

Abstract
The parietal cell (PC) plays an important role in normal gastric physiology and in common diseases of the stomach. Although the genes involved in acid secretion are well known, there is limited molecular information about other aspects of PC function. We have generated a comprehensive database of genes expressed preferentially in PCs relative to other gastric mucosal cell lineages. PCs were purified from FVB/N mouse stomachs by lectin panning. cRNA generated from PC-enriched (PC+) and PC-depleted (PC−) populations were used to query oligonucleotide-based microarrays. False-positive signals were filtered by using a new algorithm for noise reduction and selected results independently audited by real-time quantitative reverse transcription (RT)-PCR. The annotated database of 240 genes reveals previously unappreciated aspects of cellular function, including factors that may mediate PC regulation of gastric stem cell proliferation. PC+ and PC− expression profiles were also prepared from germ-free mice 2 and 8 weeks after colonization with a clinical isolate of Helicobacter pylori (Hp)—the pathogen that produces acid-peptic disease (gastritis, ulcers) in humans. Whereas PC+ gene expression was remarkably constant, the PC− fractions demonstrated a robust, evolving host response, with increased expression of genes involved in cell motility/migration, extracellular matrix interactions, and IFN responses. The consistency of PC+ gene expression allowed identification of a cohort of 92 genes enriched in PCs under all conditions studied. These genes provide a molecular profile that can be used to define this epithelial lineage under a variety of physiologic, pharmacologic, and pathologic stimuli.
Keywords: genome anatomy project‖gnotobiotic mice
In the mouse and human stomach, epithelial renewal occurs continuously in anatomically distinct, tubular-shaped mucosal invaginations known as gastric units. The multipotent gastric stem cell resides in the middle region (isthmus) of each unit (1), and gives rise to three principal descendant lineages: mucus-producing pit cells that differentiate as they migrate apically (2), zymogenic cells that differentiate during a downward migration to the base of the gastric unit (3), and parietal cells (PCs), which, unlike the other lineages, differentiate within the isthmus and then migrate apically and basally (4).
In this report, we describe an approach for retrieving PCs from the intact stomach in a fashion that allowed us to obtain a comprehensive and consistent molecular profile of PC gene expression under a variety of well-defined environmental conditions. This approach, applicable to other epithelial lineages, represents the first contribution to a mouse Gastric Genome Anatomy Project (mG-GAP) designed to provide molecular characterization of all of the principal gastric epithelial cell lineages.
The PC was selected to initiate mG-GAP for several reasons. First, this highly specialized acid-secreting cell has attracted a great deal of interest because of the common occurrence of acid peptic disease in humans. Second, although its acid-secreting pathways have been studied extensively and are the targets of numerous drugs, other PC functions remain poorly characterized. For example, genetically engineered ablation of PCs in transgenic mice blocks terminal differentiation of zymogenic cells and increases proliferation of the multipotent stem cell and its immediate committed daughters (5, 6). Genetic mosaic analysis of mice containing mixtures of gastric units with or without parietal cell ablation revealed that these effects occur only in units lacking PCs, leading to the hypothesis that PCs elaborate locally acting factors that shape the stem cell niche and regulate zymogenic cell differentiation (6). Third, PCs were selected because they are abundant in the gastric epithelium and produce unique surface glycans that can be used for purification.
One goal of functional genomics is to generate searchable, annotated databases of genes expressed in normal cells so that the full breadth of cell biological activities can be inferred (7). Another is to use this information to detect and define disordered cellular function in disease states, with the expectation that such information will lead to earlier and more accurate diagnoses, to more specific therapies, and to more precise monitoring of therapeutic responses. Therefore, our analysis of PC gene expression was extended to germ-free (GF) mice, and ex-germ-free mice that had been colonized for 2 or 8 weeks with Helicobacter pylori (Hp). Hp colonizes ≈50% of humans, and produces severe pathology, including gastric and duodenal ulcers, in a subset of its hosts (8, 9). The effect of Hp on PC function is controversial, and the relative contributions of PC and non-PC cell lineages to the pathogenesis of gastritis and ulcer disease remain unclear. Using DNA microarrays, we demonstrate the remarkable stability of PC gene expression during Hp infection, and identify a broad repertoire of host responses induced in non-acid-secreting cells.
Materials and Methods
Isolation of PC-Enriched and PC-Depleted Populations from Conventionally Raised FVB/N Mice.
Six 6- to 12-week-old animals (equivalent numbers of males and females) were used per preparation (n = 6 independent preparations). Stomachs were excised, the proximal third (forestomach) discarded, the remaining two thirds opened, and the glandular mucosa was recovered by scraping. The scraped mucosa was minced under ice-cold HBSS-Hepes [Hanks buffered saline solution/10 mM Hepes/1× minimum essential amino acids (Invitrogen)/1 mM glutamine/0.1% BSA/0.25 μg/ml amphotericin B/100 units/ml penicillin/100 μg/ml streptomycin, pH 7.4]. Minced material from three stomachs was pooled. The two pools generated from the six stomachs were then processed in parallel as follows: (i) two washes each by centrifugation (200 × g, 2 min, 4°C; 10 ml HBSS-Hepes/wash); (ii) 90 min incubation (4°C) in 10 ml of pronase (Roche) digestion solution [1 mg enzyme/ml HBSS-HCO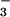 (HBSS as above with 12 mM sodium bicarbonate in lieu of Hepes)]; (iii) agitation for 10 min [orbital shaker (200 rpm), 37°C]; (iv) incubation for 5 min (23°C) to allow intact tissue fragments to settle by gravity; (v) removal of the upper 7 ml of the resulting supernatant (defined as “digestate” and collected into a fresh tube on ice); and (vi) replacement of the removed digestate with an equal volume of a solution containing 1 mg (125 units) crude type IA collagenase (Sigma) per ml HBSS-HCO
(HBSS as above with 12 mM sodium bicarbonate in lieu of Hepes)]; (iii) agitation for 10 min [orbital shaker (200 rpm), 37°C]; (iv) incubation for 5 min (23°C) to allow intact tissue fragments to settle by gravity; (v) removal of the upper 7 ml of the resulting supernatant (defined as “digestate” and collected into a fresh tube on ice); and (vi) replacement of the removed digestate with an equal volume of a solution containing 1 mg (125 units) crude type IA collagenase (Sigma) per ml HBSS-HCO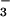 followed by another round of steps iii–v.
followed by another round of steps iii–v.
Digestates were collected from a total of 4 rounds of steps iii–vi. As each digestate was collected, it was immediately triturated, the suspension passed through a nylon filter (30 μm pore; Millipore), and enzyme activity in the filtrate quenched by transfer to 15 ml ice-cold HBSS/10% calf serum. After the pooled, quenched solution was centrifuged (1000 × g, 8 min, 4°C), the pellet was resuspended (in 4 ml ice-cold HBSS-HCO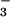 plus 4 ml 2% BSA in Ca2+/Mg2+-free HBSS) and filtered (20 μm pore).
plus 4 ml 2% BSA in Ca2+/Mg2+-free HBSS) and filtered (20 μm pore).
The filtrate was incubated (15 min, 4°C) with magnetic beads conjugated to Dolicos biflorus agglutinin (DBA, EY Laboratories; ≈50 beads per parietal cell). Beads were prepared by incubating 2.5 μM biotinylated DBA in HBSS/2% BSA with streptavidin-conjugated magnetic beads (Polysciences) for 15 min (23°C), followed by several washes in HBSS/2% BSA. DBA-magnetic bead-coated cells were pelleted (4 min; magnetic tube stand packed in ice). Two more cycles of lectin panning were performed. Cells in the final pellet were lysed by needle trituration in 350 μl RLT buffer (Qiagen, Chatsworth, CA) before RNA extraction (see below).
To generate PC-depleted (PC−) cell populations, supernatants from the first two rounds of magnetic bead-DBA lectin panning were pooled, a fresh aliquot of beads was added, and after a 10 min incubation (4°C), contaminating PCs were removed by magnetic separation. The supernatant was centrifuged (1000 × g, 8 min, 4°C), the pellet resuspended in 1.4 ml HBSS/2% BSA, and subjected to a final round of lectin panning. Cells in the supernatant were pelleted (1500 × g, 5 min) and lysed as above.
Cells from PC+ and PC− preparations were smeared onto glass slides, fixed in 70% ethanol, rehydrated in PBS, and stained using a modified Maxwell technique (10) to score the fractional representation of parietal, zymogenic, and mucus-producing cells.
Recovery of PC+ and PC− Fractions from GF and Ex-GF FVB/N Mice.
GF mice were maintained in plastic gnotobiotic isolators on a 12-h light cycle, and given free access to autoclaved chow (B & K Universal). At 6 weeks of age, GF animals were gavaged with 107 colony forming units (cfu) of a clinical H. pylori isolate (Hp1; ref. 11). The resulting ex-GF mice (referred to below as ex-GF-Hp+) were killed 2 or 8 weeks later (n = 6 mice per experiment; three independent experiments per time point). PC+ and PC− fractions were prepared from ex-GF-Hp+ and age-matched GF controls using a protocol identical to that employed for conventionally raised mice, except that a 2- × 2-mm fragment from the distal stomach (antrum) was taken to quantitate Hp1 cfu (11).
DNA Microarray Studies.
RNA was isolated from PC+ and PC− lysates using the RNeasy kit (Qiagen). Equivalent amounts of RNA from each preparation were combined to create PC+ and PC− pools representing each condition (i.e., conventionally raised, GF, ex-GF-Hp+). cRNA targets were generated (12) and hybridized to Mu11K GeneChip sets (Affymetrix, Santa Clara, CA). Overall fluorescence intensity across each chip was scaled (intensity = 150) using genechip software. Pairwise comparisons of the levels of transcripts in PC+ vs. PC− RNAs were performed as described below.
SYBR-Green-Based Real Time Quantitative Reverse Transcription (qRT)-PCR.
qRT-PCR was performed with gene-specific primers (see supporting information on the PNAS web site, www.pnas.org; or see http://gordonlab.wustl.edu/mills/parietalcells) and oligo(dT)-primed cDNA synthesized from DNase-treated RNAs. Each reaction contained 1× SYBR Green PCR Master Mix buffer (Applied Biosystems), 0.25 units UDP-N-glycosidase (Life Technologies, Grand Island, NY), 900 nM forward and reverse primers, and cDNA. A melting curve was used to identify a temperature where only amplicon, and not primer dimers, accounted for SYBR Green-bound fluorescence. Assays were performed in triplicate using an Applied Biosystems Model 7700 instrument. All data were normalized to an internal glyceraldehyde-3-phosphate dehydrogenase mRNA control (ΔΔCT analysis).
Results and Discussion
Generating a Database of Genes Preferentially Expressed in Parietal Cells.
To generate a comprehensive database of mRNAs enriched in PCs relative to other cell types in the adult FVB/N mouse stomach, we took advantage of the fact that only the PC lineage expresses GalNAcα(1–3)Gal-containing glycans recognized by DBA (13). Lectin panning with magnetic bead-DBA conjugates was used to purify PCs from dispersed gastric mucosa harvested from conventionally raised mice. In each of six independently prepared PC+ fractions, >80% of all cells were PCs. In contrast, PCs accounted for <5% of all cells in PC− fractions (Fig. 1). The PC− fraction contained pit cells (35%), neck and zymogenic cells (22 and 26%, respectively), plus other mucosal constituents (epithelial precursors, fibroblasts, and endothelial cells).
Figure 1.

Histologic characterization of parietal cell-enriched and parietal cell depleted fractions prepared by magnetic bead-lectin panning. Cells from PC+ fraction (A) and PC− fraction (B) were stained with rabbit antibodies to the noncatalytic β-subunit of mouse H+/K+-ATPase (labeled green with FITC-conjugated donkey anti-rabbit Ig). Nuclei were stained blue with bis-benzimide. Bars = 20 μm. (C) Transmission EM of two parietal cells in the PC+ fraction showing the integrity of the canalicular apparatus (closed arrow), the abundance of mitochondria (arrowhead), and binding of magnetic bead-DBA conjugates to the cell surface (open arrow). Magnification ×3300.
RNAs were pooled from the six independent preparations of each fraction (n = 6 mice per preparation), and cRNAs were used to interrogate Affymetrix Mu11K GeneChips containing probe sets representing ≈11,000 mouse genes and expressed sequence tag (EST) clusters. Chip-to-chip comparisons of PC+ to PC− samples were performed with the PC− chips designated as “Baseline.” An initial dataset of PC+-enriched transcripts was generated by extracting all mRNAs whose levels were called “Increased” over baseline by genechip software. Defining PC gene expression relative to the PC− population was appealing for several reasons. It minimized representation of genes in the database that might be up-regulated in all cells because of the isolation procedure itself, or that encoded products required for general housekeeping functions. Furthermore, because the composition of contaminating cells in the PC+ fraction was equivalent to the composition of the PC− fraction itself (data not shown), subtracting PC− transcripts from the PC+ pool alleviated the need for a completely homogeneous PC fraction.
Noise filtration.
GeneChips produce a false-positive rate of 1–2% (14, 15). The most common way to filter false-positives from genechip comparisons is to impose a threshold requirement for ≥2-fold change in the level of a transcript under the experimental conditions studied (16, 17). However, setting such a threshold is arbitrary and risks eliminating genes with biologically significant, albeit modest, differences in expression (18, 19).
In light of these concerns, we designed a system for filtering false-positives based on the signal intensities of genes whose expression level is called “changed” in chip-to-chip comparisons of an identical RNA population (n = 70 comparisons involving >30 independently generated RNA preparations from a variety of biological systems listed in ref. 20). All changes in expression in such “same-same” comparisons were defined as false-positives. The distribution of signal intensities of the false-positives in baseline and partner Mu11K chips was used to develop a series of look-up tables (LUTs). The LUTs can be used to score transcripts whose expression level is called “changed” by genechip software in comparisons of biologically distinct RNAs. LUT-derived scores range from 0 to 6, with 0 most likely and 6 least likely to represent noise (20). Eliminating transcripts with LUT scores of <4 reduces false-positives by 90% (see ref. 20 for details), and increases experiment-to-experiment reproducibility to above that obtained by imposing an arbitrary ≥2-fold change threshold (LUTs and software needed to score datasets are deposited at http://gordonlab.wustl.edu/mills).
The Database.
A total of 647 probe sets in Mu11K GeneChips were called “Increased” by genechip software in the PC+ to PC− comparison. Two hundred eighty-nine of these probe sets, representing 240 genes, had LUTs ≥4. This noise-filtered dataset was annotated and organized into functional categories. A complete list of the genes can be found in the supporting information on the PNAS web site or at http://gordonlab.wustl.edu/mills/parietalcells, with a subset presented in Table 1. Only 8% have been previously reported as expressed in PCs. Twelve percent of the database is composed of ESTs with no discernible homology to known genes.
Table 1.
Genes preferentially expressed in mouse parietal cells
| Gene | UniGene UID | Previous PC Localization |
|---|---|---|
| Energetics/metabolism-related | ||
| Non-mitochondrial | ||
| Glucose transporter 4 | Mm.10661 | New |
| Phosphoglycerate mutase (Brain form, PGAM-B) | Mm.16783 | Hkp |
| Cytosolic malate dehydrogenase (pmc-MDH-5/mdh1) | Mm.3156 | Hkp |
| Creatine kinase, brain | Mm.16831 | Hkp |
| Alpha-(nonneural) enolase | Mm.90587 | Mito |
| Lactate dehydrogenase B,H chain | Mm.9745 | Mito |
| Phosphofructokinase, muscle | Mm.26550 | Hkp |
| Galactose-1-phosphate uridyl transferase | Mm.2420 | Hkp |
| Mitochondrial: Matrix/TCA | ||
| Pyruvate dehydrogenase dihydrolipoamide acetyltransferase | Mm.38786 | Mito |
| Mitochondrial ribosomal protein S21 | Mm.24108 | Mito |
| Branched chain ketoacid dehydrogenase E1, alpha | Mm.25848 | Mito |
| Dihydrolipoamide dehydrogenase | Mm.3131 | Mito |
| Aconitase 2, mitochondrial | Mm.154581 | Mito |
| Mitochondrial: Oxidative Phosphorylation | ||
| Complex I: 7 subunits | Multiple | Mito |
| Complex II: 2 subunits | Multiple | Mito |
| Complex III: 3 subunits | Multiple | Mito |
| Complex IV: 1 subunit | Mm.2151 | Mito |
| F1F0 ATP Synthase, 4 subunits | Multiple | Mito |
| Other Mitochondrial | ||
| Mitochondrial CGI-69 | Mm.44226 | Mito |
| ATP/ADP translocase | Mm.16228 | Mito |
| Mitofilin/Motor protein | Mm.183073 | Mito |
| Acid Secretion-related | ||
| Vesicle Traffic | ||
| Sorting nexin 6/TRAF4 assoc. Factor 2 | Mm.196554 | New |
| Glioblastoma amplified sequence (Gbas)/NIPSNAP2 | Mm.12468 | New |
| Prenylated RAB acceptor 1 (PRA1) | Mm.22473 | New |
| Channel/Pump-related | ||
| Potassium voltage-gated channel, subfamily Q, member 1 | Mm.5177 | New |
| Anion exchanger 2 (Sic4a2) | Mm.4580 | PC |
| Chloride channel 3 (Clcn3) | Mm.28842 | New |
| Aquaporin 4 | Mm.1419 | PC |
| Na/K Transporting ATPase (alpha-1) | N/A | PC |
| Detoxification/degradation/scavenging | ||
| Peroxiredoxin 5 (Prdx5)/Thioredoxin peroxidase PMP20 | Mm.6587 | Mito |
| Aldehyde dehydrogenase II (AHD-2 or ALDH1) | Mm.4514 | New |
| Peroxiredoxin 2/Thioredoxin peroxidase (Tpx) | Mm.42948 | PC |
| Superoxide dismutase 2, mitochondrial | Mm.2597 | Mito |
| Cytoskeletal-associated | ||
| Mu-protocadherin/MUCDHL | Mm.29080 | New |
| Reversion Induced LIM gene | Mm.21830 | New |
| Huntingtin interacting protein 1 related | Mm.149954 | New |
| Fatty acid metabolism | ||
| Absorption/carriers | ||
| Lipoprotein lipase | Mm.1514 | New |
| Heart fatty acid binding protein (H-FABP) | Mm.22220 | PC |
| Liver fatty acid binding protein (L-FABP) | Mm.22126 | PC |
| Membrane Maintenance | ||
| MLD fatty acid desaturase/Mdes | Mm.29648 | New |
| Elongation of very long chain fatty acids-like 1 (MUEL) | Mm.29492 | New |
| Phospholipid hydroperoxide glutathione peroxidase | Mm.2400 | New |
| Synthesis | ||
| Mitochondrial glycerol-3-phosphate acyltransferase | Mm.87773 | Mito |
| Mitochondrial acetoacetyl-CoA thiolase | N/A | Mito |
| Oxidation | ||
| Propionyl CoA carboxylase, Beta subunit | Mm.21079 | Mito |
| Methylmalonyl CoA mutase | Mm.4299 | Mito |
| Isocitrate dehydrogenase 2 (NADP+) | Mm.2966 | Mito |
| AU-specific RNA-binding enoyl-CoA hydratase (AUH) | Mm.2733 | Mito |
| Hydroxylacyl-Coenzyme A dehydrogenase | Mm.2491 | Mito |
| Acetyl-Coenzyme A dehydrogenase, long-chain | Mm.2445 | Mito |
| Acyl-Coenzyme A dehydrogenase, very long chain | Mm.18630 | Mito |
| Mitochondrial Carriers | ||
| Carnitine-O-acetyltransferase | Mm.20396 | Mito |
| Calcineurin pathway-related | ||
| Calcium Stores | ||
| Calcineurin inhibitor/Down syndrome candidate region 1 | Mm.56 | New |
| Steroid Receptor | ||
| Retinol dehydrogenase 7 | Mm.6696 | New |
| shp nuclear receptor (subfamily 0, group B, member 2) | Mm.34209 | New |
| Peroxisomal phytanoyl-CoA alpha-hydroxylase (PAHX) | Mm.27066 | New |
| Census-regulating | ||
| Parathyroid hormone-related peptide | N/A | New |
| Insulin-like growth factor binding protein 2 (IGFBP-2) | N/A | New |
| Vascular endothelial growth factor B (VEGF-B) | Mm.15607 | New |
| CD36 antigen | Mm.18628 | New |
| Signal transduction | ||
| Cytoplasmic/General | ||
| Interferon-regulated gene 1 (IRG1) | Mm.2121 | New |
| Sex-regulated protein janus (HSPC141 or CDI202) | Mm.177990 | New |
| T-cell death-associated gene 51 (TDAG51) | Mm.3117 | New |
| Nuclear/Transcription Factors | ||
| Retinoblastoma-binding protein 9 | Mm.24216 | New |
| Oncostatin M-inducible protein OIG37/Gadd45 gamma | Mm.9653 | New |
| Amino-terminal enhancer of split (AES1) | Mm.2626 | New |
| Miscellaneous | ||
| Nuclear ribonuclease P subunit (RPP21) | Mm.27131 | New |
| Mouse MCJ | Mm.27692 | New |
| Prion protein | Mm.648 | New |
The core group of genes consistently expressed in a preferential fashion in PCs purified from conventionally raised, GF, and ex-GF FVB/N mice colonized for 2 or 8 weeks with H. pylori strain Hp1. See the supporting information on the PNAS web site for additional details (fold differences between PC+ and PC−, LUT scores, gene-specific annotation, reference hyperlinks, unlisted ESTs). New, not previously reported in PCs; PC, previously reported in parietal cells; Hkp, general housekeeping; Mito, mitochondrial.
The list of preferentially expressed genes provides a view of the molecular underpinnings of the biological functions carried out by PCs. For example, proton pumps distributed across the PC plasma membrane consume a great deal of energy: 35% of the entries in the database (85 genes) are involved in various aspects of cellular energy metabolism; 66 encode mitochondrial proteins. Another 14% were categorized as acid secretion/vesicular trafficking related. Some entries in the last category, such as ADP-ribosylation factor 1 (ARF1) and ezrin, are known PC products, whereas others—such as prenylated Rab acceptor 1, SARa GTPase, endobrevin, glioblastoma amplified sequence (NIPSNAP2), and sorting nexin 6—to our knowledge have not previously been reported as PC products.
PCs are also enriched in mRNAs encoding free radical scavengers that protect against potential damage from high rates of oxidative phosphorylation. Peroxiredoxin 2, glutathione peroxidase 4, mitochondrial superoxide dismutase, and catalase 1 are examples.
PCs maintain an intricate tubulovesicular membrane apparatus for exporting acid. Moreover, their large population of mitochondria use fatty acids to generate energy required for proton secretion. Accordingly, the PC+ database contains 21 genes involved in various aspects of lipid metabolism, including fatty acid absorption, synthesis, β-oxidation, and mitochondrial transport (see Table 1 and the supporting information on the PNAS web site and at http://gordonlab.wustl.edu/mills/parietalcells). Four genes have roles in maintaining membrane bilayers: fatty acid desaturase, glutathione peroxidase 4, very long chain fatty acid elongase, and oxysterol binding protein.
Calcium signaling is intimately involved in the regulation of acid secretion (21). Eleven genes preferentially expressed in PCs encode constituents of pathways involving the ubiquitous calcium-dependent protein phosphatase calcineurin (PP3). Two aspects of PP3 function are represented: regulation of calcium-related signaling pathways (e.g., calcineurin inhibitor (Dscr1); calmodulin-2, and myosin regulatory light chain), and steroid hormone signal transduction (e.g., serum glucocorticoid-regulated kinase and FK506 binding protein 4; note that PCs have recently been shown to express glucocorticoid receptors; ref. 22).
The PC+ database reveals five genes that may regulate cellular census in gastric units: none have been previously reported as being expressed in PCs. Insulin-like growth factor binding protein-2 (IGFBP-2) is a secreted protein that reduces bioavailability of IGFs (23, 24). Low molecular weight GH receptor is produced by alternative splicing and functions as a secreted GH binding protein (designated GHBP; refs. 25 and 26). PTH-like (related) peptide (PTHLP) affects cell division in a number of systems (27, 28). Elimination of PCs from the isthmal stem cell niche of gastric units (as in tox176 transgenic mice; see introduction) could alter proliferation of lineage progenitors secondary to loss of these three PC-derived growth regulators. Vascular endothelial growth factor-B (VEGF-B) mRNA is also enriched in PCs. VEGF-B stimulates angiogenesis in growing tissues and could help PCs maintain blood supply in isthmal and other regions of gastric units. CD36 is a membrane-bound lipid scavenger that functions in cell–cell communication and phagocytosis of apoptotic bodies (29, 30).
Real-time qRT-PCR provided independent confirmation that IGFBP-2, GH binding protein, PTHLP, VEGF-B, and CD36 mRNAs are enriched in PC+ compared with PC− fractions (Fig. 2). Two genes known to be preferentially expressed in PCs (the catalytic α subunit of H+/K+ ATPase; aquaporin 4), as well as an EST (AA597222) we identified as one of the most highly enriched transcripts in PC+ RNA, were used as reference controls in the qRT-PCR analysis (data not shown). Additional evidence that parietal cells are the principal sources of IGFBP, GH binding protein, and PTHLP in vivo came from the results of a qRT-PCR analysis that showed that these three mRNAs are reduced in the intact PC-ablated stomachs of tox176 mice compared with their normal littermates (Fig. 2).
Figure 2.

Expression of parietal cell-associated genes in PC+ vs. PC− fractions, and in the intact stomachs of normal mice and tox176 transgenic animals with an engineered ablation of the PC lineage. For GeneChip and qRT-PCR comparisons of expression in PC+ vs. PC− fractions, the level of each mRNA in the PC− fraction was arbitrarily set at 1. For qRT-PCR comparisons of gene expression in the stomachs of conventionally raised normal and tox176 mice, mRNA levels in tox176 RNA were set at 1. Error bar = 1 SD (n = four 14- to 16-week-old mice per group; at this age, multi and oligo-potential lineage progenitors comprise 1% of the total epithelial cell population in gastric units of normal mice and ≥10% in tox176 units).
A Gnotobiotic Mouse Model for Comparing the Responses of PC+ and PC− Populations to H. pylori Colonization.
In most humans, H. pylori (Hp) infection is associated with development of mild gastritis, but ≈10% of hosts develop severe pathology, including gastric and duodenal ulcers, gastric carcinoma, and B cell mucosa-associated lymphoid tissue (MALT) lymphoma (8, 9). The effects of Hp on PC functions during the early phases of infection remain unclear. Moreover, genetic variations among humans and their colonizing strains have made it difficult to develop hypotheses about the relative contributions of environmental, host, and bacterial factors to the destiny of Hp infection.
We reasoned that genetically and environmentally defined GF FVB/N mice, colonized with an Hp isolate, could be used in concert with the functional genomic approaches described above to obtain a comprehensive set of molecular markers of the host response. Such markers could then be used in future studies to define the impact of bacterial genotypes on the evolution of host responses or the effects of various therapeutic interventions. Therefore, we characterized gene expression in PC+ and PC− populations harvested from GF FVB/N mice inoculated with a clinical isolate of H. pylori and from untreated age-matched GF controls. This well characterized clinical isolate (Hp1) was recovered from a patient with gastritis and had been subjected to whole genome genotyping with DNA microarrays (31). A single gavage of 107 cfu of Hp1 produces persistent infection with focal gastritis in conventionally raised FVB/N mice (6, 11). We used GF rather than conventionally raised mice because, unlike humans, mice normally have an abundant and ill-defined gastric microflora. In the absence of an entrenched microflora, Hp1 colonizes the stomachs of GF FVB/N mice with 100% efficiency (n = 40), a critical feature when cell populations must be pooled from multiple stomachs immediately after sacrifice. Moreover, any changes in gene expression observed in PC+ and/or PC− populations recovered from ex-GF mice can be attributed to the introduced strain itself, rather than to perturbations in the composition of the indigenous microflora.
GF FVB/N mice were inoculated with 107 cfu of Hp1 at 6 weeks of age. The resulting ex-GF-Hp+ animals were initially killed 2 weeks later, together with a control group of 8-week-old GF mice. PC+ or PC− RNAs, pooled from ex-GF-Hp+ mice or GF controls (n = 6 mice per group; n = 3 independent experiments), were used to produce cRNAs. None of the ≈11,000 genes represented on the Mu11K GeneChips satisfied all of the following criteria: (i) increased in the PC+ populations of ex-GF-Hp+ relative to GF mice; (ii) preferentially expressed in the PC+ compared with PC− populations of ex-GF-Hp+ mice; (iii) LUT scores ≥4; and (iv) independent confirmation of i by qRT-PCR. (Criterion i was intended to identify PC+ genes whose expression increased in the presence of Hp1 whereas ii was used to verify that genes selected from i were preferentially expressed in parietal cells.)
In contrast, mRNAs identified by 275 probe sets were increased in Hp1-infected PC− compared with GF PC−, and in Hp1-infected PC− compared with Hp1-infected PC+ fractions, and also had LUT scores ≥4 (see the supporting information on the PNAS web site for a list of these probesets). Excluding transcripts encoding immunoglobulins, Hp1 induced expression of 99 unique genes. Only 16 have been previously associated with H. pylori infection (e.g., secretory leukocyte protease inhibitor, grb2, CD18, and L-selectin). These 16, and the 83 newly described host response genes, were placed into 8 categories: (i) related to IFN signaling (e.g., the endoplasmic reticulum-based IFN-γ-induced GTPase involved in protein processing in macrophages/fibroblasts); (ii) lipopolysaccharide (LPS)-regulated responses (e.g., cathepsin S); (iii) actin/cytoskeletal (e.g., prothymosin β4, regulates monomeric actin pools); (iv) extracellular matrix (ECM)/cell motility (e.g., integrin β2); (v) protein turnover (e.g., proteasome subunit α10); (vi) signal transduction (e.g., JAK3; Ets-1, an oncogene that up-regulates ECM binding proteins such as integrins and ECM degrading enzymes such as urokinase plasminogen activator; Pim1, a Serine/Threonine kinase associated with lymphomas); (vii) miscellaneous; and (viii) genes with unknown functions. One notable aspect of this early 2-week host response to Hp1 is that a number of the genes are known to be involved in the repair of damaged epithelia (cf. trefoil factors plus the constellation of proteins regulating cell movement listed in the PC− database in the supporting information on the PNAS web site; refs. 32 and 33).
To assess the specificity of the response of these 99 genes to colonization with Hp1, we examined which ones were also expressed at higher levels in conventionally raised compared with GF mice. Only 5 of the 99 were: trefoil factor 2, non-integrin laminin receptor, CD13, β-actin, melanoma-X-actin, and EST AA049952.
To assess the evolution of the host transcriptional response to Hp1, we extended our analysis to PC+ and PC− cells harvested from ex-GF-Hp+ mice colonized for 8 weeks and age-matched (14-week-old) GF controls. None of the 11,000 genes on the DNA microarrays satisfied the selection criteria described above for the PC+ fraction. Furthermore, qRT-PCR studies independently confirmed that expression of the catalytic α subunit of H+/K+ ATPase, aquaporin 4, IGFBP-2, PTHLP, VEGF-B, CD36, plus AA597222 varied <2-fold (Fig. 3; plus data not shown).
Figure 3.

qRT-PCR analysis of the fold-difference in levels of selected mRNAs in PC+ fractions or the intact stomachs of H. pylori-infected vs. germ-free FVB/N mice. Error bar = 1 SD.
Thirteen of the 99 transcripts enriched in ex-GF-Hp+ compared with GF PC− RNAs after a 2-week infection also remained enriched after an 8-week colonization (see http://gordonlab.wustl.edu/mills/parietalcells for a list of the 16 non-Ig-encoding genes increased at 8 weeks). Although the overall response of PC− cells to Hp1 was markedly attenuated after 8 compared with 2 weeks of infection, most of the functional categories listed above are represented: e.g., three genes are known to be related to Hp infection (two MHC II transcripts, secretory leukocyte protease inhibitor); 9 genes help regulate actin and mediate migration through the extracellular matrix (e.g., coronin).
The only transcripts in the 8-week Hp1-infected PC− dataset not represented in the 2-week PC− dataset are an EST, MARCKS-related protein, CEA-related protein 10, and Reg3γ. MARCKS-related protein is widely expressed and is largely responsible for coordinating actin rearrangement with cell signaling (34, 35). The function of CEA-related protein 10 remains unclear although it has been implicated in cell–cell adhesion (36). Reg3γ is a lectin expressed at high levels in the normal colon and in inflamed pancreas. It is up-regulated by bacterial lipopolysaccharide (LPS) and may play a protective role in bacterial infection (37, 38).
Further Evaluation of the Restricted PC+ Response to H. pylori Infection.
There are several reasons why the PC transcriptional response to Hp1, defined by normalized comparisons of PC+ and PC− fractions, might appear restricted. Infection could change the properties of PCs in ways that affect their survival before or during their purification, thereby producing shifts in the composition of the PC+ fraction recovered from the stomachs of GF vs. ex-GF-Hp+ mice. Two observations argue against this scenario. First, we were unable to detect compositional differences with a combination of histo- and immunohistochemical stains. Second, qRT-PCR analysis of RNAs prepared from the intact stomachs of GF and age-matched ex-GF mice colonized for 8 weeks with Hp1 (n = 3 per group), and from GF and Hp1-infected PC+ fractions, established that the relative levels of mRNAs encoding known PC markers were not significantly different (Fig. 3).
Another reason why the parietal cell-specific transcriptional response may appear restricted is that some genes are induced in both PC+ and PC− populations. A total of 12 mRNAs were increased in the direct GeneChip comparison between Hp1-infected PC+ and GF PC+ RNAs. None satisfied the criteria of being enriched in PC+ compared with PC− cells. Six of the 12 mRNAs were also increased in Hp1-infected PC− compared with GF PC−: polymeric Ig receptor (pIgR), MHC II H2-IA α, MHC II H2-IA β, Ia-associated invariant chain, L31 60s ribosomal protein, and an EST (AA547057). The PC+ fractions were not 100% pure. The responses of the MHC II H2-IA α, MHC II H2-IA β, Ia-associated invariant chain genes likely represent contributions from immune cell contaminants of the PC+ fraction. The remaining 6 members of the 12-gene list [L39 60s ribosomal protein, mitochondrial elongation factor Tu, basigin (an epithelial Ig family member involved in cell–cell contacts), protein E7 (AA518685), and a proline-rich protein (AA511380)] were not increased in PC+ vs. PC− under any of the conditions studied (germ-free, Hp1-infected, and conventionally raised).
Yet another possible reason for the restricted PC+ response is that there are sporadic changes in PCs, so that at any given time the PC population as a whole is not appreciably affected. However, the same functional genomics approach disclosed broad changes in the PC− population, indicating that, even if the response of the gastric mucosa to colonization is intermittent and/or focal, changes in host gene expression can be detected using this approach. We conclude, therefore, that the preponderance of host responses to Hp infection in this gnotobiotic model originates in the PC− fraction.
A Metaanalysis of Gene Expression in Parietal Cells.
The points raised in the preceding paragraphs about the transcription response of PCs to Hp1 infection led us to examine the consistency of expression of genes in the PC+ database under a variety of environmental conditions. We considered all mRNAs identified by the DNA microarray analysis as increased in PC+ versus PC− populations recovered from GF mice (at 8 and 14 weeks of age), from conventionally raised animals, and from ex-GF mice colonized with Hp1 for 2 or 8 weeks. Eighty-two percent of the transcripts in the conventionally raised PC+ dataset were present in datasets generated from at least one of the other conditions, while 38% were enriched in PC+ populations from all conditions (Table 1). This degree of similarity is remarkable given the degree of reproducibility reported for GeneChip data (i.e., 25–50% with duplicate chip-to-chip comparisons; refs. 20, 39, and 40). Further evidence of the constancy of gene expression was provided by qRT-PCR studies of PC+ RNAs prepared from GF and ex-GF mice. The results reveal ≤2-fold differences in levels of α-subunit of H+/K+ ATPase, IGFBP2, PTHLP, CD36, and EST AA597222 mRNAs among these preparations (data not shown).
The 92 transcripts (107 probe sets) enriched in PC+ populations recovered from all gastric environments sampled provide a functionally diverse profile of the molecular features of the parietal cell lineage. This constellation of molecular markers should be useful in defining this lineage under a variety of physiologic and/or pathologic conditions. Although our definition of parietal cell gene expression currently derives from a comparison with the multicomponent PC− population, we anticipate that a more refined molecular signature of PCs will be forthcoming as additional gastric epithelial lineages are purified and profiled.
Supplementary Material
Acknowledgments
We are indebted to Lora Hooper and Sabrina Wagoner for help in generating cRNA targets, Lisa Roberts for EM studies, and Jung Oh for assistance with RT-PCR assays. This work was supported by National Institutes of Health Grant DK58529. J.C.M. is the recipient of a Howard Hughes Medical Institute postdoctoral physician fellowship.
Abbreviations
- PC
parietal cell
- GF
germ-free
- Hp
Helicobacter pylori
- DBA
Dolicos biflorus agglutinin
- cfu
colony forming unit
- qRT-PCR
quantitative reverse transcription–PCR
- EST
expressed sequence tag
- LUT
look-up table
- IGFBP
insulin-like growth factor binding protein
- PTHLP
PTH-like (related) peptide
- VEGF-B
vascular endothelial growth factor-B
Footnotes
This paper was submitted directly (Track II) to the PNAS office.
References
- 1.Karam S M, Leblond C P. Anat Rec. 1993;236:259–279. doi: 10.1002/ar.1092360202. [DOI] [PubMed] [Google Scholar]
- 2.Karam S M, Leblond C P. Anat Rec. 1993;236:280–296. doi: 10.1002/ar.1092360203. [DOI] [PubMed] [Google Scholar]
- 3.Karam S M, Leblond C P. Anat Rec. 1993;236:297–313. doi: 10.1002/ar.1092360204. [DOI] [PubMed] [Google Scholar]
- 4.Karam S M. Anat Rec. 1993;236:314–332. doi: 10.1002/ar.1092360205. [DOI] [PubMed] [Google Scholar]
- 5.Li Q, Karam S M, Gordon J I. J Biol Chem. 1996;271:3671–3676. [PubMed] [Google Scholar]
- 6.Syder A J, Guruge J L, Li Q, Oleksiewicz C, Lorenz R G, Karam S M, Falk P G, Gordon J I. Mol Cell. 1999;3:263–274. doi: 10.1016/s1097-2765(00)80454-2. [DOI] [PubMed] [Google Scholar]
- 7.Phillips R L, Ernst R E, Brunk B, Ivanova N, Mahan M A, Deanehan J K, Moore K A, Overton G C, Lemischka I R. Science. 2000;288:1635–1640. doi: 10.1126/science.288.5471.1635. [DOI] [PubMed] [Google Scholar]
- 8.Nyren O. Semin Cancer Biol. 1998;8:275–283. doi: 10.1006/scbi.1998.0074. [DOI] [PubMed] [Google Scholar]
- 9.Feldman R. In: Helicobacter pylori: Molecular and Cellular Biology. Achtman M, Suerbaum S, editors. Norfolk, U.K.: Horizon Scientific Press; 2001. pp. 29–51. [Google Scholar]
- 10.Beinborn M, Giebel J, Linck M, Cetin Y, Schwenk M, Sewing K F. Cell Tissue Res. 1993;274:229–240. doi: 10.1007/BF00318742. [DOI] [PubMed] [Google Scholar]
- 11.Guruge J L, Falk P G, Lorenz R G, Dans M, Wirth H-P, Blaser M J, Berg D E, Gordon J I. Proc Natl Acad Sci USA. 1998;95:3925–3930. doi: 10.1073/pnas.95.7.3925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Lee C K, Klopp R G, Weindruch R, Prolla T A. Science. 1999;285:1390–1393. doi: 10.1126/science.285.5432.1390. [DOI] [PubMed] [Google Scholar]
- 13.Falk P, Roth K A, Gordon J I. Am J Physiol. 1994;266:G987–G1003. doi: 10.1152/ajpgi.1994.266.6.G987. [DOI] [PubMed] [Google Scholar]
- 14.Claverie J M. Hum Mol Genet. 1999;8:1821–1832. doi: 10.1093/hmg/8.10.1821. [DOI] [PubMed] [Google Scholar]
- 15.Lockhart D J, Winzeler E A. Nature (London) 2000;405:827–836. doi: 10.1038/35015701. [DOI] [PubMed] [Google Scholar]
- 16.Fambrough D, McClure K, Kazlauskas A, Lander E S. Cell. 1999;97:727–741. doi: 10.1016/s0092-8674(00)80785-0. [DOI] [PubMed] [Google Scholar]
- 17.Wang Y, Rea T, Bian J, Gray S, Sun Y. FEBS Lett. 1999;445:269–273. doi: 10.1016/s0014-5793(99)00136-2. [DOI] [PubMed] [Google Scholar]
- 18.Der S D, Zhou A, Williams B R, Silverman R H. Proc Natl Acad Sci USA. 1998;95:15623–15628. doi: 10.1073/pnas.95.26.15623. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Bassett D E, Jr, Eisen M B, Boguski M S. Nat Genet. 1999;21:51–55. doi: 10.1038/4478. [DOI] [PubMed] [Google Scholar]
- 20.Mills J C, Gordon J I. Nucleic Acids Res. 2001;29:e72. doi: 10.1093/nar/29.15.e72. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Urushidani T, Forte J G. J Membr Biol. 1997;159:99–111. doi: 10.1007/s002329900274. [DOI] [PubMed] [Google Scholar]
- 22.Kanemasa H, Ozawa H, Konishi H, Ito T, Nishi M, Mitsufuji S, Kodama T, Hattori T, Kawata M. Dig Dis Sci. 1999;44:2081–2087. doi: 10.1023/a:1026686705421. [DOI] [PubMed] [Google Scholar]
- 23.Pommier G J, Remacle-Bonnet M M, Tripier S G, Garrouste F L. Prog Growth Factor Res. 1995;6:197–206. doi: 10.1016/0955-2235(95)00022-4. [DOI] [PubMed] [Google Scholar]
- 24.Badinga L, Song S, Simmen R C, Clarke J B, Clemmons D R, Simmen F A. J Mol Endocrinol. 1999;23:277–285. doi: 10.1677/jme.0.0230277. [DOI] [PubMed] [Google Scholar]
- 25.Smith W C, Kuniyoshi J, Talamantes F. Mol Endocrinol. 1989;3:984–990. doi: 10.1210/mend-3-6-984. [DOI] [PubMed] [Google Scholar]
- 26.Nagano M, Chastre E, Choquet A, Bara J, Gespach C, Kelly P A. Am J Physiol. 1995;268:G431–G442. doi: 10.1152/ajpgi.1995.268.3.G431. [DOI] [PubMed] [Google Scholar]
- 27.Wysolmerski J J, Stewart A F. Annu Rev Physiol. 1998;60:431–460. doi: 10.1146/annurev.physiol.60.1.431. [DOI] [PubMed] [Google Scholar]
- 28.Dougherty K M, Blomme E A, Koh A J, Henderson J E, Pienta K J, Rosol T J, McCauley L K. Cancer Res. 1999;59:6015–6022. [PubMed] [Google Scholar]
- 29.Franc N C, Heitzler P, Ezekowitz R A, White K. Science. 1999;284:1991–1994. doi: 10.1126/science.284.5422.1991. [DOI] [PubMed] [Google Scholar]
- 30.Meszaros A J, Reichner J S, Albina J E. J Immunol. 2000;165:435–441. doi: 10.4049/jimmunol.165.1.435. [DOI] [PubMed] [Google Scholar]
- 31.Salama N, Guillemin K, McDaniel T K, Sherlock G, Tompkins L, Falkow S. Proc Natl Acad Sci USA. 2000;97:14668–14673. doi: 10.1073/pnas.97.26.14668. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Modlin I M, Poulsom R. J Clin Gastroenterol. 1997;25:S94–S100. doi: 10.1097/00004836-199700001-00016. [DOI] [PubMed] [Google Scholar]
- 33.Podolsky D K. Am J Physiol. 1999;277:G495–G499. doi: 10.1152/ajpgi.1999.277.3.G495. [DOI] [PubMed] [Google Scholar]
- 34.Umekage T, Kato K. FEBS Lett. 1991;286:147–151. doi: 10.1016/0014-5793(91)80961-2. [DOI] [PubMed] [Google Scholar]
- 35.Ramsden J J. Int J Biochem Cell Biol. 2000;32:475–479. doi: 10.1016/s1357-2725(99)00152-1. [DOI] [PubMed] [Google Scholar]
- 36.Keck U, Nedellec P, Beauchemin N, Thompson J, Zimmermann W. Eur J Biochem. 1995;229:455–464. doi: 10.1111/j.1432-1033.1995.0455k.x. [DOI] [PubMed] [Google Scholar]
- 37.Narushima Y, Unno M, Nakagawara K, Mori M, Miyashita H, Suzuki Y, Noguchi N, Takasawa S, Kumagai T, Yonekura H, Okamoto H. Gene. 1997;185:159–168. doi: 10.1016/s0378-1119(96)00589-6. [DOI] [PubMed] [Google Scholar]
- 38.Wang X, Wang B, Wu J. J Gastroenterol Hepatol. 2001;16:79–86. doi: 10.1046/j.1440-1746.2001.02389.x. [DOI] [PubMed] [Google Scholar]
- 39.Webb G C, Akbar M S, Zhao C, Steiner D F. Proc Natl Acad Sci USA. 2000;97:5773–5778. doi: 10.1073/pnas.100126597. . (First Published May 16, 2000; 10.1073/pnas.100126597) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Nadler S T, Stoehr J P, Schueler K L, Tanimoto G, Yandell B S, Attie A D. Proc Natl Acad Sci USA. 2000;97:11371–11376. doi: 10.1073/pnas.97.21.11371. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.