

Abstract
Microglia are prominent immune cells in the central nervous system (CNS) and are critical players in both neurological development and homeostasis, and in neurological diseases when dysfunctional. Our previous understanding of the phenotypes and functions of microglia has been greatly extended by a dearth of recent investigations. Distinct genetically defined subsets of microglia are now recognized to perform their own independent functions in specific conditions. The molecular profiling of single microglial cells indicates extensively heterogeneous reactions in different neurological disorders, resulting in multiple potentials for crosstalk with other kinds of CNS cells such as astrocytes and neurons. In settings of neurological diseases it could thus be prudent to establish effective cell‐based therapies by targeting entire microglial networks. Notably, activated microglial depletion through genetic targeting or pharmacological therapies within a suitable time window can stimulate replenishment of the CNS niche with new microglia. Additionally, enforced repopulation through provision of replacement cells also represents a potential means of exchanging dysfunctional with functional microglia. In each setting the newly repopulated microglia might have the potential to resolve ongoing neuroinflammation. In this review, we aim to summarize the most recent knowledge of microglia and to highlight microglial depletion and subsequent repopulation as a promising cell replacement therapy. Although glial cell replacement therapy is still in its infancy and future translational studies are still required, the approach is scientifically sound and provides new optimism for managing the neurotoxicity and neuroinflammation induced by activated microglia.
Keywords: cell replacement therapy, depletion, microglia, neuroinflammation
Main Points
Microglial depletion has mainly led to neuroprotective outcomes in neurological disease models.
Enforced microglial repopulation can replenish the empty niche in the CNS, maintain tissue integrity and potentially resolve neuroinflammation.

1. INTRODUCTION
Microglia are highly specialized and dynamic cellular components of the central nervous system (CNS) originating from embryonic precursors in the yolk sac, comprising approximately 10% of the total glial cell number in the adult brain (Ginhoux et al., 2010; Labzin, Heneka, & Latz, 2017; Li & Barres, 2017). Microglia have traditionally been considered to be in a resting and quiescent state in physiological conditions. With the advent of elegant two/multiple photon microscopy image techniques, genetic and molecular targeting tools, we now appreciate that in normal conditions microglia have a ramified morphology, are maintained by diverse signals from neurons and can continuously move their dendrites, which allows for constant active screening of the surrounding microenvironment (Kierdorf & Prinz, 2017; Nimmerjahn, Kirchhoff, & Helmchen, 2005).
Microglia are long‐lived cells with a relatively low turnover. By genetically labeling microglia in pathogen‐free mice it was recently determined that microglia can survive during the whole lifespan of an animal, and can thus exert crucial long‐lasting influences on neurodegenerative disorders (Fuger et al., 2017). However, it is well documented that microglia can be self‐regulated without contribution from peripheral myeloid cells and their turnover is tightly controlled by the coupling of apoptosis, with approximately 1% murine microglia dying in 1 day and the whole population of cells renewing several times throughout life (Askew et al., 2017; Tay et al., 2017). Although significant species differences in microglial biology such as microglial density were noted, this finding also concords with observations in humans, a recent study highlighting that more than 96% of human microglia can be slowly renewed throughout life (Reu et al., 2017).
Microglial cells are believed to play multifunctional roles in both inflammatory and physiological contexts (Grabert et al., 2016; Thompson & Tsirka, 2017). In the healthy brain microglia efficiently monitor CNS homeostasis and have a marked impact on neural development. In order to actively survey the CNS they have recently been demonstrated to require the proper activity of tandem‐pore domain halothane‐inhibited K+ channel 1, which is the main K+ channel expressed in microglial cells (Madry et al., 2017). In several pathological conditions such as epilepsy, single‐cell RNA sequencing of hippocampal microglia indicated that microglia undergo dramatically transcriptomic alterations (more than 2,000 differentially expressed genes) and immunological activation during early time points, particularly regarding mitochondrial activity and metabolic pathways (Bosco et al., 2018). As such they play an indispensable role in the inflammatory cascade. Some studies based on comprehensive single cell RNA sequencing experiments have reported that microglia do not vary considerably in the whole brain (Keren‐Shaul et al., 2017; Matcovitch‐Natan et al., 2016). However, a recent study provides further novel evidence that CD11b+ microglia in the circumventricular regions are actually maintained in the activated state even during physiological conditions (Takagi, Furube, Nakano, Morita, & Miyata, 2017). Microglia in this specific region not only display the amoeboid morphology rather than the ramified form, but also express high protein levels of activation markers in the healthy mouse brain (Takagi et al., 2017).
This recent report is consistent with the view that while microglia are uniformly distributed throughout the CNS they appear to perform characteristic functions in specific regions (De Biase et al., 2017; Marshall, Deleyrolle, Reynolds, Steindler, & Laywell, 2014). Indeed, genome‐wide transcriptional studies have reported that the bio‐energetic and immunoregulatory functions of microglia varied considerably in different anatomical regions, evidenced by cerebellar and cortical microglia displaying distinct gene expression profiles under steady‐state conditions (Grabert et al., 2016). More specifically, a recent study provides convincing evidence of an epigenetic mechanism involved in the clearance activity of microglia that differs regionally in the adult brain (Ayata et al., 2018). Variations in microglial profiles may also depend on the specific diseases states (Mastroeni et al., 2017), significantly altered transcripts having been reported in the hippocampus of Alzheimer's disease (AD) and in the substantia nigra of Parkinson's disease (PD), respectively (Mastroeni et al., 2017).
It is now well accepted that alterations in microglial activity and dysregulated microglial‐induced neuroinflammation have dual effects on many neurological diseases (Du et al., 2017; Salter & Stevens, 2017). Although the microglial field is intensively researched at present, less is still known about how microglia can be precisely targeted for optimal therapeutic efficacy. The concept of glial replacement therapy using progenitors has recently been proposed (Cartier, Lewis, Zhang, & Rossi, 2014; Shen, Li, Bao, & Wang, 2017; Srivastava, Bulte, Walczak, & Janowski, 2017). In this review we will introduce and discuss a new experimental paradigm to specifically control the excessive activation of microglia in vivo using fully differentiated and pre‐activated cells, and provide a rationale for its translation into clinical practice.
2. MULTI‐TASKING MICROGLIA: A FRIEND FOR BRAIN HOMEOSTASIS
Microglia can perform diverse functions to maintain overall tissue integrity during steady‐state conditions (Colonna & Butovsky, 2017; Kabba et al., 2017; Mosser, Baptista, Arnoux, & Audinat, 2017; Prinz, Erny, & Hagemeyer, 2017). Indeed, a growing body of evidence convincingly demonstrates that microglia are recognized for acting as “busy bees” and maintain an expanding array of functions during both early brain development and adult homeostasis (Figure 1). In particular, microglia can secrete a broad range of protective neurotrophic substances such as brain‐derived neurotrophic factor (BDNF), vascular endothelial growth factor, neuronal growth factor (NGF), insulin‐like growth factor‐1 (IGF‐1), platelet‐derived growth factors and transforming growth factor‐β (TGF‐β) (Butovsky et al., 2014; Parkhurst et al., 2013; Shibata & Suzuki, 2017; Wlodarczyk et al., 2017), thus ensuring appropriate neuronal network development and maintenance as well as enhancing memory and learning (Molteni & Rossetti, 2017; Parkhurst et al., 2013). There is a widespread consensus that microglia are also in active intimate contact with neighboring neuronal and non‐neuronal cells, thereby regulating neuronal proliferation, migration and differentiation and refining the neural circuits (Frost & Schafer, 2016; Mosser et al., 2017).
Figure 1.
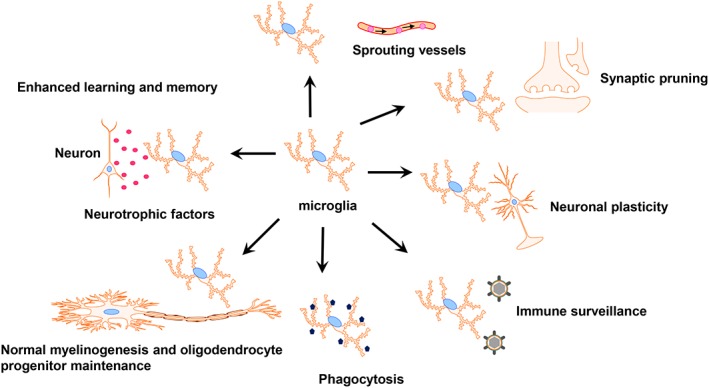
The multi‐tasking microglia in the CNS. Microglia can perform diverse functions to maintain overall tissue integrity at steady‐state conditions including: Enhancing memory and learning, maintaining oligodendrocyte progenitors and contributing to the myelinogenesis, actively screening the surroundings, involving in neural repair by phagocytic scavenging, remodeling the brain circuits through synaptic pruning and neuronal plasticity, and sprouting vessels
In order to perform these above‐mentioned functions a diverse array of receptors including TAM receptors, glutamate receptors, and purinergic receptors are used by microglia to efficiently communicate with other cells (Fourgeaud et al., 2016; York, Bernier, & MacVicar, 2017). Among these complex systems the CX3CL1/CX3CR1 and CD200‐CD200R axes play key roles in microglia–neuron contact (Eyo & Wu, 2013; Kierdorf & Prinz, 2017; Limatola & Ransohoff, 2014; Mecca, Giambanco, Donato, & Arcuri, 2018). Indeed, the CX3CR1−/− mouse exhibited profound alterations in both morphology and connectivity of the mature newborn hippocampal granule neurons (Bolos et al., 2017). The CD200−/− mouse exhibits an activated microglial phenotype accompanied by high expression of CD11b and CD45 (Hoek et al., 2000). Concurrently, microglia are critically involved in neural repair through phagocytic scavenging, such as clearing dead tissues, ingesting plaques, and apoptotic cells (Michell‐Robinson et al., 2015).
Furthermore, microglia participate in synaptogenesis by producing neurotrophic substances, such as BDNF, as well as eliminating excessive presynaptic and postsynaptic elements through synaptic pruning via the activation of the complement pathway (Masuda & Prinz, 2016; Sominsky, De Luca, & Spencer, 2017; Stevens et al., 2007; Um, 2017). In addition to the complement‐mediated mechanism, astrocyte‐derived interleukin (IL)‐33 is also physiologically required to maintain synapse homeostasis by modulating microglial synapse engulfment (Vainchtein et al., 2018). Microglia are involved in regulating and shaping both excitatory and inhibitory synapses, such as γ‐aminobutyric acid‐expressing and glycinergic synapses (Cantaut‐Belarif et al., 2017; Um, 2017). Conversely, early evidence obtained from CX3CR1 gene‐deleted mice indicated that reduced numbers of microglial cells during brain development could impair the processes of synaptic pruning, resulting in a significantly higher density of dendritic spines and immature synapses (Paolicelli et al., 2011; Shibata & Suzuki, 2017). During adult neurogenesis, microglia actively contribute to regulate the dynamics, maintenance, and functions of synapses of adult‐born neurons (Reshef et al., 2017).
In addition to their well‐described immunological roles, newly emerging neurobiological functions of microglia are currently being recognized and studied. It has been recently determined that a subpopulation of transiently activated microglia, identified in early postnatal white matter region, can directly contribute to the maintenance of oligodendrocyte progenitor numbers and subsequent myelinogenesis in the mouse, since a decreased oligodendrocyte progenitor number was noted following injection of the selective colony‐stimulating factor 1 receptor (CSF‐1R) inhibitor BLZ945 that effectively depletes microglia (Hagemeyer et al., 2017). Other recent observations provide evidence that microglia actively regulate neurovascular homeostasis, such as forming new blood vessels and the vascular branching of the retina and hindbrain (Arcuri, Mecca, Bianchi, Giambanco, & Donato, 2017; Brandenburg et al., 2016; Dudvarski Stankovic, Teodorczyk, Ploen, Zipp, & Schmidt, 2016; Zhao, Eyo, Murugan, & Wu, 2018). In a historical perspective remarkable progress has been made in deciphering many aspects of microglial biology. Further in vivo studies are still warranted to characterize and explore the versatile features of microglia.
3. THE ROLE OF MICROGLIA IN NEUROLOGICAL DISEASES: FRIEND OR FOE?
Apart from beneficial roles in the development of CNS, microglia can be widely involved in various types of neurological disorders, including stroke (Guruswamy & ElAli, 2017; Kronenberg et al., 2017), multiple sclerosis (MS) (Bogie, Stinissen, & Hendriks, 2014; Luo et al., 2017), AD (Hansen, Hanson, & Sheng, 2017; Sarlus & Heneka, 2017), PD (Subramaniam & Federoff, 2017), sleep disorders (Nadjar, Wigren, & Tremblay, 2017), amyotrophic lateral sclerosis (ALS) (Geloso et al., 2017; Liu & Wang, 2017), Huntington's disease (H. M. Yang, Yang, Huang, Tang, & Guo, 2017), epilepsy (Eyo, Murugan, & Wu, 2017; Zhao, Liao, et al., 2018), gliomas (Hambardzumyan, Gutmann, & Kettenmann, 2016; Schiffer, Mellai, Bovio, & Annovazzi, 2017), Prion diseases (Aguzzi & Zhu, 2017; Obst, Simon, Mancuso, & Gomez‐Nicola, 2017), psychiatric disorders (Mondelli, Vernon, Turkheimer, Dazzan, & Pariante, 2017; Prinz & Priller, 2014; Setiawan et al., 2018; Singhal & Baune, 2017), neuropathic pain (Inoue & Tsuda, 2018; Peng et al., 2016), adrenomyeloneuropathy (Gong et al., 2017), and traumatic brain injury (Donat, Scott, Gentleman, & Sastre, 2017). In general, microglia can be rapidly activated depending upon different stimulatory contexts and environmental changes through diverse molecular and cellular programs, subsequently transforming into the activated state and enhancing the expression of the Toll‐like receptors which sensitively bind microbial structures (Arcuri et al., 2017).
In addition to these morphological changes, activated microglia can acquire an altered gene expression pattern toward functionally distinct phenotypes. Once provoked, microglia produce pro‐inflammatory cytokines and chemokines, such as IL‐1, IL‐6, IL‐23, interferon gamma‐γ (IFN‐γ), CC chemokine ligand 2 (CCL2) and tumor necrosis factor‐α (TNF‐α) (Natoli et al., 2017; Nicolas et al., 2017; Smith, Das, Ray, & Banik, 2012), which are toxic to neighboring neurons and other glial cells such as astrocytes and oligodendrocytes. Nevertheless, activated microglia are more than simply destructive, it being widely recognized that immunoregulatory microglia are required for regulating brain repair and regeneration by secreting anti‐inflammatory factors, such as IL‐4, IL‐10, IL‐13, and TGF‐β (Cherry, Olschowka, & O'Banion, 2014; X. Jin & Yamashita, 2016).
Despite that microglia are necessary for repairing the CNS in normal conditions, activated microglia suppress the processes of brain repair by inhibiting the differentiation of oligodendrocyte precursor cells into myelinating oligodendrocytes via distinct mechanisms including heat shock protein 60 production, NO‐dependent oxidative damage and TNF‐α signaling (Li, Zhang, et al., 2017; Pang et al., 2010). Following pathological alterations, extracellular adenosine triphosphate, a source of energy metabolism, produced by dead and injured neurons can in turn activate microglia via purinergic receptors (Liu & Wang, 2017). Excessive activation of microglia can induce mitochondrial damage and decrease mitochondrial oxygen consumption depending on the degree of their activity, thus influencing total brain energy metabolism and exacerbating disease states (Ghosh, Castillo, Frias, & Swanson, 2017). Activated microglia can also be viewed as a major source of reactive oxidative stress through the release of reactive oxygen species and reactive nitrogen species, which cause oxidative damage to neurons and exacerbate the inflammatory cascade.
Moreover, excessively activated microglia have detrimental effects on neurogenesis, contribute to neuronal death and aggravate long‐term neurological deficits by hindering axonal regeneration (Kitayama, Ueno, Itakura, & Yamashita, 2011; Papageorgiou et al., 2016; Rodriguez et al., 2017). In vivo, microglia may even have a functional interaction with peripherally derived immune cells in the circulation in healthy subjects (Kanegawa et al., 2016). A recent exciting study has reported that activation of microglia plays a central role in inducing neurotoxic reactive astrocytes through production of IL‐1α, TNF, and C1q. The reactive astrocytes induced by activated microglia, termed A1, lose their homeostatic functions and consequently contribute to the death of neurons and differentiated oligodendrocytes (Liddelow et al., 2017). In addition, activated microglia have been shown to inhibit anti‐inflammatory TGF‐β signaling by downregulating TGF‐β‐regulated gene expression and this prolongs chronic microglial dysfunction, partly through the NF‐κB signaling pathway (Affram, Mitchell, & Symes, 2017). In this way microglial cells may prevent a return to normal homeostatic function. Conversely, if chronic inflammation mediated by persistent dysfunction of microglia remains after injury, this could lead to further extensive tissue damage, neuronal loss and cognitive deficits (Donat et al., 2017). In summary, although microglia are essential for the development and normal function of the CNS, dysregulated microglia contribute to disease severity in various neurological pathologies.
4. SUBSETS OF MICROGLIA: MORE THAN PRO‐INFLAMMATORY AND IMMUNOREGULATORY
Similar to macrophages in the periphery and other organs, microglia exhibit differential activation states generally associated with pro‐inflammatory or immunoregulatory functions (Mecha, Carrillo‐Salinas, Feliu, Mestre, & Guaza, 2016). The former contribute to neuroinflammation and neuronal injury through secretion of pro‐inflammatory and neurotoxic mediators, whereas the latter exert crucial influences on tissue repair through production of anti‐inflammatory cytokines (Tang & Le, 2016). We have argued that it is functional states and not in vitro induced surface marker expression phenotypes that should be used to describe activated myeloid cells (Mills, Harris, & Ley, 2015), and the classification of M1 and M2 functionally polarized states for microglia is now considered less valid (Ransohoff, 2016). Despite this conceptual development, at least nine subgroups of macrophage activation states based on transcriptome‐based network analysis have been proposed (Xue et al., 2014). It has been mostly ignored that novel subpopulations of macrophages such as CD169+ macrophages and T cell receptor positive (TCR+) macrophages may play different roles during specific conditions (Chavez‐Galan, Olleros, Vesin, & Garcia, 2015).
We are beginning to recognize that activated microglia are also quite plastic cells that may differentiate into a plethora of subsets and perform various functions in response to different stimuli and environmental changes such as obesity, diet, alcohol, and even the host microbiota (Chunchai, Chattipakorn, & Chattipakorn, 2017; Erny et al., 2015; Hanamsagar & Bilbo, 2017; Henriques et al., 2017; Johnson, 2015; Mathys et al., 2017; Menzel et al., 2017). In support of this, it has recently been reported that microglia can even respond to microbial challenges during embryogenesis, and that the absent status of a microbiome has a quite different impact on both age‐ and sex‐specific microglial gene expression (Thion et al., 2017). Furthermore, emerging data have indicated that mixed activation states are evident in later active lesions of MS whereby the activated microglia did not express purinergic receptor P2RY12, which is a novel and specific marker for homeostatic microglia (Zrzavy et al., 2017). Microglial cells may also display distinct activation patterns in the setting of AD, recently evidenced by different morphological properties between plaque‐associated and plaque‐distinct microglia using three‐dimensional cell analysis in an AD animal model (Plescher et al., 2018).
Newly emerging phenotypes of microglia continue to be described and investigated. Specifically, the existence of disease‐associated microglia during pathological states, for example in the APP‐PS1 mouse and the CX3CR1 knockout mouse, has been indicated at the ultrastructural level by transmission electron microscopy (Bisht, Sharma, Lecours, et al., 2016). Even though such disease‐associated microglia express microglial markers, such as CD11b, IBA1, and TREM2, this specific subset of cells has been suggested to be strikingly different from normal microglia since they exhibit signs of oxidative stress and transpire to be phagocytically hyperactive with highly ramified processes during pathological states. Such disease‐associated microglia can also cause abnormal interactions with synapses and lead to cognitive decline and learning deficits (Bisht, Sharma, Lacoste, & Tremblay, 2016; Bisht, Sharma, Lecours, et al., 2016). Interestingly, another unique neonatal white matter‐associated CD11c+ microglial subset has been described that specifically contributes to myelination and neurogenesis during postnatal brain development by producing neuroprotective IGF1. Although these repopulating CD11c+ microglia were evident following genetic adult microglial depletion using CX3CR1CreER iDTR mice, the newly repopulated CD11c+ microglia did not exhibit a neurogenic gene expression profile characteristic of the neonatal CD11c+ microglial subset (Bennett & Barres, 2017; Wlodarczyk et al., 2017). Consistently, the same research group also determined that CD11c+ microglia and CD11c− microglia have different roles in the contexts of cuprizone‐induced demyelination, experimental autoimmune encephalomyelitis (EAE) and neuromyelitis optica‐like disease models (Wlodarczyk et al., 2015).
Moreover, another unique type of disease‐associated microglia has been discovered in AD mice (through using massively parallel single‐cell RNA sequencing) that perform protective functions by restricting progression in neurodegenerative diseases (Keren‐Shaul et al., 2017). TREM2 is a receptor for the activation of disease‐associated microglia and the TREM2 signaling pathway has been considered as a common microglial activation pathway in several neurodegenerative conditions and aging (Deczkowska et al., 2018; Krasemann et al., 2017). Compared with normal microglia the disease‐associated microglia progressively increase lipid metabolism and expression of phagocytic genes, and are specifically localized around AD plaques, thereby engulfing neurotoxic proteins and protecting against neurodegenerative diseases (Keren‐Shaul et al., 2017). However, recent discoveries using unbiased weighted co‐expression network analysis have shed light on another distinct pro‐inflammatory subpopulation of disease‐associated microglia in the context of AD (Rangaraju et al., 2018).
Currently, it is still unknown whether other disease‐associated microglia subsets in different neurological disorders exist or not, or indeed whether specific subtypes are common to several neurological disease states. However, we are optimistic that further studies employing comprehensive combined transcriptome, proteomic and metabolomic approaches will define a wider spectrum of microglia subsets in different neurological disorders and facilitate targeting microglia with unprecedented precision (Haimon et al., 2018; Rangaraju et al., 2018). Thus further studies are warranted to characterize the scope of intrinsic mechanisms of disease‐associated microglia during diverse disease conditions.
5. DEPLETING MICROGLIA IN DISEASES: THE FRIEND IN NEED MAY NOT ALWAYS BE DESIRABLE INDEED
A novel CX3CR1CreER R26Confetti multiple reporter system has been used to demonstrate that microglia can regulate their cell number by clonally forming a cluster at the site of local injury (Madore, Baufeld, & Butovsky, 2017; Tay et al., 2017). As described above, reactive microglia can lead to tissue damage, exacerbate deleterious effects and potentially contribute to neurodegeneration (Leyns & Holtzman, 2017). As it follows that microglia play a central role in many neurological disorders (Salter & Stevens, 2017; Wendeln et al., 2018), then microglia‐directed therapy, including specific microglia depletion strategies, can be regarded as a promising immunotherapy for neurological diseases (Du et al., 2017; Feng et al., 2017; Rice et al., 2015). In order to specifically deplete microglia in an injured CNS, many approaches including pharmacological inhibition (e.g. targeting the CSF‐R1 receptor with PLX5562) and genetic targeting have been developed (Waisman, Ginhoux, Greter, & Bruttger, 2015), and we have previously reviewed the outcomes of these approaches (Han, Harris, & Zhang, 2017; Lund, Pieber, & Harris, 2017).
In support of the hypothesis, microglial depletion has thus led to a broad range of positive and neuroprotective outcomes in distinct disease conditions by reducing neuroinflammation (Table 1): ameliorating EAE (Heppner et al., 2005; Lassmann & Bradl, 2017; Nissen, Thompson, West, & Tsirka, 2018); enhancing remyelination in the cuprizone demyelination model (Beckmann et al., 2018); attenuating neurological https://www.google.se/search?q=abnormalities&spell=1&sa=X&ved=0ahUKEwi38NK94bfYAhUMMJoKHSPtDHEQvwUIJCgA and brain edema in two stroke mouse models (Li, Li, et al., 2017); reducing mRNA levels of proinflammatory cytokines such as IL‐1β and TNF‐α induced by peripheral lipopolysaccharide injection after microglial depletion (Xie et al., 2017); or acute binge ethanol withdrawal with little negative effects on behavioral functions (Walter & Crews, 2017). In one of our previous studies, we have indicated that cranial irradiation can cause transient accumulation of microglial cells followed by persistent inflammation and pronounced expression of IL‐1β and CCL2 in the hippocampus (Han et al., 2016). Indeed, neuroinflammation induced by activated resident microglia as well as infiltrating monocytes plays a pivotal role in hippocampal‐dependent severe cognitive dysfunction after both acute and long‐term irradiation (Feng et al., 2016) and microglial depletion can ameliorate these cranial radiation‐induced cognitive deficits (Acharya et al., 2016). Confirmed in another study, PLX5662‐mediated depletion can lead to protection from loss of dendritic spines, reduction of CD11b+Ly6G−Ly6Chi monocytes in the blood, inhibition of monocyte accumulation and ultimately prevention of radiation‐induced cognitive https://www.google.se/search?q=abnormalities&spell=1&sa=X&ved=0ahUKEwjGqt_3grnYAhXnApoKHQ77AlIQvwUIJCgA (Feng et al., 2016). Furthermore, PLX5562 treatment can also alleviate two different forms of Charcot–Marie–Tooth disease by reducing neuropathic features, axonal damage and promoting hindlimb grip strength (Klein et al., 2015; Scherer, 2015). One more study showed that microglial depletion by PLX5562 treatment prevented the symptom of catatonia in structural myelin protein Cnp−/− mouse by reducing neuroinflammation and neurodegeneration (Janova et al., 2017).
Table 1.
The outcomes and physiological effects of microglial depletion in different disease conditions
| Depletion ways | Disease conditions | Outcomes | Physiological effects | References |
|---|---|---|---|---|
| CD11b‐HSVTK | EAE (MS) | Beneficial | Ameliorates clinical manifestations and reduces infiltrating cells | Heppner et al. (2005) |
| PLX5622 | EAE (MS) | Beneficial | Improves mobility by increasing mature oligodendrocytes | Nissen et al. (2018) |
| BLZ945 | Cuprizone model (MS) | Beneficial | Enhances remyelination in the striatum and cortex | Beckmann et al. (2018) |
| PLX3397 | Intracerebral hemorrhage | Beneficial | Attenuates neurological https://www.google.se/search?q=abnormalities&spell=1&sa=X&ved=0ahUKEwi38NK94bfYAhUMMJoKHSPtDHEQvwUIJCgA and brain edema | Li, Li, et al. (2017) |
| PLX5622 | Charcot–Marie‐tooth | Beneficial | Improves axonal integrity and muscle weakness | Klein et al. (2015) |
| PLX3397 | AD | Beneficial | Improves the spatial and emotional memory deficits | Sosna et al. (2018) |
| GW2580 | AD | Beneficial | Prevents the progression of AD pathology | Olmos‐Alonso et al. (2016) |
| PLX5622 | AD | Beneficial | Improves the hippocampal‐dependent tasks | Dagher et al. (2015) |
| PLX3397 | AD | Beneficial | Improves contextual memory deficits but not Aβ pathology | Spangenberg et al. (2016) |
| CX3CR1CreDTR | AD | Not clear | Leads to enlargement of Aβ plaques but not number of plaques | Zhao et al. (2017) |
| PLX3397 | AD | Beneficial | Inhibits the propagation of tau and reduces the excitability | Asai et al. (2015) |
| GW2580 | ALS | Beneficial | Attenuates motor neuron cell death and extends life expectancy | Martinez‐Muriana et al. (2016) |
| CD11b‐HSVTKmt | ALS | No effect | Has little effect on motor neuron degeneration and reflex scores | Gowing et al. (2008) |
| PLX3397 | ALS | Harmful | Reduces evoked compound muscle action potentials | Spiller et al. (2018) |
| PLX5622 | AUD | Beneficial | Enhances induction of anti‐inflammatory genes | Walter et al. (2017) |
| PLX5622 | Catatonia | Beneficial | Alleviates catatonic signs and reduces white matter inflammation | Janova et al. (2017) |
| PLX5622 | Radiation‐induced memory deficits | Beneficial | Prevents memory deficits by inhibiting monocyte accumulation | Feng et al. (2016) |
| PLX5622 | Radiation‐induced memory deficits | Beneficial | Attenuates microglial activation in the irradiated hippocampus and ameliorates radiation‐induced cognitive deficits | Acharya et al. (2016) |
| PLX5622 | Neuropathic pain | Beneficial | Alleviates both mechanical and cold allodynia | Lee et al. (2018) |
| LEC | Neuropathic pain | Beneficial | Reduces initiation rather not maintenance of neuropathic pain | Wang et al. (2018) |
| PLX3397 | Neuronal injury | Beneficial | Improves recovery by modulating inflammatory signals | Rice et al. (2015) |
| PLX5622 | POCD | Beneficial | Reduces hippocampal pro‐inflammatory cytokines and inhibits CCR2‐expressing cells infiltration | Feng et al. (2017) |
| PLX3397 | Cerebral ischemia | Harmful | Promotes leukocyte infiltration and exacerbates brain infarction | Jin et al. (2017) |
| PLX3397 | Cerebral ischemia | Harmful | Causes neuronal death and increases infarct size | Szalay et al. (2016) |
| PLX3397 | PD | Harmful | Increases MPTP neurotoxicity and augments neurodeficits | Yang et al. (2018) |
| PLX5622 | Coronavirus encephalitis | Harmful | Delays virus clearance and promotes immune cells infiltration | Wheeler et al. (2018) |
AUD = Alcohol use disorder; MS = Multiple sclerosis; EAE = experimental autoimmune encephalomyelitis; AD = Alzheimer's disease; ALS = Amyotrophic lateral sclerosis; PD = Parkinson's disease; MPTP = 1‐methyl‐4‐phenyl‐1,2,3,6‐tetrahydropyridine; TDP43 = TAR DNA‐binding protein 43; LEC = Liposome‐encapsulated clodronate; POCD = Postoperative cognitive decline.
Eliminating microglia in AD mice also provided beneficial outcomes including reduced neuronal loss, improvement of memory functions and partially preventing the progression of AD pathology, but had little effects on amyloid levels and plaque loads (Asai et al., 2015; Dagher et al., 2015; Olmos‐Alonso et al., 2016; Spangenberg et al., 2016). One recent study has indicated that microglial depletion in the CX3CR1CreER DTR transgenic mouse can lead to enlargement of Aβ plaques and may cause extensive neurite damage (Zhao, Hu, Tsai, Li, & Gan, 2017). However, early long‐term pharmacologically microglial depletion could finally inhibit plaque deposition and amyloid formation in the 5XFAD mouse model of AD, together with a relatively low level of soluble fibrillar oligomers in the brain (Sosna et al., 2018). Furthermore, the administration of GW2580 (tyrosine kinase inhibitor) orally can regulate inflammation of both the CNS and peripheral nervous systems in the ALS animal model, attenuating motoneuronal cell death, slowing disease progression and extending life expectancy (Martinez‐Muriana et al., 2016). However, this has not been confirmed in other studies using different approaches to deplete microglia (Gowing et al., 2008; Spiller et al., 2018). A beneficial outcome of microglial depletion is also evident in neuropathic pain by reducing the expression of pro‐inflammatory cytokines (Lee, Shi, Fan, West, & Zhang, 2018), which was also confirmed in another study (Wang, Mao, Wu, & Wang, 2018).
Despite this intensive research, microglia depletion as a treatment paradigm is still in its infancy. It is also of importance to note the differences of microglia activation in different types of disease processes. The findings mentioned above have to be interpreted with caution until we are sufficiently knowledgeable. On the basis of the above results, some critical issues need further investigation. For example, microglial elimination has been shown to exacerbate brain neurotoxicity in the contexts of brain ischemia, PD and coronavirus encephalitis (Jin et al., 2017; Szalay et al., 2016; Wheeler, Sariol, Meyerholz, & Perlman, 2018; Yang et al., 2018). Most microglial depletion studies have been performed using CSF‐1R inhibitors several weeks before inducing the animal disease models. However, it is more reasonable from a clinical perspective to deplete the activated microglial cells after disease onset. Furthermore, current research findings employing CSF‐1R kinase inhibitors (including PLX3397, PLX5562, GW2580, and BLZ945) might target multiple cell populations other than microglia (e.g. meningeal, perivascular and choroid plexus macrophages and microglial progenitor cells) (Yang et al., 2018). Cautious interpretation of pure microglial depletion effects using systemic delivery of current available CSF‐1R inhibitors is thus warranted. Indeed, recent studies have reported that CSF‐1R inhibition can deplete IBA+ macrophages in the kidney (Chalmers et al., 2017). In addition, CSF‐1R inhibition can robustly decline the actions of nerve‐associated macrophages in the periphery (Klein et al., 2015). However, several studies have identified some potential markers that are exclusively expressed in microglia including Tmem119, P2RY12, Siglec‐H and Sal1, permitting distinction of resident microglia from other myeloid cells (Bennett et al., 2016; Butovsky et al., 2014; Buttgereit et al., 2016; Konishi et al., 2017). These markers, together with the Cre/loxP technology, may make it possible to deplete microglia with exquisite precision in the near future.
Last but not the least, the potential side‐effects of microglial depletion is another crucial aspect to be considered. Under non‐sterile conditions the depletion of microglia might induce at least a transient immunodeficiency that could be harmful both with respect to CNS infection and also in disrupting normal CNS homeostatic functionality in unaffected brain areas. Indeed, the removal of immunomodulatory microglia producing and responding to TGF‐β could be expected to be a traumatic event in itself. The selective replacement of dysfunctional or aberrantly activated microglia in affected areas would thus be the most efficient approach. Most studies have indicated that no obvious behavioral consequences in adult mice following microglial depletion, despite extensive neuronal loss in the brain (Dagher et al., 2015; Elmore, Lee, West, & Green, 2015; Elmore et al., 2014; Rice et al., 2017). Microglial depletion in the adult wide‐type C57BL/6 mouse does not influence BDNF expression, the response of astrocytes and leukocyte infiltration in the experimental setting of PD (Yang et al., 2018). By contrast, neonatal microglial depletion during early life using pharmacological strategies can have a persistent impact on related motivated behavioral domains during adulthood (Nelson & Lenz, 2017). Furthermore, new tools to specifically deplete microglia by utilizing genetic approaches are in great demand. In a recent study, we have indicated that microglia can be efficiently depleted through the administration of tamoxifen in both CX3CR1CreER DTA and CX3CR1CreER DTR transgenic mice (Figure 2a) (Lund et al., 2018). This system thus allows for depletion of microglia at different disease stages by controlling the timing of tamoxifen provision.
Figure 2.
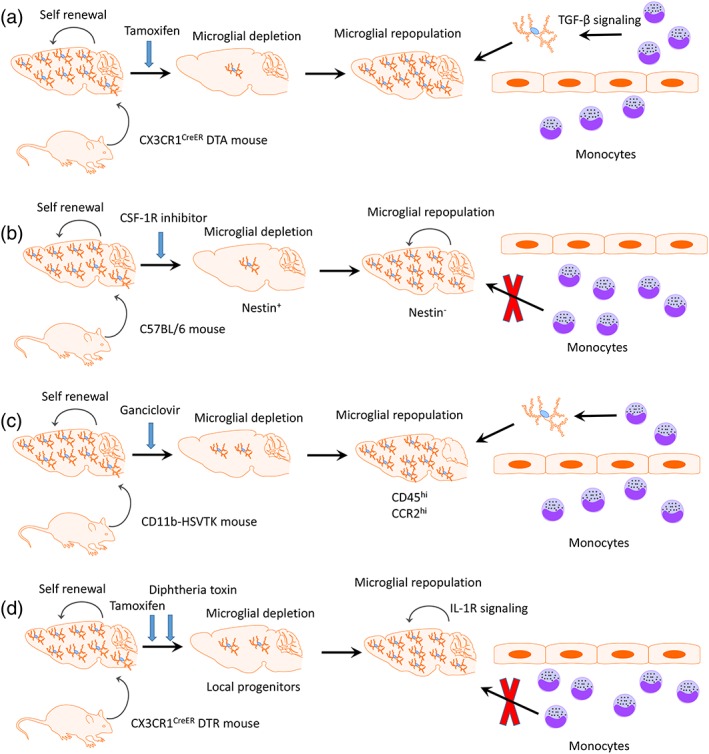
Microglial depletion and repopulation by different approaches. (a) Microglia can be efficiently depleted by the administration of tamoxifen in CX3CR1CreER DTA transgenic mouse. TGF‐β signaling is required for the peripheral myeloid cells invading the brain to colonize the functional microglial niche. (b) The nestin+ repopulated cells are transiently expressed after microglia depletion by CSF‐1R inhibitor. Newly repopulated microglia origin from resident microglial pool in the CNS rather than nestin+ progenitor cells as well as circulating monocytes. (c) Circulating monocytes, which markedly expressed CD45 and CCR2, can replace the adult CNS myeloid niche after microglial depletion in CD11b‐HSVTK transgenic mouse. (d) Resident microglia proliferation mediated by IL‐1R signaling can refill the microglial niche after partial depletion in CX3CR1CreER iDTR transgenic mouse
6. MICROGLIAL REPOPULATION RESOLVES NEUROINFLAMMATION: NEW CELLS, NEW FRIENDS
Basic and clinical research demonstrates that suppressing the immune response by depleting autoreactive immune cells may re‐establish the immune balance (Wraith, 2017). Taking MS for example, monoclonal antibodies (including Rituximab and Ocrelizumab) that selectively target and deplete CD20+ B cells have been approved for the treatment of MS (Greenfield & Hauser, 2017; Hauser et al., 2017; Sabatino, Zamvil, & Hauser, 2018; Salzer et al., 2016). Our colleagues have demonstrated that relapsing–remitting MS patients with Rituximab as the initial treatment have better relapse control and tolerability in comparison with other disease‐modifying drugs (Granqvist et al., 2018; Spelman, Frisell, Piehl, & Hillert, 2017). Available MS treatments mainly resolve the peripheral inflammation but further specific and effective cell‐depletion therapies still represent a highly unmet medical need, especially in chronic disease states.
Due to the self‐renewal ability of microglia, following microglial depletion by pharmacological therapies or genetic targeting the empty CNS niche can be entirely repopulated within a relatively short period without serious side‐effects (Elmore et al., 2015; Han et al., 2017; Varvel et al., 2012). In a previous pioneering study, Elmore and colleagues indicated that rapid repopulation of microglial cells can be noted after administration of the selective CSF‐1R inhibitor PLX3397 (Elmore et al., 2014). They claimed that following cessation of PLX3397 treatment the newly repopulated cells did not arise from bone marrow‐derived cells, but instead from local nestin+ microglial progenitor cells in the brain parenchyma (Figure 2b) (Elmore et al., 2014). However, Bruttger et al demonstrated that following partial depletion (80%) in the CX3CR1CreER iDTR mouse, microglia proliferate by themselves rather than from nestin+ microglial progenitor cells to finally refill the niche (Figure 2d) (Bruttger et al., 2015; Jakel & Dimou, 2017). This mechanism has received further support (Askew et al., 2017) including using elegant fate‐mapping approaches, single‐cell RNA sequencing and parabiosis (Huang, Xu, Xiong, Sun, et al., 2018).
The recruitment of circulating precursors does not contribute to the resident microglial pool in the healthy CNS (Ajami, Bennett, Krieger, McNagny, & Rossi, 2011; Mildner et al., 2007). However, contradictory findings have been reported by using the CD11b‐HSVTK transgenic mouse in which circulating monocytes have the ability to potentially replace the adult CNS myeloid niche after microglial depletion (Figure 2c) (Varvel et al., 2012) and the newly engrafted peripheral cells have a unique functional phenotype compared with resident microglia (Cronk et al., 2018).
By contrast, two repopulating origins following microglial depletion in the retina have been described (Huang, Xu, Xiong, Qin, et al., 2018). One is the resident central‐emerging microglia in the optic nerve and the other is the extra‐retinal periphery‐emerging microglia from the ciliary body/iris (Huang, Xu, Xiong, Qin, et al., 2018). Two distinct resources including peripheral macrophages could also contribute to robust microglial regeneration independently of irradiation (Cronk et al., 2018). However, intrinsic regulatory mechanisms that mediate the replacement of microglia‐like cells after selective depletion are not yet fully understood.
One recent study provided novel evidence that CX3CR1 signaling may actively regulate microglial compensation in the retina since CX3CR1−/− deficient mice had significantly lower numbers of repopulated microglial during early recovery when compared to CX3CR1 signaling‐sufficient mice (Zhang et al., 2018). Furthermore, we have recently demonstrated that CNS repopulated monocytes required TGF‐β signaling to colonize the functional microglial niche following microglia depletion in the CX3CR1CreER DTA transgenic mouse model (Figure 2a) (Lund et al., 2018). Specific TGF‐β signaling deficiency on the new microglia‐like cells led to development of progressive motor disease similar to ALS‐like symptoms (Lund et al., 2018).
In this context, the next critical question is whether the newly engrafted microglia‐like cells including local hyperproliferation and/or bone marrow‐derived cells could completely adapt the embryonically seeded microglial phenotypes and functions. Even though they are numerically and morphologically different from embryonically seeded microglia, the newly repopulated cells can still perform the same general functions as resident microglia including constantly surveying the microenvironment and appropriately responding to the acute events (Varvel et al., 2012; Zhang et al., 2018). Additionally, embryonically seeded microglia and the newly repopulated cells may respond differently to environmental stimuli, as evidenced by two distinct cell types showing differential motilities in response to laser burn injury in vivo (Cronk et al., 2018). Using RNA sequencing it was determined that the gene profiles of fully repopulated microglia had been little influenced by administration of PLX5622 because no inflammatory related genes were up‐regulated or down‐regulated during depletion and repopulation processes (Huang, Xu, Xiong, Qin, et al., 2018). Consistent with this idea, Elmore and colleagues determined that repopulated microglia have relatively larger cell bodies than do resident microglia. These two types of microglial cells shared the same level of mRNA gene expression as well as similar responses to lipopolysaccharide stimulation (Elmore et al., 2015). Furthermore, mice with repopulated microglia did not exhibit any cognitive or behavioral https://www.google.se/search?biw=1280&bih=615&q=abnormalities&spell=1&sa=X&ved=0ahUKEwihtbSN1bLYAhUEQZoKHUVwCk4QvwUIJCgA (Elmore et al., 2015).
More interestingly, the same research group further reported that repopulated microglia have the ability to largely resolve the pro‐inflammatory response and promote functional recovery after brain damage by replacing the active and highly swollen microglia with normal ramified microglia, downregulating the expression of reactive microglial markers and reducing the levels of inflammatory‐related genes (Rice et al., 2017). Furthermore, peripherally derived microglia‐like cells remained transcriptionally and functionally distinct from microglia arising through local proliferation (Cronk et al., 2018). Further evaluations are needed to confirm if overall brain function can be affected by mixed engrafted microglia‐like cells during diverse neurological disease models.
7. ENFORCED MICROGLIAL REPOPULATION: A NEW PROSPECT FOR FIXING A DYSFUNCTIONAL NICHE
So microglial depletion can lead to natural repopulation of the empty niche through either (a) hyperproliferation of remaining microglia, (b) stimulation of microglial precursors, or (c) infiltrating of monocytes from the circulation. An interesting aspect of myeloid cell repopulation with peripheral monocytes is that transcriptomic analyses reveal different outcomes in different tissues (Guilliams & Scott, 2017). Monocytes can thus replace Kupffer cells in the liver (Scott et al., 2016) and alveolar macrophages in the lung (van de Laar et al., 2016) with almost identical cellular phenotypes, while in the CNS bone marrow monocyte‐derived microglia only become microglia‐like cells, retaining more than 2,000 differentially expressed genes compared to resident microglia (Cronk et al., 2018). It is currently unknown why the CNS should differ in this respect to other tissues, but indicates that environmental cues must be tissue‐specific and of varied instructional consequence in different tissues (Bennett et al., 2018). It is also apparent that the repopulation process is tightly regulated, cells only occupying available tissue niches and the repopulation process (through either surviving microglia proliferation or monocyte infiltration) being halted through as yet undetermined mechanisms once the tissue is full. In certain studies an “overshoot” of repopulating cell numbers appears to be adjusted through selective loss of cells so that a homeostatic numerical occupancy is achieved. The important concept herein is that myeloid cell niches can be both efficiently depleted of resident cells and that repopulating cells can occupy the available niche with partial or total restoration of normal homeostatic functionality.
Microglial repopulation can arise from either bone marrow‐derived elements or local self‐renewal proliferation to replenish the empty niche in the CNS, which can be therapeutically targeted (Varvel et al., 2012; Waisman et al., 2015). It is also important to note that the therapeutic outcomes of the newly derived microglia can be fundamentally different during these two processes, as evidenced by recent data indicating that bone marrow‐derived microglia are functionally distinct from yolk sac‐derived microglia (Cronk et al., 2018). It follows that there might be specific advantages depending on the disease state. For example, if microglia are genetically dysfunctional, then self‐proliferation following depletion will not help. If the dysfunction extends to all myeloid cells (e.g. TREM2 in Nasu‐Hakola disease, ALS, and X‐linked adrenoleukodystrophy) then even repopulation by infiltrating monocytes will also not be beneficial. As monocyte‐derived repopulating cells would have a potentially different response to systemic stimuli compared to normal repopulating microglia, this is also an issue to contemplate. In contrast, it has been shown that bone marrow‐derived cells are much more efficient in clearing amyloid beta deposits compared to their endogenous counterparts (Kawanishi et al., 2018; Simard, Soulet, Gowing, Julien, & Rivest, 2006) and so monocyte repopulation would potentially be more efficient in AD.
A final scenario is enforced repopulation of pre‐defined myeloid cell through adoptive transfer. Natural microglia are excluded from this scenario, but blood‐derived monocytes or bone marrow‐derived macrophages, either unactivated, pre‐activated or genetically modified, have potential, as do stem cell‐derived myeloid cells. In order to successfully generate a sufficient source of renewable microglia‐like cells, several different protocols using cultured human inducible pluripotent stem cells (hiPSCs) have been recently established (Abud et al., 2017; Pocock & Piers, 2018). Specifically, hiPSCs are cultured with neuroglia differentiation media by supplement of CSF1 and IL‐34 to differentiate into Tmem119+/P2RY12+ microglia‐like cells that perform phagocytic functions (Muffat et al., 2016). Another well‐characterized method has been used to differentiate human and murine hiPSCs into microglia‐like cells through a hematopoietic progenitor‐like intermediate stage by adding defined factors and then co‐culturing with astrocytes (Pandya et al., 2017). A recently developed protocol for the derivation of microglia‐like cells from human monocytes could be the adoptive cell of most practical and functional relevance (Sellgren et al., 2017).
Considering that we have demonstrated the beneficial action of adoptively transferred immunomodulatory macrophages to prevent pathogenesis in settings of both Type 1 Diabetes (Parsa et al., 2012) and EAE (Zhang, Lund, Mia, Parsa, & Harris, 2014) and that other researchers have consequently successfully therapeutically employed our protocol in settings of spinal cord injury (Ma et al., 2015) and in wound healing (Riabov et al., 2017) then conceptually an initial transfer of immunomodulatory macrophages (either stimulated or gene‐modified) to halt the neuroinflammatory process could then be followed by transfer of microglia‐like cells. These approaches may provide a potential novel therapeutic angle for a wide array of neurological disorders and we currently actively explore this potential.
We thus propose that an immunotherapy protocol comprising total microglial ablation followed by the immediate enforced repopulation of the available niche through the adoptive transfer of myeloid cells could be considered as a means of replacing dysfunctional microglia in neurodegenerative states such as ALS and AD (Figure 3).
Figure 3.
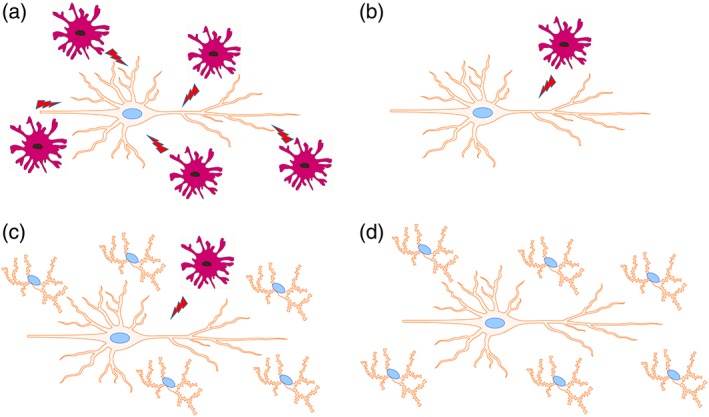
Potential scheme of microglial replacement therapy. (a) Activated microglia can be harmful to neurons at inflammatory conditions. (b) Selective ablation of microglia within suitable time window may reduce their deleterious effects. (c) Enforced repopulation through adoptively transferring nonactivated microglia or pre‐activated microglia with the desired activation phenotype (either stimulated or gene‐modified) can replenish the empty niche in the CNS. (d) The newly engrafted microglia can perform the normal functions and maintain overall tissue integrity
8. CONCLUDING REMARKS
Enforced microglia depletion and repopulation with fully differentiated cells, termed microglia‐replacement therapy, has great potential as an intervention in many neurological disease states. Optimization of timing between depletion and repopulation, long‐term effects on CNS homeostasis and long‐term therapeutic efficacy in preclinical models will provide a solid rationale for clinical translation.
CONFLICT OF INTEREST
The authors declare that there are no conflicts of interest.
ACKNOWLEDGMENTS
This work was supported by grants from the China Scholarship Council (Grant No. 201600160072 and 201700260280), the Swedish Medical Research Council, Neuroforbundet, HjärnFonden and BarnCancerFonden.
Han J, Zhu K, Zhang X‐M, Harris RA. Enforced microglial depletion and repopulation as a promising strategy for the treatment of neurological disorders. Glia. 2019;67:217–231. 10.1002/glia.23529
Funding information Barncancerfonden; China Scholarship Council, Grant/Award Numbers: 201700260280, 201600160072; Hjärnfonden; Neuroforbundet; Swedish Medical Research Council
Contributor Information
Xing‐Mei Zhang, Email: xingmei.zhang@ki.se.
Robert A. Harris, Email: robert.harris@ki.se.
REFERENCES
- Abud, E. M. , Ramirez, R. N. , Martinez, E. S. , Healy, L. M. , Nguyen, C. H. H. , Newman, S. A. , … Blurton‐Jones, M. (2017). iPSC‐derived human microglia‐like cells to study neurological diseases. Neuron, 94(2), 278–293 e279. 10.1016/j.neuron.2017.03.042 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Acharya, M. M. , Green, K. N. , Allen, B. D. , Najafi, A. R. , Syage, A. , Minasyan, H. , … Limoli, C. L. (2016). Elimination of microglia improves cognitive function following cranial irradiation. Scientific Reports, 6, 31545 10.1038/srep31545 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Affram, K. O. , Mitchell, K. , & Symes, A. J. (2017). Microglial activation results in inhibition of TGF‐beta‐regulated gene expression. Journal of Molecular Neuroscience, 63(3–4), 308–319. 10.1007/s12031-017-0980-3 [DOI] [PubMed] [Google Scholar]
- Aguzzi, A. , & Zhu, C. (2017). Microglia in prion diseases. The Journal of Clinical Investigation, 127(9), 3230–3239. 10.1172/JCI90605 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ajami, B. , Bennett, J. L. , Krieger, C. , McNagny, K. M. , & Rossi, F. M. (2011). Infiltrating monocytes trigger EAE progression, but do not contribute to the resident microglia pool. Nature Neuroscience, 14(9), 1142–1149. 10.1038/nn.2887 [DOI] [PubMed] [Google Scholar]
- Arcuri, C. , Mecca, C. , Bianchi, R. , Giambanco, I. , & Donato, R. (2017). The pathophysiological role of microglia in dynamic surveillance, phagocytosis and structural remodeling of the developing CNS. Frontiers in Molecular Neuroscience, 10, 191 10.3389/fnmol.2017.00191 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Asai, H. , Ikezu, S. , Tsunoda, S. , Medalla, M. , Luebke, J. , Haydar, T. , … Ikezu, T. (2015). Depletion of microglia and inhibition of exosome synthesis halt tau propagation. Nature Neuroscience, 18(11), 1584–1593. 10.1038/nn.4132 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Askew, K. , Li, K. , Olmos‐Alonso, A. , Garcia‐Moreno, F. , Liang, Y. , Richardson, P. , … Gomez‐Nicola, D. (2017). Coupled proliferation and apoptosis maintain the rapid turnover of microglia in the adult brain. Cell Reports, 18(2), 391–405. 10.1016/j.celrep.2016.12.041 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ayata, P. , Badimon, A. , Strasburger, H. J. , Duff, M. K. , Montgomery, S. E. , Loh, Y. E. , … Schaefer, A. (2018). Epigenetic regulation of brain region‐specific microglia clearance activity. Nature Neuroscience, 21(8), 1049–1060. 10.1038/s41593-018-0192-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beckmann, N. , Giorgetti, E. , Neuhaus, A. , Zurbruegg, S. , Accart, N. , Smith, P. , … Shimshek, D. R. (2018). Brain region‐specific enhancement of remyelination and prevention of demyelination by the CSF1R kinase inhibitor BLZ945. Acta Neuropathologica Communications, 6(1), 9 10.1186/s40478-018-0510-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bennett, F. C. , Bennett, M. L. , Yaqoob, F. , Mulinyawe, S. B. , Grant, G. A. , Hayden Gephart, M. ,… Barres, B. A. (2018). A combination of ontogeny and CNS environment establishes microglial identity. Neuron, 98(6):1170‐1183 doi: 10.1016/j.neuron.2018.05.014, 98, 1170, 1183.e8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bennett, M. L. , & Barres, B. A. (2017). A genetically distinct microglial subset promotes myelination. The EMBO Journal, 36(22), 3269–3271. 10.15252/embj.201798389 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bennett, M. L. , Bennett, F. C. , Liddelow, S. A. , Ajami, B. , Zamanian, J. L. , Fernhoff, N. B. , … Barres, B. A. (2016). New tools for studying microglia in the mouse and human CNS. Proceedings of the National Academy of Sciences of the United States of America, 113(12), E1738–E1746. 10.1073/pnas.1525528113 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bisht, K. , Sharma, K. , Lacoste, B. , & Tremblay, M. E. (2016). Dark microglia: Why are they dark? Communicative & Integrative Biology, 9(6), e1230575 10.1080/19420889.2016.1230575 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bisht, K. , Sharma, K. P. , Lecours, C. , Sanchez, M. G. , El Hajj, H. , Milior, G. , … Tremblay, M. E. (2016). Dark microglia: A new phenotype predominantly associated with pathological states. Glia, 64(5), 826–839. 10.1002/glia.22966 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bogie, J. F. , Stinissen, P. , & Hendriks, J. J. (2014). Macrophage subsets and microglia in multiple sclerosis. Acta Neuropathologica, 128(2), 191–213. 10.1007/s00401-014-1310-2 [DOI] [PubMed] [Google Scholar]
- Bolos, M. , Perea, J. R. , Terreros‐Roncal, J. , Pallas‐Bazarra, N. , Jurado‐Arjona, J. , Avila, J. , & Llorens‐Martin, M. (2017). Absence of microglial CX3CR1 impairs the synaptic integration of adult‐born hippocampal granule neurons. Brain, Behavior, and Immunity, 68, 76–89. 10.1016/j.bbi.2017.10.002 [DOI] [PubMed] [Google Scholar]
- Bosco, D. B. , Zheng, J. , Xu, Z. , Peng, J. , Eyo, U. B. , Tang, K. , … Wu, L. J. (2018). RNAseq analysis of hippocampal microglia after kainic acid‐induced seizures. Molecular Brain, 11(1), 34 10.1186/s13041-018-0376-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brandenburg, S. , Muller, A. , Turkowski, K. , Radev, Y. T. , Rot, S. , Schmidt, C. , … Vajkoczy, P. (2016). Resident microglia rather than peripheral macrophages promote vascularization in brain tumors and are source of alternative pro‐angiogenic factors. Acta Neuropathologica, 131(3), 365–378. 10.1007/s00401-015-1529-6 [DOI] [PubMed] [Google Scholar]
- Bruttger, J. , Karram, K. , Wortge, S. , Regen, T. , Marini, F. , Hoppmann, N. , … Waisman, A. (2015). Genetic cell ablation reveals clusters of local self‐renewing microglia in the mammalian central nervous system. Immunity, 43(1), 92–106. 10.1016/j.immuni.2015.06.012 [DOI] [PubMed] [Google Scholar]
- Butovsky, O. , Jedrychowski, M. P. , Moore, C. S. , Cialic, R. , Lanser, A. J. , Gabriely, G. , … Weiner, H. L. (2014). Identification of a unique TGF‐beta‐dependent molecular and functional signature in microglia. Nature Neuroscience, 17(1), 131–143. 10.1038/nn.3599 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buttgereit, A. , Lelios, I. , Yu, X. , Vrohlings, M. , Krakoski, N. R. , Gautier, E. L. , … Greter, M. (2016). Sall1 is a transcriptional regulator defining microglia identity and function. Nature Immunology, 17(12), 1397–1406. 10.1038/ni.3585 [DOI] [PubMed] [Google Scholar]
- Cantaut‐Belarif, Y. , Antri, M. , Pizzarelli, R. , Colasse, S. , Vaccari, I. , Soares, S. , … Bessis, A. (2017). Microglia control the glycinergic but not the GABAergic synapses via prostaglandin E2 in the spinal cord. The Journal of Cell Biology, 216(9), 2979–2989. 10.1083/jcb.201607048 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cartier, N. , Lewis, C. A. , Zhang, R. , & Rossi, F. M. (2014). The role of microglia in human disease: Therapeutic tool or target? Acta Neuropathologica, 128(3), 363–380. 10.1007/s00401-014-1330-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chalmers, S. A. , Wen, J. , Shum, J. , Doerner, J. , Herlitz, L. , & Putterman, C. (2017). CSF‐1R inhibition attenuates renal and neuropsychiatric disease in murine lupus. Clinical Immunology, 185, 100–108. 10.1016/j.clim.2016.08.019 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chavez‐Galan, L. , Olleros, M. L. , Vesin, D. , & Garcia, I. (2015). Much more than M1 and M2 macrophages, there are also CD169(+) and TCR(+) macrophages. Frontiers in Immunology, 6, 263 10.3389/fimmu.2015.00263 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cherry, J. D. , Olschowka, J. A. , & O'Banion, M. K. (2014). Neuroinflammation and M2 microglia: The good, the bad, and the inflamed. Journal of Neuroinflammation, 11, 98 10.1186/1742-2094-11-98 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chunchai, T. , Chattipakorn, N. , & Chattipakorn, S. C. (2017). The possible factors affecting microglial activation in cases of obesity with cognitive dysfunction. Metabolic Brain Disease, 33, 615–635. 10.1007/s11011-017-0151-9 [DOI] [PubMed] [Google Scholar]
- Colonna, M. , & Butovsky, O. (2017). Microglia function in the central nervous system during health and neurodegeneration. Annual Review of Immunology, 35, 441–468. 10.1146/annurev-immunol-051116-052358 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cronk, J. C. , Filiano, A. J. , Louveau, A. , Marin, I. , Marsh, R. , Ji, E. , … Kipnis, J. (2018). Peripherally derived macrophages can engraft the brain independent of irradiation and maintain an identity distinct from microglia. The Journal of Experimental Medicine, 215, 1627–1647. 10.1084/jem.20180247 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dagher, N. N. , Najafi, A. R. , Kayala, K. M. , Elmore, M. R. , White, T. E. , Medeiros, R. , … Green, K. N. (2015). Colony‐stimulating factor 1 receptor inhibition prevents microglial plaque association and improves cognition in 3xTg‐AD mice. Journal of Neuroinflammation, 12, 139 10.1186/s12974-015-0366-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Biase, L. M. , Schuebel, K. E. , Fusfeld, Z. H. , Jair, K. , Hawes, I. A. , Cimbro, R. , … Bonci, A. (2017). Local cues establish and maintain region‐specific phenotypes of basal ganglia microglia. Neuron, 95(2), 341–356 e346. 10.1016/j.neuron.2017.06.020 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deczkowska, A. , Keren‐Shaul, H. , Weiner, A. , Colonna, M. , Schwartz, M. , & Amit, I. (2018). Disease‐associated microglia: A universal immune sensor of neurodegeneration. Cell, 173(5), 1073–1081. 10.1016/j.cell.2018.05.003 [DOI] [PubMed] [Google Scholar]
- Donat, C. K. , Scott, G. , Gentleman, S. M. , & Sastre, M. (2017). Microglial activation in traumatic brain injury. Frontiers in Aging Neuroscience, 9, 208 10.3389/fnagi.2017.00208 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Du, L. , Zhang, Y. , Chen, Y. , Zhu, J. , Yang, Y. , & Zhang, H. L. (2017). Role of microglia in neurological disorders and their potentials as a therapeutic target. Molecular Neurobiology, 54(10), 7567–7584. 10.1007/s12035-016-0245-0 [DOI] [PubMed] [Google Scholar]
- Dudvarski Stankovic, N. , Teodorczyk, M. , Ploen, R. , Zipp, F. , & Schmidt, M. H. (2016). Microglia‐blood vessel interactions: A double‐edged sword in brain pathologies. Acta Neuropathologica, 131(3), 347–363. 10.1007/s00401-015-1524-y [DOI] [PubMed] [Google Scholar]
- Elmore, M. R. , Lee, R. J. , West, B. L. , & Green, K. N. (2015). Characterizing newly repopulated microglia in the adult mouse: Impacts on animal behavior, cell morphology, and neuroinflammation. PLoS One, 10(4), e0122912 10.1371/journal.pone.0122912 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elmore, M. R. , Najafi, A. R. , Koike, M. A. , Dagher, N. N. , Spangenberg, E. E. , Rice, R. A. , … Green, K. N. (2014). Colony‐stimulating factor 1 receptor signaling is necessary for microglia viability, unmasking a microglia progenitor cell in the adult brain. Neuron, 82(2), 380–397. 10.1016/j.neuron.2014.02.040 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Erny, D. , Hrabe de Angelis, A. L. , Jaitin, D. , Wieghofer, P. , Staszewski, O. , David, E. , … Prinz, M. (2015). Host microbiota constantly control maturation and function of microglia in the CNS. Nature Neuroscience, 18(7), 965–977. 10.1038/nn.4030 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eyo, U. B. , Murugan, M. , & Wu, L. J. (2017). Microglia‐neuron communication in epilepsy. Glia, 65(1), 5–18. 10.1002/glia.23006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eyo, U. B. , & Wu, L. J. (2013). Bidirectional microglia‐neuron communication in the healthy brain. Neural Plasticity, 2013, 456857–456810. 10.1155/2013/456857 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Feng, X. , Jopson, T. D. , Paladini, M. S. , Liu, S. , West, B. L. , Gupta, N. , & Rosi, S. (2016). Colony‐stimulating factor 1 receptor blockade prevents fractionated whole‐brain irradiation‐induced memory deficits. Journal of Neuroinflammation, 13(1), 215 10.1186/s12974-016-0671-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- Feng, X. , Valdearcos, M. , Uchida, Y. , Lutrin, D. , Maze, M. , & Koliwad, S. K. (2017). Microglia mediate postoperative hippocampal inflammation and cognitive decline in mice. JCI Insight, 2(7), e91229 10.1172/jci.insight.91229 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fourgeaud, L. , Traves, P. G. , Tufail, Y. , Leal‐Bailey, H. , Lew, E. D. , Burrola, P. G. , … Lemke, G. (2016). TAM receptors regulate multiple features of microglial physiology. Nature, 532(7598), 240–244. 10.1038/nature17630 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frost, J. L. , & Schafer, D. P. (2016). Microglia: Architects of the developing nervous system. Trends in Cell Biology, 26(8), 587–597. 10.1016/j.tcb.2016.02.006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fuger, P. , Hefendehl, J. K. , Veeraraghavalu, K. , Wendeln, A. C. , Schlosser, C. , Obermuller, U. , … Jucker, M. (2017). Microglia turnover with aging and in an Alzheimer's model via long‐term in vivo single‐cell imaging. Nature Neuroscience, 20(10), 1371–1376. 10.1038/nn.4631 [DOI] [PubMed] [Google Scholar]
- Geloso, M. C. , Corvino, V. , Marchese, E. , Serrano, A. , Michetti, F. , & D'Ambrosi, N. (2017). The dual role of microglia in ALS: Mechanisms and therapeutic approaches. Frontiers in Aging Neuroscience, 9, 242 10.3389/fnagi.2017.00242 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ghosh, S. , Castillo, E. , Frias, E. S. , & Swanson, R. A. (2017). Bioenergetic regulation of microglia. Glia, 66, 1200–1212. 10.1002/glia.23271 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ginhoux, F. , Greter, M. , Leboeuf, M. , Nandi, S. , See, P. , Gokhan, S. , … Merad, M. (2010). Fate mapping analysis reveals that adult microglia derive from primitive macrophages. Science, 330(6005), 841–845. 10.1126/science.1194637 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gong, Y. , Sasidharan, N. , Laheji, F. , Frosch, M. , Musolino, P. , Tanzi, R. , … Eichler, F. (2017). Microglial dysfunction as a key pathological change in adrenomyeloneuropathy. Annals of Neurology, 82(5), 813–827. 10.1002/ana.25085 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gowing, G. , Philips, T. , Van Wijmeersch, B. , Audet, J. N. , Dewil, M. , Van Den Bosch, L. , … Julien, J. P. (2008). Ablation of proliferating microglia does not affect motor neuron degeneration in amyotrophic lateral sclerosis caused by mutant superoxide dismutase. The Journal of Neuroscience, 28(41), 10234–10244. 10.1523/JNEUROSCI.3494-08.2008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grabert, K. , Michoel, T. , Karavolos, M. H. , Clohisey, S. , Baillie, J. K. , Stevens, M. P. , … McColl, B. W. (2016). Microglial brain region‐dependent diversity and selective regional sensitivities to aging. Nature Neuroscience, 19(3), 504–516. 10.1038/nn.4222 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Granqvist, M. , Boremalm, M. , Poorghobad, A. , Svenningsson, A. , Salzer, J. , Frisell, T. , & Piehl, F. (2018). Comparative effectiveness of rituximab and other initial treatment choices for multiple sclerosis. JAMA Neurology, 75, 320–327. 10.1001/jamaneurol.2017.4011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Greenfield, A. L. , & Hauser, S. L. (2017). B cell therapy for multiple sclerosis: Entering an era. Annals of Neurology, 83, 13–26. 10.1002/ana.25119 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guilliams, M. , & Scott, C. L. (2017). Does niche competition determine the origin of tissue‐resident macrophages? Nature Reviews. Immunology, 17(7), 451–460. 10.1038/nri.2017.42 [DOI] [PubMed] [Google Scholar]
- Guruswamy, R. , & ElAli, A. (2017). Complex roles of microglial cells in ischemic stroke pathobiology: New insights and future directions. International Journal of Molecular Sciences, 18(3), 496 10.3390/ijms18030496 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hagemeyer, N. , Hanft, K. M. , Akriditou, M. A. , Unger, N. , Park, E. S. , Stanley, E. R. , … Prinz, M. (2017). Microglia contribute to normal myelinogenesis and to oligodendrocyte progenitor maintenance during adulthood. Acta Neuropathologica, 134, 441–458. 10.1007/s00401-017-1747-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haimon, Z. , Volaski, A. , Orthgiess, J. , Boura‐Halfon, S. , Varol, D. , Shemer, A. , … Jung, S. (2018). Re‐evaluating microglia expression profiles using RiboTag and cell isolation strategies. Nature Immunology, 19(6), 636–644. 10.1038/s41590-018-0110-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hambardzumyan, D. , Gutmann, D. H. , & Kettenmann, H. (2016). The role of microglia and macrophages in glioma maintenance and progression. Nature Neuroscience, 19(1), 20–27. 10.1038/nn.4185 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Han, J. , Harris, R. A. , & Zhang, X. M. (2017). An updated assessment of microglia depletion: Current concepts and future directions. Molecular Brain, 10(1), 25 10.1186/s13041-017-0307-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Han, W. , Umekawa, T. , Zhou, K. , Zhang, X. M. , Ohshima, M. , Dominguez, C. A. , … Blomgren, K. (2016). Cranial irradiation induces transient microglia accumulation, followed by long‐lasting inflammation and loss of microglia. Oncotarget, 7(50), 82305–82323. 10.18632/oncotarget.12929 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hanamsagar, R. , & Bilbo, S. D. (2017). Environment matters: Microglia function and dysfunction in a changing world. Current Opinion in Neurobiology, 47, 146–155. 10.1016/j.conb.2017.10.007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hansen, D. V. , Hanson, J. E. , & Sheng, M. (2017). Microglia in Alzheimer's disease. The Journal of Cell Biology, 217, 459–472. 10.1083/jcb.201709069 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hauser, S. L. , Bar‐Or, A. , Comi, G. , Giovannoni, G. , Hartung, H. P. , Hemmer, B. , … Investigators, O. I. C. (2017). Ocrelizumab versus interferon Beta‐1a in relapsing multiple sclerosis. The New England Journal of Medicine, 376(3), 221–234. 10.1056/NEJMoa1601277 [DOI] [PubMed] [Google Scholar]
- Henriques, J. F. , Portugal, C. C. , Canedo, T. , Relvas, J. B. , Summavielle, T. , & Socodato, R. (2017). Microglia and alcohol meet at the crossroads: Microglia as critical modulators of alcohol neurotoxicity. Toxicology Letters, 283, 21–31. 10.1016/j.toxlet.2017.11.002 [DOI] [PubMed] [Google Scholar]
- Heppner, F. L. , Greter, M. , Marino, D. , Falsig, J. , Raivich, G. , Hovelmeyer, N. , … Aguzzi, A. (2005). Experimental autoimmune encephalomyelitis repressed by microglial paralysis. Nature Medicine, 11(2), 146–152. 10.1038/nm1177 [DOI] [PubMed] [Google Scholar]
- Hoek, R. M. , Ruuls, S. R. , Murphy, C. A. , Wright, G. J. , Goddard, R. , Zurawski, S. M. , … Sedgwick, J. D. (2000). Down‐regulation of the macrophage lineage through interaction with OX2 (CD200). Science, 290(5497), 1768–1771. [DOI] [PubMed] [Google Scholar]
- Huang, Y. , Xu, Z. , Xiong, S. , Qin, G. , Sun, F. , Yang, J. , … Peng, B. (2018). Dual extra‐retinal origins of microglia in the model of retinal microglia repopulation. Cell Discovery, 4, 9 10.1038/s41421-018-0011-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang, Y. , Xu, Z. , Xiong, S. , Sun, F. , Qin, G. , Hu, G. , … Peng, B. (2018). Repopulated microglia are solely derived from the proliferation of residual microglia after acute depletion. Nature Neuroscience, 21, 530–540. 10.1038/s41593-018-0090-8 [DOI] [PubMed] [Google Scholar]
- Inoue, K. , & Tsuda, M. (2018). Microglia in neuropathic pain: Cellular and molecular mechanisms and therapeutic potential. Nature Reviews. Neuroscience, 19(3), 138–152. 10.1038/nrn.2018.2 [DOI] [PubMed] [Google Scholar]
- Jakel, S. , & Dimou, L. (2017). Glial cells and their function in the adult brain: A journey through the history of their ablation. Frontiers in Cellular Neuroscience, 11, 24 10.3389/fncel.2017.00024 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Janova, H. , Arinrad, S. , Balmuth, E. , Mitjans, M. , Hertel, J. , Habes, M. , … Nave, K. A. (2017). Microglia ablation alleviates myelin‐associated catatonic signs in mice. The Journal of Clinical Investigation, 128, 734–745. 10.1172/JCI97032 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jin, W. N. , Shi, S. X. , Li, Z. , Li, M. , Wood, K. , Gonzales, R. J. , & Liu, Q. (2017). Depletion of microglia exacerbates postischemic inflammation and brain injury. Journal of Cerebral Blood Flow and Metabolism, 271678X17694185. doi: 10.1177/0271678X17694185, 37, 2224, 2236 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jin, X. , & Yamashita, T. (2016). Microglia in central nervous system repair after injury. Journal of Biochemistry, 159(5), 491–496. 10.1093/jb/mvw009 [DOI] [PubMed] [Google Scholar]
- Johnson, R. W. (2015). Feeding the beast: Can microglia in the senescent brain be regulated by diet? Brain, Behavior, and Immunity, 43, 1–8. 10.1016/j.bbi.2014.09.022 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kabba, J. A. , Xu, Y. , Christian, H. , Ruan, W. , Chenai, K. , Xiang, Y. , … Pang, T. (2017). Microglia: Housekeeper of the central nervous system. Cellular and Molecular Neurobiology, 38, 53–71. 10.1007/s10571-017-0504-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kanegawa, N. , Collste, K. , Forsberg, A. , Schain, M. , Arakawa, R. , Jucaite, A. , … Cervenka, S. (2016). In vivo evidence of a functional association between immune cells in blood and brain in healthy human subjects. Brain, Behavior, and Immunity, 54, 149–157. 10.1016/j.bbi.2016.01.019 [DOI] [PubMed] [Google Scholar]
- Kawanishi, S. , Takata, K. , Itezono, S. , Nagayama, H. , Konoya, S. , Chisaki, Y. , … Ashihara, E. (2018). Bone‐marrow‐derived microglia‐like cells ameliorate brain amyloid pathology and cognitive impairment in a mouse model of Alzheimer's disease. Journal of Alzheimer's Disease, 64(2), 563–585. 10.3233/JAD-170994 [DOI] [PubMed] [Google Scholar]
- Keren‐Shaul, H. , Spinrad, A. , Weiner, A. , Matcovitch‐Natan, O. , Dvir‐Szternfeld, R. , Ulland, T. K. , … Amit, I. (2017). A unique microglia type associated with restricting development of Alzheimer's disease. Cell, 169(7), 1276–1290 e1217. 10.1016/j.cell.2017.05.018 [DOI] [PubMed] [Google Scholar]
- Kierdorf, K. , & Prinz, M. (2017). Microglia in steady state. The Journal of Clinical Investigation, 127, 3201–3209. 10.1172/JCI90602 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kitayama, M. , Ueno, M. , Itakura, T. , & Yamashita, T. (2011). Activated microglia inhibit axonal growth through RGMa. PLoS One, 6(9), e25234 10.1371/journal.pone.0025234 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klein, D. , Patzko, A. , Schreiber, D. , van Hauwermeiren, A. , Baier, M. , Groh, J. , … Martini, R. (2015). Targeting the colony stimulating factor 1 receptor alleviates two forms of Charcot‐Marie‐tooth disease in mice. Brain, 138(Pt 11), 3193–3205. 10.1093/brain/awv240 [DOI] [PubMed] [Google Scholar]
- Konishi, H. , Kobayashi, M. , Kunisawa, T. , Imai, K. , Sayo, A. , Malissen, B. , … Kiyama, H. (2017). Siglec‐H is a microglia‐specific marker that discriminates microglia from CNS‐associated macrophages and CNS‐infiltrating monocytes. Glia, 65(12), 1927–1943. 10.1002/glia.23204 [DOI] [PubMed] [Google Scholar]
- Krasemann, S. , Madore, C. , Cialic, R. , Baufeld, C. , Calcagno, N. , El Fatimy, R. , … Butovsky, O. (2017). The TREM2‐APOE pathway drives the transcriptional phenotype of dysfunctional microglia in neurodegenerative diseases. Immunity, 47(3), 566–581 e569. 10.1016/j.immuni.2017.08.008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kronenberg, G. , Uhlemann, R. , Richter, N. , Klempin, F. , Wegner, S. , Staerck, L. , … Gertz, K. (2017). Distinguishing features of microglia‐ and monocyte‐derived macrophages after stroke. Acta Neuropathologica, 135, 551–568. 10.1007/s00401-017-1795-6 [DOI] [PubMed] [Google Scholar]
- Labzin, L. I. , Heneka, M. T. , & Latz, E. (2017). Innate immunity and neurodegeneration. Annual Review of Medicine, 69, 437–449. 10.1146/annurev-med-050715-104343 [DOI] [PubMed] [Google Scholar]
- Lassmann, H. , & Bradl, M. (2017). Multiple sclerosis: Experimental models and reality. Acta Neuropathologica, 133(2), 223–244. 10.1007/s00401-016-1631-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee, S. , Shi, X. Q. , Fan, A. , West, B. , & Zhang, J. (2018). Targeting macrophage and microglia activation with colony stimulating factor 1 receptor inhibitor is an effective strategy to treat injury‐triggered neuropathic pain. Molecular Pain, 14, 174480691876497 10.1177/1744806918764979 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leyns, C. E. G. , & Holtzman, D. M. (2017). Glial contributions to neurodegeneration in tauopathies. Molecular Neurodegeneration, 12(1), 50 10.1186/s13024-017-0192-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li, M. , Li, Z. , Ren, H. , Jin, W. N. , Wood, K. , Liu, Q. , … Shi, F. D. (2017). Colony stimulating factor 1 receptor inhibition eliminates microglia and attenuates brain injury after intracerebral hemorrhage. Journal of Cerebral Blood Flow and Metabolism, 37(7), 2383–2395. 10.1177/0271678X16666551 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li, Q. , & Barres, B. A. (2017). Microglia and macrophages in brain homeostasis and disease. Nature Reviews. Immunology, 18, 225–242. 10.1038/nri.2017.125 [DOI] [PubMed] [Google Scholar]
- Li, Y. , Zhang, R. , Hou, X. , Zhang, Y. , Ding, F. , Li, F. , … Wang, Y. (2017). Microglia activation triggers oligodendrocyte precursor cells apoptosis via HSP60. Molecular Medicine Reports, 16(1), 603–608. 10.3892/mmr.2017.6673 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liddelow, S. A. , Guttenplan, K. A. , Clarke, L. E. , Bennett, F. C. , Bohlen, C. J. , Schirmer, L. , … Barres, B. A. (2017). Neurotoxic reactive astrocytes are induced by activated microglia. Nature, 541(7638), 481–487. 10.1038/nature21029 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Limatola, C. , & Ransohoff, R. M. (2014). Modulating neurotoxicity through CX3CL1/CX3CR1 signaling. Frontiers in Cellular Neuroscience, 8, 229 10.3389/fncel.2014.00229 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu, J. , & Wang, F. (2017). Role of Neuroinflammation in amyotrophic lateral sclerosis: Cellular mechanisms and therapeutic implications. Frontiers in Immunology, 8, 1005 10.3389/fimmu.2017.01005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lund, H. , Pieber, M. , & Harris, R. A. (2017). Lessons learned about neurodegeneration from microglia and monocyte depletion studies. Frontiers in Aging Neuroscience, 9, 234 10.3389/fnagi.2017.00234 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lund, H. , Pieber, M. , Parsa, R. , Grommisch, D. , Ewing, E. , Kular, L. , … Harris, R. A. (2018). Fatal demyelinating disease is induced by monocyte‐derived macrophages in the absence of TGF‐beta signaling. Nature Immunology, 19(5), 1–7. 10.1038/s41590-018-0091-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luo, C. , Jian, C. , Liao, Y. , Huang, Q. , Wu, Y. , Liu, X. , … Wu, Y. (2017). The role of microglia in multiple sclerosis. Neuropsychiatric Disease and Treatment, 13, 1661–1667. 10.2147/NDT.S140634 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ma, S. F. , Chen, Y. J. , Zhang, J. X. , Shen, L. , Wang, R. , Zhou, J. S. , … Lu, H. Z. (2015). Adoptive transfer of M2 macrophages promotes locomotor recovery in adult rats after spinal cord injury. Brain, Behavior, and Immunity, 45, 157–170. 10.1016/j.bbi.2014.11.007 [DOI] [PubMed] [Google Scholar]
- Madore, C. , Baufeld, C. , & Butovsky, O. (2017). Microglial confetti party. Nature Neuroscience, 20(6), 762–763. 10.1038/nn.4570 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Madry, C. , Kyrargyri, V. , Arancibia‐Carcamo, I. L. , Jolivet, R. , Kohsaka, S. , Bryan, R. M. , & Attwell, D. (2017). Microglial ramification, surveillance, and interleukin‐1beta release are regulated by the two‐pore domain K(+) channel THIK‐1. Neuron, 97, 299–312.e6. 10.1016/j.neuron.2017.12.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marshall, G. P., 2nd , Deleyrolle, L. P. , Reynolds, B. A. , Steindler, D. A. , & Laywell, E. D. (2014). Microglia from neurogenic and non‐neurogenic regions display differential proliferative potential and neuroblast support. Frontiers in Cellular Neuroscience, 8, 180 10.3389/fncel.2014.00180 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martinez‐Muriana, A. , Mancuso, R. , Francos‐Quijorna, I. , Olmos‐Alonso, A. , Osta, R. , Perry, V. H. , … Lopez‐Vales, R. (2016). CSF1R blockade slows the progression of amyotrophic lateral sclerosis by reducing microgliosis and invasion of macrophages into peripheral nerves. Scientific Reports, 6, 25663 10.1038/srep25663 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mastroeni, D. , Nolz, J. , Sekar, S. , Delvaux, E. , Serrano, G. , Cuyugan, L. , … Coleman, P. D. (2017). Laser‐captured microglia in the Alzheimer's and Parkinson's brain reveal unique regional expression profiles and suggest a potential role for hepatitis B in the Alzheimer's brain. Neurobiology of Aging, 63, 12–21. 10.1016/j.neurobiolaging.2017.10.019 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Masuda, T. , & Prinz, M. (2016). Microglia: A unique versatile cell in the central nervous system. ACS Chemical Neuroscience, 7(4), 428–434. 10.1021/acschemneuro.5b00317 [DOI] [PubMed] [Google Scholar]
- Matcovitch‐Natan, O. , Winter, D. R. , Giladi, A. , Vargas Aguilar, S. , Spinrad, A. , Sarrazin, S. , … Amit, I. (2016). Microglia development follows a stepwise program to regulate brain homeostasis. Science, 353(6301), aad8670 10.1126/science.aad8670 [DOI] [PubMed] [Google Scholar]
- Mathys, H. , Adaikkan, C. , Gao, F. , Young, J. Z. , Manet, E. , Hemberg, M. , … Tsai, L. H. (2017). Temporal tracking of microglia activation in neurodegeneration at single‐cell resolution. Cell Reports, 21(2), 366–380. 10.1016/j.celrep.2017.09.039 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mecca, C. , Giambanco, I. , Donato, R. , & Arcuri, C. (2018). Microglia and aging: The role of the TREM2‐DAP12 and CX3CL1‐CX3CR1 axes. International Journal of Molecular Sciences, 19(1), 318 10.3390/ijms19010318 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mecha, M. , Carrillo‐Salinas, F. J. , Feliu, A. , Mestre, L. , & Guaza, C. (2016). Microglia activation states and cannabinoid system: Therapeutic implications. Pharmacology & Therapeutics, 166, 40–55. 10.1016/j.pharmthera.2016.06.011 [DOI] [PubMed] [Google Scholar]
- Menzel, F. , Kaiser, N. , Haehnel, S. , Rapp, F. , Patties, I. , Schoneberg, N. , … Bechmann, I. (2017). Impact of X‐irradiation on microglia. Glia, 66, 15–33. 10.1002/glia.23239 [DOI] [PubMed] [Google Scholar]
- Michell‐Robinson, M. A. , Touil, H. , Healy, L. M. , Owen, D. R. , Durafourt, B. A. , Bar‐Or, A. , … Moore, C. S. (2015). Roles of microglia in brain development, tissue maintenance and repair. Brain, 138(Pt 5), 1138–1159. 10.1093/brain/awv066 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mildner, A. , Schmidt, H. , Nitsche, M. , Merkler, D. , Hanisch, U. K. , Mack, M. , … Prinz, M. (2007). Microglia in the adult brain arise from Ly‐6ChiCCR2+ monocytes only under defined host conditions. Nature Neuroscience, 10(12), 1544–1553. 10.1038/nn2015 [DOI] [PubMed] [Google Scholar]
- Mills, C. D. , Harris, R. A. , & Ley, K. (2015). Macrophage polarization: Decisions that affect health. Journal of Clinical and Cellular Immunology, 6(5), 364 10.4172/2155-9899.1000364 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Molteni, M. , & Rossetti, C. (2017). Neurodegenerative diseases: The immunological perspective. Journal of Neuroimmunology, 313, 109–115. 10.1016/j.jneuroim.2017.11.002 [DOI] [PubMed] [Google Scholar]
- Mondelli, V. , Vernon, A. C. , Turkheimer, F. , Dazzan, P. , & Pariante, C. M. (2017). Brain microglia in psychiatric disorders. Lancet Psychiatry, 4(7), 563–572. 10.1016/S2215-0366(17)30101-3 [DOI] [PubMed] [Google Scholar]
- Mosser, C. A. , Baptista, S. , Arnoux, I. , & Audinat, E. (2017). Microglia in CNS development: Shaping the brain for the future. Progress in Neurobiology, 149‐150, 1–20. 10.1016/j.pneurobio.2017.01.002 [DOI] [PubMed] [Google Scholar]
- Muffat, J. , Li, Y. , Yuan, B. , Mitalipova, M. , Omer, A. , Corcoran, S. , … Jaenisch, R. (2016). Efficient derivation of microglia‐like cells from human pluripotent stem cells. Nature Medicine, 22(11), 1358–1367. 10.1038/nm.4189 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nadjar, A. , Wigren, H. M. , & Tremblay, M. E. (2017). Roles of microglial phagocytosis and inflammatory mediators in the pathophysiology of sleep disorders. Frontiers in Cellular Neuroscience, 11, 250 10.3389/fncel.2017.00250 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Natoli, R. , Fernando, N. , Madigan, M. , Chu‐Tan, J. A. , Valter, K. , Provis, J. , & Rutar, M. (2017). Microglia‐derived IL‐1beta promotes chemokine expression by Muller cells and RPE in focal retinal degeneration. Molecular Neurodegeneration, 12(1), 31 10.1186/s13024-017-0175-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nelson, L. H. , & Lenz, K. M. (2017). Microglia depletion in early life programs persistent changes in social, mood‐related, and locomotor behavior in male and female rats. Behavioural Brain Research, 316, 279–293. 10.1016/j.bbr.2016.09.006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nicolas, S. , Cazareth, J. , Zarif, H. , Guyon, A. , Heurteaux, C. , Chabry, J. , & Petit‐Paitel, A. (2017). Globular adiponectin limits microglia pro‐inflammatory phenotype through an AdipoR1/NF‐kappaB signaling pathway. Frontiers in Cellular Neuroscience, 11, 352 10.3389/fncel.2017.00352 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nimmerjahn, A. , Kirchhoff, F. , & Helmchen, F. (2005). Resting microglial cells are highly dynamic surveillants of brain parenchyma in vivo. Science, 308(5726), 1314–1318. 10.1126/science.1110647 [DOI] [PubMed] [Google Scholar]
- Nissen, J. C. , Thompson, K. K. , West, B. L. , & Tsirka, S. E. (2018). Csf1R inhibition attenuates experimental autoimmune encephalomyelitis and promotes recovery. Experimental Neurology, 307, 24–36. 10.1016/j.expneurol.2018.05.021 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Obst, J. , Simon, E. , Mancuso, R. , & Gomez‐Nicola, D. (2017). The role of microglia in prion diseases: A paradigm of functional diversity. Frontiers in Aging Neuroscience, 9, 207 10.3389/fnagi.2017.00207 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Olmos‐Alonso, A. , Schetters, S. T. , Sri, S. , Askew, K. , Mancuso, R. , Vargas‐Caballero, M. , … Gomez‐Nicola, D. (2016). Pharmacological targeting of CSF1R inhibits microglial proliferation and prevents the progression of Alzheimer's‐like pathology. Brain, 139(Pt 3), 891–907. 10.1093/brain/awv379 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pandya, H. , Shen, M. J. , Ichikawa, D. M. , Sedlock, A. B. , Choi, Y. , Johnson, K. R. , … Park, J. K. (2017). Differentiation of human and murine induced pluripotent stem cells to microglia‐like cells. Nature Neuroscience, 20(5), 753–759. 10.1038/nn.4534 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pang, Y. , Campbell, L. , Zheng, B. , Fan, L. , Cai, Z. , & Rhodes, P. (2010). Lipopolysaccharide‐activated microglia induce death of oligodendrocyte progenitor cells and impede their development. Neuroscience, 166(2), 464–475. 10.1016/j.neuroscience.2009.12.040 [DOI] [PubMed] [Google Scholar]
- Paolicelli, R. C. , Bolasco, G. , Pagani, F. , Maggi, L. , Scianni, M. , Panzanelli, P. , … Gross, C. T. (2011). Synaptic pruning by microglia is necessary for normal brain development. Science, 333(6048), 1456–1458. 10.1126/science.1202529 [DOI] [PubMed] [Google Scholar]
- Papageorgiou, I. E. , Lewen, A. , Galow, L. V. , Cesetti, T. , Scheffel, J. , Regen, T. , … Kann, O. (2016). TLR4‐activated microglia require IFN‐gamma to induce severe neuronal dysfunction and death in situ. Proceedings of the National Academy of Sciences of the United States of America, 113(1), 212–217. 10.1073/pnas.1513853113 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parkhurst, C. N. , Yang, G. , Ninan, I. , Savas, J. N. , Yates, J. R., 3rd , Lafaille, J. J. , … Gan, W. B. (2013). Microglia promote learning‐dependent synapse formation through brain‐derived neurotrophic factor. Cell, 155(7), 1596–1609. 10.1016/j.cell.2013.11.030 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parsa, R. , Andresen, P. , Gillett, A. , Mia, S. , Zhang, X. M. , Mayans, S. , … Harris, R. A. (2012). Adoptive transfer of immunomodulatory M2 macrophages prevents type 1 diabetes in NOD mice. Diabetes, 61(11), 2881–2892. 10.2337/db11-1635 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peng, J. , Gu, N. , Zhou, L. , U, B. E. , Murugan, M. , Gan, W. B. , & Wu, L. J. (2016). Microglia and monocytes synergistically promote the transition from acute to chronic pain after nerve injury. Nature Communications, 7, 12029 10.1038/ncomms12029 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Plescher, M. , Seifert, G. , Hansen, J. N. , Bedner, P. , Steinhauser, C. , & Halle, A. (2018). Plaque‐dependent morphological and electrophysiological heterogeneity of microglia in an Alzheimer's disease mouse model. Glia, 66, 1464–1480. 10.1002/glia.23318 [DOI] [PubMed] [Google Scholar]
- Pocock, J. M. , & Piers, T. M. (2018). Modelling microglial function with induced pluripotent stem cells: An update. Nature Reviews. Neuroscience, 19(8), 445–452. 10.1038/s41583-018-0030-3 [DOI] [PubMed] [Google Scholar]
- Prinz, M. , Erny, D. , & Hagemeyer, N. (2017). Ontogeny and homeostasis of CNS myeloid cells. Nature Immunology, 18(4), 385–392. 10.1038/ni.3703 [DOI] [PubMed] [Google Scholar]
- Prinz, M. , & Priller, J. (2014). Microglia and brain macrophages in the molecular age: From origin to neuropsychiatric disease. Nature Reviews. Neuroscience, 15(5), 300–312. 10.1038/nrn3722 [DOI] [PubMed] [Google Scholar]
- Rangaraju, S. , Dammer, E. B. , Raza, S. A. , Rathakrishnan, P. , Xiao, H. , Gao, T. , … Levey, A. I. (2018). Identification and therapeutic modulation of a pro‐inflammatory subset of disease‐associated‐microglia in Alzheimer's disease. Molecular Neurodegeneration, 13(1), 24 10.1186/s13024-018-0254-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ransohoff, R. M. (2016). A polarizing question: Do M1 and M2 microglia exist? Nature Neuroscience, 19(8), 987–991. 10.1038/nn.4338 [DOI] [PubMed] [Google Scholar]
- Reshef, R. , Kudryavitskaya, E. , Shani‐Narkiss, H. , Isaacson, B. , Rimmerman, N. , Mizrahi, A. , & Yirmiya, R. (2017). The role of microglia and their CX3CR1 signaling in adult neurogenesis in the olfactory bulb. eLife, 6, e30809 10.7554/eLife.30809 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reu, P. , Khosravi, A. , Bernard, S. , Mold, J. E. , Salehpour, M. , Alkass, K. , … Frisen, J. (2017). The lifespan and turnover of microglia in the human brain. Cell Reports, 20(4), 779–784. 10.1016/j.celrep.2017.07.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Riabov, V. , Salazar, F. , Htwe, S. S. , Gudima, A. , Schmuttermaier, C. , Barthes, J. , … Kzhyshkowska, J. (2017). Generation of anti‐inflammatory macrophages for implants and regenerative medicine using self‐standing release systems with a phenotype‐fixing cytokine cocktail formulation. Acta Biomaterialia, 53, 389–398. 10.1016/j.actbio.2017.01.071 [DOI] [PubMed] [Google Scholar]
- Rice, R. A. , Pham, J. , Lee, R. J. , Najafi, A. R. , West, B. L. , & Green, K. N. (2017). Microglial repopulation resolves inflammation and promotes brain recovery after injury. Glia, 65(6), 931–944. 10.1002/glia.23135 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rice, R. A. , Spangenberg, E. E. , Yamate‐Morgan, H. , Lee, R. J. , Arora, R. P. , Hernandez, M. X. , … Green, K. N. (2015). Elimination of microglia improves functional outcomes following extensive neuronal loss in the Hippocampus. The Journal of Neuroscience, 35(27), 9977–9989. 10.1523/JNEUROSCI.0336-15.2015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rodriguez, A. M. , Delpino, M. V. , Miraglia, M. C. , Costa Franco, M. M. , Barrionuevo, P. , Dennis, V. A. , … Giambartolomei, G. H. (2017). Brucella abortus‐activated microglia induce neuronal death through primary phagocytosis. Glia, 65(7), 1137–1151. 10.1002/glia.23149 [DOI] [PubMed] [Google Scholar]
- Sabatino, J. J., Jr. , Zamvil, S. S. , & Hauser, S. L. (2018). B‐cell therapies in multiple sclerosis. Cold Spring Harbor Perspectives in Medicine. 10.1101/cshperspect.a032037 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Salter, M. W. , & Stevens, B. (2017). Microglia emerge as central players in brain disease. Nature Medicine, 23(9), 1018–1027. 10.1038/nm.4397 [DOI] [PubMed] [Google Scholar]
- Salzer, J. , Svenningsson, R. , Alping, P. , Novakova, L. , Bjorck, A. , Fink, K. , … Svenningsson, A. (2016). Rituximab in multiple sclerosis: A retrospective observational study on safety and efficacy. Neurology, 87(20), 2074–2081. 10.1212/WNL.0000000000003331 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sarlus, H. , & Heneka, M. T. (2017). Microglia in Alzheimer's disease. The Journal of Clinical Investigation, 127(9), 3240–3249. 10.1172/JCI90606 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scherer, S. S. (2015). Blocking bad. Brain, 138(Pt 11), 3132–3133. 10.1093/brain/awv279 [DOI] [PubMed] [Google Scholar]
- Schiffer, D. , Mellai, M. , Bovio, E. , & Annovazzi, L. (2017). The neuropathological basis to the functional role of microglia/macrophages in gliomas. Neurological Sciences, 38(9), 1571–1577. 10.1007/s10072-017-3002-x [DOI] [PubMed] [Google Scholar]
- Scott, C. L. , Zheng, F. , De Baetselier, P. , Martens, L. , Saeys, Y. , De Prijck, S. , … Guilliams, M. (2016). Bone marrow‐derived monocytes give rise to self‐renewing and fully differentiated Kupffer cells. Nature Communications, 7, 10321 10.1038/ncomms10321 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sellgren, C. M. , Sheridan, S. D. , Gracias, J. , Xuan, D. , Fu, T. , & Perlis, R. H. (2017). Patient‐specific models of microglia‐mediated engulfment of synapses and neural progenitors. Molecular Psychiatry, 22, 170–177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Setiawan, E. , Attwells, S. , Wilson, A. A. , Mizrahi, R. , Rusjan, P. M. , Miler, L. , … Meyer, J. H. (2018). Association of translocator protein total distribution volume with duration of untreated major depressive disorder: A cross‐sectional study. Lancet Psychiatry, 5, 339–347. 10.1016/S2215-0366(18)30048-8 [DOI] [PubMed] [Google Scholar]
- Shen, Z. , Li, X. , Bao, X. , & Wang, R. (2017). Microglia‐targeted stem cell therapies for Alzheimer disease: A preclinical data review. Journal of Neuroscience Research, 95(12), 2420–2429. 10.1002/jnr.24066 [DOI] [PubMed] [Google Scholar]
- Shibata, M. , & Suzuki, N. (2017). Exploring the role of microglia in cortical spreading depression in neurological disease. Journal of Cerebral Blood Flow and Metabolism, 37(4), 1182–1191. 10.1177/0271678X17690537 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simard, A. R. , Soulet, D. , Gowing, G. , Julien, J. P. , & Rivest, S. (2006). Bone marrow‐derived microglia play a critical role in restricting senile plaque formation in Alzheimer's disease. Neuron, 49(4), 489–502. 10.1016/j.neuron.2006.01.022 [DOI] [PubMed] [Google Scholar]
- Singhal, G. , & Baune, B. T. (2017). Microglia: An Interface between the loss of neuroplasticity and depression. Frontiers in Cellular Neuroscience, 11, 270 10.3389/fncel.2017.00270 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith, J. A. , Das, A. , Ray, S. K. , & Banik, N. L. (2012). Role of pro‐inflammatory cytokines released from microglia in neurodegenerative diseases. Brain Research Bulletin, 87(1), 10–20. 10.1016/j.brainresbull.2011.10.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sominsky, L. , De Luca, S. , & Spencer, S. J. (2017). Microglia: Key players in neurodevelopment and neuronal plasticity. The International Journal of Biochemistry & Cell Biology, 94, 56–60. 10.1016/j.biocel.2017.11.012 [DOI] [PubMed] [Google Scholar]
- Sosna, J. , Philipp, S. , Albay, R., 3rd , Reyes‐Ruiz, J. M. , Baglietto‐Vargas, D. , LaFerla, F. M. , & Glabe, C. G. (2018). Early long‐term administration of the CSF1R inhibitor PLX3397 ablates microglia and reduces accumulation of intraneuronal amyloid, neuritic plaque deposition and pre‐fibrillar oligomers in 5XFAD mouse model of Alzheimer's disease. Molecular Neurodegeneration, 13(1), 11 10.1186/s13024-018-0244-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spangenberg, E. E. , Lee, R. J. , Najafi, A. R. , Rice, R. A. , Elmore, M. R. , Blurton‐Jones, M. , … Green, K. N. (2016). Eliminating microglia in Alzheimer's mice prevents neuronal loss without modulating amyloid‐beta pathology. Brain, 139(Pt 4), 1265–1281. 10.1093/brain/aww016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spelman, T. , Frisell, T. , Piehl, F. , & Hillert, J. (2017). Comparative effectiveness of rituximab relative to IFN‐beta or glatiramer acetate in relapsing‐remitting MS from the Swedish MS registry. Multiple Sclerosis, 1352458517713668. doi: 10.1177/1352458517713668, 24, 1087, 1095 [DOI] [PubMed] [Google Scholar]
- Spiller, K. J. , Restrepo, C. R. , Khan, T. , Dominique, M. A. , Fang, T. C. , Canter, R. G. , … Lee, V. M. (2018). Microglia‐mediated recovery from ALS‐relevant motor neuron degeneration in a mouse model of TDP‐43 proteinopathy. Nature Neuroscience, 21(3), 329–340. 10.1038/s41593-018-0083-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Srivastava, R. K. , Bulte, J. W. M. , Walczak, P. , & Janowski, M. (2017). Migratory potential of transplanted glial progenitors as critical factor for successful translation of glia replacement therapy: The gap between mice and men. Glia, 66, 907–919. 10.1002/glia.23275 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stevens, B. , Allen, N. J. , Vazquez, L. E. , Howell, G. R. , Christopherson, K. S. , Nouri, N. , … Barres, B. A. (2007). The classical complement cascade mediates CNS synapse elimination. Cell, 131(6), 1164–1178. 10.1016/j.cell.2007.10.036 [DOI] [PubMed] [Google Scholar]
- Subramaniam, S. R. , & Federoff, H. J. (2017). Targeting microglial activation states as a therapeutic avenue in Parkinson's disease. Frontiers in Aging Neuroscience, 9, 176 10.3389/fnagi.2017.00176 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Szalay, G. , Martinecz, B. , Lenart, N. , Kornyei, Z. , Orsolits, B. , Judak, L. , … Denes, A. (2016). Microglia protect against brain injury and their selective elimination dysregulates neuronal network activity after stroke. Nature Communications, 7, 11499 10.1038/ncomms11499 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takagi, S. , Furube, E. , Nakano, Y. , Morita, M. , & Miyata, S. (2017). Microglia are continuously activated in the circumventricular organs of mouse brain. Journal of Neuroimmunology. 10.1016/j.jneuroim.2017.10.008 [DOI] [PubMed] [Google Scholar]
- Tang, Y. , & Le, W. (2016). Differential roles of M1 and M2 microglia in neurodegenerative diseases. Molecular Neurobiology, 53(2), 1181–1194. 10.1007/s12035-014-9070-5 [DOI] [PubMed] [Google Scholar]
- Tay, T. L. , Mai, D. , Dautzenberg, J. , Fernandez‐Klett, F. , Lin, G. , Sagar, … Prinz, M. (2017). A new fate mapping system reveals context‐dependent random or clonal expansion of microglia. Nature Neuroscience, 20(6), 793–803. 10.1038/nn.4547 [DOI] [PubMed] [Google Scholar]
- Thion, M. S. , Low, D. , Silvin, A. , Chen, J. , Grisel, P. , Schulte‐Schrepping, J. , … Garel, S. (2017). Microbiome influences prenatal and adult microglia in a sex‐specific manner. Cell, 172, 500–516.e16. 10.1016/j.cell.2017.11.042 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thompson, K. K. , & Tsirka, S. E. (2017). The diverse roles of microglia in the neurodegenerative aspects of central nervous system (CNS) autoimmunity. International Journal of Molecular Sciences, 18(3), 504 10.3390/ijms18030504 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Um, J. W. (2017). Roles of glial cells in sculpting inhibitory synapses and neural circuits. Frontiers in Molecular Neuroscience, 10, 381 10.3389/fnmol.2017.00381 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vainchtein, I. D. , Chin, G. , Cho, F. S. , Kelley, K. W. , Miller, J. G. , Chien, E. C. , … Molofsky, A. V. (2018). Astrocyte‐derived interleukin‐33 promotes microglial synapse engulfment and neural circuit development. Science, 359, 1269–1273. 10.1126/science.aal3589 [DOI] [PMC free article] [PubMed] [Google Scholar]
- van de Laar, L. , Saelens, W. , De Prijck, S. , Martens, L. , Scott, C. L. , Van Isterdael, G. , … Guilliams, M. (2016). Yolk sac macrophages, fetal liver, and adult monocytes can colonize an empty niche and develop into functional tissue‐resident macrophages. Immunity, 44(4), 755–768. 10.1016/j.immuni.2016.02.017 [DOI] [PubMed] [Google Scholar]
- Varvel, N. H. , Grathwohl, S. A. , Baumann, F. , Liebig, C. , Bosch, A. , Brawek, B. , … Jucker, M. (2012). Microglial repopulation model reveals a robust homeostatic process for replacing CNS myeloid cells. Proceedings of the National Academy of Sciences of the United States of America, 109(44), 18150–18155. 10.1073/pnas.1210150109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Waisman, A. , Ginhoux, F. , Greter, M. , & Bruttger, J. (2015). Homeostasis of microglia in the adult brain: Review of novel microglia depletion systems. Trends in Immunology, 36(10), 625–636. 10.1016/j.it.2015.08.005 [DOI] [PubMed] [Google Scholar]
- Walter, T. J. , & Crews, F. T. (2017). Microglial depletion alters the brain neuroimmune response to acute binge ethanol withdrawal. Journal of Neuroinflammation, 14(1), 86 10.1186/s12974-017-0856-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang, Y. R. , Mao, X. F. , Wu, H. Y. , & Wang, Y. X. (2018). Liposome‐encapsulated clodronate specifically depletes spinal microglia and reduces initial neuropathic pain. Biochemical and Biophysical Research Communications, 499(3), 499–505. 10.1016/j.bbrc.2018.03.177 [DOI] [PubMed] [Google Scholar]
- Wendeln, A. C. , Degenhardt, K. , Kaurani, L. , Gertig, M. , Ulas, T. , Jain, G. , … Neher, J. J. (2018). Innate immune memory in the brain shapes neurological disease hallmarks. Nature, 556(7701), 332–338. 10.1038/s41586-018-0023-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wheeler, D. L. , Sariol, A. , Meyerholz, D. K. , & Perlman, S. (2018). Microglia are required for protection against lethal coronavirus encephalitis in mice. The Journal of Clinical Investigation, 128(3), 931–943. 10.1172/JCI97229 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wlodarczyk, A. , Cedile, O. , Jensen, K. N. , Jasson, A. , Mony, J. T. , Khorooshi, R. , & Owens, T. (2015). Pathologic and protective roles for microglial subsets and bone marrow‐ and blood‐derived myeloid cells in central nervous system inflammation. Frontiers in Immunology, 6, 463 10.3389/fimmu.2015.00463 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wlodarczyk, A. , Holtman, I. R. , Krueger, M. , Yogev, N. , Bruttger, J. , Khorooshi, R. , … Owens, T. (2017). A novel microglial subset plays a key role in myelinogenesis in developing brain. The EMBO Journal, 36, 3292–3308. 10.15252/embj.201696056 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wraith, D. C. (2017). The future of immunotherapy: A 20‐year perspective. Frontiers in Immunology, 8, 1668 10.3389/fimmu.2017.01668 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xie, X. , Luo, X. , Liu, N. , Li, X. , Lou, F. , Zheng, Y. , & Ren, Y. (2017). Monocytes, microglia, and CD200‐CD200R1 signaling are essential in the transmission of inflammation from the periphery to the central nervous system. Journal of Neurochemistry, 141(2), 222–235. 10.1111/jnc.13972 [DOI] [PubMed] [Google Scholar]
- Xue, J. , Schmidt, S. V. , Sander, J. , Draffehn, A. , Krebs, W. , Quester, I. , … Schultze, J. L. (2014). Transcriptome‐based network analysis reveals a spectrum model of human macrophage activation. Immunity, 40(2), 274–288. 10.1016/j.immuni.2014.01.006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang, H. M. , Yang, S. , Huang, S. S. , Tang, B. S. , & Guo, J. F. (2017). Microglial activation in the pathogenesis of Huntington's disease. Frontiers in Aging Neuroscience, 9, 193 10.3389/fnagi.2017.00193 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang, X. , Ren, H. , Wood, K. , Li, M. , Qiu, S. , Shi, F. D. , … Liu, Q. (2018). Depletion of microglia augments the dopaminergic neurotoxicity of MPTP. FASEB Journal, 32, 3336–3345. 10.1096/fj.201700833RR [DOI] [PMC free article] [PubMed] [Google Scholar]
- York, E. M. , Bernier, L. P. , & MacVicar, B. A. (2017). Microglial modulation of neuronal activity in the healthy brain. Developmental Neurobiology, 78, 593–603. 10.1002/dneu.22571 [DOI] [PubMed] [Google Scholar]
- Zhang, X. M. , Lund, H. , Mia, S. , Parsa, R. , & Harris, R. A. (2014). Adoptive transfer of cytokine‐induced immunomodulatory adult microglia attenuates experimental autoimmune encephalomyelitis in DBA/1 mice. Glia, 62(5), 804–817. 10.1002/glia.22643 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang, Y. , Zhao, L. , Wang, X. , Ma, W. , Lazere, A. , Qian, H. H. , … Wong, W. T. (2018). Repopulating retinal microglia restore endogenous organization and function under CX3CL1‐CX3CR1 regulation. Science Advances, 4(3), eaap8492 10.1126/sciadv.aap8492 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao, R. , Hu, W. , Tsai, J. , Li, W. , & Gan, W. B. (2017). Microglia limit the expansion of beta‐amyloid plaques in a mouse model of Alzheimer's disease. Molecular Neurodegeneration, 12(1), 47 10.1186/s13024-017-0188-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao, X. , Eyo, U. B. , Murugan, M. , & Wu, L. J. (2018). Microglial interactions with the neurovascular system in physiology and pathology. Developmental Neurobiology, 78, 604–617. 10.1002/dneu.22576 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao, X. , Liao, Y. , Morgan, S. , Mathur, R. , Feustel, P. , Mazurkiewicz, J. , … Huang, Y. (2018). Noninflammatory changes of microglia are sufficient to cause epilepsy. Cell Reports, 22(8), 2080–2093. 10.1016/j.celrep.2018.02.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zrzavy, T. , Hametner, S. , Wimmer, I. , Butovsky, O. , Weiner, H. L. , & Lassmann, H. (2017). Loss of 'homeostatic' microglia and patterns of their activation in active multiple sclerosis. Brain, 140(7), 1900–1913. 10.1093/brain/awx113 [DOI] [PMC free article] [PubMed] [Google Scholar]