

Abstract
Nestling birds solicit food from adults by using begging displays that appear paradoxically costly and wasteful. Theoretical work suggests that the evolution of such exuberant offspring behavior reflects parent–offspring conflict over the supply of parental investment. Originally, extravagant begging was seen as a means of psychological trickery by which offspring could wheedle additional resources from resistant parents. Subsequently, costly begging came to be viewed as the hallmark of resolved parent–offspring conflict, serving either to prevent escalated scramble competition or to enforce honest signaling. However, the theoretical assumption of costly solicitation has been called into question by the low level of energy expenditure measured empirically during begging. This finding has prompted new theoretical work that shows that begging can be cost-free and yet still resolve parent–offspring conflict. Here, I report that begging is more costly than recent work suggests. My experimental evidence from captive canaries demonstrates a marginal cost of begging through impaired growth. Furthermore, I argue that previous studies of energy expenditure during solicitation do not measure the cost of begging, as defined theoretically. More generally, my results may account for the evolution of nestling growth rates, as well as the observation that begging is typically most flamboyant in older offspring.
The evolution of exuberant nestling begging displays has been attributed to conflicts of interest over the allocation of parental investment among dependent young. In theory, optimal investment levels should differ for parents and offspring, with offspring seeking to obtain more resources than parents are prepared to supply (1, 2). Extravagant begging may have evolved originally either to facilitate offspring attempts to wheedle additional resources from resistant parents (1) or through sibling competition for limited parental resources (3). It is thought to persist today because it plays a key role in resolving parent–offspring conflict (4).
In theoretical treatments of parent–offspring conflict, costly begging generally functions to limit offspring behavior, thereby paving the way for stable conflict resolution (5–8). For example, Parker and Macnair (9) suggest that begging behavior is a form of scramble competition for resources passively doled out by parents. At equilibrium, parent–offspring conflict is resolved because costly begging prevents runaway escalation of competition. Alternatively, Godfray (5, 10) argues that begging advertises offspring need, and that parents choose offspring to provision in relation to begging intensity. According to this view, parent–offspring conflict is resolved because the high equilibrium cost of begging enforces honest signaling.
Tests of the general theoretical prediction that begging should be costly have yielded mixed results. One form of begging cost may be borne by the brood collectively as an increase in predation risk (11). For example, when taped begging calls were broadcast at artificial nests baited with eggs, more eggs were taken from noisy nests than from silent control nests (12, 13). Haskell (12) also found a marginal predation cost of begging, because fake nests that emitted calls at a higher rate were more likely to be depredated than those broadcasting calls at a lower rate. Further indirect evidence for a predation cost associated with begging vocalizations comes from a separate comparative study which found that calls were quieter in species where the risk of predation was greater (14).
In addition, chicks may suffer a personal energetic cost of exuberant begging (11), but here the experimental evidence has sparked more controversy. The usual technique is to measure oxygen consumption while chicks are begging at intensities of their own choice. For example, McCarty (15) thereby measured energetic expenditure in begging tree swallow (Tachycineta bicolor) chicks and found it to be 1.27 times the resting metabolic rate (RMR). He concluded that begging was relatively cheap. Leech and Leonard (16) likewise found expenditure on begging to be 1.28 times greater than RMR in nestling tree swallows, but concluded begging may be costly in conditions of low food availability. House wren (Troglodytes aedon) energetic expenditure also increases by a similar fraction during begging, but because other forms of exercise incur greater metabolic costs, begging was interpreted to be relatively cost-free in this species (17, 18).
These results have, in turn, spawned a new wave of theoretical work demonstrating that begging need not necessarily be costly to resolve parent–offspring conflict (3, 19, 20). There can be multiple evolutionarily stable signaling equilibria, only some of which require begging to be costly (19, 20). A characteristic of cost-free solicitation is that begging is distributed in pooling equilibria of different signal intensities, rather than the continuously variable separating equilibria predicted when begging is costly. This theoretical vision of cost-free begging may be consistent with empirical energetic measurements, but stands at odds with other evidence that begging typically varies continuously with hunger (reviewed in ref. 21, but see ref. 22). However, further theoretical work has shown that the information content of begging can be restored with sufficiently high intensities of sibling competition (23, 24).
In the past 40 years, therefore, ideas about the evolution of nestling begging displays have almost come full circle. Traditional ethologists regarded begging as a mutually cooperative, implicitly cost-free means of communicating chick hunger to parents (e.g., ref. 25). Hamilton (26) and Trivers (1) revolutionized this view with the suggestion of parent–offspring conflict and the possibility of psychologically manipulative offspring. The concept of honest signaling was restored by Godfray (10), who promoted the idea that costly begging was the hallmark of resolved parent–offspring conflict at the nest (27). The current view (20, 23, 24) abolishes even the necessity for begging costs.
In this paper, I argue that reports of the death of costly begging have been greatly exaggerated. My experiments used captive canaries (Serinus canaria), an ideal species in which to quantify the cost of begging. The return of an adult canary to the nest triggers a frenzy of nestling begging behavior. Nestlings stretch and posture, gape and call at the parent poised to feed them. Their prolonged begging bouts, as the adult empties its crop and regurgitates the contents among the brood, suggest that begging is likely to be costly in this species. Furthermore, previous experiments have established that begging functions in the manner assumed by the models that predict begging to be costly. When the brood is very hungry, begging becomes a form of scramble competition whereby offspring jostle to be closest to the feeding adult (28). When the brood has recently been fed, adults instead actively choose offspring to feed because begging reliably signals nestling hunger (28, 29).
I measured the marginal cost of begging by forcing pairs of siblings to beg for markedly different durations, but nevertheless within the range observed at the nest, to obtain the same reward. I predicted that the manipulation would have an impact on nestling growth, and I assessed its affect at three different stages of the nestling period. To assess whether a growth cost might reasonably be equated with the fitness cost described theoretically, I also examined the correlation between nestling growth and survival to independence. Finally, I investigated whether increased begging potentially incurred digestive costs by measuring the rate of fecal sac production, a known correlate of digestive efficiency (30).
Methods
Study Species.
Gloster and Fife canaries were bred at the Subdepartment of Animal Behaviour at Madingley, Cambridgeshire, United Kingdom, between March and July, 1993–1995 and 1997–2000, on a 16:8 light:dark cycle. Each pair was assigned a separate breeding cage (102 cm × 40 cm × 28 cm), which was furnished with a felt-lined plastic nestpan supported on a wooden stand, and encouraged to raise two broods. Eggs were removed on the day of laying from experimental broods and returned on the day of clutch completion, to synchronize hatching so that siblings would be similar in mass. The modal brood size was three. Breeding birds were supplied with ad libitum quantities of mixed canary seed, Nectarblend and Easisoak (all purchased from Haith's, Cleethorpes, United Kingdom), as well as cress or chick weed, grit, drinking water, and bath water.
Chick Growth Data.
I began by collecting data on daily mass gain, from hatch day (day 0) through to day 13 (n = 241 nestlings). Every evening, between 6 and 8 p.m., I weighed nestlings by using a Sartorius PT310 electronic balance. Chicks typically fledged between day 15 and 17 and were independent by day 28. Data are presented only for chicks that survived to independence.
Manipulation of Begging Costs.
I used 30 pairs of siblings matched as closely as possible in mass (difference in mass within pairs: mean = 0.025 g, variance = 0.929 g) and selected from different broods produced by 24 pairs of breeding adults. The mean mass of sibling pairs at the start of the experiment was calculated for use in later statistical analyses. Each pair of siblings was tested either on day 6 (n = 12), day 8 (n = 9), or day 10 (n = 9). Nestlings were removed from the nest, weighed, and placed in stainless steel egg cups that were insulated with nesting material. Each sibling pair was simultaneously housed in the same Brio Super Brooder IC incubator (from Southern Aviaries, Sussex, United Kingdom) at 28–32°C. They were food deprived for 40 min to induce begging, and then fed. Meals were small volumes of finely sieved Nectarblend mixed with water to a standard concentration (6.0 g Nectarblend powder with 15.0 ml warm tap water) and administered through plastic Monoject 1.0 ml syringes.
The benefits of begging were designed to be equal in each age class and treatment. Meal volume was scaled in relation to the Daily Energy Budget (DEB) estimated from mean chick mass in each age class (31), such that meal size in milliliters was 0.005⋅DEB (kJ). The meal sizes for each age class were thus: day 6, 0.15 ml; day 8, 0.20 ml; day 10, 0.22 ml.
Each sibling was assigned to one of two treatments in which begging costs were manipulated by forcing chicks to beg for different durations before they were fed. At the nest, the duration of begging bouts did not differ significantly between days 6, 8, and 10 (F2, 388 = 1.46, P = 0.23), so treatment differences in begging intensity were similar between age classes. Before feeding, one chick was induced to beg for 10 s, whereas the other had to beg for 60 s, which is within the range (7.95–65 s; mean = 19.3 s, n = 391) observed at the nest. After 20 min, and every 20 min thereafter, I raised the lid of the incubator, stimulated chicks to beg for the appropriate duration, and then fed them before closing the incubator again. The interval between feeds was chosen to mimic the rate of provisioning commonly observed both in the laboratory (unpublished data) and in the field at the nests of seed-regurgitating finches (32). After 6 h, siblings were returned to their parents, each having experienced 19 trials in their respective begging treatments. All of the chicks tested fledged successfully.
Mass Loss Attributable to Begging.
Chicks were weighed immediately before the manipulation (MS), and immediately after receiving their last meal (ME), before being returned to their nest. In addition, I weighed all meals before they were given to chicks (N) and I collected and weighed all of the fecal sacs as they were produced during the manipulation (F). From these masses, I calculated the mass loss that could be attributable only to metabolic expenditure (B) in the two treatments:
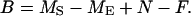 |
For each pair of siblings, I calculated B separately for the high and low begging treatments and subtracted Blow from Bhigh to derive Bdiff, a marginal cost of begging. It is possible that any differences thus calculated could be attributed merely to differences in evaporative water loss between chicks in the two treatments. To be certain that I was measuring the effect of the treatments on growth, I measured the difference between the sibling masses the evening after the experiment was completed (n = 26 sibling pairs).
Begging Intensity During the Manipulation.
Fourteen pairs of siblings were filmed during each of their 19 begging trials. I selected five consecutive trials at random for each chick and measured its postural begging intensity during each trial. From the videotape, each second I scored ordinal posture ranks, ranked in ascending order of vigor (0 = not begging; 1 = gape open; 2 = gape open, head back; 3 = as in 2 plus neck stretched; 4 = as in 3 plus back vertical) and then summed them, as is common practice in the medical and psychological literature (e.g., ref. 33). Summed posture ranks give an overall score of begging intensity known to be correlated with chick energetic expenditure in the tree swallow (16). For each chick, I summed all five measures of postural begging intensity during the experimental treatment, and then subtracted the value for chicks in the low begging treatment from their siblings in the high begging treatment to derive the marginal difference for each pair.
Statistics.
I checked whether the data met the assumptions of the statistical tests used, using Kolmogorov–Smirnov tests for deviations from normality and Bartlett's test for homogeneity of variances. The use of parametric statistics to analyze summed ordinal ranks is fully justifiable mathematically (e.g., ref. 34) and generates higher statistical power than the equivalent nonparametric test. The analysis was used merely to infer a causal relationship and not to quantify the relationship explicitly (see ref. 35). The P values reported are two-tailed.
Results
Daily Mass Gain.
In common with many other altricial species (36), canary daily mass gain peaked midway through the nestling period, at day 8, and fell off rapidly thereafter (Fig. 1). At the ages of chick testing, daily mass gain was highly correlated with mass (F1, 903 = 47.98, P < 0.0001); heavier chicks gained more weight each day.
Figure 1.

The mass gained daily by unmanipulated canary chicks during the nestling period. Filled circles indicate the days on which the cost of begging experiment was subsequently performed. Means with standard error bars are shown.
Effect of Begging Manipulation on Mass Gain.
I predicted that the impact of the begging manipulation on growth should vary in relation to daily mass gain, having the greatest effect at its peak. Given the shape of the daily mass gain curve, to assess the effect of the experimental manipulation on mass gain in relation to chick age and mean start mass, I initially fitted second order polynomial regressions and then dropped nonsignificant terms. Separate regressions were derived for Bdiff and for the difference in mass gain 24 h after the experiment because these dependent variables were correlated.
Effect on Bdiff.
There was a significant effect of age, age2, and mean start mass on Bdiff, the marginal difference between the two treatments in mass loss due to metabolic expenditure (F3, 29 = 4.75, P = 0.009). However, age and mean start mass were correlated at this stage. After fitting age and age2, mean start mass could not explain a significant amount of the variation in residual Bdiff (F1, 29 = 3.37, P = 0.08) and so was dropped from the model. Note that the resulting model yields a more conservative analysis of the data.
Together, age (t1, 29 = 2.49, P = 0.019) and age2 (t1, 29 = −2.55, P = 0.017) still explained a significant amount of variation in Bdiff (R2 = 0.21, F2, 29 = 3.50, P = 0.044; Fig. 2a). Compared with their siblings, higher begging chicks lost most weight during the experiment on day 8 (Bdiff = 0.42 ± 0.14).
Figure 2.

The marginal effect of excessive begging on mass loss due to energetic expenditure during the experiment (a) and mass gain (after controlling for start mass) during the subsequent 24 h (b), both in relation to chick age. Regression lines are shown.
Effect on difference in mass gain during the 24 h after the experiment.
Using the same procedure, only age (t1, 25 = −2.59, P = 0.016) and mean start mass (t1, 25 = 2.63, P = 0.015) significantly explained the variation in the difference between siblings in mass gain during the 24 h following the experiment (R2 = 0.27, F2, 25 = 4.27, P = 0.027). Mean start mass explained a significant amount of the variation in residual differences in mass gain (F1, 25 = 4.22, P = 0.050), after fitting age, and so was retained in the model.
Controlling for mean start mass statistically, chicks in the higher begging treatment gained less weight than their siblings, but the effect declined with increasing chick age (Fig. 2b). After controlling for age, the impact of the manipulation was greatest for heavier siblings, with potentially greater daily mass gain. These siblings showed the greatest difference in mass gain in the 24 h after the experiment.
Effect of Begging Intensity on Mass Gain.
Effect on Bdiff.
The difference in begging intensity shown by the siblings during the experiment significantly influenced Bdiff, the marginal difference in mass loss between the two treatments attributable to metabolic expenditure (F1, 13 = 4.83, P = 0.048; Fig. 3). The more chicks in the high begging treatment begged, compared with their siblings, the greater the relative mass loss that could be attributable to metabolic expenditure.
Figure 3.

The effect of increased postural begging intensity on mass loss due to energetic expenditure during the experiment.
Effect on difference in mass gain during the 24 h after the experiment.
There was no statistically significant relationship between these two variables (F1, 13 = 0.82, P = 0.386).
Effect of Nestling Growth on Offspring Survival to Independence.
I investigated whether daily mass gain was correlated with survival to independence, and how the relationship varied with chick age, using a multiple logistic regression equation (n = 241 surviving chicks, n = 86 dead nestlings with at least one current sibling surviving to independence). The greater a nestling's daily mass gain (χ2 = 76.96, P < 0.0001), the more likely it was to survive to independence. The relationship did not differ significantly between the three experimental age classes (χ2 = 2.03, P = 0.15).
Effect of Begging Intensity on Fecal Sac Production.
I assessed the effect of the manipulation on fecal sac production with a repeated measures ANOVA, with begging treatment (high or low) as the repeated measure and chick age as a between factor. Experimentally prolonging begging duration increased the number of fecal sacs produced by nestlings (F1, 27 = 5.83, P = 0.023), but this effect declined with increasing chick age (F2, 27 = 4.58, P = 0.019; Fig. 4). There was no significant effect of the manipulation on the total mass of fecal sacs produced (F1, 27 = 0.79, P = 0.381). Mean fecal sac mass was lower in the high begging treatment, but this effect also declined with chick age (F2, 27 = 4.05, P = 0.029).
Figure 4.

The interaction between chick age and begging treatment on the number of fecal sacs produced by nestlings during the experiment. Means with standard error bars are shown.
Discussion
Begging Incurs a Growth Cost.
Three lines of evidence show that begging incurs a direct growth cost in canaries. First, excessive begging retarded growth, both immediately and in the longer term, and the impact of the manipulation was greatest in chicks with the highest potential daily mass gain (Fig. 2a). Second, in older chicks that had completed most growth, the treatment had least effect on mass gain during the subsequent 24 h (Fig. 2b). Third, the greater the difference in begging intensity between siblings during the experiment, the greater the difference between them in the mass lost as a result of metabolic expenditure (Fig. 3). The growth cost of begging in canaries might reasonably be interpreted as a fitness cost, because daily mass gain was strongly correlated with the likelihood of survival to independence.
The results further suggest that begging may incur an additional indirect growth cost through its actions on digestive efficiency. Chicks that were forced to beg excessively produced a greater number of fecal sacs, although not more fecal waste in total, than their less exercised siblings. The faster rate of fecal sac production may indicate an increased digesta throughput rate, which is known to reduce digestive efficiency in other species (ref. 30; see also ref. 18).
Measuring Expenditure Has Not Measured Cost.
At first sight, the results appear to conflict with measures of energetic expenditure taken during begging in other species, which have led to the conclusion that begging incurs very low costs (15–18). However, the evidence for cheap begging stems entirely from experiments in which chicks were allowed to beg at an intensity of their own choice. Measuring expenditure in this way certainly documents the effort involved in begging, but it does not measure cost, as specified theoretically.
In the signaling models of parent–offspring conflict resolution, an individual must balance the cost and benefits of begging, and optimal signal intensity is directly related to chick condition. Only the more needy chicks can afford the higher costs of begging more intensely. Scrambling models of parent–offspring conflict resolution also require marginal begging costs, but here the tradeoff is influenced simply by the type of begging cost and the relatedness of the brood (11). The cost of begging, therefore, is not an absolute measure of effort, but a marginal measure of whether a chick can afford to spend more effort on begging. Consequently, it can be evaluated only by forcing chicks to beg for longer or shorter periods than they would themselves prefer, and then monitoring the marginal impact on a correlate of fitness (see also ref. 37). The cost of begging cannot be assessed if chicks select their own begging intensity.
In practice, applying this experimental technique to insectivorous species is likely to prove difficult. With begging bouts ranging from roughly 2–15 s in duration, there is little scope for experimental manipulation, and a high risk that experimental noise could mask any costs. Nevertheless, there may be alternative ways of testing whether there is a tradeoff between begging and growth.
In their comprehensive study of metabolic expenditure during begging by nestling house wrens, Bachmann and Chappell (17) measured expenditure at three different ages during the nestling period. The range of begging postures used by chicks in the three age classes was identical and, although the youngest chicks generally chose to beg for shorter periods than the oldest nestlings, they were capable of matching the duration of the older chick's begging displays when assuming the most vigorous begging posture.
Even so, energy expenditure on begging increased monotonically with chick mass. Because chicks in the three age groups were equally capable of begging at the same intensities, the result suggests that chicks strategically select the effort they devote to begging in relation to the growth costs they can afford to bear. Further analysis is consistent with this view. From the data Bachmann and Chappell present, the fraction of the daily energy budget devoted both to begging and to growth can be calculated. If energy for begging is traded off against energy for growth, then expenditure on begging should decrease as more of the daily energy budget is rationed to growth. Their data yield the predicted negative relationship (Fig. 5).
Figure 5.

The average proportion of the daily energy budget spent on begging by nestling house wrens in relation to the proportion of their daily energy budget spent on growth. Data were calculated from raw data presented in Table 3 in ref. 17.
Is Begging Costly Enough?
Whether the costs of begging are sufficient to prevent runaway escalation of sibling competition, or to enforce honest signaling, is a separate issue that requires an explicit experimental test. It cannot be assessed simply by reference to metabolic expenditure during other bird behaviors (e.g., ref. 15), because this gives no information about the marginal cost of intense solicitation. Even if begging proves to be sufficiently costly in canaries, it may seem unlikely that the begging costs could ever be great enough in species with shorter nest visits and consequently briefer begging displays (18). However, the higher growth rates also exhibited by these species (36) may nonetheless mean that begging could still impose a growth cost. With a much greater fraction of the daily energy budget devoted to growth, even apparently low levels of metabolic expenditure during begging could be sufficiently costly to resolve parent–offspring conflict.
A previous comparative study provides indirect support for the idea that begging growth costs may play a key role in resolving parent–offspring conflict. In general, the equilibrium cost of competing (e.g., ref. 38), or signaling (e.g., refs. 5, 10, and 39), increases with decreasing relatedness. With lower average relatedness there is a greater incentive to compete, or to be deceptive, and so a higher penalty is required for stability. Royle et al. (40) have reported that, across bird species, growth rates increase as rates of extrapair paternity go up. They interpreted the relationship as evidence of increasingly intense sibling competition to be the largest, most dominant nestling, thus capable of securing most parental resources. An equally plausible alternative view is that growth rates are scaled in relation to the growth cost required to resolve parent–offspring conflict. The faster a species grows, the greater the proportion of the daily energy budget that is devoted to growth and so the higher the marginal cost of diverting energy to begging becomes.
Why Do Older Chicks Beg So Much?
Finally, a common feature is that begging displays become increasingly flamboyant as chicks near independence. Ten days after hatching, for example, the begging displays of young canaries in the nest typically involve much more standing and wing-flapping than when chicks are 6 days old, even though younger nestlings are developmentally capable of performing such behaviors (unpublished data). In tree swallows, not surprisingly, this change in behavior with chick age is accompanied by an increase in energy expenditure during solicitation (15, 16).
Trivers (1) interpreted the vigorous displays of older juveniles as evidence of weaning conflict, with offspring attempting to solicit investment for longer than preferred by parents. Godfray (10) later suggested that the intensity of begging displays should increase as offspring condition becomes more variable, which would be the case as fledglings near independence. The results presented here offer a new explanation. Freed from the burden of devoting energy to growth, only older chicks can afford the extravagance of exuberant begging. By this stage of development, begging is energetically expensive, but virtually cost-free.
Acknowledgments
I began the work as a Junior Research Fellow at Magdalene College, Cambridge, and completed it as a Royal Society Dorothy Hodgkin Research Fellow, sponsored by the Wolfson Foundation. I am indebted to Sylvia Shelton for maintaining the canaries and I thank Tim Coulson for statistical advice, Rufus Johnstone, Janne Kotiaho, and Marty Leonard for discussion, and Nick Davies, Naomi Langmore, Kate Lessells, Doug Mock, and two anonymous referees for comments on the manuscript. I thank the Natural Environment Research Council (London) (NERC) and the Royal Society for funding the research.
Footnotes
This paper was submitted directly (Track II) to the PNAS office.
References
- 1.Trivers R. Am Zool. 1974;14:249–264. [Google Scholar]
- 2.Mock D W, Parker G A. The Evolution of Sibling Rivalry. Oxford: Oxford Univ. Press; 1997. [Google Scholar]
- 3.Rodríguez-Gironés M A, Cotton P A, Kacelnik A. Proc Natl Acad Sci USA. 1996;93:14637–14641. doi: 10.1073/pnas.93.25.14637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Godfray H C J. Nature (London) 1995;376:133–138. doi: 10.1038/376133a0. [DOI] [PubMed] [Google Scholar]
- 5.Godfray H C J. Am Nat. 1995;146:1–24. [Google Scholar]
- 6.Johnstone R A. Proc R Soc London Ser B. 1996;263:1677–1681. [Google Scholar]
- 7.Reeve H K. Proc R Soc London Ser B. 1997;264:1037–1040. [Google Scholar]
- 8.Nöldeke G, Samuelson L. J Theor Biol. 1999;197:527–539. doi: 10.1006/jtbi.1998.0893. [DOI] [PubMed] [Google Scholar]
- 9.Parker G A, Macnair M R. Anim Behav. 1979;27:1210–1235. [Google Scholar]
- 10.Godfray H C J. Nature (London) 1991;352:328–330. [Google Scholar]
- 11.Macnair M R, Parker G A. Anim Behav. 1979;27:1202–1209. [Google Scholar]
- 12.Haskell D. Proc R Soc London Ser B. 1994;257:161–164. [Google Scholar]
- 13.Leech S M, Leonard M L. Behav Ecol. 1997;8:644–646. [Google Scholar]
- 14.Briskie J V, Martin P R, Martin T E. Proc R Soc London Ser B. 1999;266:2153–2159. [Google Scholar]
- 15.McCarty J P. Auk. 1996;113:178–188. [Google Scholar]
- 16.Leech S M, Leonard M L. Proc R Soc London Ser B. 1996;263:983–987. [Google Scholar]
- 17.Bachman G C, Chappell M A. Anim Behav. 1998;55:1607–1618. doi: 10.1006/anbe.1997.0719. [DOI] [PubMed] [Google Scholar]
- 18.Chappell M A, Bachman G C. Auk. 1998;115:863–870. [Google Scholar]
- 19.Bergstrom C T, Lachmann M. Philos Trans R Soc London B. 1997;352:609–617. [Google Scholar]
- 20.Bergstrom C T, Lachmann M. Proc Natl Acad Sci USA. 1998;95:5100–5105. doi: 10.1073/pnas.95.9.5100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Kilner R, Johnstone R A. Trends Ecol Evol. 1997;12:11–15. doi: 10.1016/s0169-5347(96)10061-6. [DOI] [PubMed] [Google Scholar]
- 22.Kilner, R. M., in The Evolution of Nestling Begging: Competition, Cooperation and Communication, eds. Wright, J. & Leonard, M. L. (Kluwer, Dordrecht, The Netherlands), in press.
- 23.Johnstone R A. Proc Natl Acad Sci USA. 1999;96:12644–12649. doi: 10.1073/pnas.96.22.12644. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Rodríguez-Gironés M A. Proc R Soc London B. 1999;266:2399–2402. doi: 10.1098/rspb.1999.0937. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Armstrong E A. Bird Display and Behaviour. London: Lindsay Drummond Ltd.; 1947. [Google Scholar]
- 26.Hamilton W D. J Theor Biol. 1964;7:1–52. doi: 10.1016/0022-5193(64)90038-4. [DOI] [PubMed] [Google Scholar]
- 27.Godfray H C J, Johnstone R A. Philos Trans R Soc London B. 2000;355:1581–1591. doi: 10.1098/rstb.2000.0719. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Kilner R. Proc R Soc London Ser B. 1995;260:343–348. [Google Scholar]
- 29.Kilner R. Proc R Soc London Ser B. 1997;264:963–968. [Google Scholar]
- 30.Afik D, Karasov W H. Ecology. 1995;76:2247–2257. [Google Scholar]
- 31.Kendeigh S C, Dolnik V R, Gavrilov V M. In: Granivorous Birds in Ecosystems. Pinowski J R, Kendeigh S C, editors. Cambridge, U.K.: Cambridge Univ. Press; 1977. pp. 129–204. [Google Scholar]
- 32.Newton I. The Finches. London: Collins; 1972. [Google Scholar]
- 33.Fordham K, Stevenson-Hinde J. J Child Psychol Psychiatry. 1999;40:757–768. [PubMed] [Google Scholar]
- 34.Gaito J. Psychol Bull. 1980;87:564–567. [Google Scholar]
- 35.Stine W W. Psychol Bull. 1989;105:147–155. [Google Scholar]
- 36.Starck J M, Ricklefs R E. Avian Growth and Development. Oxford: Oxford Univ. Press; 1998. [Google Scholar]
- 37.Kotiaho, J. S., Biol. Rev., in press.
- 38.Parker G A. Anim Behav. 1985;33:519–533. [Google Scholar]
- 39.Maynard Smith J. Anim Behav. 1991;42:1034–1035. [Google Scholar]
- 40.Royle N J, Hartley I R, Owens I P F, Parker G A. Proc R Soc London Ser B. 1999;266:923–932. [Google Scholar]