

Summary
Microorganisms able to degrade aromatic contaminants constitute potential valuable biocatalysts to deal with a significant reusable carbon fraction suitable for eco‐efficient valorization processes. Metabolic engineering of anaerobic pathways for degradation and recycling of aromatic compounds is an almost unexplored field. In this work, we present the construction of a functional bzd cassette encoding the benzoyl‐CoA central pathway for the anaerobic degradation of benzoate. The bzd cassette has been used to expand the ability of some denitrifying bacteria to use benzoate as sole carbon source under anaerobic conditions, and it paves the way for future pathway engineering of efficient anaerobic biodegraders of aromatic compounds whose degradation generates benzoyl‐CoA as central intermediate. Moreover, a recombinant Azoarcus sp. CIB strain harbouring the bzd cassette was shown to behave as a valuable biocatalyst for anaerobic toluene valorization towards the synthesis of poly‐3‐hydroxybutyrate (PHB), a biodegradable and biocompatible polyester of increasing biotechnological interest as a sustainable alternative to classical oil‐derived polymers.
Introduction
Aromatic compounds are the second most abundant class of organic compounds in nature after carbohydrates. Due to the thermodynamic stability of the aromatic ring, aromatic compounds are difficult to degrade and they tend to persist in the environment for long periods of time. Many of these compounds are toxic and/or carcinogenic thus representing major persistent environmental pollutants. Therefore, removal of aromatic compounds is very important both for a balanced global carbon budget and to protect wildlife and human health. Some specialized microorganisms (bacteria, archaea and fungi) have adapted to use aromatic compounds as sole carbon and energy source (mineralization) or, at least, partially degrade these molecules to less‐toxic and persistent compounds (Carmona et al., 2009; Fuchs et al., 2011). These microorganisms constitute, thus, potential valuable biocatalysts to deal with a significant reusable carbon fraction suitable for eco‐efficient valorization processes within the framework of a sustainable knowledge‐based bio‐economy.
There are two major strategies to degrade aromatic compounds depending on the presence or absence of oxygen. In the aerobic catabolism of aromatic compounds, oxygen is not only the final electron acceptor but also a cosubstrate for two key processes, i.e. the hydroxylation and oxygenolytic ring cleavage of the aromatic ring, carried out by oxygenases. In the absence of oxygen (anaerobic catabolism), the aromatic ring is de‐aromatized by reductive reactions (Carmona et al., 2009; Fuchs et al., 2011; Boll et al., 2014; Rabus et al., 2016). A wide variety of bacteria, pathways and associated gene clusters responsible for the aerobic catabolism of aromatic compounds have been studied and characterized (Díaz et al., 2013). Moreover, genetic and metabolic engineering approaches have been applied to develop more efficient recombinant biocatalysts for the aerobic conversion of aromatic compounds to added value products, e.g. biopolymers, biofuels, commodity chemicals (Kosa and Ragauskas, 2012; Bugg and Rahmanpour, 2015; Wierckx et al., 2015; Beckham et al., 2016; Johnson et al., 2016). In contrast, the anaerobic degradation of aromatic compounds has been much less well‐studied than the aerobic degradation, especially regarding the genetic determinants that encode the anaerobic pathways, and there are only a few examples of recombinant anaerobic biodegraders (Coschigano et al., 1994; Darley et al., 2007; Zamarro et al., 2016). However, anaerobic processes may offer significant benefits compared to aerobic bioprocesses, e.g. higher yields, less heat and oxidative stress generation, reduced biomass production and lower mechanical energy input, as they do not require aeration, which can significantly reduce production costs (Cueto‐Rojas et al., 2015). Thus, metabolic engineering of anaerobic pathways for degradation and recycling of waste aromatic compounds is still an almost unexplored field of great biotechnological potential.
Similar to the very well‐known aerobic degradation strategies, the anaerobic degradation of aromatic compounds channels a wide variety of compounds into a few central intermediates through devoted peripheral degradation pathways (catabolic funnel). The different peripheral pathways converge into a few central pathways that carry out the reductive de‐aromatization and further conversion of the central intermediates to compounds of the central metabolism of the cell. Most monocyclic aromatic compounds are channelled and activated to benzoyl‐CoA. The catabolic genes encoding the benzoyl‐CoA central pathway enzymes are usually arranged in large chromosomal clusters that also contain the specific transcriptional regulators (Egland et al., 1997; Breese et al., 1998; López‐Barragán et al., 2004; Rabus et al., 2005, 2016; Wischgoll et al., 2005; Carmona et al., 2009; Holmes et al., 2012; Carlström et al., 2015; Hirakawa et al., 2015).
Azoarcus sp. CIB is a facultative anaerobic beta‐proteobacterium capable of degrading either aerobically and/or anaerobically (using nitrate as terminal electron acceptor) a wide range of aromatic compounds including some toxic hydrocarbons such as toluene (López‐Barragán et al., 2004; Martín‐Moldes et al., 2015; Zamarro et al., 2016). Several pathways involved in the anaerobic degradation of aromatic compounds, including the benzoyl‐CoA central pathway (bzd genes), have been characterized at the molecular level in strain CIB (López‐Barragán et al., 2004; Carmona et al., 2009; Juárez et al., 2013). In addition to this free‐living lifestyle, Azoarcus sp. CIB also shows an endophytic lifestyle (Fernández et al., 2014) and is able to resist some metals and metalloids, e.g. selenium oxyanions, producing nanoparticles of biotechnological interest (Fernández‐Llamosas et al., 2016). All these properties, together with the fact that the genome of strain CIB has been sequenced and annotated and different tools are available for its genetic manipulation (Martín‐Moldes et al., 2015), make Azoarcus sp. CIB a promising host for approaching metabolic engineering strategies to improve the anaerobic bioconversion of aromatic compounds.
In this work, we present the construction of a functional bzd cassette for anaerobic benzoate degradation and its application to the development of recombinant Azoarcus sp. CIB biocatalysts for toluene valorization towards the synthesis of poly‐3‐hydroxybutyrate (PHB), a biodegradable and biocompatible polyester of increasing biotechnological interest as a sustainable alternative to classical oil‐derived polymers (Rehm, 2010; Nikodinovic‐Runic et al., 2013; Madbouly et al., 2014).
Results and discussion
Construction of a functional bzd catabolic cassette for anaerobic degradation of benzoate
As indicated above, the benzoyl‐CoA central pathway involved in the anaerobic degradation of benzoate and many other aromatic compounds whose peripheral pathways converge into benzoyl‐CoA has been previously characterized in the denitrifying Azoarcus sp. CIB strain (López‐Barragán et al., 2004; Carmona et al., 2009). Benzoate becomes initially activated to benzoyl‐CoA by the benzoate‐CoA ligase (BzdA) and is then de‐aromatized by the action of a benzoyl‐CoA reductase (BzdNOPQ), the only oxygen‐sensitive enzyme within the benzoyl‐CoA pathway, that uses a low‐potential ferredoxin (BzdM) as electron donor and generates cyclohexa‐1,5‐diene‐1‐carbonyl‐CoA (1,5‐dienoyl‐CoA) (Fig. 1A). Further degradation of 1,5‐dienoyl‐CoA resembles a modified β‐oxidation pathway with addition of water to a double bond (BzdW dienoyl‐CoA hydratase), a dehydrogenation reaction (BzdX hydroxyacyl‐CoA dehydrogenase) and hydrolytic ring fission (BzdY oxoacyl‐CoA hydrolase), generating finally 3‐hydroxy‐pimelyl‐CoA that feeds into a lower pathway (Fig. 1A). The bzd genes encoding the bzd pathway enzymes are clustered together in a large operon driven by the P N promoter (Fig. 1B). The specific transcriptional regulation of the bzd operon is conducted by the BzdR repressor that is encoded immediately upstream of the catabolic operon (Fig. 1B). Induction of the bzd genes requires the binding of the effector molecule, benzoyl‐CoA, to the BzdR repressor (Durante‐Rodríguez et al., 2010). The bzdNOPQMSTUVWXYZA catabolic genes and the cognate bzdR regulatory gene have been engineered as a 19.6 kb DNA cassette into a broad‐host range vector, giving rise to plasmid pLB1 (Fig. 1B). To construct the bzd cassette, the right end of the bzd cluster (genes bzdXYZA) from plasmid pECOR8 (Table 1) was first cloned as an EcoRI fragment into the EcoRI‐digested broad‐host range pIZ1016* vector, giving rise to plasmid pIZECO (Table 1). Then, the left end of the bzd cluster (genes bzdRbzdNOPQMSTUVWX) was cloned as a XbaI/NcoI‐double digested fragment from the recombinant λBzd1 phage (López‐Barragán et al., 2004) into the XbaI/NcoI‐double digested pIZECO plasmid, giving rise to plasmid pLB1 (25.6 Kb) (Table 1). To check whether the bzd cassette was functional, it was transferred to a mutant strain, Azoarcus sp. CIBdbzdN, unable to use benzoate anaerobically because it contains a disruption insertion in the first gene of the bzd catabolic operon with avoids the expression of the rest of bzd genes (Table 1) (López‐Barragán et al., 2004). Plasmid pLB1 was transferred by biparental filter mating from E. coli S17‐1λpir (donor strain) to Azoarcus sp. CIBdbzdN (recipient strain) as previously described (López‐Barragán et al., 2004). Exconjugants harbouring the pLB1 plasmid, Azoarcus sp. CIBdbzdN (pLB1) (Table 1), were isolated aerobically on gentamicin (7.5 μg ml−1)‐containing MC medium with 10 mM glutarate as sole carbon source for counterselection of donor cells. The presence of plasmid pLB1 in Azoarcus sp. CIBdbzdN cells restored their anaerobic growth on benzoate and caused the consumption of this carbon source, as in the case of the wild‐type CIB strain containing plasmid pIZ1016 as control (Fig. 2). This result strongly suggested that the recombinant bzd cassette in plasmid pLB1 was functional. To confirm this, plasmid pLB1 was transferred to a closely related species, Azoarcus communis SWub3 (Table 1), that is an endophyte unable to degrade aromatic compounds under anaerobic conditions (Reinhold‐Hurek et al., 1993) but that can use aliphatic dicarboxylic acids, e.g. glutarate, that feed to the lower benzoyl‐CoA pathway (Fig. 1A). Remarkably, the recombinant A. communis SWub3 (pLB1) strain was able to grow anaerobically using benzoate as sole carbon and energy source (doubling time of about 15 h), confirming that the bzd cassette was functional in heterologous hosts and conferred the ability to degrade benzoate in anoxic conditions (Fig. 2).
Figure 1.
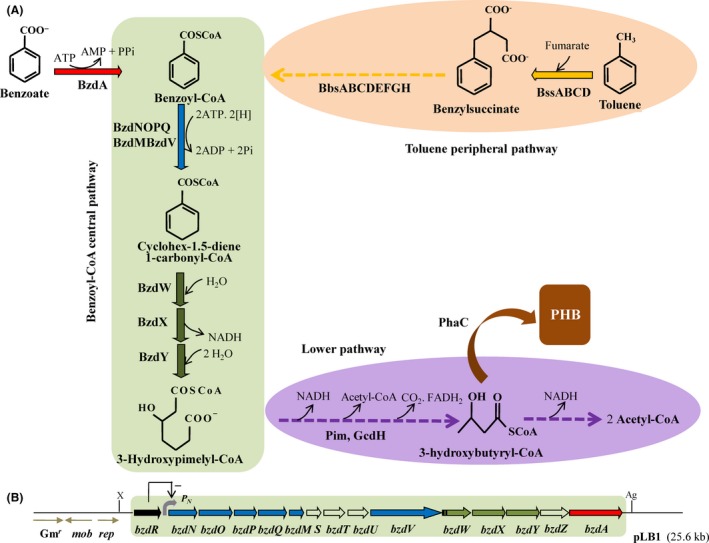
Scheme of the anaerobic metabolism of benzoate and toluene, and gene organization of the bzd cassette in Azoarcus sp. CIB.
A. Scheme of peripheral pathway for the anaerobic conversion of toluene into benzoyl‐CoA (orange), the activation of benzoate to benzoyl‐CoA (red), the benzoyl‐CoA central pathway (green), the lower pathway (violet) and the polymerization of 3‐hydroxybutyryl‐CoA to PHB (brown). Discontinuous arrows indicate that more than one enzymatic step is involved. Enzyme abbreviations: BssABCD, benzylsuccinate synthase; Bbs, enzymes involved in the modified β‐oxidation of benzylsuccinate to benzoyl‐CoA; BzdA, benzoate‐CoA ligase; BzdNOPQ, benzoyl‐CoA reductase; BzdM, ferredoxin; BzdV, putative NADPH:ferredoxin oxidoreductase; BzdW, cyclohex‐1,5‐diene‐1‐carbonyl‐CoA hydratase; BzdX, 6‐hydroxycyclohex‐1‐ene‐1‐carbonyl‐CoA dehydrogenase; BzdY, 2‐ketocyclohexane‐1‐carbonyl‐CoA hydrolase; Pim, enzymes involved in β‐oxidation of dicarboxylic acids; GcdH, glutaryl‐CoA dehydrogenase; PhaC, PHB synthase.
B. Schematic representation of the bzd genes for anaerobic benzoate degradation engineered as a mobile DNA cassette in plasmid pLB1. Genes are indicated in the same colour code than the corresponding enzymes in panel A; i.e., genes encoding enzymes involved in the initial activation, de‐aromatization and modified β‐oxidation are indicated in red, blue and green colour respectively. The bzdR gene encoding a transcriptional repressor of the catabolic PN promoter is shown in black. X and Ag, XbaI and AgeI restriction sites flanking the bzd cassette respectively. The gentamicin resistance gene (Gmr), mobilization (mob) and replication (rep) functions are also indicated.
Table 1.
Bacteria and plasmids used in this study
| Strain or plasmid | Relevant genotype and main characteristics | Reference or source |
|---|---|---|
| E. coli strains | ||
| DH5α | endA1 hsdR17 supE44 this‐1 recA1 gyrA(Nar r ) relA1 Δ/argF‐lac) U169 depRФ80dlacd(lacZ)M15 | Sambrook and Russell, (2001) |
| S17‐1λpir | Tpr Smr recA thi hsdRM + RP4׃׃2.Tc׃׃Mu׃׃Km Tn7 λpir phage lysogen | de Lorenzo and Timmis, (1994) |
| Azoarcus strains | ||
| Azoarcus sp. CIB | Wild‐type strain | López‐Barragán et al., (2004) |
| Azoarcus sp. CIBdbzdN | Kmr , Azoarcus sp. CIB with a disruption in the bzdN gene | López‐Barragán et al., (2004) |
| Azoarcus communis SWub3 | Wild‐ type strain (LMG22127) | Reinhold‐Hurek et al., (1993) |
| Azoarcus sp. CIB (pLB1) | CIB strain containing plasmid pLB1 | This work |
| Azoarcus sp. CIBdbzdN (pLB1) | CIBdbzdN strain containing plasmid pLB1 | This work |
| A. communis SWub3 (pLB1) | SWub3 strain containing plasmid pLB1 | This work |
| Plasmids | ||
| pECOR8 | Apr, pUC19 harbouring a 5.4 Kb EcoRI DNA fragment from Azoarcus sp. CIB that contains the right end of the bzd cluster | López‐Barragán et al., (2004) |
| pIZ1016 | Gmr, pBBR1MCS‐5 broad‐host range cloning vector | Moreno‐Ruiz et al., (2003) |
| pIZ1016* | Gmr, pIZ1016 derivative without the NcoI restriction site in the polylinker | This work |
| pIZECO | Gmr, 11.4 Kb‐pIZ1016* derivative harbouring the right end of the bzd cluster | This work |
| pLB1 | Gmr, 25.6 Kb‐pIZ1016* derivative harbouring the complete bzd cassette | This work |
Figure 2.
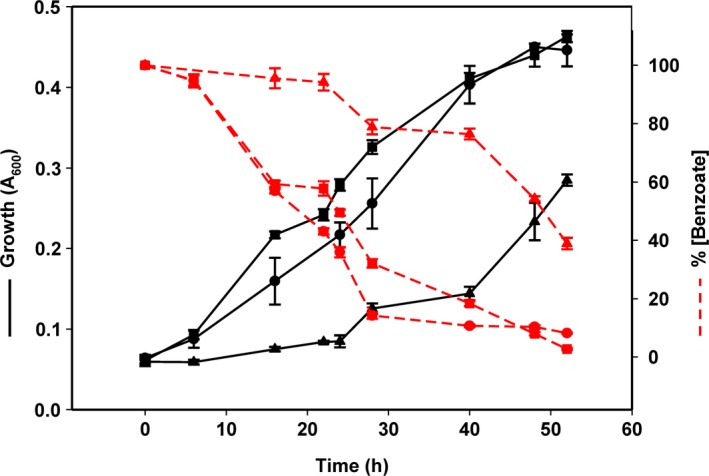
Anaerobic growth curves and benzoate consumption in different Azoarcus strains. Azoarcus sp. CIB (pIZ1016) (circles), Azoarcus sp. CIBdbzdN (pLB1) (squares) and Azoarcus communis (pLB1) (triangles) were grown anaerobically at 30°C in minimal MC medium containing 3 mM benzoate and 10 mM nitrate as sole donor and electron acceptors respectively, as previously detailed (López‐Barragán et al., 2004). Gentamicin (7.5 μg ml−1) was added to the medium to assure plasmid maintenance. Bacterial growth (black lines) was monitored by measuring the absorbance at 600 nm (A 600). The concentration of benzoate in the culture medium (red lines) was monitored spectrophotometrically at 273 nm (López‐Barragán et al., 2004) and is indicated as a percentage of the initial concentration. Values are the mean of three different experiments. Error bars indicate standard deviation.
As far as we know, this is the first report on the construction of a DNA cassette encoding a transferable benzoyl‐CoA central pathway that allows the expansion of the catabolic potential of certain facultative anaerobes towards the use of aromatic compounds under anaerobic conditions. Moreover, this result reinforces the recent thought that questions the previous idea that the Azoarcus genus comprises bacteria that fit into one of two major eco‐physiological groups, i.e. either the free‐living anaerobic biodegraders of aromatic compounds or the obligate endophytes unable to degrade aromatics under anaerobic conditions (Fernández et al., 2014). Thus, here we show that a member of the subgroup of Azoarcus strains that are obligate endophytes unable to degrade aromatics under anaerobic conditions can evolve towards the use of these carbon sources when acquiring the cognate genetic determinants.
Azoarcus sp. CIB accumulates PHB when grown anaerobically in toluene
It has been reported that some anaerobic biodegraders of aromatic hydrocarbons, such as the bacterium Aromatoleum aromaticum EbN1, are able to accumulate poly(3‐hydroxybutyrate) (PHB) up to 5.2% of the cell dry weight (CDW) during anaerobic growth on toluene using nitrate as final electron acceptor (Trautwein et al., 2008). PHB formation was predicted to contribute at two different levels, (i) enhancing consumption of surplus reducing equivalents generated during the anaerobic catabolism of aromatic hydrocarbons, and (ii) alleviating the cytotoxic effect of aromatic hydrocarbons by trapping them into the hydrophobic PHB granules (Trautwein et al., 2008; Rabus et al., 2014). To check whether Azoarcus sp. CIB was also able to accumulate PHB when grown anaerobically on toluene, PHB monomer composition and cellular PHB content of lyophilized cells were determined by gas chromatography‐tandem mass spectrometry (GC‐MS) of the methanolysed polyester as previously described (de Eugenio et al., 2010). The chromatographic profile obtained by GC‐MS of the products released showed a single peak corresponding to the 3‐hydroxy‐butyrate monomer of the PHB polymer. Figure 3A shows that Azoarcus sp. CIB cells grown anaerobically on toluene were able to accumulate PHB at a level of 13% CDW. Moreover, granules of PHB were clearly observed in Azoarcus sp. CIB cells by transmission electron microscopy (Fig. 3B).
Figure 3.
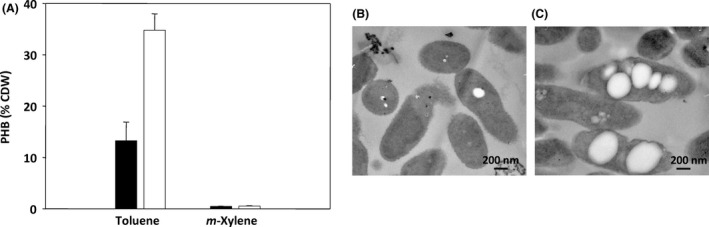
Analysis of Azoarcus sp. CIB cells that contain plasmid pIZ1016 (control) or the pLB1 plasmid grown anaerobically on aromatic hydrocarbons.
A. PHB monomer content of Azoarcus sp. CIB (pIZ1016) (black bars) and Azoarcus sp. CIB (pLB1) (white bars) cells at mid‐exponential phase of anaerobic growth at 30°C in minimal MC medium with 400 mM toluene or 275 mM m‐xylene, supplied in 2,2,4,4,6,8,8‐heptamethylnonane as an inert carrier phase, and 10 mM nitrate. PHB production was quantified by GC‐MS of the methanolysed polyester and shown as the percentage of PHB monomer with respect to the total cell dry weight (CDW). Error bars represent standard derivation found in four different experiments. Methanolysis procedure was carried out by suspending 5–10 mg of lyophilized cells in 0.5 ml of chloroform and 2 ml of methanol containing 15% sulfuric acid and 0.5 mg ml−1 of 3‐methylbenzoic acid (internal standard), and then incubated at 80°C for 7 h. After cooling, 1 ml of de‐mineralized water and 1 ml of chloroform were added and the organic phase containing the resulting methyl esters of monomers was analysed by GC‐MS (de Eugenio et al., 2010). An Agilent series 7890A coupled with 5975C MS detector (EI, 70 eV) and a split–splitless injector were used for analysis. An aliquot (1μl) of organic phase was injected into the gas chromatograph at a split ratio 1:20. Separation of compounds was achieved using an HP‐5 MS capillary column (5% phenyl‐95% methyl siloxane, 30 m × 0.25 mm film thickness). Helium was used as carrier gas at a flow rate of 1 ml min−1. The injector and transfer line temperature were set at 275 and 300°C respectively. The oven temperature programme was as follows: initial temperature 80°C for 2 min, then from 80°C up 150°C at a rate of 5°C min−1 and held for 1 min. The mass spectra were recorded in full scan mode (m/z 40–550). 3‐hydroxybutyric acid methyl ester was resolved using selected ion monitoring mode (SIM).
B and C. Representative view of Azoarcus sp. CIB (pIZ1016) (B) and Azoarcus sp. CIB (pLB1) (C) cells grown anaerobically on toluene. Cells were harvested, washed twice in PBS and fixed in 5% (w/v) glutaraldehyde. Afterwards, cells were suspended in 2.5% (w/v) OsO4 for 1 h, gradually dehydrated in ethanol solutions [30%, 50%, 70%, 90% and 100% (v/v); 30 min each) and propylene oxide (1 h), and embedded in Epon 812 resin. Ultrathin sections were cut with a microtome using a Diatome diamond knife. The sections were picked up with 400‐mesh cupper grids coated with a layer of carbon and observed using a Jeol‐1230 transmission electron microscope. PHB granules can be observed as white spheres inside the cells.
Taken together, these results show that Azoarcus sp. CIB is able to degrade toluene anaerobically and eventually reroute some of the metabolic flux towards the synthesis of a valuable biopolymer such as PHB. According to the known peripheral pathway for anaerobic toluene degradation in denitrifying bacteria, this aromatic hydrocarbon becomes activated to benzylsuccinate by the bssABCD‐encoded benzylsuccinate synthase enzyme, and then the later is converted to benzoyl‐CoA via a modified β‐oxidation pathway encoded by the bbsABCDEFJGH genes (Fig. 1A) (Heider et al., 2016; Zamarro et al., 2016). Benzoyl‐CoA is then converted to 3‐hydroxypimelyl‐CoA via the upper benzoyl‐CoA central pathway (Fig. 1A), and the later is further metabolized to the central metabolism by a dicarboxylic acid β‐oxidation pathway (lower benzoyl‐CoA pathway) (Fig. 1A) (López‐Barragán et al., 2004; Carmona et al., 2009). As one of the final metabolites of the lower benzoyl‐CoA pathway, i.e. 3‐hydroxybutyryl‐CoA, is the monomer used by the PHB synthase to produce the PHB polymer (Fig. 1A) (Rehm, 2010), the anaerobic metabolism of toluene might stimulate PHB production by increasing the 3‐hydroxybutyryl‐CoA levels in the cell. Therefore, it is tempting to speculate that increasing the flux through the benzoyl‐CoA pathway could further enhance the anaerobic conversion of toluene into PHB. In Azoarcus sp. CIB, the bss‐bbs genes are clustered together within an integrative and conjugative element ICEXTD that enhances their copy dosage in the cell (Martín‐Moldes et al., 2015; Zamarro et al., 2016), thus likely favouring the initial activation of toluene by benzylsuccinate synthase, a reaction that was shown to be a rate‐limiting step in toluene catabolism. As several gene clusters have been predicted to encode dicarboxylic acid β‐oxidation pathways in strain CIB (Martín‐Moldes et al., 2015), it appears that the single gene dosage bzd genes could represent a bottleneck when trying to enhance the metabolic flux through the benzoyl‐CoA pathway for achieving a higher conversion of toluene into PHB. Thus, the use of the pLB1 multicopy plasmid could be a rational strategy to increase the bzd gene dosage and, eventually, enhance the anaerobic production of PHB in Azoarcus sp. CIB.
The bzd cassette enhances PHB accumulation in a recombinant Azoarcus sp. CIB (pLB1) strain
To check whether the expression of the bzd genes in the multicopy plasmid pLB1 could lead to a higher anaerobic benzoate metabolism, growth and benzoate consumption in Azoarcus sp. CIB (pLB1) cells were compared with those in the wild‐type Azoarcus sp. CIB cells. As shown in Fig. 4, a slightly increase in growth and benzoate removal was observed in cells containing the pLB1 plasmid, suggesting that increasing the bzd gene dosage enhances the anaerobic degradation of benzoate in Azoarcus sp. CIB. Then, we monitored the accumulation of PHB in Azoarcus sp. CIB (pLB1) cells grown anaerobically on 400 mM toluene supplied in 2,2,4,4,6,8,8‐heptamethylnonane carrier phase. PHB production was quantified from cells harvested at mid‐exponential phase, as previously done with the wild‐type CIB strain. Interestingly, the bzd cassette caused accumulation of PHB up to 35% CDW in Azoarcus sp. CIB (pLB1) (Fig. 3A). Thus, these results show that when recombinant Azoarcus sp. CIB (pLB1) cells grow anaerobically on toluene, the bzd cassette enhances 2.7‐fold the bio‐production of PHB with respect to that observed in wild‐type cells. The higher conversion of toluene into PHB was also confirmed by transmission electron microscopy, which revealed an increase in number and size of PHB granules in recombinant cells (Fig. 3C) with respect to wild‐type cells (Fig. 3B). The level of PHB accumulation from the toxic hydrocarbon toluene was the highest described so far under anaerobic conditions, and it was even higher than that reported for the aerobic bioconversion of aromatic hydrocarbons to medium‐chain‐length polyhydroxyalkanoates (Ward et al., 2005; Nikodinovic et al., 2008; Ni et al., 2010; Narancic et al., 2012) or to PHB (Keum et al., 2008; Hori et al., 2009). As toluene is a major contaminant of high‐volume waste streams at places where it is produced or used (e.g. petrochemical industry, solvents and painting markets, biopolymers (e.g. poly(ethylene terephthalate), polyurethanes), its valorization becomes a sustainable strategy for the recycling industry (Wierckx et al., 2015).
Figure 4.
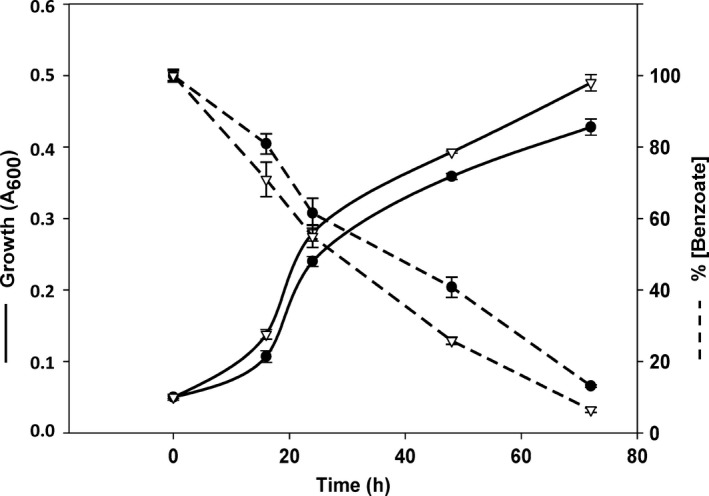
Growth and benzoate consumption of Azoarcus sp. CIB harbouring plasmid pIZ1016 (circles) or the pLB1 plasmid (triangles). Bacteria were cultivated anaerobically at 30°C in minimal MC medium containing 3 mM benzoate and 10 mM nitrate. Bacterial growth, monitored by measuring the absorbance at 600 nm (A 600), is indicated with a continuous line. The concentration of benzoate in the culture medium was monitored spectrophotometrically at 273 nm, and the percentage of benzoate remaining in the culture medium is indicated with a dashed line. Values are the mean of three different experiments. Error bars indicate standard deviation.
Azoarcus sp. CIB is also able to grow anaerobically on m‐xylene as sole carbon and energy source (Juárez et al., 2013; Zamarro et al., 2016). Although the peripheral pathway for anaerobic m‐xylene degradation is likely encoded by the same bss‐bbs genes responsible for the toluene peripheral pathway, the central intermediate of m‐xylene degradation is 3‐methylbenzoyl‐CoA rather than benzoyl‐CoA (as in the case of toluene) (Juárez et al., 2013; Zamarro et al., 2016). The 3‐methylbenzoyl‐CoA is further degraded via a specific mbd anaerobic central pathway, which is different to the common bzd pathway, and a lower pathway that does not generate 3‐hydroxybutyryl‐CoA (Juárez et al., 2013; Zamarro et al., 2016). Thus, anaerobic m‐xylene degradation might not generate as much PHB as the anaerobic catabolism of toluene. In fact, when Azoarcus sp. CIB cells were grown in 275 mM m‐xylene, supplied in 2,2,4,4,6,8,8‐heptamethylnonane carrier phase, no significant PHB accumulation was detected (< 1% CDW) (Fig. 3A). As expected, no further increase of PHB accumulation was observed in Azoarcus sp. CIB (pLB1) cells grown on m‐xylene (Fig. 3A). Therefore, these results strongly suggest that the observed effect of the multicopy bzd cassette enhancing PHB production from toluene might be due to an enhanced metabolic flux through the anaerobic benzoyl‐CoA pathway.
Conclusion
This work is a proof of concept of the potential of metabolic engineering for the anaerobic bioconversion of aromatic compounds towards the production of valuable products. We have engineered the first broad‐host range metabolic bzd cassette for the anaerobic degradation of benzoate. The bzd cassette has been used to expand the ability of some denitrifying bacteria to use benzoate as sole carbon source under anaerobic conditions, and it paves the way for future pathway engineering of efficient anaerobic biodegraders of aromatic compounds. Moreover, we have developed a recombinant Azoarcus sp. CIB strain harbouring the bzd cassette as a valuable environmentally friendly strategy for the efficient anaerobic conversion of some petrochemical waste into added value products, such as bioplastics, relevant to the circular economy.
Conflict of Interest
The authors declare no conflict of interest.
Acknowledgements
We thank A. Valencia, Fernando de la Peña and Fernando Escolar for technical assistance, Secugen S.L. for DNA sequencing and M.A. Prieto for inspiring discussions. This work was supported by grants BIO2012‐39501, BIO2016‐79736‐R and PCIN‐2014‐113 from the Ministry of Economy and Competitiveness of Spain; European Union FP7 Grant 311815; and by a grant of Fundación Ramón‐Areces XVII CN.
Microbial Biotechnology (2017) 10(6), 1418–1425
Founding Information
This work was supported by grants BIO2012‐39501, BIO2016‐79736‐R and PCIN‐2014‐113 from the Ministry of Economy and Competitiveness of Spain; European Union FP7 Grant 311815; and by a grant of Fundación Ramón‐Areces XVII CN.
References
- Beckham, GT , Johnson, CW , Karp, EM , Salvachúa, D , and Vardon, DR (2016) Opportunities and challenges in biological lignin valorization. Curr Opin Biotechnol 42: 40–53. [DOI] [PubMed] [Google Scholar]
- Boll, M , Löffler, C , Morris, BE , and Kung, JW (2014) Anaerobic degradation of homocyclic aromatic compounds via arylcarboxyl‐coenzyme A esters: organisms, strategies and key enzymes. Environ Microbiol 16: 612–627. [DOI] [PubMed] [Google Scholar]
- Breese, K , Boll, M , Alt‐Mörbe, J , Schägger, H , and Fuchs, G (1998) Genes coding for the benzoyl‐CoA pathway of anaerobic aromatic metabolism in the bacterium Thauera aromatica . Eur J Biochem 256: 148–154. [DOI] [PubMed] [Google Scholar]
- Bugg, TDH , and Rahmanpour, R (2015) Enzymatic conversion of lignin into renewable chemicals. Curr Opin Chem Biol 29: 10–17. [DOI] [PubMed] [Google Scholar]
- Carlström, CI , Loutey, D , Bauer, S , Clark, IC , Rohde, RA , Iavarone, AT , et al (2015) (Per)chlorate‐reducing bacteria can utilize aerobic and anaerobic pathways of aromatic degradation with (per)chlorate as an electron acceptor. MBio 6: e02287–14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carmona, M , Zamarro, MT , Blázquez, B , Durante‐Rodríguez, G , Juárez, JF , Valderrama, JA , et al (2009) Anaerobic catabolism of aromatic compound: a genetic and genomic view. Microbiol Mol Biol Rev 73: 71–133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coschigano, PW , Häggblom, MM , and Young, LY (1994) Metabolism of both 4‐chlorobenzoate and toluene under denitrifying conditions by a constructed bacterial strain. Appl Environ Microbiol 60: 989–995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cueto‐Rojas, HF , van Maris, AJ , Wahl, SA , and Heijnen, JJ (2015) Thermodynamics‐based design of microbial cell factories for anaerobic product formation. Trends Biotechnol 33: 534–546. [DOI] [PubMed] [Google Scholar]
- Darley, PI , Hellstern, JA , Medina‐Bellver, JI , Marqués, S , Schink, B , and Philipp, B (2007) Heterologous expression and identification of the genes involved in anaerobic degradation of 1,3‐dihydroxybenzene (resorcinol) in Azoarcus anaerobius . J Bacteriol 189: 3824–3833. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Díaz, E , Jiménez, JI , and Nogales, J (2013) Aerobic degradation of aromatic compounds. Curr Opin Biotechnol 24: 431–442. [DOI] [PubMed] [Google Scholar]
- Durante‐Rodríguez, G , Valderrama, JA , Mancheño, JM , Rivas, G , Alfonso, C , Arias‐Palomo, E , et al (2010) Biochemical characterization of the transcriptional regulator BzdR from Azoarcus sp. CIB. J Biol Chem 285: 35694–35705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Egland, PG , Pelletier, DA , Dispensa, M , Gibson, J , and Harwood, CS (1997) A cluster of bacterial genes for anaerobic benzene ring biodegradation. Proc Natl Acad Sci USA 94: 6484–6489. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Eugenio, LI , Escapa, IF , Morales, V , Dinjaski, N , Galán, B , García, JL , and Prieto, MA (2010) The turnover of medium‐chain‐length polyhydroxyalkanoates in Pseudomonas putida KT2442 and the fundamental role of PhaZ depolymerase for the metabolic balance. Environ Microbiol 12: 207–221. [DOI] [PubMed] [Google Scholar]
- Fernández, H , Prandoni, N , Fernández‐Pascual, M , Fajardo, S , Morcillo, C , Díaz, E , and Carmona, M (2014) Azoarcus sp. CIB, an anaerobic biodegrader of aromatic compounds shows an endophytic lifestyle. PLoS ONE 9: e110771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fernández‐Llamosas, H , Castro, L , Blázquez, ML , Díaz, E , and Carmona, M (2016) Biosynthesis of selenium nanoparticles by Azoarcus sp CIB. Microb Cell Fact 15: 109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fuchs, G , Boll, M , and Heider, J (2011) Microbial degradation of aromatic compounds – from one strategy to four. Nat Rev Microbiol 9: 803–816. [DOI] [PubMed] [Google Scholar]
- Heider, J , Szaleniec, M , Martins, BM , Seyhan, D , Buckel, W , and Golding, BT (2016) Structure and function of benzylsuccinate synthase and related fumarate‐adding glycyl radical enzymes. J Mol Microbiol Biotechnol 26: 29–44. [DOI] [PubMed] [Google Scholar]
- Hirakawa, H , Hirakawa, Y , Greenberg, EP , and Harwood, CS (2015) BadR and BadM proteins transcriptionally regulate two operons needed for anaerobic benzoate degradation by Rhodopseudomonas palustris . Appl Environ Microbiol 81: 4253–4262. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holmes, DE , Risso, C , Smith, JA , and Lovley, DR (2012) Genome‐scale analysis of anaerobic benzoate and phenol metabolism in the hyperthermophilic archaeon Ferroglobus placidus . ISME J 6: 146–157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hori, K , Kobayashi, A , Ikeda, H , and Unno, H (2009) Rhodococcus aeterivorans IAR1, a new bacterial strain synthesizing poly (3‐hydroxybutyrate‐co‐3‐hydroxyvalerate) from toluene. J Biosci Bioeng 107: 145–150. [DOI] [PubMed] [Google Scholar]
- Johnson, CW , Salvachúa, D , Khanna, P , Smith, H , Peterson, DJ , and Beckham, GT (2016) Enhancing muconic acid production from glucose and lignin‐derived aromatic compounds via increased protocatechuate decarboxylase activity. Metab Eng Commun 3: 111–119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Juárez, JF , Zamarro, MT , Eberlein, C , Boll, M , Carmona, M , and Díaz, E (2013) Characterization of the mbd cluster encoding the anaerobic 3‐methylbenzoyl‐CoA central pathway. Environ Microbiol 15: 148–166. [DOI] [PubMed] [Google Scholar]
- Keum, YS , Seo, JS , Li, QX , and Kim, JH (2008) Comparative metabolomic analysis of Sinorhizobium sp. C4 during the degradation of phenanthrene. Appl Microbiol Biotechnol 80: 863–872. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kosa, M , and Ragauskas, AJ (2012) Bioconversion of lignin model compounds with oleaginous Rhodococci . Appl Microbiol Biotechnol 93: 891–900. [DOI] [PubMed] [Google Scholar]
- López‐Barragán, MJ , Carmona, M , Zamarro, MT , Thiele, B , Boll, M , Fuchs, G , et al (2004) The bzd gene cluster, coding for anaerobic benzoate catabolism, in Azoarcus sp. strain CIB. J Bacteriol 186: 5762–5774. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Lorenzo, V , and Timmis, KN (1994) Analysis and construction of stable phenotypes in gram‐negative bacteria with Tn5‐ and Tn10‐derived minitransposons. Methods Enzymol 235: 386–405. [DOI] [PubMed] [Google Scholar]
- Madbouly, SA , Schrader, JA , Srinivasan, G , Liu, K , McCabe, KG , Grewell, D , et al (2014) Biodegradation behavior of bacterial‐based polyhydroxyalkanoate (PHA) and DDGS composites. Green Chem 16: 1911–1920. [Google Scholar]
- Martín‐Moldes, Z , Zamarro, MT , del Cerro, C , Valencia, A , Gómez, MJ , Arcas, A , et al (2015) Whole‐genome analysis of Azoarcus sp. strain CIB provides genetic insights to its different lifestyles and predicts novel metabolic features. Syst Appl Microbiol 38: 462–471. [DOI] [PubMed] [Google Scholar]
- Moreno‐Ruiz, E , Hernáez, MJ , Martínez‐Pérez, O , and Santero, E (2003) Identification and functional characterization of Sphingomonas macrogolitabida strain TFA genes involved in the first two steps of the tetralin catabolic pathway. J Bacteriol 185: 2026–2030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Narancic, T , Kenny, ST , Djokic, L , Vasiljevic, B , O′Connor, KE and Nikodinovic‐Runic, J (2012) Medium‐chain‐length polyhydroxyalkanoate production by newly isolated Pseudomonas sp. TN301 from a wide range of polyaromatic and monoaromatic hydrocarbons. J Appl Microbiol 113, 508–520. [DOI] [PubMed] [Google Scholar]
- Ni, YY , Kim, DY , Chung, MG , Lee, SH , Park, HY , and Rhee, YH (2010) Biosynthesis of medium‐chain‐length poly(3‐hydroxyalkanoates) by volatile aromatic hydrocarbons‐degrading Pseudomonas fulva TY16. Bioresour Technol 101: 8485–8488. [DOI] [PubMed] [Google Scholar]
- Nikodinovic, J , Kenny, ST , Babu, RP , Woods, T , Blau, WJ and O′Connor, KE (2008) The conversion of BTEX compounds by single and defined mixed cultures to medium‐chain‐length polyhydroxyalkanoate. Appl Microbiol Biotechnol 80, 665–673. [DOI] [PubMed] [Google Scholar]
- Nikodinovic‐Runic, J , Guzik, M , Kenny, ST , Babu, R , Werker, A and O′Connor, KE (2013) Carbon‐rich wastes as feedstocks for biodegradable polymer (polyhydroxyalkanoate) production using bacteria. Adv Appl Microbiol 84, 139–200. [DOI] [PubMed] [Google Scholar]
- Rabus, R , Kube, M , Heider, J , Beck, A , Heitmann, K , Widdel, F , and Reinhardt, R (2005) The genome sequence of an anaerobic aromatic‐degrading denitrifying bacterium, strain EbN1. Arch Microbiol 183: 27–36. [DOI] [PubMed] [Google Scholar]
- Rabus, R , Trautwein, K , and Wöhlbrand, L (2014) Towards habitat‐oriented systems biology of “Aromatoleum aromaticum” EbN1: chemical sensing, catabolic network modulation and growth control in anaerobic aromatic compound degradation. Appl Microbiol Biotechnol 98: 3371–3388. [DOI] [PubMed] [Google Scholar]
- Rabus, R , Boll, M , Heider, J , Meckenstock, RU , Buckel, W , Einsle, O , et al (2016) Anaerobic microbial degradation of hydrocarbons: from enzymatic reactions to the environment. J Mol Microbiol Biotechnol 26: 5–28. [DOI] [PubMed] [Google Scholar]
- Rehm, BH (2010) Bacterial polymers: biosynthesis, modification and applications. Nat Rev Microbiol 8: 578–592. [DOI] [PubMed] [Google Scholar]
- Reinhold‐Hurek, B , Hurek, T , Gillis, M , Hoste, B , Vancanneyt, M , Kersters, K , and de Ley, J (1993) Azoarcus gen. nov., nitrogen‐fixing proteobacteria associated with roots of kallar grass (Leptochloa fusca (L.) Kunth), and description of two species, Azoarcus indigens sp. nov. and Azoarcus communis sp. nov. Int J Syst Bacteriol 43: 574–584. [Google Scholar]
- Sambrook, JW , and Russell, DW (2001) Molecular Cloning: A Laboratory Manual, 3rd edn Cold Spring Harbor, NY: Cold Spring Harbor Laboratory. [Google Scholar]
- Trautwein, KS , Kühner, S , Wöhlbrand, L , Halder, T , Kutcha, K , Steinbüchel, A , and Rabus, R (2008) Solvent stress response of the denitrifying bacterium “Aromatoleum aromaticum” strain EbN1. Appl Environ Microbiol 74: 2267–2274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ward, PG , Roo, G and O′Connor, KE (2005) Accumulation of polyhydroxyalkanoate from styrene and phenylacetic acid by Pseudomonas putida CA‐3. Appl Environ Microbiol 71, 2046–2052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wierckx, N , Prieto, MA , Pomposiello, P , de Lorenzo, V , O'Connor, K , and Blank, LM (2015) Plastic waste as a novel substrate for industrial biotechnology. Microb Biotechnol 8: 900–903. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wischgoll, S , Heintz, D , Peters, F , Erxleben, A , Sarnighausen, E , Reski, R , et al (2005) Gene clusters involved in anaerobic benzoate degradation of Geobacter metallireducens . Mol Microbiol 58: 1238–1252. [DOI] [PubMed] [Google Scholar]
- Zamarro, MT , Martín‐Moldes, Z , and Díaz, E (2016) The ICEXTD of Azoarcus sp. CIB, an integrative and conjugative element with aerobic and anaerobic catabolic properties. Environ Microbiol 18: 5018–5031. [DOI] [PubMed] [Google Scholar]