

Abstract
This brief review summarizes factors associated with elite endurance performance, trends in distance running training, and participation by men and more recently women. It is framed in the context of key ideas about the physiological determinants of endurance performance but also touches on some historical and sociological factors relevant to the overall topic. Historical trends that served to increase women's participation in elite endurance events are also discussed as is the role of increased volume and intensity of training. The rapid improvement in women's world record marathon times in the 1970s and 80s are emblematic of these trends and represent a combination of increased training volume and intensity and more competitive opportunities. This occurred as bans on participation by women in endurance events were lifted. For men these same trends evolved over a much longer time frame. The main physiological factor responsible for 10–12% slower times in women compared to men at the elite level are also considered and probably centre around.
Keywords: endurance, human performance, sex differences
Introduction
This brief review on exercise performance and sex is based on a presentation given at the ‘Biomedical Basis of Elite Performance’ meeting sponsored by The Physiological Society and held at the University of Nottingham, UK in March 2016. It focuses on endurance sports in general and distance running in specific. The major reason for this focus is the availability of relatively large amounts of data on these topics in elite and sub‐elite female and male athletes. These data include laboratory‐based studies and publicly available times from competition. Additionally, there are well‐established physiological models of endurance exercise performance that can be used to frame the discussion (Joyner & Coyle, 2008). Finally, short (2–6 months) and medium (∼1 year) term training studies are available to supplement cross sectional observations in athletic subjects.
Gender vs. sex
Before considering the physiological issues, it is important to distinguish between the concepts and definitions of gender and sex (APA, 2011). These terms are frequently used interchangeably, but there are important distinctions. To clarify these distinctions the definitions provided by the American Psychological Association are clearly stated and thus useful:
Sex refers to a person's biological status and is typically categorized as male, female, or intersex (i.e. atypical combinations of features that usually distinguish male from female). There are a number of indicators of biological sex, including sex chromosomes, gonads, internal reproductive organs, and external genitalia.
Gender refers to the attitudes, feelings, and behaviours that a given culture associates with a person's biological sex. Behaviour that is compatible with cultural expectations is referred to as gender‐normative; behaviours that are viewed as incompatible with these expectations constitute gender non‐conformity.
Given that physiology is a subdiscipline of biology, the focus of this review will be on sex differences in performance. In light of several high profile issues concerning eligibility for competition at the intersection of sex and gender, the interested reader is referred to the 2015 International Olympic Committee Consensus Statement on ‘Sex Reassignment and Hyperandrogenism’ for further discussion of this topic (IOC, 2015).
History of participation and the marathon example
Depending on the sport, women were either banned from competition in endurance sports or the maximum distance for any women's event was restricted. These bans and restrictions began to be lifted in the 1960s (Cheuvront et al. 2005). Table 1 is adapted from the excellent review by Cheuvront (2005), which highlights the timeline for middle distance and distance running participation by women in the Olympics.
Table 1.
Timeline and commentary on women's historical participation in distance running
| 1928 | Several women collapse after finishing the Olympic 800 m. Women are considered too fragile for distance running and the event is banned. |
| 1960 | The women's Olympic 800 m is reinstated. |
| 1972 | A first women's Olympic 1500 m is run. Eight women ‘legally’ run the Boston Marathon. |
| 1984 | A first women's Olympic marathon is run. |
| 1998 | The Boston Marathon is host to 3500 women runners. The New York City Marathon reports 9000 women entrants. |
| 2015 | 509,000 finishers in the marathon in the US, 44% were women. |
Adapted from Cheuvront et al. 2005 and http://www.runningusa.org/marathon-report-2016?returnTo=main
Differences in the maximum distance for women's events continue into this century and as of the 2016 Olympics, the longest swimming event for women is the 800 m freestyle while the longest event for men is the 1500 m freestyle (the 800 m event is not contested for men). However, at the Fédération Internationale de Natation (FINA) World Championships both races have been held for women and men since 2000.
The initial marathon distance was about 25 miles or 40 km beginning in the 1896 Olympics. To accommodate a race start at Windsor Castle at the 1908 London Olympics, the race was extended to 26.2 miles or 42.2 km. This distance became the standard in the early 1920s.
Figure 1 shows the progression of world best marathon times for men and women. For men, there was a rapid reduction in record marathon time up to the early 1960s. This resulted from increased volume and intensity of training by top athletes. Anecdotal reports from the early 1900s indicate that top distance runners did not train daily, did not train year round, and covered only modest distances in training. This evolved during the first half of the 20th century and by the 1950s top athletes from many countries were training for several hours per day year‐round. This training also frequently included interval training or other forms of high intensity training (Bourne, 2008). While training has continued to evolve, the main features of modern training were clearly in place by this time. From a physiological perspective, the training that emerged by the 1950s was probably sufficient to maximize a given individual's biological upper limit for factors like maximal oxygen uptake and the mitochondrial content in their skeletal muscles (Joyner & Coyle, 2008). Maximal adaptations in these well‐accepted cardiovascular and metabolic determinants of performance, at least in some elite athletes, have thus probably been occurring for ∼60 years.
Figure 1. History of world records in the marathon for women and men.
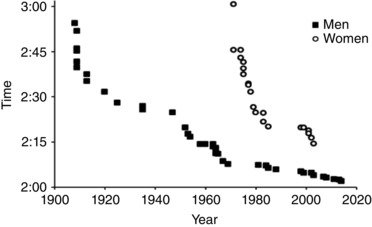
From http://www.marathonguide.com/history/records/ and other miscellaneous sources.
After the early 1960s, three factors have probably contributed to the roughly 10‐min further improvement in world record marathon times for men. These are the globalization of sport as exemplified by the emergence of East African runners since that time, professionalization starting in the late 1970s and early 1980s, and (unfortunately) doping. Of note, while marathon records continue to fall, times for the 5000 and 10,000 m races have been stable for the last 10 or so years with fewer ‘outlier’ performances as drug testing has improved (Kruse et al. 2014). Concerns about doping and record times also persist for women because many records and top marks are decades old, dating back to the era of systematic ‘state sponsored’ doping in countries such as the former East Germany and China (Franke & Berendonk, 1997; NYT, 2016). These concerns have re‐emerged in the case of Russia and the high profile doping cases involving East Africans (BBC, 2016). However, to be clear, doping is a world‐wide problem and no country is immune from its corrupting influence.
For the women's marathon, records fell much faster starting in the era of mass participation beginning in the 1970s. The simple explanation for this rapid drop in times over only 10–15 years is that for women the factors that led to the evolution of times for men over nearly a century were compressed (Pate & O'Neill, 2007). Increased training intensity and volume, globalization, more competitive opportunities and professionalization happened essentially all at once. This interpretation refutes speculation by Whipp & Ward (1992) that women might one day outrun men for the marathon based on regression analysis of records over time. Of note this paper also made similar claims for the 1500 m, but closer inspection of the key figures also suggests that the record pace by women for the marathon would one day be faster than the record pace for 1500 m which would seem unlikely under any circumstance. One interesting caveat about this paper was the extent to which the authors might or might not have been pulling an elaborate spoof on the scientific community in a high profile journal like Nature to illustrate the limits of linear regression and causal inferences.
Sex differences in performance
Women's distance running records and top times in comparable cohorts of elite athletes are typically 10–12% slower than men (Hunter et al. 2011). In swimming, the differences are in the order of ∼7% (Senefeld et al. 2016) and may be due to the viscous nature of water and its effects on the curvilinear relationship between power generation and speed. Additionally, elite female athletes generally have 5–10% more body fat than men (Pate & O'Neill, 2007). Fat is buoyant in water and compared to terrestrial events is unlikely to penalize women as much in the water. The sex differences for the rarely contested 1 h cycling record appear similar to the running records (UCI, 2015). So the key question is what explains these sex differences. To explore this topic, I will provide an overview of the physiological limiting factors for endurance exercise and then highlight sex differences in these factors.
Physiological limiting factors
The physiological limiting factors associated with endurance performance and their characteristics in elite athletes have been discussed in detail in comprehensive review articles (Joyner & Coyle, 2008). Briefly, the key factors are: (maximal oxygen consumption), lactate threshold, and efficiency. Running efficiency, in a strict engineering sense, is hard to measure and thus ‘running economy’ (the oxygen cost of a given running speed) is used as an index of efficiency.
The high values seen in elite endurance athletes are explained largely by two factors. First, as shown in Fig. 2, there is a strong relationship between total body haemoglobin and (Schmidt & Prommer, 2010). Second, there is also a strong relationship between maximum stroke volume (a key determinant of maximum cardiac output) and (Joyner & Coyle, 2008). At this time, no clear genetic signature associated with very high values has been identified and importantly no DNA variants associated with either high haemoglobin or stroke volume have been identified (Lundby et al. 2016). However, at least one elite athlete with a rare variant in his erythropoietin receptor system, which resulted in a very high red cell mass, won multiple Olympic medals in cross‐country skiing and was reported to have a very high (Juvonen et al. 1991).
Figure 2. Relationship between haemoglobin mass and in humans.
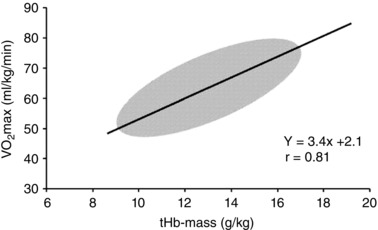
Figure adapted from Schmidt & Prommer (2010).
can be expressed in both absolute and relative terms and it is also scaled to body size (Jensen et al. 2001). For male distance runners, values in the range of 70–85 ml kg−1 min−1 are frequently seen (Joyner & Coyle, 2008). In very large cohorts of male athletes who have qualified for top level international competition in endurance sports, values in the middle to upper 70s or even higher have been reported. Fewer data are available in truly elite women, but values ∼10% lower than those seen in men are typically reported when expressed as ml kg−1 min−1. For example, a cohort of elite American female marathon runners had values of 67.1 ± 4.2 vs. 74.1 ± 2.6 ml kg−1 min−1 (Pate & O'Neill, 2007). These differences are normally attributed to a combination of higher body fat in women and lower red cell mass for a given body weight.
Consistent with the body composition differences discussed above, expressed per kilogram of fat free mass is comparable in elite endurance athletes. However, a key point is that running is weight dependent and thus expression of per kilogram is most relevant to distance running. When expressed in absolute terms (e.g. l min−1), the larger body size of men plays a critical role, so men typically have much higher values in a given athletic discipline. However, it is possible that heavy weight female rowers for example might have absolute values higher than those seen in smaller male distance runners. This caveat highlights the need to carefully consider issues like body size, scaling and absolute vs. relative values when discussing (Jensen et al. 2001). It is also interesting to note that, at least for running, the top values in elite women overlap with the lower end of the range of values seen in elite men.
An important recent caveat in this discussion is that in response to a year of prolonged intense training, the increases in stroke volume and cardiac mass observed in women were less than those observed in men (Howden et al. 2015). The relevance of this observation to elite athletes is unclear but deserves experimental follow‐up. Additionally, there is some evidence that women have ‘smaller’ lungs than men relative to body size and are more prone to arterial desaturation during heavy exercise which could limit performance (Harms et al. 1998). Thus there may be sex differences in some of the physiological determinants of in some women in comparison to men that are not explained simply on the basis of body size and composition. These differences are likely to be subtle and clearly demonstrate the need for more studies in larger numbers of truly elite female endurance athletes.
Lactate threshold
The role of lactic acid in muscle fatigue during exercise and the extent to which it is related to hypoxia in the active muscles has been an area of intense investigation for decades. Over the last 40 years much of the earlier dogma on these topics has been either reversed or is now viewed with much greater nuance (Brooks, 2001; Gladden, 2004). However, it is clear that, at least for the marathon, the so‐called lactate threshold (the running speed associated with a marked rise in blood lactate) is highly correlated with marathon performance (Farrell et al. 1979; Davis, 1985). It is also clear that elite athletes of both sexes can sustain running speeds that require ∼80% of their and are associated with a modest but not marked rise in blood lactate concentration (Pate & O'Neill, 2007; Joyner & Coyle, 2008; Poole et al. 2016).
The main physiological determinant of a high lactate threshold is probably the ability of the mitochondria in the trained skeletal muscles to increase in volume in response to training (Holloszy & Coyle, 1984). This adaptation permits more pyruvate to be oxidized at a given rate of glycolysis. It also shifts the regulation of muscle metabolism away from carbohydrates at any absolute level of exercise and permits a faster running speed or higher power output to be sustained at a given rate of glycogen utilization. An increase in capillary density in the trained muscles also probably plays a role in how long subjects might be able to exercise at intensities higher than their lactate thresholds (Coyle et al. 1988). In general, an hour or more of training per day is required to maximize mitochondrial adaptations in muscle (Holloszy & Coyle, 1984; Joyner & Coyle, 2008), and there is no evidence that the adaptability of women is less than that of men. At the fast speeds and high power outputs associated with elite endurance sports carbohydrate availability is a key for performance. While greater fat oxidation can spare glycogen and potentially prolong the time to fatigue it can also increase the oxygen cost of a given workload (Hawley & Leckey, 2015). How these considerations interact at the limits of elite human performance are currently the focus of intense investigation (L. Burke, personal communication).
Running economy
Of the three main factors associated with elite endurance running performance, less is known about running economy than or the lactate threshold. It is clear that the running speed generated for a given submaximal can vary by 15–20%, and among relatively homogeneous groups of highly trained male runners, running economy is a definitive factor in determining race performance (Conley & Krahenbuhl, 1980; Joyner, 1991). However, less is known about the fundamental biological determinants of running economy, and this is true for both sexes but especially women. Biomechanical factors including vertical displacement and braking probably play a role as do the elastic properties of muscles and connective tissue. Additionally, fibre type might also play a role (Joyner & Coyle, 2008). Running economy does appear (at least somewhat) trainable and at least one elite female athlete showed remarkable improvement during late adolescence and early adulthood and her values were excellent compared to men (Jones, 1998; Moore, 2016). East African runners have also been noted to have exceptional running economy and newer data suggest that very short ground contact times contribute to their exceptional economy (Larsen & Sheel, 2015; Santos‐Concejero et al. 2017).
Integration of factors
Based on the ideas outlined above, it appears that the main physiological difference between women and men that affects differing performance achievements and records in elite endurance competitions is . By contrast, lactate threshold and running economy probably do not explain much, if any, of the observed sex differences in performance. Other factors such as sex differences in thermoregulation could conceivably play a role, but these might only be apparent when environmental conditions are suboptimal for record‐setting performances. Thus their relevance to world records is questionable. Additionally, there is evidence of ongoing (but diminishing) sex differences in field depth in key competitions. In other words, the best performances of men have typically been more tightly clustered than those of women (Hunter et al. 2015). Finally, among non‐elite runners, there is strong evidence that women are much better at pacing themselves to avoid catastrophic slowing during the second half of marathon races (Deaner et al. 2015). However, pacing patterns for faster runners of both sexes are similar and marked by relatively even pacing and minimal rates of catastrophic slowing. This latter observation has led to speculation about sex differences in common sense, planning and risk taking.
Summary and personal perspective
Compared to other endurance sports, distance running has been studied much more intensively for both men and women. For almost all of the issues outlined above there are either fewer or much fewer data on elite women vs. elite men. The above discussion also focuses primarily on steady‐state physiological responses, which while still relevant might be less so in events like cycling or cross‐country skiing that are frequently marked by varied terrain and thus not typically steady‐state efforts. Thus more data are clearly needed in elite athletes of both sexes, but especially women. The advent of obtaining data during competitive events in elite athletes of both sexes using calibrated field monitors also needs much wider utilization. These data will be critical to ask both general and sex‐specific questions and generate new hypotheses and insights about the limits of human performance.
On a personal note, starting in the late 1970s and into the 1980s I was lucky enough to participate in distance running as a college undergraduate and into medical school and beyond. During this time, distance running became widely open to women and I participated on successful co‐ed teams. In this context, it was an incredible privilege to have a ‘front row seat’ as my female teammates made what was once thought to be dangerous and impossible, routine. The larger lesson is that it is a grave mistake to underestimate or limit any individual or group of people on the basis of superficial preconceived notions and biases. I am forever grateful to these women for teaching me this lesson.
Additional information
Competing interests
None declared.
Acknowledgements
I would like to acknowledge my teammate and friend of more than 40 years Marjorie (Kaput) Becklund, J.D., who won a bronze medal in the marathon at the 1983 World University Games. She is one of the most tenacious and determined competitors I have ever met.
Biography
Michael J. Joyner is the Caywood Professor of Anesthesiology at Mayo Clinic where he was named Distinguished Investigator in 2010. His research focuses on exercise physiology, human performance, blood pressure regulation, metabolism and transfusion practices. He is also interested in the limits of scientific reductionism and the de‐hyping of biomedical communications. The latter interest has led to a number of thought provoking and critical essays on the so‐called Precision Medicine narrative in the popular press and key scientific journals. Professor Joyner attended the University of Arizona where he graduated with a BS in 1981 and an MD in 1987. As an undergraduate he participated in athletics with a marathon best of 2:25:44. After leaving Arizona he completed his residency training in anesthesiology at the Mayo Clinic in 1993 and then developed an independent research laboratory that has been funded by NIH since that time. His former fellows have established research programmes at leading institutions throughout the world and he has held leadership positions at Mayo, in the extramural research community, and with leading journals.

This review was presented as a plenary lecture which took place at the meeting of The Biomedical Basis of Elite Performance in Nottingham, UK, 6–8 March 2016.
References
- APA (2011). Definition of Terms: Sex, Gender, Gender Identity, Sexual Orientation. American Psychological Association, Washington, DC. [Google Scholar]
- BBC (2016). http://www.bbc.com/news/world-europe-36578239.
- Bourne N (2008). Fast science: A history of training theory and methods for elite runners through 1975. University of Texas at Austin; http://hdl.handle.net/2152/17760. [Google Scholar]
- Brooks GA (2001). Lactate doesn't necessarily cause fatigue: why are we surprised? J Physiol 536, 1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheuvront SN, Carter R, Deruisseau KC & Moffatt RJ (2005). Running performance differences between men and women: an update. Sports Med 35, 1017–1024. [DOI] [PubMed] [Google Scholar]
- Conley DL & Krahenbuhl GS (1980). Running economy and distance running performance of highly trained athletes. Med Sci Sports Exerc 12, 357–360. [PubMed] [Google Scholar]
- Coyle EF, Coggan AR, Hopper MK & Walters TJ (1988). Determinants of endurance in well‐trained cyclists. J Appl Physiol (1985) 64, 2622–2630. [DOI] [PubMed] [Google Scholar]
- Davis JA (1985). Anaerobic threshold: review of the concept and directions for future research. Med Sci Sports Exerc 17, 6–21. [PubMed] [Google Scholar]
- Deaner RO, Carter RE, Joyner MJ & Hunter SK (2015). Men are more likely than women to slow in the marathon. Med Sci Sports Exerc 47, 607–616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Farrell P, Wilmore JH, Coyle EF, Billing JE & Costill DL (1979). Plasma lactate accumulation and distance running performance. Med Sci Sports 25, 1091–1097. [DOI] [PubMed] [Google Scholar]
- Franke WW & Berendonk B (1997). Hormonal doping and androgenization of athletes: a secret program of the German Democratic Republic government. Clin Chem 43, 1262–1279. [PubMed] [Google Scholar]
- Gladden LB (2004). Lactate metabolism: a new paradigm for the third millennium. J Physiol 558, 5–30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harms CA, McClaran SR, Nickele GA, Pegelow DF, Nelson WB & Dempsey JA (1998). Exercise‐induced arterial hypoxaemia in healthy young women. J Physiol 507, 619–628. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hawley JA & Leckey JJ (2015). Carbohydrate dependence during prolonged, intense endurance exercise. Sports Med 45 (Suppl. 1), 5–12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holloszy JO & Coyle EF (1984). Adaptations of skeletal muscle to endurance exercise and their metabolic consequences. J Appl Physiol Respir Environ Exerc Physiol 56, 831–838. [DOI] [PubMed] [Google Scholar]
- Howden EJ, Perhonen M, Peshock RM, Zhang R, Arbab‐Zadeh A, Adams‐Huet B & Levine BD (2015). Females have a blunted cardiovascular response to one year of intensive supervised endurance training. J Appl Physiol (1985) 119, 37–46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hunter SK, Joyner MJ & Jones AM (2015). The two‐hour marathon: What's the equivalent for women? J Appl Physiol (1985) 118, 1321–1323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hunter SK, Stevens AA, Magennis K, Skelton KW & Fauth M (2011). Is there a sex difference in the age of elite marathon runners? Med Sci Sports Exerc 43, 656–664. [DOI] [PubMed] [Google Scholar]
- IOC (2015). IOC Consensus Meeting on Sex Reassignment and Hyperandrogenism November 2015 . International Olympic Committee, https://stillmed.olympic.org/Documents/Commissions_PDFfiles/Medical_commission/2015-11_ioc_consensus_meeting_on_sex_reassignment_and_hyperandrogenism-en.pdf.
- Jensen K, Johansen L & Secher NH (2001). Influence of body mass on maximal oxygen uptake: effect of sample size. Eur J Appl Physiol 84, 201–205. [DOI] [PubMed] [Google Scholar]
- Jones AM (1998). A five year physiological case study of an Olympic runner. Br J Sports Med 32, 39–43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Joyner MJ (1991). Modeling: optimal marathon performance on the basis of physiological factors. J Appl Physiol (1985) 70, 683–687. [DOI] [PubMed] [Google Scholar]
- Joyner MJ & Coyle EF (2008). Endurance exercise performance: the physiology of champions. J Physiol 586, 35–44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Juvonen E, Ikkala E, Fyhrquist F & Ruutu T (1991). Autosomal dominant erythrocytosis caused by increased sensitivity to erythropoietin. Blood 78, 3066–3069. [PubMed] [Google Scholar]
- Kruse TN, Carter RE, Rosedahl JK & Joyner MJ (2014). Speed trends in male distance running. PLos One 9, e112978. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Larsen HB & Sheel AW (2015). The Kenyan runners. Scand J Med Sci Sports 25 (Suppl. 4), 110–118. [DOI] [PubMed] [Google Scholar]
- Lundby C, Montero D& Joyner MJ (2016). Biology of VO2max: looking under the physiology lamp. Acta Physiol (Oxf) (in press; DOI: 10.1111/apha.12827). [DOI] [PubMed] [Google Scholar]
- Moore IS (2016). Is there an economical running technique? a review of modifiable biomechanical factors affecting running economy. Sports Med 46, 793–807. [DOI] [PMC free article] [PubMed] [Google Scholar]
- NYT (2016). http://www.nytimes.com/2016/02/25/world/asia/china-olympics-doping-ma-junren.html?_r=0.
- Pate RR & O'Neill JR (2007). American women in the marathon. Sports Med 37, 294–298. [DOI] [PubMed] [Google Scholar]
- Poole DC, Burnley M, Vanhatalo A, Rossiter HB & Jones AM (2016). Critical power: an important fatigue threshold in exercise physiology. Med Sci Sports Exerc 48, 2320–2334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Santos‐Concejero J, Tam N, Coetzee DR, Oliván J, Noakes TD & Tucker R (2017). Are gait characteristics and ground reaction forces related to energy cost of running in elite Kenyan runners? J Sports Sci 35, 531–538. [DOI] [PubMed] [Google Scholar]
- Senefeld J, Joyner MJ, Stevens A & Hunter SK (2016). Sex differences in elite swimming with advanced age are less than marathon running. Scand J Med Sci Sports 26, 17–28. [DOI] [PubMed] [Google Scholar]
- Schmidt W & Prommer N (2010). Impact of alterations in total hemoglobin mass on VO2max. Exerc Sport Sci Rev 38, 68–75. [DOI] [PubMed] [Google Scholar]
- UCI (2015). Timeline of modern UCI Hour Record In UCI Union Cycliste Internationale, http://www.uci.ch/track/news/article/timeline-modern-uci-hour-record/. [Google Scholar]
- Whipp BJ & Ward SA (1992). Will women soon outrun men? Nature 355, 25. [DOI] [PubMed] [Google Scholar]