

Abstract
Collisions with glass are a serious threat to avian life and are estimated to kill hundreds of millions of birds per year in the United States. We monitored 22 buildings at the Virginia Tech Corporate Research Center (VTCRC) in Blacksburg, Virginia, for collision fatalities from October 2013 through May 2015 and explored possible effects exerted by glass area and surrounding land cover on avian mortality. We documented 240 individuals representing 55 identifiable species that died due to collisions with windows at the VTCRC. The relative risk of fatal collisions at all buildings over the study period were estimated using a Bayesian hierarchical zero-inflated Poisson model adjusting for percentage of tree and lawn cover within 50 m of buildings, as well as for glass area. We found significant relationships between fatalities and surrounding lawn area (relative risk: 0.96, 95% credible interval: 0.93, 0.98) as well as glass area on buildings (RR: 1.30, 95% CI [1.05–1.65]). The model also found a moderately significant relationship between fatal collisions and the percent land cover of ornamental trees surrounding buildings (RR = 1.02, 95% CI [1.00–1.05]). Every building surveyed had at least one recorded collision death. Our findings indicate that birds collide with VTCRC windows during the summer breeding season in addition to spring and fall migration. The Ruby-throated Hummingbird (Archilochus colubris) was the most common window collision species and accounted for 10% of deaths. Though research has identified various correlates with fatal bird-window collisions, such studies rarely culminate in mitigation. We hope our study brings attention, and ultimately action, to address this significant threat to birds at the VTCRC and elsewhere.
Keywords: Windows, Collisions, Bird conservation, Birds, Advocacy
Introduction
In the United States, glass is responsible for an estimated 365–988 million bird deaths per year (Klem, 1990; Klem, 2009; Loss et al., 2014). In considering direct anthropogenic impacts to avian life in the US, the number of birds killed by glass is second only to the number killed each year (1.3–4.0 billion) by free-ranging domestic cats (Loss, Will & Marra, 2013). The literature has well documented the hazards of glass to birds (Banks, 1976; Klem, 1979; Klem, 1989; Klem, 1990; Dunn, 1993; Blem & Willis, 1998; O’Connell, 2001; Drewitt & Langston, 2008; Hager et al., 2008; Bayne, Scobie & Rawson-Clark, 2012; Machtans, Wedeles & Bayne, 2013; Hager & Craig, 2014; Kummer, Bayne & Machtans, 2016; Barton, Riding & Loss, 2017). However, studies of bird-window collision fatalities rarely lead to mitigating hazards.
Prior research has revealed several factors that influence avian mortality at windows and suggests that collisions can result from the interaction of these variables. Bird strikes are shown to increase in spring and fall when many birds are migrating (Borden et al., 2010; Ocampo-Peñuela et al., 2016), though year-round data is scant (Kummer, Bayne & Machtans, 2016). Building characteristics, such as the amount of glass, affect the likelihood that a collision will occur (Collins & Horn, 2008; Hager et al., 2013). The majority of fatal collisions in the US occur at low-rise buildings (Loss et al., 2014) and the positive correlation between building size and mortality is particularly strong in regions of low urbanization (Hager et al., 2017). Landscape features around buildings can affect collision rate, with extensive vegetation drawing birds in and reflecting in windows (Klem, 2009; Hager et al., 2008). Greenery mirrored by windows presents a false image of suitable habitat to birds that fly towards it, unable to distinguish the image from reality. Nocturnal light emitted by buildings attracts and disorients migrating birds (Ogden, 1996) and collisions typically peak during the morning and early afternoon hours (Klem, 1989; Kahle, Flannery & Dumbacher, 2016). Particularly vulnerable taxonomic families include Parulidae, Turdidae, and Emberizidae and long-distant migrants are generally at greater risk (Arnold & Zink, 2011). At the species level, disproportionate colliders include Ruby-throated Hummingbird, Black-throated Blue Warbler (Setophaga caerulescens), Ovenbird (Seiurus aurocapilla) Brown Creeper (Certhia Americana), Swamp Sparrow (Melospiza georgiana), and White-throated Sparrow (Zonotrichia albicollis) (Ogden, 1996; Hager et al., 2008; Borden et al., 2010; Loss et al., 2014).
Despite the recent research and attention documenting bird-window collisions at Duke University (Ocampo-Peñuela et al., 2016) and the Minnesota Vikings Stadium (Grant, 2014), building managers are often reluctant to take precautionary measures to reduce the threat of windows and glass to birds. Various deterrent materials are available for making windows and glass bird-friendly (e.g., window films, UV liquid, hanging cords), but large-scale implementation is lacking. Studies that evaluate the effectiveness of these products on a large scale are needed. Current legislation aimed at reducing bird-window collisions is insufficient. Only one state, Minnesota, and a few cities across the US, have mandated building standards for preventing bird strikes (American Bird Conservancy, 2012). The Federal Bird-Safe Buildings Act (H.R. 2280), introduced in May 2015, would require bird-friendly design standards on buildings newly constructed, acquired, or substantially renovated, and if passed would apply only to public buildings.
This study was initiated after a bird flew into the office window of R Schneider at the Virginia Tech Corporate Research Center (VTCRC) in mid-September 2013. VTCRC management granted permission to carry out the study and surveys began on October 19, 2013. The purpose of our research was to investigate all buildings at the VTCRC to determine the extent of bird-window collisions and examine possible effects of season, building characteristics, and surrounding landscape. We expected to observe an increase in fatal collisions at buildings with a greater surface area of glass, near extensive vegetation, and during spring and fall migration. Surveys were conducted year-round due to the lack of data gathered outside of the spring and fall migration (C Sheppard, pers. comm., 2013). If particularly hazardous buildings at the VTCRC were identified, we sought to apply and assess the effectiveness of collision deterrents on windows.
Methods
Study area
The VTCRC is located in Blacksburg, Virginia, United States (Fig. 1). At the time of the study, the office park consisted of 28 buildings on approximately 230 acres. The first building was constructed in 1988 and future plans include the construction of an additional 16 buildings (VTCRC, 2016). Buildings were assigned identification numbers that correspond with a pre-existing VTCRC building map (VTCRC, 2016). The majority of the buildings are two-story and only two buildings in the office park (buildings 12 and 17) have three stories. There is one glass walkway connecting buildings 3 and 14. Two man-made ponds are located in the VTCRC, the largest of which is located adjacent to buildings 20 and 21. Scattered ornamental trees are present throughout the office park, with larger forested patches on the western boundary and northeast corner of the property. Highly reflective and mirror-like windows comprise the majority of windows at the VTCRC (Fig. 2). Twenty-two buildings were monitored when schedules and weather allowed. The remaining six buildings were not surveyed due to ownership, or because the type of work performed in the buildings prohibited surveys.
Figure 1. VTCRC map.

Buildings surveyed for bird-window collisions at the VTCRC, Blacksburg, VA. Background image source USDA NAIP 2014.
Figure 2. VTCRC windows.

Typical highly reflective windows at the VTCRC, Blacksburg, VA (A) Building 2 (B) Building 15.
Collision surveys
We conducted year-round surveys opportunistically from October 19, 2013 until May 27, 2015. Methods were adapted from Hager & Cosentino (2014a) and Hager & Cosentino (2014b). A group of volunteers, comprised mostly of undergraduate students from Virginia Tech, carried out the surveys documenting all evidence of bird-window collisions. No carcasses were removed or collected; only photos were taken to document evidence of bird-window collisions (e.g., carcasses, feather piles). Each surveyor made one pass, walking slowly, around the perimeter of each building. The search area around each building was the width of the surveyor’s arms (approximately 2 m) held out horizontally from the building wall. Common substrates surveyed included lawn, pavement, and mulch. Surveyors also searched on top, behind, and inside of the shrubs next to the buildings. In addition to species information, date, start survey time, end survey time, building number, and building side were recorded. Surveys were not conducted during a specific time of day or in adverse weather conditions. Data was reported by volunteers every day or at minimum, once a week. We compared all findings, by photo or in the field, to previous findings to avoid double counting.
Data measurement and analysis
Land cover was classified within a 50 m buffer for each of the 22 buildings surveyed. Virginia Geographic Information Network (VGIN) data was used to classify surrounding land cover to the following classes: forest, lawn (including pasture and turfgrass), impervious, water, and tree. To better classify scattered ornamental trees present at the VTCRC, the ‘tree’ land cover class was used to distinguish between sparse tree coverage and forest. We estimated glass area for each building using the program ImageJ (Rasband, 2016) following methods detailed by Hager & Cosentino (2014b). Glass area was centered at 0 and scaled when we used it our Bayesian regression model (Supplemental Information). Meteorological seasons were used to report seasonality of our results. Fall was defined as September–November, winter as December–February, spring as March–May, and summer as June–August.
Linear regression analyses were performed in the program R Core Team (2017), with the RStudio Team (2015), to explore possible relationships between the number of bird deaths per building and predictor variables, glass area and surrounding land cover. We relied on negative binomial regression for this analysis because Poisson distribution led to overdispersion in the data. In order to compare the number of bird deaths across buildings despite uneven survey effort, Poisson regression was performed with an offset for the number of observations. These results informed the Bayesian hierarchical regression model we used to estimate the relative risk of fatal collisions at the 22 buildings across the 20 months. We assumed a zero-inflated Poisson (ZIP) distribution for the number of fatal collisions observed at each building within each month. A ZIP model is common for handling excess zero counts in data and assumes these zeroes are generated by a process separate than the count values (Lambert, 1992). The model contains two components, a binomial distribution for the excess zeroes and a Poisson distribution for the count values (which may contain zeroes). The Poisson distribution assumes a mean of the product of the relative risk θit and the expected count eit at building i in month t. The expected counts are the product of the overall fatal collision rate rt in each month and the number of site observations at each building in each month.
We modeled the log of the relative risk θit for building i at month t as:
| (1) |
where δt is a temporal random effect. The parameter β1 is the log relative risk associated with a percent increase in the tree area within 50 m of a building and, similarly, β2 is the log relative risk associated with a percent increase in lawn area while β3 is the effect of a standard deviation increase in glass area on a building (SD = 175.63 m2). We chose these variables based on known relationships and significant relationships found in the exploratory analyses. The temporal term δt captured variation in the collisions across time not explained by the variables.
For the binomial distribution, we specified the probability of excess zeroes at building i as:
| (2) |
where α1 is the increase in the log-odds of an excess zero for each percent increase in tree area, α2 is the change in the log-odds for each percentage increase in lawn area, and α3 is the increase in log-odds associated for each 175.63 m2 of glass area on a building.
Prior distributions were required to complete the Bayesian hierarchical model. All parameters for both the Poisson and binomial model were given non-informative normal priors centered at 0 with variance 10 and the temporal term δt followed a random walk prior of order one. This prior allows each δt term to be correlated with the previous δt−1 term with variance 1,000 and assumes that fatal collisions are more associated in consecutive months than collisions in non-consecutive months.
We estimated the model parameters using Markov chain Monte Carlo (MCMC) in WinBUGS (Spiegelhalter et al., 2002) (Supplemental Information). We ran one MCMC chain for 30,000 iterations with a burn-in of 15,000. The remaining sample was thinned to every 3 iterations, yielding a final posterior sample of size 5,000 for computing posterior summaries. We assessed convergence through visual inspection of trace and density plots for each parameter and Geweke’s diagnostic (Geweke, 1992). For parameter estimates, we sampled from the joint posterior distribution and report the mean posterior estimates and the 95% credible intervals on the relative risk scale. We also plotted the estimated relative risks for each building month to month.
Results
We documented a total of 240 collision deaths representing 55 identifiable species at the VTCRC during the course of the study (Table 1). Forty-one (17%) of the bird deaths were unidentifiable remains. The average number of surveys per building for the study period was 136. All 22 buildings surveyed had at least one recorded collision death. Of the 298 total survey days, 47 occurred on days in 2013, 198 days in 2014, and 53 in 2015. We conducted 92 surveys in the fall, 75 in the winter, 96 in the spring, and 35 in the summer.
Table 1. Bird-window collision numbers.
Species observed as window collision deaths at the VTCRC (October 2013–May 2015).
| Family | Common name | Scientific name | Fall (9/1-11/30) | Winter (12/1-2/28) | Spring (3/1-5/31) | Summer (6/1-8/31) | Total collisions |
|---|---|---|---|---|---|---|---|
| Charadriidae | Killdeer | Charadrius vociferus | – | – | 1 | – | 1 |
| Scolopacidae | American Woodcocka | Scolopax minor | – | 1 | – | – | 1 |
| Columbidae | Mourning Dove | Zenaida macroura | 5 | 2 | 4 | 3 | 14 |
| Cuculidae | Yellow-billed Cuckooa | Coccyzus americanus | 1 | – | 4 | – | 5 |
| Caprimulgidae | Common Nighthawk | Chordeiles minor | 1 | – | – | – | 1 |
| Trochilidae | Ruby-throated Hummingbird | Archilochus colubris | 15 | – | 7 | 2 | 24 |
| Tyrannidae | Great Crested Flycatcher | Myiarchus crinitus | – | – | – | 1 | 1 |
| Tyrannidae | Eastern Kingbirda | Tyrannus tyrannus | – | – | 2 | – | 2 |
| Picidae | Northern Flicker | Colaptes auratus | – | – | 2 | 2 | 4 |
| Picidae | Yellow-bellied Sapsucker | Sphyrapicus varius | 1 | 1 | – | – | 2 |
| Picidae | Unknown Woodpecker | Woodpecker spp. | 1 | – | – | – | 1 |
| Vireonidae | Red-eyed Vireo | Vireo olivaceus | 1 | – | – | – | 1 |
| Vireonidae | Blue-headed Vireo | Vireo solitarius | 1 | – | – | – | 1 |
| Vireonidae | Yellow-throated Vireo | Vireo flavifrons | – | – | – | 1 | 1 |
| Paridae | Carolina Chickadee | Poecile carolinensis | 1 | – | – | – | 1 |
| Paridae | Tufted Titmouse | Baeolophus bicolor | 1 | – | – | – | 1 |
| Sittidae | White-breasted Nuthatch | Sitta carolinensis | 1 | – | – | – | 1 |
| Certhiidae | Brown Creeper | Certhia americana | 1 | – | – | – | 1 |
| Regulidae | Golden-crowned Kinglet | Regulus satrapa | 7 | 2 | – | – | 9 |
| Regulidae | Ruby-crowned Kinglet | Regulus calendula | – | 1 | 1 | – | 2 |
| Turdidae | American Robin | Turdus migratorius | 2 | – | 11 | 8 | 21 |
| Turdidae | Hermit Thrush | Catharus guttatus | 3 | – | 1 | – | 4 |
| Turdidae | Unknown Thrush | Thrush spp. | 1 | – | – | – | 1 |
| Turdidae | Eastern Bluebird | Sialia sialis | – | – | 2 | – | 2 |
| Mimidae | Brown Thrasher | Toxostoma rufum | – | – | 2 | – | 2 |
| Mimidae | Gray Catbirda | Dumetella carolinensis | 2 | – | 10 | 3 | 15 |
| Mimidae | Northern Mockingbird | Mimus polyglottos | 1 | – | – | 1 | 2 |
| Sturnidae | European Starling | Sturnus vulgaris | – | – | 2 | – | 2 |
| Motacillidae | American Pipit | Anthus rubescens | – | 1 | – | – | 1 |
| Bombycillidae | Cedar Waxwing | Bombycilla cedrorum | 2 | – | 3 | 2 | 7 |
| Parulidae | American Redstart | Setophaga ruticilla | – | – | 1 | – | 1 |
| Parulidae | Black-throated Blue Warbler | Setophaga caerulescens | 1 | – | – | – | 1 |
| Parulidae | Black-throated Green Warbler | Setophaga virens | 2 | – | – | – | 2 |
| Parulidae | Chestnut-sided Warbler | Setophaga pensylvanica | 1 | – | – | – | 1 |
| Parulidae | Common Yellowthroat | Geothlypis trichas | 2 | – | – | 1 | 3 |
| Parulidae | Ovenbird | Seiurus aurocapilla | – | – | – | 3 | 3 |
| Parulidae | Yellow-rumped Warbler | Setophaga coronata | – | – | 1 | – | 1 |
| Parulidae | Louisiana Waterthrush | Parkesia motacilla | – | – | – | 1 | 1 |
| Parulidae | Northern Waterthrush | Parkesia noveboracensis | 1 | – | – | – | 1 |
| Parulidae | Palm Warbler | Setophaga palmarum | 1 | – | – | – | 1 |
| Parulidae | Tennessee Warbler | Oreothlypis peregrina | 1 | – | – | – | 1 |
| Parulidae | Worm-eating Warbler | Helmitheros vermivorum | – | – | 1 | – | 1 |
| Parulidae | Yellow Warbler | Setophaga petechia | 1 | – | – | – | 1 |
| Emberizidae | Dark-eyed Junco | Junco hyemalis | 3 | 2 | 4 | – | 9 |
| Emberizidae | Eastern Towheea | Pipilo erythrophthalmus | 1 | 1 | – | – | 2 |
| Emberizidae | Field Sparrowa | Spizella pusilla | – | – | – | 1 | 1 |
| Emberizidae | Fox Sparrow | Passerella iliaca | 1 | – | – | – | 1 |
| Emberizidae | Song Sparrow | Melospiza melodia | 1 | – | 2 | 3 | 6 |
| Emberizidae | White-throated Sparrow | Zonotrichia albicollis | 5 | 2 | 2 | – | 9 |
| Emberizidae | White-crowned Sparrow | Zonotrichia leucophrys | – | 1 | – | – | 1 |
| Emberizidae | Savannah Sparrow | Passerculus sandwichensis | – | – | 1 | – | 1 |
| Cardinalidae | Indigo Bunting | Passerina cyanea | 1 | – | 2 | – | 3 |
| Cardinalidae | Northern Cardinal | Cardinalis cardinalis | 1 | 1 | 1 | 2 | 5 |
| Cardinalidae | Scarlet Tanager | Piranga olivacea | 1 | – | – | – | 1 |
| Icteridae | Red-winged Blackbird | Agelaius phoeniceus | – | – | 1 | 2 | 3 |
| Fringillidae | American Goldfinch | Spinus tristis | 1 | – | 4 | – | 5 |
| Fringillidae | House Finch | Haemorhous mexicanus | 1 | – | – | 2 | 3 |
| Unknown | Unidentified | 16 | 2 | 11 | 12 | 41 | |
| TOTAL | 90 | 17 | 83 | 50 | 240 |
Notes.
Virginia Species of Greatest Conservation Need (VDGIF 2015).
Our model converged based on visual inspection of trace and density plots for the parameters and according to Geweke’s Diagnostics (Table 2). Overall, we found significant effects for percent lawn area within 50 m of a building and glass area on a building. Specifically, we found a significantly increased relative risk of 1.30 (95% CRI [1.05–1.65]) for fatal collisions for each 175.6 m2 of additional glass area on a building; and a significantly reduced risk of 0.96 (95% CI [0.93–0.98]) for collisions for each percent increase in lawn area within 50 m of a building. We also found a marginally significant relationship between collisions and the percent area of trees within 50 m of a building. However, we found no significant relationships between the variables and the log-odds of no fatal collision (the excess zeroes in the model). We also observed decreased relative risk of collisions between December and March when we plotted the relative risks (Fig. 3).
Table 2. Relative risk.
Posterior relative risk and log-odds estimates for fatal collisions. The β parameters are on the relative risk scale, and the α parameters are on the log-odds scale. A parameter is considered significant if the credible interval does not contain 1. Geweke’s Diagnostic should be between −1.96 and 1.96 for a parameter to be considered converged.
| Effect | Posterior mean | 95% credible interval | Geweke’s diagnostic |
|---|---|---|---|
| PercentTree, β1 | 1.02 | 1.00, 1.05 | 0.23 |
| PercentLawn, β2 | 0.96 | 0.93, 0.98 | −0.53 |
| GlassArea, β3 | 1.30 | 1.05, 1.65 | −1.29 |
| PercentTree, α1 | 0.99 | 0.95, 1.03 | −0.54 |
| PercentLawn, α2 | 0.99 | 0.97, 1.00 | 0.53 |
| GlassArea, α3 | 0.85 | 0.56, 1.32 | −1.48 |
Figure 3. Relative risk.
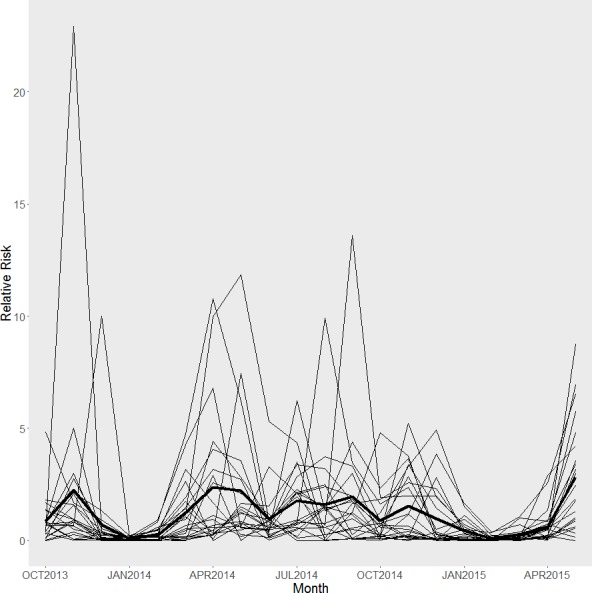
Estimated relative risk of fatal collisions for each building at each month. The thicker black line represents the average relative risk across time.
Glass area and surrounding land cover
Seven buildings were responsible for the majority (57.1%) of window collision deaths (Fig. 4). Buildings 11 and 12 had the highest avian mortality and accounted for a quarter (24.6%) of all documented collision deaths. Although we were unable to obtain window specifications on all buildings to make quantitative comparisons of reflectivity and glass type, observations from our surveys suggest that window reflectivity plays a large role in the frequency of collisions at the VTCRC. The amount of glass on buildings ranges from 18 m2 (building 8) to 693 m2 (building 9). Based on our model, this feature played a significant role in fatal collisions. Building 8, with the least amount of glass, had only one recorded collision death. The two buildings that killed the most birds, 11 and 12, have an estimated 451 m2 and 492 m2 of glass, respectively. Buildings 1 and 2 are almost identical buildings located adjacent to each other. Building 1, with 219 m2 glass area, killed 15 birds; building 2, with 191 m2 glass area, killed nine. A notable difference between the two buildings is the amount of forest area (840 m2) on one side of building 1, while building 2 has no surrounding forest area.
Figure 4. Collisions by building.
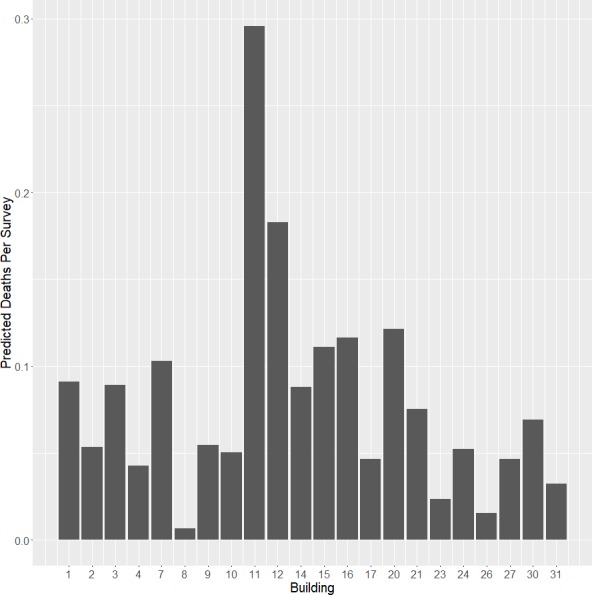
Mean number of collision deaths per survey for each building surveyed at the VTCRC (October 2013–May 2015).
Impervious area (51%) consisting of parking lots, sidewalks, and roads dominate the surrounding 50 m of all buildings in the VTCRC. Lawn (35%), tree (10%), forest (3%), and open water (1%) comprise the remaining area. Our model indicated that lawn area mitigated the relative risk of fatal collisions while tree area was associated with a slight increase in risk of collision. Building 11 has the highest percentage of surrounding forest cover (25.2%), while building 12 has no surrounding forest area. Building 11 also has the least amount of lawn area compared to the other buildings.
Seasonality
July, May, and September had the highest mortality respectively when correcting for uneven survey effort (Fig. 5). Collision deaths were lowest in the winter months. The species with the most collision deaths in the fall were Ruby-throated Hummingbird (n = 15), Golden-crowned Kinglet (Regulus satrapa) (n = 7), Mourning Dove (Zenaida macroura) (n = 5), and White-throated Sparrow (n = 5). In the winter, White-throated Sparrow (n = 2), Mourning Dove (n = 2), Golden-crowned Kinglet (n = 2), and Dark-eyed Junco (Junco hyemalis) (n = 2) were the most frequent colliders. American Robin (Turdus migratorius) (n = 11), Gray Catbird (Dumetella carolinensis) (n = 10), and Ruby-throated Hummingbird (n = 7) dominated in the spring. In the summer, American Robin (n = 8), Mourning Dove (n = 3), Gray Catbird (n = 3), Ovenbird (n = 3), and Song Sparrow (Melospiza melodia) (n = 3) were most frequently detected.
Figure 5. Collisions by month.
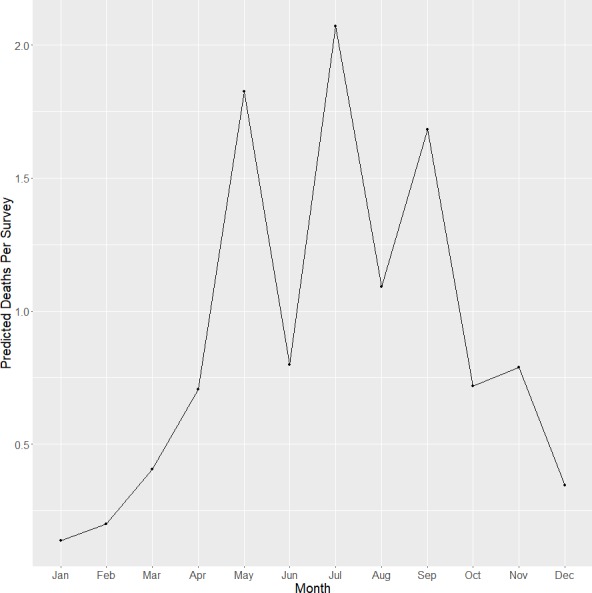
Mean number of collision deaths per survey for each month at the VTCRC (October 2013–May 2015). Months were combined across years.
Species
The Ruby-throated Hummingbird was the most common window collision species and accounted for 10% of all deaths recorded during our entire study. The majority (29.2%) of hummingbird deaths were at building 11. The next three most common species, American Robin, Gray Catbird, and Mourning Dove, accounted for 21% of identifiable deaths combined. Nine of the 55 species we documented are disproportionately vulnerable to collisions at all building types (Loss et al., 2014). Six Virginia Species of Greatest Conservation Need (Virginia Department of Game and Inland Fisheries, 2015) were documented as window kills, including American Woodcock (Scolopax minor), Eastern Towhee (Pipilo erythrophthalmus), and Field Sparrow (Spizella pusilla) (Table 1).
The most common families were Emberizidae (13% of collision deaths), Turdidae (12%), Trochilidae (10%), Mimidae (8%) and Parulidae (8%). Emberizidae collisions were mostly Dark-eyed Junco (n = 9) and White-throated Sparrow (n = 9). American Robin (n = 21) represented the majority of Turdidae collisions. Ruby-throated Hummingbird was the only species within Trochilidae. Within Mimidae, Gray Catbird (n = 15) was the most common species. Thirteen species comprised Parulidae, with Common Yellowthroat (Geothlypis trichas) and Ovenbird (n = 3) comprising most of the collisions.
Discussion
Our documentation of 240 birds over 20 months suggest that avian fatalities are a significant problem at the VTCRC. We found collision victims year-round because the reflective windows did not present a barrier to avoid. Results of our study indicate a positive correlation between the number of fatal collisions and glass area. Glass or window area is one of the primary factors affecting collisions as previous studies have shown (Klem et al., 2009; Borden et al., 2010; Hager et al., 2013; Cusa, Jackson & Mesure, 2015; Ocampo-Peñuela et al., 2016) and our findings provide additional evidence for this. In addition to glass area, we speculate that collision frequency at the VTCRC buildings is related to the configuration of glass on building façades. Several buildings (e.g., 12, 11, and 7) contain a large portion of glass concentrated in one area. Additionally, many of the buildings in this office park have darkly tinted, though nonetheless reflective, windows. The buildings that experienced a high number of collisions, including building 11, exhibit a mirror-like reflection of the immediate environment that can confuse birds by projecting a false image of habitat. Building 11 also had the highest amount of surrounding forest cover, a factor that may have contributed to a higher number of collisions. Measuring the degree of reflectivity, a trait that can vary by type of glass, cloud cover, angle of the sun, time of day, and by façade, proved to be beyond the scope of our study.
In our analyses of land cover characteristics, lawn area was negatively correlated with the number of bird fatalities in both our exploratory linear regression and the reported Bayesian model. This finding may be attributed to these areas attracting fewer birds than more heavily vegetated, resource-laden areas in the vicinity. Small sample size limited our ability to fully explore the relationship between nearby vegetation and the number of fatal collisions. Of the 22 buildings surveyed, only six had forest cover within 50 m; therefore, we were unable to quantify possible effects of this variable. While the amount of impervious surface in the vicinity of buildings was not significantly related to collisions, ornamental trees were found to be marginally significant to the number of collisions we recorded. Our analyses did not take into account potential effects of variables such as building size, window reflectivity, nocturnal lighting, and local avian abundance, all of which would be pertinent focuses of future research at this site (Wittig et al., 2017).
Our results indicate that birds strike windows year-round at the VTCRC. Mitigating efforts should be implemented across the breeding season and both spring and fall migration to be most effective. Although we documented the highest number of fatalities during May and September, July had the most bird deaths per survey. The number of collisions in July is notable because the majority of related studies focus survey efforts during the spring and fall migration and little research is conducted during the breeding season (O’Connell, 2001; Gelb & Delacretaz, 2006; Hager et al., 2008; Klem et al., 2009; Borden et al., 2010; Bayne, Scobie & Rawson-Clark, 2012; Loss et al., 2014). Bird deaths in July are likely due to the number of newly fledged birds that are unfamiliar with the surrounding landscape (Hager, 2014; Kahle, Flannery & Dumbacher, 2016). Similar to Hager & Craig (2014), the species most frequently detected during summer was the American Robin.
Consistent with other studies showing their tendency to be frequent colliders (Klem, 1989; Hager & Craig, 2014; Loss et al., 2014), we found that Ruby-throated Hummingbirds fatally struck windows more often than any other species. Hummingbirds may be at increased risk of collision compared with other species because of their small size and delicate stature, as well as their high flight velocities (Kahle, Flannery & Dumbacher, 2016). This combination could make them less likely to survive an impact with a solid barrier. Additional research is needed to assess how relative species size and robustness correlate with strike susceptibility in hummingbirds and other species. Our results support the finding that migrants are especially prone to collisions, but suggests that resident species are also at risk. Notably, six of the species we documented are listed as Virginia Species of Greatest Conservation Need (Virginia Department of Game and Inland Fisheries, 2015), a designation signifying their decline in the state and the need for conservation action.
The actual number of bird-window collision deaths is likely higher than documented at the VTCRC (O’Connell, 2001). Predators and scavengers undoubtedly played a role in the detection of carcasses (Hager, Cosentino & McKay, 2012; Kummer et al., 2016), especially since the time interval between surveys varied. Evidence of raccoons (e.g., scat piles) was common on survey routes and the VTCRC managed a feral cat colony at the time of this study. Ground crews hindered detectability when mowing and leaf collection occurred. We did not account for searcher detection error, and we acknowledge that some carcasses likely went undetected during our study (Smallwood, 2007; Parkins, Elbin & Barnes, 2015). Detectability at buildings varied due to surrounding substrate; building 9, which contains the highest amount of glass, also has heavy ivy groundcover along the building perimeter that likely contributed to lower detectability.
Mitigation measures that appear most effective are window films or netting applied to the outside of windows (Klem, 1990; Ocampo-Peñuela et al., 2016). Due to the highly reflective quality of the windows in the VTCRC, collision deterrents must be applied to window exteriors in order to increase glass visibility. An efficient approach would target buildings shown to be most hazardous to birds, including buildings 11, 12, 20, 16, and 15. Closing interior blinds has little to no effect on the external appearance of the glass at many VTCRC windows. Due to the press this project received (Supplemental Information), we made contact with several architects to suggest that plans for VTCRC Phase II incorporate bird-window collision mitigation measures and less reflective glass. Options for new construction at the VTCRC include using less reflective glass, fritted glass, smaller windowpanes, and angling windows upon installation so they do not reflect the sky or adjacent vegetation (Klem et al., 2004).
VTCRC management can reduce the number of birds that die each year by retrofitting the most hazardous buildings to increase visibility to birds. Although we identified specific “problem” buildings that experienced a higher number of collisions, ultimately management did not grant permission when we requested to field-test collision deterrents. The reasons given for not altering windows included general aesthetics, potential damage to windows, interference with heating and cooling systems, policy, and cost. Our communications with VTCRC management ended when they stated they would follow Virginia Tech policy addressing bird-window collisions. Coincidentally, there is no Virginia Tech policy addressing avian collisions. A number of Virginia Tech offices on campus were cooperative in investigating bird-window collisions, and we conducted a small pilot study on the university campus from fall 2014 to spring of 2015 (Supplemental Information). When we requested to test mitigation measures at one building, Fralin Biotechnology Center, we were not permitted due to aesthetic reasons.
We must consider the cumulative impacts of bird-window collisions nationwide. As defined by the National Environmental Protection Act (1969), “Cumulative impacts can result from individually minor but collectively significant actions taking place over a period of time”. In the near future, we hope studies that ignite mitigation actions will become more commonplace, as with those implemented at Duke University (Ocampo-Peñuela et al., 2016). College campuses can include bird-safe buildings as a component of their sustainability campaigns. Bird-window collision studies performed on campuses provide students with an opportunity to investigate human impacts on the natural world where they live, work, and study.
While we acknowledge the overwhelming scope of bird-window collisions (Loss et al., 2014; Machtans & Thogmartin, 2014), we view the findings of our research as a call to action. To protect avian life at the VTCRC and the Virginia Tech campus in the future, testing and implementing permissible mitigating strategies should be an important component of bird-window collision research. Applying mitigation measures such as window films to the few buildings with high avian mortality would be a reasonable approach. Ultimately, a University policy on minimizing bird-window collisions would help guide future construction, encourage modification to buildings known to be particularly threatening to birds, and make the school a leader in sustainable, bird-friendly building.
Supplemental Information
Raw data.
Raw data.
Raw data.
Raw data.
Raw data.
Raw data.
Official survey and incidental reports of bird-window collisions on the Virginia Tech campus (September 2014–May 2015).
Acknowledgments
First, we thank the volunteers who made this study possible: Julia Magill, Aylett Lipford, Mary Ratliff, Pat Polentz, Catherine Kim, Katie Anderson, Katy Miller, Julie Chang, Elizabeth Fazio, Cindy Alcazar, Laura Veru, Lea Sarment, Lauren Bowman, Justin Hall, Kris Schneider, Becky Keller, Eric Hallerman, Susan Hutson, Michael St. Germain, and The Virginia Tech Student Chapter of The Wildlife Society. Many thanks to John Gerwin (North Carolina Museum of Natural Sciences) and Donald Mackler (Virginia Tech) for their valuable knowledge identifying bird remains. We thank Kevin McGuckin, Ingrid Mans, and Verl Emrick of the Conservation Management Institute for GIS analyses. Travis Belote provided valuable statistical advice and knowledge. Collaborators from the Department of Statistics at Virginia Tech, Yafei Zhang, Sierra Merkes, and Wenyu Gao supplied additional statistical guidance. Laura Lawson and two anonymous reviewers provided helpful comments on the manuscript. Lastly, we thank Dr. Daniel Klem and Dr. Christine Sheppard for their work and guidance.
Funding Statement
On 01/06/14, $1,784 was raised via Experiment.com. The majority of the funds went to purchase field guides and field supplies for volunteers. The remaining funds went to fellowships for two volunteers. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Additional Information and Declarations
Competing Interests
The authors declare there are no competing interests.
Author Contributions
Rebecca M. Schneider conceived and designed the experiments, performed the experiments, analyzed the data, prepared figures and/or tables, authored or reviewed drafts of the paper, approved the final draft.
Christine M. Barton performed the experiments, analyzed the data, prepared figures and/or tables, authored or reviewed drafts of the paper, approved the final draft.
Keith W. Zirkle analyzed the data, prepared figures and/or tables, authored or reviewed drafts of the paper, approved the final draft.
Caitlin F. Greene performed the experiments, authored or reviewed drafts of the paper, approved the final draft.
Kara B. Newman performed the experiments, authored or reviewed drafts of the paper.
Data Availability
The following information was supplied regarding data availability:
The raw data have been provided as Supplemental Files and here: Zirkle, Keith; rmurray@vt.edu (2018): Files for Schneider et al. (2018). Figshare. https://doi.org/10.6084/m9.figshare.5982394.v1.
References
- American Bird Conservancy (2012).American Bird Conservancy Legislation. 2012. http://collisions.abcbirds.org/legislation.html. [23 February 2018]. http://collisions.abcbirds.org/legislation.html
- Arnold & Zink (2011).Arnold TW, Zink RM. Collision mortality has no discernible effect on population trends of North American birds. PLOS ONE. 2011;6(9):e24708. doi: 10.1371/journal.pone.0024708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Banks (1976).Banks RC. Reflective plate glass-a hazard to migrating birds. BioScience. 1976;26(6):414. [Google Scholar]
- Barton, Riding & Loss (2017).Barton CM, Riding CS, Loss SR. Magnitude and correlates of bird collisions at glass bus shelters in an urban landscape. PLOS ONE. 2017;12(6):e0178667. doi: 10.1371/journal.pone.0178667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bayne, Scobie & Rawson-Clark (2012).Bayne EM, Scobie CA, Rawson-Clark M. Factors influencing the annual risk of bird–window collisions at residential structures in Alberta, Canada. Wildlife Research. 2012;39:583–592. doi: 10.1071/WR11179. [DOI] [Google Scholar]
- Blem & Willis (1998).Blem CR, Willis BA. Seasonal variation of human–caused mortality of birds in the Richmond area. The Raven. 1998;69(1):3–8. [Google Scholar]
- Borden et al. (2010).Borden WC, Lockhart OM, Jones AW, Lyons MS. Seasonal, taxonomic, and local habitat components of bird-window collisions on an urban university campus in Cleveland, OH. The Ohio Journal of Science. 2010;110:44–52. [Google Scholar]
- Collins & Horn (2008).Collins KA, Horn DJ. Bird-window collisions and factors influencing their frequency at Millikin University in Decatur, Illinois. 68th midwest fish and wildlife conference abstract.2008. [Google Scholar]
- Cusa, Jackson & Mesure (2015).Cusa M, Jackson DA, Mesure M. Window collisions by migratory bird species: urban geographical patterns and habitat associations. Urban Ecosystems. 2015;18(4):1427–1446. doi: 10.1007/s11252-015-0459-3. [DOI] [Google Scholar]
- Drewitt & Langston (2008).Drewitt AL, Langston RH. Collision effects of wind-power generators and other obstacles on birds. Annals of the New York Academy of Sciences. 2008;1134(1):233–266. doi: 10.1196/annals.1439.015. [DOI] [PubMed] [Google Scholar]
- Dunn (1993).Dunn EH. Bird mortality from striking residential windows in winter. Journal of Field Ornithology. 1993;64:302–309. [Google Scholar]
- Gelb & Delacretaz (2006).Gelb Y, Delacretaz N. Avian window strike mortality at an urban office building. The Kingbird. 2006;56:190–198. [Google Scholar]
- Geweke (1992).Geweke J. Evaluating the accuracy of sampling-based approaches to the calculation of posterior moments. In: Bernardo JM, Berger JO, Dawid AP, Smith AFM, editors. Bayesian statistics 4. Oxford University Press; Oxford: 1992. pp. 169–193. [Google Scholar]
- Grant (2014).Grant T. Green Monsters: examining the environmental impact of sports stadiums. Villanova Environmental Law Journal. 2014;25(1):149–176. [Google Scholar]
- Hager & Cosentino (2014a).Hager SB, Cosentino BJ. Surveying for bird carcasses resulting from window collisions: a standardized protocol. PeerJ. 2014a;2:e406v401. doi: 10.7287/peerj.preprints.406v1. [DOI] [Google Scholar]
- Hager & Cosentino (2014b).Hager SB, Cosentino BJ. Evaluating the drivers of bird-window collisions in North America/EREN bird-window collisions project. 2014b. https://sites.google.com/a/augustana.edu/eren-bird-window-collisions-project/protocols/c-measuring-windows. [19 August 2017]. https://sites.google.com/a/augustana.edu/eren-bird-window-collisions-project/protocols/c-measuring-windows
- Hager et al. (2017).Hager SB, Cosentino BJ, Aguilar-Gómez MA, Anderson ML, Bakermans M, Boves TJ, Brandes D, Butler MW, Butler EM, Cagle NL, Calderón-Parra R, Capparella AP, Chen A, Cipollini K, Conkey AA, Contreras TA, Cooper RI, Corbin CE, Curry RL, Dosch JJ, Drew MG, Dyson K, Foster C, Francis CD, Fraser E, Furbush R, Hagemeyer NDG, Hopfensperger KN, Klem D, Lago E, Lahey A, Lamp K, Lewis G, Loss SR, Machtans CS, Madosky J, Maness TJ, McKay KJ, Menke SB, Muma KB, Ocampo-Peñuela N, O’Connell TJ, Ortega-Álvarez R, Pitt AL, Puga-Caballero AL, Quinn JE, Varian-Ramos CW, Riding CS, Roth AR, Saenger PG, Schmitz RT, Schnurr J, Simmons M, Smith AD, Sokoloski DR, Vigliotti J, Walters EL, Walters LA, Weir JT, Winnett-Murray K, Withey JC, Zuria I. Continent-wide analysis of how urbanization affects bird-window collision mortality in North America. Biological Conservation. 2017;212(A):209–215. doi: 10.1016/j.biocon.2017.06.014. [DOI] [Google Scholar]
- Hager, Cosentino & McKay (2012).Hager SB, Cosentino BJ, McKay KJ. Scavenging affects persistence of avian carcasses resulting from window collisions in an urban landscape. Journal of Field Ornithology. 2012;83:203–211. doi: 10.1111/j.1557-9263.2012.00370.x. [DOI] [Google Scholar]
- Hager et al. (2013).Hager SB, Cosentino BJ, McKay KJ, Monson C, Zuurdeeg W, Blevins B. Window area and development drive spatial variation in bird-window collisions in an urban landscape. PLOS ONE. 2013;8(1):e53371. doi: 10.1371/journal.pone.0053371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hager & Craig (2014).Hager SB, Craig ME. Bird-window collisions in the summer breeding season. PeerJ. 2014;2:e460. doi: 10.7717/peerj.460. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hager et al. (2008).Hager SB, Trudell H, McKay KJ, Crandall SM, Mayer L. Bird density and mortality at windows. Wilson Journal Ornithology. 2008;120:550–564. doi: 10.1676/07-075.1. [DOI] [Google Scholar]
- Kahle, Flannery & Dumbacher (2016).Kahle LQ, Flannery ME, Dumbacher JP. Bird-window collisions at a West-Coast Urban Park Museum: analyses of bird biology and window attributes from Golden Gate Park, San Francisco. PLOS ONE. 2016;11(1):e0144600. doi: 10.1371/journal.pone.0144600. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klem (1979).Klem D. Ph.D dissertation. 1979. Biology of collisions between birds and windows. [Google Scholar]
- Klem (1989).Klem D. Bird-window collisions. Wilson Bulletin. 1989;101:606–620. [Google Scholar]
- Klem (1990).Klem D. Collisions between birds and windows: mortality and prevention. Journal of Field Ornithology. 1990;61:120–128. [Google Scholar]
- Klem (2009).Klem D. Avian mortality at windows: the second largest human source of bird mortality on earth. In: Rich TD, Arizmendi C, Demarest D, Thompson C, editors. Tundra to tropics: proceedings of the fourth international partners in flight conference; McAllen. 2009. pp. 244–251. [Google Scholar]
- Klem et al. (2009).Klem Jr D, Farmer CJ, Delacretaz N, Gelb Y, Saenger PG. Architectural and landscape risk factors associated with bird-glass collisions in an urban environment. The Wilson Journal of Ornithology. 2009;121:126–134. doi: 10.1676/08-068.1. [DOI] [Google Scholar]
- Klem et al. (2004).Klem D, Keck DC, Marty KL, Miller Ball AJ, Niciu EE, Platt CT. Effects of window angling, feeder placement and scavengers on avian mortality at plate glass. The Wilson Bulletin. 2004;116:69–73. doi: 10.1676/0043-5643(2004)116[0069:EOWAFP]2.0.CO;2. [DOI] [Google Scholar]
- Kummer, Bayne & Machtans (2016).Kummer JA, Bayne EM, Machtans CS. Use of citizen science to identify factors affecting bird–window collision risk at houses. The Condor: Ornithological Applications. 2016;118:624–639. doi: 10.1650/CONDOR-16-26.1. [DOI] [Google Scholar]
- Kummer et al. (2016).Kummer JA, Nordell CJ, Berry TM, Collins CV, Tse CRL, Bayne EM. Use of bird carcass removals by urban scavengers to adjust bird-window collision estimates. Avian Conservation and Ecology. 2016;11(2) doi: 10.5751/ACE-00927-110212. Article 12. [DOI] [Google Scholar]
- Lambert (1992).Lambert D. Zero-inflated poisson regression, with an application to defects in manufacturing. Technometrics. 1992;34(1):1–14. doi: 10.2307/1269547. [DOI] [Google Scholar]
- Loss et al. (2014).Loss SR, Will T, Loss SS, Marra PP. Bird-building collisions in the United States: estimates of annual mortality and species vulnerability. The Condor. 2014;116:8–23. doi: 10.1650/CONDOR-13-090.1. [DOI] [Google Scholar]
- Loss, Will & Marra (2013).Loss SR, Will T, Marra PP. The impact of free-ranging domestic cats on wildlife of the United States. Nature Communications. 2013;4:1–7. doi: 10.1038/ncomms2380. [DOI] [PubMed] [Google Scholar]
- Machtans & Thogmartin (2014).Machtans CS, Thogmartin WE. Understanding the value of imperfect science from national estimates of bird mortality from window collisions. The Condor. 2014;116(1):3–7. doi: 10.1650/CONDOR-13-134.1. [DOI] [Google Scholar]
- Machtans, Wedeles & Bayne (2013).Machtans CS, Wedeles CH, Bayne EM. A first estimate for canada of the number of birds killed by colliding with building windows. Avian Conservation and Ecology. 2013;8(2) doi: 10.5751/ACE-00568-080206. Article 6. [DOI] [Google Scholar]
- National Environmental Policy Act (1969).National Environmental Policy Act 1969. Cumulative impact. Title 40: Protection of Environment, Part 1508.7. https://ceq.doe.gov/
- Ocampo-Peñuela et al. (2016).Ocampo-Peñuela N, Winton RS, Wu CJ, Zambello E, Wittig TW, Cagle NL. Patterns of bird-window collisions inform mitigation on a university campus. PeerJ. 2016;4:e1652. doi: 10.7717/peerj.1652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O’Connell (2001).O’Connell TJ. Avian window strike mortality at a suburban office park. The Raven. 2001;72:141–149. [Google Scholar]
- Ogden (1996).Ogden LJE. Collision course: the hazards of lighted structures and windows to migrating birds. World Wildlife Fund Canada and the Fatal Light Awareness Program; Toronto: 1996. [Google Scholar]
- Parkins, Elbin & Barnes (2015).Parkins KL, Elbin SB, Barnes E. Light, glass, and bird-building collisions in an urban park. Northeastern Naturalist. 2015;22:84–94. doi: 10.1656/045.022.0113. [DOI] [Google Scholar]
- R Core Team (2017).R Core Team . R Foundation for Statistical Computing; Vienna: 2017. [Google Scholar]
- Rasband (2016).Rasband WS. US National Institutes of Health; Bethesda: 1997–2016. [Google Scholar]
- RStudio Team (2015).RStudio Team . RStudio, Inc; Boston: 2015. [Google Scholar]
- Smallwood (2007).Smallwood SK. Estimating wind turbine-caused bird mortality. The Journal of Wildlife Management. 2007;71(8):2781–2791. doi: 10.2193/2007-006. [DOI] [Google Scholar]
- Spiegelhalter et al. (2002).Spiegelhalter DJ, Best NG, Carlin BP, Van Der Linde A. Bayesian measures of model complexity and fit. Journal of the Royal Statistical Society: Series B (Statistical Methodology) 2002;64:583–639. doi: 10.1111/1467-9868.00353. [DOI] [Google Scholar]
- Virginia Department of Game and Inland Fisheries (2015).Virginia Department of Game and Inland Fisheries Virginia’s 2015 wildlife action plan. 2015. http://www.bewildvirginia.org/wildlife-action-plan. [31 March 2016]. http://www.bewildvirginia.org/wildlife-action-plan
- VTCRC (2016).Virginia Tech Corporate Research Center (VTCRC) Virginia tech corporate research center website. 2016. http://www.vtcrc.com/about/ [5 March 2016]. http://www.vtcrc.com/about/
- Wittig et al. (2017).Wittig TW, Cagle NL, Ocampo-Peñuela N, Winton SR, Zambello E, Lichtneger Z. Species traits and local abundance affect bird-window collision frequency. Avian Conservation and Ecology. 2017;12(1) doi: 10.5751/ACE-01014-120117. Article 17. [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Raw data.
Raw data.
Raw data.
Raw data.
Raw data.
Raw data.
Official survey and incidental reports of bird-window collisions on the Virginia Tech campus (September 2014–May 2015).
Data Availability Statement
The following information was supplied regarding data availability:
The raw data have been provided as Supplemental Files and here: Zirkle, Keith; rmurray@vt.edu (2018): Files for Schneider et al. (2018). Figshare. https://doi.org/10.6084/m9.figshare.5982394.v1.