

Abstract
Background
Measurement of densities of host-seeking malaria vectors is important for estimating levels of disease transmission, for appropriately allocating interventions, and for quantifying their impact. The gold standard for estimating mosquito—human contact rates is the human landing catch (HLC), where human volunteers catch mosquitoes that land on their exposed body parts. This approach necessitates exposure to potentially infectious mosquitoes, and is very labour intensive. There are several safer and less labour-intensive methods, with Centers for Disease Control light traps (LT) placed indoors near occupied bed nets being the most widely used.
Methods
This paper presents analyses of 13 studies with paired mosquito collections of LT and HLC to evaluate these methods for their consistency in sampling indoor-feeding mosquitoes belonging to the two major taxa of malaria vectors across Africa, the Anopheles gambiae sensu lato complex and the Anopheles funestus s.l. group. Both overall and study-specific sampling efficiencies of LT compared with HLC were computed, and regression methods that allow for the substantial variations in mosquito counts made by either method were used to test whether the sampling efficacy varies with mosquito density.
Results
Generally, LT were able to collect similar numbers of mosquitoes to the HLC indoors, although the relative sampling efficacy, measured by the ratio of LT:HLC varied considerably between studies. The overall best estimate for An. gambiae s.l. was 1.06 (95% credible interval: 0.68–1.64) and for An. funestus s.l. was 1.37 (0.70–2.68). Local calibration exercises are not reproducible, since only in a few studies did LT sample proportionally to HLC, and there was no geographical pattern or consistent trend with average density in the tendency for LT to either under- or over-sample.
Conclusions
LT are a crude tool at best, but are relatively easy to deploy on a large scale. Spatial and temporal variation in mosquito densities and human malaria transmission exposure span several orders of magnitude, compared to which the inconsistencies of LT are relatively small. LT, therefore, remain an invaluable and safe alternative to HLC for measuring indoor malaria transmission exposure in Africa.
Electronic supplementary material
The online version of this article (doi:10.1186/s12936-015-0761-9) contains supplementary material, which is available to authorized users.
Keywords: Human landing catch, CDC light traps, Anopheles gambiae, Anopheles funestus
Background
Direct estimates of the rates at which humans are bitten by vector mosquitoes are invaluable indicators of humans’ risk of exposure to vector-borne pathogens. The numbers of host-seeking mosquitoes caught by trapping methods which catch the majority of vectors attacking surveyed hosts can be used to estimate the human biting rate (HBR), a basic parameter in assessing transmission of any mosquito-borne disease and malaria in particular [1]. The HBR, when multiplied by the prevalence of sporozoites in mosquitoes, gives an estimate of the entomological inoculation rate (EIR), a measure of human exposure to malaria transmission [2].
The human landing catch (HLC), in which human volunteers catch mosquitoes that land on their exposed body parts, is considered the ‘gold standard’ method for measuring exposure of humans to mosquito bites [3, 4]. However, this method is difficult to justify for most applications due to the deliberate exposure of humans to potentially infectious bites from vectors of human disease. While it is possible to protect HLC participants from Plasmodium falciparum with anti-malarial drug prophylaxis [5], the same cannot be said of most arboviruses for which preventative drugs and vaccines are not available. It is also uncomfortable, and labour-intensive to the point that it is impossible to sustain on a large scale.
Several other methods that do not require human exposure have been developed as alternatives to HLC for estimating the HBR [6]. For African malaria vectors, placing Centers for Disease Control Miniature Light Traps (LT) near human-occupied bed nets inside houses is by far the most widely used and evaluated method [7–11] but other alternatives include bed net traps [12–14], tent traps [15–19] and odour-baited traps [20].
In general, the numbers of host-seeking mosquitoes of a given taxon captured by different methods are strongly correlated but data from two methods can be highly correlated without the methods necessarily sampling from the same mosquito population [21]. If two trapping methods are sampling from the same mosquito population, albeit potentially with different trapping efficiencies, the numbers of mosquitoes they report should be proportionate over the whole range of mosquito densities. However, traps often capture zero mosquitoes during the night and mosquito densities vary over many orders of magnitude. This makes standard methods for assessing agreement between different measurement techniques [21, 22] inapplicable. The way in which the zero observations are handled can make important differences to inferences about proportionality in numbers caught [23].
The sampling efficacy for malaria vectors of LT as compared to HLC has been evaluated in several areas with diverse outcomes [8–10, 24–31]. Discrepancies between studies might be the result of differences in statistical approach, or other methodological differences, for instance in placement of the trap [32] or spatial and temporal variations in environmental conditions or the behaviour of local mosquito populations. The pooled analysis presented here is an attempt to overcome these ambiguities by analysing proportionality in numbers of mosquitoes caught and the relative trapping efficacy of LT, compared to HLC, using data from 13 separate studies from 12 diverse settings across five African countries, encompassing a wide range of vector densities.
Methods
Study sites
A dataset allowing direct comparison of indoor LT to indoor HLC in multiple sites across rural Africa (Figure 1) was compiled, mainly from multi-country studies of malaria transmission and epidemiology: the Malaria Transmission Intensity and Mortality Burden Across Africa (MTIMBA) [33] and Malaria Transmission Consortium (MTC) [34] projects. The MTIMBA studies were conducted between 2001 and 2004, including studies in Burkina Faso (Oubritenga, Kourweogo and Nouna), Tanzania (Ulanga and Rufiji) and Ghana (Navrongo). The MTC studies were conducted between 2006 and 2010, at sites in Zambia (Chisoba and Nyamumba) and Kenya (Aduoyo Miyare, Songo Rota, Kirindo and Kobala) (Table 1). Additional data for validation were obtained from earlier studies in Tanzania and from Bioko, Equatorial Guinea [24].
Figure 1.
Location of study sites. Map of Africa showing the location of study sites where data for this analysis were collected.
Table 1.
Study details
| Site | Country | Platform | Year(s) | Dominant species | References | ||
|---|---|---|---|---|---|---|---|
| An. gambiae complex | An. funestus group | ||||||
| 1 | Aduoyo Miyare | Kenya | MTC | 2009 | arabiensis | funestus | [19] |
| 2 | Chisobe | Zambia | MTC | 2009–2010 | quadriannulatus | funestus | [18] |
| 3 | Kirindo | Kenya | MTC | 2009 | arabiensis | funestus | [19] |
| 4 | Kobala | Kenya | MTC | 2009 | arabiensis | funestus | [19] |
| 5 | Kourweogo | Burkina Faso | MTIMBA | 2001–2004 | gambiae | funestus | [71] |
| 6 | Navrongo | Ghana | MTIMBA | 2001–2004 | gambiae | funestus | [62] |
| 7 | Nouna | Burkina Faso | MTIMBA | 2001–2004 | gambiae | funestus | [72] |
| 8 | Nyamumba | Zambia | MTC | 2009–2010 | quadriannulatus | funestus | [18] |
| 9 | Oubritenga | Burkina Faso | MTIMBA | 2001–2004 | gambiae | funestus | [71] |
| 10 | Rufiji | Tanzania | MTIMBA | 2001–2004 | gambiae | funestus | [73] |
| 11 | Sango Rota | Kenya | MTC | 2009 | arabiensis | funestus | [19] |
| 12 | Ulanga 2004 | Tanzania | MTIMBA | 2004 | gambiae | funestus | [74] |
| 13 | Ulanga 2006 | Tanzania | MTIMBA | 2006 | gambiae | funestus | [74] |
MTC Malaria Transmission Consortium, MTIMBA Malaria Transmission Intensity and Mortality Burden across Africa.
Light trap calibration studies
For each study, standardized mosquito sampling protocols were used [6, 19]. LT were hung beside a sleeping place where one human volunteer slept covered by a bed net, the treatment status of which was not controlled and apparently had little effect [31, 35]. The LT was hung at the foot of the sleeping place at about 1.5 m above the floor [32]. The volunteers switched the LT on before going to bed, while the mosquito collectors switched the traps off in the morning. HLCs were done by volunteers that sat indoors and outdoors collecting mosquitoes which landed on their exposed limbs, using torchlight and aspirators [3]. For most of the MTIMBA studies, two pairs of volunteers conducted the HLC at each sampling point, with one pair replacing the other after the sixth hour (e.g., at midnight if catches started at 18.00 h). Within a pair, the volunteers interchanged positions (indoors or outdoors) hourly. In the MTC studies [18, 19] and the MTIMBA studies at the Ulanga site, only one pair of volunteers conducted HLC throughout the night, indoors and outdoors, without exchanging positions. Within each hour, they collected mosquitoes for 45 min and rested for 15 min [36]. The mosquito counts were not adjusted for the resting breaks of the volunteers.
In most of the MTIMBA studies, data included up to 3 years of daily indoor LT collections together with occasional HLC collections carried out for 48 nights of trapping (i.e., 24 periods of two consecutive nights of collection) spread over a year. The standard procedure involved classification of the human population into geographical clusters of about 100 people who were living in the same area, based on each study’s demographic database. Each month, at least 30 clusters (defined as groups of people geographically centred on index persons) were selected by simple random sampling from the demographic database. Each of the selected clusters was visited in a logistically convenient order each month. Each month, the index person and three additional people recruited from a randomized list of households in the same cluster were assigned LT for each collection night. The nearest compound to the index person was selected for indoor and outdoor HLC [15, 18, 19, 37]. Collection intensity and duration varied among studies. In the Ulanga study, an initial MTIMBA study in 2004 and two distinct additional studies, separated only by a few days at the end of 2006 in the same village, were all conducted using essentially identical methodology.
Protection of human subjects and ethical approval
Ethical clearance was obtained from respective local ethical review bodies. Participants were educated on the study procedures and were made aware of the health risks involved by their participation. As precautionary measure, study participants were screened regularly for malaria infection by microscopy or rapid diagnostic test, followed by treatment of positive cases as per the local malaria treatment guidelines. In MTC studies, volunteers were given malaria prophylaxis, specifically Lariam® (mefloquine) was provided in Kenya, Malarone® (atovaquone and proguanil hydrochloride) in Tanzania and Deltaprim® (dapsone and pyrimethamine) in Zambia.
Data analysis
Agreement between the two methods was analysed by an extension of the method initially described previously [27]. Only trap counts for mosquitoes identified as members of the Anopheles gambiae complex and Anopheles funestus group that include the most important primary vectors in Africa, were included in the analysis. For each of these two important taxa, only strata (collections by two methods matched by location and night) where at least one mosquito was captured by one of the sampling methods (indoor HLC and LT) were included in the analysis. Data on outdoor HLC were excluded from the analysis as these are, in principle, not directly comparable with indoor collections by either HLC or LT. The number of strata included in the analysis varied by mosquito species taxon and study (Table 2). Those studies where less than ten strata were available per vector taxon were excluded from the analysis (Table 2).
Table 2.
Data used in the modelling and model results
| Site/species | Number of traps nights | Number of mosquitoes | Model 1 | Model 2 | |||
|---|---|---|---|---|---|---|---|
| LT | HLC | LT | HLC | αs | αs | γs | |
| An. gambiae s.l. | |||||||
| Aduoyo Miyare | 31 | 31 | 141 | 181 | 0.79 (0.63, 0.98) | 0.36 (0.15, 0.63) | 1.80 (1.29, 2.61) |
| Chisobe | 44 | 44 | 507 | 275 | 1.83 (1.58, 2.12) | 22.3 (12.3, 37.0) | 0.55 (0.45, 0.68) |
| Kirindo | 28 | 28 | 162 | 71 | 2.2 (1.69, 2.89) | 0.57 (0.27, 0.89) | 3.80 (2.16, 7.02) |
| Kobala | 17 | 18 | 18 | 25 | 0.84 (0.48, 1.43) | 0.57 (0.26, 0.95) | 2.09 (1.19, 4.43) |
| Kourweogo | 78 | 81 | 662 | 637 | 1.06 (0.95, 1.18) | 0.40 (0.27, 0.56) | 1.58 (1.38, 1.81) |
| Navrongo | 76 | 76 | 3,316 | 3,865 | 0.86 (0.82, 0.90) | 0.26 (0.19, 0.35) | 1.39 (1.29, 1.48) |
| Nouna | 78 | 80 | 1,834 | 812 | 2.39 (2.20, 2.60) | 0.37 (0.29, 0.48) | 1.58 (1.49, 1.67) |
| Nyamumba | 43 | 43 | 277 | 130 | 2.09 (1.71, 2.57) | 4.66 (2.51, 7.47) | 0.75 (0.57, 1.01) |
| Oubritenga | 108 | 113 | 771 | 791 | 0.97 (0.88, 1.08) | 0.66 (0.49, 0.88) | 1.18 (1.06, 1.32) |
| Rufiji | 6 | 6 | 27 | 24 | NA | NA | |
| Sango Rota | 32 | 32 | 31 | 87 | 0.41 (0.27, 0.60) | 0.40 (0.16, 0.87) | 0.62 (0.34, 0.99) |
| Ulanga 2004 | 22 | 44 | 1,005 | 6,403 | 0.33 (0.31, 0.36) | 1.22 (0.41, 2.72) | 0.75 (0.63, 0.91) |
| Ulanga 2006 | 36 | 18 | 4,008 | 1,477 | 1.36 (1.28, 1.44) | 0.06 (0.03, 0.11) | 2.92 (2.52, 3.40) |
| An. funestus s.l. | |||||||
| Aduoyo Miyare | 29 | 29 | 41 | 45 | 0.97 (0.65, 1.44) | 0.69 (0.40, 0.96) | 5.98 (1.84, 17.5) |
| Chisobe | 52 | 52 | 1,692 | 1,101 | 1.53 (1.43, 1.66) | 9.8 (5.95, 15.56) | 0.69 (0.6, 0.78) |
| Kirindo | 5 | 5 | 1 | 4 | NA | NA | NA |
| Kobala | 3 | 3 | 4 | 1 | NA | NA | NA |
| Kourweogo | 34 | 33 | 23 | 14 | 1.68 (0.97, 2.99) | 1.48 (0.89, 3.84) | 2.06 (0.84, 5.41) |
| Navrongo | 75 | 75 | 4,373 | 2,018 | 2.19 (2.08, 2.30) | 0.06 (0.04, 0.08) | 5.56 (5.03, 6.13) |
| Nouna | 57 | 56 | 819 | 267 | 3.20 (2.78, 3.69) | 0.42 (0.29, 0.57) | 1.96 (1.76, 2.20) |
| Nyamumba | 55 | 55 | 938 | 648 | 1.45 (1.31, 1.60) | 4.33 (2.68, 6.59) | 0.75 (0.65, 0.87) |
| Oubritenga | 62 | 61 | 41 | 56 | 0.83 (0.55, 1.22) | 0.72 (0.41, 1.05) | 1.42 (0.62, 4.75) |
| Rufiji | 5 | 5 | 37 | 1 | NA | NA | NA |
| Sango Rota | 1 | 1 | 0 | 1 | NA | NA | NA |
| Ulanga 2004 | 22 | 40 | 41 | 70 | 1.13 (0.79, 1.61) | 0.63 (0.35, 0.97) | 5.49 (1.46, 13.3) |
| Ulanga 2006 | 22 | 11 | 24 | 4 | 2.20 (1.11, 5.21) | 1.45 (0.56, 3.02) | 3.25 (0.98, 18.1) |
Numbers between parentheses are 95% credible intervals.
site specific sampling efficacy, exponent testing proportionality, NA not analysed because less than ten trap nights of data were available.
In order to estimate the sampling efficiencies of LT relative to HLC, the following statistical model was used:
| 1 |
where is the expected number of mosquitoes of a given taxon caught using LT in stratum i; is the expected number of mosquitoes of the same taxon caught using the human landing method in the same stratum i; is the relative sampling efficacy corresponding to study s, compared to HLC for which the value is set to unity. The underlying mosquito density is assumed to have a log-normal distribution, i.e., , Poisson errors were assumed in the observed numbers of mosquitoes caught by any of the two methods so that: and: and the model therefore assumes the distribution of the numbers of mosquitoes caught by any method to be a log-normal mixture of Poisson distributions, and that HLC gives an unbiased estimate of the true mosquito density.
To allow for stochastic variation between studies, and to obtain an estimate of the overall average sampling efficacy across studies, the logarithms of the study-specific sampling efficacy, , were assumed to vary normally about the overall average of the species complex, , i.e.: , thus leading to a hierarchical statistical model which was fitted using a Bayesian Markov chain Monte Carlo algorithm in the software WinBUGS version 1.4 [38]. The parameters , , , , and were assigned weakly informative prior distributions which constrained them to be positive but otherwise did not impose important constraints. The choice of prior between several different distributional forms was found to make negligible difference to the results.
To examine whether the sampling efficacy varied with the average mosquito density, the following extended model was also fitted:
| 2 |
where is an exponent corresponding to study s. A value of different from unity indicates a lack of proportionality between the mosquito sampling methods. In addition, will differ from if is different from unity.
The validity of the calibration was checked using published data sets from three sites on Bioko Island in Equatorial Guinea with an An. gambiae sensu stricto (s.s.) and Anopheles melas mixture [24] and for an urban site (Dar es Salaam in Tanzania) (99% An. gambiae s.s. and 1% Anopheles arabiensis) [15] by comparing with the credible intervals of models fitted to the pooled multi-study data resource described above.
Results and discussion
The most abundant vector taxon sampled by either method across the sites and studies was An. gambiae sensu lato (s.l.) (Table 2). In the sites in Zambia (Chisoba and Nyamumba) An. funestus s.l. was more often sampled than An. gambiae s.l. For An. gambiae s.l., 12 studies were included, whereas for An. funestus s.l., only nine studies provided sufficient data. Forest plots (Figure 2) show point and interval estimates from the linear model of the relative sampling efficacy (Eq. 1) in each study, as well as the estimate of the overall average sampling efficacy. LT often collected greater numbers of mosquitoes than HLC (in seven out of 12 studies, LT caught more An. gambiae s.l. than HLC, and in seven out of nine studies, LT caught more An. funestus s.l. than HLC), although this relation varied between studies, even between sites adjacent to each other in Kenya or Zambia, and between sequential studies within a single site in Ulanga, Tanzania. In a comparison between the An. gambiae complex and the An. funestus group, the relative sampling efficacy of LT was greater for An. funestus s.l. than for An. gambiae s.l. for the studies in Navrongo and Ulanga 2004, but lesser for Nyamumba. For the other six studies where a comparison between species complexes was possible, the 95% credible intervals of the sampling efficacy estimates for these two vector taxa overlapped. The overall estimate of sampling efficacy of LT relative to HLC, estimated by pooling data from all studies and allowing for stochastic variability between studies, represented by the dashed vertical lines in Figure 2, was close to unity for both taxa [ = 1.06 (0.68–1.64) for An. gambiae s.l. and = 1.37 (0.70–2.68) for An. funestus s.l.], meaning that on average it makes little difference whether LT collections are assumed equivalent to human biting rates, or if a conversion factor is used.
Figure 2.
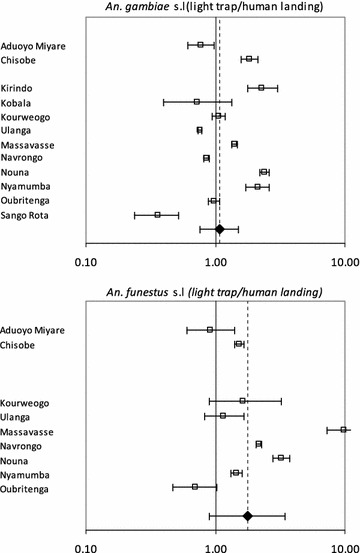
Study specific and overall estimates of sampling efficacy. Forest plots giving the estimated sampling efficacy of LT relative to HLC on a logarithmic scale, point estimates and 95% credible intervals of model (1) for An. gambiae s.l. (top panel) and An. funestus s.l. (bottom panel). The dashed vertical lines indicate the best estimates of the overall average sampling efficiencies.
Funnel plots (Figure 3) [39] were used to examine evidence for selection bias in these results. The points in these plots are expected to form a triangular pattern centred on the best estimate of the average sampling efficacy, which corresponds to the vertical line. An asymmetric funnel would indicate a relationship between treatment effect and study size, suggesting either a selection bias or a systematic difference between smaller and larger studies. In Figure 3, a large proportion of the points fall outside the dashed triangles, indicating that there was much more variation between studies in the estimated sampling efficacy than was expected if the true value of the efficacy was the same in each study. However, there is no indication of any systematic bias either upwards or downwards in the averages, since there are points scattered either side of the vertical lines, more or less independently of the standard error.
Figure 3.
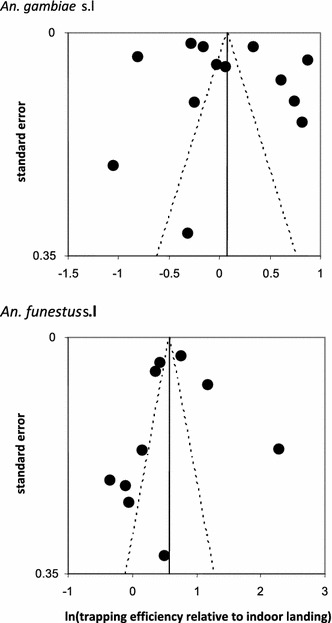
Representativeness of studied sites. Funnel plots giving the logarithm of the estimated sampling efficacy for each study (horizontal axis), standard error (se) of this estimate (vertical axis). The vertical line corresponds to the estimated overall average sampling efficacy. The dashed lines correspond to 95% pseudo-confidence limits calculated as 1.96 ± se within which 95% of the points are expected to occur in the event that the differences between studies arise only because of sampling variation.
Figure 4 panels a and b and plots in Additional file 1 show the fitted linear relationships (Eq. 1) for each study, indicating how the variation shown in Figure 2, translates into different lines relating the numbers of mosquitoes caught by the two methods. Figure 4 panels c and d and plots in Additional file 1 show the fitted non-linear relationship (Eq. 2) between both methods in the numbers of mosquitoes collected. The comparisons between the lines in Figure 4a and the curves in Figure 4c indicate how much the relative sampling efficiencies for An. gambiae s.l. depend on mosquito density. Similarly, Figure 4b and d allow the corresponding comparisons for An. funestus. There is considerable variation among studies in the shapes of the curves in Figure 4c or d and few of the curves for the individual studies are close to being straight lines. For An. gambiae s.l., out of 12 studies included in the analysis, the only study in which the 95% credible interval estimate for included the value of unity, so proportionality could not be excluded, was Nyamumba in Zambia. For eight of the 12 studies, the LT:HLC ratio increased as the number of mosquitoes increased (corresponding to ), and for three studies (Chisobe, Sango Rota and Ulanga 2004), it decreased (Table 2). Interestingly, this ratio increased with mosquito density in the Ulanga 2006 study, even though this study was done in the same location as the Ulanga 2004 study in which it decreased. Similarly, while this ratio increased with mosquito density at three of the studies in the same region of western Kenya (Aduoyo Miyare, Kirindo, Kobala), it decreased in the fourth (Sango Rota).
Figure 4.
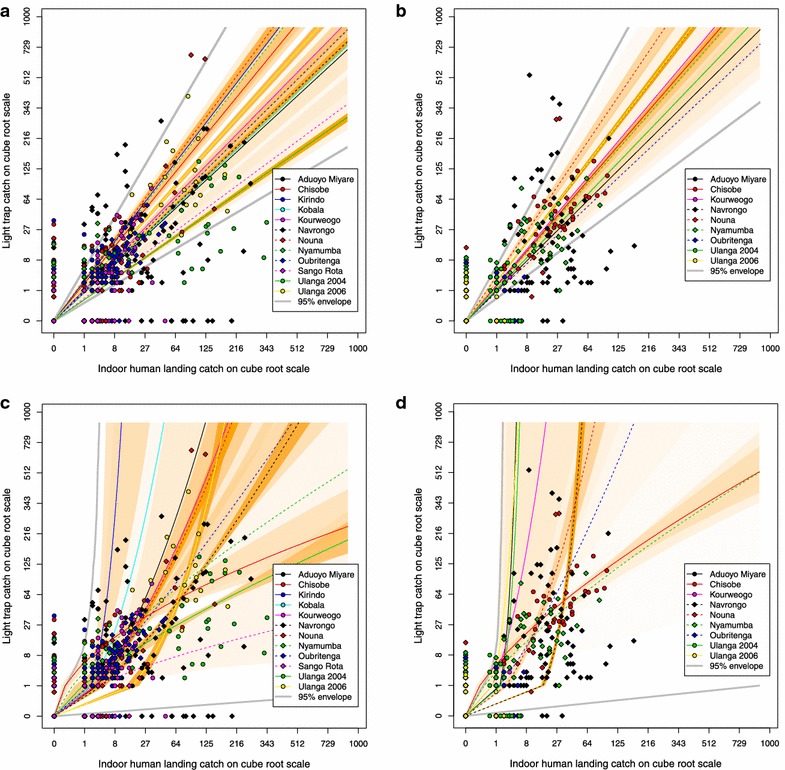
Fitted sampling efficiencies of light traps relative to human landing catches. Each point corresponds to a single matched set of mosquito collections; the shaded orange polygons correspond to 95% credible intervals for the best fitting curves, estimated for each site, with the transparency of the orange colouring inversely proportional to the surface area of the polygon. The grey lines demarcate the envelope within which 95% of fitted curves of unobserved studies are expected to fall based the variation observed in the studies used in this analysis (based on the joint posterior distributions of and ). Panel a shows the linear model for An. gambiae; panel b shows the linear model for An. funestus; panel c shows the power model for An. gambiae, and panel d shows the power model for An. funestus.
For An. funestus s.l., out of the nine studies included in the analysis, the 95% credible interval estimate for included the value of unity for only three studies (Kourweogo, Oubritenga and Ulanga 2006) indicating that proportionality could not be excluded for these studies (Table 2). In four other studies (Aduoyo Miyare, Navrongo, Nouna and Ulanga 2004), the relative sampling efficacy of LT increased with increasing mosquito density (i.e., the slope in Figure 4 increased as the density increased). In the two remaining studies (Chisobe and Nyamumba), the slope decreased as mosquito density increased. No consistent geographical pattern was observed in the relative sampling efficacy of LT in sampling either species. In addition to the contrasting results for An. gambiae s.l. of the two sequential studies at the same village in Ulanga, the relationship varied also strongly among studies in geographically close sites, such as those located in neighbouring districts of western Kenya and Burkina Faso. Where was significantly different from unity for both species complexes, this was in the same direction for both species ( for Aduoyo Miyare, Navrongo and Nouna, and for Chisobe).
Despite the very limited precision of LT and inconsistent relationship with HLC, they may nevertheless be useful for comparing vector densities across different locations and time periods: mean catch per trap per night with LT was closely correlated to the mean catch per person per night by HLC for both An. gambiae s.l. and An. funestus s.l. The potential utility of the hierarchical models, assuming either simple linear proportionality or density-dependent relative sensitivity curves, was therefore explored to determine how best this multi-study dataset of unprecedented size might be exploited to generate approximate but broadly applicable calibration functions, for LT catches compared with HLC, with which to estimate human indoor exposure rates to malaria mosquitoes.
The hierarchical models allowing for deviations from proportionality clearly fit better than the simpler linear relationships, with the deviance information criterion (DIC) being much lower for the non-linear model (Table 3). While the Bayesian approach does not formally provide p values, these DIC values indicate that the improved fit of the power model cannot be attributed to chance alone. However, the variation in shapes of the multiple fitted curves within this hierarchical model (Figure 4; Table 2) result in very wide credibility intervals. After comparison of the results of these multi-study hierarchical models with the Bioko and Dar es Salaam data sets, it is clear that while the credibility intervals of the simple linear models are very wide, they are nevertheless precise enough to distinguish one vector population that responds to LT in a typical fashion and three that do not. The data from An. melas in Arena Blanca (Luba), Bioko, fit within the credible interval of the linear hierarchical model and are, therefore, very approximately consistent with the levels of relative trapping efficacy observed in these 13 studies from other rural areas (Additional file 2). However, in the two other sites on Bioko, Mongola and Riaba, and also in urban Dar es Salaam, where relative sensitivity of LT was so poor that it forced local surveillance teams to develop their own tent trap [15–17, 40], all the data lie below the lower boundary of the credibility interval for the linear model (Figures 2.2–2.4 in Additional file 2). In Mongola, Riaba and Dar es Salaam, LT are inadequate as a method for measuring host-seeking anopheline densities, unlike in the 13 rural studies to which the models were fitted. The site in Mongola is lit up at night due to a lit road and an airport, and also urban Dar es Salaam is lit up. However, Riaba is a rural site so night lights do not appear to be a likely explanation for the poor performance of LT at this site.
Table 3.
Summary of fit of statistical models
| Model | Deviance (ergodic average) | Effective number of parameters () | Deviance information criterion (DIC) [75] |
|---|---|---|---|
| Model 1 | 26,579.7 | 854.4 | 27,434.2 |
| Model 2 | 22,656.8 | 867.9 | 23,524.6 |
The model with the lower DIC (Model 2) is the best fitting.
Even though the simple linear hierarchical model fit is also imprecise, the linear model can be used to compare to calibration data from other study sites, to determine whether LT catch malaria vector mosquitoes with sufficient efficacy to be useful and to very approximately relate these to biting rates experienced by exposed humans sleeping indoors. An R script is provided with an example (Additional files 3, 4) to allow other investigators to determine whether their locally-relevant calibration data are consistent with the observation of very approximate simple mean equivalence across these 13 geographically diverse studies.
As with other comparisons of LT with HLC in Africa [8, 9, 24], these results lead to a conclusion that the numbers of mosquitoes caught in LT placed beside human-occupied bed nets are only a very approximate indicators of the rates at which unprotected humans are exposed to mosquito bites. There were wide variations in the relative sampling efficiencies of LT and HLC, both across studies, even if they were very close to each other, and within study sites as time passes. While imprecision of the observations made with either method are to be expected based on the strong aggregation or overdispersion that is typical of mosquito catch data [35, 41–43], the matched experimental designs applied in each of these studies should yield data that are unbiased with respect to sampled times, locations and houses, natural variations in attractiveness [44–46] and technical skills of individual of human volunteers within studies, providing the relative sampling efficacy remains constant.
With both methods, it is challenging to trap mosquitoes in truly representative locations. The original procedures for HLC recognized the need to allow for differences in location between mosquito collectors and the general human population [47] (in particular in terms of whether they are inside or outside houses), but studies that adjust estimates of exposure for the use of interventions (such as repellents, house-screening and bed nets) [48, 49] have been the exception. Although it remains the standard, there is no obvious reason why HLC should be more reproducible than LT, especially as they are not automated and rely heavily on manual execution by volunteers working at times of the night when human performance of any active task is least effective [50, 51]. The substantial variations between studies in relative trapping efficacy are therefore unsurprising given the fundamentally crude nature of both techniques as applied in real-world situations [4, 6] and the difficulties in standardizing them [52]. Subtle differences in procedures could therefore explain some of the variation between studies.
Different Anopheles species are known to respond differently to LT. Studies of Cuban [25], Brazilian [26], Papua New Guinean (PNG) [27, 53] and Venezuelan [28] vectors have found LT to catch far fewer Anopheles than HLC, while with other vectors (in particular in a study of Anopheles fluviatilis in India [29] and another study of Malaysian vectors [30]) relative trapping efficacy has proven highly variable. In general, LT have been used mainly to capture African malaria vectors for which they appear to be relatively reliable, but a single mosquito taxon may react to LT very differently in nearby locations. In coastal Tanzania, LT appear to work well in all rural areas [10, 54] (Figure 4) but not in Dar es Salaam [15, 16], even though the most common malaria vector in all these studies are predominantly An. gambiae s.s., for which no geographic gene flow barriers are apparent along the mainland coast [55]. This suggests that temporal and geographic variations in the general environment and the dynamic mosquito populations it supports differentially affects the performance of the different methods, and/or that that there are subtle variations within the same taxon in terms of the innate mosquito characteristics that affect relative trapping efficacy. Most method comparison studies do not distinguish between species within the An. gambiae s.l. complex or the An. funestus s.l. group, even though they may well behave quite differently when they encounter either a LT beside an occupied bed net or an alert human sitting on a stool with a torch and aspirator. Sibling species have been reported as differentially sampled by the two techniques [15] and there may well also be relevant genetic variations within sibling species. For instance, LT may be biased towards sampling mosquitoes that are infected with malaria sporozoites [9, 10, 27], although infection rates are too low for this to explain most of the variation observed between studies here. The curvilinear relationships of relative trapping efficacy with density imply that some factor(s) affecting trapping efficacy vary with density within a mosquito population, although it is not known to what extent the sub-species composition changed over the seasons. This conclusion is reinforced by the different results of the two different Ulanga studies in the same place, separated only by a short period and using essentially identical methodologies. Given the dynamic nature of mosquito populations, and the demographic inevitability that peak mosquito densities are associated with recent emergences and relatively young populations, while low densities are often associated with aging populations [56], it may well be that much of the highly variable density-dependence observed here results from differential responsiveness of mosquitoes of different ages and/or physiological status.
Fortunately, it is not necessary to measure mosquito biting densities or EIR with any great degree of precision and even a very approximate estimate can be remarkably informative as an explanatory variable in epidemiological studies. Observed annual mean values for these indices of human exposure to mosquito bites and sporozoite inoculation exposure, respectively, vary spectacularly across logarithmic scales, spanning approximately four orders of magnitude [2, 57, 58]. Compared with this enormous range of variation in mean true values, even the variations in trapping efficacy described above are relatively small, so for most practical purposes it is reasonable to assume that LT catches are proportional, and approximately equivalent to the exposure of unprotected humans sleeping indoors. Over much of the range of transmission intensities of malaria in Africa there is saturation of prevalence in human populations so epidemiologically relevant measures of malaria prevalence, incidence, mortality and transmission stability are relatively insensitive to small changes in exposure. At measurable levels of EIR, large changes in transmission are required to achieve substantial changes in epidemiologically relevant indicators of malaria morbidity, mortality and parasite populations [2, 57, 58], so even the very loose approximation to proportionality suggested in Figure 4 is probably satisfactory for estimating transmission exposure in the vast majority of cases. So when HBR are estimated from LT catches, it is unlikely to make much practical difference how any conversion factor may be selected.
Malaria control and elimination require an indicator of malaria transmission exposure with sufficient precision to select appropriate intervention efforts. Considering the scales over which variations in the true mean EIR and the entomological measurement precision occur, the indicator on the entomological scale does not need not be very precise. The problem is analogous to selecting a rifle for hunting on the basis of the anticipated size of game animal pursued. A rifle selected to shoot a duck will suffice for any duck, regardless of whether it is a large or small one, spanning a three- to four-fold range of body mass. Similarly, a gun selected to shoot an impala, buffalo or elephant will suffice for any animal within the natural size range of that species. However, it is literally vitally important to distinguish between these four different target species which differ in size by approximately an order of magnitude. LT may, therefore, be very useful for measuring approximate biting or inoculation rates, and also inter-annual and spatial variation in these. Where available, they also provide better measures of seasonality in transmission than do indices measured in humans such as prevalence (where the oscillations are dampened owning to persistence of infection), or clinical incidence (where seasonality also depends on acquired immunity). The approximate proportionality in LT and HLC on average suggests that seasonal patterns estimated by different entomological approaches are likely to concur.
LT have proven invaluable for measuring malaria transmission in a wide diversity of contexts. Most of the EIR estimates used to support modern quantitative understanding of malaria were measured with LT and paint a remarkably consistent picture despite their limited precision [2, 57–59]. LT provide a low-cost way of achieving high sampling intensities for malaria vectors, most recently in longitudinal, community-based trapping schemes using solar-powered battery chargers [60]. They have been used many times to map vector-biting densities on very fine spatial scales [35, 41–43, 61–63]. Exposures estimated from LT data have been related with epidemiological outcomes in various settings in Africa [21, 60, 64–68] and provide valuable entomological measures of the impact of vector control on human exposure [69, 70].
Conclusions
Overall, LT are an imprecise tool for measuring human exposure to indoor-biting African malaria vectors: variability between studies appears so important that no single effort to calibrate LT against HLC can be considered reliable: even where a single research team conducted sequential studies at the same site (Ulanga, Tanzania and both Zambian sites) or parallel studies at several neighbouring locations (all Zambian and Kenyan study sites), none of these studies yielded results that were consistent with each other. However, it is equally true to say that no consistent difference between the trapping efficacy of these two methods for estimating human exposure could be demonstrated, nor could any consistent relationship between relative trapping efficacy and vector biting density be demonstrated. The most practical and parsimonious approach to relating indoor LT catches to indoor human exposure appears to be to simply consider them as very approximately proportional and equivalent.
Local calibration exercises could be used to validate or invalidate the assumption of proportionality and equivalence by comparing the locally estimated mean sampling efficacy with the credibility intervals of the hierarchical simple linear model fitted to data from these 13 calibration studies carried out in rural areas distributed across Africa. If the local calibration estimate is below the lower limit of these multi-study credibility intervals, as was the case in one outlier study from an urban setting, LT most probably lack sufficient sensitivity for routine use in that context and alternative trapping methods should be evaluated. However, given that only large variations in transmission exposure have appreciable epidemiological relevance, and exposure may vary over four orders of magnitude in endemic areas of Africa, even such imprecise, approximate entomological estimates of biting or inoculation rates may nevertheless be useful. The clear track record of LT documented in the literature confirms that they remain an invaluable tool for studying malaria vector ecology and transmission epidemiology, so long as the substantial inconsistencies in trapping efficacy they exhibit are carefully considered in experimental design and data interpretation. Except where proven otherwise by comparison with the simple linear hierarchical model fit provided here (Additional files 2, 3, 4), LT may be considered a safe and approximately equivalent alternative to HLC for measuring indoor exposure to bites and malaria transmission by African vectors.
Authors’ contributions
OJTB, BJH, TAS and GFK drafted the manuscript; OJTB, BJH and TAS performed the analysis; JEG, NB, AS, CHS, NG, DAD, and SA planned and implemented the field studies. All authors read and approved the final manuscript.
Acknowledgements
The MTIMBA project was initiated by the International Network for the Demographic Evaluation of Populations and Their Health (INDEPTH) and was financed by the Multinational Initiative for Malaria/The Special Programme for Research and Training in Tropical Diseases Research and the Roll Back Malaria Programme. The MTC project was funded by the Bill and Melinda Gates Foundation (Award 45114, coordinated by Professor Frank Collins and Dr Neil Lobo at Notre Dame University, South Bend, IN, USA). Studentship for BH was financed by the Stipendienkommission für Nachwuchs-Kräfte aus Entwicklungsländern, Basel, Switzerland. The authors thank Japhet Kihonda for help with the field studies. OJTB was supported through the Malaria Modelling Project #OPP1032350, funded by the Bill and Melinda Gates Foundation. This manuscript has been published with permission from the director, KEMRI. The findings and conclusions in this study are those of the authors and do not necessarily represent the views of the Centers for Disease Control and Prevention.
Compliance with ethical guidelines
Competing interests The authors declare that they have no competing interests.
Additional files
Site specific plots.
Plots for additional data.
R script.
Sample data.
Contributor Information
Olivier J T Briët, Email: olivier.briet@unibas.ch.
Bernadette J Huho, Email: bjohn@ihi.or.tz.
John E Gimnig, Email: hzg1@cdc.gov.
Nabie Bayoh, Email: NBayoh@kemricdc.org.
Aklilu Seyoum, Email: Aklilu_Seyoum@africairs.net.
Chadwick H Sikaala, Email: chadsikaala@gmail.com.
Nicodem Govella, Email: govella@ihi.or.tz.
Diadier A Diallo, Email: ddiallo@path.org.
Salim Abdullah, Email: sabdulla@ihi.or.tz.
Thomas A Smith, Email: thomas.a-smith@unibas.ch.
Gerry F Killeen, Email: Gerry.Killeen@lstmed.ac.uk.
References
- 1.Macdonald G. The epidemiology and control of malaria. London: Oxford University Press; 1957. [Google Scholar]
- 2.Beier JC, Killeen G, Githure JI. Entomologic inoculation rates and Plasmodium falciparum malaria prevalence in Africa. Am J Trop Med Hyg. 1999;61:109–113. doi: 10.4269/ajtmh.1999.61.109. [DOI] [PubMed] [Google Scholar]
- 3.WHO . Manual on practical entomology in malaria. Part II: methods and techniques. Geneva: World Health Organization; 1975. [Google Scholar]
- 4.Service MW. A critical review of procedures for sampling populations of adult mosquitoes. Bull Entomol Res. 1977;67:343–382. doi: 10.1017/S0007485300011184. [DOI] [Google Scholar]
- 5.Gimnig JE, Walker ED, Otieno P, Kosgei J, Olang G, Ombok M, et al. Incidence of malaria among mosquito collectors conducting human landing catches in western Kenya. Am J Trop Med Hyg. 2013;88:301–308. doi: 10.4269/ajtmh.2012.12-0209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Silver JB. Mosquito ecology: field sampling methods. 3. New York: Springer; 2008. [Google Scholar]
- 7.Odetoyinbo JA. Preliminary investigation on the use of a light-trap for sampling malaria vectors in the Gambia. Bull World Health Organ. 1969;40:547–560. [PMC free article] [PubMed] [Google Scholar]
- 8.Lines J, Curtis CF, Wilkes TJ, Njunju EM. Monitoring human-biting mosquitoes (Diptera: Culicidae) in Tanzania with light traps hung beside mosquito nets. Bull Entomol Res. 1991;81:77–84. doi: 10.1017/S0007485300053268. [DOI] [Google Scholar]
- 9.Mbogo CN, Glass GE, Forster D, Kabiru EW, Githure JI, Ouma J, et al. Evaluation of light traps for sampling anopheline mosquitoes in Kilifi, Kenya. J Am Mosq Control Assoc. 1993;9:260–263. [PubMed] [Google Scholar]
- 10.Davis JR, Hall T, Chee EM, Majala A, Minjas J, Shiff CJ. Comparison of sampling anopheline mosquitoes by light-trap and human-bait collections indoors at Bagamoyo, Tanzania. Med Vet Entomol. 1995;9:249–255. doi: 10.1111/j.1365-2915.1995.tb00130.x. [DOI] [PubMed] [Google Scholar]
- 11.Costantini C, Sagnon NF, Sanogo E, Merzagora L, Coluzzi M. Relationship to human biting collections and influence of light and bednet in CDC light-trap catches of West African malaria vectors. Bull Entomol Res. 1998;88:503–511. doi: 10.1017/S000748530002602X. [DOI] [Google Scholar]
- 12.Mathenge EM, Killeen G, Oulo DO, Irungu LW, Ndegwa PN, Knols B. Development of an exposure-free bednet trap for sampling Afrotropical malaria vectors. Med Vet Entomol. 2002;16:67–74. doi: 10.1046/j.0269-283x.2002.00350.x. [DOI] [PubMed] [Google Scholar]
- 13.Mathenge EM, Omweri GO, Irungu LW, Ndegwa PN, Walczak E, Smith TA, et al. Comparative field evaluation of the Mbita trap, the Centers for Disease Control light trap, and the human landing catch for sampling of malaria vectors in western Kenya. Am J Trop Med Hyg. 2004;70:33–37. [PubMed] [Google Scholar]
- 14.Mathenge EM, Misiani GO, Oulo DO, Irungu LW, Ndegwa PN, Smith TA, et al. Comparative performance of the Mbita trap, CDC light trap and the human landing catch in the sampling of Anopheles arabiensis, An. funestus and culicine species in a rice irrigation in western Kenya. Malar J. 2005;4:7. doi: 10.1186/1475-2875-4-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Govella NJ, Chaki PP, Geissbuhler Y, Kannady K, Okumu F, Charlwood JD, et al. A new tent trap for sampling exophagic and endophagic members of the Anopheles gambiae complex. Malar J. 2009;8:157. doi: 10.1186/1475-2875-8-157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Govella NJ, Chaki PP, Mpangile JM, Killeen GF. Monitoring mosquitoes in urban Dar es Salaam: evaluation of resting boxes, window exit traps, CDC light traps, Ifakara tent traps and human landing catches. Parasit Vectors. 2011;4:40. doi: 10.1186/1756-3305-4-40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Chaki PP, Mlacha Y, Msellemu D, Muhili A, Malishee AD, Mtema ZJ, et al. An affordable, quality-assured community-based system for high-resolution entomological surveillance of vector mosquitoes that reflects human malaria infection risk patterns. Malar J. 2012;11:172. doi: 10.1186/1475-2875-11-172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Sikaala CH, Killeen GF, Chanda J, Chinula D, Miller JM, Russell TL, et al. Evaluation of alternative mosquito sampling methods for malaria vectors in Lowland South-East Zambia. Parasit Vectors. 2013;6:91. doi: 10.1186/1756-3305-6-91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Wong J, Bayoh N, Olang G, Killeen GF, Hamel MJ, Vulule JM, et al. Standardizing operational vector sampling techniques for measuring malaria transmission intensity: evaluation of six mosquito collection methods in western Kenya. Malar J. 2013;12:143. doi: 10.1186/1475-2875-12-143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Jawara M, Awolola TS, Pinder M, Jeffries D, Smallegange RC, Takken W, et al. Field testing of different chemical combinations as odour baits for trapping wild mosquitoes in The Gambia. PLoS One. 2011;6:e19676. doi: 10.1371/journal.pone.0019676. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Bland JM, Altman DG. Statistical methods for assessing agreement between two methods of clinical measurement. Lancet. 1986;1:307–310. doi: 10.1016/S0140-6736(86)90837-8. [DOI] [PubMed] [Google Scholar]
- 22.Bland JM, Altman DG. Measuring agreement in method comparison studies. Stat Methods Med Res. 1999;8:135–160. doi: 10.1191/096228099673819272. [DOI] [PubMed] [Google Scholar]
- 23.Smith T. Proportionality between light trap catches and biting densities of malaria vectors [letter; comment] J Am Mosq Control Assoc. 1995;11:377–378. [PubMed] [Google Scholar]
- 24.Overgaard HJ, Saebo S, Reddy MR, Reddy VP, Abaga S, Matias A, et al. Light traps fail to estimate reliable malaria mosquito biting rates on Bioko Island, Equatorial Guinea. Malar J. 2012;11:56. doi: 10.1186/1475-2875-11-56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Marquetti MC, Navarro A, Bisset J, Garcia FA. Comparison of three catching methods for collecting anopheline mosquitoes. Mem Inst Oswaldo Cruz. 1992;87:457–458. doi: 10.1590/S0074-02761992000300023. [DOI] [PubMed] [Google Scholar]
- 26.Lima JB, Rosa-Freitas MG, Rodovalho CM, Santos F, Lourenco-de-Oliveira R. Is there an efficient trap or collection method for sampling Anopheles darlingi and other malaria vectors that can describe the essential parameters affecting transmission dynamics as effectively as human landing catches?—A review. Mem Inst Oswaldo Cruz. 2014;109:685–705. doi: 10.1590/0074-0276140134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Hii JL, Smith T, Mai A, Ibam E, Alpers MP. Comparison between anopheline mosquitoes (Diptera: Culicidae) caught using different methods in a malaria endemic area of Papua New Guinea. Bull Entomol Res. 2000;90:211–219. doi: 10.1017/S000748530000033X. [DOI] [PubMed] [Google Scholar]
- 28.Rubio-Palis Y. Evaluation of light traps combined with carbon dioxide and 1-octen-3-ol to collect anophelines in Venezuela. J Am Mosq Control Assoc. 1996;12:91–96. [PubMed] [Google Scholar]
- 29.Gunasekaran K, Jambulingam P, Sadanandane C, Sahu SS, Das PK. Reliability of light trap sampling for Anopheles fluviatilis, a vector of malaria. Acta Trop. 1994;58:1–11. doi: 10.1016/0001-706X(94)90116-3. [DOI] [PubMed] [Google Scholar]
- 30.Parsons RE, Dondero TJ, Cheong WH. Comparison of CDC miniature light traps and human biting collections for mosquito catches during malaria vector surveys in peninsular Malaysia. Mosquito News. 1974;34:211–213. [Google Scholar]
- 31.Fornadel CM, Norris LC, Norris DE. Centers for Disease Control light traps for monitoring Anopheles arabiensis human biting rates in an area with low vector density and high insecticide-treated bed net use. Am J Trop Med Hyg. 2010;83:838–842. doi: 10.4269/ajtmh.2010.10-0088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Mboera LE, Kihonda J, Braks MA, Knols BG. Influence of centers for disease control light trap position, relative to a human-baited bed net, on catches of Anopheles gambiae and Culex quinquefasciatus in Tanzania. Am J Trop Med Hyg. 1998;59:595–596. doi: 10.4269/ajtmh.1998.59.595. [DOI] [PubMed] [Google Scholar]
- 33.Huho B, Briet O, Seyoum A, Sikaala C, Bayoh N, Gimnig J, et al. Consistently high estimates for the proportion of human exposure to malaria vector populations occurring indoors in rural Africa. Int J Epidemiol. 2013;42:235–247. doi: 10.1093/ije/dys214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Malaria Transmission Consortium (2015). https://mtc.crc.nd.edu. Accessed 1 Jun 2015
- 35.Magbity EB, Magbity EB, Lines JD, Marbiah MT, David K, Peterson E. How reliable are light traps in estimating biting rates of adult Anopheles gambiae s.l. (Diptera: Culicidae) in the presence of treated bed nets? Bull Entomol Res. 2002;92:71–76. doi: 10.1079/BER2002200. [DOI] [PubMed] [Google Scholar]
- 36.Killeen GF, Kihonda J, Lyimo E, Oketch FR, Kotas ME, Mathenge E, et al. Quantifying behavioural interactions between humans and mosquitoes: evaluating the protective efficacy of insecticidal nets against malaria transmission in rural Tanzania. BMC Infect Dis. 2006;6:161. doi: 10.1186/1471-2334-6-161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Okumu FO, Kotas ME, Kihonda J, Mathenge EM, Killeen GF, Moore SJ. Comparative evaluation of methods used for sampling malaria vectors in the Kilombero valley, South eastern Tanzania. Open Trop Med J. 2008;1:51–55. doi: 10.2174/1874315300801010051. [DOI] [Google Scholar]
- 38.Spiegelhalter DJ, Thomas A, Best N, Lunn D (2003) Winbugs Version 1.4. MRC-BSU, Cambridge, England
- 39.Egger M, Davey Smith G, Schneider M, Minder C. Bias in meta-analysis detected by a simple, graphical test. BMJ. 1997;315:629–634. doi: 10.1136/bmj.315.7109.629. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Govella NJ, Moore JD, Killeen GF. An exposure-free tool for monitoring adult malaria mosquito populations. Am J Trop Med Hyg. 2010;83:596–600. doi: 10.4269/ajtmh.2010.09-0682. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Smith T, Charlwood JD, Takken W, Tanner M, Spiegelhalter DJ. Mapping the densities of malaria vectors within a single village. Acta Trop. 1995;59:1–18. doi: 10.1016/0001-706X(94)00082-C. [DOI] [PubMed] [Google Scholar]
- 42.Lindsay SW, Armstrong-Schellenberg JR, Zeiler HA, Daly RJ, Salum FM, Wilkins HA. Exposure of Gambian children to Anopheles gambiae malaria vectors in an irrigated rice production area. Med Vet Entomol. 1995;9:50–58. doi: 10.1111/j.1365-2915.1995.tb00116.x. [DOI] [PubMed] [Google Scholar]
- 43.Ribeiro JM, Seulu F, Abose T, Kidane G, Teklehaimanot A. Temporal and spatial distribution of anopheline mosquitos in an Ethiopian village: implications for malaria control strategies. Bull World Health Organ. 1996;74:299–305. [PMC free article] [PubMed] [Google Scholar]
- 44.Lindsay SW, Adiamah JH, Miller JE, Pleass RJ, Armstrong JR. Variation in attractiveness of human subjects to malaria mosquitoes (Diptera: Culicidae) in The Gambia. J Med Entomol. 1993;30:368–373. doi: 10.1093/jmedent/30.2.368. [DOI] [PubMed] [Google Scholar]
- 45.Mukabana WR, Takken W, Killeen GF, Knols BG. Allomonal effect of breath contributes to differential attractiveness of humans to the African malaria vector Anopheles gambiae. Malar J. 2004;3:1. doi: 10.1186/1475-2875-3-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Knols BG, De Jong R, Takken W. Differential attractiveness of isolated humans to mosquitoes in Tanzania. Trans R Soc Trop Med Hyg. 1995;89:604–606. doi: 10.1016/0035-9203(95)90406-9. [DOI] [PubMed] [Google Scholar]
- 47.Garrett-Jones C (1964) A method for estimating the man-biting rate. World Health Organization, Geneva, WHO/Mal/450
- 48.Elliott R. Studies on man vector contact in some malarious areas in Colombia. Bull World Health Organ. 1968;38:239–253. [PMC free article] [PubMed] [Google Scholar]
- 49.Killeen GF, Tami A, Kihonda J, Okumu FO, Kotas ME, Grundmann H, et al. Cost-sharing strategies combining targeted public subsidies with private-sector delivery achieve high bednet coverage and reduced malaria transmission in Kilombero Valley, southern Tanzania. BMC Infect Dis. 2007;7:121. doi: 10.1186/1471-2334-7-121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Shen J, Botly LC, Chung SA, Gibbs AL, Sabanadzovic S, Shapiro CM. Fatigue and shift work. J Sleep Res. 2006;15:1–5. doi: 10.1111/j.1365-2869.2006.00493.x. [DOI] [PubMed] [Google Scholar]
- 51.Boivin DB, James FO. Light treatment and circadian adaptation to shift work. Ind Health. 2005;43:34–48. doi: 10.2486/indhealth.43.34. [DOI] [PubMed] [Google Scholar]
- 52.Kelly-Hope LA, McKenzie FE. The multiplicity of malaria transmission: a review of entomological inoculation rate measurements and methods across sub-Saharan Africa. Malar J. 2009;8:19. doi: 10.1186/1475-2875-8-19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Bryan JH. Vectors of Wuchereria bancrofti in the Sepik Provinces of Papua New Guinea. Trans R Soc Trop Med Hyg. 1986;80:123–131. doi: 10.1016/0035-9203(86)90210-5. [DOI] [PubMed] [Google Scholar]
- 54.Shiff CJ, Minjas JN, Hall T, Hunt RH, Lyimo S, Davis JR. Malaria infection potential of anopheline mosquitoes sampled by light trapping indoors in coastal Tanzanian villages. Med Vet Entomol. 1995;9:256–262. doi: 10.1111/j.1365-2915.1995.tb00131.x. [DOI] [PubMed] [Google Scholar]
- 55.Maliti D, Ranson H, Magesa S, Kisinza W, Mcha J, Haji K, et al. Islands and stepping-stones: comparative population structure of Anopheles gambiae sensu stricto and Anopheles arabiensis in Tanzania and implications for the spread of insecticide resistance. PLoS One. 2014;9:e110910. doi: 10.1371/journal.pone.0110910. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Briet OJ. A simple method for calculating mosquito mortality rates, correcting for seasonal variations in recruitment. Med Vet Entomol. 2002;16:22–27. doi: 10.1046/j.0269-283x.2002.00335.x. [DOI] [PubMed] [Google Scholar]
- 57.Smith DL, Dushoff J, Snow RW, Hay SI. The entomological inoculation rate and Plasmodium falciparum infection in African children. Nature. 2005;438:492–495. doi: 10.1038/nature04024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Killeen GF. A second chance to tackle African malaria vector mosquitoes that avoid houses and don’t take drugs. Am J Trop Med Hyg. 2013;88:809–816. doi: 10.4269/ajtmh.13-0065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Hay SI, Rogers DJ, Toomer JF, Snow R. Annual Plasmodium falciparum entomological inoculation rates (EIR) across Africa: literature survey, Internet access and review. Trans R Soc Trop Med Hyg. 2000;94:113–127. doi: 10.1016/S0035-9203(00)90246-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Sikaala CH, Chinula D, Chanda J, Hamainza B, Mwenda M, Mukali I, et al. A cost-effective, community-based, mosquito-trapping scheme that captures spatial and temporal heterogeneities of malaria transmission in rural Zambia. Malar J. 2014;13:225. doi: 10.1186/1475-2875-13-225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Amek N, Bayoh N, Hamel M, Lindblade KA, Gimnig JE, Odhiambo F, et al. Spatial and temporal dynamics of malaria transmission in rural Western Kenya. Parasit Vectors. 2012;5:86. doi: 10.1186/1756-3305-5-86. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Kasasa S, Asoala V, Gosoniu L, Anto F, Adjuik M, Tindana C, et al. Spatio-temporal malaria transmission patterns in Navrongo demographic surveillance site, northern Ghana. Malar J. 2013;12:63. doi: 10.1186/1475-2875-12-63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Rumisha SF, Smith T, Abdulla S, Masanja H, Vounatsou P. Modelling heterogeneity in malaria transmission using large sparse spatio-temporal entomological data. Glob Health Action. 2014;7:22682. doi: 10.3402/gha.v7.22682. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Charlwood JD, Smith T, Lyimo E, Kitua A, Masanja H, Booth M, et al. Incidence of Plasmodium falciparum infection in infants in relation to exposure to sporozoite-infected anophelines. Am J Trop Med Hyg. 1998;59:243–251. doi: 10.4269/ajtmh.1998.59.243. [DOI] [PubMed] [Google Scholar]
- 65.McElroy PD, Beier JC, Oster CN, Beadle C, Sherwood JA, Oloo AJ, et al. Predicting outcome in malaria: correlation between rate of exposure to infected mosquitoes and level of Plasmodium falciparum parasitemia. Am J Trop Med Hyg. 1994;51:523–532. [PubMed] [Google Scholar]
- 66.McElroy PD, Beier JC, Oster CN, Onyango FK, Oloo AJ, Lin X, et al. Dose- and time-dependent relations between infective Anopheles inoculation and outcomes of Plasmodium falciparum parasitemia among children in western Kenya. Am J Epidemiol. 1997;145:945–956. doi: 10.1093/oxfordjournals.aje.a009054. [DOI] [PubMed] [Google Scholar]
- 67.Smith T, Maire N, Dietz K, Killeen GF, Vounatsou P, Molineaux L, et al. Relationship between the entomologic inoculation rate and the force of infection for Plasmodium falciparum malaria. Am J Trop Med Hyg. 2006;75(2 Suppl):11–18. doi: 10.4269/ajtmh.2006.75.2_suppl.0750011. [DOI] [PubMed] [Google Scholar]
- 68.Rumisha SF, Smith TA, Masanja H, Abdulla S, Vounatsou P. Relationship between child survival and malaria transmission: an analysis of the malaria transmission intensity and mortality burden across Africa (MTIMBA) project data in Rufiji demographic surveillance system, Tanzania. Malar J. 2014;13:124. doi: 10.1186/1475-2875-13-124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Lengeler C. Insecticide-treated bed nets and curtains for preventing malaria. Cochrane Database Syst Rev. 2004;2:CD000363. doi: 10.1002/14651858.CD000363.pub2. [DOI] [PubMed] [Google Scholar]
- 70.Pluess B, Tanser FC, Lengeler C, Sharp BL. Indoor residual spraying for preventing malaria. Cochrane Database Syst Rev. 2010;4:CD006657. doi: 10.1002/14651858.CD006657.pub2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Ilboudo-Sanogo E, Cuzin-Ouattara N, Diallo DA, Cousens SN, Esposito F, Habluetzel A, et al. Insecticide-treated materials, mosquito adaptation and mass effect: entomological observations after five years of vector control in Burkina Faso. Trans R Soc Trop Med Hyg. 2001;95:353–360. doi: 10.1016/S0035-9203(01)90179-8. [DOI] [PubMed] [Google Scholar]
- 72.Ye Y, Hoshen M, Kyobutungi C, Louis VR, Sauerborn R (2009) Local scale prediction of Plasmodium falciparum malaria transmission in an endemic region using temperature and rainfall. Glob Health Action 2. doi:10.3402/gha.v2i0.1923 [DOI] [PMC free article] [PubMed]
- 73.Kigadye ES, Nkwengulila G, Magesa SM, Abdulla S. Diversity, spatial and temporal abundance of Anopheles gambiae complex in the Rufiji River basin, south-eastern Tanzania. Tanzan J Health Res. 2010;12:68–72. doi: 10.4314/thrb.v12i1.56320. [DOI] [PubMed] [Google Scholar]
- 74.Killeen GF, Smith TA, Ferguson HM, Mshinda H, Abdulla S, Lengeler C, et al. Preventing childhood malaria in Africa by protecting adults from mosquitoes with insecticide-treated nets. PLoS Med. 2007;4:e229. doi: 10.1371/journal.pmed.0040229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Spiegelhalter DJ, Best N, Carlin BP, van der Linde A. Bayesian measures of model complexity and fit (with discussion) J Roy Statist Soc B. 2002;64:583–640. doi: 10.1111/1467-9868.00353. [DOI] [Google Scholar]