

Abstract
Despite its importance during viral or bacterial infections, transcriptional regulation of the interferon-β gene (Ifnb1) in activated macrophages is only partially understood. Here we report that TRIM33 deficiency results in high, sustained expression of Ifnb1 at late stages of toll-like receptor-mediated activation in macrophages but not in fibroblasts. In macrophages, TRIM33 is recruited by PU.1 to a conserved region, the Ifnb1 Control Element (ICE), located 15 kb upstream of the Ifnb1 transcription start site. ICE constitutively interacts with Ifnb1 through a TRIM33-independent chromatin loop. At late phases of lipopolysaccharide activation of macrophages, TRIM33 is bound to ICE, regulates Ifnb1 enhanceosome loading, controls Ifnb1 chromatin structure and represses Ifnb1 gene transcription by preventing recruitment of CBP/p300. These results characterize a previously unknown mechanism of macrophage-specific regulation of Ifnb1 transcription whereby TRIM33 is critical for Ifnb1 gene transcription shutdown.
 Transcriptional regulation of the interferon-β gene (Ifnb1) in macrophages is a critical immune event. Here, Ferri et al. show that, at late phases of macrophages activation, TRIM33 bound to a distal repressor element suppresses Ifnb1 transcription by preventing recruitment of CBP/p300.
Transcriptional regulation of the interferon-β gene (Ifnb1) in macrophages is a critical immune event. Here, Ferri et al. show that, at late phases of macrophages activation, TRIM33 bound to a distal repressor element suppresses Ifnb1 transcription by preventing recruitment of CBP/p300.
In response to viral or bacterial infections, germ line-encoded pattern recognition receptors induce type I interferon (IFN-I, encoded by 14 Ifna and 1 Ifnb genes in mice) gene transcription in myeloid and non-myeloid cells1. The secreted type I IFNs act early and transiently during the innate immune response to prime adaptive immunity and then type I Ifn gene transcription is shut down2. This transient expression of IFN-I is physiologically important. Whereas early IFN-I expression is important to control infection, pathogens that can overcome this initial control benefit from IFN-I mediated immune system suppression3,4.
Among type I Ifn genes, Ifnb1 gene regulation has been a model system to study inducible transcription. After pattern recognition receptor-mediated activation, a group of transcription factors, including IRF3/IRF7, NF-κB and ATF2/c-jun, cooperatively binds to a regulatory sequence located between −102 and −47 bp upstream of the Ifnb1 transcriptional start site (TSS) to form the Ifnb1 enhanceosome. This enhanceosome recruits CBP/p300 for acetylation of histone H3 and permits recruitment of chromatin-remodeling complexes for initiation of transcription5.
Monocyte/macrophage-specific regulation of Ifnb1 gene transcription has only been observed in the modulation of the transcriptional activity of IRF3. In monocytes, a PU.1-binding site located in the enhanceosome is constitutively bound by a IRF8/PU.1 myeloid complex that creates a preformed activation complex. This myeloid complex can recruit IRF3 through direct IRF3/IRF8 interaction and is involved in the rapid transcription of Ifnb1 observed in monocytes after pathogenic stimulation6. Conversely, transcriptional regulator MafB can impair the interactions between IRF3 bound to the enhanceosome and co-activators, and thus negatively regulates Ifnb1 transcription7. Finally, binding of transcriptional regulator YY1 to different sites within the enhanceosome has been shown to result in activation (at the beginning) or repression (at the end) of Ifnb1 gene transcription in fibroblasts8,9,10. Altogether, these results point to the enhanceosome as a regulator of both initiation and termination of the Ifnb1 transcription cycle and of monocyte/macrophage-specific regulation of Ifnb1 transcription.
The tripartite motif (TRIM) protein family is characterized by a shared N-terminal structure consisting of a RING, two B-box domains, a coiled-coil domain and, for most of the TRIM family members, an E3 ubiquitin ligase activity11. TRIM33, together with TRIM24, TRIM28 and TRIM66, form a sub-family of TRIM proteins characterized by a C-terminal plant homeodomain juxtaposed to a bromodomain12. These TRIM proteins do not directly bind to DNA but can be recruited by DNA-binding proteins, such as nuclear receptors13, SMAD14, PU.1 or TAL-1 (refs 15, 16), to act as transcriptional regulators.
A large number of TRIM proteins are innate immune enhancers and act at multiple levels in signalling pathways, including production and action of IFN-β to confer antiviral and antibacterial cytokine production17,18. As for the TRIM33 sub-family, TRIM24 represses the viral-defense IFN response in hepatocytes as a consequence of repression of VL30 transcription13,19. Although these TRIM24 mediated repressions are potentiated by TRIM33, TRIM33 deficiency alone cannot elicit the IFN response in hepatocytes13,19. TRIM24 also acts as a negative regulator of the IFN/signal transducers and activators of the transcription signalling pathway13,19, and TRIM33 is essential for cytosolic RNA-induced NLRP3 inflammasome activation20. However, up to now, no TRIM protein has been shown to directly regulate Ifnb1 gene transcription after recruitment to an Ifnb1 regulatory sequence.
We have previously shown that TRIM33 can interact with PU.1 to repress its transcriptional activity during haematopoiesis and suggested that TRIM33 may regulate myeloid fate and have a role in macrophages15. Here, we show that TRIM33 deficiency in macrophages results in sustained expression of Ifnb1 during the late phases of macrophages activation. Our results indicate that TRIM33, recruited by PU.1 on a distal regulatory element of the Ifnb1 gene, regulates Ifnb1 enhanceosome loading and shuts down Ifnb1 gene transcription during the late stages of lipopolysaccharide (LPS) activation of macrophages by preventing recruitment of CBP/p300.
Results
TRIM33 controls Ifnb1 expression in activated macrophages
Trim33fl/fl mice were crossed with mice expressing the Cre recombinase from the endogenous Lysozyme2 locus (LyzCre) to generate Trim33−/− mice. Trim33−/− mice did not display developmental abnormalities and were healthy. PCR genotyping of purified haematopoietic cell populations showed deletion of Trim33 in monocytes, bone-marrow-derived macrophages (BMDM), peritoneal macrophages (PMs) and neutrophils but not in myeloid progenitors, nor in B lymphocytes (Fig. 1a, upper panels and Supplementary Fig. 1a). The deletion of the Trim33 gene allele was associated with a loss of TRIM33 expression in BMDM (Fig. 1a, lower panel).
Figure 1. TRIM33 regulates Ifnb1 gene expression in activated macrophages.

(a) Genomic PCR detection of Trim33 in BMDM and PM (top) and western blot analysis of TRIM33 in BMDM from WT and Trim33−/− mice (bottom). (b) Kinetics of Ifnb1 mRNA levels in WT and Trim33−/− BMDM (left panel) or PM (right panel) after activation with LPS. Mean±s.e.m., n=3 to 6. (c) IFN-β protein levels in supernatants of non-activated and LPS-activated WT and Trim33−/− BMDM. Mean±s.e.m., n=4. (d) Relative mRNA levels of Ifnb1 and IFN-β target genes in Trim33−/− BMDM versus WT BMDM treated with LPS (0.1 ng ml−1) for 3 or 24 h. Mean±s.e.m., n=3. (e) Kinetics of Ifna mRNA levels in WT and Trim33−/− BMDM activated with LPS. Mean±s.e.m., n=3. (f) Kinetics of Ifnb1 mRNA levels in NIH3T3 cells expressing a shRNA targeting Trim33 (NIH3T3 TRIM33−) or luciferase (NIH3T3 TRIM33+) and activated with poly(I:C). Mean±s.e.m., n=3. (g) Kinetics of Ifnb1 mRNA levels in WT and Trim33−/− iM treated for the indicated times with LPS. Mean±s.e.m., n=3. (h) Relative expression of Ifnb1 mRNA 0, 2 and 8 h after LPS treatment of Trim33−/− iM transduced with a lentivirus coding for GFP (iM Trim33−/− +GFP), for full-length flag-TRIM33 and GFP (iM Trim33−/− +flag-TRIM33) or for flag-TRIM33 mutants lacking the coiled-coil domain (iM Trim33−/− +flag-ΔCC), the ubiquitin ligase activity (iM Trim33−/− +flag-ΔUb) or the SMAD-interaction domain (iM Trim33−/− +flag-ΔSMAD). Mean±s.e.m., n=5. All qRT-PCR data are normalized to HPRT and, unless otherwise indicated, presented relative to expression of untreated WT cells. *P<0.05; **P<0.01 and ***P<0.001, Mann–Whitney test.
The effects of Trim33 deletion on Ifnb1 expression in macrophages were studied by comparing kinetics of Ifnb1 mRNA levels in wild-type (WT) or Trim33−/− BMDM and PM, upon TLR4 activation by LPS or upon TLR3 activation by poly(I:C). These two activation protocols mimic bacterial (LPS) or viral (poly(I:C)) infections. A rapid early increase of Ifnb1 mRNA levels was observed in WT and Trim33−/− BMDM or PM activated by LPS (Fig. 1b). Thereafter, Ifnb1 mRNA levels decreased from 2 h in WT BMDM (or 4 h in WT PM) and returned to basal levels by 6 h in WT BMDM and PM. In contrast, although Ifnb1 mRNA levels decreased between 2 and 4 h, Ifnb1 mRNA levels began to rise significantly by 6 h and remained high 24 h after LPS activation in Trim33−/− BMDM or PM (Fig. 1b). This late sustained expression of Ifnb1 was also observed when BMDM and PM were stimulated by poly(I:C) with Ifnb1 mRNA levels being much higher at all late time points in poly(I:C) treated Trim33−/− BMDM compared with WT (Supplementary Fig. 1b). The increased Ifnb1 mRNA levels in Trim33−/− BMDM activated with LPS correlated with increased levels of secreted IFN-β (Fig. 1c). Finally, using a sub-optimal concentration of LPS (0.1 ng ml−1) which did not activate Ifnb1 in WT BMDM, we showed, in LPS-activated Trim33−/− BMDM, an increased expression of genes whose transcription is dependent on IFN-β and occurs late during LPS activation (Fig. 1d). This increased expression was not accounted for by different responses of WT and Trim33−/− BMDM to IFN-β (Supplementary Fig. 1c) and could be obtained after addition of IFN-β during the late phases of LPS activation of WT BMDM (Supplementary Fig. 1d). Altogether, these results show that TRIM33 deficiency is associated with increased Ifnb1 mRNA level associated with IFN-β secretion during the late stages of LPS activation of BMDM.
To study if TRIM33 deficiency affects the expression of other Ifn genes, quantitative RT-PCR was performed and showed similar kinetics of Ifna mRNA levels in WT or Trim33−/− BMDM with no late increased expression of Ifna mRNA levels in Trim33−/− BMDM (Fig. 1e). Finally, highly efficient shRNA-mediated depletion of TRIM33 in NIH3T3 cells (NIH3T3 TRIM33−; Supplementary Fig. 1e) did not modify kinetics of Ifnb1 mRNA levels in response to poly(I:C) (Fig. 1f), indicating cell-specificity in the regulation of Ifnb1 expression by TRIM33.
Potential direct or indirect roles for TRIM33 in Ifnb1 expression were addressed in ‘rescue' experiments in which TRIM33 protein was added back to LPS-activated immortalized Trim33−/− macrophages (iM)21. As anticipated, Trim33−/− iM bore the expected deletion of Trim33 gene and these cells expressed no TRIM33 protein (Supplementary Fig. 1f). Ifnb1 mRNA levels increased 1 h after LPS activation of WT and Trim33−/− iM and peaked at 2 h with twofold higher observed expression in WT iM than in Trim33−/− iM (Fig. 1g). As anticipated, Ifnb1 mRNA levels in WT iM then dropped, whereas in Trim33−/− iM, Ifnb1 mRNA levels remained high (Fig. 1g). Thus, LPS activation of Trim33−/− iM mimicked the late phase of the Ifnb1 mRNA kinetics seen in Trim33−/− BMDM. To address whether expression of ectopic TRIM33 could rescue Ifnb1 downregulation at late time points, a flag-TRIM33-IRES-GFP (or GFP alone) construct was expressed in Trim33−/− iM and cells were stimulated with LPS (Fig. 1h). The full-length exogenous TRIM33 protein restored Ifnb1 downregulation at late time points of LPS activation of Trim33−/− iM (Fig. 1h) indicating a role for TRIM33 in the regulation of Ifnb1 gene expression during LPS activation of macrophage.
To determine the TRIM33 protein domains involved in Ifnb1 regulation, mutant flag-TRIM33-IRES-GFP constructs lacking the coiled-coil domain (flag-ΔCC), the ubiquitin ligase activity (flag-ΔUb) or the Smad-interaction domain (flag-ΔSmad) were expressed in Trim33−/− iM (Supplementary Fig. 1g). The flag-ΔUb and the flag-ΔSmad rescued normal Ifnb1 expression at late time points of LPS-activated Trim33−/− iM whereas the flag-ΔCC did not rescue normal profile of Ifnb1 expression in activated Trim33−/− iM (Fig. 1h). These results indicate that the Smad-interaction and ubiquitin ligase domains are not necessary for TRIM33 regulation of Ifnb1 expression and suggest that TRIM33 is part of a regulatory complex that regulates Ifnb1 transcription.
Altogether, these results reveal a role for TRIM33 in the regulation of Ifnb1 expression specifically in downregulating its expression during the late stages of macrophage activation.
TRIM33 binds a distal Ifnb1 regulatory element
TRIM33 ChIP-seq data obtained from the RAW 264.7 murine macrophage cell line or from BMDM treated with LPS for 24 h showed a single TRIM33 peak at the Ifnb1 locus that overlapped with a PU.1 peak located 15 kb upstream from the Ifnb1 TSS (Fig. 2a). Interestingly, no TRIM33 occupancy was observed at the PU.1 peak located just upstream the Ifnb1 promoter corresponding to the enhanceosome-binding sites. We called this −15 kb region the Ifnb1 Control Element (ICE). Recently, the human homologue of ICE, named L2, has been shown to be a virus-inducible enhancer of Ifnb1 in non-myeloid cells where it displays promoter and enhancer activities22.
Figure 2. TRIM33 is bound to a distal Ifnb1 gene regulatory element (ICE) in macrophages.

(a) TRIM33 ChIP-seq data in RAW 264.7 cells and BMDM activated for 24 h with LPS showing TRIM33 binding to ICE, located 15 kb upstream the Ifnb1 TSS. Blue boxes indicate PU.1 peaks in BMDM (data from ref. 43). Green boxes indicate positions of ICE and the enhanceosome. (b) ChIP-qPCR data for TRIM33 binding on ICE in WT BMDM (left panel) and for PU.1 binding on Ifnb1 promoter and on ICE (right panels) in WT and Trim33−/− BMDM at indicated time points after LPS activation. Data represent the enrichment over a negative control region in the β-globin promoter. Mean±s.e.m., n=3. (c) ChIP-qPCR analysis of TRIM33 binding at the Ifnb1 promoter and on ICE in NIH3T3 cells. Mean±s.e.m., n=3. (d) Kinetics of Ifnb1 mRNA levels in NIH3T3 cells expressing PU.1 and a shRNA targeting Trim33 (NIH3T3 PU.1+ TRIM33−) or PU.1 and an shRNA targeting luciferase (NIH3T3 PU.1+ TRIM33+) and activated with poly(I:C). Mean±s.e.m., n=2. (e) Luciferase reporter assay in RAW 264.7 cells transfected with reporter constructs containing the Ifnb1 promoter alone (Ifnb1 Prom-luc) or ICE cloned upstream of the Ifnb1 promoter (ICE-Ifnb1 Prom-Luc) and activated with LPS. Mean±s.e.m., n=3. (f,g) Fold increase of Ifnb1 mRNA levels in ICE+/− versus WT RAW 264.7 cells after LPS activation (f) and in ICE+/− versus WT NIH3T3 cells after poly(I:C) activation (g). Mean±s.e.m., n=3. (h) Fold increase of Ifnb1 mRNA in ICE−/− versus WT RAW 264.7 cells after LPS (left) or poly(I:C) (right) activation. Mean±s.e.m., n=3 to 5. Clones carrying different deletions at the PU.1/TRIM33 site in ICE are indicated in brackets. *P<0.05 and **P<0.01, Mann–Whitney test.
Constitutive TRIM33 occupancy at ICE was observed in BMDM although occupancy appeared to increase at late time points (24 h) following LPS activation (Fig. 2b, left panel). PU.1 binding at the Ifnb1 promoter and at ICE was constitutive and detected in WT and Trim33−/− BMDM (Fig. 2b, middle and right panels) indicating that PU.1 binding to ICE was not TRIM33-dependent. Finally, no TRIM33 binding was observed on the Ifnb1 promoter or ICE in NIH3T3 cells (Fig. 2c), indicating macrophage specificity of TRIM33 in binding to ICE.
To determine if TRIM33-mediated regulation of Ifnb1 in myeloid cells can be obtained in NIH3T3 by simultaneous expression of PU.1 and TRIM33, NIH3T3 cells were transduced with a PU.1 expressing lentivirus (Supplementary Fig. 2a). PU.1 expressing NIH3T3 were then transduced with the lentivirus that promoted efficient shRNA-mediated depletion of TRIM33 (Supplementary Fig. 1e) and NIH3T3 PU.1+TRIM33+ and NIH3T3 PU.1+TRIM33− responses to poly(I:C) were studied. Whereas TRIM33 deficiency did not modify kinetics of Ifnb1 mRNA levels in response to poly(I:C) in normal NIH3T3 cells (Fig. 1f), TRIM33 deficiency increased Ifnb1 mRNA levels in NIH3T3 PU.1+ cells treated with poly(I:C) but did not result in sustained expression of Ifnb1 during the late stages of poly(I:C) activation (Fig. 2d). These results show that TRIM33 can repress Ifnb1 expression in presence of PU.1 and indicate that additional factors that are not present in NIH3T3 cells are required for TRIM33-mediated late downregulation of Ifnb1 expression.
To study the effect of ICE on Ifnb1 promoter activity in myeloid cells, we used RAW 264.7 cells in which LPS regulates Ifnb1 gene expression (Supplementary Fig. 2b). RAW 264.7 cells were transfected with luciferase reporter constructs containing a 391 bp fragment of the Ifnb1 promoter preceded by a 408 bp fragment corresponding to the central region of ICE (ICE-Ifnb1 Prom-Luc) or the fragment containing the Ifnb1 promoter alone as a control sequence (Ifnb1 Prom-Luc). Transcriptional activity of ICE-reporter (ICE-Ifnb1 Prom-Luc) showed four to tenfold less activity than the promoter reporter in LPS-activated RAW 264.7 cells indicating that ICE functions as a cis-acting transcriptional repressor element in RAW 264.7 cells (Fig. 2e).
To study the role of ICE in RAW 264.7 and NIH3T3 cells, we used a CRISPR/Cas9 lentiviral system to deliver Cas9 and gRNA (Supplementary Fig. 2c). Using this system, we had to screen 200 RAW 264.7 clones to get one RAW 264.7 clone with monoallelic deletion of ICE (Supplementary Fig. 2d). This low efficiency of recombination in RAW 264.7 cells does not favour the use of transient transfection for CRISPR/Cas9-mediated deletion of ICE, transient transfection efficiency in RAW 264.7 being 15% compared with 90% using the lentiviral system. RAW 264.7 and NIH3T3 clones bearing this deletion on one allele (termed RAW ICE+/− and NIH3T3 ICE+/−, respectively; Supplementary Fig. 2d) were analysed for Ifnb1 expression after LPS or poly(I:C) activation. In LPS-activated RAW WT cells, Ifnb1 mRNA levels increased during the first 4 h and then gradually decreased (Supplementary Fig. 2e). In contrast, whereas Ifnb1 mRNA levels increased comparably in RAW ICE+/− cells, they subsequently remained stable (Fig. 2f and Supplementary Fig. 2e). In NIH3T3 WT and ICE+/− cells activated with poly(I:C), kinetics of Ifnb1 mRNA levels were similar (Fig. 2g and Supplementary Fig. 2f). These results indicate that ICE plays a role in the negative regulation of Ifnb1 expression at late stages of LPS activation in RAW cells.
Sequential deletions centered on the PU.1/TRIM33-binding site of ICE were performed in RAW 264.7 cells using CRISPR/Cas9 technology (Supplementary Fig. 2g). Of the 96 clones screened, two carried a biallelic deletion of ICE (Supplementary Fig. 2d). Two RAW 264.7 clones bearing a 100 or 176 bp deletion on the two Ifnb1 alleles (termed #34 and #15; Supplementary Fig. 2g) were analysed for Ifnb1 expression after LPS or poly(I:C) activation. In the two clones high and sustained Ifnb1 mRNA levels were found but only after LPS or poly(I:C) activation, with the highest increase being obtained during the late phase of activation (Fig. 2h).
Altogether, these results show that ICE functions as a cis-acting transcriptional repressor element of the Ifnb1 gene activation in macrophages and indicate that TRIM33 and PU.1 bound to ICE may have an important role in Ifnb1 gene transcription shutdown during the late phase of macrophage activation.
ICE chromatin structure and interaction with Ifnb1 in BMDM
Comparison of reported human and mouse ICE sequences showed 70% sequence homology around the TRIM33/PU.1 peak with consensus binding motifs for AP-1, NF-κB and IRF transcription factors (Supplementary Fig. 3a, left panel). Furthermore, c-JUN, the p65 subunit of NF-κB and IRF3, three transcription factors that form the Ifnb1 enhanceosome and regulate its transcriptional activity, were also bound to ICE three hours after LPS activation of BMDM (Supplementary Fig. 3a, right panel; data from ref. 23). We studied a possible TRIM33 dependency of c-jun and p65 binding to ICE but found that TRIM33 deficiency did not modify the kinetics of their binding to ICE (Fig. 3a).
Figure 3. ICE chromatin structure and interaction with the Ifnb1 promoter during activation of macrophages.

(a) ChIP-qPCR analysis of c-jun or p65 binding at ICE in WT and Trim33−/− BMDM treated for the indicated times with LPS. Mean±s.e.m., n=2 to 4. (b) ChIP-qPCR analysis of RNA Pol II (upper left panel) and H3K4me3 (lower left panel) at ICE in WT and Trim33−/− BMDM treated for the indicated times with LPS. Mean±s.e.m., n=3 to 5. (Right panel) UCSC genome browser images showing H3K4me3 ChIP-seq profiles at ICE in WT and Trim33−/− BMDM treated for the indicated times with LPS. (c) ChIP-qPCR analysis of acetylated histone H3 (left panel) and CBP/p300 (right panel) at ICE in WT and Trim33−/− BMDM treated for the indicated times with LPS. Mean±s.e.m., n=4. **P<0.01, Mann–Whitney test. (d) DNA looping at the Ifnb1 locus was determined by 3C-seq performed before or 24 h after LPS activation of WT and Trim33−/− BMDM using either ICE (upper panel) or Ifnb1 gene (lower panel) as viewpoints (shown by an eye and a yellow band). Data represent normalized reads per million (r.p.m.) per restriction fragment. The x axis shows the genomic coordinates of the Ifnb1 locus. The positions of the EcoRI restriction sites and the TRIM33 ChIP-seq profile in BMDM are indicated on the top.
Analysis of ICE chromatin structure in macrophages showed that this region is characterized by high levels of H3K4me3 and H3K27ac modification, flanked by regions enriched for H3K4me1, all features of transcriptionally active promoters (Supplementary Fig. 3b, red graph, data from ref. 24). These epigenetic modifications are present in short-term haematopoietic stem cells (ST-HSCs) and in myeloid cells, but not in mature erythroid and lymphoid cells (Supplementary Fig. 3b). We studied the effect of TRIM33 deficiency on the chromatin structure of ICE in non-activated and LPS-activated BMDM to ask whether TRIM33 has a role in histone modification of this region. Before and during LPS activation of WT and Trim33−/− BMDM, no significant differences in H3K4me1 levels on ICE could be detected (Supplementary Fig. 3c). ChIP and ChIP-seq analyses showed a constitutive binding of RNA Polymerase II (Pol II) and constitutively high levels of H3K4me3 on ICE (Fig. 3b). Prior to LPS activation, histone H3 acetylation (H3ac) on ICE was similar in WT and Trim33−/− BMDM (Fig. 3c, left panel). During LPS activation, H3ac on ICE did not change in WT BMDM but was transiently increased 10 h after LPS activation in Trim33−/− BMDM (Fig. 3c, left panel). This increased H3ac on ICE in Trim33−/− BMDM was associated with transient increase of CBP/p300 histone H3 acetyl-transferase (HAT) -binding on ICE 10 h after LPS activation (Fig. 3c, right panel). Collectively, database analyses and our results suggest that, in BMDM, ICE exhibits a promoter-like chromatin signature established early during myeloid differentiation. TRIM33 deficiency did not modify ICE chromatin signature and only led to transient increases of H3ac and CBP/p300 recruitment on ICE whereas a sustained high expression of Ifnb1 was observed in LPS-activated Trim33−/− BMDM suggesting that TRIM33 bound to ICE might act at a distance.
To investigate how TRIM33 bound to ICE, that is located 15 kb upstream of the Ifnb1 transcription unit, may regulate Ifnb1 transcription, chromosome conformation capture experiments followed by deep sequencing (3C-seq) were performed in WT and Trim33−/− BMDM before and after LPS activation. ICE (Fig. 3d, top) and the Ifnb1 gene (Fig. 3d, bottom), including its promoter, were used as viewpoints. In WT BMDM, both viewpoints showed a loop between ICE and the Ifnb1 gene before LPS activation and a twofold stronger interaction 24 h after LPS activation (Fig. 3d, blue curves). Interestingly, Trim33 deletion did not significantly modify DNA looping in this region, even after LPS treatment (Fig. 3d, compare red (Trim33−/−) and blue (WT) curves). These results indicate that ICE interacts with the Ifnb1 proximal region in a constitutive and TRIM33-independent manner, but that this interaction is strengthened following LPS stimulation.
TRIM33 represses Ifnb1 by preventing CBP/p300 recruitment
To characterize how TRIM33 might regulate Ifnb1 transcription, we studied the effect of TRIM33 deficiency on temporal binding of p65 and c-jun to the enhanceosome as these two factors are major regulators of its activity. As anticipated, in WT BMDM, c-jun and p65 were recruited to the Ifnb1 promoter at early times following LPS activation (1 h), but their binding returned to basal levels after 24 h (Fig. 4a). In contrast, in the absence of TRIM33, p65 and c-jun binding to the Ifnb1 promoter was maintained even 24 h after activation (Fig. 4a). Thus, TRIM33 deficiency maintains enhanceosome loading by c-jun and p65 during the late phases of LPS activation.
Figure 4. TRIM33 regulates Ifnb1 gene expression through inhibition of CBP/p300 recruitment.

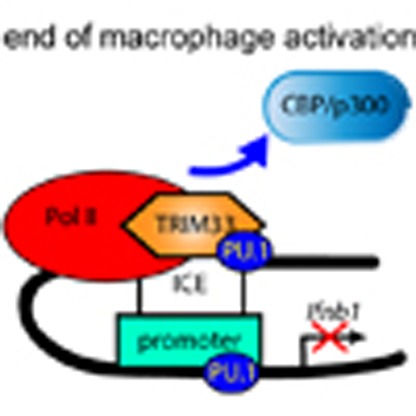
(a) ChIP-qPCR of p65 (left panel) and c-jun (right panel) at the Ifnb1 promoter in WT and Trim33−/− BMDM treated with LPS. Mean±s.e.m., n=2 to 4. (b, top) ChIP-qPCR of RNA Pol II at the Ifnb1 promoter (left panel) and at a region located +2.5 kb from TSS (middle panel) in WT and Trim33−/− BMDM treated with LPS. Mean±s.e.m., n=3 to 5. (Bottom) ChIP-qPCR (left panel) and ChIP-seq (right panel) of H3K4me3 at the Ifnb1 promoter in WT and Trim33−/− BMDM treated with LPS. Mean±s.e.m., n=3. (c) ChIP-qPCR of H3ac at the Ifnb1 promoter in WT and Trim33−/− BMDM treated with LPS. Mean±s.e.m., n=4. (d) ChIP-qPCR of H3ac at the Ifnb1 promoter in Trim33−/− iM expressing GFP (iM Trim33−/− +GFP), full-length flag-TRIM33 and GFP (iM Trim33−/−+flag-TRIM33), or flag-TRIM33 lacking the coiled-coil domain and GFP (iM Trim33−/−+flag-ΔCC), and treated for 0 or 7 h with LPS. Mean±s.e.m., n=3. (e, left) Relative expression of Ifnb1 mRNA in LPS-activated WT BMDM in presence or absence of Tricostatin A (TSA), added 4 h after LPS stimulation. Mean±s.e.m., n=3. (Right) ChIP-qPCR of CBP/p300 binding at the Ifnb1 promoter in WT and Trim33−/− BMDM treated with LPS. Mean±s.e.m., n=4. (f) Western blot of CBP in WT and Trim33−/− BMDM treated with LPS. β-actin is shown as control. n=4 independent experiments. (g) Kinetics of Ifnb1 mRNA levels in LPS-activated WT and Trim33−/− BMDM, in the presence (dotted lines) or absence (continuous lines) of C646. C646 was added 2 h after LPS stimulation and Ifnb1 mRNA levels were determined. Mean±s.e.m., n=8. (h) ChIP-qPCR of H3ac at the Ifnb1 promoter in LPS-activated WT and Trim33−/− BMDM, in the presence (hatched columns) or absence (solid columns) of C646. Data are presented relative to H3ac binding in untreated WT BMDM. Mean±s.e.m., n=2. (i) Schematic representation depicting the role of TRIM33 in the macrophage-restricted Ifnb1 gene transcription shut down. *P<0.05 and **P<0.01, Mann–Whitney test.
We then determined the effect of TRIM33 deficiency on Ifnb1 transcription and chromatin structure during LPS activation of BMDM. On the Ifnb1 TSS, low levels of paused Pol II were observed before LPS stimulation in both WT and Trim33−/− cells (Fig. 4b, upper left panel). In WT BMDM, Pol II occupancy at TSS and in Ifnb1 3′ region increased after 1 h of stimulation and then was lost at 4 h (Fig. 4b, upper panels). In Trim33−/− BMDM, Pol II occupancy similarly increased after 1 h of stimulation but its levels were maintained up to 24 h (Fig. 4b, upper panels). H3K4me3 levels were transiently increased in WT BMDM after 4 h of LPS stimulation, whereas in Trim33−/− BMDM, persistent and high levels of H3K4me3 were also observed up to 24 h (Fig. 4b, lower panel and right panel). These observations suggest that the paused Pol II seen in the absence of LPS treatment is used for rapid Ifnb1 transcription immediately following LPS stimulation in WT and Trim33−/− BMDM. Then, in WT BMDM, Pol II drops off and Ifnb1 transcription ceases, whereas in Trim33−/− BMDM, continued Pol II occupancy and H3K4me3 promote sustained Ifnb1 transcription.
Prior to LPS activation, histone H3ac on the Ifnb1 promoter was similar in WT and Trim33−/− BMDM (Fig. 4c). This H3ac increased at 1 h after LPS activation of WT and Trim33−/− BMDM. It then returned to basal levels in WT BMDM, whereas a 5- to 8-fold increase in H3ac was observed after 10 and 24 h in Trim33−/− BMDM (Fig. 4c). H3ac on the Ifnb1 promoter in LPS-activated Trim33−/− iM mimicked that observed in Trim33−/− BMDM (Fig. 4d). Expression of ectopic full-length flag-TRIM33 in Trim33−/− iM led to decreased H3ac on the Ifnb1 promoter 7 h after LPS stimulation (Fig. 4d), whereas the flag-ΔCC that could not rescue normal Ifnb1 expression in Trim33−/− iM (Fig. 1h) did not decrease H3ac on the Ifnb1 promoter 7 h after LPS stimulation (Fig. 4d). These results indicate a link between TRIM33 and the regulation of H3ac on the Ifnb1 promoter.
Increased H3ac on the Ifnb1 promoter in LPS-activated Trim33−/− BMDM indicated a possible role for TRIM33 in regulating the equilibrium between recruitment and/or activity of HAT and histone deacetylases (HDAC). Treatment of LPS-activated WT BMDM with trichostatine A, an inhibitor of HDACs, did not result in sustained Ifnb1 expression, whereas it did alter expression of genes regulated by HDACs (Fig. 4e, left panel and Supplementary Fig. 4a). Conversely, the increased H3ac was found to be associated with CBP/p300 HAT binding on the Ifnb1 promoter 10 and 24 h after LPS activation in Trim33−/− BMDM (Fig. 4e, right panel). As total CBP protein levels were similar in WT and Trim33−/− BMDM (Fig. 4f), these results indicated that TRIM33 deficiency promotes CBP recruitment. To determine the importance of CBP/p300 activity in the phenotype of Trim33−/− BMDM, we used C646, a small-molecule inhibitor that efficiently competes with CBP/p300 substrates25. Adding C646 2 h after LPS induction did not modify Ifnb1 expression in WT BMDM, but reversed the high sustained Ifnb1 transcription level in Trim33−/− BMDMs at both 10 and 24 h (Fig. 4g). This reversion was associated with decreased levels of H3ac (Fig. 4h) and reduced Pol II occupancy within the Ifnb1 promoter region in Trim33−/− BMDM (Supplementary Fig. 4b).
Altogether, these results show that enhanced CBP/p300 recruitment and activity at late times of activation is required for sustained Ifnb1 expression in Trim33−/− BMDM and that TRIM33 may regulate Ifnb1 expression at late times by preventing the recruitment of CBP/p300.
Discussion
Although IFN-α and IFN-β share the same receptor, IFN-β seems to be the major immune-suppressive IFN26 and macrophage-restricted synthesis of IFN-β can contribute to protective or pathological immune responses in the lung27. Surprisingly, despite the importance of IFN-β production by macrophages, no cis- or trans-acting factor that might confer a myeloid specificity to Ifnb1 gene transcription has been characterized. The data presented in this study provide evidence of a previously unappreciated regulation of Ifnb1 transcription in macrophages through a macrophage-restricted TRIM33 recruitment on a regulatory sequence, designated as ICE, located 15 kb upstream from the Ifnb1 TSS.
ICE has an open-chromatin structure highly conserved during myeloid differentiation and characterized by Pol II occupancy and tri-methylation of H3K4, two features that are not typical marks of an enhancer or a repressor. In addition to a PU.1-binding site, ICE contains DNA-binding sites for transcriptional regulators such as IRF3, c-jun or p65 that are also part of the enhanceosome. ICE has been recently shown to interact with the Ifnb1 promoter in human fibroblasts in the absence of any viral infection22. Here, we show that ICE can also interact with the Ifnb1 promoter in non-activated macrophages. These combined findings indicate that ICE/Ifnb1 loop is cell-type independent, stable and independent of viral or bacterial stimulation. Such enhancer or repressor loop stability has been described during development where they are associated with paused polymerase28 and in human fibroblasts where TNF-α-responsive enhancers contact their target promoters before TNF-α activation29.
Transient transfection in fibroblast cell lines shows that ICE acts as an enhancer of Ifnb1 promoter activity, but only after viral infection22. In contrast, in macrophages, ICE acts as a repressor of Ifnb1 promoter activity during LPS activation. Furthermore, the high and sustained expression of Ifnb1 in LPS- or poly(I:C)-activated RAW cells where both alleles of ICE are deleted using lentiviral delivery of Cas9 and gRNA indicates that ICE has a repressor activity when these cells are stimulated. Although the lentiviral system used might increase the frequency of off-target effects30,31, the similar results obtained with three different clones and with different gRNAs make off-target effects less likely. Use of recombinant Cas9 protein32 or highly efficient transient transfection of RAW cells will reduce off-target effects and definitively show that ICE is a unique element that acts as an enhancer in fibroblasts, but displays promoter marks and acts as a repressor at late stages of LPS activation of macrophages. These opposite roles of ICE in fibroblasts and macrophages might be linked to recruitment of distinct proteins in the two cell types. Indeed, PU.1, which is not expressed in fibroblasts, is bound to ICE in macrophages and might be involved in the cell-specific function of ICE. Ectopic PU.1 expression in TRIM33+ and TRIM33− NIH3T3 cells indicates that TRIM33-mediated repression of Ifnb1 is dependent on PU.1 expression. However, in fibroblasts activated with poly(I:C), PU.1 expression associated with TRIM33 deficiency does not result in the sustained Ifnb1 expression observed in poly(I:C)-activated Trim33−/− BMDM. This result suggests that factors present in BMDM, but not in fibroblasts, are required for Ifnb1 repression during the late stages of BMDM activation. Amongst haematopoietic-restricted PU.1 partners, IRF8 and IRF3 that regulate Ifnb1 activation in BMDM6,33 are good candidates to cooperate with TRIM33 for Ifnb1 repression during the late stages of BMDM activation.
Several TRIM proteins regulate production and action of IFN-β during innate immune response. TRIM6 is a positive regulator of IFN-β and is critical for IFN-β-mediated antiviral response34, TRIM25 is essential for RIG-I-mediated-type I IFN induction35, TRIM21 regulates IRF3-dependent IFN-β expression36 and TRIM56 modulates STING and subsequent IFN-β induction37. Here, we show the implication of TRIM33 in the transcriptional regulation of Ifnb1. Interestingly, TRIM33 deficiency did not alter the initial activation of Ifnb1 transcription after LPS activation of macrophages but impaired repression of Ifnb1 expression during the late stages of this activation. Together with the continuous binding of TRIM33 on ICE, the complementation of the Ifnb1 phenotype in macrophages by re-expression of TRIM33 in Trim33−/− immortalized macrophages suggested a direct role of TRIM33 in Ifnb1 transcription and indicated that the activity and/or substrate availability of TRIM33 regulated Ifnb1 expression at the end of LPS activation of macrophages.
Two lines of evidence suggest that, during the late stages of LPS activation of BMDM, TRIM33 represses Ifnb1 expression through modulation of CBP/p300 recruitment and/or activity at the Ifnb1 promoter. First, TRIM33 inactivation led to recruitment of CBP/p300 at the Ifnb1 promoter and enhanced H3ac. Second, CBP/p300 activity is critical for enhanced Ifnb1 transcription at late stages as chemical inhibition of CBP/p300 enzymatic activity could restore the regulated expression of Ifnb1 gene in Trim33−/− macrophages. As CBP/p300 acts as a switch to initially turn on Ifnb1 gene transcription38, these results indicated that TRIM33 might turn off Ifnb1 transcription during the late stages of LPS activation of macrophages by impairing the CBP/p300 recruitment and activity at the Ifnb1 promoter. This TRIM33-dependent recruitment and activity of CBP/p300 could be direct and/or mediated by trans-acting factors such as p65 that are, in Trim33−/− BMDM, loaded onto the enhanceosome during the late phases of LPS activation. However, as PU.1 can interact with CBP/p300 (refs 39, 40) and TRIM33, we propose a model where CBP/p300 and TRIM33 compete for PU.1 binding during LPS activation of BMDM. During the initial stages of LPS activation, CBP/p300 recruited by the enhanceosome might activate Ifnb1 transcription, whereas during the late stages of LPS activation, TRIM33 might interact with PU.1, impair PU.1/CBP/p300 interaction and shut down Ifnb1 transcription (Fig. 4i).
Numerous studies have shown the complexity of Ifnb1 gene transcriptional regulation. Our data describe a new regulatory layer through identification of cis- and trans-acting elements that regulate Ifnb1 gene transcription at specific steps of macrophage activation, but not in fibroblasts. These findings not only provide important insights into the cell-specific regulation of Ifnb1 gene transcription but also extend the role of TRIM proteins as critical regulators of the innate immune response.
Methods
Mice
To generate deletion of Trim33 in mature myeloid cells, Trim33fl/fl C57Bl/6-CD45.2 mice15 were crossed with Lysozyme-Cre C57Bl/6-CD45.2 mice (strain name: B6.129P2-Lyz2tm1(cre)Ifo/J, The Jackson Laboratory). Male and female mice of 8–12 months of age were used. Experiments were performed in compliance with European legislation and with the Ethics Committee of the French Ministry of Agriculture (Agreement B9203202).
Cell culture
For BMDM generation, mouse bone marrow (BM) was flushed out of the tibiae, femora and humeri using a syringe with PBS, filtered through a 70-μm mesh filter (Becton Dickinson) to remove debris and pelleted by centrifugation. Adherent cells were removed by incubating BM cells on culture dishes for 3 h in IMDM supplemented with 10% foetal calf serum (FCS), 1% penicillin/streptomycin (PS; Life Technologies) and 10 μM thioglycerol (Sigma Aldrich), then non-adherent cells were cultured for 7 days on culture dishes in the same medium supplemented with 25 ng ml−1 mouse CSF1 (Miltenyi Biotec). On day 7, BMDM were activated with LPS or poly(I:C) in IMDM supplemented with 2.5% FCS and 1% PS. For PMs, cells were collected with cold PBS and cultured for 2 h in RPMI 1640 medium (Life Technologies), supplemented with 10% FCS. All adherent cells expressed F4/80 and were considered as PMs. PM were activated with LPS or poly(I:C) in RPMI 1640 medium supplemented with 2.5% FCS and 1% PS. WT and Trim33−/− iM were generated as previously described21. Briefly, BM cells isolated from WT and Trim33−/− mice were cultured in IMDM supplemented with 10% FCS and 50 ng ml−1 mouse CSF1 (Peprotech). After overnight culture, the cells were transduced with a lentiviral vector expressing the SV40 large T antigen (SFFV.Tag LV). Cells were then cultured in a chemically defined medium (SFM-macrophage medium, Life Technologies) supplemented with CSF1 (50 ng ml−1). Single clones were isolated that expressed F4/80, CD11b but not Gr1. The resulting clones, termed iM, were maintained in IMDM supplemented with 20% foetal bovine serum, 1% PS and 50 ng ml−1 CSF1. iM were activated with LPS in IMDM supplemented with 2.5% FCS and 1% PS. For cell lines, RAW 264.7 and NIH3T3 cells were grown in DMEM (Life Technologies) supplemented with 10% FCS and 1% PS. RAW 264.7 cells were activated with LPS in DMEM supplemented with 1% FCS. Unless otherwise indicated, LPS (Sigma Aldrich) was used at 100 ng ml−1, poly(I:C) at 30 μg ml−1, TSA (Sigma Aldrich) at 10 nM, and C646 (Merck Millipore) at 10 μM. NIH3T3 were transfected with 10 μg ml−1 poly(I:C) (Sigma Aldrich).
Chromatin Immunoprecipitation (ChIP)
Cells were fixed with 1% formaldehyde for 10 min at 37 °C, lysed in SDS lysis buffer (50 mM Tris pH8, 10 mM EDTA, 1% SDS, protease inhibitor cocktail (Roche)) and sonicated. Supernatant was diluted 10 times in IP dilution buffer (16.7 mM Tris pH8, 167 mM NaCl, 1.2 mM EDTA, 1.1% TritonX-100, 0.01% SDS, protease inhibitor cocktail (Roche)) and immunoprecipitations were carried out overnight with specific antibodies. Immunoprecipitated chromatin was collected using Protein A Agarose/Salmon Sperm DNA beads (Millipore) and, after washing and elution, reverse cross-linking was carried out with 0.2 M NaCl at 65 °C overnight. The chromatin was then digested by 20 μg of Proteinase K (Invitrogen) for 1 h at 45 °C and isolated by phenol–chloroform extraction. PCR reactions were performed using SYBR Green PCR Master Mix (Applied Biosystems) and specific primers sequences are listed below.
For sequencing, 10 ng of purified DNA from ChIP was adapter-ligated, PCR amplified and sequenced on the Illumina Genome Analyzer IIx as single-end 50 base reads following Illumina's instructions. Sequence reads were mapped to reference genome mm9/NCBI37 using Bowtie v0.12.7.
3C-seq
3C-seq was performed as previously described41, using EcoRI as first cutter and DpnII as second cutter. Briefly, cells were fixed with 2% formaldehyde for 10 min at room temperature, and nuclei were prepared by lysing cells in 10 mM Tris pH8, 10 mM NaCl, 0.2% NP-40. Nuclei were lyzed in 0.5 ml 1 × EcoRI buffer supplemented with 0.3% SDS for 1 h at 37 °C. Triton X-100 was added at 2% final for another 1 h at 37 °C. DNA was then digested with 400 units of EcoRI overnight at 37 °C. Samples were incubated for 25 min at 65 °C with 1.6% SDS, diluted to 7 ml with ligase buffer, supplemented with 1% Triton X-100 and incubated at 37 °C for 1 h. Ligation was performed for 4 h at 16 °C with 100 u T4 DNA ligase, then 300 μg proteinase K were added and samples were decrosslinked overnight at 65 °C. RNase (300 μg) was added for 45 min at 37 °C, and samples were extracted with phenol/chloroform/isoamyl alcohol (25:24:1; PCI). After precipitation, samples were digested with DpnII, extracted with PCI and precipitated, and ligated in 15 ml ligation buffer at 16 °C for 4 h before another PCI extraction and precipitation. Samples were amplified by PCR with the primers listed below, and PCR products were purified on QIAprep columns (Qiagen). The resulting libraries were sequenced on a Illumina HiSeq 2000 as single-end 100 bp reads. After trimming, reads were aligned on the mm9 version of the mouse genome using Bowtie. Results are presented as the number of reads per million reads per EcoRI fragment, in the middle of each fragment.
Assay for IFN-β
WT and Trim33−/− BMDM supernatants were collected after LPS stimulation. ELISA was performed with LegendMax Mouse IFN-β kit (Biolegend), according to manufacturer's instructions.
Quantitative RT-PCR
Total RNA was extracted with the RNeasy Plus Micro kit (Qiagen) and reverse transcribed with random primers and Superscript III (Life Technologies). Quantitative PCR was performed using the Power SYBR green PCR master mix (Applied Biosystems) in the 7900HT Fast Real-Time or the StepOne PCR Systems (Applied Biosystems). Primer sequences are listed below.
Lentiviral vectors for rescue and knockdown
For rescue experiments in Trim33−/− iM, the lentiviral vector was constructed by inserting full-length flag-TRIM33 from the pSG5-flag-TRIM33 (ref. 15) into the pTRIP/ΔU3-MND-IRES-GFP vector, downstream the MND promoter, using the Gateway technology (Life Technologies). Deletions of the coiled-coil domain (aa 321–457), the ubiquitin ligase activity (aa 141–145) and the SMAD-interaction (aa 555–876) domain were generated by PCR. The pTRIP/ΔU3-MND-GFP was used in control experiments. NIH3T3 PU1+ cells were obtained by transducing a lentiviral vector pTRIP/ΔU3-MND-PU.1-IRES-GFP. For TRIM33 knocking-down in NIH3T3 or NIH3T3 PU.1+, the sequence of the shRNA targeting mouse Trim33 was obtained from Sigma Aldrich (5′- CCGGCGGACTTAAATCGGTTGTTAACTCGAGTTAACAACCGATTTAAGTCCGTTTTTG -3′) and cloned in the pSuper vector (OligoEngine) downstream the H1 promoter. H1-shRNATRIM33 fragment was then cloned in the pTRIP/ΔU3-EF1α-GFP, where the GFP coding sequence is under the control of the EF1α promoter. All the constructs were verified by sequencing. Lentiviral vector production and cells transduction were carried out as described42. GFP-positive cells were sorted and used in functional assays.
Luciferase assays
A 391 bp DNA fragment of murine Ifnb1 promoter was obtained by PCR on genomic DNA and cloned into the pGL3 Basic vector (Promega) using KpnI and XhoI sites. A 408 bp DNA fragment spanning the −15 kb PU.1/TRIM33-binding site was obtained by PCR of genomic DNA and cloned upstream of the 391 bp DNA fragment of murine Ifnb1 promoter into the pGL3 Basic vector. After sequencing of the different constructs, RAW 264.7 cells were transfected using JetPEI (PolyPlus-transfection) with the indicated constructs and the TK-renilla reporter vector (Promega), as an internal control for normalization. Twenty-four hours after transfection, cells were activated with LPS (100 ng ml−1) and luciferase activity was determined using the dual-luciferase reporter assay (Promega). Firefly luciferase activity was normalized with respect to renilla luciferase activity.
Genomic deletions of ICE with the CRISPR/CAS9 system
Two guides RNA (gRNA) were designed to delete a 1,361 bp fragment containing the −15 kb region using the CRISPR Design Tool (http://tools.genome-engineering.org), to minimize the number of off-target sites. Sequences for gRNA 5A2 and 3A are 5′- TTCTCGTTCATTGTTAGCGA -3′ and 5′- CTCTAGTTTAGACGTTTAAC -3′, respectively. Singles gRNA were then cloned into the LentiCRISPRv2 (Addgene) vector, which contains the human CAS9-coding sequence, using BsmBI cloning sites. Lentivirus were produced and co-transduced into RAW 264.7 and NIH3T3. After puromycin selection (3 μg ml−1, Sigma Aldrich), cells were clonally isolated by fluorescence-activated cell sorting and individual clones were screened for deletion by PCR using the following primers: F5-1:5′- TGACGACAAATGTGGTACTGG -3′; F3-1:5′- CAGGCGAAAGGGAAACTAAA -3′; and R3-1:5′- TAGGAGTGGCAGATGGGAAG -3′.
Deletions of 100 or 176 bp were carried out using one gRNA guide (5′- AAACACGTTAGTTGTCAGAC -3′) flanking the PU.1/TRIM33 site at the ICE and cloned into the LentiCRISPRv2 vector as above. After puromycin selection and clonal isolation, WT and ICE−/− RAW 264.7 cells were identified by PCR using the following primers: F: 5′- AGCAAAGCCGAAGAAGACAC -3′ and R: 5′- GGAAGAGGACGAGAGAACCA -3′.
All the clones were verified by sequencing of a 2 kb genomic region containing the ICE.
Protein extraction and western blot
Total proteins from BMDM or iM were extracted using lysis buffer (50 mM Tris pH8, 300 mM NaCl, 1% Triton X-100, 0.1% sodium dodecyl sulfate, 1 mM EDTA, phosphatase inhibitor cocktail-1 and -2 (Sigma Aldrich) and complete protease inhibitor cocktail (Roche)). Proteins were separated on 3–8% Tris-Acetate gel or 4–12% Bis Tris gel (Life Technologies), transferred on a nitrocellulose membrane (Amersham) and blotted with the indicated antibodies. Following staining with HRP-coupled secondary antibodies, the proteins were detected with ECL Prime by chemiluminescence (Amersham).
Antibodies
The following antibodies were used in western blot and ChIP: TRIM33 (3E9, Euromedex for WB (1:1,000); A301-059A, Bethyl for ChIP (1:100)), c-JUN (sc-1694X, Santa Cruz (1:200)), p65 (sc-372X, Santa Cruz (1:200)), RNA Pol II (sc-899X, Santa Cruz (1:200)), H3K4me3 (07-473, Millipore (1:500)), PU.1 (sc-352X, Santa Cruz (1:10,000 for western blot; 1:500 for ChIP)), p300 (sc-585X, Santa Cruz), AcH3 (06-599, Millipore (1:500)), H3K4me1 (ab-8895, Abcam (1:500)), Flag (M2 monoclonal, Sigma Aldrich (1:1,000)), CBP (A-22, sc-369, Santa Cruz (1:1,000)), β-actin (A5441, Sigma Aldrich (1:10,000)).
Primer sequences
The following primer sequences were used in quantitative RT-PCR, ChIP and 3C-seq: Ifnb1 F: 5′- CTGGCTTCCATCATGAACAA -3′; Ifnb1 R: 5′- AGAGGGCTGTGGTGGAGAA -3′; Ifna F: 5′- GGATGTGACCTTCCTCAGACTC -3′; Ifna R: 5′- ACCTTCTCCTGCGGGAATCCAA -3′; Hprt F: 5′- GCTGGTGAAAAGGACCTCT -3′; Hprt R: 5′- CACAGGACTAGAACACCTGC -3′; Ifnb1 promoter F: 5′- CCACCTGTTGTTCATGATGG -3′; Ifnb1 promoter R: 5′- CATTCTCACTGCAGCCTTTG -3′; Ifnb1 ICE F: 5′- CCAAGGGTTGCGTAATGAAC -3′; Ifnb1 ICE R: 5′- CCCGATCTTCAAATCCAGTC -3′; Ifnb1 +2,5 kb F: 5′- TAGCTTCCATGCCCAGTTTC -3′; Ifnb1 +2,5 kb R: 5′- CCAAAAGTATGCCCATCACC -3′; β-globin major promoter F: 5′- GAAGCCTGATTCCGTAGAGC -3′; β-globin major promoter R: 5′- CAACTGATCCTACCTCACCTTATATGC -3′; ICE EcoRI primer: 5′- AATGATACGGCGACCACCGAACACTCTTTCCCTACACGACGCTCTTCCGATCT -3′ followed by a 4nt tag and 5′- GATGGCACACCTTGAATTC -3′; ICE DpnII primer: 5′- CAAGCAGAAGACGGCATACGAGACGACAAATGTGGTACTG -3′; Ifnb1 EcoRI primer: 5′- AATGATACGGCGACCACCGAACACTCTTTCCCTACACGACGCTCTTCCGATCT -3′ followed by a 4nt tag and 5′- GGGTGTTCTATTTTATCAATAC -3′; Ifnb1 DpnII primer: 5′- CAAGCAGAAGACGGCATACGATTTCTAGAGTTTCCGACTCTG -3′.
Additional information
Accession codes: High throughput sequencing data have been deposited in the Gene Expression Omnibus (GEO) database (www.ncbi.nlm.nih.gov/geo), under accession codes GSE73322 (3c-seq) and GSE43654 (ChIP-seq).
How to cite this article: Ferri, F. et al. TRIM33 switches off Ifnb1 gene transcription during the late phase of macrophage activation. Nat. Commun. 6:8900 doi: 10.1038/ncomms9900 (2015).
Supplementary Material
Supplementary Figures 1-4 and Supplementary References
Acknowledgments
We are grateful to the staff of the iRCM animal facility for excellent support during the mouse studies and to N. Dechamps and J. Baijer of the iRCM cytometry platform for excellent support during FACS experiments. F.F. and A.P. are supported by fellowships from Dim Stem Pole and La Ligue contre le Cancer. This project was supported by grants to P.-H.R. from La Ligue contre le Cancer (équipe labelisée), ANR (ANR-12-BSV1-0001-02), INCA (PL BIO 013031), Inserm, CEA/DSV, Paris-Diderot and Paris-Sud.
Footnotes
Author contributions P.-H.R., F.F., I.D. and G.R. designed the project, analysed data and wrote the manuscript. A.P., V.P., A.-S.G., F.F., G.R., M.D. and D.L. organized, performed and analysed most experiments and wrote the manuscript. E.S., A.v.d.H. and P.K. assisted with experimental design and data analysis for 3C-Seq and wrote the manuscript. M.L.S. and M.D.P. assisted with experimental design for iM production and wrote the manuscript. All authors approved the final version of the manuscript.
References
- Trinchieri G. Type I interferon: friend or foe? J. Exp. Med. 207, 2053–2063 (2010) . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stetson D. B. & Medzhitov R. Type I interferons in host defense. Immunity 25, 373–381 (2006) . [DOI] [PubMed] [Google Scholar]
- Teijaro J. R. et al. Persistent LCMV infection is controlled by blockade of type I interferon signaling. Science 340, 207–211 (2013) . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilson E. B. et al. Blockade of chronic type I interferon signaling to control persistent LCMV infection. Science 340, 202–207 (2013) . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ford E. & Thanos D. The transcriptional code of human IFN-beta gene expression. Biochim. Biophys. Acta. 1799, 328–336 (2010) . [DOI] [PubMed] [Google Scholar]
- Li P. et al. IRF8 and IRF3 cooperatively regulate rapid interferon-beta induction in human blood monocytes. Blood 117, 2847–2854 (2011) . [DOI] [PubMed] [Google Scholar]
- Kim H. & Seed B. The transcription factor MafB antagonizes antiviral responses by blocking recruitment of coactivators to the transcription factor IRF3. Nat. Immunol. 11, 743–750 (2010) . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Siednienko J. et al. Nuclear factor kappaB subunits RelB and cRel negatively regulate Toll-like receptor 3-mediated beta-interferon production via induction of transcriptional repressor protein YY1. J. Biol. Chem. 286, 44750–44763 (2011) . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Josse T. et al. Association of the interferon-beta gene with pericentromeric heterochromatin is dynamically regulated during virus infection through a YY1-dependent mechanism. Nucleic. Acids. Res. 40, 4396–4411 (2012) . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weill L., Shestakova E. & Bonnefoy E. Transcription factor YY1 binds to the murine beta interferon promoter and regulates its transcriptional capacity with a dual activator/repressor role. J. Virol. 77, 2903–2914 (2003) . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hatakeyama S. TRIM proteins and cancer. Nat. Rev. Cancer 11, 792–804 (2011) . [DOI] [PubMed] [Google Scholar]
- Herquel B., Ouararhni K. & Davidson I. The TIF1alpha-related TRIM cofactors couple chromatin modifications to transcriptional regulation, signaling and tumor suppression. Transcription 2, 231–236 (2011) . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herquel B. et al. Trim24-repressed VL30 retrotransposons regulate gene expression by producing noncoding RNA. Nat. Struct. Mol. Biol. 20, 339–346 (2013) . [DOI] [PubMed] [Google Scholar]
- Xi Q. et al. A poised chromatin platform for TGF-beta access to master regulators. Cell 147, 1511–1524 (2011) . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kusy S. et al. Adult hematopoiesis is regulated by TIF1gamma, a repressor of TAL1 and PU.1 transcriptional activity. Cell Stem Cell 8, 412–425 (2011) . [DOI] [PubMed] [Google Scholar]
- Bai X. et al. TIF1gamma controls erythroid cell fate by regulating transcription elongation. Cell 142, 133–143 (2010) . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ozato K., Shin D. M., Chang T. H. & Morse H. C. 3rd TRIM family proteins and their emerging roles in innate immunity. Nat. Rev. Immunol. 8, 849–860 (2008) . [DOI] [PMC free article] [PubMed] [Google Scholar]
- McNab F. W., Rajsbaum R., Stoye J. P. & O'Garra A. Tripartite-motif proteins and innate immune regulation. Curr. Opin. Immunol. 23, 46–56 (2011) . [DOI] [PubMed] [Google Scholar]
- Tisserand J. et al. Tripartite motif 24 (Trim24/Tif1 alpha) tumor suppressor protein is a novel negative regulator of interferon (IFN)/signal transducers and activators of transcription (STAT) signaling pathway acting through retinoic acid receptor alpha (Rar alpha) inhibition. J. Biol. Chem. 286, 33369–33379 (2011) . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weng L. et al. The E3 ubiquitin ligase tripartite motif 33 is essential for cytosolic RNA-induced NLRP3 inflammasome activation. J. Immunol. 193, 3676–3682 (2014) . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Squadrito M. L. et al. Endogenous RNAs modulate microRNA sorting to exosomes and transfer to acceptor cells. Cell Rep. 8, 1432–1446 (2014) . [DOI] [PubMed] [Google Scholar]
- Banerjee A. R., Kim Y. J. & Kim T. H. A novel virus-inducible enhancer of the interferon-beta gene with tightly linked promoter and enhancer activities. Nucleic Acids Res. 42, 12537–12554 (2014) . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Uhlenhaut N. H. et al. Insights into negative regulation by the glucocorticoid receptor from genome-wide profiling of inflammatory cistromes. Mol. Cell 49, 158–171 (2013) . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lara-Astiaso D. et al. Immunogenetics. Chromatin state dynamics during blood formation. Science 345, 943–949 (2014) . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bowers E. M. et al. Virtual ligand screening of the p300/CBP histone acetyltransferase: identification of a selective small molecule inhibitor. Chem. Biol. 17, 471–482 (2010) . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ng C. T. et al. Blockade of interferon Beta, but not interferon alpha, signaling controls persistent viral infection. Cell Host Microbe 17, 653–661 (2015) . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hogner K. et al. Macrophage-expressed IFN-beta contributes to apoptotic alveolar epithelial cell injury in severe influenza virus pneumonia. PLoS Pathog. 9, e1003188 (2013) . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ghavi-Helm Y. et al. Enhancer loops appear stable during development and are associated with paused polymerase. Nature 512, 96–100 (2014) . [DOI] [PubMed] [Google Scholar]
- Jin F. et al. A high-resolution map of the three-dimensional chromatin interactome in human cells. Nature 503, 290–294 (2013) . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pattanayak V., Guilinger J. P. & Liu D. R. Determining the specificities of TALENs, Cas9, and other genome-editing enzymes. Methods Enzymol. 546, 47–78 (2014) . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang X. et al. Unbiased detection of off-target cleavage by CRISPR-Cas9 and TALENs using integrase-defective lentiviral vectors. Nat. Biotechnol. 33, 175–178 (2015) . [DOI] [PubMed] [Google Scholar]
- Kim S., Kim D., Cho S. W., Kim J. & Kim J. S. Highly efficient RNA-guided genome editing in human cells via delivery of purified Cas9 ribonucleoproteins. Genome Res. 24, 1012–1019 (2014) . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mancino A. et al. A dual cis-regulatory code links IRF8 to constitutive and inducible gene expression in macrophages. Genes Dev. 29, 394–408 (2015) . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rajsbaum R. et al. Unanchored K48-linked polyubiquitin synthesized by the E3-ubiquitin ligase TRIM6 stimulates the interferon-IKKepsilon kinase-mediated antiviral response. Immunity 40, 880–895 (2014) . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Inn K. S. et al. Linear ubiquitin assembly complex negatively regulates RIG-I- and TRIM25-mediated type I interferon induction. Mol. Cell 41, 354–365 (2011) . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Higgs R. et al. The E3 ubiquitin ligase Ro52 negatively regulates IFN-beta production post-pathogen recognition by polyubiquitin-mediated degradation of IRF3. J. Immunol. 181, 1780–1786 (2008) . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsuchida T. et al. The ubiquitin ligase TRIM56 regulates innate immune responses to intracellular double-stranded DNA. Immunity 33, 765–776 (2010) . [DOI] [PubMed] [Google Scholar]
- Merika M., Williams A. J., Chen G., Collins T. & Thanos D. Recruitment of CBP/p300 by the IFN beta enhanceosome is required for synergistic activation of transcription. Mol. Cell 1, 277–287 (1998) . [DOI] [PubMed] [Google Scholar]
- Yamamoto H., Kihara-Negishi F., Yamada T., Hashimoto Y. & Oikawa T. Physical and functional interactions between the transcription factor PU.1 and the coactivator CBP. Oncogene 18, 1495–1501 (1999) . [DOI] [PubMed] [Google Scholar]
- Bai Y., Srinivasan L., Perkins L. & Atchison M. L. Protein acetylation regulates both PU.1 transactivation and Ig kappa 3' enhancer activity. J. Immunol. 175, 5160–5169 (2005) . [DOI] [PubMed] [Google Scholar]
- Stadhouders R. et al. Multiplexed chromosome conformation capture sequencing for rapid genome-scale high-resolution detection of long-range chromatin interactions. Nat. Protoc. 8, 509–524 (2013) . [DOI] [PubMed] [Google Scholar]
- Kusy S. et al. NKX3.1 is a direct TAL1 target gene that mediates proliferation of TAL1-expressing human T cell acute lymphoblastic leukemia. J. Exp. Med. 207, 2141–2156 (2010) . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heinz S. et al. Simple combinations of lineage-determining transcription factors prime cis-regulatory elements required for macrophage and B cell identities. Mol. Cell 38, 576–589 (2010) . [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary Figures 1-4 and Supplementary References