10.1111/j.1365-2958.2007.06020.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Wendisch V. F., de Graaf A. A., Sahm H., Eikmanns B. J. Quantitative determination of metabolic fluxes during coutilization of two carbon sources: comparative analyses with Corynebacterium glutamicum during growth on acetate and/or glucose. Journal of Bacteriology. 2000;182(11):3088–3096. doi: 10.1128/JB.182.11.3088-3096.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Bogaard P. T. C. Van den, Kleerebezem M., Kuipers O. P., Vos W. M. Control of lactose transport, β-galactosidase activity, and glycolysis by CcpA in Streptococcus thermophilus: evidence for carbon catabolite repression by a non-phosphoenolpyryvate-dependent phosphotransferase system sugar. Journal of Bacteriology. 2000;182:5982–5989. doi: 10.1128/jb.182.21.5982-5989.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Parche S., Beleut M., Rezzonico E., et al. Lactose-over-glucose preference in Bifidobacterium longum NCC2705: glcP, encoding a glucose transporter, is subject to lactose repression. Journal of Bacteriology. 2006;188:1260–1265. doi: 10.1128/JB.188.4.1260-1265.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Collier D. N., Hager P. W., Phibbs P. V., Jr. Catabolite repression control in the Pseudomonads . Research in Microbiology. 1996;147(6-7):551–561. doi: 10.1016/0923-2508(96)84011-3. [DOI] [PubMed] [Google Scholar]
- 104.Liu M., Durfee T., Cabrera J. E., Zhao K., Jin D. J., Blattner F. R. Global transcriptional programs reveal a carbon source foraging strategy by Escherichia coli . The Journal of Biological Chemistry. 2005;280(16):15921–15927. doi: 10.1074/jbc.M414050200. [DOI] [PubMed] [Google Scholar]
- 105.Blencke H. M., Homuth G., Ludwig H., Mäder U., Hecker M., Stülke J. Transcriptional profiling of gene expression in response to glucose in Bacillus subtilis: regulation of the central metabolic pathways. Metabolic Engineering. 2003;5(2):133–149. doi: 10.1016/S1096-7176(03)00009-0. [DOI] [PubMed] [Google Scholar]
- 106.Moreno M. S., Schneider B. L., Maile R. R., Weyler W., Saier M. H., Jr. Catabolite repression mediated by the CcpA protein in Bacillus subtilis: novel modes of regulation revealed by whole-genome analyses. Molecular Microbiology. 2001;39(5):1366–1381. doi: 10.1111/j.1365-2958.2001.02328.x. [DOI] [PubMed] [Google Scholar]
- 107.Görke B., Stülke J. Carbon catabolite repression in bacteria: many ways to make the most out of nutrients. Nature Reviews Microbiology. 2008;6(8):613–624. doi: 10.1038/nrmicro1932. [DOI] [PubMed] [Google Scholar]
- 108.Winkler H. H., Wilson T. H. Inhibition of β-galactoside transport by substrates of the glucose transport system in Escherichia coli . Biochimica et Biophysica Acta. 1967;135(5):1030–1051. doi: 10.1016/0005-2736(67)90073-9. [DOI] [PubMed] [Google Scholar]
- 109.Hogema B. M., Arents J. C., Bader R., Postma P. W. Autoregulation of lactose uptake through the LacY permease by enzyme IIAGlc of the PTS in Escherichia coli K-12. Molecular Microbiology. 1999;31(6):1825–1833. doi: 10.1046/j.1365-2958.1999.01319.x. [DOI] [PubMed] [Google Scholar]
- 110.Nelson S. O., Wright J. K., Postma P. W. The mechanism of inducer exclusion. Direct interaction between purified IIIGlc of the phosphoenolpyruvate:sugar phosphotransferase system and the lactose carrier of Escherichia coli . The EMBO Journal. 1983;2:715–720. doi: 10.1002/j.1460-2075.1983.tb01490.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Smirnova I., Kasho V., Choe J. Y., Altenbach C., Hubbell W. L., Kaback H. R. Sugar binding induces an outward facing conformation of LacY. Proceedings of the National Academy of Sciences of the United States of America. 2007;104(42):16504–16509. doi: 10.1073/pnas.0708258104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Titgemeyer F., Mason R. E., Saier M. H., Jr. Regulation of the raffinose permease of Escherichia coli by the glucose-specific enzyme IIA of the phosphoenolpyruvate:sugar phosphotransferase system. Journal of Bacteriology. 1994;176(2):543–546. doi: 10.1128/jb.176.2.543-546.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Misko T. P., Mitchell W. J., Meadow N. D., Roseman S. Sugar transport by the bacterial phosphotransferase system. Reconstitution of inducer exclusion in Salmonella typhimurium membrane vesicles. The Journal of Biological Chemistry. 1987;262(33):16261–16266. [PubMed] [Google Scholar]
- 114.Djordjevic G. M., Tchieu J. H., Saier M. H., Jr. Genes involved in control of galactose uptake in Lactobacillus brevis and reconstitution of the regulatory system in Bacillus subtilis . Journal of Bacteriology. 2001;183(10):3224–3236. doi: 10.1128/JB.183.10.3224-3236.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Poolman B., Knol J., Mollet B., Nieuwenhuis B., Sulter G. Regulation of bacterial sugar-H+ symport by phosphoenolpyruvate-dependent enzyme I/HPr-mediated phosphorylation. Proceedings of the National Academy of Sciences of the United States of America. 1995;92(3):778–782. doi: 10.1073/pnas.92.3.778. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Gunnewijk M. G. W., Poolman B. Phosphorylation state of Hpr determines the level of expression and the extent of phosphorylation of the lactose transport protein of Streptococcus thermophilus . The Journal of Biological Chemistry. 2000;275(44):34073–34079. doi: 10.1074/jbc.M003512200. [DOI] [PubMed] [Google Scholar]
- 117.Plumbridge J. Expression of ptsG, the gene for the major glucose PTS transporter in Escherichia coli, is repressed by Mlc and induced by growth on glucose. Molecular Microbiology. 1998;29(4):1053–1063. doi: 10.1046/j.1365-2958.1998.00991.x. [DOI] [PubMed] [Google Scholar]
- 118.de Reuse H., Danchin A. The ptsH, ptsI, and crr genes of the Escherichia coli phosphoenolpyruvate-dependent phosphotransferase system: a complex operon with several modes of transcription. Journal of Bacteriology. 1988;170(9):3827–3837. doi: 10.1128/jb.170.9.3827-3837.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Bettenbrock K., Fischer S., Kremling A., Jahreis K., Sauter T., Gilles E. D. A quantitative approach to catabolite repression in Escherichia coli . The Journal of Biological Chemistry. 2006;281(5):2578–2584. doi: 10.1074/jbc.M508090200. [DOI] [PubMed] [Google Scholar]
- 120.Boris G. Carbon catabolite repression in bacteria: many ways to make the most out of nutrients. Nature Reviews Microbiology. 2008;6(8):613–624. doi: 10.1038/nrmicro1932. [DOI] [PubMed] [Google Scholar]
- 121.Park Y. H., Lee B. R., Seok Y. J., Peterkofsky A. In vitro reconstitution of catabolite repression in Escherichia coli . The Journal of Biological Chemistry. 2006;281(10):6448–6454. doi: 10.1074/jbc.M512672200. [DOI] [PubMed] [Google Scholar]
- 122.Tanaka Y., Kimata K., Aiba H. A novel regulatory role of glucose transporter of Escherichia coli: membrane sequestration of a global repressor Mlc. The EMBO Journal. 2000;19(20):5344–5352. doi: 10.1093/emboj/19.20.5344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Lee S. J., Boos W., Bouché J. P., Plumbridge J. Signal transduction between a membrane-bound transporter, PtsG, and a soluble transcription factor, Mlc of Escherichia coli . The EMBO Journal. 2000;19(20):5353–5361. doi: 10.1093/emboj/19.20.5353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Bettenbrock K., Sauter T., Jahreis K., Kremling A., Lengeler J. W., Gilles E. D. Correlation between growth rates, EIIACrr phosphorylation, and intracellular cyclic AMP levels in Escherichia coli K-12. Journal of Bacteriology. 2007;189(19):6891–6900. doi: 10.1128/JB.00819-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Hogema B. M., Arents J. C., Bader R., et al. Inducer exclusion in Escherichia coli by non-PTS substrates: the role of the PEP to pyruvate ratio in determining the phosphorylation state of enzyme IIAGlc . Molecular Microbiology. 1998;30(3):487–498. doi: 10.1046/j.1365-2958.1998.01053.x. [DOI] [PubMed] [Google Scholar]
- 126.Inada T., Kimata K., Aiba H. Mechanism responsible for glucose-lactose diauxie in Escherichia coli: challenge to the cAMP model. Genes to Cells. 1996;1(3):293–301. doi: 10.1046/j.1365-2443.1996.24025.x. [DOI] [PubMed] [Google Scholar]
- 127.Kimata K., Takahashi H., Inada T., Postma P., Aiba H. cAMP receptor protein-cAMP plays a crucial role in glucose-lactose diauxie by activating the major glucose transporter gene in Escherichia coli . Proceedings of the National Academy of Sciences of the United States of America. 1997;94(24):12914–12919. doi: 10.1073/pnas.94.24.12914. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Deutscher J., Francke C., Postma P. W. How phosphotransferase system-related protein phosphorylation regulates carbohydrate metabolism in bacteria. Microbiology and Molecular Biology Reviews. 2006;70(4):939–1031. doi: 10.1128/MMBR.00024-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Aulkemeyer P., Ebner R., Heilenmann G., et al. Molecular analysis of two fructokinases involved in sucrose metabolism of enteric bacteria. Molecular Microbiology. 1991;5(12):2913–2922. doi: 10.1111/j.1365-2958.1991.tb01851.x. [DOI] [PubMed] [Google Scholar]
- 130.Kornberg H. L. Fructose transport by Escherichia coli . Philosophical Transactions of the Royal Society B: Biological Sciences. 1990;326(1236):505–513. doi: 10.1098/rstb.1990.0028. [DOI] [PubMed] [Google Scholar]
- 131.Kornberg H. L. Routes for fructose utilization by Escherichia coli . Journal of Molecular Microbiology and Biotechnology. 2001;3(3):355–359. [PubMed] [Google Scholar]
- 132.Titgemeyer F., Hillen W. Global control of sugar metabolism: a gram-positive solution. Antonie van Leeuwenhoek. 2002;82(1-4):59–71. doi: 10.1023/A:1020628909429. [DOI] [PubMed] [Google Scholar]
- 133.Warner J. B., Lolkema J. S. A Crh-specific function in carbon catabolite repression in Bacillus subtilis . FEMS Microbiology Letters. 2003;220(2):277–280. doi: 10.1016/S0378-1097(03)00126-5. [DOI] [PubMed] [Google Scholar]
- 134.Henkin T. M., Grundy F. J., Nicholson W. L., Chambliss G. H. Catabolite repression of α-amylase gene expression in Bacillus subtilis involves a trans-acting gene product homologous to the Escherichia coli lacI and galR repressors. Molecular Microbiology. 1991;5(3):575–584. doi: 10.1111/j.1365-2958.1991.tb00728.x. [DOI] [PubMed] [Google Scholar]
- 135.Galinier A., Kravanja M., Engelmann R., et al. New protein kinase and protein phosphatase families mediate signal transduction in bacterial catabolite repression. Proceedings of the National Academy of Sciences of the United States of America. 1998;95(4):1823–1828. doi: 10.1073/pnas.95.4.1823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Reizer J., Hoischen C., Titgemeyer F., et al. A novel protein kinase that controls carbon catabolite repression in bacteria. Molecular Microbiology. 1998;27(6):1157–1169. doi: 10.1046/j.1365-2958.1998.00747.x. [DOI] [PubMed] [Google Scholar]
- 137.Jault J. M., Fieulaine S., Nessler S., et al. The HPr kinase from Bacillus subtilis is a homo-oligomeric enzyme which exhibits strong positive cooperativity for nucleotide and fructose 1,6-bisphosphate binding. The Journal of Biological Chemistry. 2000;275(3):1773–1780. doi: 10.1074/jbc.275.3.1773. [DOI] [PubMed] [Google Scholar]
- 138.Mijakovic I., Poncet S., Galinier A., et al. Pyrophosphate-producing protein dephosphorylation by HPr kinase/phosphorylase: a relic of early life? Proceedings of the National Academy of Sciences of the United States of America. 2002;99(21):13442–13447. doi: 10.1073/pnas.212410399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Seidel G., Diel M., Fuchsbauer N., Hillen W. Quantitative interdependence of coeffectors, CcpA and cre in carbon catabolite regulation of Bacillus subtilis . FEBS Journal. 2005;272(10):2566–2577. doi: 10.1111/j.1742-4658.2005.04682.x. [DOI] [PubMed] [Google Scholar]
- 140.Schumacher M. A., Seidel G., Hillen W., Brennan R. G. Structural mechanism for the fine-tuning of CcpA function by the small molecule effectors glucose 6-phosphate and fructose 1,6-bisphosphate. Journal of Molecular Biology. 2007;368(4):1042–1050. doi: 10.1016/j.jmb.2007.02.054. [DOI] [PubMed] [Google Scholar]
- 141.van Wezel G. P., König M., Mahr K., et al. A new piece of an old jigsaw: glucose kinase is activated posttranslationally in a glucose transport-dependent manner in Streptomyces coelicolor A3(2) Journal of Molecular Microbiology and Biotechnology. 2006;12(1-2):67–74. doi: 10.1159/000096461. [DOI] [PubMed] [Google Scholar]
- 142.Arndt A., Eikmanns B. J. The alcohol dehydrogenase gene adhA in Corynebacterium glutamicum is subject to carbon catabolite repression. Journal of Bacteriology. 2007;189(20):7408–7416. doi: 10.1128/JB.00791-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Gerstmeir R., Cramer A., Dangel P., Schaffer S., Eikmanns B. J. RamB, a novel transcriptional regulator of genes involved in acetate metabolism of Corynebacterium glutamicum . Journal of Bacteriology. 2004;186(9):2798–2809. doi: 10.1128/JB.186.9.2798-2809.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Bott M., Brocker M. Two-component signal transduction in Corynebacterium glutamicum and other corynebacteria: on the way towards stimuli and targets. Applied Microbiology and Biotechnology. 2012;94(5):1131–1150. doi: 10.1007/s00253-012-4060-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Müller C., Petruschka L., Cuypers H., Burchhardt G., Herrmann H. Carbon catabolite repression of phenol degradation in Pseudomonas putida is mediated by the inhibition of the activator protein PhIR. Journal of Bacteriology. 1996;178(7):2030–2036. doi: 10.1128/jb.178.7.2030-2036.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Moreno R., Ruiz-Manzano A., Yuste L., Rojo F. The Pseudomonas putida Crc global regulator is an RNA binding protein that inhibits translation of the AlkS transcriptional regulator. Molecular Microbiology. 2007;64(3):665–675. doi: 10.1111/j.1365-2958.2007.05685.x. [DOI] [PubMed] [Google Scholar]
- 147.Moreno R., Rojo F. The target for the Pseudomonas putida Crc global regulator in the benzoate degradation pathway is the BenR transcriptional regulator. Journal of Bacteriology. 2008;190(5):1539–1545. doi: 10.1128/JB.01604-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Petersen S., Young G. M. Essential role for cyclic AMP and its receptor protein in Yersinia enterocolitica virulence. Infection and Immunity. 2002;70(7):3665–3672. doi: 10.1128/IAI.70.7.3665-3672.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Ramström H., Bourotte M., Philippe C., Schmitt M., Haiech J., Bourguignon J. J. Heterocyclic bis-cations as starting hits for design of inhibitors of the bifunctional enzyme histidine-containing protein kinase/phosphatase from Bacillus subtilis . Journal of Medicinal Chemistry. 2004;47(9):2264–2275. doi: 10.1021/jm021043o. [DOI] [PubMed] [Google Scholar]
- 150.Curtiss R., Kelly S. M. Salmonella typhimurium deletion mutants lacking adenylate cyclase and cyclic AMP receptor protein are avirulent and immunogenic. Infection and Immunity. 1987;55(12):3035–3043. doi: 10.1128/iai.55.12.3035-3043.1987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Romeo T. Global regulation by the small RNA-binding protein CsrA and the non- coding RNA molecule CsrB. Molecular Microbiology. 1998;29(6):1321–1330. doi: 10.1046/j.1365-2958.1998.01021.x. [DOI] [PubMed] [Google Scholar]
- 152.Jonas K., Edwards A. N., Ahmad I., Romeo T., Römling U., Melefors O. Complex regulatory network encompassing the Csr, c-di-GMP and motility systems of Salmonella typhimurium . Environmental Microbiology. 2010;12(2):524–540. doi: 10.1111/j.1462-2920.2009.02097.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Yakhnin H., Pandit P., Petty T. J., Baker C. S., Romeo T., Babitzke P. CsrA of Bacillus subtilis regulates translation initiation of the gene encoding the flagellin protein (hag) by blocking ribosome binding. Molecular Microbiology. 2007;64(6):1605–1620. doi: 10.1111/j.1365-2958.2007.05765.x. [DOI] [PubMed] [Google Scholar]
- 154.Romeo T., Gong M. Genetic and physical mapping of the regulatory gene csrA on the Escherichia coli K-12 chromosome. Journal of Bacteriology. 1993;175(17):5740–5741. doi: 10.1128/jb.175.17.5740-5741.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Romeo T., Gong M., Liu M. Y., Brun-Zinkernagel A. M. Identification and molecular characterization of csrA, a pleiotropic gene from Escherichia coli that affects glycogen biosynthesis, gluconeogenesis, cell size, and surface properties. Journal of Bacteriology. 1993;175(15):4744–4755. doi: 10.1128/jb.175.15.4744-4755.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Romeo T. Post-transcriptional regulation of bacterial carbohydrate metabolism: evidence that the gene product CsrA is global mRNA decay factor. Research in Microbiology. 1996;147(6-7):505–512. doi: 10.1016/0923-2508(96)84004-6. [DOI] [PubMed] [Google Scholar]
- 157.Baker C. S., Eöry L. A., Yakhnin H., Mercante J., Romeo T., Babitzke P. CsrA inhibits translation initiation of Escherichia coli hfq by binding to a single site overlapping the Shine-Dalgarno sequence. Journal of Bacteriology. 2007;189(15):5472–5481. doi: 10.1128/JB.00529-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Dubey A. K., Baker C. S., Romeo T., Babitzke P. RNA sequence and secondary structure participate in high-affinity CsrA-RNA interaction. RNA. 2005;11(10):1579–1587. doi: 10.1261/rna.2990205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Suzuki K., Wang X., Weilbacher T., et al. Regulatory circuitry of the CsrA/CsrB and BarA/UvrY systems of Escherichia coli . Journal of Bacteriology. 2002;184(18):5130–5140. doi: 10.1128/JB.184.18.5130-5140.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Weilbacher T., Suzuki K., Dubey A. K., et al. A novel sRNA component of the carbon storage regulatory system of Escherichia coli . Molecular Microbiology. 2003;48(3):657–670. doi: 10.1046/j.1365-2958.2003.03459.x. [DOI] [PubMed] [Google Scholar]
- 161.Baker C. S., Morozov I., Suzuki K., Romeo T., Babitzke P. CsrA regulates glycogen biosynthesis by preventing translation of glgC in Escherichia coli . Molecular Microbiology. 2002;44(6):1599–1610. doi: 10.1046/j.1365-2958.2002.02982.x. [DOI] [PubMed] [Google Scholar]
- 162.Mckee A. E., Rutherford B. J., Chivian D. C., et al. doi: 10.1186/1475-2859-11-79. Microbial Cell Factories. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Yakandawala N., Romeo T., Friesen A. D., Madhyastha S. Metabolic engineering of Escherichia coli to enhance phenylalanine production. Applied Microbiology and Biotechnology. 2008;78(2):283–291. doi: 10.1007/s00253-007-1307-z. [DOI] [PubMed] [Google Scholar]
- 164.Moat A. G., Foster J. W., Spector M. P. Microbiology. 4th. New York, NY, USA: Wiley-Liss; 2002. [Google Scholar]
- 165.Saier M. H., Jr., Ramseier T. M., Reizer J. Regulation of carbon utilization. In: Neidhardt C., Curtiss R. III, Ingraham J. L., et al., editors. Escherichia coli and Salmonella: Cellular and Molecular Biology. Washington, DC, USA: ASM Press; 1996. pp. 1325–1343. [Google Scholar]
- 166.Saier M. H., Jr., Ramseier T. M. The catabolite repressor/activator (Cra) protein of enteric bacteria. Journal of Bacteriology. 1996;178(12):3411–3417. doi: 10.1128/jb.178.12.3411-3417.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Chin A. M., Feldheim D. A., Saier M. H., Jr. Altered transcriptional patterns affecting several metabolic pathways in strains of Salmonella typhimurium which overexpress the fructose regulon. Journal of Bacteriology. 1989;171(5):2424–2434. doi: 10.1128/jb.171.5.2424-2434.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Lee J. H., Lee D. E., Lee B. U., Kim H. S. Global analyses of transcriptomes and proteomes of a parent strain and an l-threonine-overproducing mutant strain. Journal of Bacteriology. 2003;185(18):5442–5451. doi: 10.1128/JB.185.18.5442-5451.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Ramseier T. M., Bledig S., Michotey V., Feghali R., Saier M. H., Jr. The global regulatory protein FruR modulates the direction of carbon flow in Escherichia coli . Molecular Microbiology. 1995;16(6):1157–1169. doi: 10.1111/j.1365-2958.1995.tb02339.x. [DOI] [PubMed] [Google Scholar]
- 170.Ramseier T. M., Chien S. Y., Saier M. H., Jr. Cooperative interaction between Cra and Fnr in the regulation of the cydAB operon of Escherichia coli . Current Microbiology. 1996;33(4):270–274. doi: 10.1007/s002849900112. [DOI] [PubMed] [Google Scholar]
- 171.Saier H. M., Jr., Ramseier T., Reizer J. Regulation of carbon utilization. In: Neidhardt F. C., editor. Escherichia coli and Salmonella typhimurium: Cellular and Molecular Biology. Washington, DC, USA: ASM Press; 1997. [Google Scholar]
- 172.Ryu S., Ramseier T. M., Michotey V., Saier M. H., Jr., Garges S. Effect of the FruR regulator on transcription of the pts operon in Escherichia coli . The Journal of Biological Chemistry. 1995;270(6):2489–2496. doi: 10.1074/jbc.270.6.2489. [DOI] [PubMed] [Google Scholar]
- 173.Mikulskis A., Aristarkhov A., Lin E. C. C. Regulation of expression of the ethanol dehydrogenase gene (adhE) in Escherichia coli by catabolite repressor activator protein Cra. Journal of Bacteriology. 1997;179(22):7129–7134. doi: 10.1128/jb.179.22.7129-7134.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.Prost J. F., Nègre D., Oudot C., et al. Cra-dependent transcriptional activation of the icd gene of Escherichia coli . Journal of Bacteriology. 1999;181(3):893–898. doi: 10.1128/jb.181.3.893-898.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Cortay J.-C., Negre D., Scarabel M., et al. In vitro asymmetric binding of the pleiotropic regulatory protein , FruR, to the ace operator controlling glyoxylate shunt enzyme synthesis. Journal of Biological Chemictry. 1994;269:14885–14891. [PubMed] [Google Scholar]
- 176.Sarkar D., Shimizu K. Effect of cra gene knockout together with other genes knockouts on the improvement of substrate consumption rate in Escherichia coli under microaerobic condition. Biochemical Engineering Journal. 2008;42(3):224–228. doi: 10.1016/j.bej.2008.06.019. [DOI] [Google Scholar]
- 177.Sarkar D., Siddiquee K. A. Z., Araúzo-Bravo M. J., Oba T., Shimizu K. Effect of cra gene knockout together with edd and iclR genes knockout on the metabolism in Escherichia coli . Archives of Microbiology. 2008;190(5):559–571. doi: 10.1007/s00203-008-0406-2. [DOI] [PubMed] [Google Scholar]
- 178.Phue J. N., Noronha S. B., Hattacharyya R., Wolfe A. J., Shiloach J. Glucose metabolism at high density growth of E. coli B and E. coli K: differences in metabolic pathways are responsible for efficient glucose utilization in E. coli B as determined by microarrays and Northern blot analyses. Biotechnology and Bioengineering. 2005;90(7):805–820. doi: 10.1002/bit.20478. [DOI] [PubMed] [Google Scholar]
- 179.Yao R., Hirose Y., Sarkar D., Nakahigashi K., Ye Q., Shimizu K. Catabolic regulation analysis of Escherichia coli and its crp, mlc, mgsA, pgi and ptsG mutants. Microbial Cell Factories. 2011;10, article 67 doi: 10.1186/1475-2859-10-67. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Yan D. Protection of the glutamate pool concentration in enteric bacteria. Proceedings of the National Academy of Sciences of the United States of America. 2007;104(22):9475–9480. doi: 10.1073/pnas.0703360104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181.Ninfa A. J., Jiang P., Atkinson M. R., Peliska J. A. Integration of antagonistic signals in the regulation of nitrogen assimilation in Escherichia coli . Current Topics in Cellular Regulation. 2000;36:31–75. doi: 10.1016/s0070-2137(01)80002-9. [DOI] [PubMed] [Google Scholar]
- 182.Reitzer L. Nitrogen assimilation and global regulation in Escherichia coli . Annual Reviews in Microbiology. 2003;57:155–176. doi: 10.1146/annurev.micro.57.030502.090820. [DOI] [PubMed] [Google Scholar]
- 183.Fischer E., Sauer U. Metabolic flux profiling of Escherichia coli mutants in central carbon metabolism using GC-MS. European Journal of Biochemistry. 2003;270(5):880–891. doi: 10.1046/j.1432-1033.2003.03448.x. [DOI] [PubMed] [Google Scholar]
- 184.Zhang X., Jantama K., Moore J. C., Shanmugam K. T., Ingram L. O. Production of L-alanine by metabolically engineered Escherichia coli . Applied Microbiology and Biotechnology. 2007;77(2):355–366. doi: 10.1007/s00253-007-1170-y. [DOI] [PubMed] [Google Scholar]
- 185.Commichau F. M., Forchhammer K., Stülke J. Regulatory links between carbon and nitrogen metabolism. Current Opinion in Microbiology. 2006;9(2):167–172. doi: 10.1016/j.mib.2006.01.001. [DOI] [PubMed] [Google Scholar]
- 186.Mao X. J., Huo Y. X., Buck M., Kolb A., Wang Y. P. Interplay between CRP-cAMP and PII-Ntr systems forms novel regulatory network between carbon metabolism and nitrogen assimilation in Escherichia coli . Nucleic Acids Research. 2007;35(5):1432–1440. doi: 10.1093/nar/gkl1142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 187.Sauer U., Eikmanns B. J. The PEP-pyruvate-oxaloacetate node as the switch point for carbon flux distribution in bacteria. FEMS Microbiology Reviews. 2005;29(4):765–794. doi: 10.1016/j.femsre.2004.11.002. [DOI] [PubMed] [Google Scholar]
- 188.Hua Q., Yang C., Baba T., Mori H., Shimizu K. Responses of the central metabolism in Escherichia coli to phosphoglucose isomerase and glucose-6-phosphate dehydrogenase knockouts. Journal of Bacteriology. 2003;185(24):7053–7067. doi: 10.1128/JB.185.24.7053-7067.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189.Hua Q., Yang C., Oshima T., Mori H., Shimizu K. Analysis of gene expression in Escherichia coli in response to changes of growth-limiting nutrient in chemostat cultures. Applied and Environmental Microbiology. 2004;70(4):2354–2366. doi: 10.1128/AEM.70.4.2354-2366.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 190.Nanchen A., Schicker A., Revelles O., Sauer U. Cyclic AMP-dependent catabolite repression is the dominant control mechanism of metabolic fluxes under glucose limitation in Escherichia coli . Journal of Bacteriology. 2008;190(7):2323–2330. doi: 10.1128/JB.01353-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 191.Kumar R., Shimizu K. Metabolic regulation of Escherichia coli and its gdhA, glnL, gltB, D mutants under different carbon and nitrogen limitations in the continuous culture. Microbial Cell Factories. 2010;9:8. doi: 10.1186/1475-2859-9-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192.Jiang P., Ninfa A. J. Escherichia coli PII signal transduction protein controlling nitrogen assimilation acts as a sensor of adenylate energy charge in vitro . Biochemistry. 2007;46(45):12979–12996. doi: 10.1021/bi701062t. [DOI] [PubMed] [Google Scholar]
- 193.Ninfa A. J., Jiang P. PII signal transduction proteins: sensors of α-ketoglutarate that regulate nitrogen metabolism. Current Opinion in Microbiology. 2005;8(2):168–173. doi: 10.1016/j.mib.2005.02.011. [DOI] [PubMed] [Google Scholar]
- 194.Gosset G., Zhang Z., Nayyar S., Cuevas W. A., Saier M. H., Jr. Transcriptome analysis of Crp-dependent catabolite control of gene expression in Escherichia coli . Journal of Bacteriology. 2004;186(11):3516–3524. doi: 10.1128/JB.186.11.3516-3524.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 195.Rahman M., Hasan M. R., Shimizu K. Growth phase-dependent changes in the expression of global regulatory genes and associated metabolic pathways in Escherichia coli . Biotechnology Letters. 2008;30(5):853–860. doi: 10.1007/s10529-007-9621-1. [DOI] [PubMed] [Google Scholar]
- 196.Helling R. B. Pathway choice in glutamate synthesis in Escherichia coli . Journal of Bacteriology. 1998;180(17):4571–4575. doi: 10.1128/jb.180.17.4571-4575.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 197.Liang A., Houghton L. Coregulation of oxidized nicotinamide adenine dinucleotide (phosphate) transhydrogenase and glutamate dehydrogenase activities in enteric bacteria during nitrogen limitation. Journal of Bacteriology. 1981;146(3):997–1002. doi: 10.1128/jb.146.3.997-1002.1981. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 198.Willis R. C., Iwata K. K., Furlong C. E. Regulation of glutamine transport in Escherichia coli . Journal of Bacteriology. 1975;122(3):1032–1037. doi: 10.1128/jb.122.3.1032-1037.1975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 199.Claverie-Martin F., Magasanik B. Role of integration host factor in the regulation of the glnHp2 promoter of Escherichia coli . Proceedings of the National Academy of Sciences of the United States of America. 1991;88(5):1631–1635. doi: 10.1073/pnas.88.5.1631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 200.Blauwkamp T. A., Ninfa A. J. Physiological role of the GlnK signal transduction protein of Escherichia coli: survival of nitrogen starvation. Molecular Microbiology. 2002;46(1):203–214. doi: 10.1046/j.1365-2958.2002.03153.x. [DOI] [PubMed] [Google Scholar]
- 201.Magasanik B. Regulation of nitrogen utilization. In: Neidhardt F. C., editor. Escherichia coli and Salmonella typhimurium: Cellular and Molecular Biology. Washington, DC, USA: ASM Press; 1996. pp. 1344–1356. [Google Scholar]
- 202.Riba L., Becerril B., Servin-Gonzalez L., Valle F., Bolivar F. Identification of a functional promoter for the Escherichia coli gdhA gene and its regulation. Gene. 1988;71(2):233–246. doi: 10.1016/0378-1119(88)90040-6. [DOI] [PubMed] [Google Scholar]
- 203.Camarena L., Poggio S., García N., Osorio A. Transcriptional repression of gdhA in Escherichia coli is mediated by the Nac protein. FEMS Microbiology Letters. 1998;167(1):51–56. doi: 10.1016/S0378-1097(98)00366-8. [DOI] [PubMed] [Google Scholar]
- 204.Reitzer L. J., Magasanik B. Expression of glnA in Escherichia coli is regulated at tandem promoters. Proceedings of the National Academy of Sciences of the United States of America. 1985;82(7):1979–1983. doi: 10.1073/pnas.82.7.1979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 205.Atkinson M. R., Ninfa A. J. Mutational analysis of the bacterial signal-transducing protein kinase/phosphatase nitrogen regulator II (NR(II) or NtrB) Journal of Bacteriology. 1993;175(21):7016–7023. doi: 10.1128/jb.175.21.7016-7023.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 206.Atkinson M. R., Blauwkamp T. A., Ninfa A. J. Context-dependent functions of the PII and GlnK signal transduction proteins in Escherichia coli . Journal of Bacteriology. 2002;184(19):5364–5375. doi: 10.1128/JB.184.19.5364-5375.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 207.van Heeswijk W. C., Hoving S., Molenaar D., Stegeman B., Kahn D., Westerhoff H. V. An alternative P(II) protein in the regulation of glutamine synthetase in Escherichia coli . Molecular Microbiology. 1996;21(1):133–146. doi: 10.1046/j.1365-2958.1996.6281349.x. [DOI] [PubMed] [Google Scholar]
- 208.Pahel G., Rothstein D. M., Magasanik B. Complex glnA-glnL-glnG operon of Escherichia coli . Journal of Bacteriology. 1982;150(1):202–213. doi: 10.1128/jb.150.1.202-213.1982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 209.Paul L., Mishra P. K., Blumenthal R. M., Matthews R. G. Integration of regulatory signals through involvement of multiple global regulators: control of the Escherichia coli gltBDF operon by Lrp, IHF, Crp, and ArgR. BMC Microbiology. 2007;7, article 2 doi: 10.1186/1471-2180-7-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 210.Shapiro B. M., Stadtman E. R. Glutamine synthetase deadenylylating enzyme. Biochemical and Biophysical Research Communications. 1968;30(1):32–37. doi: 10.1016/0006-291x(68)90708-0. [DOI] [PubMed] [Google Scholar]
- 211.Stadtman E. R. Discovery of glutamine synthetase cascade. Methods in Enzymology. 1990;182:793–809. doi: 10.1016/0076-6879(90)82062-7. [DOI] [PubMed] [Google Scholar]
- 212.Jaggi R., van Heeswijk W. C., Westerhoff H. V., Ollis D. L., Vasudevan S. G. The two opposing activities of adenylyl transferase reside in distinct homologous domains, with intramolecular signal transduction. The EMBO Journal. 1997;16(18):5562–5571. doi: 10.1093/emboj/16.18.5562. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 213.Maheswaran M., Forchhammer K. Carbon-source-dependent nitrogen regulation in Escherichia coli is mediated through glutamine-dependent GlnB signalling. Microbiology. 2003;149(8):2163–2172. doi: 10.1099/mic.0.26449-0. [DOI] [PubMed] [Google Scholar]
- 214.Merrick M. J., Edwards R. A. Nitrogen control in bacteria. Microbiological Reviews. 1995;59(4):604–622. doi: 10.1128/mr.59.4.604-622.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 215.Ninfa A. J., Atkinson M. R. PII signal transduction proteins. Trends in Microbiology. 2000;8(4):172–179. doi: 10.1016/S0966-842X(00)01709-1. [DOI] [PubMed] [Google Scholar]
- 216.Jiang P., Peliska J. A., Ninfa A. J. The regulation of Escherichia coli glutamine synthetase revisited: role of 2-ketoglutarate in the regulation of glutamine synthetase adenylylation state. Biochemistry. 1998;37(37):12802–12810. doi: 10.1021/bi980666u. [DOI] [PubMed] [Google Scholar]
- 217.Kustu S., Santero E., Keener J., Popham D., Weiss D. Expression of sigma 54 (ntrA)-dependent genes is probably united by a common mechanism. Microbiological Reviews. 1989;53(3):367–376. doi: 10.1128/mr.53.3.367-376.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 218.Lamarche M. G., Wanner B. L., Crépin S., Harel J. The phosphate regulon and bacterial virulence: a regulatory network connecting phosphate homeostasis and pathogenesis. FEMS Microbiology Reviews. 2008;32(3):461–473. doi: 10.1111/j.1574-6976.2008.00101.x. [DOI] [PubMed] [Google Scholar]
- 219.Ishige T., Krause M., Bott M., Wendisch V. F., Sahm H. The phosphate starvation stimulon of Corynebacterium glutamicum determined by DNA microarray analyses. Journal of Bacteriology. 2003;185(15):4519–4529. doi: 10.1128/JB.185.15.4519-4529.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 220.Baek J. H., Lee S. Y. Novel gene members in the Pho regulon of Escherichia coli . FEMS Microbiology Letters. 2006;264(1):104–109. doi: 10.1111/j.1574-6968.2006.00440.x. [DOI] [PubMed] [Google Scholar]
- 221.Wendisch V. F. Genetic regulation of Corynebacterium glutamicum metabolism. Journal of Microbiology and Biotechnology. 2006;16(7):999–1009. [Google Scholar]
- 222.Damoglou A. P., Dawes E. A. Studies on the lipid content and phosphate requirement of glucose-and acetate-grown Escherichia coli . Biochemical Journal. 1968;110(4):775–781. doi: 10.1042/bj1100775. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 223.Wanner B. L. Gene regulation by phosphate in enteric bacteria. Journal of Cellular Biochemistry. 1993;51(1):47–54. doi: 10.1002/jcb.240510110. [DOI] [PubMed] [Google Scholar]
- 224.Stock J. B., Ninfa A. J., Stock A. M. Protein phosphorylation and regulation of adaptive responses in bacteria. Microbiological Reviews. 1989;53(4):450–490. doi: 10.1128/mr.53.4.450-490.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 225.Parkinson J. S. Signal transduction schemes of bacteria. Cell. 1993;73(5):857–871. doi: 10.1016/0092-8674(93)90267-T. [DOI] [PubMed] [Google Scholar]
- 226.Baek J. H., Lee S. Y. Transcriptome analysis of phosphate starvation response in Escherichia coli . Journal of Microbiology and Biotechnology. 2007;17(2):244–252. [PubMed] [Google Scholar]
- 227.Harris R. M., Webb D. C., Howitt S. M., Cox G. B. Characterization of PitA and PitB from Escherichia coli . Journal of Bacteriology. 2001;183(17):5008–5014. doi: 10.1128/JB.183.17.5008-5014.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 228.Blanco A. G., Sola M., Gomis-Rüth F. X., Coll M. Tandem DNA recognition by PhoB, a two-component signal transduction transcriptional activator. Structure. 2002;10(5):701–713. doi: 10.1016/S0969-2126(02)00761-X. [DOI] [PubMed] [Google Scholar]
- 229.Torriani A., Ludke D. N. The pho regulon of Escherichia coli . In: Schaechter M., Neidhardt F. C., Ingraham J., Kjeldgaard N. O., editors. The Molecular Biology of Bacterial Growth. Boston, Mass, USA: Jones and Bartlett; 1985. [Google Scholar]
- 230.Makino K., Shinagawa H., Nakata A. Regulation of the phosphate regulon of Escherichia coli K-12: regulation and role of the regulatory gene phoR. Journal of Molecular Biology. 1985;184(2):231–240. doi: 10.1016/0022-2836(85)90376-6. [DOI] [PubMed] [Google Scholar]
- 231.Makino K., Shinagawa H., Amemura M., Kimura S., Nakata A., Ishihama A. Regulation of the phosphate regulon of Escherichia coli. Activation of pstS transcription by PhoB protein in vitro . Journal of Molecular Biology. 1988;203(1):85–95. doi: 10.1016/0022-2836(88)90093-9. [DOI] [PubMed] [Google Scholar]
- 232.Makino K., Shinagawa H., Amemura M., Kawamoto T., Yamada M., Nakata A. Signal transduction in the phosphate regulon of Escherichia coli involves phosphotransfer between PhoR and PhoB proteins. Journal of Molecular Biology. 1989;210(3):551–559. doi: 10.1016/0022-2836(89)90131-9. [DOI] [PubMed] [Google Scholar]
- 233.Shinagawa H., Makino K., Amemura M., Nakata A. Structure and function of the regulatory gene for the phosphate regulon in Escherichia coli . In: Torriani-Gorini A., Rothman F. G., Silver S., Wright A., Yagil E., editors. Phosphate Metabolism and Cellular Regulation in Microorganisms. Washington, DC, USA: American Society for Microbiology; 1987. pp. 20–25. [Google Scholar]
- 234.Wanner B. L. Phosphate regulon of gene expression in Escherichia coli . In: Neidhardt F. C., lngraham J. L., Low K. B., Schaechter M., Umbarger H. E., editors. Escherichia coli and Salmonella typhimurium: Cellular and Molecular Biology. Washington, DC, USA: American Society for Microbiology; 1987. pp. 1326–1333. [Google Scholar]
- 235.Amemura M., Makino K., Shinagawa H., Nakata A. Cross talk to the phosphate regulon of Escherichia coli by PhoM protein: PhoM is a histidine protein kinase and catalyzes phosphorylation of PhoB and PhoM-open reading frame 2. Journal of Bacteriology. 1990;172(11):6300–6307. doi: 10.1128/jb.172.11.6300-6307.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 236.Hsieh Y. J., Wanner B. L. Global regulation by the seven-component Pi signaling system. Current Opinion in Microbiology. 2010;13(2):198–203. doi: 10.1016/j.mib.2010.01.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 237.van Dien S. J., Keasling J. D. A dynamic model of the Escherichia coli phosphate-starvation response. Journal of Theoretical Biology. 1998;190(1):37–49. doi: 10.1006/jtbi.1997.0524. [DOI] [PubMed] [Google Scholar]
- 238.Wanner B. L. Phosphate signaling and the control of gene expression in Escherichia coli . In: Silver S., William W., editors. Metal Ions in Gene Regulation. New York, NY, USA: Chapman & Hall; 1997. pp. 104–128. [Google Scholar]
- 239.Oganesyan V., Oganesyan N., Adams P. D., et al. Crystal structure of the “PhoU-like” phosphate uptake regulator from Aquifex aeolicus . Journal of Bacteriology. 2005;187(12):4238–4244. doi: 10.1128/JB.187.12.4238-4244.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 240.Nesmeianova M. A., Gonina S. A., Kulaev I. S. Biosynthesis of Escherichia coli polyphosphatases under control of the regulatory genes usual for alkaline phosphatase. Doklady Akademii Nauk SSSR. 1975;224(3):710–712. [PubMed] [Google Scholar]
- 241.Pratt C., Torriani A. Complementation test between alkaline phosphatase regulatory mutations phoB and phoRc in Escherichia coli . Genetics. 1977;85(2):203–208. doi: 10.1093/genetics/85.2.203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 242.Zuckier G., Ingenito E., Torriani A. Pleiotropic effects of alkaline phosphatase regulatory mutations phoB and phoT on anaerobic growth of and polyphosphate synthesis in Escherichia coli . Journal of Bacteriology. 1980;143(2):934–941. doi: 10.1128/jb.143.2.934-941.1980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 243.Guan C. D., Wanner B., Inouye H. Analysis of regulation of phoB expression using a phoB-cat fusion. Journal of Bacteriology. 1983;156(2):710–717. doi: 10.1128/jb.156.2.710-717.1983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 244.Yamada M., Makino K., Amemura M., Shinagawa H., Nakata A. Regulation of the phosphate regulon of Escherichia coli: analysis of mutant phoB and phoR genes causing different phenotypes. Journal of Bacteriology. 1989;171(10):5601–5606. doi: 10.1128/jb.171.10.5601-5606.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 245.Kimura S., Makino K., Shinagawa H., Amemura M., Nakata A. Regulation of the phosphate regulon of Escherichia coli: characterization of the promoter of the pstS gene. Molecular and General Genetics. 1989;215(3):374–380. doi: 10.1007/BF00427032. [DOI] [PubMed] [Google Scholar]
- 246.Nakata A., Amemura M., Shinagawa H. Regulation of the phosphate regulon in Escherichia coli K-12: regulation of the negative regulatory gene phoU and identification of the gene product. Journal of Bacteriology. 1984;159(3):979–985. doi: 10.1128/jb.159.3.979-985.1984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 247.Tsfasman I. M., Nesmeyanova M. A. Membrane proteins in Escherichia coli: effect of orthophosphate and mutations on regulatory genes of secreted alkaline phosphatase. Molekulyarnaya Biologiya. 1981;15(2):298–309. [PubMed] [Google Scholar]
- 248.Ault-Riché D., Fraley C. D., Tzeng C. M., Kornberg A. Novel assay reveals multiple pathways regulating stress-induced accumulations of inorganic polyphosphate in Escherichia coli . Journal of Bacteriology. 1998;180(7):1841–1847. doi: 10.1128/jb.180.7.1841-1847.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 249.Lazzaroni J. C., Portalier R. C. Regulation of the lkyB gene expression in Escherichia coli K-12 strains carrying an lkyB-lacZ gene fusion. Molecular and General Genetics. 1985;201(2):323–328. doi: 10.1007/BF00425678. [DOI] [PubMed] [Google Scholar]
- 250.Shin P. K., Seo J. H. Analysis of E. coliphoA-lacZ fusion gene expression inserted into a multicopy plasmid and host cell's chromosome. Biotechnology and Bioengineering. 1990;36(11):1097–1104. doi: 10.1002/bit.260361104. [DOI] [PubMed] [Google Scholar]
- 251.Marzan L. W., Shimizu K. Metabolic regulation of Escherichia coli and its phoB and phoR genes knockout mutants under phosphate and nitrogen limitations as well as at acidic condition. Microbial Cell Factories. 2011;10:39. doi: 10.1186/1475-2859-10-39. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 252.Makino K., Amemura M., Kim S. K., Nakata A., Shinagawa H. Role of the σ70 subunit of RNA polymerase in transcriptional activation by activator protein PhoB in Escherichia coli . Genes and Development. 1993;7(1):149–160. doi: 10.1101/gad.7.1.149. [DOI] [PubMed] [Google Scholar]
- 253.Gentry D. R., Hernandez V. J., Nguyen L. H., Jensen D. B., Cashel M. Synthesis of the stationary-phase sigma factor σ s is positively regulated by ppGpp. Journal of Bacteriology. 1993;175(24):7982–7989. doi: 10.1128/jb.175.24.7982-7989.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 254.Ruiz N., Silhavy T. J. Constitutive activation of the Escherichia coli Pho regulon upregulates rpoS translation in an Hfq-dependent fashion. Journal of Bacteriology. 2003;185(20):5984–5992. doi: 10.1128/JB.185.20.5984-5992.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 255.Taschner N. P., Yagil E., Spira B. A differential effect of σ s on the expression of the PHO regulon genes of Escherichia coli . Microbiology. 2004;150(9):2985–2992. doi: 10.1099/mic.0.27124-0. [DOI] [PubMed] [Google Scholar]
- 256.Spira B., Silberstein N., Yagil E. Guanosine 3′,5′-bispyrophosphate (ppGpp) synthesis in cells of Escherichia coli starved for Pi . Journal of Bacteriology. 1995;177(14):4053–4058. doi: 10.1128/jb.177.14.4053-4058.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 257.Spira B., Yagil E. The integration host factor (IHF) affects the expression of the phosphate-binding protein and of alkaline phosphatase in Escherichia coli . Current Microbiology. 1999;38(2):80–85. doi: 10.1007/s002849900407. [DOI] [PubMed] [Google Scholar]
- 258.Taschner N. P., Yagil E., Spira B. The effect of IHF on σ s selectivity of the phoA and pst promoters of Escherichia coli . Archives of Microbiology. 2006;185(3):234–237. doi: 10.1007/s00203-005-0082-4. [DOI] [PubMed] [Google Scholar]
- 259.Schurdell M. S., Woodbury G. M., McCleary W. R. Genetic evidence suggests that the intergenic region between pstA and pstB plays a role in the regulation of rpoS translation during phosphate limitation. Journal of Bacteriology. 2007;189(3):1150–1153. doi: 10.1128/JB.01482-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 260.Wanner B. L., Wilmes M. R., Young D. C. Control of bacterial alkaline phosphatase synthesis and variation in an Escherichia coli K-12 phoR mutant by adenyl cyclase, the cyclic AMP receptor protein, and the phoM operon. Journal of Bacteriology. 1988;170(3):1092–1102. doi: 10.1128/jb.170.3.1092-1102.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 261.Kang Y., Weber K. D., Qiu Y., Kiley P. J., Blattner F. R. Genome-wide expression analysis indicates that FNR of Escherichia coli K-12 regulates a large number of genes of unknown function. Journal of Bacteriology. 2005;187(3):1135–1160. doi: 10.1128/JB.187.3.1135-1160.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 262.Gunsalus R. P. Control of electron flow in Escherichia coli: coordinated transcription of respiratory pathway genes. Journal of Bacteriology. 1992;174(22):7069–7074. doi: 10.1128/jb.174.22.7069-7074.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 263.Alexeeva S., Hellingwerf K. J., de Mattos M. J. T. Requirement of ArcA for redox regulation in Escherichia coli under microaerobic but not anaerobic or aerobic conditions. Journal of Bacteriology. 2003;185(1):204–209. doi: 10.1128/JB.185.1.204-209.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 264.Zhu J., Shalel-Levanon S., Bennett G., San K. Y. Effect of the global redox sensing/regulation networks on Escherichia coli and metabolic flux distribution based on C-13 labeling experiments. Metabolic Engineering. 2006;8(6):619–627. doi: 10.1016/j.ymben.2006.07.002. [DOI] [PubMed] [Google Scholar]
- 265.Shalel-Levanon S., San K. Y., Bennett G. N. Effect of oxygen, and ArcA and FNR regulators on the expression of genes related to the electron transfer chain and the TCA cycle in Escherichia coli . Metabolic Engineering. 2005;7(5-6):364–374. doi: 10.1016/j.ymben.2005.07.001. [DOI] [PubMed] [Google Scholar]
- 266.Zhu J., Shimizu K. The effect of pfl gene knockout on the metabolism for optically pure D-lactate production by Escherichia coli . Applied Microbiology and Biotechnology. 2004;64(3):367–375. doi: 10.1007/s00253-003-1499-9. [DOI] [PubMed] [Google Scholar]
- 267.Zhu J., Shimizu K. Effect of a single-gene knockout on the metabolic regulation in Escherichia coli for D-lactate production under microaerobic condition. Metabolic Engineering. 2005;7(2):104–115. doi: 10.1016/j.ymben.2004.10.004. [DOI] [PubMed] [Google Scholar]
- 268.Alexeeva S., de Kort B., Sawers G., Hellingwerf K. J., de Mattos M. J. T. Effects of limited aeration and of the ArcAB system on intermediary pyruvate catabolism in Escherichia coli . Journal of Bacteriology. 2000;182(17):4934–4940. doi: 10.1128/JB.182.17.4934-4940.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 269.Kessler D., Knappe J. Anaerobic dissimilation of pyruvate. In: Neidhardt F. C., Curtiss R., Ingraham J. I., et al., editors. E. Coli and Salmonella: Cellular and Molecular Biology. 2nd. Vol. 1. Washington, DC, USA: ASM Press; 1996. pp. 199–205. [Google Scholar]
- 270.Sawers G., Watson G. A glycyl radical solution: oxygen-dependent interconversion of pyruvate formate-lyase. Molecular Microbiology. 1998;29(4):945–954. doi: 10.1046/j.1365-2958.1998.00941.x. [DOI] [PubMed] [Google Scholar]
- 271.Wagner A. F. V., Schultz S., Bomke J., Pils T., Lehmann W. D., Knappe J. YfiD of Escherichia coli and Y061 of bacteriophage T4 as autonomous glycyl radical cofactors reconstituting the catalytic center of oxygen-fragmented pyruvate formate-lyase. Biochemical and Biophysical Research Communications. 2001;285(2):456–462. doi: 10.1006/bbrc.2001.5186. [DOI] [PubMed] [Google Scholar]
- 272.Kato M., Mizuno T., Shimizu T., Hakoshima T. Insights into multistep phosphorelay from the crystal structure of the C-terminal HPt domain of ArcB. Cell. 1997;88(5):717–723. doi: 10.1016/S0092-8674(00)81914-5. [DOI] [PubMed] [Google Scholar]
- 273.Ishige K., Nagasawa S., Tokishita S. I., Mizuno T. A novel device of bacterial signal transducers. The EMBO Journal. 1994;13(21):5195–5202. doi: 10.1002/j.1460-2075.1994.tb06850.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 274.Iuchi S., Lin E. C. C. Mutational analysis of signal transduction by ArcB, a membrane sensor protein responsible for anaerobic repression of operons involved in the central aerobic pathways in Escherichia coli . Journal of Bacteriology. 1992;174(12):3972–3980. doi: 10.1128/jb.174.12.3972-3980.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 275.Tsuzuki M., Ishige K., Mizuno T. Phosphotransfer circuitry of the putative multi-signal transducer, ArcB, of Escherichia coli: in vitro studies with mutants. Molecular Microbiology. 1995;18(5):953–962. doi: 10.1111/j.1365-2958.1995.18050953.x. [DOI] [PubMed] [Google Scholar]
- 276.Georgellis D., Lynch A. S., Lin E. C. C. In vitro phosphorylation study of the Arc two-component signal transduction system of Escherichia coli . Journal of Bacteriology. 1997;179(17):5429–5435. doi: 10.1128/jb.179.17.5429-5435.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 277.Iuchi S. Phosphorylation/dephosphorylation of the receiver module at the conserved aspartate residue controls transphosphorylation activity of histidine kinase in sensor protein ArcB of Escherichia coli . The Journal of Biological Chemistry. 1993;268(32):23972–23980. [PubMed] [Google Scholar]
- 278.Matsushika A., Mizuno T. A dual-signaling mechanism mediated by the ArcB hybrid sensor kinase containing the histidine-containing phosphotransfer domain in Escherichia coli . Journal of Bacteriology. 1998;180(15):3973–3977. doi: 10.1128/jb.180.15.3973-3977.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 279.Georgellis D., Kwon O., Lin E. C. C. Amplification of signaling activity of the Arc two-component system of Escherichia coli by anaerobic metabolites. An in vitro study with different protein modules. The Journal of Biological Chemistry. 1999;274(50):35950–35954. doi: 10.1074/jbc.274.50.35950. [DOI] [PubMed] [Google Scholar]
- 280.Iuchi S., Lin E. C. C. arcA (dye), a global regulatory gene in Escherichia coli mediating repression of enzymes in aerobic pathways. Proceedings of the National Academy of Sciences of the United States of America. 1988;85(6):1888–1892. doi: 10.1073/pnas.85.6.1888. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 281.Lynch A. S., Lin E. C. C. Transcriptional control mediated by the ArcA two-component response regulator protein of Escherichia coli: characterization of DNA binding at target promoters. Journal of Bacteriology. 1996;178(21):6238–6249. doi: 10.1128/jb.178.21.6238-6249.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 282.Park S. J., McCabe J., Turna J., Gunsalus R. P. Regulation of the citrate synthase (gltA) gene of Escherichia coli in response to anaerobiosis and carbon supply: role of the arcA gene product. Journal of Bacteriology. 1994;176(16):5086–5092. doi: 10.1128/jb.176.16.5086-5092.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 283.Park S. J., Tseng C. P., Gunsalus R. P. Regulation of succinate dehydrogenase (sdhCDAB) operon expression in Escherichia coli in response to carbon supply and anaerobiosis: role of ArcA and Fnr. Molecular Microbiology. 1995;15(3):473–482. doi: 10.1111/j.1365-2958.1995.tb02261.x. [DOI] [PubMed] [Google Scholar]
- 284.Park S. J., Gunsalus R. P. Oxygen, iron, carbon, and superoxide control of the fumarase fumA and fumC genes of Escherichia coli: role of the arcA, fnr, and soxR gene products. Journal of Bacteriology. 1995;177(21):6255–6262. doi: 10.1128/jb.177.21.6255-6262.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 285.Park S. J., Chao G., Gunsalus R. P. Aerobic regulation of the sucABCD genes of Escherichia coli, which encode α-ketoglutarate dehydrogenase and succinyl coenzyme A synthetase: roles of ArcA, Fnr, and the upstream sdhCDAB promoter. Journal of Bacteriology. 1997;179(13):4138–4142. doi: 10.1128/jb.179.13.4138-4142.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 286.Iuchi S., Chepuri V., Fu H. A., Gennis R. B., Lin E. C. C. Requirement for terminal cytochromes in generation of the aerobic signal for the arc regulatory system in Escherichia coli: study utilizing deletions and lac fusions of cyo and cyd. Journal of Bacteriology. 1990;172(10):6020–6025. doi: 10.1128/jb.172.10.6020-6025.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 287.Lynch A. S., Lin E. C. C. Responses to molecular oxygen. In: Neidhardt F. C., editor. Escherichia coli and Salmonella typhimurium: Cellular and Molecular Biology. 2nd. Washington, DC, USA: ASM press; 1996. pp. 1526–1538. [Google Scholar]
- 288.Cunningham L., Georgellis D., Green J., Guest J. R. Co-regulation of lipoamide dehydrogenase and 2-oxoglutarate dehydrogenase synthesis in Escherichia coli: characterisation of an ArcA binding site in the lpd promoter. FEMS Microbiology Letters. 1998;169(2):403–408. doi: 10.1016/S0378-1097(98)00506-0. [DOI] [PubMed] [Google Scholar]
- 289.Cotter P. A., Gunsalus R. P. Contribution of the fnr and arcA gene products in coordinate regulation of cytochrome o and d oxidase (cyoABCDE and cydAB) genes in Escherichia coli . FEMS Microbiology Letters. 1992;91(1):31–36. doi: 10.1016/0378-1097(92)90558-6. [DOI] [PubMed] [Google Scholar]
- 290.Drapal N., Sawers G. Promoter 7 of the Escherichia coli pfl operon is a major determinant in the anaerobic regulation of expression by ArcA. Journal of Bacteriology. 1995;177(18):5338–5341. doi: 10.1128/jb.177.18.5338-5341.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 291.Tseng C. P., Albrecht J., Gunsalus R. P. Effect of microaerophilic cell growth conditions on expression of the aerobic (cyoABCDE and cydAB) and anaerobic (narGHJI, frdABCD, and dmsABC) respiratory pathway genes in Escherichia coli . Journal of Bacteriology. 1996;178(4):1094–1098. doi: 10.1128/jb.178.4.1094-1098.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 292.Ramseier T. M., Chien S. Y., Saier M. H., Jr. Cooperative interaction between Cra and Fnr in the regulation of the cydAB operon of Escherichia coli . Current Microbiology. 1996;33:270–274. doi: 10.1007/s002849900112. [DOI] [PubMed] [Google Scholar]
- 293.Georgellis D., Kwon O., Lin E. C. C. Quinones as the redox signal for the arc two-component system of bacteria. Science. 2001;292(5525):2314–2316. doi: 10.1126/science.1059361. [DOI] [PubMed] [Google Scholar]
- 294.Malpica R., Franco B., Rodriguez C., Kwon O., Georgellis D. Identification of a quinone-sensitive redox switch in the ArcB sensor kinase. Proceedings of the National Academy of Sciences of the United States of America. 2004;101(36):13318–13323. doi: 10.1073/pnas.0403064101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 295.Nizam S. A., Zhu J., Ho P. Y., Shimizu K. Effects of arcA and arcB genes knockout on the metabolism in Escherichia coli under aerobic condition. Biochemical Engineering Journal. 2009;44(2-3):240–250. doi: 10.1016/j.bej.2008.12.017. [DOI] [Google Scholar]
- 296.Vemuri G. N., Eiteman M. A., Altman E. Increased recombinant protein production in Escherichia coli strains with overexpressed water-forming NADH oxidase and a deleted ArcA regulatory protein. Biotechnology and Bioengineering. 2006;94(3):538–542. doi: 10.1002/bit.20853. [DOI] [PubMed] [Google Scholar]
- 297.Vemuri G. N., Altman E., Sangurdekar D. P., Khodursky A. B., Eiteman M. A. Overflow metabolism in Escherichia coli during steady-state growth: transcriptional regulation and effect of the redox ratio. Applied and Environmental Microbiology. 2006;72(5):3653–3661. doi: 10.1128/AEM.72.5.3653-3661.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 298.Nizam S. A., Shimizu K. Effects of arc A and arc B genes knockout on the metabolism in Escherichia coli under anaerobic and microaerobic conditions. Biochemical Engineering Journal. 2008;42:229–236. [Google Scholar]
- 299.Peng L., Shimizu K. Effect of fadR gene knockout on the metabolism of Escherichia coli based on analyses of protein expressions, enzyme activities and intracellular metabolite concentrations. Enzyme and Microbial Technology. 2006;38:512–520. [Google Scholar]
- 300.Schuetz R., Kuepfer L., Sauer U. Systematic evaluation of objective functions for predicting intracellular fluxes in Escherichia coli . Molecular Systems Biology. 2007;3(119) doi: 10.1038/msb4100162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 301.Crack J. C., Brun N. E. Le, Thomson A. J., Green J., Jervis A. J. Reactions of Nitric Oxide and Oxygen with the Regulator of Fumarate and Nitrate Reduction, a Global Transcriptional Regulator, during Anaerobic Growth of Escherichia coli . Methods in Enzymology. 2008;437:191–209. doi: 10.1016/S0076-6879(07)37011-0. [DOI] [PubMed] [Google Scholar]
- 302.Spiro S., Guest J. R. Adaptive responses to oxygen limitation in Escherichia coli . Trends in Biochemical Sciences. 1991;16:310–314. doi: 10.1016/0968-0004(91)90125-f. [DOI] [PubMed] [Google Scholar]
- 303.Green J., Guest J. R. Activation of FNR-dependent transcription by iron: an in vitro switch for FNR. FEMS Microbiology Letters. 1993;113(2):219–222. doi: 10.1016/0378-1097(93)90272-4. [DOI] [PubMed] [Google Scholar]
- 304.Kiley P. J., Reznikoff W. S. fnr mutants that activate gene expression in the presence of oxygen. Journal of Bacteriology. 1991;173(1):16–22. doi: 10.1128/jb.173.1.16-22.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 305.Salmon K., Hung S. P., Mekjian K., Baldi P., Hatfield G. W., Gunsalus R. P. Global gene expression profiling in Escherichia coli K12: The effects of oxygen availability and FNR. The Journal of Biological Chemistry. 2003;278(32):29837–29855. doi: 10.1074/jbc.M213060200. [DOI] [PubMed] [Google Scholar]
- 306.Williams R., Bell A., Sims G., Busby S. The role of two surface exposed loops in transcription activation by the Escherichia coli CRP and FNR proteins. Nucleic Acids Research. 1991;19(24):6705–6712. doi: 10.1093/nar/19.24.6705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 307.Ziegelhoffer E. C., Kiley P. J. In vitro analysis of a constitutively active mutant form of the Escherichia coli global transcription factor FNR. Journal of Molecular Biology. 1995;245(4):351–361. doi: 10.1006/jmbi.1994.0029. [DOI] [PubMed] [Google Scholar]
- 308.Bell A. I., Gaston K. L., Cole J. A., Busby S. J. W. Cloning of binding sequences for the Escherichia coli transcription activators, FNR and CRP: location of bases involved in discrimination between FNR and CRP. Nucleic Acids Research. 1989;17(10):3865–3874. doi: 10.1093/nar/17.10.3865. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 309.Li B., Wing H., Lee D., Wu H. C., Busby S. Transcription activation by Escherichia coli FNR protein: similarities to, and differences from, the CRP paradigm. Nucleic Acids Research. 1998;26(9):2075–2081. doi: 10.1093/nar/26.9.2075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 310.Unden G., Achebach S., Holighaus G., Tran H. Q., Wackwitz B., Zeuner Y. Control of Fnr function of Escherichia coli by O2 and reducing conditions. Journal of Molecular Microbiology and Biotechnology. 2002;4(3):263–268. [PubMed] [Google Scholar]
- 311.Marzan L. W., Siddiquee K. A. Z., Shimizu K. Metabolic regulation of fnr gene knockout Escherichia coli under oxygen limitation. Bioengineered Bugs. 2011;2(6):331–337. doi: 10.4161/bbug.2.6.16350. [DOI] [PubMed] [Google Scholar]
- 312.Greenberg J. T., Demple B. A global response induced in Escherichia coli by redox-cycling agents overlaps with that induced by peroxide stress. Journal of Bacteriology. 1989;171(7):3933–3939. doi: 10.1128/jb.171.7.3933-3939.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 313.Pomposiello P. J., Demple B. Redox-operated genetic switches: the SoxR and OxyR transcription factors. Trends in Biotechnology. 2001;19(3):109–114. doi: 10.1016/S0167-7799(00)01542-0. [DOI] [PubMed] [Google Scholar]
- 314.Gaudu P., Weiss B. SoxR, a [2Fe–2S] transcription factor, is active only in its oxidized form. Proceedings of the National Academy of Sciences of the United States of America. 1996;93(19):10094–10098. doi: 10.1073/pnas.93.19.10094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 315.Amabile-Cuevas C. F., Demple B. Molecular characterization of the soxRS genes of Escherichia coli: two genes control a superoxide stress regulon. Nucleic Acids Research. 1991;19(16):4479–4490. doi: 10.1093/nar/19.16.4479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 316.Fawcett W. P., Wolf R. E. Genetic definition of the Escherichia coli zwf “soxbox”, the DNA binding site for SoxS-mediated induction of glucose 6-phosphate dehydrogenase in response to superoxide. Journal of Bacteriology. 1995;177(7):1742–1750. doi: 10.1128/jb.177.7.1742-1750.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 317.Gaudu P., Weiss B. Flavodoxin mutants of Escherichia coli K-12. Journal of Bacteriology. 2000;182(7):1788–1793. doi: 10.1128/JB.182.7.1788-1793.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 318.Liochev S. I., Fridovich I. Fumarase C, the stable fumarase of Escherichia coli, is controlled by the soxRS regulon. Proceedings of the National Academy of Sciences of the United States of America. 1992;89(13):5892–5896. doi: 10.1073/pnas.89.13.5892. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 319.Gruer M. J., Guest J. R. Two genetically-distinct and differentially-regulated aconitases (AcnA and AcnB) in Escherichia coli . Microbiology. 1994;140(10):2531–2541. doi: 10.1099/00221287-140-10-2531. [DOI] [PubMed] [Google Scholar]
- 320.Liochev S. I., Hausladen A., Fridovich I. Nitroreductase A is regulated as a member of the soxRS regulon of Escherichia coli . Proceedings of the National Academy of Sciences of the United States of America. 1999;96(7):3537–3539. doi: 10.1073/pnas.96.7.3537. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 321.Flint D. H., Tuminello J. F., Emptage M. H. The inactivation of Fe–S cluster containing hydro-lyases by superoxide. The Journal of Biological Chemistry. 1993;268(30):22369–22376. [PubMed] [Google Scholar]
- 322.Liochev S. I., Fridovich I. The role of O2. − in the production of HO.: in vitro and in vivo. Free Radical Biology and Medicine. 1994;16(1):29–33. doi: 10.1016/0891-5849(94)90239-9. [DOI] [PubMed] [Google Scholar]
- 323.Greenberg J. T., Monach P., Chou J. H., Josephy P. D., Demple B. Positive control of a global antioxidant defense regulon activated by superoxide-generating agents in Escherichia coli . Proceedings of the National Academy of Sciences of the United States of America. 1990;87(16):6181–6185. doi: 10.1073/pnas.87.16.6181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 324.Storz G., Tartaglia L. A., Ames B. N. Transcriptional regulator of oxidative stress-inducible genes: direct activation by oxidation. Science. 1990;248(4952):189–194. doi: 10.1126/science.2183352. [DOI] [PubMed] [Google Scholar]
- 325.Kabir M. M., Shimizu K. Investigation into the effect of soxR and soxS genes deletion on the central metabolism of Escherichia coli based on gene expressions and enzyme activities. Biochemical Engineering Journal. 2006;30(1):39–47. doi: 10.1016/j.bej.2006.01.015. [DOI] [Google Scholar]
- 326.Wolf R. E., Prather D. M., Shea F. M. Growth-ratedependent alteration of 6-phosphogluconate dehydrogenase and glucose 6-phosphate dehydrogenase levels in Escherichia coli K-12. Journal of Bacteriology. 1979;139:1093–1096. doi: 10.1128/jb.139.3.1093-1096.1979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 327.Tsaneva I. R., Weiss B. soxR, a locus governing a superoxide response regulon in Escherichia coli K-12. Journal of Bacteriology. 1990;172:4197–4205. doi: 10.1128/jb.172.8.4197-4205.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 328.Hanson R. L., Rose C. Effects of an insertion mutation in a locus affecting pyridine nucleotide transhydrogenase (pnt::Tn5) on the growth of Escherichia coli . Journal of Bacteriology. 1980;141(1):401–404. doi: 10.1128/jb.141.1.401-404.1980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 329.Smith J. L. The role of gastric acid in preventing foodborne disease and how bacteria overcome acid conditions. Journal of Food Protection. 2003;66(7):1292–1303. doi: 10.4315/0362-028x-66.7.1292. [DOI] [PubMed] [Google Scholar]
- 330.Castanie-Cornet M. P., Penfound T. A., Smith D., Elliott J. F., Foster J. W. Control of acid resistance in Escherichia coli . Journal of Bacteriology. 1999;181(11):3525–3535. doi: 10.1128/jb.181.11.3525-3535.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 331.Lin J., Lee I. S., Frey J., Slonczewski J. L., Foster J. W. Comparative analysis of extreme acid survival in Salmonella typhimurium, Shigella flexneri, and Escherichia coli . Journal of Bacteriology. 1995;177(14):4097–4104. doi: 10.1128/jb.177.14.4097-4104.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 332.Rektorschek M., Buhmann A., Weeks D., et al. Acid resistance of Helicobacter pylori depends on the UreI membrane protein and an inner membrane proton barrier. Molecular Microbiology. 2000;36(1):141–152. doi: 10.1046/j.1365-2958.2000.01835.x. [DOI] [PubMed] [Google Scholar]
- 333.Foster J. W. Escherichia coli acid resistance: tales of an amateur acidophile. Nature Reviews Microbiology. 2004;2(11):898–907. doi: 10.1038/nrmicro1021. [DOI] [PubMed] [Google Scholar]
- 334.Stincone A., Daudi N., Rahman A. S., et al. A systems biology approach sheds new light on Escherichia coli acid resistance. Nucleic Acids Research. 2011;39(17):7512–7528. doi: 10.1093/nar/gkr338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 335.Richard H. T., Foster J. W. Acid resistance in Escherichia coli . Advances in Applied Microbiology. 2003;52:167–186. doi: 10.1016/S0065-2164(03)01007-4. [DOI] [PubMed] [Google Scholar]
- 336.Richard H. T., Foster J. W. Escherichia coli glutamate- and arginine-dependent acid resistance systems increase internal pH and reverse transmembrane potential. Journal of Bacteriology. 2004;86(18):6032–6041. doi: 10.1128/JB.186.18.6032-6041.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 337.Gong S., Richard H., Foster J. W. YjdE (AdiC) is the arginine:agmatine antiporter essential for arginine-dependent acid resistance in Escherichia coli . Journal of Bacteriology. 2003;185(15):4402–4409. doi: 10.1128/JB.185.15.4402-4409.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 338.Iyer R., Williams C., Miller C. Arginin-agmatine antiporter in extreme acid resistance in Escherichia coli . Journal of Bacteriology. 2003;185:6556–6561. doi: 10.1128/JB.185.22.6556-6561.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 339.Castanié-Cornet M. P., Foster J. W. Escherichia coli acid resistance: cAMP receptor protein and a 20 bp cis-acting sequence control pH and stationary phase expression of the gadA and gadBC glutamate decarboxylase genes. Microbiology. 2001;147:709–715. doi: 10.1099/00221287-147-3-709. [DOI] [PubMed] [Google Scholar]
- 340.Tanabe M., Nishio K., Iko Y., et al. Rotation of a Complex of the γ Subunit and c Ring of Escherichia coli ATP Synthase: the Rotor and Stator are Interchangeable. Journal of Biological Chemistry. 2001;276:15269–15274. doi: 10.1074/jbc.M100289200. [DOI] [PubMed] [Google Scholar]
- 341.Martin-Galiano A. J., Ferrandiz M. J., de La Campa A. G. The promoter of the operon encoding the F0F1 ATPase of Streptococcus pneumonia is inducible by pH. Molecular Microbiology. 2001;41:327–338. doi: 10.1046/j.1365-2958.2001.02597.x. [DOI] [PubMed] [Google Scholar]
- 342.Castanié-Cornet M. P., Cam K., Bastiat B., Cros A., Bordes P., Gutierrez C. Acid stress response in Escherichia coli: mechanism of regulation of gadA transcription by RcsB and GadE. Nucleic Acids Research. 2010;38(11):3546–3554. doi: 10.1093/nar/gkq097.gkq097 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 343.Marzan L. W., Hasan C. M. M., Shimizu K. Effect of acidic condition on the metabolic regulation of Escherichia coli and its phoB mutant. Microbiology. In press. [DOI] [PubMed]
- 344.Eguchi Y., Itou J., Yamane M., et al. B1500, a small membrane protein, connects the two-component systems EvgS/EvgA and PhoQ/ PhoP in Escherichia coli . Proceedings of the National Academy of Sciences. 2007;104:18712–18717. doi: 10.1073/pnas.0705768104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 345.Castelli M. E., García Véscovi E., Soncini F. C. The phosphatase activity is the target for Mg2+ regulation of the sensor protein PhoQ in Salmonella . The Journal of Biological Chemistry. 2000;275(30):22948–22954. doi: 10.1074/jbc.M909335199. [DOI] [PubMed] [Google Scholar]
- 346.Masuda N., Church G. M. Regulatory network of acid resistance genes in Escherichia coli . Molecular Microbiology. 2003;48(3):699–712. doi: 10.1046/j.1365-2958.2003.03477.x. [DOI] [PubMed] [Google Scholar]
- 347.Ma Z., Gong S., Richard H., Tucker D. L., Conway T., Foster J. W. GadE (YhiE) activates glutamate decarboxylase-dependent acid resistance in Escherichia coli K-12. Molecular Microbiology. 2003;49(5):1309–1320. doi: 10.1046/j.1365-2958.2003.03633.x. [DOI] [PubMed] [Google Scholar]
- 348.Tucker D. L., Tucker N., Ma Z., et al. Genes of the GadX-GadW regulon in Escherichia coli . Journal of Bacteriology. 2003;185(10):3190–3201. doi: 10.1128/JB.185.10.3190-3201.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 349.Sato M., Machida K., Arikado E., Saito H., Kakegawa T., Kobayashi H. Expression of outer membrane proteins in Escherichia coli growing at acid pH. Applied and Environmental Microbiology. 2000;66(3):943–947. doi: 10.1128/AEM.66.3.943-947.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 350.Sužiedeliene E., Sužiedelis K., Garbenčiute V., Normark S. The acid-inducible asr gene in Escherichia coli: transcriptional control by the phoBR operon. Journal of Bacteriology. 1999;181(7):2084–2093. doi: 10.1128/jb.181.7.2084-2093.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 351.Bunch P. K., Mat-Jan F., Lee N., Clark D. P. The ldhA gene encoding the fermentative lactate dehydrogenase of Escherichia coli . Microbiology. 1997;143:1, 187–195. doi: 10.1099/00221287-143-1-187. [DOI] [PubMed] [Google Scholar]
- 352.Hockney R. C. Recent developments in heterologous protein production in Escherichia coli . Trends in Biotechnology. 1994;12(11):456–463. doi: 10.1016/0167-7799(94)90021-3. [DOI] [PubMed] [Google Scholar]
- 353.Hoffmann F., Weber J., Rinas U. Metabolic adaptation of Escherichia coli during temperature-induced recombinant protein production: 1. Readjustment of metabolic enzyme synthesis. Biotechnology and Bioengineering. 2002;80(3):313–319. doi: 10.1002/bit.10379. [DOI] [PubMed] [Google Scholar]
- 354.Philippidis G. P., Smith T. K., Wyman C. E. Study of the enzymatic hydrolysis of cellulose for production of fuel ethanol by the simultaneous saccharification and fermentation process. Biotechnology and Bioengineering. 1993;41(9):846–853. doi: 10.1002/bit.260410903. [DOI] [PubMed] [Google Scholar]
- 355.Sternberg N. Properties of a mutant of Escherichia coli defective in bacteriophage λ head formation (groE). II. The propagation of phage λ . Journal of Molecular Biology. 1973;76(1):25–44. doi: 10.1016/0022-2836(73)90079-x. [DOI] [PubMed] [Google Scholar]
- 356.Chirico W. J., Waters M. G., Blobel G. 70 K heat shock related proteins stimulate protein translocation into microsomes. Nature. 1988;332(6167):805–810. doi: 10.1038/332805a0. [DOI] [PubMed] [Google Scholar]
- 357.Taylor W. E., Straus D. B., Grossman A. D. Transcription from a heat-inducible promoter causes heat shock regulation of the sigma subunit of E. coli RNA polymerase. Cell. 1984;38(2):371–381. doi: 10.1016/0092-8674(84)90492-6. [DOI] [PubMed] [Google Scholar]
- 358.Goff S. A., Casson L. P., Goldberg A. L. Heat shock regulatory gene htpR influences rates of protein degradation and expression of the lon gene in Escherichia coli . Proceedings of the National Academy of Sciences of the United States of America. 1984;81(21 I):6647–6651. doi: 10.1073/pnas.81.21.6647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 359.VanBogelen R. A., Vaughn V., Neidhardt F. C. Gene for heat-inducible lysyl-tRNA synthetase (lysU) maps near cadA in Escherichia coli . Journal of Bacteriology. 1983;153(2):1066–1068. doi: 10.1128/jb.153.2.1066-1068.1983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 360.Yura T., Nagai H., Mori H. Regulation of the heat-shock response in bacteria. Annual Review of Microbiology. 1993;47:321–350. doi: 10.1146/annurev.mi.47.100193.001541. [DOI] [PubMed] [Google Scholar]
- 361.Chuang S. E., Blattner F. R. Characterization of twenty-six new heat shock genes of Escherichia coli . Journal of Bacteriology. 1993;175(16):5242–5252. doi: 10.1128/jb.175.16.5242-5252.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 362.Craig E. A., Gross C. A. Is hsp70 the cellular thermometer? Trends in Biochemical Sciences. 1991;16(4):135–140. doi: 10.1016/0968-0004(91)90055-z. [DOI] [PubMed] [Google Scholar]
- 363.Gamer J., Bujard H., Bukau B. Physical interaction between heat shock proteins DnaK, DnaJ, and GrpE and the bacterial heat shock transcription factor σ 32 . Cell. 1992;69(5):833–842. doi: 10.1016/0092-8674(92)90294-M. [DOI] [PubMed] [Google Scholar]
- 364.Liberek K., Galitski T. P., Zylicz M., Georgopoulos C. The DnaK chaperone modulates the heat shock response of Escherichia coli by binding to the σ 32 transcription factor. Proceedings of the National Academy of Sciences of the United States of America. 1992;89(8):3516–3520. doi: 10.1073/pnas.89.8.3516. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 365.Tilly K., McKittrick N., Zylicz M., Georgopoulos C. The dnaK protein modulates the heat-shock response of Escherichia coli . Cell. 1983;34(2):641–646. doi: 10.1016/0092-8674(83)90396-3. [DOI] [PubMed] [Google Scholar]
- 366.Mogk A., Schlieker C., Friedrich K. L., Schönfeld H. J., Vierling E., Bukau B. Refolding of substrates bound to small Hsps relies on a disaggregation reaction mediated most efficiently by ClpB/DnaK. The Journal of Biological Chemistry. 2003;278(33):31033–31042. doi: 10.1074/jbc.M303587200. [DOI] [PubMed] [Google Scholar]
- 367.Schlieker C., Bukau B., Mogk A. Prevention and reversion of protein aggregation by molecular chaperones in the E. coli cytosol: implications for their applicability in biotechnology. Journal of Biotechnology. 2002;96(1):13–21. doi: 10.1016/S0168-1656(02)00033-0. [DOI] [PubMed] [Google Scholar]
- 368.Kitagawa M., Miyakawa M., Matsumura Y., Tsuchido T. Escherichia coli small heat shock proteins, IbpA and IbpB, protect enzymes from inactivation by heat and oxidants. European Journal of Biochemistry. 2002;269(12):2907–2917. doi: 10.1046/j.1432-1033.2002.02958.x. [DOI] [PubMed] [Google Scholar]
- 369.Sørensen H. P., Mortensen K. K. Soluble expression of recombinant proteins in the cytoplasm of Escherichia coli . Microbial Cell Factories. 2005;4, article 1 doi: 10.1186/1475-2859-4-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 370.Rosen R., Ron E. Z. Proteome analysis in the study of the bacterial heat-shock response. Mass Spectrometry Reviews. 2002;21(4):244–265. doi: 10.1002/mas.10031. [DOI] [PubMed] [Google Scholar]
- 371.Richmond C. S., Glasner J. D., Mau R., Jin H., Blattner F. R. Genome-wide expression profiling in Escherichia coli K-12. Nucleic Acids Research. 1999;27(19):3821–3835. doi: 10.1093/nar/27.19.3821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 372.Gadgil M., Kapur V., Hu W. S. Transcriptional response of Escherichia coli to temperature shift. Biotechnology Progress. 2005;21(3):689–699. doi: 10.1021/bp049630l. [DOI] [PubMed] [Google Scholar]
- 373.Kao C. M. Functional genomics technologies: creating: new paradigms for fundamental and applied biology. Biotechnology Progress. 1999;15:304–311. doi: 10.1021/bp990027q. [DOI] [PubMed] [Google Scholar]
- 374.Hasan C. M., Shimizu K. Effect of temperature up-shift on fermentation and metabolic characteristics in view of gene expressions in Escherichia coli . Microbial Cell Factories. 2008;7:35. doi: 10.1186/1475-2859-7-35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 375.Gross C. A., Straus D. B., Erickson J. W., Yura T. The function and regulation of heat shock proteins in Escherichia coli . In: Morimoto R. I., Tissières A., Cold Spring Harbor Laboratory, editors. Stress Proteins in Biology and Medicine. Cold Spring Harbor, NY, USA: Cold Spring Harbor Laboratory Press; 1990. pp. 167–189. [Google Scholar]
- 376.Yura T., Nakahigashi K. Regulation of the heat-shock response. Current Opinion in Microbiology. 1999;2(2):153–158. doi: 10.1016/S1369-5274(99)80027-7. [DOI] [PubMed] [Google Scholar]
- 377.Tilly K., Erickson J., Sharma S., Georgopoulos C. Heat shock regulatory gene rpoH mRNA level increases after heat shock in Escherichia coli . Journal of Bacteriology. 1986;168(3):1155–1158. doi: 10.1128/jb.168.3.1155-1158.1986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 378.Skelly S., Coleman T., Fu C. F., Brot N., Weissbach H. Correlation between the 32-kDa sigma factor levels and in vitro expression of Escherichia coli heat shock genes. Proceedings of the National Academy of Sciences of the United States of America. 1987;84(23):8365–8369. doi: 10.1073/pnas.84.23.8365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 379.Grossman A. D., Erickson J. W., Gross C. A. The htpR gene product of E. coli is a sigma factor for heat-shock promoters. Cell. 1984;38(2):383–390. doi: 10.1016/0092-8674(84)90493-8. [DOI] [PubMed] [Google Scholar]
- 380.Cowing D. W., Bardwell J. C. A., Craig E. A., Woolford C., Hendrix R. W., Gross C. A. Consensus sequence for Escherchia coli heat shock gene promoter. Proceedings of the National Academy of Sciences of the United States of America. 1985;82(9):2679–2683. doi: 10.1073/pnas.82.9.2679. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 381.Tilly K., Spence J., Georgopoulos C. Modulation of stability of the Escherichia coli heat shock regulatory factor σ 32 . Journal of Bacteriology. 1989;171(3):1585–1589. doi: 10.1128/jb.171.3.1585-1589.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 382.Arsène F., Tomoyasu T., Bukau B. The heat shock response of Escherichia coli . International Journal of Food Microbiology. 2000;55(1-3):3–9. doi: 10.1016/S0168-1605(00)00206-3. [DOI] [PubMed] [Google Scholar]
- 383.Shin D., Lim S., Seok Y. J., Ryu S. Heat shock RNA polymerase (Eσ 32) is involved in the transcription of mlc and Crucial for Induction of the Mlc Regulon by Glucose in Escherichia coli . The Journal of Biological Chemistry. 2001;276(28):25871–25875. doi: 10.1074/jbc.M101757200. [DOI] [PubMed] [Google Scholar]
- 384.Hosono K., Kakuda H., Ichihara S. Decreasing accumulation of acetate in a rich medium by Escherichia coli on introduction of genes on a multicopy plasmid. Bioscience, Biotechnology and Biochemistry. 1995;59(2):256–261. doi: 10.1271/bbb.59.256. [DOI] [PubMed] [Google Scholar]
- 385.Cho S., Shin D., Geun E. J., Heu S., Ryu S. High-level recombinant protein production by overexpression of Mlc in Escherichia coli . Journal of Biotechnology. 2005;119(2):197–203. doi: 10.1016/j.jbiotec.2005.03.008. [DOI] [PubMed] [Google Scholar]
- 386.Plumbridge J. Control of the expression of the manXYZ operon in Escherichia coli: Mlc is a negative regulator of the mannose PTS. Molecular Microbiology. 1998;27(2):369–380. doi: 10.1046/j.1365-2958.1998.00685.x. [DOI] [PubMed] [Google Scholar]
- 387.Kim S. Y., Nam T. W., Shin D., Koo B. M., Seok Y. J., Ryu S. Purification of Mlc and analysis of its effects on the pts expression in Escherichia coli . The Journal of Biological Chemistry. 1999;274(36):25398–25402. doi: 10.1074/jbc.274.36.25398. [DOI] [PubMed] [Google Scholar]
- 388.Kimata K., Inada T., Tagami H., Aiba H. A global repressor (Mlc) is involved in glucose induction of the ptsG gene encoding major glucose transporter in Escherichia coli . Molecular Microbiology. 1998;29(6):1509–1519. doi: 10.1046/j.1365-2958.1998.01035.x. [DOI] [PubMed] [Google Scholar]
- 389.Tanaka Y., Kimata K., Inada T., Tagami H., Aiba H. Negative regulation of the pts operon by Mlc: mechanism underlying glucose induction in Escherichia coli . Genes to Cells. 1999;4(7):391–399. doi: 10.1046/j.1365-2443.1999.00268.x. [DOI] [PubMed] [Google Scholar]
- 390.Nam T. W., Cho S. H., Shin D., et al. The Escherichia coli glucose transporter enzyme IICBGlc recruits the global repressor Mlc. The EMBO Journal. 2001;20(3):491–498. doi: 10.1093/emboj/20.3.491. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 391.Kumari S., Beatty C. M., Browning D. F., et al. Regulation of acetyl coenzyme A synthetase in Escherichia coli . Journal of Bacteriology. 2000;182(15):4173–4179. doi: 10.1128/JB.182.15.4173-4179.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 392.Lin H., Bennett G. N., San K. Y. Chemostat culture characterization of Escherichia coli mutant strains metabolically engineered for aerobic succinate production: a study of the modified metabolic network based on metabolite profile, enzyme activity, and gene expression profile. Metabolic Engineering. 2005;7(5-6):337–352. doi: 10.1016/j.ymben.2005.06.002. [DOI] [PubMed] [Google Scholar]
- 393.Browning D. F., Beatty C. M., Wolfe A. J., Cole J. A., Busby S. J. W. Independent regulation of the divergent Escherichia coli nrfA and acsP1 promoters by a nucleoprotein assembly at a shared regulatory region. Molecular Microbiology. 2002;43(3):687–701. doi: 10.1046/j.1365-2958.2002.02776.x. [DOI] [PubMed] [Google Scholar]
- 394.Beatty C. M., Browning D. F., Busby S. J. W., Wolfe A. J. Cyclic AMP receptor protein-dependent activation of the Escherichia coliacsP2 promoter by a synergistic class III mechanism. Journal of Bacteriology. 2003;185(17):5148–5157. doi: 10.1128/JB.185.17.5148-5157.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 395.Browning D. F., Beatty C. M., Sanstad E. A., Gunn K. E., Busby S. J. W., Wolfe A. J. Modulation of CRP-dependent transcription at the Escherichia coliacsP2 promoter by nucleoprotein complexes: anti-activation by the nucleoid proteins FIS and IHF. Molecular Microbiology. 2004;51(1):241–254. doi: 10.1046/j.1365-2958.2003.03824.x. [DOI] [PubMed] [Google Scholar]
- 396.Rahman M., Shimizu K. Altered acetate metabolism and biomass production in several Escherichia coli mutants lacking rpoS-dependent metabolic pathway genes. Molecular BioSystems. 2008;4(2):160–169. doi: 10.1039/b712023k. [DOI] [PubMed] [Google Scholar]
- 397.Soini J., Falschlehner C., Mayer C., et al. Transient increase of ATP as a response to temperature up-shift in Escherichia coli . Microbial Cell Factories. 2005;4, article 9 doi: 10.1186/1475-2859-4-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 398.Hoffmann F., Rinas U. Plasmid amplification in Escherichia coli after temperature upshift is impaired by induction of recombinant protein synthesis. Biotechnology Letters. 2001;23(22):1819–1825. doi: 10.1023/A:1012718200638. [DOI] [Google Scholar]
- 399.Weber J., Hoffmann F., Rinas U. Metabolic adaptation of Escherichia coli during temperature-induced recombinant protein production: 2. Redirection of metabolic fluxes. Biotechnology and Bioengineering. 2002;80(3):320–330. doi: 10.1002/bit.10380. [DOI] [PubMed] [Google Scholar]
- 400.Wittmann C., Weber J., Betiku E., Krömer J., Böhm D., Rinas U. Response of fluxome and metabolome to temperature-induced recombinant protein synthesis in Escherichia coli . Journal of Biotechnology. 2007;132(4):375–384. doi: 10.1016/j.jbiotec.2007.07.495. [DOI] [PubMed] [Google Scholar]
- 401.Privalle C. T., Fridovich I. Induction of superoxide dismutase in Escherichia coli by heat shock. Proceedings of the National Academy of Sciences of the United States of America. 1987;84(9):2723–2726. doi: 10.1073/pnas.84.9.2723. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 402.Benov L., Fridovich I. Superoxide dismutase protects against aerobic heat shock in Escherichia coli . Journal of Bacteriology. 1995;177(11):3344–3346. doi: 10.1128/jb.177.11.3344-3346.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 403.Gunasekera T. S., Csonka L. N., Paliy O. Genome-wide transcriptional responses of Escherichia coli K-12 to continuous osmotic and heat stresses. Journal of Bacteriology. 2008;190(10):3712–3720. doi: 10.1128/JB.01990-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 404.Farmer W. R., Liao J. C. Reduction of aerobic acetate production by Escherichia coli . Applied and Environmental Microbiology. 1997;63(8):3205–3210. doi: 10.1128/aem.63.8.3205-3210.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 405.van de Walle M., Shiloach J. Proposed mechanism of acetate accumulation in two recombinant Escherichia coli strains during high density fermentation. Biotechnology and Bioengineering. 1998;57(1):71–78. doi: 10.1002/(SICI)1097-0290(19980105)57:1&lt;71::AID-BIT9&gt;3.0.CO;2-S. [DOI] [PubMed] [Google Scholar]
- 406.Cronan J. E., Subrahmanyam S. FadR, transcriptional co-ordination of metabolic expediency. Molecular Microbiology. 1998;29(4):937–943. doi: 10.1046/j.1365-2958.1998.00917.x. [DOI] [PubMed] [Google Scholar]
- 407.DiRusso C. C., Heimert T. L., Metzger A. K. Characterization of FadR, a global transcriptional regulator of fatty acid metabolism in Escherichia coli . The Journal of Biological Chemistry. 1992;267(12):8685–8691. [PubMed] [Google Scholar]
- 408.DiRusso C. C., Nyström T. The fats of Escherichia coli during infancy and old age: regulation by global regulators, alarmones and lipid intermediates. Molecular Microbiology. 1998;27(1):1–8. doi: 10.1046/j.1365-2958.1998.00645.x. [DOI] [PubMed] [Google Scholar]
- 409.Fraenkel D. G. Glycolysis. In: Neidhardt F. C., editor. Escherichia coli and Salmonella: Cellular and Molecular Biology. Washington, DC, USA: AMS, Mira Digital Publishing; 1999. [Google Scholar]
- 410.Morita T., El-Kazzaz W., Tanaka Y., Inada T., Aiba H. Accumulation of glucose 6-phosphate or fructose 6-phosphate is responsible for destabilization of glucose transporter mRNA in Escherichia coli . The Journal of Biological Chemistry. 2003;278(18):15608–15614. doi: 10.1074/jbc.M300177200. [DOI] [PubMed] [Google Scholar]
- 411.D'Alessio G., Josse J. Glyceraldehyde phosphate dehydrogenase, phosphoglycerate kinase, and phosphoglyceromutase of Escherichia coli. Simultaneous purification and physical properties. The Journal of Biological Chemistry. 1971;246(13):4319–4325. [PubMed] [Google Scholar]
- 412.Nakano M. M., Zuber P., Sonenshein A. L. Anaerobic regulation of Bacillus subtilis Krebs cycle genes. Journal of Bacteriology. 1998;180(13):3304–3311. doi: 10.1128/jb.180.13.3304-3311.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 413.Murai T., Tokushige M., Nagai J., Katsuki H. Physiological functions of NAD- and NADP-linked malic enzymes in Escherichia coli . Biochemical and Biophysical Research Communications. 1971;43(4):875–881. doi: 10.1016/0006-291x(71)90698-x. [DOI] [PubMed] [Google Scholar]
- 414.Yoon S. H., Han M. J., Lee S. Y., Jeong K. J., Yoo J. S. Combined transcriptome and proteome analysis of Escherichia coli during high cell density culture. Biotechnology and Bioengineering. 2003;81(7):753–767. doi: 10.1002/bit.10626. [DOI] [PubMed] [Google Scholar]
- 415.Campbell J. W., Cronan J. E. Escherichia coli fadR positively regulates transcription of the fabB fatty acid biosynthetic gene. Journal of Bacteriology. 2001;183(20):5982–5990. doi: 10.1128/JB.183.20.5982-5990.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 416.Farewell A., Diez A. A., DiRusso C. C., Nyström T. Role of the Escherichia coli FadR regulator in stasis survival and growth phase-dependent expression of the uspA, fad, and fab genes. Journal of Bacteriology. 1996;178(22):6443–6450. doi: 10.1128/jb.178.22.6443-6450.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 417.Arnold C. N., Mcelhanon J., Lee A., Leonhard R., Siegele D. A. Global analysis of Escherichia coli gene expression during the acetate-induced acid tolerace response. Journal of Bacteriology. 2001;183(7):2178–2186. doi: 10.1128/JB.183.7.2178-2186.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 418.Kirkpatrick C., Maurer L. M., Oyelakin N. E., Yoncheva Y. N., Maurer R., Slonczewski J. L. Acetate and formate stress: opposite responses in the proteome of Escherichia coli . Journal of Bacteriology. 2001;183(21):6466–6477. doi: 10.1128/JB.183.21.6466-6477.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 419.Aronis H. R. Regulation of gene expression during entry into stationary phase. In: Neidhardt F. C., Curtiss R. III, Ingram J. L., et al., editors. Escherichia coli and Salmonella: Cellular and Molecular Biology. 2nd. Washington, DC, USA: ASM Press; 1996. pp. 1497–1512. [Google Scholar]
- 420.Aronis R. H. Signal transduction and regulatory mechanisms involved in control of the σ s (RpoS) subunit of RNA polymerase. Microbiology and Molecular Biology Reviews. 2002;66(3):373–395. doi: 10.1128/MMBR.66.3.373-395.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 421.AronisAronis H. R. Stationary phase gene regulation: what makes an Escherichia coli promoter σ s-selective? Current Opinion in Microbiology. 2002;5(6):591–595. doi: 10.1016/S1369-5274(02)00372-7. [DOI] [PubMed] [Google Scholar]
- 422.Lacour S., Landini P. σ S-dependent gene expression at the onset of stationary phase in Escherichia coli: function of σ s-dependent genes and identification of their promoter sequences. Journal of Bacteriology. 2004;186(21):7186–7195. doi: 10.1128/JB.186.21.7186-7195.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 423.Vijayakumar S. R. V., Kirchhof M. G., Patten C. L., Schellhorn H. E. RpoS-regulated genes of Escherichia coli identified by random lacZ fusion mutagenesis. Journal of Bacteriology. 2004;186(24):8499–8507. doi: 10.1128/JB.186.24.8499-8507.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 424.Wei B., Shin S., LaPorte D., Wolfe A. J., Romeo T. Global regulatory mutations in csrA and rpoS cause severe central carbon stress in Escherichia coli in the presence of acetate. Journal of Bacteriology. 2000;182(6):1632–1640. doi: 10.1128/JB.182.6.1632-1640.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 425.Rahman M., Hasan M. R., Oba T., Shimizu K. Effect of rpoS gene knockout on the metabolism of Escherichia coli during exponential growth phase and early stationary phase based on gene expressions, enzyme activities and intracellular metabolite concentrations. Biotechnology and Bioengineering. 2006;94(3):585–595. doi: 10.1002/bit.20858. [DOI] [PubMed] [Google Scholar]
- 426.Booth I. R. Regulation of cytoplasmic pH in bacteria. Microbiological Reviews. 1985;49(4):359–378. doi: 10.1128/mr.49.4.359-378.1985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 427.Weber H., Polen T., Heuveling J., Wendisch V. F., Hengge R. Genome-wide analysis of the general stress response network in Escherichia coli: σ s-dependent genes, promoters, and sigma factor selectivity. Journal of Bacteriology. 2005;187(5):1591–1603. doi: 10.1128/JB.187.5.1591-1603.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 428.Maeda H., Fujita N., Ishihama A. Competition among seven Escherichia coliσ subunits: relative binding affinities to the core RNA polymerase. Nucleic Acids Research. 2000;28(18):3497–3503. doi: 10.1093/nar/28.18.3497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 429.Kusano S., Ding Q., Fujita N., Ishihama A. Promoter selectivity of Escherichia coli RNA polymerase Eσ 70 and Eσ 38 holoenzymes: effect of DNA supercoiling. The Journal of Biological Chemistry. 1996;271(4):1998–2004. doi: 10.1074/jbc.271.4.1998. [DOI] [PubMed] [Google Scholar]
- 430.Loewen P. C., Ossowski I. V., Switala J., Mulvey M. R. KatF (σ S) synthesis in Escherichia coli is subject to posttranscriptional regulation. Journal of Bacteriology. 1993;175(7):2150–2153. doi: 10.1128/jb.175.7.2150-2153.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 431.Cortay J. C., Negre D., Galinier A., Duclos B., Perriere G., Cozzone A. J. Regulation of the acetate operon in Escherichia coli: purification and functional characterization of the IclR repressor. The EMBO Journal. 1991;10(3):675–679. doi: 10.1002/j.1460-2075.1991.tb07996.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 432.Jordan P. A., Tang Y., Bradbury A. J., Thomson A. J., Guest J. R. Biochemical and spectroscopic characterization of Escherichia coli aconitases (AcnA and AcnB) Biochemical Journal. 1999;344(3):739–746. doi: 10.1042/0264-6021:3440739. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 433.Lu C., Bentley W. E., Rao G. Comparisons of oxidative stress response genes in aerobic Escherichia coli fermentations. Biotechnology and Bioengineering. 2003;83(7):864–870. doi: 10.1002/bit.10732. [DOI] [PubMed] [Google Scholar]
- 434.Tang Y., Quail M. A., Artymiuk P. J., Guest J. R., Green J. Escherichia coli aconitases and oxidative stress: post-transcriptional regulation of sodA expression. Microbiology. 2002;148(4):1027–1037. doi: 10.1099/00221287-148-4-1027. [DOI] [PubMed] [Google Scholar]
- 435.Varghese S., Tang Y., Imlay J. A. Contrasting sensitivities of Escherichia coli aconitases A and B to oxidation and iron depletion. Journal of Bacteriology. 2003;185(1):221–230. doi: 10.1128/JB.185.1.221-230.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 436.Sprenger G. A., Schorken U., Sprenger G., Sahm H. Transaldolase B of Escherichia coli K-12: cloning of its gene, talB, and characterization of the enzyme from recombinant strains. Journal of Bacteriology. 1995;177(20):5930–5936. doi: 10.1128/jb.177.20.5930-5936.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 437.Zhao G., Winkler M. E. An Escherichia coli K-12 tktA tktB mutant deficient in transketolase activity requires pyridoxine (vitamin B6) as well as the aromatic amino acids and vitamins for growth. Journal of Bacteriology. 1994;176(19):6134–6138. doi: 10.1128/jb.176.19.6134-6138.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 438.Datta A. G., Racker E. Mechanism of action of transketolase. I. Properties of the crystalline yeast enzyme. The Journal of Biological Chemistry. 1961;236:617–623. [PubMed] [Google Scholar]
- 439.Jung I. L., Phyo K. H., Kim I. G. RpoS-mediated growth-dependent expression of the Escherichia colitkt genes encoding transketolases isoenzymes. Current Microbiology. 2005;50(6):314–318. doi: 10.1007/s00284-005-4501-1. [DOI] [PubMed] [Google Scholar]
- 440.Stryer L. Biochemistry. New York, NY, USA: W. H. Freeman; 1988. [Google Scholar]
- 441.Tseng C. P., Yu C. C., Lin H. H., Chang C. Y., Kuo J. T. Oxygen- and growth rate-dependent regulation of Escherichia coli fumarase (FumA, FumB, and FumC) activity. Journal of Bacteriology. 2001;183(2):461–467. doi: 10.1128/JB.183.2.461-467.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 442.Krapp A. R., Rodriguez R. E., Poli H. O., Paladini D. H., Palatnik J. F., Carrillo N. The flavoenzyme ferredoxin (flavodoxin)-NADP(H) reductase modulates NADP(H) homeostasis during the soxRS response of Escherichia coli . Journal of Bacteriology. 2002;184(5):1474–1480. doi: 10.1128/JB.184.5.1474-1480.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 443.Cronan J. E., Jr., LaPorte D. Tricarboxylic acid cycle and glyoxylate bypass. In: Neidhardt F. C., Curtiss R. III, Ingraham J. L., et al., editors. Escherichia coli and Salmonella: Cellular and Molecular Biology. 2nd. Vol. 1. Washington, DC, USA: American Society for Microbiology; 1996. pp. 206–216. [Google Scholar]
- 444.Shin S., Song S. G., Lee D. S., Pan J. G., Park C. Involvement of iclR and rpoS in the induction of acs, the gene for acetyl coenzyme A synthetase of Escherichia coli K-12. FEMS Microbiology Letters. 1997;146(1):103–108. doi: 10.1016/S0378-1097(96)00461-2. [DOI] [PubMed] [Google Scholar]
- 445.DiRusso C. C., Metzger A. K., Heimert T. L. Regulation of transcription of genes required for fatty acid transport and unsaturated fatty acid biosynthesis in Escherichia coli by FadR. Molecular Microbiology. 1993;7(2):311–322. doi: 10.1111/j.1365-2958.1993.tb01122.x. [DOI] [PubMed] [Google Scholar]
- 446.Marrakchi H., Zhang Y. M., Rock C. O. Mechanistic diversity and regulation of type II fatty acid synthesis. Biochemical Society Transactions. 2002;30(6):1050–1055. doi: 10.1042/BST0301050. [DOI] [PubMed] [Google Scholar]
- 447.Maloy S. R., Nunn W. D. Genetic regulation of the glyoxylate shunt in Escherichia coli K-12. Journal of Bacteriology. 1982;149(1):173–180. doi: 10.1128/jb.149.1.173-180.1982. [DOI] [PMC free article] [PubMed] [Google Scholar]