

Abstract
Selenium (Se) is an essential element in poultry nutrition and its bio-efficacy depends on its chemical form. A growing body of research proves that organic forms of Se, mainly selenomethionine (SeMet), in poultry diets have a range of important advantages over traditional sodium selenite. In fact, the organic Se concept considers SeMet as a storage form of Se in the chicken body. As chickens are not able to synthesize SeMet, its provision through diet is a key strategy to fight commercially relevant stresses. Indeed, in stress conditions, when increased selenoprotein expression requires additional Se, while its provision via feed usually decreases due to a reduction in feed consumption, Se reserves in the body (mainly in the muscles) could help maintain an effective antioxidant defense and prevent detrimental consequences of stresses. The poultry industry is looking for the most effective sources of organic Se for commercial use. In this review, advantages and disadvantages of main organic Se sources for poultry (Se-yeast, SeMet, and OH-SeMet) are analyzed, and future directions for the development of new Se sources are identified.
Keywords: chicken, OH-SeMet, poultry, selenium, SeMet, Se-yeast
Introduction
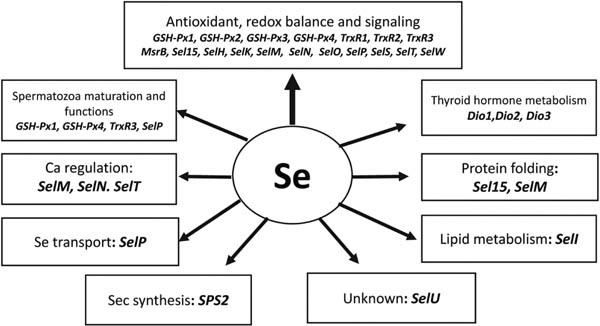
Selenium (Se) is an essential element for poultry and in farm animal nutrition that was discovered by the Swedish chemist Berzelius 200 years ago. Interest in this enigmatic element is growing since 1957, when its essentiality was described by Schwarz and Foltz (1957, 1958; Fig. 1). Severe Se deficiency has been shown to be associated with the development of various disorders that do no longer occur in modern commercial poultry industry. However, decreased productive and reproductive performance due to suboptimal levels of dietary Se and inadequate antioxidant defenses in stress conditions can still be observed in farms. The introduction of modern genetics to the poultry industry has substantially improved the growth rates of chickens. However, a major downside of such improvements in performance is that the birds are often highly sensitive to various stresses. Therefore, in modern poultry production, there is an important movement from prevention of Se deficiency to meeting the exact Se requirement of birds for optimizing their performance. In particular, the discovery and characterization of major selenoproteins, as well as a better understanding of the relationships between different antioxidants of the antioxidant system, provide new insights in this area. It is generally accepted that in biological systems, Se participates in various biochemical pathways and physiological functions as an integral part of a range of important selenoproteins. In chickens, 26 genes encoding different selenocysteine (SeCys) -containing proteins have been identified (Lei, 2017; Zhao et al., 2017). Interestingly, more than half of the known selenoproteins are directly or indirectly involved in antioxidant defenses and maintaining redox balance in the cell (Fig. 2). For example, selenoproteins are involved in glutathione-dependent hydroperoxide removal, reduction of thioredoxins, selenophosphate synthesis, activation and inactivation of thyroid hormones, thioredox-independent repair of oxidized methionine residues, endoplasmic reticulum-associated protein degradation, and other important biochemical processes. This explains the role of Se in animal health, including gut health and immune system regulation. Selenoprotein expression is characterised by high tissue specificity, depends on Se availability, can be regulated by hormones, and contributes to various pathological conditions if compromised (Surai, 2006).
Fig. 1.
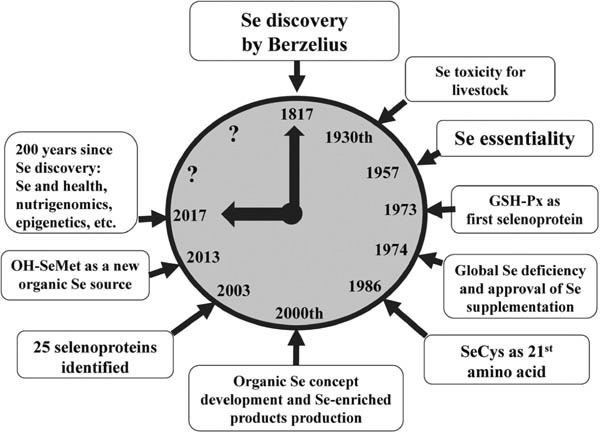
History of Se research and usage. Selenium was discovered by the Swedish chemist Berzelius in 1817. In 1930, Se toxicity for livestock was described (Surai, 2006). In 1957, Se essentiality was discovered by Schwarz and Foltz (1957, 1958). The first selenoprotein, GSH-Px was described by Rotruck et al. (1973). In 1970, a global Se deficiency in livestock was admitted and the FDA approved Se supplements for poultry and swine in 1974 in the form of selenite or selenate. In 1986, SeCys was identified as the 21st amino acid encoded by the stop codon TGA (Chambers et al., 1986). The organic Se concept was developed in 2000 and a range of Se-enriched products appeared on the market (Surai, 2006). In 2003, mammalian proteomes were characterised and 25 selenoproteins were identified (Kryukov et al., 2003). Later, 26 genes encoding different selenoproteins were identified (Lei, 2017; Zhao et al., 2017). Organic selenium sources found their way into animal/poultry nutrition, and a new effective source of organic Se (OH-SeMet) combining major advantages of Se-yeast and pure SeMet was successfully tested and found its way to poultry/animal industry (Briens et al., 2013, 2014; Jlali et al., 2013). Two hundred years have passed since the discovery of Se, and interest in the chemistry, biochemistry, and practical application of this element in poultry/animal industry increases steadily.
Fig. 2. Established and suggested selenoprotein functions.
During the last 40 years, a mandatory approach to regulate Se supplementation in commercial feed has been used in the poultry industry worldwide. To meet Se requirements, the poultry industry has relied completely on supplemental Se delivered with premixes (usually at 0.2–0.3 mg/kg diet), while the Se levels in the feed ingredients have not been taken into account (Surai, 2006; Surai and Fisinin, 2014).
Organic Se Concept Development
A growing number of studies over the last three decades have suggested that the dietary form of Se is a major determinant of its efficiency in meeting the Se requirement in poultry. There are two major Se sources for poultry: inorganic Se, mainly selenite or selenate, and organic Se, mainly in the form of selenomethionine (SeMet). In major feed ingredients, including grains, soya, and oilseed, Se is found exclusively in organic forms (with SeMet comprising more than 50% of total Se), while the major commercial Se supplements in use for the last 40 years are selenite and selenate (Surai, 2006). Studies have suggested that organic Se naturally occurs in the form of various seleno-amino acids in poultry diets and it seems likely that the digestive system adapted to these organic Se forms during evolution, thereby explaining principal differences in assimilation and metabolism between organic and inorganic forms of Se (Surai, 2006; Surai and Fisinin, 2014).
In 1974, the FDA approved Se supplements for poultry and swine in the form of selenite or selenate (Surai, 2006). Since then, a great body of evidence showing disadvantages of using inorganic Se in poultry nutrition has been accumulated.
Firstly, nutrients in the feed interact with each other, and sodium selenite can be reduced to the unavailable form (elemental Se) by various nutrients possessing reducing activity. For example, the chemical reaction between sodium selenite and ascorbic acid (AA) in premix, feed, or gut can lead to selenite reduction to elemental Se, which cannot be absorbed in the digestive tract of chickens, and the oxidation of AA, which thereby loses its biological activity (Robinson et al., 1985; Ip, 1986; Gosetti et al., 2007). In fact, pink particles in the premix after long storage very often represent elemental Se produced as a result of the aforementioned reaction. This was clearly demonstrated when solutions of selenite and AA were mixed in proportions similar to those in supplements: only about 50% selenite was recovered after 2 h, and selenite was not detectable after 24 h storage (Gosetti et al., 2007). A similar reaction was shown to occur in the chicken gut (Mykkänen and Mutanen, 1983). When premixes containing sodium selenite were prepared with glucose monohydrate, cornstarch, or sucrose and stored at room temperature, changes in odor and/or color were observed (Groce et al., 1973). The odor was musty and sweetish in character, while pink to dark-red particles appeared in the originally white matrix. Furthermore, when pigs were fed old premix, Se retention decreased, while Se excretion substantially increased. Decreased Se selenite availability due to its reduction has been also demonstrated in other studies. For example, in rats, the protective effect of selenite on tumorigenesis was prevented by vitamin C, due to the formation of unavailable elemental Se, while the chemopreventive action of SeMet was not affected (Ip, 1986). In a human trial, when selenite and AA were administered together, Se availability was dramatically reduced (Robinson et al., 1985). In contrast, it seems likely that AA can enhance SeMet assimilation from the diet (Mutanen and Mykkänen, 1985).
Secondly, sodium selenite can be lost as a vapor after its conversion to a volatile form. For example, it has been shown that selenite can be dissolved when dispersed in feeds with relatively high water activity. When dissolved, it may form selenious acid and disperse as a vapor (Eisenberg et al., 2007).
Thirdly, pro-oxidant effects of sodium selenite (Spallholz, 1997; Drake, 2006; Xiang et al., 2009; Brozmanová et al., 2010) can have detrimental consequences in the chicken gut. It has been recently shown that sodium selenite at 0.3 ppm can damage the gut structure in chickens. Chickens fed 0.3 ppm sodium selenite showed vacuolar degeneration in the epithelial cells lining of the intestinal crypts of the duodenum and excess mononuclear cell infiltration and aggregation in between degenerated and necrotic intestinal glands in the ileum (Attia et al., 2010). In great contrast to sodium selenite, SeMet has been suggested to possess antioxidant properties (Schrauzer, 2000; Suryo Rahmanto and Davies, 2011).
Fourthly, sodium selenite is poorly transferred to the egg and developing embryo (Surai, 2006) and thus has a limited ability to improve antioxidant defenses against hatching-imposed oxidative stress (Surai et al., 2016).
Finally, sodium selenite cannot be stored in Se reserves in the body, which are needed to maintain effective antioxidant defenses in stress conditions (Surai and Fisinin, 2014).
Thus, the use of sodium selenite in poultry diets has been questioned recently, and the organic Se concept has been developed and successfully introduced in the poultry industry (Surai, 2006; Fisinin et al., 2008; Surai and Fisinin, 2014). Compared with inorganic Se, organic sources of Se appear to better meet the needs of modern poultry. Main advantages of organic Se sources for poultry have been recently reviewed (Surai and Fisinin 2014). In particular, advantages for breeders include better transfer to the egg and developing embryo, which results in stronger antioxidant defenses and benefits in terms of hatchability and viability of newly hatched chicks (Fisinin et al., 2008; Surai and Fisinin, 2014). It has been suggested that Se delivered to the egg might have an epigenetic effect on the developing progeny, which awaits detailed investigation (Fisinin et al., 2016). Beneficial effects of organic Se in layers are associated with the higher efficacy of Se transfer to the egg (Jlali et al., 2013; Tufarelli et al., 2016), which has a positive effect on internal egg quality (Haugh units) via the activation of methionine sulfoxide reductase B (MsrB), a selenoprotein responsible for the prevention of protein oxidation and for maintaining the water-holding capacity of albumen. It has been suggested that the reversal methionine oxidation in proteins can alter their structure and functions, to regulate redox signaling (Kaya et al., 2015). Therefore, the reduction of oxidized methionine into an active form by MsrB is considered to be key to the prevention/repair of detrimental consequences of oxidative stress on protein structure and functions (Lee, 2016). Furthermore, Se has a positive effect on eggshell quality, including shell breaking strength, via the modulation of organic matrix formation (Surai, 2006). Advantages of organic Se in the broiler diet include increased Se transfer to the muscles and the build-up of Se reserves in the body, which is expected to result in improved chicken resistance to various stresses and to have a positive effect on immunity, gut health, and meat quality (Surai, 2006; Fisinin et al., 2008; Surai and Fisinin, 2014). In particular, it has been recently shown that Se deficiency in chickens promotes gut inflammation (Gao et al., 2016; Wu et al., 2016). Furthermore, specific emphasis has been placed on protein oxidation in relation to meat quality, including water-holding capacity of meat and drip loss. Indeed, increased activity of MsrB in the muscles as a result of improved Se status owing to the usage of hydroxyselenomethionine (OH-SeMet) in the diet led to a decrease in protein oxidation (Zhao et al., 2017) and could reduce drip loss. Se has been suggested to have immunomodulatory properties as well, which remain to be explored (Surai, 2006).
There are two important questions to be answered. First, what is an optimal Se concentration in the chicken egg to provide maximal protection against oxidative stress caused by the process of hatching? Second, how to achieve that concentration in commercial poultry production? The first question might be answered by analyzing the Se concentration in egg yolks from wild birds having a free choice of food. Se concentrations of 100–200 ng/g (Pappas et al., 2005) or 300 ng/g wet yolk (Surai, 2000) have been found in egg yolk of chickens maintained on a standard diet (without supplementary Se and containing approximately 0.1 or 0.17 ppm Se respectively; Pappas et al., 2005; Surai, 2000). In this case, Se is derived from the basic dietary constituents including wheat, maize, and soybean meal, and the levels in the body are quite variable depending on dietary composition and the place of origin of the feed ingredients. In our study, the level of Se in avian yolks was found to differ widely among free-living species, and yolk Se concentrations were substantially higher in eggs of wild species than in those of domestic chickens (Pappas et al., 2006a). However, after organic Se supplementation at 0.4–0.5ppm (Surai, 2000; Pappas et al., 2005), Se levels in the chicken egg yolk were quite close to the levels found in wild birds, suggesting that there is scope for breeders to increase Se supplementation. Additional evidence of high Se concentrations in eggs from wild birds was found in little egrets, black-crowned night herons, and bridled terns from coastal areas of Hong Kong (Lam et al., 2005). Furthermore, comparatively high Se levels were found in eggs from tree swallow, bank swallow, and house wren (Dickerson et al., 2002). Similarly, in tissues of birds from various areas, including the Barents Sea, Alaska (Savinov et al., 2003), and arctic Russia (Stout et al., 2002), as well as in bald eagles from Adak Island, Alaska (Stout and Trust, 2002), Se concentrations were shown to be several fold higher than those found in domestic chickens. It is clear that effective Se dietary supplements are needed to meet the Se requirement of modern breeds of egg and meat types of poultry.
Accordingly, a range of supplements claimed to be organic Se sources can be found on the world market (Table 1). However, strictly speaking, only those products providing SeMet or its precursor could be considered as sources of organic Se, as SeMet is the only form that allows building Se reserves in the body (mainly in the muscles) (Surai, 2006). As avian species as well as other animals cannot synthesize SeMet, it has to be provided through diet. True organic Se sources include Se-enriched yeast (Se-yeast), SeMet, Zn-SeMet, and OH-SeMet, while Se chelates, glycinates, and proteinates should not be included into this category of supplements.
Table 1. Sources of organic Se available on the market.
| Source | Comments | SeMet* | References |
|---|---|---|---|
| Se-yeast | Well researched, 50–70% SeMet | Yes | Schrauzer, 2006 |
| Se-Met | Not stable, >95% SeMet | Yes | Liu et al., 2017 |
| Zn-SeMet | Not stable, >95% SeMet | Yes | Geraert et al., 2015 |
| OH-SeMet | Stable, >95% OH-SeMet | Yes | Liu et al., 2017 |
| Se-proteinates | Chemistry is not proven | No | Liu et al., 2017 |
| Se-glycinates | Chemistry is not proven | No | Liu et al., 2017 |
| Se-chelates | Chemistry is not proven | No | Kubachka et al., 2017 |
| Se-homolanthionine | Lack of research data | No | Anan et al., 2011 |
| Other (nano-Se, etc.) | Lack of data on molecular mechanisms of action | No | Pelyhe and Mezes, 2013; Sarkar et al., 2015 |
As only SeMet can be non-specifically incorporated into proteins in measurable amounts, it is an active component of organic Se sources.
Of interest, Se supplements available for humans include mainly sodium selenite, Se-yeast, and pure l-SeMet (Surai, 2006). Similar to animal studies, studies in humans have indicated that SeMet is the major metabolizable form of Se for humans (Schrauzer, 2000, 2001, 2003; Schrauzer and Surai, 2009). Sodium selenite, Se-yeast, and SeMet have shown cancer-preventive effects in various clinical trials (Surai, 2006). However, the most comprehensive clinical trial on cancer prevention by using Se (SELECT), where Se was supplemented in the form of l-SeMet (200 µg/day), did not show any benefit and was terminated prematurely (Nicastro and Dunn, 2013). The main lesson from the SELECT trial is that there is a need for further research to better understand the molecular mechanisms of Se action, elucidate an optimal dosage of Se supplementation, and determine the most effective forms of Se for dietary supplementation.
Building Se Reserves in the Body
After matrix digestion, similar to Met, ingested SeMet released from selenium-enriched yeast or pure SeMet is absorbed in the small intestine via a single, Na+-dependent, carrier-mediated process (Wolffram et al., 1989). Part of the absorbed SeMet is trans-selenated to SeCys, which is converted to hydrogen selenide (H2Se) by β-lyase, and H2Se is further converted to selenophosphate (HSePO32−) by selenophosphate synthetase (SPS). SPS provides the active Se donor in the form of selenophosphate for the synthesis of SeCys, the 21st amino acid in the genetic code. The terminal reaction of SeCys synthesis is catalyzed by SeCys synthase when serine or phosphoserine is converted into SeCys (Dobosz-Bartoszek and Simonovic, 2016). SeCys synthesis involves complex cellular machinery, including specialized tRNA. In fact, selenophosphate first reacts with tRNA-bound serinyl residues to form SeCys -bound tRNA, and the incorporation of SeCys into selenoproteins occurs via UGA codon recognition (Gladishev et al., 2016). In contrast to the other amino acids, no free pool of SeCys exists in the cell and only newly synthesized SeCys is incorporated into selenoproteins (Surai, 2006),
As mentioned above, humans and higher animals, including poultry, cannot synthesize SeMet, and it is the only seleno-amino acid that can be significantly stored in organs and tissues, building Se reserves in the body (Schrauzer, 2003, 2006). In contrast, there is no evidence of non-specific incorporation of Se into plasma proteins when administered as selenate or as SeCys (Surai, 2006). A growing body of evidence shows that SeMet is metabolized as a constituent of the methionine pool. In fact, in humans, approximately 46.9% of the total Se in the body is located in skeletal muscles, while only 4% of body Se is found in the kidneys (Oster et al., 1988). In chicken breast and leg muscles, SeMet is the major Se form, comprising more than half (66.7% and 56.1%) of total Se (Bierla et al., 2008). The authors also showed that in chickens fed a diet containing high doses of organic Se, the SeMet fraction in the muscles increased up to 99%, which confirmed non-specific incorporation of SeMet into muscle proteins. In muscles, the content of Met is always in large excess over that of SeMet. For example, in humans, the SeMet/Met ratio in skeletal muscle is approximately 1:7,000, which is similar to the ratio of 1: >6,000 found in chicken breast muscle (Schrauzer and Surai, 2009). We calculated that in the egg yolk, the SeMet/Met ratio is approximately 1:160,000, and in egg white, it is close to 1:87,000. When organic Se is included in the breeder diet, this ratio can change substantially. It is interesting to note that SeMet comprises 53–71% of total Se in the egg albumen, while its proportion in the egg yolk (12–19%) is much lower (Lipiec et al., 2010). This explains why albumin response to dietary organic Se is substantially stronger than that in the egg yolk (Surai, 2006). SeMet reportedly has a lower turnover rate in the body than sodium selenite and is characterised by greater Se re-utilization efficiency (Swanson et al., 1991). The confirmation of chicken muscle being a storage site for Se came from a recent study by Brandt-Kjelsen et al. (2014) who showed that organic selenium has the longest half-life in muscles (12 days) and in the brain and lungs (13 days), while the shortest half-lives were found in the liver, kidneys, and pancreas (about 4 days). In comparison, the average half-lives of SeMet and selenite in the human body are 252 and 102 days, respectively (Patterson et al., 1989). It was proven long ago that Se from both selenite and SeMet is readily available for the synthesis of the selenoenzyme glutathione peroxidase (GSH-Px) in rat tissues (Pierce and Tappel, 1977). Furthermore, it has been shown recently in vitro and in vivo that the Se availability from SeMet and sodium selenite is similar, while SeMet was more efficiently transported than sodium selenite in an in vitro membrane permeability study using Caco-2 cells (Takahashi et al., 2017).
The idea that Se stored in tissues in the form of SeMet is available for selenoprotein synthesis has been proven in a range of studies (for review see Surai, 2006). In particular, chicks hatched from eggs enriched with Se were characterised by increased liver GSH-Px activity not only at hatching, but also at 5 days posthatch (Surai, 2000). Similarly, in quail (Surai et al., 2006) and chickens (Pappas et al., 2005), the Se concentration in the livers of the progeny was alleviated up to 2–4 weeks posthatch owing to organic Se in the breeder diet and increased Se levels in the egg. In addition, GSH-Px activity was increased in chicken muscle several weeks posthatch owing to maternal organic Se supplementation (Pappas et al., 2005). In an elegant study with chickens, it was proven that endogenous Se accumulated in tissues through organic dietary Se supplementation was available for maintaining the GSH-Px activity when Se supplementation ceased (Payne and Southern, 2005). Similarly, in SeMet or Se-yeast supplemented mice, GSH-Px activities in the liver decreased slower during Se depletion than in mice given selenite (Spallholz and Rafferty, 1987). The half-life of GSH-Px was calculated to be 4.2 and 9.1 days in rats that received 3 ppm Se as selenite or SeMet, respectively (Ip and Hayes, 1989). In a human study, once supplements were withdrawn, GSH-Px activity in platelets (Levander et al., 1983) or selenoprotein P concentration (Persson-Maschos et al., 1998) decreased slower in the group given wheat Se than in the selenite-supplemented group. In New Zealanders who consumed high-Se bread, plasma Se remained elevated when Se supplementation ceased (Robinson et al., 1985). Furthermore, Se-yeast was shown to provide a long-lasting body pool of Se in children (Alfthan et al., 2000).
It seems likely that the protective effect of organic Se is most pronounced in stress conditions. Indeed, our previous study (Pappas et al., 2006b) showed that mortality of chick embryos in week 3 of egg incubation was 3.5% for the control group and 10.6% in fish-oil supplemented (stressed) 27-week-old breeders. Inclusion of Se-yeast (0.4 ppm) in the breeder diets decreased mortality to 3.0% and 6.2%, respectively. In addition, stress associated with fish oil supplementation in the breeder diet reduced both hatchability and 1-day-old chick weight (Pappas et al., 2006b). Again, the inclusion of organic Se into the breeder diet prevented some of these adverse effects. Therefore, Se reserves in the chicken body (mainly in the muscles) built as a result of non-specific SeMet incorporation into the proteins in place of Met are considered to be an important adaptation of chickens to various stresses. This strategy might prevent decreases in productive and reproductive performance in stressful commercial conditions of chicken production (Surai and Fisinin, 2016a, b). In general, the metabolic pathway of Se in birds (quails) is the same as that in rodents, but the metabolic capacity for Se seems to be larger in quails than in rodents. Indeed, the concentrations of exogenous Se in all organs and tissues of SeMet-administered quails were significantly higher than those in selenite-administered quails 3 h after the administration, suggesting that SeMet is more rapidly and/or efficiently incorporated into the quail body than selenite (Anan et al., 2014). It should be noted that only a small portion of the methionine pool can be replaced with SeMet, and protein turnover prevents accumulation of SeMet to toxic levels in the organism (Schrauzer, 2003). A model of organic Se action in poultry is shown in Fig. 3.
Fig. 3.
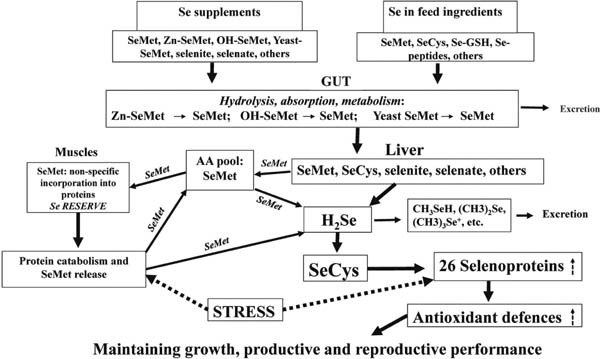
Organic Se in action. A range of Se-containing compounds, including selenite, selenate, SeMet, Zn-SeMet, OH-SeMet, Se-yeast, SeCys, Se-GSH, and Se-peptides, can be included into premixes. All those Se forms come to the intestine where initial hydrolysis (Se-Met will be released from Se-yeast or Zn-SeMet; OH-SeMet will be converted into SeMet) and some metabolic changes will take place. This includes excretion of Se metabolites via bile, feces, and urine. Further, selenite, selenate, SeMet and some other Se forms will be delivered to the liver for metabolization and distribution. In parallel, a fraction of SeMet will go to the free amino acid pool and be used for building Se reserves mainly in muscles. The next step of Se assimilation and metabolism includes the conversion of all major forms of Se into H2Se, from which SeCys will be synthesized and incorporated into 26 newly synthesized selenoproteins that are integral part of the antioxidant system of the body. Under stress conditions, protein catabolism will take place, which will release some SeMet incorporated into those proteins, and this SeMet will be converted into H2Se and further into newly synthesized SeCys and 26 selenoproteins. Additional sources of Se will be responsible for the upregulation of selenoprotein genes and additional synthesis of selenoproteins, which will upregulate antioxidant defenses and will aid the body to adapt to and overcome the stress with minimal negative consequences. When only selenite is present in the diet, Se reserves in the muscles will not be built and therefore, the ability of the body to adapt to stress will be restricted.
The mechanisms regulating SeMet conversion to H2Se and further to SeCys and respective selenoproteins are not clear at present, but it seems likely that changes in redox status of the cells/tissues and activation of proteasomal protein degradation are involved. Indeed, ATP- and ubiquitin-independent proteolysis by the 20S proteasome is responsible for the selective degradation of oxidized proteins. In vitro, the 20S proteasome shows increased proteolytic activity toward oxidized polypeptides. In cells overexpressing GSH-Px-1, chymotrypsin-like activity of the proteasome was decreased by 30% (Kretz-Remy and Arrigo, 2003). This observation was correlated with a 2-fold increase in the half-life of IκB-α, a protein whose basal turnover is 20S proteasome-dependent. Furthermore, following exposure to H2O2, human T47D cells overexpressing GSH-Px showed Se-dependent decreases in intracellular ROS accumulation and 20S proteasome chymotrypsin-like activity. Moreover, exposure of HeLa cells to antioxidant compounds reduced the proteasome 20S chymotrypsin-like activity. These results suggest that GSH-Px activity or pro-reducing conditions can downregulate basal 20S proteasome activity (Kretz-Remy and Arrigo, 2003). This implies that selenium is a key element in proteasome activity control. There might be a feedback mechanism of recognition of SeMet as a source of Se for selenoprotein synthesis. In stress conditions, some amino acids inside muscle proteins can be oxidized (Davies, 2016), and this will trigger an increase in proteasome activity to degrade such proteins (Pajares et al., 2015) and release SeMet to be available as an additional source of Se for selenoprotein synthesis. When the antioxidant/pro-oxidant equilibrium is restored, increased GSH-Px activity would decrease proteasome activity and protein degradation.
Se-yeast: Advantages and Disadvantages
It is generally accepted that chemical and physical properties of Se and sulphur are very similar. Enzymatic systems in plants are not able to distinguish between these two elements; SeMet instead of Met was synthesized when sulphur in the incubation medium was replaced with Se (Surai, 2006). The commercial technology of production of Se-yeast is based on the aforementioned features. Saccharomyces cerevisiae is the main yeast strain used for aerobic fermentation in a Se-enriched medium containing beet or cane molasses, vitamins, nutritional salts, and sodium selenite as the Se source to produce Se-yeast (Esmaeili and Khosravi-Darani, 2014). An important limitation to Seyeast production is the toxicity of sodium selenite to yeast cells, which results in dramatic reductions in yeast growth. Recent results obtained by scanning electron microscopy confirmed a damaging effect of increasing concentrations of sodium selenite to yeast cells (Rajashree and Muthukumar, 2013). Therefore, industrial Se-yeast production requires a long selection process for Se-tolerant yeast strains. Theoretically, Se-yeast can accumulate up to 6,000 ppm Se; however, full replacement of Met in the yeast cells is not possible and therefore, Se concentrations in current commercial products vary between 1,000 and 3,000 ppm (Schrauzer, 2006). In many cases, industrial Se-yeast production technologies are patented and differ between producers, which represents an additional source of variation in the composition of Se-yeast supplements found on the world market.
After harvest, Se-yeast cream is usually pasteurized and then dried. Final product characteristics include water content (5–7%), protein (40–50%), carbohydrate (11–48%), fat (2–8%), and residual ash (5–10%) (EFSA, 2008). The technology of Se-yeast manufacturing was developed more than four decades ago (for review, see Schrauzer, 2006) and today, Se-yeast is produced by a number of companies worldwide and widely used in poultry production (for review see Surai 2006; Surai and Fisinin 2014). As mentioned above, SeMet is an active component of Se-yeast, but its proportion in Se-yeast is not shown on the label, as only total Se and the inorganic proportion in Se-yeast are officially regulated.
Consequently, there are several points that remain to be addressed for commercial usage of Se-yeast. Firstly, based on modern Se-yeast production technology, it is not possible to guarantee an exact percentage of SeMet in the final product, as its composition depends on fermentation conditions, including yeast strain, source of inorganic Se, Se concentration, protocol of Se addition, base medium, energy sources (molasses), pH, temperature, shaking speed, aeration, inoculum size, and incubation time (Esmaeili and Khosravi-Darani, 2014). Indeed, the Se-metabolite profile of Se-yeast is influenced by cultivation conditions (Rao et al., 2010). For example, it has been recently shown that Se toxicity and Se-metabolite profile in Se-yeast are significantly affected by the amount of sulphur provided during the fermentation (Mapelli et al., 2012). Secondly, analytical limitations to SeMet determination in Se-yeast further complicate this issue (Kubachka et al., 2017). Furthermore, there are principal differences in composition and probably in assimilation efficacy of yeasts produced by different technologies/companies. Indeed, the fraction of water-soluble Se in various yeasts varies from 11.5% up to 28.0% (Encinar et al., 2003). It has been confirmed that significant differences exist between Se-yeast products in terms of Se deposition in different cell fractions and Se bioavailability (Fagan et al., 2015).
Se-yeast has been reported to contain more than 60 (Arnaudguilhem et al., 2012) or even 100 (Gilbert-Lopez et al., 2017) unique Se species, but only SeMet has been well researched and has been proven to be an active compound of Se-yeast. Possible roles and effects of these other Se compounds in animals/poultry await further investigation, but based on current knowledge, it can be concluded that their bio-potency and efficacy in most cases are not different from those of sodium selenite. Variability in SeMet proportion in Se-yeast is the major concern for poultry nutritionists. In fact, it would best to balance Se in the poultry diet not based on total Se, but on SeMet, which is impossible when using Se-yeast because of the aforementioned limitations in Seyeast production and analysis. For example, Rayman (2004) reported percentages of SeMet of 84, 69, 75, 81, 83, 61, and 60 in in seven different supplements. The author concluded that, despite the fact that in commercial Se-yeast products found on the market nearly all Se is in the organic form, only 55–75% is SeMet. Indeed, great variability in SeMet content of Se-yeast products has been reported: 54–60% (Larsen et al., 2004); 60–61% (Kotrebai et al., 2000), 63% (EFSA, 2016), 63. 4–66. 6% (Burdock and Cousins, 2010), 64% (McSheehy et al., 2005), 65% (Wrobel et al., 2003), 73% (Wolf et al., 2001), 74. 8% (Yoshida et al., 2002), 84% (Uden et al., 2004), 85% (Ip et al., 2000; Fan et al., 2003). In general, the criterion for commercial Se-yeast is > 60% SeMet (Bierla et al., 2012). When Se-yeast samples of five leading manufacturers worldwide were analyzed (Casal et al., 2010), the proportion of water-extractable Se varied between 16–35%, and the SeMet faction also greatly varied. Recently, considerable incorporation of SeCys in proteins of the yeast proteome, despite the absence of the UGA codon, was demonstrated (Bierla et al., 2013). The authors showed that 10–15% of Se present in Se-yeast is SeCys. This means that if all the Se in Se-yeast is accounted for, the maximum SeMet fraction would not exceed 85%, but in many cases will be much lower than that. In accordance herewith, in a batch production analysis, SeMet in the product varied between 58–69%, and SeCys accounted for 15–22% (EFSA, 2016). However, dietary SeCys, similar to sodium selenite, is not effective in increasing the tissue Se concentration and building Se reserves in monogastric animals as shown in rats by Deagen et al. (1987). Using ICP-MS, the UT2A Lab in France analyzed 12 Se-yeast products of eight commercial brands, collected from end users in the Asia-Pacific region. The results revealed that Se-yeast products largely vary in terms of total Se and SeMet contents. For a number of samples, SeMet accounted for approximately 60% of total Se. However, certain products showed extremely high variation in or even lacked SeMet, being a simple mixture of yeast and mineral selenium (Liu et al., 2017). Recently, a range of commercially available Se supplements was evaluated, and discrepancies between labeled ingredients and detected species were noted (Kubachka et al., 2017).
It should be noted that absorption of dietary Se (organic Se) is generally believed to be good (∼80%; Reilly, 2006). Absorption and retention of Se from Se-yeast, measured in 12 volunteers fed 77 Se-labeled Se-yeast was shown to be 75–90%; however, other Se-yeasts gave different results (50–60%) (Sloth et al., 2003). Intestinal absorption of SeMet in women was shown to be 95.7–97.3% of the administered dose (Griffiths et al., 1976). The fact that Se-yeast efficacy is determined by the fraction of SeMet was proven in a study in broilers using two commercially available Se-yeast preparations, containing SeMet at 63% and 56.7% (Simon et al., 2013). Se content in the muscles of broilers fed a diet supplemented with sodium selenite was 133 µg/kg, while that in broilers fed a diet supplemented with Se-yeast containing 26% or 69% SeMet was 161 and 267 µg/kg Se, respectively, indicating that high SeMet content results in higher Se deposition in muscle tissues (Van Beirendonck et al., 2016a, b). Similar results have been reported by Liu et al. (2017), who compared two Se-yeast products with significantly different SeMet content (74% vs. 33%). In brief, Se accumulation in the muscles is proportional to the level of SeMet consumption through diet.
SeMet and Zn-SeMet
SeMet exists in various forms, including the naturally occurring l-SeMet and two synthetic forms, d-SeMet and dl-SeMet (Schrauzer, 2003). Pure SeMet can also be used as a dietary supplement (Schrauzer 2000, 2001, 2003; Schrauzer and Surai, 2009). There is a range of publications showing the beneficial effects of SeMet in poultry diets. For example, dietary l-SeMet is more effectively transferred to the egg than Se-yeast (Delezie et al., 2014). It seems likely that both l-SeMet and d-SeMet can be used as Se sources for poultry with better efficacy than sodium selenite (Wang et al., 2011). However, l-SeMet is more effective in improving antioxidant defenses in chickens as well as average daily gain and feed conversion ratio (FCR) than d-SeMet. On the other hand, a study analyzing major indexes of growth, development, and antioxidant defense system in plasma and breast muscles in broilers supplemented with either sodium selenite or dl-SeMet at 0.15mg/kg showed that dl-SeMet was not different from sodium selenite in terms of daily gain, FCR, total antioxidant activity, GSH-Px activity (Jiang et al., 2009). Only total superoxide dismutase activity and GSH levels in breast muscle were significantly upregulated upon organic Se supplementation (Jiang et al., 2009). dl-SeMet was more effective than sodium selenite in improving chicken immunity (Wang et al., 2016). More recently, DLSeMet in the chicken diet was shown to more strongly increase GSH-Px and total antioxidant activity in thigh muscle and liver than sodium selenite (Bakhshalinejad et al., 2017). There is a need for more studies comparing the efficiencies of l-, d-, dl-SeMet and other forms of organic Se in poultry diets.
Unfortunately, scientific information on Zn-SeMet is quite limited and this product is not registered for use in the EU, although it is available on other markets, including Asian countries. This product is sometimes called “chelated selenium,” but in fact, the Zn is chelated by SeMet. Based on the chemical structure of the product, it seems likely that it does not differ from pure SeMet in terms of stability, though its absorption efficiency might be lower (Geraert et al., 2015). Inclusion of Zn-SeMet into the laying hen diet at 0.2 and 0,4 mg/kg was shown to have positive effects on egg production and quality in comparison to non-supplemented layers, and GSH-Px activity in layer plasma was increased only at 0.2 ppm Zn-SeMet supplementation (Laika and Jahanian, 2015). When Zn-SeMet was compared to sodium selenite at 0.4 ppm supplementation in breeder diet, the reproductive performance of birds in both groups was the same. The only significant difference induced by Zn-SeMet was a heavier weight from hatchling until egg production peak (33 weeks) (Urso et al., 2015). Similarly, Se source (sodium selenite or Zn-SeMet at 0.3 mg/kg) in layer diets did not affect egg production and quality parameters (FCR, egg weight, Haugh units, and eggshell thickness). Replacement of sodium selenite in the layer diet with Zn-SeMet was associated with an increase in the Se level in the egg albumin (1.92 vs. 1.35mg/kg), but did not affect Se level in the egg yolk (Chantiratikul et al., 2008). SeMet and Se-yeast efficacies have been compared in dogs using six biochemical markers, including intraprostatic dihydrotestosterone, testosterone, dihydrotestosterone: testosterone ratio, and epithelial cell DNA damage, proliferation, and apoptosis (Waters et al., 2012). By comparing dogs that achieved equivalent intraprostatic Se concentration, it was concluded that there was no significant difference in potency of the tested Se supplements. Indeed, SeMet is the main active ingredient of Se-yeast.
However, SeMet in purified form can be easily oxidized. For example, in freeze-dried oyster samples, total Se and the Se species detected were shown to be stable for at least 12 months; however, after purification of Se species, SeMet in the enzymatic extracts is only stable for 10 days if stored at 4°Cin Pyrex containers (Moreno et al., 2002). SeMet is also partially (∼20%) lost after storage in water solution for 30 days at 20°C (Lindemann et al., 2000). SeMet was identified as the major compound in gastrointestinal extract and its oxidation product (selenoxide, SeMetO) was the main degradation product formed after medium and long-term sample storage, respectively (Reyes et al., 2006). Indeed, pure SeMet is chemically oxidized to SeMetO in the small intestine (Lavu et al., 2016). According to a report by the EFSA (2013a), there is a problem with the stability of SeMet in premixtures containing trace elements; SeMet recovery from premixes after 3, 6, and 9 months of storage was 55%, 54%, and 37%, respectively.
OH-SeMet
Recently, a new organic Se source, OH-SeMet (2-hydroxy-4-melhylselenobutanoic acid or HMSeBA), has been developed (Briens et al., 2013, 2014). It is a precursor of SeMet and after its dietary consumption, OH-SeMet is easily converted into SeMet and metabolized in the same way as pure SeMet, including building Se reserves in muscles. OH-SeMet is synthesized in seven steps, and the solid form of the product is manufactured by spraying a 40% solution of OH-SeMet on a carrier (e.g. silica) and mixing. The liquid form of additive is prepared by dissolving solid OH-SeMet in distilled water (5% solution; EFSA, 2013b). In commercial products, OH-SeMet is shown to be more stable than pure SeMet (Geraert et al., 2015). It seems likely that OH-SeMet during storage behaves like the hydroxylated analog of Met, which is a stable product.
In a comparison of the effect of OH-SeMet with that of sodium selenite and Se-yeast, all three Se sources increased muscle Se compared with control chickens, with a significant (P<0.05) source effect in the order OH-SeMet > Se-yeast > sodium selenite and the effect of OH-SeMet being 1.5-fold stronger than that of Se-yeast (Briens et al., 2014). Selenium speciation in tissues indicates that muscle Se is exclusively present as SeMet or SeCys, indicating a full conversion of OH-SeMet to active Se species in the bird. These results corroborate the higher bioavailability of organic Se than that of mineral Se. Based on muscle Se levels, it is clear that OH-SeMet is characterised by a better bio-efficiency than Se-yeast; OH-SeMet was 39% more available than Se-yeast. It seems likely that the higher SeMet content in OH-SeMet supplement (nearly 100% in OH-SeMet vs. 60–70% in Seyeast) is responsible for the aforementioned differences. Interestingly, OH-SeMet in chicken diet increased the breast-muscle Se concentration compared to hatch value at day 21, while Se-yeast supplement only maintained the original Se concentration and sodium selenite significantly decreased the muscle Se concentration (Couloigner et a1., 2015). Furthermore, OH-SeMet in turkey diet improved GSH-Px activity in thigh muscles and decreased lipid peroxidation (Briens et al., 2016). In addition, compared with sodium selenite or Seyeast, OH-SeMet demonstrated an increased efficacy to enrich SeMet and total Se depositions, to induce mRNA expression of selenoprotein S and MsrB, thioredoxin reductase activity, and protein expression of GSH-Px4, selenoprotein P, and selenoprotein U in the tissues of chicks (Zhao et al., 2017). Similar to growing chickens, breeders would benefit from OH-SeMet supplementation, as laying hens fed a diet with 0.2 ppm Se as OH-SeMet transferred more Se to their eggs (+28.8%) and built bigger Se reserves in muscles (+28%) than birds fed the diet supplemented with 0.2 ppm Se in the Se-yeast form (Jlali et al., 2013).
Recently, in the EU, supplementation with Se-yeast has been limited to 0.2 mg Se/kg complete feed for reasons of consumer safety (Commission Implementing Regulation No. 427/2013 of 8 May 2013). Taking into account research data on the effects of organic Se on breeders, which are mainly related to dietary supplementation at 0.3 ppm, and previous commercial usage of such levels of Se supplementation, commercial premix and feed companies are looking for the most effective sources of organic Se. The idea is to deliver the same amount of Se to the egg and build the same Se reserves in muscles using 0.2 ppm Se dietary supplementation as can be achieved with 0.3 ppm in the form of Se-yeast. Indeed, the aforementioned results clearly showed that OH-SeMet supplied in the same dose as Se-yeast in the chicken diet would provide additional benefit in terms of Se reserves in the muscles as well as Se transfer to the egg and probably, to the developing embryo. This improvement in Se status could potentially be translated into higher selenoprotein expression and better antioxidant protection in stressful conditions of commercial poultry production (Zhao et al., 2017; Lei et al., 2017).
Se-homolanthionine (SeHLan)
A specific Se-yeast is produced based on Torula yeast (Cyberlindnera jadinii). Its specificity lies in the fact that it contains mainly SeHLan. However, it is not clear at present how SeHLan functions in the body. When pure SeMet and SeHLan were compared in a study in mice, Se-HLan was significantly less effective in liver Se accumulation than SeMet (Anan and Ogra, 2013). Indeed, SeHLan can be metabolized in poultry/animals similar to SeMet, but it cannot build Se reserves in the body (Anan et al., 2011) as only SeMet can be non-specifically incorporated into body proteins without altering their functions (Schrauzer and Surai, 2009).
Chelated Se Products
There is a range of products on the market that are claimed to contain chelated Se (including Se-glycinates, Se-proteinates, and Se-amino acid complexes); however, the chemical position of Se in the periodic table of elements indicates that Se is not a true metal, more a metalloid, and therefore, its chelating ability is questioned. Indeed, attempts to detect chelated Se in such products resulted in the detection of only inorganic Se (selenite and selenate) (Amoako et al., 2009; Kubachka et al., 2017). Recent bioassay results confirmed that those products (glycinates and proteinates) have a similar efficacy as sodium selenite (Liu et al., 2017). Similarly, Givens et al. (2004) showed that a chelated selenium–amino acid complex in cow diet was not different from sodium selenite when the efficacy of Se transfer to the milk was assessed. Indeed, chelated Se products are not related to SeMet and should probably not be included in the organic Se category.
Selenium Nanoparticle (nano-Se) Products
Recently, nano-Se have received substantial attention as potential novel nutritional supplements because of their low toxicity and novel characteristics, such as great specific surface area, high surface activity, and high catalytic efficiency (Pelyhe and Mezes, 2013; Sarkar et al., 2015; Wadhwani et al., 2016; Skalickova et al., 2017). In addition, it has been suggested that nano-Se can have important applications in the field of medicine owing to their antibacterial and anticancer properties (Wang et al., 2007; Ahmed et al., 2014; Wadhwani et al., 2016). Nano-Se has an ability to increase selenoenzyme activity comparable to that of SeMet. However, the question remains how elemental Se can be involved in SeCys synthesis and selenoprotein expression. There are several reports of successful testing of nano-Se in broiler nutrition (Wang, 2009; Zhou and Wang, 2011; Cai et al., 2012; Gulyas et al., 2017). In many cases, its low Se toxicity is considered the main advantage of nano-Se. However, one should realize that Se toxicity is not a major problem in poultry industry and Se in the form of sodium selenite or organic Se (SeMet, Se-yeast or other preparations) is an essential part of premixes produced worldwide. While it seems likely that nano-Se could be a new chemopreventive agent for the treatment of various diseases (Zhang et al., 2008), including cancer, its value as a feed supplement for the poultry industry is questionable.
Conclusion
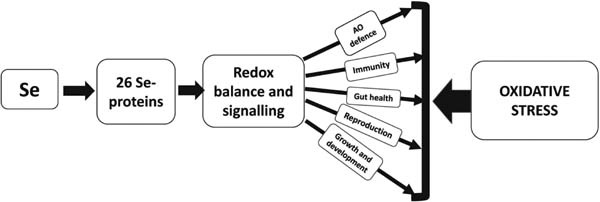
Se is an essential element in animal/poultry nutrition and its dietary supplementation in an optimal form and amount is key to maintaining animal health, productive, and reproductive performance. It has been proven that organic Se (mainly SeMet) in animal/poultry diets has a range of advantages over traditional sodium selenite. In fact, the organic Se concept considers SeMet as a storage form of Se in the chicken body. As animals/poultry are not able to synthesize SeMet, its provision through diet is a key strategy to fight commercially relevant stresses. Indeed, in stress conditions, when increased selenoprotein expression requires additional Se, but its provision with feed is decreased because of reduced feed consumption, Se reserves in the body (mainly in muscles) help maintain an effective antioxidant defense. It is well appreciated that via selenoprotein expression, Se is involved in the protection against oxidative stress and regulation of cell growth, apoptosis, and modification of cell signaling systems and transcription factors (Fig. 4). Therefore, its adequate dietary supply is crucial for various physiological processes in the animal/chicken body. Furthermore, it seems likely that increased muscle reserves of Se may enhance chicken resistance to stress and disease.
Fig. 4. Protective effects of Se in poultry.
The poultry industry is looking for the most effective sources of organic Se for commercial use. Several selenium compounds have been authorized as feed additives in the EU, including sodium selenite, sodium selenate, Se-yeast, l-SeMet, dl-SeMet, and OH-SeMet. In accordance with EU regulations, the total dietary Se is limited to 0.5mg/kg and the addition of the aforementioned organic Se compounds is limited to 0.2mg Se/kg (EFSA, 2014). Se-yeast has received substantial attention and is widely used in commercial products as a reliable source of organic Se. Generally speaking, there are three generations of Se supplements on the worldwide market. The first generation includes inorganic Se sources, mainly selenite and selenate. We would consider this generation of Se supplements as outdated. They are on the market since 1970 and helped animal and poultry industry get rid of real Se deficiency, but with today's precision nutrition concept, they have limited value and their only advantage is their comparatively low price. The second generation of Se supplements for poultry includes Se-yeast, SeMet, and Zn-SeMet. Usage of such Se supplements revolutionized Se nutrition over the last 20 years. However, a main disadvantage of Se-yeast is that the level of the active compound (SeMet) is quite variable (60% on average). There is no proof that the remaining fraction (30–50%) of Se, e.g., in the form of SeCys or MeSeCys, has any additional benefits to poultry in comparison to sodium selenite. From the production point of view, it is difficult to guarantee a certain percentage of SeMet in Se-yeast, as a range of production-related factors, including yeast strain, medium composition, selenite concentration, and temperature, affect the composition of the end product. Analytical difficulties in precisely determining the SeMet content in yeast-based products further complicate the issue. The main disadvantage of pure SeMet is its instability when included in premixes and feeds. The third generation of Se supplements for poultry is represented by OH-SeMet, which combined advantages of both Se-yeast (stability) and Se-Met (> 95% of active Se compound, i.e., SeMet). Clearly, there is a need for more research devoted to organic Se sources in poultry and farm animal nutrition.
There is a need for more research devoted to comparative studies of different forms of organic Se in poultry/animal nutrition to better understand the advantages and limitations of each form. In addition, there is a need for further clarification in which form Se is present in products such as Se-chelates, Se-proteinates, Se-glycinates to place them in a proper category of Se supplements. Indeed, Se-homolanthionine is commercially available but awaits more research to explain the molecular mechanisms of its metabolization and possible advantages to inorganic forms of Se. A quickly developing area of nano-Se research and application urgently needs well-designed studies with usage of modern molecular techniques to explain how metallic nano-Se particles are metabolized in the chicken/animal/human body to become a source of Se for synthesis of selenoproteins, which represent biological activity involving main active Se forms in biological systems.
Acknowledgments
Peter F. Surai and Ivan I. Kochish received a grant of the Government of Russian Federation (Grant No. 14.W03.31. 0013).
References
- Ahmed HH, Khalil WK and, Hamza AH. Molecular mechanisms of nano-selenium in mitigating hepatocellular carcinoma induced by N-nitrosodiethylamine (NDEA) in rats. Toxicology Mechanisms and Methods, 24: 593-602. 2014. [DOI] [PubMed] [Google Scholar]
- Alfthan G, Xu GL, Tan WH, Aro A, Wu J, Yang YX, Liang WS, Xue WL and Kong LH. Selenium supplementation of children in a selenium-deficient area in China: blood selenium levels and glutathione peroxidase activities. Biological Trace Element Research, 73: 113-125. 2000. [DOI] [PubMed] [Google Scholar]
- Amoako PO, Uden PC and Tyson JF. Speciation of selenium dietary supplements; formation of S-(methylseleno)cysteine and other selenium compounds. Analytica Chimica Acta, 652: 315-323. 2009. [DOI] [PubMed] [Google Scholar]
- Anan Y, Mikami T, Tsuji Y, Ogra Y. Distribution and metabolism of selenohomolanthionine labeled with a stable isotope. Analytical Bioanalitical Chemistry, 399: 1765-1772. 2011. [DOI] [PubMed] [Google Scholar]
- Anan Y and Ogra Y. Toxicological and pharmacological analysis of selenohomolanthionine in mice. Toxicology Research, 2: 115-122. 2013. [Google Scholar]
- Anan Y, Ohbo A, Tani Y and Ogra Y. Metabolic pathway of inorganic and organic selenocompounds labeled with stable isotope in Japanese quail. Analytical and Bioanalytical Chemistry, 406: 7959-7966. 2014. [DOI] [PubMed] [Google Scholar]
- Arnaudguilhem C, Bierla K, Ouerdane L, Preud'homme H, Yiannikouris A and Lobinski R. Selenium metabolomics in yeast using complementary reversed-phase/hydrophilic ion interaction (HILIC) liquid chromatography– electrospray hybrid quadrupole trap/Orbitrap mass spectrometry. Analytica Chimica Acta, 757: 26-38. 2012. [DOI] [PubMed] [Google Scholar]
- Attia YA, Abdalah AA, Zeweil HS, Bovera F, Tag El-Din AA and Araft MA. Effect of inorganic or organic selenium supplementation on productive performance, egg quality and some physiological traits of dual-purpose breeding hens. Czech Journal of Animal Science, 55: 505-519. 2010. [Google Scholar]
- Bakhshalinejad R, Akbari Moghaddam Kakhki R, Zoidis E. Effects of different dietary sources and levels of selenium supplements on growth performance, antioxidant status and immune parameters in Ross 308 broiler chickens. British Poultry Science (In Press). 2017 doi: 10.1080/00071668.2017.1380296. [DOI] [PubMed] [Google Scholar]
- Bierla K, Dernovics M, Vacchina V, Szpunar J, Bertin G and Lobinski R. Determination of selenocysteine and selenomethionine in edible animal tissues by 2D size-exclusion reversed-phase HPLC–ICP MS following carbamidomethylation and proteolytic extraction. Analytical and Bioanalytical Chemistry, 390: 1789-1798. 2008. [DOI] [PubMed] [Google Scholar]
- Bierla K, Szpunar J, Yiannikouris A and Lobinski R. Comprehensive speciation of selenium in selenium-rich yeast. Trends in Analytical Chemistry, 41: 122-132. 2012. [Google Scholar]
- Bierla K, Bianga J, Ouerdane L, Szpunar J, Yiannikouris A and Lobinski R. A comparative study of the Se/S substitution in methionine and cysteine in Se-enriched yeast using an inductively coupled plasma mass spectrometry (ICP MS)-assisted proteomics approach. Journal of Proteomics, 87: 26-39. 2013. [DOI] [PubMed] [Google Scholar]
- Brandt-Kjelsen A, Govasmark E, Haug A and Salbu B. Turnover of Se in adequately fed chickens using Se-75as a tracer. Journal of Animal Physiology and Animal Nutrition, 98: 547-558. 2014. [DOI] [PubMed] [Google Scholar]
- Briens M, Mercier Y, Rouffineau F, Vacchina V and Geraert PA. Comparative study of a new organic selenium source v. selenoyeast and mineral selenium sources on muscle selenium enrichment and selenium digestibility in broiler chickens. British Journal of Nutrition, 110: 617-624. 2013. [DOI] [PubMed] [Google Scholar]
- Briens M, Mercier Y, Rouffineau F and Geraert PA. 2-Hydroxy-4-methylselenobutanoic acid induces additional tissue selenium enrichment in broiler chicken compared to other selenium sources. Poultry Science, 93: 85-93. 2014. [DOI] [PubMed] [Google Scholar]
- Briens M, Faure M, Coiloigner F, Garet J, Maucotel T, Tommasino N, Gatelier P, Durand D, Garaert PA and Mercier Y. Hydroxyselenomethionine contributes to maintain color stability of turkey meat. Proceedings of Australian Poultry Science. Symposium, Sydney, Australia, pp. 149-152. 2016. [Google Scholar]
- Brozmanová J, Mániková D, Vlčková V and Chovanec M. Selenium: a double-edged sword for defense and offence in cancer. Archives of Toxicology, 84: 919-938. 2010. [DOI] [PubMed] [Google Scholar]
- Burdock GA and Cousins RJ. Amendment to the dossier in support of the generally recognozed as safe (GRAS) status of Sel-Plex as a food ingredient. https://www.fda.gov/downloads/Food/IngredientsPackagingLabeling/GRAS/NoticeInventory/ucm267426.pdf. 2010. Accessed August, 2017. [Google Scholar]
- Cai SJ, Wu CX, Gong LM, Song T, Wu H and Zhang LY. Effects of nano-selenium on performance, meat quality, immune function, oxidation resistance, and tissue selenium content in broilers. Poultry Science, 91: 2532-2539. 2012. [DOI] [PubMed] [Google Scholar]
- Casal SG, Far J, Bierla K, Ouerdane L and Szpunar J. Study of the Se-containing metabolomes in Se-rich yeast by size-exclusioncation-exchange HPLC with the parallel ICP MS and electrospray orbital ion trap detection. Metallomics, 2: 535-548. 2010. [DOI] [PubMed] [Google Scholar]
- Chambers I, Frampton J, Goldfarb P, Affara N, McBain W and Harrison PR. The structure of the mouse glutathione peroxidase gene: the selenocysteine in the active site is encoded by the 'termination’ codon, TGA. EMBO Journal, 5: 1221-1227. 1986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chantiratikul A, Chinrasri O, Chantiratikul P. Effect of sodium selenite and zinc-l-selenomethionine on performance and selenium concentrations in eggs of laying hens. Asian-Australasian Journal of Animal Sciences, 21: 1048-1052. 2008. [Google Scholar]
- Couloigner F, Jlali M, Briens M, Rouffineau F, Geraert PA and Mercier Y. Selenium deposition kinetics of different selenium sources in muscle and feathers of broilers. Poultry Science, 94: 2708-2714. 2015. [DOI] [PubMed] [Google Scholar]
- Davies MJ. Protein oxidation and peroxidation. Biochemical Journal, 473: 805-825. 2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deagen JT, Butler JA, Beilstein MA and Whanger PD. Effects of dietary selenite, selenocystine and selenomethionine on selenocysteine lyase and glutathione peroxidase activities and on selenium levels in rat tissues. Journal of Nutrition, 117: 91-98. 1987. [DOI] [PubMed] [Google Scholar]
- Delezie E, Rovers M, Van der Aa A, Ruttens A, Wittocx S, Segers L. Comparing responses to different selenium sources and dosages in laying hens. Poultry Science, 93: 3083-3090. 2014. [DOI] [PubMed] [Google Scholar]
- Dickerson K, Custer TW, Custer CM and Allen K. Bioavailability and exposure assessment of petroleum hydrocarbons and trace elements in birds nesting near the north platter river, Casper, Wyoming. Contaminants Report Number: R6/716C/00. U.S. Fish and Wildlife Service, Region 6, pp 1-72. 2002. [Google Scholar]
- Dobosz-Bartoszek M and Simonovic M. Structure and Mechanism of Selenocysteine Synthase. In: Selenium. Its Molecular Biology and Role in Human Health (Hatfield DL, Schweizer U, Tsuji PA and Gladyshev VN eds.). pp. 101-112. Springer International Publishing, New York. 2016. [Google Scholar]
- Drake EN. Cancer chemoprevention: selenium as a prooxidant, not an antioxidant. Medical Hypotheses, 67: 318-322. 2006. [DOI] [PubMed] [Google Scholar]
- EFSA. Selenium-enriched yeast as source for selenium added for nutritional purposes in foods for particular nutritional uses and foods (including food supplements) for the general population. Scientific Opinion of the Panel on Food Additives. EFSA Journal, 766: 1-42. 2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- EFSA. Scientific Opinion on the safety and efficacy of L-selenomethionine as feed additive for all animal species. EFSA Journal, 11: 3219. 2013a. [Google Scholar]
- EFSA. Scientific Opinion on safety and efficacy of hydroxyanalogue of selenomethionine as feed additive for all species. EFSA Journal, 11: 3046. 2013b. [Google Scholar]
- EFSA. Scientific Opinion on the safety and efficacy of DL-selenomethionine as a feed additive for all animal species. EFSA Journal, 12: 3567. 2014. [Google Scholar]
- EFSA. Selenium-enriched yeast (Alkosel®3000) for all animal species. EFSA Journal, 14: 4624. 2016. [Google Scholar]
- Eisenberg S. Relative stability of selenites and selenates in feed premixes as a function of water activity. Journal of AOAC International, 90: 349-353, 2007. [PubMed] [Google Scholar]
- Encinar JR, Ruzik R, Buchmann W, Tortajada J, Lobinski R and Szpunar J. Detection of selenocompounds in a tryptic digest of yeast selenoprotein by MALDI time-of-flight MS prior to their structural analysis by electrospray ionization triple quadrupole MS. Analyst, 128: 220-224. 2003. [DOI] [PubMed] [Google Scholar]
- Esmaeili S and Khosravi-Darani K. Selenium-enriched yeast: As selenium source for nutritional purpose. Current Nutrition & Food Science, 10: 49-56. 2014. [Google Scholar]
- Fagan S, Owens R, Ward P, Connolly C, Doyle S and Murphy R. Biochemical comparison of commercial selenium yeast preparations. Biological Trace Element Research, 166: 245-259. 2015. [DOI] [PubMed] [Google Scholar]
- Fan XY, Guo XN, Fu XH, He XP, Wang CL and Zhang BR. The breeding and culture condition optimization of a high-biomass, selenium-enriched yeast strain. Sheng Wu Gong Cheng Xue Bao 19: 720-724. 2003. [PubMed] [Google Scholar]
- Fisinin VI, Papazyan TT and Surai PF. Selenium in poultry nutrition. In: Current Advances in Se Research and Applications (Surai PF and Taylor-Pickard J eds.). Vol. 1. pp. 221-261. Wageningen Academic Publishers, Wageningen, The Netherlands. 2008. [Google Scholar]
- Fisinin VI, Shatskikh EV, Latipova EN and Surai PF. Maternal effect in poultry – from vitamins to vitagenes and epigenetics. Ptitza and Ptitzeproducti (Poultry and Poultry Products), Moscow, 1: 29-33. 2016. [Google Scholar]
- Gao X, Zhang Z, Xing H, Yu J, Zhang N and Xu S. Selenium deficiency induced inflammation and increased expression of regulating inflammatory cytokines in the chicken gastrointestinal tract. Biological Trace Element Research, 173: 210-218. 2016. [DOI] [PubMed] [Google Scholar]
- Geraert PA, Briens M, Mercier Y, Liu YJ. Comparing organic selenium sources. Asian Poultry Magazine, August, 26-29. 2015. [Google Scholar]
- Gilbert-López B, Dernovics M, Moreno-González D, Molina-Díaz A and García-Reyes JF. Detection of over 100 selenium metabolites in selenized yeast by liquid chromatography electrospray time-of-flight mass spectrometry. Journal of chromatography. B, Analytical Technologies in the Biomedical and Life Sciences, 1060: 84-90. 2017. [DOI] [PubMed] [Google Scholar]
- Givens DI, Allison R, Cottrill B, Jonathon S and Blake JS. Enhancing the selenium content of bovine milk through alteration of the form and concentration of selenium in the diet of the dairy cow. Journal of Science in Food and Agriculture, 84: 811-817. 2004. [Google Scholar]
- Gladishev VN, Carlson BA, Hatfield DL. Pathways in de novo biosynthesis of selenocysteine and cysteine in eukaryotes. In: Selenium. Its Molecular Biology and Role in Human Health (Hatfield DL, Schweizer U, Tsuji PA and Gladyshev VN eds.). pp. 39-45. Springer International Publishing, New York. 2016. [Google Scholar]
- Gosetti F, P. Frascarolo P, Polati S, Medana C, Gianotti V, Palma P, Aigotti R, Baiocchi C and Gennaro MC. Speciation of selenium in diet supplements by HPLC–MS/MS methods. Food Chemistry, 105: 1738-1747. 2007. [Google Scholar]
- Griffiths NM, Stewart RD and Robinson MF. The metabolism of [75Se] in four women. British Journal of Nutrition, 35: 373-382. 1976. [DOI] [PubMed] [Google Scholar]
- Groce AW, Miller ER, Hitchcock JP, Ullrey DE and Magee WT. Selenium balance in the pig as affected by selenium source and vitamin E. Journal of Animal Science, 37: 942-947. 1973. [DOI] [PubMed] [Google Scholar]
- Gulyas G, Csosz E, Prokisch J, Javor A, Mezes M, Erdelyi M, Balogh K, Janaky T, Szabo Z, Simon A and Czegledi L. Effect of nano-sized, elemental selenium supplement on the proteome of chicken liver. Journal of Animal Physiology and Animal Nutrition, 101: 502-510. 2017. [DOI] [PubMed] [Google Scholar]
- Ip C. Interaction of vitamin C and selenium supplementation in the modification of mammary carcinogenesis in rats. Journal of the National Cancer Institute, 77: 299-303. 1986. [PubMed] [Google Scholar]
- Ip C and Hayes C. Tissue selenium levels in selenium-supplemented rats and their relevance in mammary cancer protection. Carcinogenesis, 10: 921-925. 1989. [DOI] [PubMed] [Google Scholar]
- Ip C, Birringer M, Block E, Kotrebai M, Tyson JF, Uden PC and Lisk DJ. Chemical speciation influences comparative activity of selenium-enriched garlic and yeast in mammary cancer prevention. Journal of Agricultural and Food Chemistry, 48: 2062-2070. 2000. [DOI] [PubMed] [Google Scholar]
- Jiang Z, Lin Y, Zhou G, Luo L, Jiang S, Chen F. Effects of dietary selenomethionine supplementation on growth performance, meat quality and antioxidant property in yellow broilers. Journal of Agricultural and Food Chemistry, 57: 9769-9772. 2009. [DOI] [PubMed] [Google Scholar]
- Jlali M, Briens M, Rouffineau F, Mercerand F, Geraert PA and Mercier Y. Effect of 2-hydroxy-4-methylselenobutanoic acid as a dietary selenium supplement to improve the selenium concentration of table eggs. Journal of Animal Science, 91: 1745-1752. 2013. [DOI] [PubMed] [Google Scholar]
- Kaya A, Lee BC, Gladyshev VN. Regulation of protein function by reversible methionine oxidation and the role of selenoprotein MsrB1. Antioxidants & Redox Signaling, 23: 814-822. 2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kotrebai M, Birringer M, Tyson JF, Block E and Uden PC. Selenium speciation in enriched and natural samples by HPLCICP-MS and HPLC-ESI-MS with perfluorinated carboxylic acid ion-pairing agents. Analyst, 125: 71-78. 2000. [DOI] [PubMed] [Google Scholar]
- Kretz-Remy C and Arrigo AP. Modulation of the chymotrypsin-like activity of the 20S proteasome by intracellular redox status: effects of glutathione peroxidase-1 overexpression and antioxidant drugs. Biological Chemistry, 384: 589-595. 2003. [DOI] [PubMed] [Google Scholar]
- Kryukov GV, Castellano S, Novoselov SV, Lobanov AV, Zehtab O, Guigó R and Gladyshev VN. Characterization of mammalian selenoproteomes. Science, 300: 1439-1443. 2003. [DOI] [PubMed] [Google Scholar]
- Kubachka KM, Hanley T, Mantha M, Wilson RA, Falconer TM, Kassa Z, Oliveira A, Landero J and Caruso J. Evaluation of selenium in dietary supplements using elemental speciation. Food Chemistry, 218: 313-320. 2017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laika M, Jahanian R. Dietary supplementation of organic selenium could improve performance, antibody response, and yolk oxidative stability in laying hens fed on diets containing oxidized fat. Biological Trace Element Research, 165: 195-205. 2015. [DOI] [PubMed] [Google Scholar]
- Lam JC, Tanabe S, Lam MH and Lam PK. Risk to breeding success of water birds by contaminants in Hong Kong: evidence from trace elements in eggs. Environmental Pollution, 135: 481-490. 2005. [DOI] [PubMed] [Google Scholar]
- Larsen EH, Hansen M, Paulin H, Moesgaard S, Reid M and Rayman M. Speciation and bioavailability of selenium in yeast-based intervention agents used in cancer chemoprevention studies. Journal of AOAC International, 87: 225-232. 2004. [PubMed] [Google Scholar]
- Lavu RV, Van De Wiele T, Pratti VL, Tack F and Du Laing G. Selenium bioaccessibility in stomach, small intestine and colon: Comparison between pure Se compounds, Se-enriched food crops and food supplements. Food Chemistry, 197 (Pt A): 382-387. 2016. [DOI] [PubMed] [Google Scholar]
- Lee BC. Biochemistry and Function of Methionine Sulfoxide Reductase. In: Selenium. Its Molecular Biology and Role in Human Health (Hatfield DL, Schweizer U, Tsuji PA and Gladyshev VN eds.). pp. 287-292. Springer International Publishing, New York. 2016. [Google Scholar]
- Lei X. Avian selenogenome: Response to dietary Se and protection against oxidative insults. Poultry Science, 96 (E-supplement-1): 220. 2017. [Google Scholar]
- Levander OA, Alfthan G, Arvilommi H, Gref CG, Huttunen JK, Kataja M, Koivistoinen P and Pikkarainen J. Bioavailability of selenium to Finnish men as assessed by platelet glutathione peroxidase activity and other blood parameters. American Journal of Clinical Nutrition, 37: 887-897. 1983. [DOI] [PubMed] [Google Scholar]
- Lindemann T, Prange A, Dannecker W and Neidhart B. Stability studies of arsenic, selenium, antimony and tellurium species in water, urine, fish and soil extracts using HPLC/ICP-MS. Fresenius Journal of Analytical Chemistry, 368: 214-220. 2000. [DOI] [PubMed] [Google Scholar]
- Lipiec E, Siara G, Bierla K, Ouerdane L and Szpunar J. Determination of selenomethionine, selenocysteine, and inorganic selenium in eggs by HPLC-inductively coupled plasma mass spectrometry. Analytical and Bioanalytical Chemistry, 397: 731-741. 2010. [DOI] [PubMed] [Google Scholar]
- Liu YG, Geraert PA and Briens M. How to compare organic selenium sources? Proceedings of 28th Annual Australian Poultry Science Symposium, Sydney, New South Wales, pp. 217-221. 2017. [Google Scholar]
- Mapelli V, Hillestrom PR, Patil K, Larsen EH and Olsson L. The interplay between sulphur and selenium metabolism influences the intracellular redox balance in Saccharomyces cerevisiae. FEMS Yeast Research, 12: 20-32. 2012. [DOI] [PubMed] [Google Scholar]
- McSheehy S, Yang L, Sturgeon R and Mester Z. Determination of methionine and selenomethionine in selenium-enriched yeast by species-specific isotope dilution with liquid chromatography– mass spectrometry and inductively coupled plasma mass spectrometry detection. Analytical Chemistry, 77: 344-349, 2005. [DOI] [PubMed] [Google Scholar]
- Moreno P, Quijano MA, Gutierrez AM, Perez-Conde MC and Camara C. Stability of total selenium and selenium species in lyophilised oysters and in their enzymatic extracts. Analytical and Bioanalytical Chemistry, 374: 466-476. 2002. [DOI] [PubMed] [Google Scholar]
- Mutanen M, Mykkänen HM. Effect of ascorbic acid supplementation on selenium bioavailability in humans. Human Nutrition. Clinical Nutrition, 39: 221-226. 1985. [PubMed] [Google Scholar]
- Mykkänen HM and Mutanen ML. Effect of ascorbic acid on the intestinal absorption of 75Se-selenite in chicks. Nutrition Reports International, 28: 67-73. 1983. [Google Scholar]
- Nicastro HL, Dunn BK. Selenium and prostate cancer prevention: insights from the selenium and vitamin E cancer prevention trial (SELECT). Nutrients, 5: 1122-1148. 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oster O, Schmiedel G and Prellwitz W. The organ distribution of selenium in German adults. Biological Trace Element Research. 15: 23-45. 1988. [DOI] [PubMed] [Google Scholar]
- Pajares M, Jiménez-Moreno N, Dias IH, Debelec B, Vucetic M, Fladmark KE, Basaga H, Ribaric S, Milisav I and Cuadrado A. Redox control of protein degradation. Redox Biology, 6: 409-420. 2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pappas AC, Karadas F, Surai PF and Speake BK. The selenium intake of the female chicken influences the selenium status of her progeny. Comparative Biochemistry and Physiology B, Biochemistry and Molecular Biology, 142: 465-474. 2005. [DOI] [PubMed] [Google Scholar]
- Pappas AC, Acamovic T, Sparks NH, Surai PF and McDevitt RM. Effects of supplementing broiler breeder diets with organoselenium compounds and polyunsaturated fatty acids on hatchability. Poultry Science, 85: 1584-1593. 2006b. [DOI] [PubMed] [Google Scholar]
- Pappas AC, Karadas F, Surai PF, Wood N, Cassey P and Speake BK Interspecies variation in yolk selenium concentrations among eggs of free-living birds. Journal of Trace Elements in Medicine and Biology, 20: 155-160. 2006a. [DOI] [PubMed] [Google Scholar]
- Patterson BH, Levander OA, Helzlsouer K, McAdam PA, Lewis SA, Taylor PR, Veillon C and Zech LA. Human selenite metabolism: a kinetic model. American Journal of Physiology, 257: R556-R567. 1989. [DOI] [PubMed] [Google Scholar]
- Payne RL and Southern LL. Changes in glutathione peroxidase and tissue selenium concentrations of broilers after consuming a diet adequate in selenium. Poultry Science, 84: 1268-1276. 2005. [DOI] [PubMed] [Google Scholar]
- Pelyhe C and Mezes M. Myths and facts about the effects of nanoselenium in farm animals-Mini-Review. European Chemical Bulletin, 2: 1049-1052. 2013. [Google Scholar]
- Persson-Moschos M, Alfthan G and Akesson B. Plasma selenoprotein P levels of healthy males in different selenium status after oral supplementation with different forms of selenium. European Journal of Clinical Nutrition, 52: 363-367. 1998. [DOI] [PubMed] [Google Scholar]
- Pierce S and Tappel AL. Effects of selenite and selenomethionine on glutathione peroxidase in the rat. Journal of Nutrition, 107: 475-479. 1977. [DOI] [PubMed] [Google Scholar]
- Rajashree K and Muthukumar T. Preparation of selenium tolerant yeast Saccharomyces cerevisiae. Journal of Microbiology and Biotechnology Research, 3: 46-53. 2013. [Google Scholar]
- Rao Y, McCooeye M, Windust A, Bramanti E, D'Ulivo A and Mester Z. Mapping of selenium metabolic pathway in yeast by liquid chromatography-Orbitrap mass spectrometry. Analytical Chemistry, 82: 8121-8130. 2010. [DOI] [PubMed] [Google Scholar]
- Rayman MP. The use of high-selenium yeast to raise selenium status: how does it measure up? British Journal of Nutrition, 92: 557-573. 2004. [DOI] [PubMed] [Google Scholar]
- Reilly C. Selenium in food and health. Springer Science + Business Media, LLC, USA. 2006. [Google Scholar]
- Reyes LH, Marchante-Gayón JM, Alonso JIJ and Sanz-Medel A. Application of isotope dilution analysis for the evaluation of extraction conditions in the determination of total selenium and selenomethionine in yeast-based nutritional supplements. Journal of Agricultural and Food Chemistry, 54: 1557-1563. 2006. [DOI] [PubMed] [Google Scholar]
- Robinson MF, Thomson CD and Huemmer PK. Effect of a megadose of ascorbic acid, a meal and orange juice on the absorption of selenium as sodium selenite. New Zealand Medical Journal, 98: 627-629. 1985. [PubMed] [Google Scholar]
- Rotruck JT, Pope AL, Ganther HE, Swanson AB, Hafeman DG and Hoekstra WG. Selenium: biochemical role as a component of glutathione peroxidase. Science, 179: 588-590. 1973. [DOI] [PubMed] [Google Scholar]
- Sarkar B, Bhattacharjee S, Daware A, Tribedi P, Krishnani KK and Minhas PS. Selenium Nanoparticles for Stress-Resilient Fish and Livestock. Nanoscale Research Letters, 10: 371. 2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Savinov VM, Gabrielsen GW and Savinova TN. Cadmium, zinc, copper, arsenic, selenium and mercury in seabirds from the Barents Sea: levels, inter-specific and geographical differences. The Science of the Total Environment, 306: 133-158. 2003. [DOI] [PubMed] [Google Scholar]
- Schrauzer GN. Selenomethionine: a review of its nutritional significance, metabolism and toxicity. Journal of Nutrition, 130: 1653-1656. 2000. [DOI] [PubMed] [Google Scholar]
- Schrauzer GN. Nutritional selenium supplements: product types, quality, and safety. Journal of the American College of Nutrition, 20: 1-4. 2001. [DOI] [PubMed] [Google Scholar]
- Schrauzer GN. The nutritional significance, metabolism and toxicology of selenomethionine. Advances in Food and Nutrition Research, 47: 73-112. 2003. [DOI] [PubMed] [Google Scholar]
- Schrauzer GN. Selenium yeast: Composition, quality, analysis, and safety. Pure and Applied Chemistry, 78, 1: 105-109. 2006. [Google Scholar]
- Schrauzer GN and Surai PF. Selenium in human and animal nutrition: resolved and unresolved issues. A partly historical treatise in commemoration of the fiftieth anniversary of the discovery of the biological essentiality of selenium, dedicated to the memory of Klaus Schwarz (1914–1978) on the occasion of the thirtieth anniversary of his death. Critical Reviews in Biotechnology, 29: 2-9. 2009. [DOI] [PubMed] [Google Scholar]
- Schwarz K and Foltz CM. Selenium as an integral part of factor 3 against dietary liver degeneration. Journal of American Chemical Society, 79: 3292-3293. 1957. [Google Scholar]
- Schwarz K and Foltz CM. Factor 3 activity of selenium compounds. Journal of Biological Chemistry, 233: 245-251. 1958. [PubMed] [Google Scholar]
- Skalickova S, Milosavljevic V, Cihalova K, Horky P, Richtera L and Adam V. Selenium nanoparticles as a nutritional supplement. Nutrition, 33: 83-90. 2017. [DOI] [PubMed] [Google Scholar]
- Simon E, Ballet N, Francesch M and Brufau J. Comparison between the dietary supplementation of sodium selenite and Selenium-yeast on meat Se accumulation in broiler. World’ s Poultry Science Journal, 69 (Suppl.): 1-5. 2013. [Google Scholar]
- Sloth JJ, Larsen EH, Bügel SH and Moesgaard S. Determination of total selenium and 77Se in isotopically enriched human samples by ICP-dynamic reaction cell-MS. Journal of Analytical Atomic Spectrometry, 18: 317-322. 2003. [Google Scholar]
- Spallholz JE. Free radical generation by selenium compounds and their prooxidant toxicity. Biomedical and Environmental Sciences, 10: 260-270. 1997. [PubMed] [Google Scholar]
- Spallholz J E and Rafferty A. Nutritional, chemical and toxicological evaluation of high-selenium yeast. In: Selenium in Biology and Medicine, Part A (Combs GF, Spallholz JE, Levander OA, Oldfield JE eds.). pp. 516-529. Avi-Van Nostrand Reinhold New York, NY. 1987. [Google Scholar]
- Stout JH and Trust KA. Elemental and organochlorine residues in bald eagles from Adak Island, Alaska. Journal of Wildlife Diseases, 38: 511-517. 2002. [DOI] [PubMed] [Google Scholar]
- Stout JH, Trust KA, Cochrane JF, Suydam RS and Quakenbush LT.Environmental contaminants in four eider species from Alaska and arctic Russia. Environmental Pollution, 119: 215-226. 2002. [DOI] [PubMed] [Google Scholar]
- Surai PF. Effect of the selenium and vitamin E content of the maternal diet on the antioxidant system of the yolk and the developing chick. British Poultry Science 41: 235-243. 2000. [DOI] [PubMed] [Google Scholar]
- Surai PF. Selenium in Nutrition and Health. Nottingham University Press, Nottingham. 2006. [Google Scholar]
- Surai PF and Fisinin VI. Selenium in poultry breeder nutrition: An update. Animal Feed Science and Technology, 191: 1-15. 2014. [Google Scholar]
- Surai PF and Fisinin VI. Vitagenes in poultry production. Part 1. Technological and environmental stresses. World's Poultry Science Journal, 72: 721-733. 2016a. [Google Scholar]
- Surai PF and Fisinin VI. Vitagenes in poultry production. Part 2. Nutritional and Internal stresses. World's Poultry Science Journal 72: 761-772. 2016b. [Google Scholar]
- Surai PF, Fisinin VI and Karadas F. Antioxidant Systems in Chick Embryo Development. Part 1. Vitamin E, Carotenoids and Selenium. Animal Nutrition, 2: 1-11. 2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Surai PF, Karadas F, Pappas AC and Sparks NH. Effect of organic selenium in quail diet on its accumulation in tissues and transfer to the progeny. British Poultry Science, 47: 65-72. 2006. [DOI] [PubMed] [Google Scholar]
- Suryo Rahmanto A and Davies MJ. Catalytic activity of selenomethionine in removing amino acid, peptide, and protein hydroperoxides. Free Radical Biology & Medicine, 51: 2288-2299. 2011. [DOI] [PubMed] [Google Scholar]
- Swanson CA, Patterson BH, Levander OA, Veillon C, Taylor PR, Helzlsouer K, McAdam PA and Zech LA. Human [74Se] selenomethionine metabolism: a kinetic model. American Journal of Clinical Nutrition 54: 917-926. 1991. [DOI] [PubMed] [Google Scholar]
- Takahashi K, Suzuki N and Ogra Y. Bioavailability Comparison of Nine Bioselenocompounds In Vitro and In Vivo. International Journal of Molecular Sciences, 18:3. 2017. doi: 10.3390/ijms18030506. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tufarelli V, Ceci E and Laudadio V. 2-Hydroxy-4-Methylselenobutanoic Acid as New Organic Selenium Dietary Supplement to Produce Selenium-Enriched Eggs. Biological Trace Element Research, 171: 453-458. 2016. [DOI] [PubMed] [Google Scholar]
- Uden PC, Boakye HT, Kahakachchi C, Hafezi R, Nolibos P, Block E, Johnson S and Tyson, J.F. Elemental selective characterization of stability and reactivity of Selenium species in selenized yeast. Journal of Analytical Atomic Spectrometry, 19: 65-73. 2004. [Google Scholar]
- Urso UR, Dahlke F, Maiorka A, Bueno IJ, Schneider AF, Surek D, Rocha C. Vitamin E and selenium in broiler breeder diets: Effect on live performance, hatching process, and chick quality. Poultry Science, 94: 976-983. 2015. [DOI] [PubMed] [Google Scholar]
- Van Beirendonck S, Driessen B, Rovers M, Segers L, Ruttens A and Du Laing G. Relation between selenomethionine content in dietary selenium sources and selenium deposition in broiler muscle tissue. Proceedings of XXV World's Poultry Congress, Beijing, China, pp. 63. 2016a. [Google Scholar]
- Van Beirendonck S, Driessen B, Rovers M, Segers L, Elsin I., Ruttens A and Du Laing G. Relation between selenomethionine in dietary selenium sources and selenium deposition in broiler. Book of Abstracts of the 67th Annual Meeting of the European Association for Animal production, pp. 662. 2016b. [Google Scholar]
- Wadhwani SA, Shedbalkar UU, Singh R and Chopade BA. Biogenic selenium nanoparticles: current status and future prospects. Applied Microbiology and Biotechnology, 100: 2555-2566. 2016. [DOI] [PubMed] [Google Scholar]
- Wang H, Zhang J and Yu H. Elemental selenium at nano size possesses lower toxicity without compromising the fundamental effect on selenoenzymes: comparison with selenomethionine in mice. Free Radical Biology & Medicine 42: 1524-1533. 2007. [DOI] [PubMed] [Google Scholar]
- Wang Y. Differential effects of sodium selenite and nano-Se on growth performance, tissue se distribution, and glutathione peroxidase activity of avian broiler. Biological Trace Element Research, 128: 184-190. 2009. [DOI] [PubMed] [Google Scholar]
- Wang Y, Zhan X, Zhang X, Wu R, Yuan D. Comparison of different forms of dietary selenium supplementation on growth performance, meat quality, selenium deposition, and antioxidant property in broilers. Biological Trace Element Research, 143: 261-273. 2011. [DOI] [PubMed] [Google Scholar]
- Wang Y, Wang H, Zhan X. Effects of different dl-selenomethionine and sodium selenite levels on growth performance, immune functions and serum thyroid hormones concentrations in broilers. Journal of Animal Physiology and Animal Nutrition 100: 431-439. 2016. [DOI] [PubMed] [Google Scholar]
- Waters DJ, Shen S, Kengeri SS, Chiang EC, Combs Jr GF, Morris JS and Bostwick DG. Prostatic response to supranutritional selenium supplementation: comparison of the target tissue potency of selenomethionine vs. selenium-yeast on markers of prostatic homeostasis. Nutrients, 4: 1650-1663. 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wolf WR, Zainal H and Yager B. Selenomethionine content of candidate reference materials. Fresenius Journal of Analytical Chemistry, 370: 286-290. 2001. [DOI] [PubMed] [Google Scholar]
- Wolffram S, Berger B, Grenacher B and Scharrer E. Transport of selenoamino acids and their sulfur analogues across the intestinal brush border membrane of pigs. Journal of Nutrition, 119: 706-712. 1989. [DOI] [PubMed] [Google Scholar]
- Wrobel K, Kannamkumarath SS, Wrobel K and Caruso JA. Hydrolysis of proteins with methanesulfonic acid for improved HPLC-ICP-MS determination of seleno-methionine in yeast and nuts. Analytical and Bioanalytical Chemistry, 375: 133-138. 2003. [DOI] [PubMed] [Google Scholar]
- Wu C, Xu Z and Huang K. Effects of Dietary Selenium on Inflammation and Hydrogen Sulfide in the Gastrointestinal Tract in Chickens. Biological Trace Element Research, 174: 428-435. 2016. [DOI] [PubMed] [Google Scholar]
- Yoshida M, Abe M, Fukunaga K and Kikuchi K. Bioavailability of selenium in the defatted dark muscle of tuna. Food Additives and Contaminants, 19: 990-995. 2002. [DOI] [PubMed] [Google Scholar]
- Xiang N, Zhao R and Zhong W. Sodium selenite induces apoptosis by generation of superoxide via the mitochondrial-dependent pathway in human prostate cancer cells. Cancer Chemotherapy and Pharmacology, 63: 351-362. 2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang J, Wang X and Xu T. Elemental selenium at nano size (Nano-Se) as a potential chemopreventive agent with reduced risk of selenium toxicity: comparison with se-methylselenocysteine in mice. Toxicological Sciences, 101: 22-31. 2008. [DOI] [PubMed] [Google Scholar]
- Zhao L, Sun LH, Huang JQ, Briens M, Qi DS, Xu SW and Lei XG. A novel organic selenium compound exerts unique regulation of selenium speciation, selenogenome, and selenoproteins in broiler chicks. Journal of Nutrition 147: 789-797. 2017. [DOI] [PubMed] [Google Scholar]
- Zhou X and Wang Y. Influence of dietary nano elemental selenium on growth performance, tissue selenium distribution, meat quality, and glutathione peroxidase activity in Guangxi Yellow chicken. Poultry Science, 90: 680-686. 2011. [DOI] [PubMed] [Google Scholar]