

Abstract
This study establishes a relationship between desiccation tolerance and the transfer of amphiphilic molecules from the cytoplasm into lipids during drying, using electron paramagnetic resonance spectroscopy of amphiphilic spin probes introduced into imbibed radicles of pea (Pisum sativum) and cucumber (Cucumis sativa) seeds. Survival following drying and a membrane integrity assay indicated that desiccation tolerance was present during early imbibition and lost in germinated radicles. In germinated cucumber radicles, desiccation tolerance could be re-induced by an incubation in polyethylene glycol (PEG) before drying. In desiccation-intolerant radicles, partitioning of spin probes into lipids during dehydration occurred at higher water contents compared with tolerant and PEG-induced tolerant radicles. The difference in partitioning behavior between desiccation-tolerant and -intolerant tissues could not be explained by the loss of water. Consequently, using a two-phase model system composed of sunflower or cucumber oil and water, physical properties of the aqueous solvent that may affect the partitioning of amphiphilic spin probes were investigated. A significant relationship was found between the partitioning of spin probes and the viscosity of the aqueous solvent. Moreover, in desiccation-sensitive radicles, the rise in cellular microviscosity during drying commenced at higher water contents compared with tolerant or PEG-induced tolerant radicles, suggesting that the microviscosity of the cytoplasm may control the partitioning behavior in dehydrating seeds.
Anhydrobiotic organisms have the ability to withstand the removal of most of their cellular water (Crowe et al., 1992, 1997; Leprince et al., 1993; Vertucci and Farrant, 1995). There is a general consensus that survival in the dried state involves the protection of proteins and retention of membrane functional integrity upon desiccation and rehydration (Crowe et al., 1992, 1997; Hoekstra et al., 1997). Several strategies for coping with desiccation have been identified in seeds and pollens. Among molecules that are able to offer protection, the oligo- and disaccharide family (Crowe et al., 1992) and the so-called late embryogenesis abundant proteins (Close, 1996; Kermode, 1997) have received the most attention. It has been proposed recently that a concerted down-regulation of energy metabolism during desiccation (Leprince and Hoekstra, 1998; Leprince et al., 1999) is required to ensure survival in the dried state. A repressed metabolism, together with the combined free-radical processing systems, is thought to alleviate the chances of producing toxic reactive O2 species that are responsible for free-radical injury (Senaratna and McKersie, 1986; Leprince et al., 1994, 1995; Leprince and Hoekstra, 1998).
Current evidence indicates that tolerance of desiccation is a multifactorial trait. Organisms will only be able to survive drying if all the mechanisms and conditions necessary for desiccation tolerance are present in the system (Leprince et al., 1993). There are a number of reports in which desiccation tolerance is achieved without the co-operation of one of the protecting factors (Blackman et al., 1992; Hoekstra et al., 1994, 1997; Black et al., 1999; Pammenter and Berjak, 1999). There are also examples showing that desiccation-sensitive tissues may accumulate significant amounts of late embryogenesis abundant proteins and/or sugars (Vertucci and Farrant, 1995; Black et al., 1999; Pammenter and Berjak, 1999). In the light of these data, it is sensible to assume that, considering the complexity of desiccation tolerance, many mechanisms yet to be discovered are likely to participate in the acquisition of desiccation tolerance.
It is well established that many small amphiphilic molecules partition into membranes with considerable biological effects both in the hydrated and dry state. For instance, the transfer of molecules from the cytoplasm to the membranes is the basis for the drug potency of anesthetic compounds and antibiotic peptides (Janoff et al., 1981; Herbette et al., 1983; Go and Ngiam, 1997). A wide range of secondary metabolites with amphiphilic properties (e.g. carvacrol, thymol) have been found to possess strong antimicrobial activity (Ultee et al., 1998, 1999). Thermodynamic principles underlying partitioning into lipid bilayers have been applied to predict the insertion and folding of proteins into membranes (for review, see White and Wimley, 1994). In anhydrobiotes, the physiological significance of partitioning has been recently recognized (Golovina et al., 1998; Hoekstra et al., 1999). Using electron paramagnetic resonance (EPR) spectroscopy on dehydrating cattail (Typha latifolia) pollen, these authors observed the transfer of amphiphilic nitroxide spin probes from the cytoplasm into lipids during drying, indicative of a redistribution of these molecules accompanying changes in water content. In the hydrated state, the spin probes are partitioned between the aqueous cytoplasmic and lipid phase in a certain ratio of concentrations corresponding to a chemical equilibrium (Marshall, 1978; Chan and Dill, 1997). Upon dehydration, the amounts of amphiphilic spin probe present in the lipids increased (Golovina et al., 1998). In the dried state, the spin probes preferentially exist in the lipid phase. Upon rehydration, the spin probes partitioned back from the lipids into the cytoplasm.
In dry systems, the partitioning of nitroxide spin probes and endogenous amphiphilic substances extracted from pollen was found to fluidize membranes and disturb the packing order of membrane components, resulting in an increased membrane permeability (Hoekstra et al., 1997; Golovina et al., 1998; Oliver et al., 1998). This change in packing order was proposed to result in a depression of the membrane transition temperature, which could be a mechanism to prevent imbibitional injury in pollen (Hoekstra et al., 1997). Some endogenous biologically relevant amphipaths that may undergo similar partitioning effects as nitroxide spin probes are phenolic acids, flavonoids, and alkaloids (Hoekstra et al., 1997; Golovina et al., 1998). These molecules are present in high concentrations in dry seeds, pollen, and resurrection plants (Larson, 1988; Golovina et al., 1998; Oliver et al., 1998; Shirley, 1998). Furthermore, they are known to be potent anti-oxidants (Larson, 1988; Saija et al., 1995; Rice-Evans and Miller, 1996; Rice-Evans et al., 1997). Characteristically, desiccation-tolerant organisms are capable of evading damage from oxidation and peroxidative reactions during drying (Senaratna and McKersie, 1986; Leprince et al., 1994, 1995). Therefore, partitioning of these amphiphilic anti-oxidants from the cytoplasm into the membranes during drying may prevent desiccation-induced oxidative damage.
In the hydrated state, partitioning of amphiphilic molecules into membranes leads to disturbance of membrane functions. For instance, amphiphilic molecules are known to alter membrane-bound enzyme activities, such as oxidative phosphorylation (Ravanel et al., 1989; Takahashi et al., 1998) and Ca pumps (Herbette et al., 1983). The antimicrobial properties of carvacrol, which is a natural amphiphilic compound present in oregano, have been found to be mediated by altering both membrane fluidity and membrane potential of Bacillus cereus that was grown on various media (Ultee et al., 1998, 1999). Thus, the above data tentatively suggest that partitioning into lipids may be beneficial when it has occurred in the dried state, whereas it appears to be detrimental in the hydrated state.
Increase in membrane permeability, loss of membrane function, and disturbances in metabolism upon drying are characteristics of desiccation-sensitive tissues (Crowe et al., 1992; Leprince et al., 1993, 1999, 2000; Vertucci and Farrant, 1995). Therefore, this raises the question if the redistribution of endogenous amphiphilic substances during drying is associated with the loss of membrane integrity and desiccation tolerance. Prior to investigating this hypothesis, it is essential to document the behavior of the redistribution of amphipaths during drying in association with desiccation tolerance. Using EPR spectroscopy, we studied in situ the effects of drying on the partitioning of nitroxide spin probes as model amphiphilic molecules in relation with desiccation tolerance. To this purpose, imbibed pea (Pisum sativum) and cucumber (Cucumis sativa) seeds were chosen as representatives of starchy and oily seeds, respectively. Furthermore, in germinated cucumber seeds that are desiccation sensitive, desiccation tolerance can conveniently be re-induced by an osmotic stress for several days (Bruggink and van der Toorn, 1995). This physiological trait was used in this study to ascertain the relationship between desiccation tolerance and partitioning.
This study aims to also identify the possible mechanisms underlying observed differences in partitioning behavior. The molecular mechanisms that account for the partitioning of molecules between heterogeneous aqueous solvents (such as cytoplasm) and the lipid phase are complex (Simon et al., 1979; Janoff et al., 1981; White and Wimley, 1994; Chan and Dill, 1997; Rowe et al., 1998). To gain insights into the factors responsible for the desiccation-induced partitioning of amphipaths into biological tissues, a two-phase model system was developed using oil extracted from cucumber and sunflower seeds and an aqueous phase in which the physical properties of the solvent were manipulated by the addition of cosolutes.
RESULTS
Stages of Desiccation Tolerance in Imbibed Pea and Cucumber Radicles
Loss of desiccation tolerance was assessed using viability and membrane integrity assays. After 42 h of imbibition, cucumber seeds had not yet started to germinate and exhibited a high survival percentage after drying and rehydration (Table I). This was previously confirmed by an EPR spin probe technique that assessed the integrity of plasma membranes upon rehydration after different periods of drying (Leprince et al., 2000). In 42-h-old imbibed radicles, membrane permeability did not change significantly before and after drying (Leprince et al., 2000). After 72 h of imbibition, 96.2% of seeds had germinated, and their radicles were not able to survive drying (Table I). From the relation between membrane permeability and water contents during drying (extracted from data of Leprince et al., 2000), we found that the rise in membrane permeability of 72-h-old imbibed radicles occurred only when the tissues were dried below 0.3 g water/g dry weight (g/g) (Table I). This water content is referred to as the critical water content below, which membranes become sensitive to drying. Re-induction of desiccation tolerance in cucumber radicles can be brought about by exposing germinated seeds to an osmotic and chilling stress (Bruggink and van der Toorn, 1995; Leprince et al., 2000). After incubating seeds at a water potential of −1.5 MPa at 10°C for 7 d prior to dehydration, the membranes of treated radicles remained intact throughout drying and the radicles did not exhibit a critical water content (Table I).
Table I.
Germination, desiccation tolerance, and critical water content in pea and cucumber radicles during imbibition
| Species | Imbibition Treatment | % of Germinated Seeds | % of Desiccation Tolerance | Critical Water Content |
|---|---|---|---|---|
| Pea | 24 h in Water | 0 | 96 ± 4 | NAa |
| 72 h in Water | 94 ± 2 | 0 | 1.2 | |
| Cucumber | 42 h in Water | 0 | 98 ± 4 | NA |
| 72 h in Water | 91 ± 3 | 1 ± 1 | 0.3 | |
| 72 h in Water + 7 days at −1.5 MPa, 10°C | NA | 92 ± 2 | NA |
For each species, the imbibition treatment resulting in desiccation-tolerant and -intolerant radicles is shown. Germination and desiccation tolerance represent the average (±se) of three separate experiments based on a population of 30 to 50 seeds. The critical water contents correspond to the onset of membrane damage during drying and were obtained from Leprince et al. (2000). Water content is expressed as g water/g dry wt.
A, Not applicable.
In pea, 24-h-old and 72-h-old imbibed radicles were chosen to represent the desiccation-tolerant and -intolerant stages, respectively. During dehydration of the 24-h-old radicles the membrane permeability remained unchanged (Leprince et al., 2000). In contrast, the 72-h-old radicles started to lose their membrane integrity when the tissues were dried below 1.2 g/g (Table I).
Drying Induces a Redistribution of Amphiphilic Molecules from Cytoplasm to Lipids
Amphiphilic molecules, such as the nitroxide spin probes TEMPONE (4-oxo-2, 2, 6, 6-tetramethyl-1-piperidinyloxy) and TEMPO (2, 2, 6, 6-tetramethyl-1-piperidinyloxy), are distributed between the aqueous and lipid phase so that chemical equilibrium is achieved (Marshall, 1978; Tinocco et al., 1995). During drying, the EPR spectrum of TEMPONE in cucumber radicles exhibited an increase in the amplitude of the lipid peak and a decrease in the cytoplasmic (aqueous) peak (Fig. 1). Thus, drying induced movement of TEMPONE molecules from the cytoplasm into the lipids. In this paper, such movement will be referred to as a transfer process in accordance with the literature (Simon et al., 1979; White and Wimley, 1994; Chan and Dill, 1997; Go and Ngiam, 1997).
Figure 1.

EPR spectra of TEMPONE introduced in desiccation-tolerant cucumber radicles dried to various water contents (expressed on a dry weight basis). The amplitudes of the lipid (L) component and the aqueous cytoplasmic component (W) are indicated in the high-field region of the top spectrum.
The effect of drying on the transfer process of TEMPONE and TEMPO that were introduced in desiccation-tolerant tissues was studied (Fig. 2). The transfer process during drying was assessed by plotting the partitioning value as a function of water content. In hydrated cucumber radicles, the partitioning value of TEMPONE molecules was 30, whereas in hydrated pea radicles, it was approximately 8. Such difference is attributable to the higher amount of storage lipids in cucumber compared with pea (Golovina et al., 1997, 1998). During the onset of drying, no significant changes in the transfer process occurred in each species. When a water content of approximately 0.8 g/g was reached, the TEMPONE molecules progressively moved into the lipid phase with further drying as indicated by an increase in the partitioning value (Fig. 2). Note that the onset of transfer of TEMPONE in pea and cucumber radicles occurred at approximately the same water content. Below 0.2 g/g, a powder spectrum appeared, indicating that part of the spin probe became immobilized (data not shown). This made it impossible to determine the partitioning value accurately; subsequently, spectra of samples containing less that 0.2 g/g were not analyzed.
Figure 2.

Effect of drying on the partitioning value of two amphiphilic nitroxides TEMPO and TEMPONE that were introduced in desiccation-tolerant radicles of pea and cucumber before drying (TEMPO, pea [▾]; TEMPONE, pea [●]; and TEMPONE, cucumber [○]). Data represent two to four different drying experiments that were pooled together.
The effect of polarity of the spin probes on the transfer process during drying was studied in pea radicles (Fig. 2). TEMPO is a more apolar molecule than TEMPONE. Because TEMPO has a higher chemical affinity with hydrophobic solvent than TEMPONE, a higher amount of TEMPO molecules is expected to be present in the lipid phase. This is illustrated in Figure 2 where the partitioning value of TEMPO in hydrated radicles of pea was 40, compared with 8 for TEMPONE. Figure 2 shows that the partitioning behavior of TEMPO is also sensitive to the loss of water. In cucumber axes (data not shown), the partitioning value for TEMPO in the hydrated state was already 90, which made it impossible to follow the transfer process during drying. It can be observed that the moisture content at which an endogenous natural amphipath moves from the cytoplasm to the lipid phase during drying depends on its polarity.
Desiccation-Tolerant and -Intolerant Tissues Exhibit Differential Transfer of Amphiphilic Spin Probes during Drying
The desiccation-induced transfer of amphiphilic molecules in desiccation-tolerant radicles prompted the question of whether this redistribution was similar in desiccation-intolerant tissues. Because partitioning of amphipaths into membranes affects the physical properties and functionality of membranes (Herbette et al., 1983; Ravanel et al., 1989; Golovina et al., 1998; Rowe et al., 1998; Takahashi et al., 1998), the onset of the transfer of these molecules into the lipids during drying might be important to desiccation tolerance.
To assess the relation between partitioning and desiccation tolerance, the transfer process of TEMPONE into the lipids during drying was compared in desiccation-tolerant and -intolerant radicles of pea and cucumber (Fig. 3). In desiccation-intolerant radicles of pea, the curve representing the relation between partitioning value and water content during drying was significantly above that of desiccation-tolerant pea radicles (Fig. 3A). Thus, the transfer of TEMPONE occurred at higher water contents during drying in desiccation-intolerant tissues than in tolerant ones. When tolerant and sensitive tissues were dried below 0.4 g/g, almost all TEMPONE molecules had partitioned into the lipids.
Figure 3.

Partitioning values of TEMPONE introduced in radicles of pea (A) and cucumber (B) as a function of water content during drying of desiccation-tolerant (●), -intolerant (○), and PEG-induced tolerant radicles (▴). For each treatment data of two to four independent drying experiments are shown together.
In desiccation-tolerant cucumber radicles, drying did not substantially affect partitioning values until a water content of 0.8 g/g was reached. Reducing the water content below this value resulted in a steep increase in the transfer process of TEMPONE. In desiccation-sensitive radicles, initial drying did not induce noticeable changes in partitioning value until the tissue water content reached 1.4 g/g. Thereafter, partitioning values increased steeply with decreasing water content. Comparison of the partitioning behavior of desiccation-sensitive radicles with desiccation-tolerant tissues revealed that <1.0 g/g, the partitioning curve of sensitive tissues was positioned above that of desiccation-tolerant radicles, and that the onset of the transfer process commenced at higher water contents in desiccation-sensitive tissues (Fig. 3).
Figure 3 shows that, at the onset of drying, the partitioning value in desiccation-sensitive cucumber radicles was lower than in tolerant radicles. This difference in partitioning value between hydrated desiccation-sensitive and -tolerant tissues likely originated from differences in lipid content (33.0 ± 2.9% and 25 ± 2.9% dry weight for desiccation-tolerant and -sensitive tissues, respectively). This interpretation can also be deduced from Figure 2, where cucumber seeds (high oil content) are compared with pea seeds (low oil content). The partitioning value of TEMPONE in hydrated cucumber radicles is higher than that in pea radicles with similar water contents.
An osmotic treatment that re-induced desiccation tolerance in sensitive radicles of cucumber was used to further establish the relationship between desiccation tolerance and the differential transfer of amphipaths into the lipid environment upon drying (Table I, Fig. 3B). At the onset of drying, the transfer process of TEMPONE in polyethylene glycol (PEG)-treated radicles was similar to that of desiccation-intolerant tissues. However, the onset of the partitioning commenced at lower water contents compared with desiccation-intolerant tissues. Note that correction of the water contents due to changes in oil contents (25 ± 2.9% and 22.0 ± 1.7% dry weight for desiccation-intolerant and PEG-treated tolerant radicles) did not significantly change the position of the partitioning curves, and therefore could not account for the differential transfer. At water contents <0.8 g/g, the curve representing partitioning values as a function of water content of the PEG-induced tolerant radicles overlapped that of desiccation-tolerant tissues. Thus, our data indicate that the onset of the transfer process of amphipaths during drying occurred at higher water contents in desiccation-sensitive radicles than in tolerant ones.
The difference in partitioning behavior between tolerant and intolerant tissues cannot stem from changes in tissue dry weight during germination. Changes in dry weight would affect the expression of the water content on a dry-weight basis, resulting in an horizontal shift of the partitioning curves. The actual changes in dry weight between the different stages did not exceed 7% (data not shown). When the curves of the partitioning values in Figure 3 were corrected for the changes in dry weight between the different treatments, no significant changes in the outcome of the results were found (data not shown). We calculated that to account for the difference in partitioning values found in this study the decrease in dry weight in desiccation-intolerant tissues should have been approximately 40%.
Causes of Desiccation-Induced Transfer of Amphiphilic Molecules
The differential onset of partitioning of amphiphilic spin probes during drying between desiccation-tolerant and -intolerant tissues led us to investigate possible causes for such a difference. The causes for transfer of amphipaths into lipids have been extensively studied in model systems using thermodynamics and molecular dynamics (De Young and Dill, 1988; Chan and Dill, 1997; Rowe et al., 1998). The roles of chemical affinity (i.e. hydrophobicity and/or polarity of both amphipaths and solvents), the changes in solvent volume and the composition of the solvent, in partitioning (Marshall, 1978; Chan and Dill, 1997) have been well established. Using a two-phase model system composed of extracted cucumber oil and water, we studied how changes in physical properties of the aqueous solvent affect the transfer process of TEMPONE into oil.
The change in volume of free water in the cytoplasm has been considered as a factor involved in desiccation-induced partitioning of TEMPONE introduced in pollen (Golovina et al., 1998). To check this possibility in our material, the transfer process of TEMPONE was monitored in a cucumber oil/water system in which the volume of the water was reduced by decreasing the water/oil ratio (Fig. 4). A reduction of water volume from 5 to 0.5 induced an increase in the partitioning value of TEMPONE from 5 to 40, indicating that the reduction of water volume in our biological material will induce the transfer process of amphiphilic molecules during drying. However, a reduction of water volume cannot explain the observed differences in water contents corresponding to the onset of the partitioning value in desiccation-tolerant and -intolerant tissues during drying (Fig. 3). A reduction in water volume during drying can be translated into a reduction of water content. Yet, Figure 3 demonstrates that differences in transfer behavior between desiccation-tolerant and intolerant tissues still persist when the partitioning values are expressed as a function of tissue water content.
Figure 4.

Effect of decreasing aqueous volumes on the partitioning value of TEMPONE in a two-phase model system consisting of various amounts of water and a constant amount of storage oil extracted from cucumber seeds.
Changes in intrinsic properties of the aqueous solvent consequently were thought to be responsible for differences in partitioning in vivo. Therefore, the properties of the aqueous solvent of a two-phase model system consisting of water and sunflower oil were altered by the addition of a range of different co-solutes that are known to affect polarity, pH, conductivity, and viscosity (Table II). The amount of spin probe and oil/water ratio was kept constant. Table II shows that changes in the solvent conductivity using CaCl2 and CsCl2 or changes in pH had no or very little effect on the transfer process of TEMPONE. However, high concentrations of glycerol, dimethyl sulfoxide (DMSO), or sugars (Glc or Suc) were able to significantly induce the transfer of TEMPONE into the oil. In contrast to Tris (tris-[hydroxymethyl]aminomethane) and salts, high amounts of dissolved DMSO, glycerol, Glc, and Suc have a substantial effect on the solution viscosity (Table II). Thus, the increase in viscosity may induce the transfer of TEMPONE molecules into the oil phase. This was confirmed in Figure 5 using glycerol and Suc solutions of known viscosity in the water/sunflower oil system. For both compounds molecules, a significant linear relationship was found between the viscosity of the aqueous solution and the partitioning value. However, the slopes of the regressions were found to be different, suggesting that an additional factor is responsible for the transfer of TEMPONE into the oil phase.
Table II.
Effect of various co-solutes on the partitioning of TEMPONE in a two-phase model system (water:sunflower oil, 1:1 [v:v]) at 22°C
| Co-Solute | Concentration | Mole Fraction | Solvent Properties Altered | Relative Viscosity η/ηo | Increase in P Value (ΔP) |
|---|---|---|---|---|---|
| None | 0 | 1.00 | 0 (10.8 ± 0.01) | ||
| CaCl2 | 25% (w/v) | 0.04 | Conductivity | 2.75 | 3.30 |
| CsCl2 | 1% (w/v) | 0.01 | Conductivity | 0.99 | 0.94 |
| CsCl2 | 15% (w/v) | 0.02 | Conductivity | 0.96 | 0.98 |
| Tris, pH 2.6 | 50 mm | 8×10−4 | Ionization | 1.01 | 0.39 |
| Tris, pH 5.3 | 50 mm | 8×10−4 | Ionization | 1.01 | 0.44 |
| Tris, pH 6.6 | 50 mm | 8×10−4 | Ionization | 1.01 | 0.39 |
| Tris, pH 9.1 | 50 mm | 8×10−4 | Ionization | 1.01 | 0.57 |
| DMSO | 75% (v/v) | 0.408 | Hydrogen bonding | 1.53 | 6.81 |
| Glycerol | 40% (v/v) | 0.061 | Hydrogen bonding | 2.01 | 6.00 |
| Glycerol | 75% (v/v) | 0.369 | Hydrogen bonding | 27.57 | 45.1 |
| Glc | 60% (w/v) | 0.13 | Hydrogen bonding | 3.76 | 33.2 |
| Suc | 75% (w/v) | 0.14 | Hydrogen bonding | 10.61 | 61.2 |
Solutes were chosen according to the nature of their concentrative effect on the dielectric constant of aqueous solution (conductance, hydrogen bonding, and ionization). The amounts of added co-solutes are expressed both on a volume and a mole fraction basis. Data are the average of duplicates (sd < 1%). Partitioning data are expressed as an increase in partitioning (P) value compared with values of the water control (average of seven replicates [±sd] is shown in brackets). Relative viscosity of the aqueous solutions were determined experimentally using EPR and Eq. 2 (see “Materials and Methods”) except for CaCl2 and CsCl2, which were taken from Wolf et al. (1986).
Figure 5.

Effect of viscosity on the partitioning value of TEMPONE in a two-phase model system consisting of equal volumes of sunflower oil and aqueous solvent. Viscosity in the aqueous phase was manipulated by adding various amounts of glycerol (●) or Suc (○). For glycerol, two independent experiments are shown together, whereas for Suc, the average of duplicate samples (±sd) is shown. Viscosity was experimentally determined using TEMPONE EPR spectroscopy (Eq. 2) in the presence and the absence of the compounds in the solution. Viscosity values are expressed as a relative value compared to the viscosity of pure water (η0). Data were fitted with linear regressions (r2 indicated).
Changes in Cellular Viscosity in Relation to Desiccation Tolerance
To assess whether changes in viscosity during drying may account for the desiccation-induced transfer process of TEMPONE in our biological material, the microviscosity of the cytoplasm was measured using the polar spin probe 3-carboxy-proxyl (CP). Being a polar spin probe, CP was not present in the lipid phase of the cells within the range of water contents studied here (data not shown). In pea, the cellular microviscosity in both desiccation-tolerant and -intolerant radicles increased upon drying (Fig. 6A). Before drying, the cellular microviscosity was higher in desiccation-intolerant radicles than in tolerant tissues. At any given water content during drying, the cytoplasm of desiccation-intolerant radicles exhibited a viscosity approximately 2-fold higher than that of tolerant tissues.
Figure 6.

Effect of drying on the cellular microviscosity in pea (A) and cucumber (B) radicles at different stages of germination (desiccation-tolerant [●], -intolerant [○], and PEG-induced tolerant [▴]). Microviscosity was measured by EPR spectroscopy using CP as a polar spin probe. Data represent two to four independent drying experiments that are pooled together. Data were fitted with fourth-order polynomial regressions (r2 > 0.96), except for desiccation-tolerant cucumber, where an asymmetric function was used (r2 = 0.87).
In cucumber, the cellular microviscosity remained constant during the onset of drying from 1.2 g/g to 0.6 g/g and from 2.6 g/g to 1.6 g/g for desiccation-tolerant and -intolerant radicles, respectively (Fig. 6B). When tissues were dried below these values, microviscosity of the desiccation-intolerant cytoplasm of cucumber radicles was considerably higher than that of tolerant radicles. In PEG-induced tolerant cucumber radicles, the desiccation-induced increase in microviscosity was intermediate between tolerant and intolerant tissues (Fig. 6B). In these tissues, microviscosity remained constant during drying until a water content of 0.9 g/g was reached, thereafter it increased sharply. The pattern of increase resembled that of 42 h-imbibed radicles, suggesting that cellular viscosity could be linked to desiccation tolerance, as suggested earlier (Leprince and Hoekstra, 1998).
On account of our observations from the two-phase model system (Fig. 5), the differential increase in viscosity between desiccation-tolerant and -sensitive tissues may explain the difference in partitioning behavior. This was assessed by plotting the values of cellular microviscosity obtained during drying (Fig. 6) as a function of partitioning values of TEMPONE (Fig. 3) in pea and cucumber. For pea, the relationships between partitioning values and cellular microviscosity for both desiccation-tolerant and -intolerant tissues were overlapping each other (Fig. 7A). Similarly, the relation obtained from desiccation-intolerant and PEG-induced tolerant radicles of cucumber (Fig. 7B) overlapped each other perfectly, suggesting that intracellular viscosity is regulating the transfer process of amphiphilic molecules during drying. However, partitioning values of desiccation-tolerant radicles of cucumber increased at a much lower viscosity than those of sensitive and PEG-induced tolerant tissues. This difference can be traced back in Figure 3, which shows that the partitioning value in hydrated tissues is 3.2-fold higher in tolerant than in sensitive axes. Such difference corresponds approximately to the shift found in the viscosity/partitioning relationship between desiccation-tolerant and -sensitive tissues (Fig. 7B). Therefore, it is the initial partitioning value of the hydrated desiccation tolerant tissues that is responsible for the differences exhibited in Figure 7B. However, at viscosities rising from 0.02 and 0.08 n m−2 s, both relationships have a similar slope, reinforcing the argument that the shift in the viscosity/partitioning curve is due to a difference in partitioning value before drying. This difference in absolute partitioning values between hydrated tolerant and sensitive cucumber axes originates from differences in oil contents, as demonstrated earlier.
Figure 7.

The relation between partitioning value of TEMPONE (derived from Fig. 3) as a function of the cellular microviscosity in pea (A) and cucumber (B) during drying of desiccation-tolerant (●), -intolerant (○), and PEG-induced-tolerant (▴) radicles. Values of the cellular viscosity were obtained from water content values by interpolation of the fitted curves shown in Figure 6.
DISCUSSION
Using EPR spectroscopy of nitroxide spin probes introduced into imbibed radicles of pea and cucumber, this study assessed the transfer process of amphiphilic molecules into lipids in dehydrating desiccation-tolerant and -intolerant tissues. As previously shown for dehydrating pollen (Golovina et al., 1998), the water removal from seed tissues induced the transfer of both TEMPO and TEMPONE (amphiphilic spin probes of different polarity) into the lipids (Figs. 2 and 3). Figure 3 shows that the onset of the transfer process occurred at higher water contents in desiccation-intolerant radicles than in tolerant radicles during drying. The relationship between desiccation tolerance and early partitioning during drying was ascertained with cucumber seeds using PEG treatments, which can re-induce desiccation tolerance in germinated sensitive radicles (Bruggink and van der Toorn, 1995; Table I).
Transfer of Amphiphilic Compounds into the Lipids during Drying Relates to Increase in Viscosity
The observation that desiccation-tolerant and -sensitive tissues exhibited a differential transfer process of TEMPONE upon drying raised the question of what factors can be responsible for this difference. As mentioned before, changes in dry weight during germination cannot account for the different partitioning behavior. In addition, the observed changes in lipid contents of the different stages did not account for the differences in partitioning behavior between tolerant and intolerant tissues. In desiccation-intolerant radicles of cucumber, the amount of lipid is reduced by 25% compared with tolerant tissues. Our in vitro partitioning study on two-phase model system (Fig. 4) shows that the lower the amounts of lipids, the lower the partitioning value. Considering that the lipid content decreased in intolerant tissues, this would result in a decrease in partitioning value, which is contrary to our observations (Fig. 3). Therefore, the difference in partitioning behavior between desiccation-tolerant and -intolerant radicles is not an artifact due to differences in lipid contents or in water volume to lipid volume ratio between these tissues.
This study has identified three main factors that drive the transfer process of nitroxide spin probes during desiccation: (a) polarity of the amphiphilic compounds (Fig. 2); (b) change in water volume during drying (Fig. 4); and (c) cellular viscosity (Table II; Figs. 6 and 7, A and B). Various thermodynamic models have demonstrated that both polarity (i.e. chemical affinity) and change in water volume (i.e. change in mole fraction of solutes) control the equilibrium concentration of amphipaths between oil and aqueous phases (Tinocco et al., 1995; Chan and Dill, 1997; Go and Ngiam, 1997). The importance of the change in volume in the transfer process is illustrated in the in vitro model system where the volumes of the aqueous (Fig. 4) or lipid (Fig. 7C) phase were manipulated. Both polarity and reduction in volume of free water were assumed to be responsible for transfer of spin probes from and into lipids in cattail pollen subjected to drying and rehydration regimes (Golovina et al., 1998).
However, in our material, neither polarity nor change of water volume could account for the differential transfer of TEMPONE between desiccation-tolerant and -sensitive radicles. This can be clearly seen in Figure 3, where partitioning values are plotted as a function of water content, which can be translated into volume. Figures 5 and 6 and Table II indicate that the rise in cellular viscosity during drying may be regarded as a key factor driving the transfer process of amphiphilic compounds into the lipids of seed tissues. Furthermore, Figure 5 and Table II indicate that an additional factor besides viscosity is influencing the transfer process. Because the co-solutes that affect viscosity contain several hydroxyl groups, this additional factor could be related to hydrogen bonding between co-solutes and TEMPONE.
At the molecular level, the link between cellular viscosity and transfer behavior of TEMPONE may be supported by theoretical considerations derived from model systems. For many years, the assumption has been made that the mole fraction of solvent and solutes (so-called entropy of mixing [i.e. the change in entropy resulting from the introduction of the amphipaths in each phase]) and chemical affinity between solute and solvent can be used as thermodynamic parameters to fully describe a partition phenomenon (Marshall, 1978; White and Wimley, 1994). However, there is increasing evidence suggesting that molecular interactions between solutes and solvents and additional forces (such as conformation of molecules) may influence partitioning behavior of amphiphilic molecules (De Young and Dill, 1988; White and Wimley, 1994; Chan and Dill, 1997; Go and Ngiam, 1997; Rowe et al., 1998). For instance, steric interference of the solutes or any molecular process that restricts rotational and translational freedom has a significant effect on partitioning. This is particularly relevant when the solvents contain polymeric molecules such as polypeptides, which is probably the case for cytoplasm and lipids (White and Wimley, 1994; Chan and Dill, 1997). The EPR spectroscopy method used in this study to assess viscosity gives access to values of local viscosity (i.e. molecular range) as opposed to viscosity at a macromolecular scale (i.e. dynamic flow; Marsh, 1981; Roozen et al., 1991; Leprince et al., 1999). High local viscosity restricts rotational and translational movements of the spin probes in each media. This restriction of movement would then result in an energetically unfavorable situation that will induce the transfer process into the lipids.
Therefore, in the light of these observations and extrapolating the data from our model system studies (Table II), we suggest that the impact of viscosity on partitioning behavior of amphipaths in radicles may stem from the rise in molecular crowding within the cytoplasm and changes in hydrogen bonding during drying. Our suggestion echoes that of Wolkers (1998) who suggested that the control of molecular interactions through hydrogen bonding during drying is important to achieve desiccation tolerance. It is likely that desiccation-tolerant tissues may have the capability of controlling both cellular microviscosity and hydrogen bonding properties by changing their cytoplasmic composition. The above suggestions reinforce the consideration of cellular viscosity as an emerging mechanism of desiccation tolerance (Leprince and Hoekstra, 1998). Previously we showed that viscosity depresses O2 diffusion and high metabolism rates during drying, both of which have been implicated in the desiccation sensitivity of seeds (Leprince and Hoekstra, 1998; Leprince et al., 1999).
Desiccation Tolerance Is Related to Differential Transfer of Amphipaths during Drying
The onset of desiccation-induced transfer of amphiphilic molecules occurred at higher water contents in desiccation-sensitive tissues than in tolerant ones (Fig. 3). This observation raises the question whether there would be a possible relationship between partitioning and desiccation tolerance. It is well documented that, at high water contents, amphipath partitioning leads to injurious effects to prokaryotes, plant, and animal systems. The interactions of amphiphilic molecules with membranes result in a direct damaging effect by perturbation of the membrane properties (Herbette et al., 1983; Maher and Singer, 1984; Saija et al., 1995; Hoekstra et al., 1997; Golovina et al., 1998; Oliver et al., 1998; Ultee et al., 1998, 1999). Flavonoids, a large family of amphipaths that are present in seeds, induced a decrease in membrane permeability of lipid vesicles that was proportional to the amount of amphipaths introduced in the lipid bilayers (Golovina et al., 1998; Oliver et al., 1998; Hoekstra et al., 1999). In addition, the perturbing effects of amphipaths on membranes are known to cause metabolic dysfunction. Ultee et al. (1999) have shown that carvacrol affects the membrane fluidity of prokaryotes in the hydrated state, leading to a dissipation of the proton gradient, which in turn induced a depletion of ATP and a drop in pH. Many amphipaths such as flavonoid aglycones are able to alter oxidative phosphorylation or impair electron transfer in isolated plant mitochondria (Ravanel et al., 1989; Takahashi et al., 1998). It is noteworthy that various symptoms of desiccation-induced metabolic dysfunction have been observed at high moisture contents and before the onset of loss of membrane integrity in recalcitrant seeds and germinating orthodox seeds that had lost their tolerance (Leprince et al., 1999, 2000). By analogy with these studies, it could be argued that partitioning at too high water contents (where metabolism is still highly active) of endogenous amphipaths into the lipid phase during drying may initiate a cascade of degradative reactions. The high tissue water content would allow these reactions to propagate during drying and may ultimately lead to cell death in the dry state.
In this study, the water content region where a difference in partitioning is evident does not match the critical water content for membrane integrity (Table I). However, this observation is not in contradiction with a possible link between partitioning and desiccation sensitivity. Figure 2 shows that the transfer process of amphiphilic molecules into the lipid phase is also dependent on the polarity of the molecule. Thus, each type of molecule will redistribute at different water contents during drying according to their polarity. It follows that the range of water contents for which the partitioning is different in desiccation-tolerant and -intolerant tissues will depend on the polarity of the amphiphilic molecule studied. In respect with desiccation tolerance, the water content range at which endogenous and potentially harmful amphipaths may partition into membranes needs further investigation. Nonetheless, regardless of its polarity, any endogenous amphiphilic molecule will redistribute to the lipids at a higher water content during drying in desiccation-intolerant tissues than in desiccation-tolerant tissues.
During the latter stages of drying, an increasing amount of amphipaths present in the cytoplasm will partition into the lipids, irrespective of the initial difference in the onset of the transfer process between desiccation-intolerant and -tolerant tissues. A transfer process that occurs at a low-water content, at which metabolism is dramatically reduced because of a high-cellular viscosity (Leprince and Hoekstra, 1998), is unlikely to induce metabolic dysfunction due to membrane disturbances. In effect, a transfer process occurring at very low-water contents could be advantageous for the long-term stabilization of membranes in the dried state, considering the anti-oxidant potential and/or membrane fluidizing properties of some endogenous amphipaths such as flavonoids. Future studies are pending to ascertain whether there is a link between partitioning into membranes during drying and desiccation-induced damage.
MATERIALS AND METHODS
Plant Material and Treatments
Seeds of cucumber (Cucumis sativa cv Alfaris, Novartis, Enkhuizen, The Netherlands) and pea (Pisum sativum cv Karina, Nunhems Zaden, The Netherlands) were imbibed for up to 72 h on wet filter papers at 20°C and 15°C, respectively. To obtain desiccation-tolerant and -intolerant tissues, pea and cucumber seeds were removed at different intervals of imbibition. Pea seeds were subsequently dried at room temperature under a flow of dry air (approximately 3% relative humidity [RH]) for 24 h and cucumber seeds were dried in a container at 33% RH achieved by a saturated K2CO3 solution. Desiccation-tolerant cucumber and pea radicles were obtained by imbibing seeds, respectively, for 42 and 24 h, whereas intolerant radicles of both species were obtained after 72 h of imbibition. To induce desiccation tolerance in germinated cucumber radicles, 72-h-imbibed seeds with a radicle length ranging from 2 to 4 mm were submitted to an osmotic treatment (PEG 8000 solution having a water potential of −1.5 MPa) at 10°C for 7 d (Bruggink and van der Toorn, 1995). After the osmotic treatment and drying, seeds were rinsed briefly in water then re-imbibed on wet filter paper at 25°C. Growth of the radicle was scored as a marker for desiccation tolerance. Water contents were assessed gravimetrically by comparing the sample weights before and after drying for 40 h at 96°C and are expressed on a dry weight basis.
Determination of Partitioning Behavior
TEMPONE and TEMPO are amphiphilic nitroxide spin probes. According to their polarity characteristics, the spin probes will distribute between the lipid and the aqueous phase. This distribution can be assessed by EPR spectroscopy because of differences in the isotropic hyperfine splitting constants of EPR spectra in the hydrophilic and hydrophobic environments (Marsh, 1981). Thus, the partition behavior of these amphiphilic molecules between lipid (storage oil and membranes) and cytoplasm can be easily monitored in situ on excised radicles. The spectrum of TEMPO(NE) introduced into the seed tissues is the result of a superposition of two spectra, corresponding to the spin probe present in the cytoplasm (W) and lipids (L). The spectra superposition is resolved in the high-field region as two peaks (Fig. 1). Radicles of cucumber and pea were incubated for 30 min in 1 mm of TEMPO or TEMPONE solution at room temperature. After 15 min, 120 mm K-ferricyanide was added to remove the spin probe signal from the outside of the tissues and the intercellular spaces.
For in vitro partitioning experiments, we used two-phase model systems consisting of oil (extracted from our cucumber seeds or commercial sunflower oil) that was added, in different proportions, to aqueous solutions containing TEMPONE alone or codissolved with various concentrations of respectively CaCl2, CsCl2, DMSO, Tris, glycerol, Suc, and Glc (Sigma, St. Louis). The amount of TEMPONE was adjusted to obtain a constant concentration of 250 μm in the total mixture volume. Three to 5 μL of the oil and aqueous solution was transferred into a 200-μL EPR capillary that was sealed. The two phases were mixed by centrifuging twice the solutions from top to bottom. After 24 h, spectra of the two-phase system were recorded when the partitioning of the spin probe had reached equilibrium (data not shown).
The distribution or partitioning of amphiphilic spin probes at equilibrium between a lipid and a water phase was quantified from the EPR spectra using the partitioning value (Marsh, 1981):
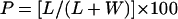 |
1 |
where L and W are the amplitudes of the high-field peaks of the spectrum originating from the spin probe being present in the lipid or aqueous cytoplasmic phase, respectively (Fig. 1). The amplitudes of the respective peaks were considered to be a good approximation of the amount dissolved in the lipid phase and water phase, respectively, as demonstrated by Golovina et al. (1998).
Viscosity Measurements
The polar spin probe CP was used to determine the microviscosity of the cytoplasmic environment of the cells according to the methods described in Leprince et al. (1999). Being very polar, CP was detected solely in the cytoplasm (Buitink et al., 1998). Isolated radicles of pea and cucumber were labeled for 60 min in a 1-mm CP solution at room temperature. After 45 min of incubation, 200 mm K-ferricyanide was added to remove the signal from the outside of the tissues and the intercellular spaces. To obtain the microviscosity, the rotational correlation time (τR), corresponding to the lifetime of the spin probe in a given orientation, was obtained from the EPR spectrum using the equation developed by Knowles et al. (1976):
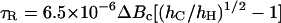 |
2 |
where Bc is the width of the center field component in Tesla, and hC and hH are the amplitudes of the central and high-field components of the three-line nitroxide radical spectrum, respectively. The rotational motion of the spin probe was assumed to be isotropic. From τR, one can derive the microviscosity, i.e. the local viscosity of the matrix in which the spin probe is rotating, according to the modified Stokes-Einstein equation (Roozen et al., 1991):
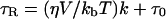 |
3 |
where τR is the rotational correlation time, η is the solvent viscosity, kb is Boltzmann's constant, V the volume of the rotating molecule (1.8 × 10−28 m3 for CP), T the absolute temperature, τ0 is the rotational correlation time at zero viscosity (which is negligible), and k is a dimension-less slip parameter (0.09 for CP; Roozen et al., 1991).
EPR Spectroscopy
After the physiological treatments and labeling with the appropriate nitroxide spin probe, isolated radicles were dried on the bench (30%–40% RH, 20°C–22°C) for different time intervals after which three pea axes or four cucumber axes per drying treatment were sealed in EPR glass capillaries. EPR spectra were recorded with a Bruker X-band EPR spectrometer (model 300E; Bruker Analytik, Rheinstetten, Germany) using a microwave power of 2 mW and a modulation amplitude of 0.03 milliteslas (for TEMPO and TEMPONE) or 0.1 milliteslas (for CP). After measurements, the samples were retrieved from the capillaries, and their water contents were determined as described above.
Lipid Extraction and Estimation of Storage Oil Content
Total lipids were extracted from 72-h-imbibed cotyledons of cucumber with CHCl3-methanol using a modified Folch's procedure described by Hamilton et al. (1992). Total lipids were used for in vitro oil-water partitioning experiments. The storage oil content in radicles was estimated by differential scanning calorimetry using known amounts of isolated oil bodies as standards. In seeds, the storage oil is located in small spherical intracellular organelles, termed “oil body.” Oil bodies were extracted and purified by flotation according to Murphy and Cummins (1989), then dried under 3% RH. Because oil bodies are surrounded by a phospholipid monolayer and proteins, oil bodies retain their in vivo integrity after drying (Murphy and Cummins, 1989). Three dried radicles or known amounts of dried oil bodies were sealed in aluminum pans and scanned from −90°C to 25°C at 10°C/min in a Pyris-1 differential scanning calorimeter (Perkin-Elmer, Norwalk, CT). The absence of water makes the melting of storage lipids clearly visible. From the heating thermograms, the melting enthalpies of lipids were determined using the Pyris-1 software. The amount of storage lipid in cucumber radicles was then estimated from lipid melting enthalpies using the relation between the known weight of isolated oil bodies and their respective lipid melting enthalpies.
Experimental Design and Statistics
Tolerance of desiccation was tested at least three times independently on 30 to 50 seeds per treatment. For the EPR spectroscopy on biological material, the experimental unit is a sample of three to four isolated radicles dried to a certain moisture content. Experiments were replicated at least three times to obtain an appropriate range of water contents. Data from all experiments were pooled together in a single graph. For the EPR spectroscopy on two-phase systems, the various mixtures containing oil and aqueous solutions were prepared and analyzed in duplicate.
ACKNOWLEDGMENTS
The authors thank Dr. E.A. Golovina and Prof. A.N. Tikhonov (Moscow University) for stimulating discussions and attracting our attention to the pitfalls of the spin probe EPR spectroscopy, Dr. F.A.M. Leermakers (Laboratory of Physical Chemistry and Colloid Science, Wageningen University) for discussing the relations between viscosity and partitioning and Dr. T. Bruggink (Novartis, Enkhuizen, The Netherlands) for the generous gift of cucumber seeds.
Footnotes
This work was supported by the Netherlands Technology Foundation (Stichting Technische Wetenschappen) and was coordinated by the Life Sciences Foundation.
LITERATURE CITED
- Black M, Corbineau F, Gee H, Côme D. Water content, raffinose, and dehydrins in the induction of desiccation tolerance in immature wheat embryos. Plant Physiol. 1999;120:463–471. doi: 10.1104/pp.120.2.463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blackman SA, Obendorf RL, Leopold AC. Maturation proteins and sugars in desiccation tolerance of developing soybean seeds. Plant Physiol. 1992;100:225–230. doi: 10.1104/pp.100.1.225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bruggink T, van der Toorn P. Induction of desiccation tolerance in germinated seeds. Seed Sci Res. 1995;5:1–4. [Google Scholar]
- Buitink J, Claessens MMAE, Hemminga MA, Hoekstra FA. Influence of water content and temperature on molecular mobility and intracellular glasses in seeds and pollen. Plant Physiol. 1998;118:531–541. doi: 10.1104/pp.118.2.531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chan HS, Dill KA. Solvation: how to obtain microscopic energies from partitioning and solvation experiments. Annu Rev Biophys Biomol Struct. 1997;26:425–429. doi: 10.1146/annurev.biophys.26.1.425. [DOI] [PubMed] [Google Scholar]
- Close TJ. Dehydrins: emergence of a biochemical role of a family of plant dehydration proteins. Physiol Plant. 1996;97:795–803. [Google Scholar]
- Crowe JH, Crowe LM, Carpenter JF, Prestrelski SJ, Hoekstra FA, De Araujo PS, Panek AD. Anhydrobiosis: cellular adaptations to extreme dehydration. In: Dantzler WH, editor. Handbook of Physiology, Section 13, Comparative Physiology. II. Oxford: Oxford University Press; 1997. pp. 1445–1477. [Google Scholar]
- Crowe JH, Hoekstra FA, Crowe LM. Anhydrobiosis. Annu Rev Physiol. 1992;54:570–599. doi: 10.1146/annurev.ph.54.030192.003051. [DOI] [PubMed] [Google Scholar]
- De Young LR, Dill KA. Solute partitioning into lipid bilayer membranes. Biochemistry. 1988;27:5281–5289. doi: 10.1021/bi00414a050. [DOI] [PubMed] [Google Scholar]
- Go M-L, Ngiam T-L. Thermodynamics of partitioning of the antimalarial drug mefloquine in phospholipid bilayers and bulk solvents. Chem Pharmacol Bull. 1997;45:2055–2060. doi: 10.1248/cpb.45.2055. [DOI] [PubMed] [Google Scholar]
- Golovina EA, Hoekstra FA, Hemminga MA. Drying increases intracellular partitioning of amphiphilic substances into the lipid phase: impact on membrane permeability and significance for desiccation tolerance. Plant Physiol. 1998;118:975–986. doi: 10.1104/pp.118.3.975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Golovina EA, Tikhonov AN, Hoekstra FA. An electron paramagnetic resonance spin probe study of membrane permeability changes with seed aging. Plant Physiol. 1997;114:383–389. doi: 10.1104/pp.114.1.383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hamilton S, Hamilton RJ, Sewell PA. Extraction of lipids and derivative formation. In: Hamilton RS, Hamilton S, editors. Lipid Analysis. A Practical Approach. Oxford: IRL Press; 1992. pp. 13–64. [Google Scholar]
- Herbette L, Katz AM, Sturtevant JM. Comparisons of the interaction of propranolol and timolol with model and biological membrane systems. Mol Pharmacol. 1983;24:259–269. [PubMed] [Google Scholar]
- Hoekstra FA, Golovina EA, Van Aelst AC, Hemminga MA. Imbibitional leakage from anhydrobiotes revisited. Plant Cell Environ. 1999;22:1121–1131. [Google Scholar]
- Hoekstra FA, Haigh AM, Tetteroo FAA, Van Roekel T. Changes in soluble sugars in relation to desiccation tolerance in cauliflower seeds. Seed Sci Res. 1994;4:143–147. [Google Scholar]
- Hoekstra FA, Wolkers WF, Buitink J, Golovina EA, Crowe JH, Crowe LM. Membrane stabilization in the dried state. Comp Biochem Physiol. 1997;117A:335–341. [Google Scholar]
- Janoff AS, Pringle MJ, Miller KW. Correlation of general anesthetic potency with solubility in membranes. Biochim Biophys Acta. 1981;649:125–128. doi: 10.1016/0005-2736(81)90017-1. [DOI] [PubMed] [Google Scholar]
- Kermode AR. Approaches to elucidate the basis of desiccation tolerance in seeds. Seed Sci Res. 1997;7:75–95. [Google Scholar]
- Knowles PF, Marsh D, Rattle HWE. Magnetic Resonance of Biomolecules: An Introduction to the Theory and Practice of NMR and ESR in Biological Systems. London: John Wiley & Sons; 1976. [Google Scholar]
- Larson RA. The antioxidants of higher plants. Phytochemistry. 1988;27:969–978. [Google Scholar]
- Leprince O, Atherton NM, Deltour R, Hendry GAF. The involvement of respiration in free radical processes during loss of desiccation tolerance in germinating Zea mays L.: an electron paramagnetic resonance study. Plant Physiol. 1994;104:1333–1339. doi: 10.1104/pp.104.4.1333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leprince O, Buitink J, Hoekstra FA. Radicles and cotyledons of recalcitrant seeds of Castanea sativa Mill. exhibit contrasting responses of respiration to drying in relation to desiccation sensitivity. J Exp Bot. 1999;338:1515–1524. [Google Scholar]
- Leprince O, Harren FJM, Buitink J, Alberda M, Hoekstra FA. Metabolic dysfunction and unabated respiration precede the loss of membrane integrity during dehydration of germinating radicles. Plant Physiol. 2000;122:597–608. doi: 10.1104/pp.122.2.597. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leprince O, Hendry GAF, McKersie BD. The mechanisms of desiccation tolerance in developing seeds. Seed Sci Res. 1993;3:231–246. [Google Scholar]
- Leprince O, Hoekstra FA. The response of cytochrome redox state and energy metabolism to dehydration support a role for cytoplasmic viscosity in desiccation tolerance. Plant Physiol. 1998;118:1253–1264. doi: 10.1104/pp.118.4.1253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leprince O, Vertucci CW, Hendry GAF, Atherton NM. The expression of desiccation-induced damage in orthodox seeds is a function of oxygen and temperature. Physiol Plant. 1995;94:233–240. [Google Scholar]
- Marsh D. Electron spin resonance: spin labels. In: Grell E, editor. Membrane Spectroscopy: Molecular Biology, Biochemistry and Biophysics. Vol. 31. Berlin: Springer-Verlag; 1981. pp. 51–142. [DOI] [PubMed] [Google Scholar]
- Marshall AG. Biophysical Chemistry: Principles, Techniques, and Applications. New York: John Wiley & Sons; 1978. [Google Scholar]
- Maher P, Singer SJ. Structural changes in membranes produced by the binding of small amphipathic molecules. Biochemistry. 1984;23:232–240. doi: 10.1021/bi00297a010. [DOI] [PubMed] [Google Scholar]
- Murphy DJ, Cummins I. Seed oil-bodies: isolation, composition and role of oil-body apolipoproteins. Phytochemistry. 1989;28:2063–2069. [Google Scholar]
- Oliver AE, Hincha DK, Crowe LM, Crowe JH. Interactions of arbutin with dry and hydrated bilayers. Biochim Biophys Acta. 1998;1370:87–97. doi: 10.1016/s0005-2736(97)00246-0. [DOI] [PubMed] [Google Scholar]
- Pammenter NW, Berjak P. A review of recalcitrant seed physiology in relation to desiccation tolerance mechanisms. Seed Sci Res. 1999;9:13–37. [Google Scholar]
- Ravanel P, Creuzet S, Tissut M. Inhibitory effect of hydroxyflavones on the exogenous NADH dehydrogenase of plant mitochondrial inner membranes. Phytochemistry. 1989;29:441–445. [Google Scholar]
- Rice-Evans CA, Miller NJ. Antioxidant activities of flavonoids as bioactive components of food. Biochem Soc Trans. 1996;24:790–795. doi: 10.1042/bst0240790. [DOI] [PubMed] [Google Scholar]
- Rice-Evans CA, Miller NJ, Paganga G. Antioxidant properties of phenolic compounds. Trends Plant Sci. 1997;2:152–159. [Google Scholar]
- Roozen MJGW, Hemminga MA, Walstra P. Molecular motion in glassy water-malto-oligosaccharide (matrodextrin) mixtures as studied by conventional and saturation-transfer spin-probe ESR spectroscopy. Carbohydr Res. 1991;215:229–237. [Google Scholar]
- Rowe ES, Zhang F, Leung TW, Parr JS, Guy PT. Thermodynamics of membrane partitioning for a series of n-alcohols determined by titration calorimetry: role of hydrophobic effects. Biochemistry. 1998;37:2430–2440. doi: 10.1021/bi9721602. [DOI] [PubMed] [Google Scholar]
- Saija A, Scalese M, Lanza M, Marzullo D, Bonina F, Castelli F. Flavonoids as antioxidant agents: importance of their interaction with biomembranes. Free Radic Biol Med. 1995;19:481–486. doi: 10.1016/0891-5849(94)00240-k. [DOI] [PubMed] [Google Scholar]
- Senaratna T, McKersie BD. Loss of desiccation tolerance during seed germination: a free radical mechanism of injury. In: Leopold AC, editor. Membranes, Metabolism and Dry Organisms. Ithaca, NY: Cornell University Press; 1986. pp. 85–101. [Google Scholar]
- Shirley BW. Flavonoids in seeds and grains: physiological function, agronomic importance and the genetics of biosynthesis. Seed Sci Res. 1998;8:415–422. [Google Scholar]
- Simon SA, Stone WL, Bennett PB. Can regular solutes theory be applied to lipid bilayer membranes? Biochim Biophys Acta. 1979;550:38–47. doi: 10.1016/0005-2736(79)90113-5. [DOI] [PubMed] [Google Scholar]
- Takahashi L, Sert MA, Kelmer-Bracht AM, Bracht A, Ishii-Iwamoto EL. Effects of rutin and quercetin on mitochondrial metabolism and on ATP levels in germinating tissues of Glycine max. Plant Physiol Biochem. 1998;36:495–501. [Google Scholar]
- Tinocco I, Jr, Sauer K, Wang JC. Physical Chemistry: Principles and Applications in Biological Sciences. Englewood Cliffs, NJ: Prentice-Hall; 1995. [Google Scholar]
- Ultee A, Gorris LGM, Smid EJ. Bactericidal activity of carvacrol towards the foodborne pathogen Bacillus cereus. J Appl Microbiol. 1998;85:211–218. doi: 10.1046/j.1365-2672.1998.00467.x. [DOI] [PubMed] [Google Scholar]
- Ultee A, Kets EPW, Smid EJ. Mechanisms of action of carvacrol on the foodborne pathogen Bacillus cereus. Appl Environ Microbiol. 1999;65:4606–4610. doi: 10.1128/aem.65.10.4606-4610.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vertucci CW, Farrant JM. Acquisition and loss of desiccation tolerance. In: Kigel J, Galili G, editors. Seed Development and Germination. New York: Marcel Dekker; 1995. pp. 237–271. [Google Scholar]
- White SH, Wimley WC. Peptides in lipid bilayers: structural and thermodynamic basis for partitioning and folding. Curr Opin Struct Biol. 1994;4:79–86. [Google Scholar]
- Wolf AV, Brown MG, Prentiss PG. Concentrative properties of aqueous solutions: conversion tables. In: Weast RC, Astle MJ, Beyer WH, editors. CRC Handbook of Chemistry and Physics. 67th Ed. Boca Raton, FL: CRC Press; 1986. pp. D219–D271. [Google Scholar]
- Wolkers WF. The role of macromolecular stability in desiccation tolerance. PhD thesis. The Netherlands: Wageningen University; 1998. [Google Scholar]