

Abstract
Quaternary climatic cycles have influenced marine organisms’ spatial distribution and population dynamics. This study aimed to elucidate the evolutionary influences of contemporary and glacial physical barriers on the population structure, demography and colonization history of the mudskipper (Periophthalmus modestus) based on a mitochondrial gene segment (ND5) from 131 individual fish sampled in the northwestern Pacific Ocean. The current Kuroshio Current and the glacial exposure of the Taiwan Strait appeared to have restricted migration among the South China Sea, coastal East China and Japan. However, genetic homogeneity (Nm>1) also suggested contemporary larval transportation by sea circulation between the East China Sea and the South China Sea or historical dispersal along the glacial exposed shoreline among China, Japan and the Ryukyu Islands. Evolutionary signals of the strengthened East Asian Summer Monsoon in the mid-Pleistocene and regional difference in intertidal primary productions were indicated by a late-Pleistocene population expansion of P. modestus with a higher effective population size in the South China Sea than in the East China Sea. Furthermore, a potential colonization origin from the South China Sea was consistently inferred by different clues, including the populations’ coalescence times, the ancestral haplotype distribution, the number of private haplotypes and species/genetic diversity.
Historical climatic changes are believed to have greatly influenced the coastal environment1 and the evolutionary history of its biota2,3. With periodic sea-level fluctuations, marine organisms have experienced repeated habitat expansions and contractions. These historic events were imprinted in the evolutionary processes, affecting the distribution and dynamics of populations4,5. It has been hypothesized that a rising sea level and range expansion could result in genetic homogeneity and rapid population growth6,7,8,9, whereas a lowering sea level and habitat fragmentation could lead to a heterogeneous population structure4 and a genetic bottleneck for many marine organisms10.
The marginal seas of the northwestern Pacific Ocean (such as Indonesian seas, China seas, and the Sea of Japan/East Sea) have attracted considerable attention in phylogeographical studies due to their complicated topography1,11 and high genetic diversity12,13. China seas (including the South China Sea, the East China Sea, the Yellow Sea and Bohai) provide a good dynamic physical model with which to test the evolutionary responses of marine organisms to the periodic geomorphologic and oceanographic changes during the Quaternary (Figs. 1, 2). The isolated glacial sea basins (viz. semi-enclosed South China Sea and the narrow Okinawa Trough), due to the exposure of the shallow continental shelves14 (Fig. 1a) and sub-habitats divided by the branching Kuroshio Currents (Fig. 1b), likely separated marine species into different populations, so lineage diversification and population differentiation could be expected (Fig. 2a,b). Nevertheless, these isolated populations would be remixed due to absence of the Kuroshio Current in the glacial semi-closed Okinawa Trough (Fig. 1a) or by postglacial coastal sea circulations driven by East Asian Monsoons between the East China Sea and South China Sea (Fig. 1b). Thus, a genetic homogeneous population could also be expected for some marine species in the China seas (Fig. 2c,d).
Figure 1. The sampling sites of P. modestus and oceanographic conditions in the China seas during the glacial53,54 (a) and contemporary39,40 (b) periods.
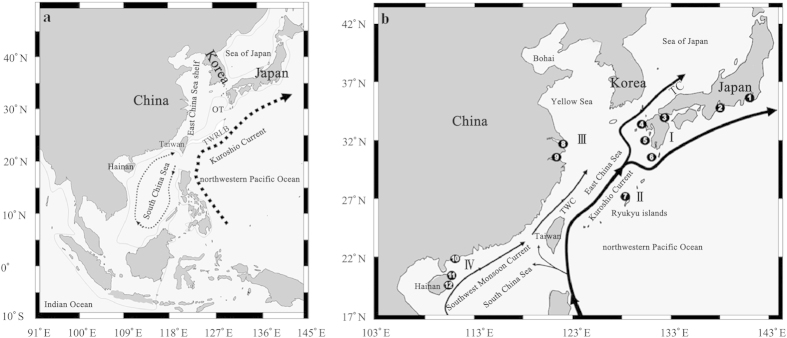
Two glacial sea basins (Okinawa Trough (OT) and South China Sea) were isolated by the exposure of the Taiwan Strait and the Taiwan-Ryukyu land bridge (TWRLB). The glacial shoreline and ocean current are denoted by the light solid line and dotted arrows, respectively. The contemporary surface circulations, including the summer Southwest Monsoon Current, the Taiwan Warm Current (TWC), and the main course of the Kuroshio Current and Tsushima Current (TC), are shown by solid arrows. The names of the numbered sampling localities and four geographic groups (I, II, III and IV) corresponding to different regions are listed in Table 1. The map was created in the Generic Mapping Tools (GMT v5.1.2; http://gmt.soest.hawaii.edu/) software package.
Figure 2. Evolutionary patterns of marine organisms in the China seas: Regional vicariance by glacial sea basins (a) or postglacial sea currents (b) and population connectivity through glacial dispersal (c) or present-time gene flow driven by sea currents (d).
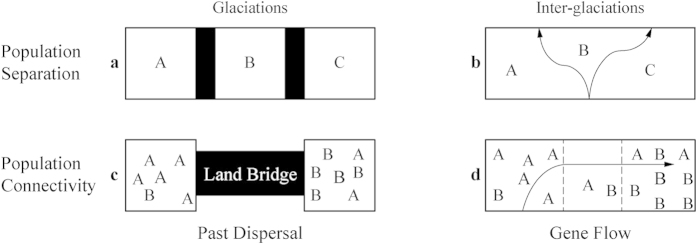
Black boxes indicate an exposed shallow shelf, and white boxes indicate regional populations (A–C). The arrows mark sea currents.
Several previous studies on different species showed the two above-mentioned contrasting evolutionary patterns in the China seas15. Deep lineage differentiation or heterogeneous populations (Fig. 2a,b) were observed in turban shell (Turbo cornutus)16, Pandaka gobies17, tideland snail (Cerithidea cingulata)18, mudskipper (Boleophthalmus pectinirostris)19, mullet (Chelon haematocheilus)20, acorn barnacles (Tetraclita squamosa and T. japonica)21,22, and mitten crabs (Eriocheir sensu stricto)23,24. Yet another pattern of population expansion with genetic homogeneity (Fig. 2c,d) was also found in tideland snail (Batillaria zonalis)25, spotted sea bass (Lateolabrax maculatus)26, Japanese anchovy (Engraulis japonicus)27, demersal fish Nibea albiflora28, neon damselfish (Pomacentrus coelestis)29, swimming crab Portunus trituberculatus30 and mud crab Scylla paramamosain31. The two contrasting phylogeographic patterns among these species could be attributed to their different evolutionary histories or dispersal capabilities. Therefore, to reveal the general evolutionary consequence of eustatic oscillations in the China seas, studies on phylogeographic histories of different species are highly desirable.
As other gobies are13,17,32,33, mudskipper Periophthalmus modestus (Cantor, 1842; Gobioidei: Gobiidae) is a good species for studying the evolutionary effects of sea-level fluctuations and physical barriers in the coastal northwestern Pacific Ocean because of its amphibious life history, which depends on the intertidal mudflat habitat34,35 and limited adult dispersal capability. This species is endemic in this region, including the coasts of the China seas, the Korean peninsula and Japan36. Mukai and Sugimoto37 found two divergent lineages and a significant genetic differentiation of P. modestus between the main islands of Japan (Honshu and Kyushu) and the Ryukyu Islands (Tanegashima and Okinawajima), but they could not elucidate the overall phylogeographic history of this species because of lack of samples from the Asian continental coast. In the present study, sequences of a mitochondrial gene from P. modestus along the coast of China were combined with those from the previous study to address three evolutionary questions: first, the evolutionary influence of barriers (e.g., the glacial exposure of the Taiwan Strait and contemporary branches of the Kuroshio Current) on population genetic structure; second, the demographic response to glacial cycles; and third, the colonization origin for this species in the China seas. This study will shed light on the understanding of relationships among Pleistocene climatic cycles, oceanographic conditions and the evolution of marine organisms.
Results
Phylogenetic analyses
A dataset with 76 haplotypes (772 bp) was obtained through sequence alignment. No indels or stop codons were found. In contrast to Mukai’s and Sugimoto’s result37, no different lineages or subspecies were identified in P. modestus through phylogenetic analyses (Fig. 3). A single lineage and a consistent branching pattern with a South China Sea haplotype (N66) basal to all other haplotypes was revealed in the neighbour-joining (NJ), maximum likelihood (ML) and Bayesian inference (BI) trees, although nodal supports for grouping the haplotypes other than N66 were not strong (BP = 62/58 for NJ/ML and posterior probability (PP) = 0.54 for BI). Likelihood mapping analysis corroborated the internal branch of the ingroup, excluding N66 with a strong support value (99.7%, Fig. S1), suggesting N66's basal position. Furthermore, N59 and N60 were rejected as potential ancestral haplotypes by the Shimodaira-Hasegawa (SH) test (∆lnL = 1.054, P < 0.001). Median-joining network (MJN) analysis yielded a star-like network without an obvious geographic structure. Some newly derived terminal haplotypes were generally endemic or private, and interior haplotypes were widely distributed and shared across different regions (Fig. 3, Table S1).
Figure 3. Phylogenetic trees (NJ, ML and BI) and network (MJN) of haplotypes. Sampled haplotypes are indicated by names (N01–N76), and the hollow ovals represent unsampled haplotypes (mv1–9) in MJN.

The numbers on the branches show the bootstrap values over 50% (NJ/ML) or posterior probability values over 0.5 (BI) and the number of substitutions over 2 in MJN. The 12 sampling localities are distinguished by different colours (grey for 1–7, black for 8–12) and styles, and the size of the circles is proportional to the frequency of haplotypes in MJN. The basal haplotype N66 is indicated using a star. The four clusters a, b, c, d were shown.
Genetic diversity and population differentiation
Among the four geographic groups defined (see Methods), decreasing trends in genetic diversity and the proportion of private haplotypes were revealed from the South China Sea group (IV: H/Π, 0.991/3.19%; Np/N, 0.78) through the coastal East China group (Ш: H/Π, 0.978/2.61%; Np/N, 0.54) to the coastal Japan group (І: H/Π, 0.819/2.22%; Np/N, 0.31) (Tables 1, S1). Significant genetic differentiation (Bonferroni correction, P < 0.017) based on the haplotype frequency and the TrN + G distance was detected between the coastal Japan group (I) and China groups (III and IV; Table 2). Furthermore, a high endemism index within these three groups (0.77–0.88) also indicated their historical isolations and limited migration. However, high gene flow (Nm>1) was observed in all pairwise comparisons among three groups (I, III and IV).
Table 1. Sampling localities, sample size, number of haplotypes and genetic diversity of P. modestus.
| Population ID | Locality | Coordinates | N/Nh/Np | Proportion of private haplotypes (Np/N) | Endemism indices (Np/Nh) | H/Π (%) |
|---|---|---|---|---|---|---|
| I (1~6) | Coastal Japan group | − | 42/16/13 | 0.31 | 0.81 | 0.819/ 2.22 |
| 1 | Edogawa, Japan | 35˚42′05′′N 139˚55′18′′E | 7/4/2 | 0.29 | 0.50 | 0.714/ 1.88 |
| 2 | Ibigawa, Japan | 35˚05′07′′N 136˚41′01′′E | 12/6/4 | 0.33 | 0.67 | 0.682/ 0.15 |
| 3 | Yamagunigawa, Japan | 33˚36′29′′N 131˚11′02′′E | 6/3/1 | 0.17 | 0.33 | 0.733/ 0.81 |
| 4 | Imarigawa, Japan | 33˚17′16′′N 129˚50′01′′E | 4/2/0 | 0 | 0 | 0.500/ 2.79 |
| 5 | Rokkakugawa, Japan | 33˚11′23′′N 130˚12′03′′E | 9/7/4 | 0.44 | 0.57 | 0.917/ 3.55 |
| 6 | Tanegashima Island, Ryukyu Islands | 30˚26′29′′N 130˚57′10′′E | 4/2/1 | 0.25 | 0.50 | 0.500/ 0.08 |
| II (7) | Okinawajima Island, Ryukyu Islands | 26˚39′02′′N 127˚58′33′′E | 6/2/1 | 0.17 | 0.50 | 0.600/ 0.09 |
| III (8, 9) | Coastal East China group | − | 37/26/20 | 0.54 | 0.77 | 0.978/ 2.61 |
| 8 | Chongming Island, Shanghai, China | 31˚31′30′′N 121˚57′30′′E | 32/23/18 | 0.56 | 0.78 | 0.976/ 2.51 |
| 9 | Nanhui, Shanghai, China | 30˚51′36′′N 121˚54′36′′E | 5/4/2 | 0.40 | 0.50 | 0.900/ 2.98 |
| IV (10 ~ 12) | South China Sea group | − | 46/41/36 | 0.78 | 0.88 | 0.991/ 3.19 |
| 10 | Hailing Island, Yangjiang, Guangdong, China | 21˚39′0′′N 111˚57′36′′E | 33/31/27 | 0.82 | 0.87 | 0.996/ 3.46 |
| 11 | Longlou, Wenchang, Hainan, China | 19˚40′48′′N 111˚0′0′′E | 6/6/4 | 0.67 | 0.67 | 1.0/ 0.68 |
| 12 | Dongjiao, Wenchang, Hainan, China | 19˚33′36′′N 110˚49′48′′E | 7/7/4 | 0.57 | 0.57 | 1.0/ 3.54 |
N, sampling size; Nh, number of haplotypes; Np, number of private haplotypes; H, haplotype diversity; Π, nucleotide diversity.
See Mukai and Sugimoto37 for localities 1–7; localities 8–12 are from this study.
Table 2. Pairwise distance (below diagonal) and Nm (above diagonal) between three geographic groups based on haplotype frequency (FST) and Tamura-Nei distance (ΦST). See Table 1 for population ID.
| ID | I | III | IV | I | III | IV |
|---|---|---|---|---|---|---|
| Frequency | TrN + G | |||||
| I | 0 | 6.050 | 5.352 | 0 | 3.933 | 2.858 |
| III | 0.076* | 0 | 98.589 | 0.113* | 0 | 11.074 |
| IV | 0.085* | 0.005 | 0 | 0.149* | 0.043 | 0 |
*Level of significance, P ≤ 0.017 for Bonferroni correction.
Demographic analyses
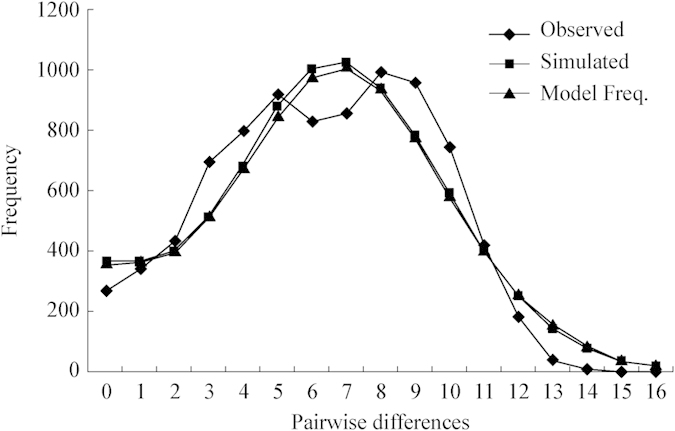
A unimodal curve was observed in mismatch distribution analyses for all sequences of P. modestus (Fig. 4), and it was not significantly different from the expected sudden expansion model (SSD = 0.003, P = 0.672). The population expansion of P. modestus was also supported by two neutrality tests (Tajima’s D = −1.923, P = 0.005 and Fu’s Fs = −24.992, P = 0). Based on expansion parameter τ (τ = 7.438), the expansion time was inferred to be 248 thousand years ago (kya).
Figure 4. Mismatch distribution analysis for the whole population of P. modestus.
Bayesian Skyline Plot (BSP) presented a more detailed demographic history (Fig. 5b). The time to the recent common ancestor (TMRCA) of P. modestus was estimated to be 365 (539–210) kya, consistent with the inferred population expansion time (248 kya). A slow population growth during 365–120 kya and a slight bottleneck during 120–70 kya followed by rapid expansion were revealed after 70 kya. The plot of the East China Sea group showed a relative constant population size for a long time and a recent rapid expansion after 60 kya (Fig. 5b). The BSP of the South China Sea group revealed a slow population growth in 350–140 kya and a slight bottleneck in 140–100 kya followed by rapid expansion after 100 kya (Fig. 5b). Through comparison, the South China Sea group showed an older TMRCA and earlier expansion than the East China Sea group. Furthermore, the effective population size in the South China Sea was higher than that in the East China Sea since 300 kya.
Figure 5. History of the East Asian Summer Monsoon indicated by magnetic susceptibility (MS).
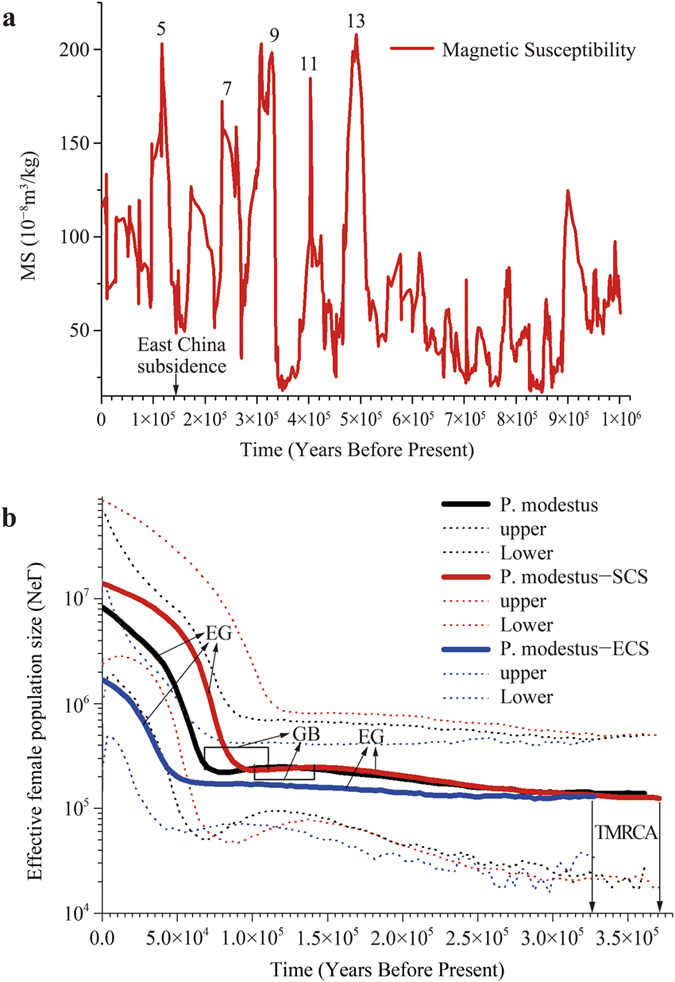
60 (a) and the demographic history of P. modestus, the East China Sea population and the South China Sea population (b). Thick lines show the medians of population sizes, and thin dotted lines indicate upper and lower 95% credibility intervals. Expansion growth (EG) and genetic bottleneck (GB) are indicated. The last five interglaciations (isotopic stages 5, 7, 9, 11 and 13) are labelled in (a).
Discussion
The genetic signal of glacial isolation was suggested by high endemism indexes within the coastal East China (III, 0.77) and the South China Sea (IV, 0.88) groups (Table 1). These glacial isolated sea basins would have caused private haplotypes, heterogeneous populations or divergent lineages between the East China Sea (Okinawa Trough) and the South China Sea20,23,24 due to the exposure of the Taiwan Strait (Figs. 1a, 2a). However, the physical separation was likely disturbed by a fluctuating sea level. The postglacial rising sea level and the re-flooded Taiwan Strait were likely responsible for the contemporary high gene flow of P. modestus (Table 2, Nm>11) between these once fragmented habitats (Fig. 1b). Moreover, a seasonal change in monsoon circulations38 could drive planktonic larvae of marine organisms to migrate between the East China Sea and South China Sea31 (Fig. 2d).
The significant genetic differentiation and high endemism indexes revealed between/within the geographic groups of coastal Japan (I) and China (III and IV; Tables 1, 2) indicate a negative influence of the contemporary Kuroshio Current on the connectivity of marine organisms in the East China Sea (Fig. 2b). The postglacial Kuroshio Current branches into the East China Sea, the Sea of Japan and the northwestern Pacific regions39 (Fig. 1b) and thus divides the sea area into heterogeneous sub-habitats with different temperatures and salinities40. The Kuroshio Current was shown to act as a dispersal barrier to promote lineage diversification or population differentiation in some marine organisms16,21,22,41,42,43,44,45,46 (Fig. 2b). In this study, the branches of the Kuroshio Current also seem to have influenced the population structure of P. modestus.
The high gene flow (Nm>1) observed between the coasts of China and Japan (Table 2) likely indicates a past dispersal instead of ongoing migration in the China seas (Fig. 2c). The isolation time (<10 kyr) of P. modestus linked to the postglacial Kuroshio Current seems to be insufficient in accumulating a deep genetic divergence between mainland China and the main islands of Japan. Similar historical population dispersals at times of a lower sea level across a long distance were observed in some West Pacific marine taxa47,48,49. Due to the inability to discriminate among contemporary gene flow and historical events in most traditional population structure analyses50, the inferred genetic connectivity of P. modestus between coastal China and the main islands of Japan might be attributed to a historical coastline connection and a long-distance dispersal instead of contemporary gene flow51 (Fig. 2c). The endemic distribution of terminal haplotypes and the sharing of interior haplotypes (Fig. 3, MJN) further indicate historical range expansion50 across coastal China, Japan and the Ryukyu Islands. During the Last Glaciation Maximum, the sea level dropped ca. 130–150 m in the East China Sea52, and a land bridge connecting Taiwan and the Ryukyu Islands blocked the entry of Kuroshio Current into the East China Sea53,54 (Fig. 1a). Migration and population admixture became possible when the East China Sea was reduced in size to the elongated Okinawa Trough with a continuous coastline between mainland China and Japan (Figs. 1a, 2c). As an amphibious fish, the larvae of P. modestus develop in open water with a planktonic stage of approximately 50 days55,56. The glacial eastward Kuroshio Current54 (Fig. 1a) might also have contributed to its range expansion through larval transport from coastal China to Japan.
The signal of the demographic expansion of P. modestus was detected through a mismatch distribution analysis (Fig. 4), the two neutrality tests, and BSP (Fig. 5b). The results indicate a rapid population expansion in the whole population of P. modestus since ca. 70 kya. Given the uncertainty of the molecular clock, a late Pleistocene expansion since the last interglacial sea-level highstand (<133 kya) can also be inferred for P. modestus using a slower molecular rate (e.g., the conventional 1% per million years (/myr) of teleostean mitochondrial Cyt b rate57). The shoreline enlargement due to East China subsidence occurred in the late Pleistocene11,58. Furthermore, the strengthening of the East Asian Summer Monsoon in the mid-Pleistocene59,60 (Fig. 5a) caused high precipitation during the subsequent inter-glaciations and interstades61,62. The increased rainfall and runoff generally parallelized higher nutrient input into the intertidal habitat63, which could have been responsible for late Pleistocene population growth of coastal organisms.
The South China Sea population of P. modestus showed a larger historical effective population size and earlier growth relative to the East China Sea population (Fig. 5b). The difference in regional population dynamics is closely related to difference in the primary production between the northern South China Sea and the East China Sea64. A decreasing trend in glacial and interglacial mean terrestrial net primary production was observed from coastal South China to East China65,66,67,68,69,70,71,72. Furthermore, heavier precipitation in low latitude relative to high latitude73 can transport much more terrestrial organic matter to intertidal mudflats and coastal regions74,75. Then, the terrigenous nutrients can be integrated into the food web by benthic microalgae76,77. Although no obvious global trend in the spatial distribution of intertidal microphytobenthic biomass was revealed at this point, the higher effective population size of mudskipper in the northern South China Sea relative to the East China Sea was likely influenced by regional difference in the intertidal nutrient and primary production78. Furthermore, for mudflat-dependent mudskipper, the later population expansion in the East China Sea was likely attributed to the late Pleistocene development of the muddy shoreline derived from the Changjiang Delta79,80,81 due to high microalgae biomass in muddy sediments82,83. A similar higher population size and earlier expansion were also observed in mitten crabs of Eriocheir hepuensis from the coastal northern South China Sea relative to E. sinensis from the East China Sea23.
Two glacial sea basins (South China Sea and Okinawa Trough) could have served as refugia for marine species in the China seas (Fig. 1a). The question is which one acted as the colonization origin of the species. The present study supports the South China Sea as the origin, based on following clues: First, an older coalescence time (TMRCA) is inferred for the South China Sea population relative to the East China Sea population (Fig. 5b). TMRCA indicates the divergence time within a lineage or population, and the South China Sea population is thus believed to have experienced a longer evolutionary history than the East China Sea population. The difference in the evolutionary time is also consistent with the history of the sea basins: The South China Sea was opened in the Oligocene-middle Miocene (ca. 30–15 million years ago, mya)84, whereas the Okinawa Trough formed only in late Miocene and Pliocene (ca. 10–3 mya) as shallow freshwater and brackish water lakes85. The reopening and large-scale depression of the Okinawa Trough occurred after the early (<1 mya) and middle Pleistocene (<0.7 mya), respectively86,87. Although there is uncertainty regarding the evolutionary rate of the mitochondrial ND5 gene, the relative coalescence times of the two geographic populations are apparent. Second, a putative ancestral haplotype (N66) is found only in site 10 from the South China Sea (Figs. 1b, 3 and Table S1). Several other haplotypes (e.g., N65, N67–76) were also inferred as old, close to basal haplotype N66 (Fig. 3). The South China Sea population thus possesses more older haplotypes than the East China Sea population (8 versus 5) even if the sampling size is lower in the former (46 versus 85; Table 1). Third, a descending proportion of private haplotypes from the South China Sea group (IV, 0.78) through the coastal East China group (III, 0.54) to the coastal Japan group (I, 0.31) indicates this species’ colonization origin in the South China Sea because a lower proportion of private haplotypes is expected in the recolonized region88. Fourth, genetic diversity is the highest in the South China Sea group (H/Π, 0.991/3.19%) compared to values in the other groups from the East China Sea (e.g., coastal East China group, 0.978/2.61%; coastal Japan group, 0.819/2.22%; and Okinawa group, 0.600/0.09%; Table 1). Similarly, a decreasing trend in genetic diversity from coastal China to Japanese sites is also observed in an estuarine fish (Salanx ariakensis)51. The colonization origin or glacial refuge is expected to possess higher genetic diversity because of its longer evolutionary time, and the newly colonized regions generally exhibit lower genetic diversity due to the founder effect5. Fifth, higher species diversity of the genus Periophthalmus occurs in the South China Sea than in the East China Sea (10 versus 3)36. Similarly, a higher species diversity of congeners in the South China Sea is observed from the hairtails Trichiurus89, mud crabs Scylla90 and fiddler crabs Uca91. The East Indies Triangle, including the South China Sea, is generally believed to operate as a centre of origin due to the higher species diversity in this area relative to neighbouring regions of the Indo-West Pacific12. P. modestus is distributed only in the marginal seas of the northwestern Pacific, including the China seas, the Sea of Japan and the eastern Japanese coast36. Therefore, through postulating that the South China Sea is the earlier refuge or colonization origin for P. modestus, minimum inter-sea basins migration are inferred92. P. modestus might have dispersed northward into the East China Sea and other marginal seas following the interglacial rising sea level. Subsequently, the northern population would have retreated back into two potential refugia (the Okinawa Trough and the South China Sea) during the period of glaciation. The repeated range expansions and contractions thus caused a gradually declining diversity distribution from the South China Sea through the East China Sea to the coast of Japan.
In summary, this study indicated the negative influence of the postglacial Kuroshio Current and the glacial exposure of the Taiwan Strait on the population structure of P. modestus. Although a similar genetic homogeneity was observed among the South China Sea group, the coastal East China Group and the coastal Japan group, contemporary gene flow through the Taiwan Strait and historical dispersal across the Okinawa Trough could be responsible for their population admixture. The demographic history is likely correlated with the mid-Pleistocene strengthened East Asian Summer Monsoon and the difference in primary production between the coastal South China Sea and the East China Sea. As a species distributed in the marginal seas of the northwestern Pacific Ocean, P. modestus is inferred to have colonized northward from the South China Sea through the East China Sea to the coasts of Korea and Japan.
Methods
Sampling and data collection
A total of 131 individual P. modestus fish were evaluated in this study, including those reported by Mukai and Sugimoto37. Eighty-three individuals were newly collected from five coastal sites in the East China Sea (sites 8, 9) and the South China Sea (sites 10–12; Fig. 1b) and were preserved in 95% ethanol for molecular analysis. Total genomic DNA was extracted from each specimen using a standard phenol-chloroform extraction method93. A segment (approximately 970 bp) of the mitochondrial gene NADH dehydrogenase 5 subunit (ND5) was amplified using the primer pair, L12321-Leu and H13396–ND5M94. Initial denaturation was 4 min at 95 °C, followed by 35 cycles of 1 min at 95 °C, 1 min at 55 °C, 2 min at 72 °C, and a final extension of 4 min at 72 °C. PCR products were separated on 1.5% agarose gel and purified with a Gel Extraction Mini Kit (Watson BioTechnologies, Shanghai, China). Purified products were sequenced with the primer H13396 on an ABI Prism 3730 automatic sequencer (Applied Biosystems, Thermo Fisher Scientific Corporation, USA). These sequences were deposited in GenBank with accession numbers HQ453212-HQ453269. ND5 sequences of P. modestus and an outgroup species P. argentilineatus (GenBank/EMBL/DDBJ accession numbers: AB257605–AB257627) collected from the main islands of Japan and the Ryukyu Islands37 were included in the analysis (Table 1). All sequences were aligned using ClustalX95 with default parameters. The evolutionary models for the datasets including and excluding outgroups were determined using Modeltest96. Two TrN+G models97 with different parameters were selected under the Akaike Information Criterion (AIC) for all sequences (gamma = 0.1103; base frequencies A = 0.2845, C = 0.2723, G = 0.1374, T = 0.3057; rate matrix, R[A–C] = R[A–T] = R[C–G] = R[G–T] = 1, R[C–T] = 7.8281, R[A–G] = 12.9280) and hierarchical likelihood ratio tests (hLRTs) for ingroup sequences (gamma = 0.0163; base frequencies A = 0.2831, C = 0.2638, G = 0.1428, T = 0.3104; rate matrix, R[A–C] = R[A–T] = R[C–G] = R[G–T] = 1, R[C–T] = 7.1746, R[A–G] = 13.9593), respectively. A parameters-complicated TIM+I model selection under the AIC for ingroup sequences was not used for subsequent population analyses because of model limitations in those programs.
Phylogenetic analyses
Four tree-construction methods, NJ98, ML99, BI100 and MJN101, were used to recover the intraspecific evolutionary relationship using PAUP (ver. 4.0b10)102, MrBayes (ver. 3.2.1)103 and NETWORK (ver. 4.613; fluxus-engineering.com), respectively. For NJ analysis, maximum likelihood distances were used. The ML analysis was conducted using a heuristic search with the random addition of sequences (nreps = 10). The nodal supports were assessed using non-parameter bootstrap sampling with 10,000 and 1,000 pseudoreplicates for NJ and ML analysis, respectively. BI was performed with a six-parameter model (GTR+G) similar to TrN+G. These parameters were estimated in the program using the following settings: ngen = 7,000,000; samplefreq = 1000; burnin = 1,750; Nchains = 4; and Nruns = 2. The convergence of independent runs was achieved when white noise was seen in the overlay plot of generation versus the log probability for both runs with the potential scale reduction factor (PSRF) approaching 1 and a low standard deviation of split frequencies (0.005838 < 0.01) after 7,000,000 generations.
Due to uncertainty in the phylogenetic trees, the branching order of the ingroup was assessed based on the inferred phylogenetic tree and network (Fig. 3): (i) The support for the internal branches of four clusters, including a (outgroup), b (N66), c (N65, N67, N68, N69, N70, N71, N72, N73, N74, N75, N76), and d (the rest haplotypes), was estimated using likelihood mapping in TREE-PUZZLE-5.2104,105. (ii) An alternative scenario to enforce the monophyly of the outgroup and two haplotypes, N59 and N60, with multiple substitutions to others was compared with the estimated ML tree using the SH test106 in PAUP and Seq-Gen v1.3.3107 with partial optimization and 1000 simulated datasets.
Population structure analyses
To avoid artificial statistical bias due to a low sample size from some localities, some neighbouring sites were combined into four geographic groups, including the coastal Japan group (I, sites 1–6), the Okinawa Island group (II, site 7), the coastal East China group (III, sites 8 and 9), and the South China Sea group (IV, sites 10–12), in the following population structure analyses, according to some historical (e.g., glacial exposure of the Taiwan Strait) and/or present (e.g., branches of the Kuroshio Current) barriers to gene flow (Fig. 1). The proportion of private haplotypes, the endemism index, the haplotype diversity (H) and the nucleotide diversity (Π) were estimated for each locality and geographic group using ARLEQUIN version 3.5108. Excluding insufficient sampling group II, the pairwise genetic divergence (FST and ΦST) and gene flow (Nm) among three geographic groups (I, Ш and IV) were assessed based on the haplotype frequency and the ingroup’s TrN + G model in ARLEQUIN, respectively. The significance of the F statistics for the geographic group comparisons was evaluated using 10,000 permutations, and the Bonferroni correction for multiple testings109 was applied with a lower threshold for the nominal significance level (k = 3, P1 = 0.05/3, and P ≤ 0.017).
Demographic history
The demographic history of P. modestus was inferred through a mismatch distribution analysis110 and two neutral tests, Tajima’s D111,112 and Fu’s Fs113, using Arlequin. Both neutrality tests are sensitive to population growth in the absence of selection, and significant negative values generally suggest population expansion111,112,113. The significance of the neutrality tests was assessed in Arlequin by 10,000 permutations. For mismatch analysis, a multimodal distribution is expected for populations in demographic equilibrium, whereas a unimodal distribution usually indicates a recent demographic expansion110,114. The validity of the estimated stepwise expansion model was tested using the sum of square deviations (SSD) between the observed and expected mismatch as a statistic to infer the significance with the parameter bootstrap approach (10,000 replicates). The expansion time (t) was estimated through the equation t = τ/2 μm, where τ is the mutational timescale, m is the segment length (m = 772 for the present data), and 2 μ is the pairwise mutational rate of the fragment under study. There is no general mitochondrial DNA evolutionary rate for teleosts; an approximate pairwise molecular clock (2 μ) of 3.8%/myr for the ND5 gene from related gobies (Rhinogobius species)32 was thus used in this study.
A more accurate coalescent model, BSP, implemented in BEAST v1.8.2115 and visualized in TRACER v1.6116, was also used to estimate the divergence time (TMRCA)117 and effective population size changes over time for all sequences of P. modestus. Furthermore, the ingroup sequences of P. modestus were divided into two geographic groups corresponding to two identified marine eco-regions118, the East China Sea (sites 1–9) and the South China Sea (sites 10–12). Subsequently, the population dynamics of two geographic groups from the East China Sea and the South China Sea were further compared using BSP. These analyses were run using the following parameters: 6 × 107 generations, a burn-in of 6 × 106 generations, sampling per 10,000, and 8 groups for the East China Sea; 9 × 107 generations, a burn-in of 9 × 106 generations, sampling per 10,000, and 15 groups for the South China Sea; and 6 × 107 generations, a burn-in of 6 × 106 generations, sampling per 10,000, and 20 groups for all sequences. The effective sample sizes of all runs were over 200. An evolutionary rate (u) of 1.9%/myr32 was used to plot population size with respect to time.
Additional Information
Accession Codes: DNA sequence accession numbers: HQ453212-HQ453269 in GenBank.
How to cite this article: He, L. et al. Biogeographical role of the Kuroshio Current in the amphibious mudskipper Periophthalmus modestus indicated by mitochondrial DNA data. Sci. Rep. 5, 15645; doi: 10.1038/srep15645 (2015).
Supplementary Material
Acknowledgments
We sincerely thank Xiaoyan Wu, Jianzhong GE, Jianyi Liu and Xuesen Cui for their kind encouragement and support during the study and manuscript preparation. Several colleagues, including Jinzhou Du, Bing Deng, Guosen Zhang, Zhuoyi Zhu, Ruifeng Zhang, Heng Zhang, Chengqi Fan and Hequan Gu, provided help in field sampling. This research was supported by the National Natural Science Foundation of China (30800117), the Key Project of Chinese National Programs for Fundamental Research and Development (973 Program; 2011CB409801), 111 Project (B08022), Ministry of Education, China, the Postdoctoral Science Foundation of China (2013M540347 and 2014T70406), project No. 2007M04 from a special research fund for national non-profit institutes (East China Sea Fisheries Research Institute), and the State Key Laboratory of Estuarine and Coastal Research funding, East China Normal University.
Footnotes
Author Contributions L.J.H. designed the project, collected samples, performed the molecular experiment, conducted data analyses, and wrote the manuscript. K.H.C. analysed the data and revised the manuscript. T.M. and J.Z. contributed reagents/materials/analysis tools, assisted in sample collection and revised the manuscript. Q.M. was involved in the fieldwork. All authors read and approved the final manuscript.
References
- Wang P. X. & Sun X. J. Last glacial maximum in China: comparison between land and Sea. Catena 23, 341–353 (1994).
- Avise J. C. Phylogeography: The History and Formation of Species. Harvard University Press, Cambridge, Massachusetts (2000).
- Hewitt G. M. The genetic legacy of the Quaternary ice ages. Nature 405, 907–913 (2000). [DOI] [PubMed] [Google Scholar]
- Hewitt G. M. Some genetic consequences of ice ages, and their role in divergence and speciation. Biol. J. Linn. Soc. 58, 247–276 (1996). [Google Scholar]
- Hewitt G. M. Post-glacial re-colonization of European biota. Biol. J. Linn. Soc. 68, 87–112 (1999). [Google Scholar]
- Barry J. P., Baxter C. H., Sagarin R. D. & Gilman S. E. Climate-related, long-term faunal changes in a California rocky intertidal community. Science 267, 627–675 (1995). [DOI] [PubMed] [Google Scholar]
- Rex M. A. An oblique slant on deep-sea biodiversity. Nature 385, 577–578 (1997). [Google Scholar]
- Kitamura A., Omote H. & Oda M. Molluscan response to early Pleistocene rapid warming in the Sea of Japan. Geology 28, 723–726 (2000). [Google Scholar]
- Ruban D. A. Jurassic transgressions and regressions in the Caucasus (northern Neotethys Ocean) and their influences on the marine biodiversity. Palaeogeogr. Palaeocl. 251, 422–436 (2007). [Google Scholar]
- Brett C. E., Hendy A. J. W., Bartholomew A. J., Bonelli J. J. R. & Mclaughlin P. I. Response of shallow marine biotas to sea-level fluctuations: A review of faunal replacement and the process of habitat tracking. Palaios 22, 228–244 (2007). [Google Scholar]
- Wang P. X. Cenozoic deformation and the history of sea-land interactions in Asia in Continent-Ocean Interactions in the East Asian Marginal Seas (eds. Clift P., Wang P., Kuhnt W. & Hayes D.) 1–22 (Geophysical Monograph, AGU, 149, 2004). [Google Scholar]
- Briggs J. C. The marine East Indies: diversity and speciation. J. Biogeogr. 32, 1517–1522 (2005). [Google Scholar]
- Polgar G. et al. Phylogeography and demographic history of two widespread Indo-Pacific mudskippers (Gobiidae: Periophthalmus). Mol. Phylogenet. Evol. 73, 161–176 (2014). [DOI] [PubMed] [Google Scholar]
- Wang P. X. The Ice age China Sea: Status and problems. Quaternary Sci. 10, 111–124 (1990). [Google Scholar]
- Ni G., Li Q., Kong L. F. & Yu H. Comparative phylogeography in marginal seas of the northwestern Pacific. Mol. Ecol. 23, 534–548 (2014). [DOI] [PubMed] [Google Scholar]
- Kojima S., Segawa R. & Hayashi I. Genetic differentiation among populations of Japanese turban shell Turbo (Batillus) cornutus corresponding to warm currents. Mar. Ecol.-Prog. Ser. 15, 149–155 (1997). [Google Scholar]
- Mukai T., Suzuki T. & Nishida M. Genetic and geographical differentiation of Pandaka gobies in Japan. Ichthyol. Res. 51, 222–227 (2004). [Google Scholar]
- Kojima S. et al. Molecular phylogeny and population structure of tideland snails in the genus Cerithidea around Japan. Mar. Biol. 149, 525–535 (2006). [Google Scholar]
- Kanemori Y., Takegaki T. & Natsukari Y. Genetic population structure of Boleophthalmus pectinirostris inferred from mitochondrial DNA sequences. Jap. J. Ichthyol. 53, 133–141 (2006). [Google Scholar]
- Liu J. X., Gao T. X., Wu S. F. & Zhang Y. P. Pleistocene isolation in the Northwestern Pacific marginal seas and limited dispersal in a marine fish, Chelon haematocheilus (Temminck & Schlegel, 1845). Mol. Ecol. 16, 275–288 (2007). [DOI] [PubMed] [Google Scholar]
- Chan B. K. K., Tsang L. M. & Chu K. H. Morphological and genetic differentiation of the acorn barnacle Tetraclita squamosa (Crustacea: Cirripedia) in East Asia and description of a new species of Tetraclita. Zool. Scr. 36, 79–91 (2007). [Google Scholar]
- Tsang L. M., Chan B. K. K., Ma K. Y. & Chu K. H. Genetic differentiation, hybridization and adaptive divergence in two subspecies of the acorn barnacle Tetraclita japonica in the northwestern Pacific. Mol. Ecol. 17, 4157–4163 (2008). [DOI] [PubMed] [Google Scholar]
- Xu J. W., Chan T. Y., Tsang L. M. & Chu K. H. Phylogeography of the mitten crab Eriocheir sensu stricto in East Asia: Pleistocene isolation, population expansion and secondary contact. Mol. Phylogenet. Evol. 52, 45–56 (2009). [DOI] [PubMed] [Google Scholar]
- Xu J. W. & Chu K. H. Genome scan of the mitten crab Eriocheir sensu stricto in East Asia: Population differentiation, hybridization and adaptive speciation. Mol. Phylogenet. Evol. 64, 118–129 (2012). [DOI] [PubMed] [Google Scholar]
- Kojima S. et al. Phylogeography of the endangered tideland snail Batillaria zonalis in the Japanese and Ryukyu Islands. Ecol. Res. 20, 686–694 (2005). [Google Scholar]
- Liu J. X., Gao T. X., Yokogawa K. & Zhang Y. P. Differential population structuring and demographic history of two closely related fish species, Japanese sea bass (Lateolabrax japonicus) and spotted sea bass (Lateolabrax maculatus) in northwestern Pacific. Mol. Phylogenet. Evol. 39, 799–811 (2006). [DOI] [PubMed] [Google Scholar]
- Liu J. X. et al. Late Pleistocene divergence and subsequent population expansion of two closely related fish species, Japanese anchovy (Engraulis japonicus) and Australian anchovy (Engraulis australis). Mol. Phylogenet. Evol. 40, 712–723 (2006). [DOI] [PubMed] [Google Scholar]
- Han Z. Q., Gao T. X., Yanagimoto T. & Sakurai Y. Genetic population structure of Nibea albiflora in Yellow Sea and East China Sea. Fisheries Sci. 74, 544–552 (2008). [Google Scholar]
- Liu S. Y. V., Kokita T. & Dai C. F. Population genetic structure of the neon damselfish (Pomacentrus coelestis) in the northwestern Pacific Ocean. Mar. Biol. 154, 745–753 (2008). [Google Scholar]
- Liu Y. et al. Genetic differentiation between populations of swimming crab Portunus trituberculatus along the coastal waters of the East China Sea. Hydrobiologia 618, 125–137 (2009). [Google Scholar]
- He L. J. et al. Late Pleistocene expansion of Scylla paramamosain along the coast of China: a population dynamic response to the Last Interglacial sea level highstand. J. Exp. Mar. Biol. Ecol. 385, 20–28 (2010). [Google Scholar]
- Mukai T., Nakamura S., Suzuki T. & Nishida M. Mitochondrial DNA divergence in yoshinobori gobies (Rhinogobius species complex) between the Bonin Islands and the Japan–Ryukyu Archipelago. Ichthyol. Res. 52, 410–413 (2005). [Google Scholar]
- Mukai T., Nakamura S. & Nishida M. Genetic population structure of a reef goby, Bathygobius cocosensis, in the northwestern Pacific. Ichthyol. Res. 56, 380–387 (2009). [Google Scholar]
- Gordon M. S., Ng W. S. & Yip A. Y. Aspects of the physiology of terrestrial life in amphibious fishes. III. The Chinese mudskipper Periophthalmus cantonensis. J. Exp. Biol. 72, 57–77 (1978). [DOI] [PubMed] [Google Scholar]
- Takita T., Agusnimar & Ali A. B. Distribution and habitat requirements of oxudercine gobies (Gobiidae: Oxudercinae) along the Straits of Malacca. Ichthyol. Res. 46, 131–138 (1998). [Google Scholar]
- Murdy E. O. A taxonomic revision and cladistic analysis of the oxudercine gobies (Gobiidae: Oxudercinae). Rec. Aust. Mus., 11 (Suppl), 1–93 (1989). [Google Scholar]
- Mukai T. & Sugimoto M. Genetic population structure of the mudskipper, Periophthalmus modestus, in Japan inferred from mitochondrial DNA sequence variations. Jpn. J. Ichthyol. 53, 151–158 (2006). [Google Scholar]
- Fang G. H. et al. A note on the South China Sea shallow interocean circulation. Adv. Atmos. Sci. 22, 946–954 (2005). [Google Scholar]
- Xu X. & Oda M. (1999) Surface-water evolution of the eastern China Sea during the last 36,000 years. Mar. Geol. 156, 285–304. [Google Scholar]
- Ujiié Y., Ujiié H., Taira A., Nakamura T. & Oguri K. Spatial and temporal variability of surface water in the Kuroshio source region, Pacific Ocean, over the past 21,000 years: evidence from planktonic foraminifera. Mar. Micropaleontol. 49, 335–364 (2003). [Google Scholar]
- Kojima S., Segawa R. & Hayashi I. Stability of the courses of the warm coastal currents along the Kyushu Island suggested by the population structure of the Japanese Turban shell, Turbo (Batillus) cornutus. J. Oceanogr. 56, 601–604 (2000). [Google Scholar]
- Ogoh K. & Ohmiya Y. Biogeography of luminous marine ostracod driven irreversibly by the Japan current. Mol. Biol. Evol. 22, 1543–1545 (2005). [DOI] [PubMed] [Google Scholar]
- Kuriiwa K., Chiba S. N., Motomura H. & Matsuura K. Phylogeography of Blacktip Grouper, Epinephelus fasciatus (Perciformes: Serranidae), and influence of the Kuroshio Current on cryptic lineages and genetic population structure. Ichthyol. Res. 61, 361–374 (2014). [Google Scholar]
- Virginie M. P. & Jaeger J. J. Island biogeography of the Japanese terrestrial mammal assemblages: an example of a relict fauna. J. Biogeogr. 26, 959–972 (1999). [Google Scholar]
- Ota H. The current geographic faunal pattern of reptiles and amphibians of the Ryukyu archipelago and adjacent regions. Tropics 10, 51–62 (2000). [Google Scholar]
- Nakamura K., Suwa R., Denda T. & Yokota M. Geohistorical and current environmental influences on floristic differentiation in the Ryukyu Archipelago, Japan. J. Biogeogr. 36, 919–928 (2009). [Google Scholar]
- Benzie J. A. H. & Williams S. T. Gene flow among giant clam (Tridacna maxima) populations in the west Pacific is not consistent with dispersal by present-day ocean currents. Evolution 51, 768–783 (1997). [DOI] [PubMed] [Google Scholar]
- Williams S. T. & Benzie J. A. H. Indo-West Pacific patterns of genetic differentiation in the high-dispersal starfish Linckia laevigata. Mol. Ecol. 6, 559–573 (1997). [Google Scholar]
- Benzie J. A. H. Genetic structure of marine organisms and SE Asian biogeography in Biogeography and geological evolution of SE Asia (eds Hall R. & Holloway J. D.) 197–209 (Backhuys Publishers, Leiden, The Netherlands, 1998).
- Templeton A. R., Routman E. & Phillips C. Separating population structure from population history: a cladistic analysis of the geographical distribution of mitochondrial DNA haplotypes in the Tiger Salamander, Ambystoma tigrinum. Genetics 140, 767–782 (1995). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hua X. et al. Phylogeographical analysis of an estuarine fish, Salanx ariakensis (Osmeridae: Salanginae) in the north-western Pacific. J. Fish Biol. 75, 354–367 (2009). [DOI] [PubMed] [Google Scholar]
- Xie C., Jian Z. & Zhao Q. Paleogeographic maps of the China Seas at the last glacial maximum in WESTPAC Paleogeographic Maps 75. (UNESCO/IOC Publications, Shanghai,1995). [Google Scholar]
- Kimura M. Palaeogeography of the Ryukyu Islands. Tropics 10, 5–24 (2000). [Google Scholar]
- Diekmann B. et al. Detrital sediment supply in the southern Okinawa Trough and its relation to sea-level and Kuroshio dynamics during the late Quaternary. Mar. Geol. 255, 83–95 (2008). [Google Scholar]
- Ishimatsu A. et al. Mudskippers brood their eggs in air but submerge them for hatching. J. Exp. Biol. 210, 3946–3954 (2007). [DOI] [PubMed] [Google Scholar]
- Kobayashi T., Dotsu Y. & Miura N. Egg development and rearing experiments of the larvae of the mud skipper, Periophthalmus cantonensis. Bulletin of the Faculty of Fish., Nagasaki University 33, 49–62 (1972). [Google Scholar]
- Bowen B. W., Bass A. L., Rocha L. A., Grant W. S. & Robertson D. R. Phylogeography of the trumpetfishes (Aulostomus): ring species complex on a global scale. Evolution 55, 1029–1039 (2001). [DOI] [PubMed] [Google Scholar]
- Sun X. J., Luo Y., Huang F., Tian J. & Wang P. Deep-sea pollen from the South China Sea: Pleistocene indicators of East Asian monsoon. Mar. Geol. 201, 97–118 (2003). [Google Scholar]
- Xiao J. & An Z. Three large shifts in East Asian monsoon circulation indicated by loess-paleosol sequences in China and late Cenozoic deposits in Japan. Paleogeogr Paleocl. 154, 179–189 (1999). [Google Scholar]
- Han W. X., Fang X. M. & Berger A. Tibet forcing of mid-Pleistocene synchronous enhancement of East Asian winter and summer monsoons revealed by Chinese loess record. Quaternary Res. 78, 174–184 (2012). [Google Scholar]
- Xiao J. L. et al. East Asian monsoon variation during the last 130,000 years: Evidence from the Loess Plateau of central China and Lake Biwa of Japan. Quaternary Sci. Rev. 18, 147–157 (1999). [Google Scholar]
- Wang Y. J. et al. A high-resolution absolute-dated late Pleistocene monsoon record from Hulu Cave, China. Science 294, 2345–2348 (2001). [DOI] [PubMed] [Google Scholar]
- Martin R. E. The fossil record of biodiversity: nutrients, productivity, habitat area and differential preservation. Lethaia 36, 179–194 (2003). [Google Scholar]
- He L.J. et al. (2014) Demographic response of cutlassfish (Trichiurus japonicus and T. nanhaiensis) to fluctuating palaeo-climate and regional oceanographic conditions in the China seas. Sci. Rep.-UK 4, 6380 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Melillo J. M. et al. Global climate-change and terrestrial net primary production. Nature 363, 234–240 (1993). [Google Scholar]
- Xiao Q. G., Chen W. Y., Sheng Y. W. & Guo L. Estimating the net primary productivity in China using meteorological satellite data. Acta Bot. Sin. 38, 35–39 (1996). [Google Scholar]
- Field C.B., Behrenfeld M. J., Randerson J. T. & Falkowski P. Primary production of the biosphere: integrating terrestrial and oceanic components. Science 281, 237–240 (1998). [DOI] [PubMed] [Google Scholar]
- Xiao X. et al. Net primary production of terrestrial ecosystems in China and its equilibrium responses to changes in climate and atmospheric CO2 concentration. Acta Phytoeco. Sin. 22, 97–118 (1998). [Google Scholar]
- Fang J. Y. Forest productivity in China and its responses to global climate change. Acta Phytoecoligica Sinica 24, 513–517 (2000). [Google Scholar]
- Tao B., Li K. R., Shao X. M. & Cao M. K. The temporal and spatial patterns of terrestrial net primary productivity in China. J. Geogr. Sci. 13, 163–171 (2003). [Google Scholar]
- Yi S., Saito Y., Zhao Q. H. & Wang P. X. Vegetation and climate changes in the Changjiang (Yangtze River) Delta, China, during the past 13,000 years inferred from pollen records. Quaternary Sci. Rev. 22, 1501–1519 (2003). [Google Scholar]
- Crucifix M., Betts R. A. & Hewitt C. D. Pre-industrial-potential and Last Glacial Maximum global vegetation simulated with a coupled climate-biosphere model: Diagnosis of bioclimatic relationships. Global Planet Change 45, 295–312 (2005). [Google Scholar]
- Barron E. J., Hay W. W. & Thompson S. The hydrologic cycle: A major variable during earth history. Palaeogeogr. Palaeoclimatol. Palaeoecol. 75, 157–174 (1989). [Google Scholar]
- Herbeck L. S., Unger D., Krumme U., Liu S.M. & Jennerjahn T. C. Typhoon-induced precipitation impact on nutrient and suspended matter dynamics of a tropical estuary affected by human activities in Hainan, China. Estuar Coast Shelf S. 93, 375–388 (2011). [Google Scholar]
- Zhou W. H., Yin K. D., Harrison P. J. & Lee J. H. W. The influence of late summer typhoons and high river discharge on water quality in Hong Kong waters. Estuar. Coast. Shelf S. 111, 35–47 (2012). [Google Scholar]
- Newell R. I. E., Marshall N., Sasekumar A. & Chong V. C. Relative importance of benthic microalgae, phytoplankton, and mangroves as sources of nutrition for penaeid prawns and other coastal invertebrates from Malaysia. Mar. Biol. 123, 595–606 (1995). [Google Scholar]
- Sauriau P. & Kang C. Stable isotope evidence of benthic microalgae-based growth and secondary production in the suspension feeder Cerastoderma edule (Mollusca, Bivalvia) in the Marennes-Olerin Bay. Hydrobiologia, 440, 317–329 (2000). [Google Scholar]
- Ning X. R., Cai Y. M., Liu Z. L., Hu X. G. & Liu C. G. Standing crop and primary production of benthic microalgae on tidal flats in the Sanggou and Jiaozhou Bays, China. Acta Oceanol. Sin. 22, 75–87 (2003). [Google Scholar]
- Li C. X., Chen Q. Q., Zhang J. Q., Yang S. Y. & Fan D. D. Stratigraphy and paleoenvironmental changes in the Yangtze Delta during the Late Quaternary. J. Asian Earth Sci. 18, 453–469 (2000). [Google Scholar]
- Hori K., Saito Y., Zhao Q. H. & Wang P. X. Architecture and evolution of the tide-dominated Changjiang (Yangtze) River delta, China. Sediment. Geol. 146, 249–264 (2002). [Google Scholar]
- Liu J. P. et al. Flux and fate of Yangtze River sediment delivered to the East China Sea. Geomorphology 85, 208–224 (2007). [Google Scholar]
- Cartaxana P., Mendes C. R., van Leeuwe M.A. & Brotas V. Comparative study on microphytobenthic pigments of muddy and sandy intertidal sediments of the Tagus estuary. Estuar. Coast. Shelf S. 66, 225–230 (2006). [Google Scholar]
- Du G.Y., Son M., Yun M., An S. & Chung I. K. Microphytobenthic biomass and species composition in intertidal flats of the Nakdong River estuary, Korea. Estuar. Coast. Shelf S. 82, 663–672 (2009). [Google Scholar]
- Taylor B. & Hayes D. Origin and history of the South China Sea basin. AGU Geophys. Monogr. 27, 23–56 (1983). [Google Scholar]
- Letouzey J. & Kimura M. Okinawa Trough genesis; structure and evolution of a backarc basin developed in a continent. Mar. Petrol. Geol. 2, 111–130 (1985). [Google Scholar]
- Kimura M. Genesis and formation of the Okinawa Trough, Japan. Memoir of Geological Society, Japan 34, 77–88 (1990). [Google Scholar]
- Tanaka G. & Nomura S. Late Miocene and Pliocene Ostracoda from the Shimajiri Group, Kume-jima Island, Japan: Biogeographical significance of the timing of the formation of back-arc basin (Okinawa Trough). Palaeogeogr. Palaeocl. 276, 56–68 (2009). [Google Scholar]
- Maggs C. A. et al. Evaluating signatures of glacial refugia for North Atlantic benthic marine taxa. Ecology 89, S108–122 (2008). [DOI] [PubMed] [Google Scholar]
- Wang K. L., Liu L. Y., You F. & Xu C. Studies on the genetic variation and systematics of the hairtails fishes from the South China Sea. Mar. Sci. 2, 69–72 (1992). [Google Scholar]
- Keenan C. P., Davies P. J. F. & Mann D. L. A revision of the genus Scylla de Haan, 1833 (Crustacea: Decapoda: Brachyura: Portunidae). Raffles B. Zool. 46, 217–245 (1998). [Google Scholar]
- Shih H. T., Ng P. K. L., Fang S. H., Chan B. K. K. & Wong K. J. H. Diversity and distribution of fiddler crabs (Brachyura: Ocypodidae: Uca) from China, with new records from Hainan Island in the South China Sea. Zootaxa 2640, 1–19 (2010). [Google Scholar]
- Cann R. L., Stoneking M. & Wilson A. C. Mitochondrial DNA and human evolution. Nature 325, 31–36 (1987). [DOI] [PubMed] [Google Scholar]
- Zhang D. X. & Hewitt G. M. Isolation of animal cellular total DNA in Molecular Tools for Screening Biodiversity: Plants and Animals (eds. Karp A., Isaac P. G. & Ingram D. S.) 5–9 (Chapman & Hall, 1998).
- Miya M. & Nishida M. Use of Mitogenomic information in teleostean molecular phylogenetics: a tree-based exploration under the maximum parsimony optimality criterion. Mol. Phylogenet. Evol. 17, 437–455 (2000). [DOI] [PubMed] [Google Scholar]
- Thompson J. D., Gibson T. J., Plewniak F., Jeanmougin F. & Higgins D. G. The ClustalX windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 25, 4876–4882 (1997). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Posada D. & Crandall K. A. Modeltest: testing the model of DNA substitution. Bioinformatics 14, 817–818 (1998). [DOI] [PubMed] [Google Scholar]
- Tamura K. & Nei M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 10, 512–526 (1993). [DOI] [PubMed] [Google Scholar]
- Saitou N. & Nei M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 4, 406–425 (1987). [DOI] [PubMed] [Google Scholar]
- Felsenstein J. Evolutionary trees from DNA sequences: A maximum likelihood approach. J. Mol. Evol. 17, 368–376 (1981). [DOI] [PubMed] [Google Scholar]
- Huelsenbeck J. P. & Ronquist F. MRBAYES: Bayesian inference of phylogeny. Bioinformatics 17, 754–755 (2001). [DOI] [PubMed] [Google Scholar]
- Bandelt H. J., Forster P. & Röhl A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 16, 37–48 (1999). [DOI] [PubMed] [Google Scholar]
- Swofford D. L. PAUP*: phylogenetic analysis using parsimony and other methods, beta version 4b10. Sinauer Associates, Sunderland, Massachusetts (2002). [Google Scholar]
- Ronquist F. et al. MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 61, 539–542 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Strimmer K. & von Haeseler A. Likelihood-mapping: A simple method to visualize phylogenetic content of a sequence alignment. Proc. Natl. Acad. Sci. USA. 94, 6815–6819 (1997). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schmidt H.A., Strimmer K., Vingron M. & von Haeseler A. TREE-PUZZLE: maximum likelihood phylogenetic analysis using quartets and parallel computing. Bioinformatics 18, 502–504 (2002). [DOI] [PubMed] [Google Scholar]
- Shimodaira H. & Hasegawa M. Multiple comparisons of loglikelihoods with applications to phylogenetic inference. Mol. Biol. Evol. 16, 1114–1116 (1999). [Google Scholar]
- Rambaut A. & Grassly N. C. Seq-Gen: An application for the Monte Carlo simulation of DNA sequence evolution along phylogenetic trees. Comput. Appl. Biosci. 13, 235–238 (1997). [DOI] [PubMed] [Google Scholar]
- Excoffier L. & Lischer H. E. L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 10, 564–567 (2010). [DOI] [PubMed] [Google Scholar]
- Rice W. R. Analyzing tables of statistical tests. Evolution 43, 223–225 (1989). [DOI] [PubMed] [Google Scholar]
- Rogers A. R. & Harpending H. Population growth makes waves in the distribution of pairwise genetic divergences. Mol. Biol. Evol. 9, 552–569 (1992). [DOI] [PubMed] [Google Scholar]
- Tajima F. Statistical method for testing the neutral mutation hypothesis by DNA polymorphism. Genetics 123, 585–595 (1989a). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tajima F. The effect of change in population size on population DNA polymorphism. Genetics 123, 597–601 (1989b). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fu Y. X. Statistical tests of neutrality of mutations against population growth, hitchhiking and background selection. Genetics 147, 915–925 (1997). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Slatkin M. & Hudson R. H. Pairwise comparisons of mitochondrial DNA sequences in stable and exponentially growing populations. Genetics 129, 555–562 (1991). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Drummond A. J., Suchard M. A., Xie D. & Rambaut A. Bayesian phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 29, 1969–1973 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rambaut A., Suchard M. & Drummond A. J. Tracer v1.6. Available from http:// tree.bio.ed.ac.uk/software/tracer/ (2013) Date of access: 11/12/2013.
- Rosenberg N. A. & Feldman M. W. The relationship between coalescence times and population divergence times in Modern Developments in Theoretical Population Genetics (eds. Slatkin M. & Veuille M.) 130–164. (Oxford University Press, New York, 2002)
- Spalding M. D. et al. Marine ecoregions of the world: A bioregionalization of coastal and shelf areas. Bioscience 57, 573–593 (2007). [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.