

ABSTRACT
The eukaryotic replicative DNA helicase, Mcm2-7, is loaded in inactive form as a double hexameric complex around double-stranded DNA. To ensure that replication origins fire no more than once per S phase, activation of the Mcm2-7 helicase is temporally separated from Mcm2-7 loading in the cell cycle. This 2-step mechanism requires that inactive Mcm2-7 complexes be maintained for variable periods of time in a topologically bound state on chromatin, which may create a steric obstacle to other DNA transactions. We have recently found in the budding yeast, Saccharomyces cerevisiae, that Mcm2-7 double hexamers can respond to collisions with transcription complexes by sliding along the DNA template. Importantly, Mcm2-7 double hexamers remain functional after displacement along DNA and support replication initiation from sites distal to the origin. These results reveal a novel mechanism to specify eukaryotic replication origin sites and to maintain replication origin competence without the need for Mcm2-7 reloading.
KEYWORDS: chromosomes, DNA replication, Mcm2-7, ORC, replication origin
Introduction
Eukaryotic chromosomes form the template for numerous protein complexes that transcribe, repair, or replicate the genome, or that organize the chromosomal DNA into distinct chromatin domains. The concurrence of these events on the same DNA template can lead to mutual interference with potentially deleterious effects on genome stability. A prominent example is transcription-replication conflict, in which collisions between replication forks and transcription complexes can lead to DNA breaks and genome instability.1 Eukaryotic replication origins may be particularly susceptible to interference due to the prolonged retention of the replicative DNA helicase at the origin prior to origin firing. This derives from the fact that eukaryotes have evolved a 2-step origin activation mechanism that prevents repeated origin firing within 1 cell cycle.2 Central to this re-replication control strategy is the loading cycle of the eukaryotic replicative DNA helicase, Mcm2-7. Unlike bacterial replicative DNA helicases, the Mcm2-7 complex is loaded in inactive form at the origin, and activation of the Mcm2-7 helicase does not occur until a later stage in the cell cycle when origins fire3 (Fig. 1). Specifically, Mcm2-7 loading can only occur at the end of mitosis and during G1 phase, while activation of the Mcm2-7 helicase is restricted to the subsequent S phase. Since replication origin firing genome-wide occurs throughout S phase, with some origins firing early and others late in S phase, individual Mcm2-7 complexes must reside for variable periods of time at the origin.4 This delay in helicase activation may explain why Mcm2-7 complexes, unlike bacterial replicative DNA helicases, are loaded around double-stranded DNA, as such a mechanism avoids the generation and persistence of damage-sensitive single-stranded DNA at replication origins in the absence of ongoing DNA synthesis. On the other hand, this configuration locks Mcm2-7 complexes in a topologically bound state around DNA that could act as a steric block to other forms of DNA metabolism.
Figure 1.
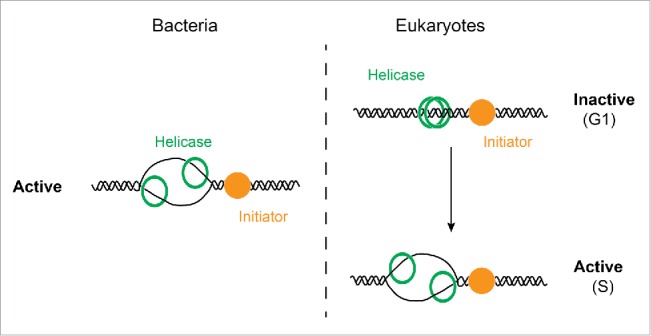
Activity states of the replicative DNA helicase at bacterial and eukaryotic replication origins. In bacteria the replicative DNA helicase is loaded in active form around single-stranded DNA at the origin (left). In eukaryotes, the replicative DNA helicase is loaded in inactive form around double-stranded DNA at the origin; activation of the helicase is temporally separated from helicase loading in the cell cycle (right). The initiator protein(s) direct the loading of the replicative DNA helicase. G1 and S indicate the G1 phase and S phase cell cycle stages in eukaryotes.
Various lines of evidence demonstrate the exceptional stability of origin-bound Mcm2-7 complexes prior to activation. For example, native origin-bound Mcm2-7 complexes functionally resist high salt elution from budding yeast and Xenopus chromatin,5,6 as do Mcm2-7 complexes loaded around DNA in vitro with purified budding yeast proteins,7,8 while fluorescence recovery after photo-bleaching (FRAP) experiments in live hamster and C. elegans cells demonstrate that chromatin-bound Mcm2-7 complexes exchange unusually slowly with the soluble pool of Mcm2-7 in G1 phase.9-11 Contrary to these observations, Mcm2-7 complexes are rapidly lost from chromatin in G1 phase-arrested budding yeast cells upon depletion of the Mcm2-7 loading factors ORC and Cdc6, suggesting that Mcm2-7 complexes need to be constantly reloaded to be maintained on chromosomes under this condition.12-15 The mechanism by which Mcm2-7 complexes are removed from chromosomes in G1-arrested budding yeast cells is not known. Nonetheless, Mcm2-7 complexes in all eukaryotic cells must remain stably chromosome-bound during both normal S phase and checkpoint-induced S phase arrest, as reloading at this stage cannot occur due to re-replication control mechanisms.2,16
These observations raise the question how Mcm2-7 complexes are maintained at origins in vivo, where they face the risk of disruption by other forms of DNA metabolism. To maintain Mcm2-7 complexes at origins is important, because DNA replication has to initiate from a sufficiently high number of origin sites to ensure timely completion of genome duplication before the onset of mitosis, and to reduce the chance of replication fork failure, which increases with the distance replication forks have to travel between adjacent replication origins.17,18
To characterize collisions between origin-bound Mcm2-7 and other protein complexes we have recently studied the effects of transcriptional interference at budding yeast replication origins.19 Surprisingly, we found that Mcm2-7 complexes can slide ahead of elongating RNA polymerases and subsequently initiate replication from sites distal to the loading site at the origin. These observations demonstrate (i) a molecular mechanism for the maintenance of Mcm2-7 complexes on DNA that is not dependent on the reloading of Mcm2-7, and (ii) reveal that eukaryotic replication origins, unlike those of bacteria, are inherently flexible and can be specified even after the replicative DNA helicase has been loaded on the DNA.
Mcm2-7 complexes are loaded as double hexameric rings around DNA
Mcm2-7 proteins assemble into ring-shaped hetero-hexameric complexes with a defined subunit order. As members of the AAA+ family of ATPases they form composite ATP-binding sites at subunit interfaces and utilize the energy of ATP-binding and –hydrolysis to thread DNA through the central channel of the Mcm2-7 ring during DNA unwinding.20 The Mcm2-7 ring thus has to be transiently opened and closed around DNA during Mcm2-7 origin loading. This is achieved by an incompletely understood ATP-dependent remodeling mechanism that involves the coordinated action of a set of loading factors, including the origin recognition complex (ORC), Cdc6, and Cdt1, in addition to being dependent on ATP hydrolysis by Mcm2-7.21 ORC and Cdc6 assemble into a complex at the origin and cooperatively recruit the Mcm2-7 complex, which at this stage is bound by Cdt1. Cdc6 and Cdt1 act transiently during the loading reaction and are released from the origin during the Mcm2-7 loading reaction. Although ORC remains bound at the origin following Mcm2-7 loading, it does not maintain physical contact with Mcm2-7 22,23 and can be eluted from DNA in vitro without disrupting origin activity.6-8, 19,24
Intriguingly, in the course of the reaction 2 individual Mcm2-7 rings are loaded in opposite orientation into a stable, double-hexameric complex around double-stranded DNA. The individual hexamers are held together at the conserved N-terminal domains of their subunits, while the C-terminal AAA+ motor domains, which are proximal to the fork during DNA unwinding, are facing outwards.22,25 A long continuous channel that is wide enough to accommodate double-stranded DNA runs lengthwise through the Mcm2-7 double hexamer (DH), which is consistent with electron microscopic images of DNA-bound Mcm2-7 DHs after tungsten rotary shadow casting that suggest that double-stranded DNA passes longitudinally through the complex.22,26 The head-to-head configuration of the Mcm2-7 DH thus provides a molecular mechanism for the establishment of bidirectional DNA synthesis at eukaryotic origins.27
Mcm2-7 DHs are mobile
Mcm2-7 loading onto DNA has been reconstituted with purified budding yeast proteins,22,26,28 and reconstituted budding yeast Mcm2-7 DHs are functional to support regulated origin firing in vitro.7,8, 29,30 Intriguingly, by comparing the residence times of reconstituted Mcm2-7 DHs on linear versus circular DNA it was found that Mcm2-7 DHs, similar to other proteins encircling DNA,31,32 can slide along and off the ends of DNA.22,26
Mcm2-7 sliding appears to depend on high salt concentrations in the reaction buffer 22; in solutions of lower ionic strength Mcm2-7 DHs remain stably associated with linear DNA fragments,7,19 indicating that Mcm2-7 DHs are resistant to spontaneous sliding at physiological salt concentrations. Mcm2-7 DHs are large, approximately 1.2 MDa, protein complexes of ∼20 nm length, enabling them to encircle ∼60 bp of B-form DNA. Intriguingly, the central channel is not straight, but exhibits a kink at the hexamer-hexamer interface that is caused by an off-register stacking of the 2 Mcm2-7 rings, which may force the DNA to bend at the channel midpoint.22,25 This DNA bending may oppose spontaneous Mcm2-7 DH sliding and in a physiological setting Mcm2-7 DH sliding may thus require coupling to an energy consuming motor, as would occur upon collision with other DNA translocases.
Mcm2-7 DH relocalization after collision with transcription complexes
To determine the fate of Mcm2-7 DHs at replication origins after collision with other DNA translocases we initially studied collisions between transcription complexes and reconstituted budding yeast Mcm2-7 DHs in vitro.19 To model such a collision event in vitro we employed T7 RNA polymerase (RNAP), a monomeric RNAP that affords tight control of the transcription reaction due to its dependence on specific short promoter and terminator sequences to initiate and terminate transcription, respectively. While Mcm2-7 DHs remained stably bound to circular closed DNA molecules upon transcription through the origin site, they were displaced efficiently from linear DNA molecules, demonstrating that T7 RNAP can push Mcm2-7 DHs off the free ends of DNA. Importantly, the DNA templates remained competent for replication in vitro when transcription through the origin occurred after Mcm2-7 loading. However, transcription induced a shift in the positions of Mcm2-7 DHs and initiation sites by up to several kilobase pairs, demonstrating that Mcm2-7 DHs can initiate DNA replication from non-origin sites after displacement by T7 RNAP in vitro.
To investigate the effect of collisions of RNAP with replication origins in budding yeast cells we exploited the fact that the majority of budding yeast origins is located between convergent or tandemly oriented genes.33 Due to the small size of the intergenic regions in budding yeast this results in the majority of the origins being located near the 3′ end of genes. To increase collisions between RNAP II and origins downstream of genes we conditionally delayed transcription termination by RNAP II using a temperature-sensitive variant of the transcription termination factor Rat1.34,35 Using ChIP-seq to map the chromosomal positions of both ORC and Mcm2-7 across the genome we found that ORC binding was not altered at non-permissive temperature in rat1-1 cells, whereas Mcm2-7 positions around origins were shifted by up to 2 kilobase pairs in a direction that correlated with the direction of transcription around the respective origin sites. The fact that ORC positions did not change upon defective termination by RNAP II implies that Mcm2-7 redistribution under these conditions is not a consequence of alternative Mcm2-7 loading, but due to Mcm2-7 complexes being pushed ahead of elongating RNAP II.
We mapped replication initiation sites genome-wide in rat1-1 cells using Okazaki fragment sequencing.36 Unexpectedly, origin activity was dramatically affected across all chromosomes at non-permissive temperature in rat1-1 cells, exhibiting changes in both efficiency and position. Initiation site shifts correlated with the shift in Mcm2-7 distribution and with the prevailing direction of transcription around the origins, demonstrating that RNAP II can push Mcm2-7 DHs for up to 2 kb from an origin through chromatin, and that Mcm2-7 remain competent for initiation from origin distal sites in vivo.
Previous studies in S. cerevisae have demonstrated that transcription through an origin interferes with the stability of autonomously replicating plasmids, as well as with the activity of replication origins at their native chromosomal location.37-40 At first sight these studies seem at odds with our finding that replication origins can shift upon transcriptional interference. However, it has to be noted that plasmid maintenance studies are performed under conditions of chronic transcription stress over many cell generations, which may thus interfere with the initial loading of the Mcm2-7 complex. Gros et al.,19 on the other hand, analyzed the effect of transcriptional interference restricted to 2 h in an asynchronous yeast culture, which allows detection of interference at origins both before and after Mcm2-7 loading. Moreover, previous studies analyzed origin activity exclusively at known origin sites, such that shifts in origin position would have gone unnoticed, whereas Gros et al. 19 employed unbiased genome-wide approaches to monitor origin activity at every chromosomal position.
It should ne noted that the ability of Mcm2-7 DHs to initiate DNA replication after collision with RNAP is not without limits. For example, we observed that origin efficiency was globally reduced in rat1-1 cells at non-permissive temperature.19 Moreover, at several sites we observed repositioning of Mcm2-7 without a corresponding shift in initiation site position, indicating that the Mcm2-7 complexes had lost their replication competence after displacement by RNAP. The reason for this is not known, but in a previous study we had identified DNA templates that supported Mcm2-7 loading, but were defective for DNA replication in vitro, indicating that Mcm2-7 DHs loaded around certain DNA sequences cannot initiate DNA replication.7 Reduced origin efficiency in rat1-1 cells may thus be due to a combination of factors, including reduced initiation efficiency of Mcm2-7 DHs repositioned around non-canonical origin sites and reduced Mcm2-7 loading efficiency at origins that are transcribed prior to Mcm2-7 loading. The ability of Mcm2-7 DHs to slide upon collision with other proteins may therefore be most relevant under normal growth conditions, when interference levels at origins are low. On the other hand, the fact that Okazaki fragments mapped to both strands of the chromosomal DNA over wide regions of the genome in rat1-1 cells at non-permissive temperature may also indicate that initiation sites were simply more widely dispersed than can be detected by the population average-based methods of ChIP-seq and Okazaki fragment sequencing.
DNA sequence-independent Mcm2-7 activation
The observation that DNA replication can initiate from non-origin sites in budding yeast is surprising given that replication origin function in budding yeast, in striking contrast to other eukaryotes,17 depends on specific DNA sequences.41-44 However, specific origin sequences seem to be important for Mcm2-7 loading, rather than Mcm2-7 activation, in budding yeast. The only conserved sequence element at budding yeast origins, the ARS consensus sequence (ACS), forms part of the ORC binding site,45 while additional origin sequences adjacent to the ACS, known as B elements, though important for full origin activity in vivo,42 are not conserved. The DNA across B elements exhibits a significant nucleotide skew, suggesting that the structure of DNA at B elements, rather than specific DNA sequences, may be important for origin activity, possibly to establish a nucleosome-free region around chromosomal origins that is permissive for Mcm2-7 loading on chromatin.46,47 Naked DNA should, therefore, bypass the requirement for generation of a nucleosome-free region at the origin and permit Mcm2-7 loading at sites that might otherwise not be accessible within chromatin, which is supported by in vitro replication studies.7,30 The lack of conserved sequences outside of the ACS thus implies that budding yeast Mcm2-7 are loaded around non-conserved DNA sequences at the origin, which is consistent with specific DNA sequences not being mechanistically required for Mcm2-7 DH activation throughout eukaryotes.
Two levels of eukaryotic replication origin flexibility
Surprisingly, while budding yeast chromosomes are normally replicated from specific origin sites, deletion of these sites does not disrupt the replication competence of derivatives of chromosome III and VI in vivo.48,49 Although a potential involvement of alternative, e.g. recombination-based, replication mechanisms in the maintenance of origin-depleted chromosome derivatives has not been directly investigated, the stability of origin-depleted chromosome III derivatives is severely impaired in orc2-1 mutant cells,49 indicating that DNA replication initiates by the canonical mechanism from ectopic sites in this setting. This finding is consistent with observations made with purified budding yeast proteins in vitro, in which ORC was found to normally direct Mcm2-7 loading at a canonical origin site present in the DNA template, but would direct Mcm2-7 loading at lower affinity non-canonical sites if the canonical origin site was mutated.7 Targeting ORC to specific high affinity sites in vivo may increase the efficiency of Mcm2-7 loading in the budding yeast genome, where intergenic space is sparse, while the ability to direct replication initiation from non-canonical sites may increase the robustness of the replication competence of chromosomes when canonical sites get disrupted. Budding yeast replication origins are thus intrinsically flexible not only due to Mcm2-7 DH mobility, but also due to the flexibility of ORC to direct the loading of Mcm2-7 at non-canonical sites (Fig. 2).
Figure 2.
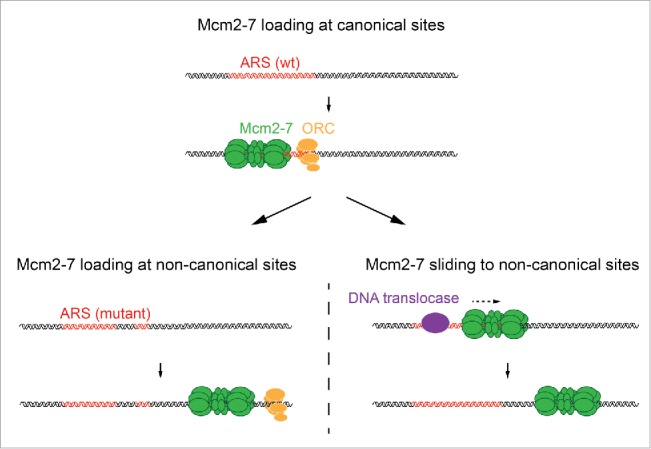
Two levels of budding yeast origin flexibility. Mcm2-7 complexes are preferentially loaded at canonical origin sites that contain a high-affinity ORC binding site (top). Mutation of the high-affinity ORC binding site at a canonical origin can redirect ORC to lower-affinity binding sites in the template, resulting in Mcm2-7 loading at non-canonical origin sites (bottom left). Collisions between Mcm2-7 DHs and other DNA translocases at replication origins can induce the sliding of Mcm2-7 to non-canonical origin sites (bottom right)/ ARS: Autonomously replicating sequence.
Origin specification by transcription
Transcription has long been recognized to influence the distribution of replication origins along eukaryotic chromosomes. For example, DNA replication initiates promiscuously along the chromosomes of higher eukaryotes until the onset of transcription at the mid-blastula transition, when initiation sites are restricted to non-transcribed intergenic regions.50 Another well-studied example is the initiation zone located between the convergent dihydrofolate reductase (DHFR) and 2BE2121 genes in CHO cells, the boundaries of which appear to be similarly circumscribed by ongoing transcription.51,52 The incompatibility of transcription and replication initiation thus explains the predominantly intergenic positioning of replication origins from yeast to human.17,53 It is commonly presumed that transcription inhibits the licensing of replication origins by preventing the loading of Mcm2-7 complexes. Our data suggest that transcription complexes may, in addition, clear transcription units from replication origins by redistributing Mcm2-7 DHs from their sites of loading to regions downstream. This model is further supported by the recent observation that Mcm2-7 complexes, but not ORC, are redistributed along chromosomes in HU arrested Drosophila cells in a manner that correlates with the local transcription landscape.54 Because transcription termination occurs stochastically in eukaryotes,55,56 collisions between RNAP and downstream replication origins could lead to a dispersive redistribution of Mcm2-7 DHs, which may contribute to the establishment of the broad initiation zones typically observed in higher eukaryotes.
The small intergenic regions in budding yeast are incompatible with the formation of broad initiation zones. Yet, stochastic transcription termination, pervasive transcription,57 and heterogenous mRNA transcription 58 may all lead to frequent collisions between transcription complexes and Mcm2-7 DHs at budding yeast origins, and transcripts have indeed been mapped to a large fraction of budding yeast origins.37 The stochastic nature of such collisions and the potentially limited ensuing shift in origin positions may hinder detection of origin shifts in the budding yeast genome during normal growth.
Conclusions
Replication origins in the budding yeast, S. cerevisiae, have been thought to fundamentally differ from those in higher eukaryotes due to their dependence on specific DNA sequences. Recent findings mitigate this notion by demonstrating that budding yeast origins exhibit a significant degree of flexibility. Firstly, ORC can direct functional Mcm2-7 loading at non-canonical sites if canonical origins are not available, and secondly, Mcm2-7 DHs can respond to collisions with DNA translocases by sliding away from the origin and initiating DNA replication from origin distal sites. This indicates that eukaryotic replication origins are not static genetic elements, but are mobile physical entities in the form of Mcm2-7 DHs bound around DNA. Adoption of a flexible strategy to specify and maintain replication origins, instead of a strictly deterministic one, might increase the robustness of the replication competence of eukaryotic chromosomes.
While we have focused our discussion here primarily on collisions with the transcription machinery, it has to be noted that Mcm2-7 double hexamers can be pushed along DNA by a variety of DNA translocases.19 It will, therefore, be interesting to determine how other forms of DNA metabolism may influence eukaryotic origin positions in vivo. Moreover, it will be interesting to learn if Mcm2-7 DHs can be pushed ahead of replication forks, where they could potentially serve as a local Mcm2-7 reservoir to support replication restart at sites of replication fork failure. While the benefits of enhanced mobility for Mcm2-7 DH stability on chromatin may seem obvious in light of the genomic traffic occurring on eukaryotic chromosomes, the flip side of such increased Mcm2-7 DH stability may be that it could pose a barrier for the removal of unused Mcm2-7 DHs at dormant origins from chromatin during S phase progression. While a regulated ubiquitin-dependent degradation mechanism has been proposed to mediate the removal of active Mcm2-7 helicase complexes from chromatin during the termination of DNA replication,59,60 it will be interesting to find out if unused Mcm2-7 DHs are similarly removed from chromatin by a regulated unloading mechanism, or by simple stochastic disintegration upon collision between opposing replication forks.
Disclosure of potential conflicts of interest
No potential conflicts of interest were disclosed.
Funding
Work in the Remus lab is funded by National Institute of General Medical Sciences grant R01GM107239 (D.R.) and National Institutes of Health grant P30 CA008748 (Memorial Sloan Kettering Cancer Center).
References
- [1].Gaillard H, Aguilera A. Transcription as a Threat to Genome Integrity. Ann Rev Biochem 2016; PMID:27023844 [DOI] [PubMed] [Google Scholar]
- [2].Siddiqui K, On KF, Diffley JF. Regulating DNA replication in eukarya. Cold Spring Harb Perspect Biol 2013; 5; PMID:23838438; http://dx.doi.org/ 10.1101/cshperspect.a012930 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [3].Remus D, Diffley JF. Eukaryotic DNA replication control: lock and load, then fire. Curr Opin Cell Biol 2009; 21:771-7; PMID:19767190; http://dx.doi.org/ 10.1016/j.ceb.2009.08.002 [DOI] [PubMed] [Google Scholar]
- [4].Rhind N, Gilbert DM. DNA replication timing. Cold Spring Harb Perspect Biol 2013; 5:a010132; PMID:23838440; http://dx.doi.org/ 10.1101/cshperspect.a010132 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [5].Donovan S, Harwood J, Drury LS, Diffley JF. Cdc6p-dependent loading of Mcm proteins onto pre-replicative chromatin in budding yeast. Proc Natl Acad Sci U S A 1997; 94:5611-6; PMID:9159120; http://dx.doi.org/ 10.1073/pnas.94.11.5611 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [6].Rowles A, Tada S, Blow JJ. Changes in association of the Xenopus origin recognition complex with chromatin on licensing of replication origins. J Cell Sci 1999; 112 ( Pt 12):2011-8; PMID:10341218 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [7].Gros J, Devbhandari S, Remus D. Origin plasticity during budding yeast DNA replication in vitro. EMBO J 2014; 33:621-36; PMID:24566988; http://dx.doi.org/ 10.1002/embj.201387278 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [8].Yeeles JT, Deegan TD, Janska A, Early A, Diffley JF. Regulated eukaryotic DNA replication origin firing with purified proteins. Nature 2015; 519:431-5; PMID:25739503; http://dx.doi.org/ 10.1038/nature14285 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [9].Kuipers MA, Stasevich TJ, Sasaki T, Wilson KA, Hazelwood KL, McNally JG, Davidson MW, Gilbert DM. Highly stable loading of Mcm proteins onto chromatin in living cells requires replication to unload. J Cell Biol 2011; 192:29-41; PMID:21220507; http://dx.doi.org/ 10.1083/jcb.201007111 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [10].McNairn AJ, Okuno Y, Misteli T, Gilbert DM. Chinese hamster ORC subunits dynamically associate with chromatin throughout the cell-cycle. Experimental cell research 2005; 308:345-56; PMID:15950218; http://dx.doi.org/ 10.1016/j.yexcr.2005.05.009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [11].Sonneville R, Querenet M, Craig A, Gartner A, Blow JJ. The dynamics of replication licensing in live Caenorhabditis elegans embryos. J Cell Biol 2012; 196:233-46; PMID:22249291; http://dx.doi.org/ 10.1083/jcb.201110080 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [12].Aparicio OM, Weinstein DM, Bell SP. Components and dynamics of DNA replication complexes in S. cerevisiae: redistribution of MCM proteins and Cdc45p during S phase. Cell 1997; 91:59-69; PMID:9335335; http://dx.doi.org/ 10.1016/S0092-8674(01)80009-X [DOI] [PubMed] [Google Scholar]
- [13].Chen S, de Vries MA, Bell SP. Orc6 is required for dynamic recruitment of Cdt1 during repeated Mcm2-7 loading. Gen Dev 2007; 21:2897-907; PMID:18006685; http://dx.doi.org/ 10.1101/gad.1596807 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [14].Cocker JH, Piatti S, Santocanale C, Nasmyth K, Diffley JF. An essential role for the Cdc6 protein in forming the pre-replicative complexes of budding yeast. Nature 1996; 379:180-2; PMID:8538771; http://dx.doi.org/ 10.1038/379180a0 [DOI] [PubMed] [Google Scholar]
- [15].Semple JW, Da-Silva LF, Jervis EJ, Ah-Kee J, Al-Attar H, Kummer L, Heikkila JJ, Pasero P, Duncker BP. An essential role for Orc6 in DNA replication through maintenance of pre-replicative complexes. EMBO J 2006; 25:5150-8; PMID:17053779; http://dx.doi.org/ 10.1038/sj.emboj.7601391 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [16].Zegerman P, Diffley JF. DNA replication as a target of the DNA damage checkpoint. DNA repair 2009; 8:1077-88; PMID:19505853; http://dx.doi.org/ 10.1016/j.dnarep.2009.04.023 [DOI] [PubMed] [Google Scholar]
- [17].Hyrien O. Peaks cloaked in the mist: the landscape of mammalian replication origins. J Cell Biol 2015; 208:147-60; PMID:25601401; http://dx.doi.org/ 10.1083/jcb.201407004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [18].Alver RC, Chadha GS, Blow JJ. The contribution of dormant origins to genome stability: from cell biology to human genetics. DNA repair 2014; 19:182-9; PMID:24767947; http://dx.doi.org/ 10.1016/j.dnarep.2014.03.012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [19].Gros J, Kumar C, Lynch G, Yadav T, Whitehouse I, Remus D. Post-licensing Specification of Eukaryotic Replication Origins by Facilitated Mcm2-7 Sliding along DNA. Mol Cell 2015; 60:797-807; PMID:26656162; http://dx.doi.org/ 10.1016/j.molcel.2015.10.022 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [20].Abid Ali F, Costa A. The MCM Helicase Motor of the Eukaryotic Replisome. J Mol Biol. 2016 May 8;428(9 Pt B):1822-32. [DOI] [PubMed] [Google Scholar]
- [21].Deegan TD, Diffley JF. MCM: one ring to rule them all. Curr Opin Struct Biol 2016; 37:145-51; PMID:26866665; http://dx.doi.org/ 10.1016/j.sbi.2016.01.014 [DOI] [PubMed] [Google Scholar]
- [22].Remus D, Beuron F, Tolun G, Griffith JD, Morris EP, Diffley JF. Concerted loading of Mcm2-7 double hexamers around DNA during DNA replication origin licensing. Cell 2009; 139:719-30; PMID:19896182; http://dx.doi.org/ 10.1016/j.cell.2009.10.015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [23].Sun J, Fernandez-Cid A, Riera A, Tognetti S, Yuan Z, Stillman B, Speck C, Li H. Structural and mechanistic insights into Mcm2-7 double-hexamer assembly and function. Gen Dev 2014; 28:2291-303; PMID:25319829; http://dx.doi.org/ 10.1101/gad.242313.114 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [24].Hua XH, Newport J. Identification of a preinitiation step in DNA replication that is independent of origin recognition complex and cdc6, but dependent on cdk2. J Cell Biol 1998; 140:271-81; PMID:9442103; http://dx.doi.org/ 10.1083/jcb.140.2.271 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [25].Li N, Zhai Y, Zhang Y, Li W, Yang M, Lei J, Tye BK, Gao N. Structure of the eukaryotic MCM complex at 3.8 A. Nature 2015; 524:186-91; PMID:26222030; http://dx.doi.org/ 10.1038/nature14685 [DOI] [PubMed] [Google Scholar]
- [26].Evrin C, Clarke P, Zech J, Lurz R, Sun J, Uhle S, Li H, Stillman B, Speck C. A double-hexameric MCM2-7 complex is loaded onto origin DNA during licensing of eukaryotic DNA replication. Proc Natl Acad Sci U S A 2009; 106:20240-5; PMID:19910535; http://dx.doi.org/ 10.1073/pnas.0911500106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [27].Huberman JA, Riggs AD. On the mechanism of DNA replication in mammalian chromosomes. J Mol Biol 1968; 32:327-41; PMID:5689363; http://dx.doi.org/ 10.1016/0022-2836(68)90013-2 [DOI] [PubMed] [Google Scholar]
- [28].Kawasaki Y, Kim HD, Kojima A, Seki T, Sugino A. Reconstitution of Saccharomyces cerevisiae prereplicative complex assembly in vitro. Gen Cells 2006; 11:745-56; http://dx.doi.org/ 10.1111/j.1365-2443.2006.00975.x [DOI] [PubMed] [Google Scholar]
- [29].Froelich CA, Kang S, Epling LB, Bell SP, Enemark EJ. A conserved MCM single-stranded DNA binding element is essential for replication initiation. eLife 2014; 3:e01993; PMID:24692448; http://dx.doi.org/ 10.7554/eLife.01993 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [30].On KF, Beuron F, Frith D, Snijders AP, Morris EP, Diffley JF. Prereplicative complexes assembled in vitro support origin-dependent and independent DNA replication. EMBO J 2014; 33:605-20; PMID:24566989; http://dx.doi.org/ 10.1002/embj.201387369 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [31].Gorman J, Chowdhury A, Surtees JA, Shimada J, Reichman DR, Alani E, Greene EC. Dynamic basis for one-dimensional DNA scanning by the mismatch repair complex Msh2-Msh6. Mol Cell 2007; 28:359-70; PMID:17996701; http://dx.doi.org/ 10.1016/j.molcel.2007.09.008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [32].Stukenberg PT, Studwell-Vaughan PS, O'Donnell M. Mechanism of the sliding beta-clamp of DNA polymerase III holoenzyme. J Biol Chem 1991; 266:11328-34; PMID:2040637 [PubMed] [Google Scholar]
- [33].MacAlpine DM, Bell SP. A genomic view of eukaryotic DNA replication. Chromosome Res 2005; 13:309-26; PMID:15868424; http://dx.doi.org/ 10.1007/s10577-005-1508-1 [DOI] [PubMed] [Google Scholar]
- [34].Kim M, Krogan NJ, Vasiljeva L, Rando OJ, Nedea E, Greenblatt JF, Buratowski S. The yeast Rat1 exonuclease promotes transcription termination by RNA polymerase II. Nature 2004; 432:517-22; PMID:15565157; http://dx.doi.org/ 10.1038/nature03041 [DOI] [PubMed] [Google Scholar]
- [35].Luo W, Johnson AW, Bentley DL. The role of Rat1 in coupling mRNA 3′-end processing to transcription termination: implications for a unified allosteric-torpedo model. Gen Dev 2006; 20:954-65; PMID:16598041; http://dx.doi.org/ 10.1101/gad.1409106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [36].McGuffee SR, Smith DJ, Whitehouse I. Quantitative, genome-wide analysis of eukaryotic replication initiation and termination. Mol Cell 2013; 50:123-35; PMID:23562327; http://dx.doi.org/ 10.1016/j.molcel.2013.03.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [37].Looke M, Reimand J, Sedman T, Sedman J, Jarvinen L, Varv S, Peil K, Kristjuhan K, Vilo J, Kristjuhan A. Relicensing of transcriptionally inactivated replication origins in budding yeast. J Biol Chem 2010; 285:40004-11; PMID:20962350; http://dx.doi.org/ 10.1074/jbc.M110.148924 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [38].Mori S, Shirahige K. Perturbation of the activity of replication origin by meiosis-specific transcription. J Biol Chem 2007; 282:4447-52; PMID:17170106; http://dx.doi.org/ 10.1074/jbc.M609671200 [DOI] [PubMed] [Google Scholar]
- [39].Snyder M, Sapolsky RJ, Davis RW. Transcription interferes with elements important for chromosome maintenance in Saccharomyces cerevisiae. Mol Cell Biol 1988; 8:2184-94; PMID:3290652; http://dx.doi.org/ 10.1128/MCB.8.5.2184 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [40].Nieduszynski CA, Blow JJ, Donaldson AD. The requirement of yeast replication origins for pre-replication complex proteins is modulated by transcription. Nucleic Acids Res 2005; 33:2410-20; PMID:15860777; http://dx.doi.org/ 10.1093/nar/gki539 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [41].Deshpande AM, Newlon CS. The ARS consensus sequence is required for chromosomal origin function in Saccharomyces cerevisiae. Mol Cell Biol 1992; 12:4305-13; PMID:1406623; http://dx.doi.org/ 10.1128/MCB.12.10.4305 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [42].Marahrens Y, Stillman B. A yeast chromosomal origin of DNA replication defined by multiple functional elements. Science 1992; 255:817-23; PMID:1536007; http://dx.doi.org/ 10.1126/science.1536007 [DOI] [PubMed] [Google Scholar]
- [43].Rivier DH, Rine J. An origin of DNA replication and a transcription silencer require a common element. Science 1992; 256:659-63; PMID:1585179; http://dx.doi.org/ 10.1126/science.1585179 [DOI] [PubMed] [Google Scholar]
- [44].Stinchcomb DT, Struhl K, Davis RW. Isolation and characterisation of a yeast chromosomal replicator. Nature 1979; 282:39-43; PMID:388229; http://dx.doi.org/ 10.1038/282039a0 [DOI] [PubMed] [Google Scholar]
- [45].Bell SP, Stillman B. ATP-dependent recognition of eukaryotic origins of DNA replication by a multiprotein complex. Nature 1992; 357:128-34; PMID:1579162; http://dx.doi.org/ 10.1038/357128a0 [DOI] [PubMed] [Google Scholar]
- [46].Breier AM, Chatterji S, Cozzarelli NR. Prediction of Saccharomyces cerevisiae replication origins. Genome Biol 2004; 5:R22; PMID:15059255; http://dx.doi.org/ 10.1186/gb-2004-5-4-r22 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [47].Eaton ML, Galani K, Kang S, Bell SP, MacAlpine DM. Conserved nucleosome positioning defines replication origins. Gen Dev 2010; 24:748-53; PMID:20351051; http://dx.doi.org/ 10.1101/gad.1913210 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [48].Bogenschutz NL, Rodriguez J, Tsukiyama T. Initiation of DNA replication from non-canonical sites on an origin-depleted chromosome. PloS one 2014; 9:e114545; PMID: 25486280; http://dx.doi.org/ 10.1371/journal.pone.0114545 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [49].Dershowitz A, Snyder M, Sbia M, Skurnick JH, Ong LY, Newlon CS. Linear derivatives of Saccharomyces cerevisiae chromosome III can be maintained in the absence of autonomously replicating sequence elements. Mol Cell Biol 2007; 27:4652-63; PMID:17452442; http://dx.doi.org/ 10.1128/MCB.01246-06 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [50].Hyrien O, Maric C, Mechali M. Transition in specification of embryonic metazoan DNA replication origins. Science 1995; 270:994-7; PMID:7481806; http://dx.doi.org/ 10.1126/science.270.5238.994 [DOI] [PubMed] [Google Scholar]
- [51].Mesner LD, Hamlin JL. Specific signals at the 3′ end of the DHFR gene define one boundary of the downstream origin of replication. Gen Dev 2005; 19:1053-66; PMID:15879555; http://dx.doi.org/ 10.1101/gad.1307105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [52].Saha S, Shan Y, Mesner LD, Hamlin JL. The promoter of the Chinese hamster ovary dihydrofolate reductase gene regulates the activity of the local origin and helps define its boundaries. Gen Dev 2004; 18:397-410; PMID:14977920; http://dx.doi.org/ 10.1101/gad.1171404 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [53].Petryk N, Kahli M, d'Aubenton-Carafa Y, Jaszczyszyn Y, Shen Y, Silvain M, Thermes C, Chen CL, Hyrien O. Replication landscape of the human genome. Nat Commun 2016; 7:10208; PMID:26751768; http://dx.doi.org/ 10.1038/ncomms10208 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [54].Powell SK, MacAlpine HK, Prinz JA, Li Y, Belsky JA, MacAlpine DM. Dynamic loading and redistribution of the Mcm2-7 helicase complex through the cell cycle. EMBO J 2015; 34:531-43; PMID:25555795; http://dx.doi.org/ 10.15252/embj.201488307 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [55].Fong N, Brannan K, Erickson B, Kim H, Cortazar MA, Sheridan RM, Nguyen T, Karp S, Bentley DL. Effects of Transcription Elongation Rate and Xrn2 Exonuclease Activity on RNA Polymerase II Termination Suggest Widespread Kinetic Competition. Mol Cell 2015; 60:256-67; PMID:26474067; http://dx.doi.org/ 10.1016/j.molcel.2015.09.026 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [56].Mischo HE, Proudfoot NJ. Disengaging polymerase: terminating RNA polymerase II transcription in budding yeast. Biochimica et biophysica acta 2013; 1829:174-85; PMID:23085255; http://dx.doi.org/ 10.1016/j.bbagrm.2012.10.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [57].Jensen TH, Jacquier A, Libri D. Dealing with pervasive transcription. Mol Cell 2013; 52:473-84; PMID:24267449; http://dx.doi.org/ 10.1016/j.molcel.2013.10.032 [DOI] [PubMed] [Google Scholar]
- [58].Pelechano V, Wei W, Steinmetz LM. Extensive transcriptional heterogeneity revealed by isoform profiling. Nature 2013; 497:127-31; PMID:23615609; http://dx.doi.org/ 10.1038/nature12121 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [59].Maric M, Maculins T, De Piccoli G, Labib K. Cdc48 and a ubiquitin ligase drive disassembly of the CMG helicase at the end of DNA replication. Science 2014; 346:1253596; PMID:25342810; http://dx.doi.org/ 10.1126/science.1253596 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [60].Moreno SP, Bailey R, Campion N, Herron S, Gambus A. Polyubiquitylation drives replisome disassembly at the termination of DNA replication. Science 2014; 346:477-81; PMID:25342805; http://dx.doi.org/ 10.1126/science.1253585 [DOI] [PubMed] [Google Scholar]