

Abstract
The goal of the present paper is to establish and validate the link between cancer diagnosis and therapy by microRNAs detection. The induction in vitro of some specific microRNAs after treatment with MDR ligands has been outlined. Starting from the results obtained by in vitro induction of MDCK and MDCK-MDR1 cells treated by a MDR1 ligand, a new scenario in the early diagnosis and chemotherapy could be disclosed. To corroborate this perspective a short overview on pancreatic cancer diagnosis and chemotherapeutic treatment has been reported.
Keywords: microRNAs, MDR, early diagnosis, MDCK-MDR1, P-glycoprotein
1. Introduction
MultiDrug Resistance (MDR) is the major obstacle to a successful cancer therapy and is often associated with an increased efflux of anticancer drugs due to some proteins belonging to the ATP-Binding Cassette (ABC) transporters family such as P-gp, BCRP and MRP1 [1,2,3,4,5]. P-gp, exerting a physiological activity as first line of defense of our body and together with protein ABCG2/BCRP, is localized at the apical level in cells membranes of different cellular compartments such as liver, intestine, kidney, placenta [1,2,3,4,5]. This strategic localization gives P-gp a crucial role as responsible for drugs absorption and accumulation. MRP1 (ABCC1) also belonging to ABC transporters, localized at the basolateral level in cells membranes, facilitates the transport of the substrates in the blood [1,2,3,4,5].
The involvement of P-gp, BCRP and MRP1 in MDR is due to their overexpression in resistant tumor cells [2]. In particular MDR1/P-gp behaves as “protein scavenger” able to “capture” drugs and transport them out of cells.
MicroRNAs (miRNAs) are small endogenous non-coding RNAs responsible for the post-transcriptional regulation of target genes by the interaction with specific sequences in their 3' untranslated region (3'UTR), leading to mRNA degradation and/or translational inhibition [6]. MiRNAs play important roles in many cellular processes such as proliferation, apoptosis and differentiation, and in many physiological processes such as metabolism, cardiogenesis, development and function of the nervous and immune systems [6,7,8]. The miRNAs dysregulation in cancers are widely reported and recent literatures revealed a correlation of miRNAs levels in biological fluids with chemotherapy response [8]. To et al. reported a double regulation of MDR, direct and indirect, mediated by miRNAs [8]. Moreover, a recent study demonstrated that a miRNAs dysregulation and an altered expression of Dicer and Argonaute, enzymes involved in miRNA maturation process, have been found in a doxorubicin-resistant breast cancer cell line, MCF-7/DOX [9]. Several miRNAs have been found to regulate drug resistance genes such as ABCG2 [10,11,12,13] and MDR1 [9,14] and the modulation of miRNAs expression or function has been reported to alter the sensitivity of cancer cells to anticancer drugs [15,16].
MiR-27a and miR-451 are involved in the activation of MDR1 expression [14], and in particular miR-27a has been found able to modulate MDR1 expression by the inhibition of FZD7/β-catenin pathway in hepatocellular carcinoma cells [17]. MiR-298 has been found able to increase MDR1 expression and so responsible for doxorubicin-resistance of metastatic breast cancer [18].
The hypothesis that miRNAs could be involved in the modulation of P-gp expression is supported by suggestive evidence: (i) miR-27a and miR-451 induce an increased P-gp expression in ovarian cancer cells [9,14], (ii) miR-451 also down-regulates MDR1 expression in MCF7/DOX-resistant cells [9]; (iii) changes in miR-381 and miR-495 expression are inversely correlated to MDR1 expression and the development of MDR [19].
In the first part of this paper, we monitored the induction of some specific miRNAs by the treatment of wild type cell line (MDCK cells) and in the corresponding cell line where P-gp is overexpressed (MDCK-MDR1 cells) unsensitive to antineoplastic treatment. In particular we stimulated MDCK cells with a specific P-gp substrate 8c and a selective P-gp inhibitor 8a, as reported in Table 1 [20], verifying the down- or up-regulation of miRNAs with respect to untreated cells.
Table 1.
Biological evaluation of tariquidar derivatives 8a and 8c.
| Compound | R | MDR1 a EC50, µM | Pappa | MDR1 interacting mechanism a |
| 8a | 3-OH | 0.25 ± 0.02 | 0.9 | inhibitor |
| 8c | 4-NO2 | 0.51 ± 0.032 | 2.7 | substrate |
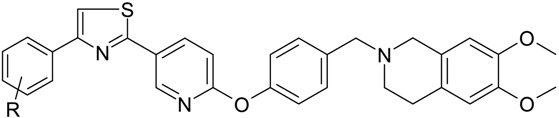
a All data are reported in reference [20].
Preliminary eight microRNAs have been identified as highly expressed in MDCK-wild type and are also reported in many human tumors as glioblastoma, bladder cancer, breast cancer, renal cancer as reported in the literature [12,13,14]. Aims of this study are: (i) to probe a link between some miRNAs and MDR P-gp-mediated; (ii) to improve the knowledge of the pathways involved in cancers characterized by MDR; (iii) to verify if different miRNAs are modulated by P-gp inhibitor and substrate; (iv) to suggest some miRNAs as probes to diagnose MDR and to address chemotherapy.
2. Results and Discussion
Eight microRNAs have been identified as highly expressed in MDCK-wild type and MDCK-MDR1 as listed in Table 2.
Table 2.
Frequence of miRNAs higly expressed in MDCK-wt and MDCK-MDR1.
| miR-Dog | miR-Homo | Freq. MDCK-wt | RPM Freq. MDCK-wt | Freq. MDCK-MDR1 | RPM Freq. MDCK-MDR1 |
|---|---|---|---|---|---|
| cfa-miR-182 | hsa-miR-182-5p | 835,232 | 32,507,727 | 785,269 | 44,147.288 |
| cfa-miR-21 | hsa-miR-21-5p | 767,174 | 29,858.950 | 582,432 | 32,744.188 |
| cfa-miR-10a | hsa-miR-10a-5p | 1,610,023 | 62,662.126 | 944,420 | 53,094.446 |
| cfa-miR-27b | hsa-miR-27a-3p | 612,930 | 23,855.867 | 319,758 | 17,977.170 |
| cfa-miR-30a | hsa-miR-30a-5p | 515,654 | 20,069.943 | 312,771 | 17,584.374 |
| cfa-miR-26a | hsa-miR-26a-5p | 545,522 | 21,232.388 | 359,038 | 20,185.414 |
| cfa-miR-10b | hsa-miR-10a-5p | 2,127,806 | 82,813.929 | 1,602,329 | 90,080.803 |
| cfa-miR-191 | hsa-miR-191-5p | 636,561 | 24,775.571 | 376,207 | 21,150.622 |
Freq: Frequence for each miRNA higly expressed; RPM Freq: normalization of frequence for each miRNA higly expressed.
These miRNAs represent the background and so they cannot be considered for our studies. By contrast, the expression of five miRNAs, modulated only by compound 8c displaying MDR substrate activity, has been detected in MDCK-wt and MCDK-MDR1 treated with 8c. As expected, P-gp inhibitor 8a did not exert any effect on miRNAs expression because it has been tested as a negative control. Cfa-miR-181d (hsa-miR-181a-5p), cfa-miR-218 (hsa-miR-218-5p) were found up-regulated in 8c-treated MDCK-MDR1 cells with respect to untreated MDCK-MDR1 cells, while cfa-miR-454 (hsa-miR-130a-3p), cfa-miR-424 (hsa-miR-424-3p), cfa- miR-1307 were found down-regulated, as depicted in Table 3.
Table 3.
Expression of five microRNAs known and selected by miRDeep2 in MDR, regulated by compound 8c.
| miR-Dog | miR-Homo | RPM-MDCK-MDR1 | RPM–Substrate-8c-MDCK-MDR1 | Substrate-8c-MDCK-MDR1/MDCK-wt |
|---|---|---|---|---|
| cfa-miR-181d | hsa-miR-181a-5p | 0.000 | 122.319 | 6.93450378 |
| cfa-miR-218 | hsa-miR-218-5p | 8.814 | 40.113 | 2.186153174 |
| cfa-miR-454 | hsa-miR-130a-3p | 9.264 | 0.000 | −3.211643438 |
| cfa-miR-424 | hsa-miR-424-3p | 11.963 | 0.000 | −3.580449275 |
| cfa-miR-1307 | - | 31.864 | 0.000 | −4.993842191 |
2.1. MiRNAs and MDR Pathway
2.1.1. cfa-miR-424 (hsa-miR-424-3p)
MiRNA-424, identified as down-regulated in 8c-treated MDR1-MDCK cells, is involved in different pathways leading to MDR up-regulation. In particular, it has been reported that miR-424 mediates the induction of the hypoxia inducing factors HIF-1α and HIF-2α [21]. HIF-1α is responsible for MDR1 up-regulation. Moreover, miR-424 modulates also the expression of protein cullin 2 (CUL2), a scaffolding protein displaying a pivotal role in the assembly of the ubiquitin ligase system, thereby stabilizing HIF-1α [21]. Considering these pathways, compound 8c inducing a miR-424 down-regulation could lead to a consequent reduction of HIF1α and consequently of MDR P-gp-mediated.
2.1.2. cfa-miR-454 (hsa-miR-130a-3p)
miR-130a-3p (corresponding to miR-454 in the dog) expression in humans is reduced in response to the action of the substrate 8c, as reported in Table 3. A recent study identified the miRNAs differentially expressed in a tumor ovarian carcinoma cell line (SKOV3) and in the parental cell line insensitive to cisplatin (SKOV3/CIS). This different miRNAs profile suggests that these miRNAs and their target proteins could play a crucial role in the development of cisplatin resistance in ovarian cancer [22]. The expression levels of miR-130a were higher in SKOV3/CIS cells than in SKOV3 cells; also P-gp expression level was very high in SKOV3/CIS and absent in SKOV3 cells. Overall, these results demonstrated that miR-130a up-regulation in resistant ovarian cells was correlated to MDR1-mediated cisplatin resistance. The MDR induction is probably due to a direct modulation of PI3K /Akt, PTEN and mTOR, by miR-130a-3p up-regulation [22].
Another study by Xu et al. demonstrated that the up-regulation of miR-130a, induced by cisplatin, leads to cisplatin resistance by the inhibition of RUNX3 and the activation of Wnt signaling [23]. Therefore, also in this case the down-regulation of miR-130a could lead to a decrese of MDR.
2.1.3. cfa-miR-218 (hsa-miR-218-5p)
MiR-218-5p was detected as up-regulated in 8c-treated cells; its down-regulation is reported in etoposide resistant MCF7VP breast cancer cell line compared to MCF7 wild type [24]. This effect has been considered an indicator of etoposide-resistance in this cell line since it is also accompanied to an increased expression of ABCC1 and ABCC6. Therefore, its down-regulation by MDR1 substrates could lead to a decreased expression of MDR gene.
2.1.4. miR-181a-5p
MiR-181a has been considered as tumor suppressor in human acute monocytic leukemia (AML) and its overexpression has been demonstrated to induce apoptosis of AML blasts [25,26,27]. This effect is probably linked to the modulation of the expression of the antiapoptotic protein BCL2. Apoptosis induced by the antineoplastic agent, daunorubicin (DNR), can be blocked by BCL-2 overexpression and so the suppression of BCL-2 expression enhance DNR-induced apoptosis [28]. BCL-2 has been identified in several studies as one of the potential targets of miR-181a and so the down-regulation of miR-181a induced in K562 cells resistance to DNR and the up-regulation of miR-181 sensitized the K562/A02 cells to DNR. Therefore, miR-181a might sensitize K562/A02 cells to DNR by repressing the BCL-2 protein expression [27].
2.1.5. miR-1307
cfa-miR-1307 identified by the program miRDeep2 in MDR1-MDCK cells treated with the substrate 8c, do not correspond to any human miRNA, and it is probably unknown. However, this miRNA is down regulated in the MDR1-MDCK cells treated with the substrate 8c compared to the parental MDCK wild type cells. Moreover, TargetScanHuman (a software for the prediction of microRNA targets) identified as probable cfa-miR-1307 target, the BCL2 gene, responsible for the corresponding anti-apoptotic protein.
2.2. Perspective of microRNAs in MDR Diagnosis
Several studies have demonstrated that some miRNAs are associated with tumor promotion (oncogenes) or could inhibit tumors by reducing cell proliferation, cell survival (tumor suppressors) and cellular differentiation; thus, restoring the expression of some miRNAs in tumor cells could lead to differentiation and induce malignant cells into a normal state.
Recently, the deregulation of miRNA expression in pancreatic cancer tumors has been reported [29,30,31,32,33,34,35] and the monitoring of this deregulation has been performed at different stages of cancer. It has already reported that miRNAs expression varied in different solid tumors (breast, colon, lung, pancreas, prostate, and stomach) giving information on the response of cancer cells to different treatment (Table 4). For example, a study, performed on the human pancreatic cancer cell line BxPC-3, demonstrated that the inhibition of proliferation induced by trichostatin A (TSA) was time and dose-dependent and this effect was accompanied by a different miRNAs expression profile [36]; TSA treatment induced the expression of tumor suppressor miR-200c and miR-21 leading to growth arrest. Similarly, curcumin induced up-regulation of miR-22 and down-regulation of miR-199a in human pancreatic cells [30], 5-aza-2'-deoxycytidine exposure induced the up-regulation of several miRNAs (miR-107, miR-103, miR-29a, miR29b and miR-320) in MiaPACA-2 and Panc-1 cell lines [37]. Moreover, the natural treatment with 3,3-diindolylmethane (DIM) or isoflavone induced the down-regulation of miR-200b, miR-200c, let-7b, let-7c, let-7d, and let-7e in gemcitabine-resistant cells [38].
Table 4.
Panel of microRNAs in normal and pancreatic cancer.
| microRNA | Normal tissue | Pancreatic cancer | PDAC |
|---|---|---|---|
| miR-103 | Basal level | Up-regulated | Basal level |
| miR-107 | Basal level | Up-regulated | Basal level |
| miR-155 | Basal level | Down-regulated | Basal level |
| miR-216 | Basal level | Basal level | High level |
| miR-217 | Basal level | Basal level | High level |
| miR-133a | Basal level | Basal level | Absent |
| miR-99 | Not detected | Detected | Detected |
| miR-100 | Not detected | Detected | Detected |
| Mir-100-1,2 | Not detected | Detected | Detected |
| miR-125a | Not detected | Detected | Detected |
| miR-125b-1 | Not detected | Detected | Detected |
| miR199a-1 | Not detected | Detected | Detected |
| miR199a-2 | Not detected | Detected | Detected |
| miR-21 | Basal level | Detected | Detected |
| miR-221 | Not detected | Detected | Detected |
| miR-222 | Not detected | Detected | Detected |
| miR-181a | Not detected | Detected | Detected |
| miR-181b | Not detected | Detected | Detected |
| miR-181d | Not detected | Detected | Detected |
| miR-196a | Basal level | Detected | Detected |
The miRNA expression pattern may discriminate normal pancreas from tumors; indeed, the up-regulation of miR-103 and miR-107, and the down-regulation of miR-155 can distinguish tumors from the normal pancreas [37]. Moreover, the presence of miR-216 and -217 and the lack of miR-133a expression was a characteristic feature of the pancreatic tissue.
This different expression profile can be useful for miRNAs-based diagnosis [34]. The up-regulation of miR-99, miR-100, miR-100-1/2, miR-125a, miR-125b-1, miR-199a-1, and miR-199a-2 was detected in cancers and in chronic pancreatitis but not in normal pancreatic tissue; the up-regulation of miR-21, miR-221, miR-222, miR-181a, miR-181b, miR-181d, and miR-155 was found in tumors compared to benign pancreatic tissue [32,33]. Moreover, the up-regulation of miR-21 was predictive of a poorer outcome and may be an important biomarker for prognosis of pancreatic cancer [39]. MiR-196a and miR-217 can discriminate between healthy tissue, PDAC, and chronic pancreatitis [40]. The expression of miR-21, miR-210, miR-155, and miR-196a in the plasma of patients revealed the cancer and among them overexpression of miR-155 and miR196a is due to the progression of the pathology. Therefore, the identification of specific miRNAs in plasma can provide sensitive and specific blood-based biomarkers for pancreatic cancer diagnosis [41]. Moreover, the high levels of miR-155, miR-203, miR-210 and miR-222 in pancreatic tumors were also associated with increased risk of death compared to patients with tumors with low expression of the same miRNAs [42].
2.3. Usefulness of miRNAs Recognition in Multidrug Resistance Chemotherapy: On Overview on Pancreatic Cancer
To date, there are different therapeutic approaches for pancreatic cancer, including single or multi-agents chemotherapy, chemotherapy followed by chemoradiation, or immediate concurrent chemoradiation. In patients affected by pancreatic metastatic cancer, the overall survival is of 6 months from the time of diagnosis. Gemcitabine has been the standard treatment (from 1997). Recently, the combination of the antineoplastic agents 5-fluorouracil, oxaliplatin, irinotecan and leucovorin (FOLFIRINOX) have been used as alternative strategy [43] leading to an improved response of advanced metastatic disease compared with gemcitabine alone and other historical treatments [44,45,46,47,48].
Folfirinox showed a median overall survival (11 months) higher than that of gemcitabine (6.8 months) and a median progression-free survival almost doubled (6.4 months) with respect to gemcitabine (3.3 months) [49]. However, folfirinox treatment in some clinical trials has showed high toxicity with respect to gemcitabine regimen (neutropenia, diarrhea, fatigue) [49].
A relationship between the expression of some miRNAs and survival of patients with pancreatic adenocarcinoma has been already reported and miR-21, miR-155, miR-203, miR-210 and miR-222 have been identified as potential predictors of survival [42,50].
The expression of miR-155, miR-203, miR-210, miR-216, miR-217 and miR-222, differentially expressed in pancreatic tumors compared to normal tissues, has showed an high correlation with the overall survival. In particular, patients with elevated expression of all four miRNAs had a 6.2-fold increased risk of tumor-related death with respect to patients with a lower expression of these miRNAs [51].
High miR-21 expression has been detected in patients with a significantly shorter overall survival (OS) [50] and a correlation between miR-21 expression and gemcitabine resistance has been proved. Also miR-200 is involved in chemosensitivity of pancreatic cancer cells and its decreased expression together with an increased expression of miR-21 are linked to gemcitabine resistance in pancreatic cancer cell lines [51].
Therefore, since miR-21 is expressed in all pancreatic adenocarcinoma forms and it is significantly up-regulated after gemcitabine treatment, miR-21 could be a prognostic factor leading pancreatic cancer therapy [50].
3. Experimental
3.1. Cells Lines
Materials: Cell culture reagents were purchased from Celbio s.r.l. (Milano, Italy).
Cell cultures MDCK-MDR1 and MDCK cells are a gift of P. Borst, NKI-AVL Institute, Amsterdam, Nederland. All MDCK cells were grown in DMEM high glucose supplemented with 10% fetal bovine serum, 2 mM glutamine, 100 U/mL penicillin, 100 µg/mL streptomycin, in a humidified incubator at 37 °C with a 5% CO2 atmosphere. MDCK cells were cultured to 80% confluence, the medium removed and the cells rinsed in PBS. The cells were scraped and suspended in PBS, then centrifuged at 1500 rpm for 15 min. The supernatant was discarded and the pellet used for the analysis.
Treatment with 8a and 8c. MDCK-MDR1 and MDCK cells were treated with 0.5 µM 8a and 8c (solved in medium buffer) at 48h of exposure. The concentration has been selected taking into account the activity values (EC50 = 0.25 and 0.51 µM, respectively) of the two compounds towards P-gp [20]. The exposure time has been determined considering the induction of involved pathways into the cells
3.2. Data Processing
In a first step of this work, we used indexed genome of dog to map reads. In this operation miRdeep2 used an algorithm which divided sequences that map to those that do not map to the genome of dog, so we discarded sequences that don’t map on genome of dog. In the second step we used a script of miRDeep2 able to extract all the sequences including the respective name given in miRBase [52] in relation to the human species and the species of the dog, so as to compare them in the next step. For this step we have needed of a script in perl that extracts microRNA with their name for each species from miRBase. So we have obtained a file for human, a file for dog and a file for the other species. So in the next phase miRDeep2 used these files of reference and compared, previously selected reads, with sequences having a distinct name miR extracted from miRBase. So it is possible to assign a name to each sequence as compared with miR of man and dog shows in miRBase. The final phase includes the acquisition of two files for each experiment, the first file contains all the known sequences of miR of dog with same “seed” in human and the second file contains unknown miR for dog and for other species. But we focused on known miR that have a name in miRBase both for dog that for human because characterized by same seed. We have obtained these outputs: (1) miRNAs known for MDCK-wt and MDCK-MDR1 (2) miRNAs known for each cell line treated respectively with the compound 8a (MDR1-inhibitor) and 8c (MDR1-substrate). We filtered known miRNAs according to high score to eliminate repeated sequences and a shorter sequence of the same microRNA. The next step was to compare all the miRNAs obtained in each experiment with the source file in which we have calculated the frequency with which each sequence of microRNA is expressed in each experiment. At this point we have created a script in perl to calculate a “fold change” between:
(i) MDCK- MDR1 vs. MDCK-wt; (ii) MDCK-MDR1 treated with compound 8a and 8c vs. untreated MDCK- MDR1. “Fold change” is the ratio between the expression of miRNAs in each cell line and its control, where the control is represented by MDCK-wt or MDCK-overexpressing MDR1 (MDCK-MDR1) (See Tables S2 and S3 in supplementary materials).
4. Conclusions
In this article the role of miRNAs in MDR has been pointed out. For this purpose, an brief experimental section has been carried out where the link between some miRNAs and the induced MDR has been demonstrated. This finding confirmed that miRNAs could be routinely employed both for the diagnosis and to drive the pharmacological treatment since the experimental section demonstrated that the over-and/or down-expression of miRNAs can be drug-sensitive. This preliminary study aims to hit both the diagnostic and therapeutic items, and it could drive the chemotherapeutic strategy taking into account the level of drug-sensitive miRNAs. To corroborate this perspective a short overview on pancreatic cancer diagnosis and chemotherapeutic treatment has been reported.
Supplementary Materials
Supplementary materials can be accessed at: http://www.mdpi.com/1420-3049/19/5/5611/s1.
Author Contributions
Andreas Gisel, Mirna Valvano deal with the experimental section (microRNA detection) and bioinformatic analysis; Patrizia Nardulli, Amalia Azzariti, Imane Ghafir El Idrissi and Antonio Carrieri deal with the perspective in pancreatic cancer; Marialessandra Contino and Nicola Antonio Colabufo performed the experimental section (tumor cells treatment, synthesis of the tested compounds and results analysis) and edited the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
Footnotes
Sample Availability: Compounds 8a and 8c are available from the authors.
References
- 1.Pharm A.N., Penchala S., Graf R.A., Wang J., Huang Y. Pharmacogenomic Characterization of ABC Transporters. Involved in Multidrug Resistance. In: Colabufo N.A., editor. Multidrug Resistance: Biological and Pharmaceutical Advance in the Antitumour Treatment. Research Signpost; Kerala, India: 2008. pp. 19–62. [Google Scholar]
- 2.Gimenez-Bonafe P., Guillen Canovas A., Ambrosio S., Tortosa A., Perez Tomas R. Drugs Modulating MDR. In: Colabufo N.A., editor. Multidrug Resistance: Biological and Pharmaceutical Advance in the Antitumour Treatment. Research Signpost; Kerala, India: 2008. pp. 63–99. [Google Scholar]
- 3.Perez-Tomas R. Multidrug resistance: Retrospect and prospects in anti-cancer drug treatment. Curr. Med. Chem. 2006;13:1859–1876. doi: 10.2174/092986706777585077. [DOI] [PubMed] [Google Scholar]
- 4.Teodori E., Dei S., Martelli S., Scapecchi F., Gualtieri F. The functions and structure of ABC transporters: Implications for the design of new inhibitors of Pgp and MRP1 to control multidrug resistance (MDR) Curr. Drug Targets. 2006;7:893–909. doi: 10.2174/138945006777709520. [DOI] [PubMed] [Google Scholar]
- 5.Avendãno C., Menéndez J.C. Inhibitors of multidrug resistance to antitumor agents (MDR) Curr. Med. Chem. 2002;9:159–119. doi: 10.2174/0929867023371175. [DOI] [PubMed] [Google Scholar]
- 6.Bartel D.P. MicroRNAs: Target recognition and regulatory functions. Cell. 2009;136:215–233. doi: 10.1016/j.cell.2009.01.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Kuzhandai V., Ramesh R., Srinivasan A.R. Ciruclating MicroRNAs as biomarkers in health and disease. J. Clin. Diagn. Res. 2012;6:1791–1795. doi: 10.7860/JCDR/2012/4901.2653. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.To K.K. MicroRNA: A prognostic biomarker and a possible druggable target for circumventing multidrug resistance in cancer chemotherapy. J. Biomed. Sci. 2013;20:1–19. doi: 10.1186/1423-0127-20-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Kovalchuk O., Fikowski J., Meservy J., Ilnytskyy Y., Tryndyak V.P., Chekhun V.F., Pogribny I.P. Involvement of microRNA-451 in resistance of the MCF-7 breast cancer cells to chemotherapeutic drug doxorubicin. Mol. Cancer Ther. 2008;7:2152–2159. doi: 10.1158/1535-7163.MCT-08-0021. [DOI] [PubMed] [Google Scholar]
- 10.To K.K., Zhan Z., Litman T., Bates S.E. Regulation of ABCG2 expression at the 3'untranslated region of its mRNA through modulation of transcript stability and protein translation by a putative microRNA in the S1 colon cancer cell line. Mol. Cell Biol. 2008;28:5147–5161. doi: 10.1128/MCB.00331-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.To K.K., Robey R.W., Knutsen T., Zhan Z., Ried T., Bates S.E. Escape from hsa-miR-519c enables drug-resistant cells to maintain high expression of ABCG2. Mol. Cancer Ther. 2009;8:2959–2968. doi: 10.1158/1535-7163.MCT-09-0292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Pan Y.Z., Morris M.E., Yu A.M. MicroRNA-328 negatively regulates the expression of breast cancer resistance protein (BCRP/ABCG2) in human cancer cells. Mol. Pharmacol. 2009;75:1374–1379. doi: 10.1124/mol.108.054163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Yamamoto Y., Yoshioka Y., Minoura K., Takahashi R.U., Takeshita F., Taya T., Horii R., Fukuoka Y., Kato T., Kosaka N., et al. An integrative genomic analysis revealed therelevance of microRNA and gene expression for drug-resistance in human breast cancer cells. Mol. Cancer. 2011;10:135. doi: 10.1186/1476-4598-10-135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Zhu H., Wu H., Liu X., Evans B.R., Medina D.J., Liu C.G., Yang J.M. Role of MicroRNA mir27a and mir 451 in the regulation of MDR1/P glycoprotein. Biochem. Pharmacol. 2008;76:582–588. doi: 10.1016/j.bcp.2008.06.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Allen K.E., Weiss G.J. Resistance may not be futile: MicroRNA biomarkers for chemoresisatnce and potential therapeutics. Mol. Cancer Ther. 2010;9:3126–3136. doi: 10.1158/1535-7163.MCT-10-0397. [DOI] [PubMed] [Google Scholar]
- 16.Rodrigues A.C., Li X., Radecki L., Pan Y.Z., Winter J.C., Huang M., Yu A.M. MicroRNA expression is differentially altered by xenobiotic drugs in different human cell lines. Biopharm. Drug Dispos. 2011;32:355–367. doi: 10.1002/bdd.764. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Chen Z., Ma T., Huang C., Zhang L., Lv X., Xu T., Hu T., Li J. MiR-27a modulates the MDR1/P-glycoprotein expression by inhibiting FZD7/β-catenin pathway in hepatocellular carcinoma cells. Cell Signal. 2013;25:2693–2701. doi: 10.1016/j.cellsig.2013.08.032. [DOI] [PubMed] [Google Scholar]
- 18.Bao L., Hazari S., Mehra S., Kaushal D., Moroz K., Dash S. Increased expression of P-glycoprotein and doxorubicin chemoresistance of metastatic breast cancer is regulated by miR-298. Am. J. Pathol. 2012;180:2490–2503. doi: 10.1016/j.ajpath.2012.02.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Xu Y., Ohms S.J., Li Z., Wang Q., Gong G., Hu Y., Mao Z., Shannon M.F., Fan J.Y. Changes in the expression of miR-381 and miR-495 are inversely associated with the expression of the MDR1 gene and development of multi-drug resistance. PLoS One. 2013;8:e82062. doi: 10.1371/journal.pone.0082062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Contino M., Zinzi L., Perrone M.G., Leopoldo M., Berardi F., Perrone R., Colabufo N.A. Potent and selective tariquidar bioisosters as potential PET. Bioorg. Med. Chem. Lett. 2013;23:1370–1374. doi: 10.1016/j.bmcl.2012.12.084. [DOI] [PubMed] [Google Scholar]
- 21.Ghosh G., Subramanian I.V., Adhikari N., Zhang X., Joshi H.P., Basi D., Chandrashekhar Y.S., Hall J.L., Roy S., Zeng Y., et al. Hypoxia-induced microRNA-424 expression in human endothelial cells regulates HIF-αisoforms and promotes angiogenesis. J. Clin. Invest. 2010;120:4141–4154. doi: 10.1172/JCI42980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Yang L., Li N., Wang H., Jia X., Wang X., Luo J. Altered microRNA expression in cisplatin-resistant ovarian cancer cells and upregulation of miR-130a associated with MDR1/P-glycoprotein-mediated drug resistance. Oncol. Rep. 2012;28:592–600. doi: 10.3892/or.2012.1823. [DOI] [PubMed] [Google Scholar]
- 23.Xu N., Shen C., Luo Y., Xia L., Xue F., Xia Q., Zhang J. Upregulated miR-130a increases drug resistance by regulating RUNX3 and Wnt signaling in cisplatin-treated HCC cell. Biochem. Biophys Res. Commun. 2012;425:468–472. doi: 10.1016/j.bbrc.2012.07.127. [DOI] [PubMed] [Google Scholar]
- 24.Moitra K., Im K., Limpert K., Borsa A., Sawitzke J., Robey R., Yuhki N., Savan R., Huang da W., Lempicki R.A., et al. Differential gene and microRNA expression between etoposide resistant and etoposide sensitive MCF7 breast cancer cell lines. PLoS One. 2012;7:e45268. doi: 10.1371/journal.pone.0045268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Shi L., Cheng Z., Zhang J., Li R., Zhao P., Fu Z., You Y. hsa-mir-181a and hsa-mir-181b function as tumor suppressors in human glioma cells. Brain Res. 2008;1236:185–193. doi: 10.1016/j.brainres.2008.07.085. [DOI] [PubMed] [Google Scholar]
- 26.Fanini F., VI, Faabbri M. MicroRNAs: Tiny players with a big role in the pathogenesis of leukemias and lymphomas. Hematol. Rev. 2009;1:40–45. [Google Scholar]
- 27.Li H., Hui L., Xu W. miR-181a sensitizes a multidrug-resistant leukemia cell line K562/A02 to daunorubicin by targeting BCL-2. Acta Biochim. Biophys. Sin. (Shanghai) 2012;44:269–277. doi: 10.1093/abbs/gmr128. [DOI] [PubMed] [Google Scholar]
- 28.Kim Y.H., Park J.W., Lee J.Y., Surh Y.J., Kwon T.K. Bcl-2 overexpression prevents daunorubicin-induced apoptosis through inhibition of XIAP and Akt degradation. Biochem. Pharmacol. 2003;66:1779–1786. doi: 10.1016/S0006-2952(03)00545-8. [DOI] [PubMed] [Google Scholar]
- 29.Roldo C., Missiaglia E., Hagan J.P., Falconi M., Capelli P., Bersani S., Calin G.A., Volinia S., Liu C.G., Scarpa A., et al. MicroRNA expression abnormalities in pancreatic endocrine and acinar tumors are associated with distinctive pathologic features and clinical behavior. J. Clin. Oncol. 2006;24:4677–4684. doi: 10.1200/JCO.2005.05.5194. [DOI] [PubMed] [Google Scholar]
- 30.Sun M., Estrov Z., Ji Y., Coombes K.R., Harris D.H., Kurzrock R. Curcumin (diferuloylmethane) alters the expression profiles of microRNAs in human pancreatic cancer cells. Mol. Cancer Ther. 2008;7:464–473. doi: 10.1158/1535-7163.MCT-07-2272. [DOI] [PubMed] [Google Scholar]
- 31.Bloomston M., Frankel W.L., Petrocca F., Volinia S., Alder H., Hagan J.P., Liu C.G., Bhatt D., Taccioli C., Croce C.M. MicroRNA expression patterns to differentiate pancreatic adenocarcinoma from normal pancreas and chronic pancreatitis. JAMA. 2007;297:1901–1908. doi: 10.1001/jama.297.17.1901. [DOI] [PubMed] [Google Scholar]
- 32.Gironella M., Seux M., Xie M.J., Cano C., Tomasini R., Gommeaux J., Garcia S., Nowak J., Yeung M.L., Jeang K.T., et al. Tumor protein 53-induced nuclear protein 1 expression is repressed by miR-155, and its restoration inhibits pancreatic tumor development. Proc. Natl. Acad. Sci. USA. 2007;104:16170–16175. doi: 10.1073/pnas.0703942104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Lee E.J., Gusev Y., Jiang J., Nuovo G.J., Lerner M.R., Frankel W.L., Morgan D.L., Postier R.G., Brackett D.J., Schmittgen T.D. Expression profiling identifies microRNA signature in pancreatic cancer. Int. J. Cancer. 2007;120:1046–1054. doi: 10.1002/ijc.22394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Szafranska A.E., Davison T.S., John J., Cannon T., Sipos B., Maghnouj A., Labourier E., Hahn S.A. MicroRNA expression alterations are linked to tumorigenesis and non-neoplastic processes in pancreatic ductal adenocarcinoma. Oncogene. 2007;26:4442–4452. doi: 10.1038/sj.onc.1210228. [DOI] [PubMed] [Google Scholar]
- 35.Tsuda N., Ishiyama S., Li Y., Ioannides C.G., Abbruzzese J.L., Chang D.Z. Synthetic microRNA designed to target glioma-associated antigen 1 transcription factor inhibits division and induces late apoptosis in pancreatic tumor cells. Clin. Cancer Res. 2006;12:6557–6564. doi: 10.1158/1078-0432.CCR-06-0588. [DOI] [PubMed] [Google Scholar]
- 36.Zhang S., Cai X., Huang F., Zhong W., Yu Z. Effect of trichostatin a on viability and microRNA expression in human pancreatic cancer cell line BxPC-3. Exp. Oncol. 2008;30:265–268. [PubMed] [Google Scholar]
- 37.Lee K.H., Lotterman C., Karikari C., Omura N., Feldmann G., Habbe N., Goggins M.G., Mendell J.T., Maitra A. Epigenetic silencing of MicroRNA miR-107 regulates cyclin-dependent kinase 6 expression in pancreatic cancer. Pancreatology. 2009;9:293–301. doi: 10.1159/000186051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Li Y., VandenBoom T.G., Kong D., Wang Z., Ali S., Philip P.A., Sarkar F.H. Up-regulation of miR-200 and let-7 by natural agents leads to the reversal of epithelial-to-mesenchymal transition in gemcitabine-resistant pancreatic cancer cells. Cancer Res. 2009;69:6704–6712. doi: 10.1158/0008-5472.CAN-09-1298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Dillhoff M., Liu J., Frankel W., Croce C., Bloomston M. MicroRNA-21 is overexpressed in pancreatic cancer and a potential predictor of survival. J. Gastrointest. Surg. 2008;12:2171–2176. doi: 10.1007/s11605-008-0584-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Szafranska A.E., Doleshal M., Edmunds H.S., Gordon S., Luttges J., Munding J.B., Barth R.J., Jr., Gutmann E.J., Suriawinata A.A., Marc Pipas J., et al. Analysis of microRNAs in pancreatic fine-needle aspirates can classify benign and malignant tissues. Clin. Chem. 2008;54:1716–1724. doi: 10.1373/clinchem.2008.109603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Wang J., Chen J., Chang P., LeBlanc A., Li D., Abbruzzesse J.L., Frazier M.L., Killary A.M., Sen S. MicroRNAs in plasma of pancreatic ductal adenocarcinoma patients as novel blood-based biomarkers of disease. Cancer Prev. Res. 2009;2:807–813. doi: 10.1158/1940-6207.CAPR-09-0094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Greither T., Grochola L., Udelnow A., Lautenschlager C., Wurl P., Taubert H. Elevated expression of microRNAs 155, 203, 210 and 222 in pancreatic tumours associates with poorer survival. Int. J. Cancer. 2010;102:731–737. doi: 10.1038/sj.bjc.6605520. [DOI] [PubMed] [Google Scholar]
- 43.Boone B.A, Steve J., Krasinskas A.M., Zureikat A.H., Lembersky B.C., Gibson M.K., Stoller R.G., Zeh H.J., Bahary N. Outcomes with FOLFIRINOX for borderline resectable and locally unresectable pancreatic cancer. J. Surg. Oncol. 2013;108:236–224. doi: 10.1002/jso.23392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Ychou M., Conroy T., Seitz J.F., Gourgou S., Hua A., Mery-Mignard D., Kramar A. An open phase I study assessing the feasibility of the triple combination: Oxaliplatin plus irinotecan plus leucovorin/5-fluorouracil every 2 weeks in patients with advanced solid tumors. Ann. Oncol. 2003;14:481–489. doi: 10.1093/annonc/mdg119. [DOI] [PubMed] [Google Scholar]
- 45.Assaf E., Verlinde-Carvalho M., Delbaldo C., Grenier J., Sellam Z., Pouessel D., Bouaita L., Baumgaertner I., Sobhani I., Tayar C., et al. 5-Fluorouracil/leucovorin combined with irinotecanand oxaliplatin (FOLFIRINOX) as second-line chemotherapy in patients with metastatic pancreatic adenocarcinoma. Oncology. 2011;80:301–306. doi: 10.1159/000329803. [DOI] [PubMed] [Google Scholar]
- 46.Conroy T., Paillot B., Francois E., Bugat R., Jacob J.H., Stein U., Nasca S., Metges J.P., Rixe O., Michel P., et al. Irinotecan plus oxaliplatin and leucovorin-modulated fluorouracil in advanced pancreatic cancer-A groupe tumeurs digestives of the federation nationale des centres de lutte contre le cancer study. J. Clin. Oncol. 2005;23:1228–1236. doi: 10.1200/JCO.2005.06.050. [DOI] [PubMed] [Google Scholar]
- 47.Peddi P.F., Lubner S., McWilliams R., Tan B.R., Picus J., Sorscher S.M., Suresh R., Lockhart A.C., Wang J., Menias C., et al. Multi-institutional experience with FOLFIRINOX in pancreatic adenocarcinoma. JOP. 2012;13:497–501. doi: 10.6092/1590-8577/913. [DOI] [PubMed] [Google Scholar]
- 48.Gourgou-Bourgade S., Bascoul-Mollevi C., Desseigne F., Ychou M., Bouché O., Guimbaud R., Bécouarn Y., Adenis A., Raoul J.L., Boige V., et al. Impact of FOLFIRINOX compared with gemcitabine on quality of life in patients with metastatic pancreatic cancer: Results from the PRODIGE 4/ACCORD 11 randomized trial. J. Clin. Oncol. 2013;31:23–29. doi: 10.1200/JCO.2012.44.4869. [DOI] [PubMed] [Google Scholar]
- 49.Costello E., Greenhalf W., Neoptolemos J.P. New biomarkers and targets in pancreatic cancer and their application to treatment. Nat. Rev. Gastroenterol. Hepatol. 2012;9:435–444. doi: 10.1038/nrgastro.2012.119. [DOI] [PubMed] [Google Scholar]
- 50.Giovannetti E., Funel N., Peters G.J., Del Chiaro M., Erozenci L.A., Vasile E., Leon L.G., Pollina L.E., Groen A., Falcone A., et al. MicroRNA-21 in pancreatic cancer: Correlation with clinical outcome and pharmacologic aspects underlying its role in the modulation of gemcitabine activity. Cancer Res. 2010;70:4528–4538. doi: 10.1158/0008-5472.CAN-09-4467. [DOI] [PubMed] [Google Scholar]
- 51.Ali S., Ahmad A., Banerjee S., Padhye S., Dominiak K., Schaffert J.M., Wang Z., Philip P.A., Sarkar F.H. Gemcitabine sensitivity can be induced in pancreatic cancer cells through modulation of miR-200 and miR-21 expression by curcumin or its analogue CDF. Cancer Res. 2010;70:3606–3617. doi: 10.1158/0008-5472.CAN-09-4598. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 52.miRBase: The microRNA database. [(accessed on 28 April 2014)]. Available online: http://www.mirbase.org/
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.