

Abstract
ON bipolar neurons in retina detect the glutamate released by rods and cones via metabotropic glutamate receptor 6 (mGluR6), whose cascade is unknown. The trimeric G-protein Go might mediate this cascade because it colocalizes with mGluR6. To test this, we studied the retina in mice negative for the α subunit of Go (Gαo−/−). Retinal layering, key cell types, synaptic structure, and mGluR6 expression were all normal, as was the a-wave of the electroretinogram, which represents the rod and cone photocurrents. However, the b-wave of the electroretinogram, both rod- and cone-driven components, was entirely missing. Because the b-wave represents the massed response of ON bipolar cells, its loss in the Gαo null mouse establishes that the light response of the ON bipolar cell requires Go. This represents the first function to be defined in vivo for the α subunit of the most abundant G-protein of the brain .
Keywords: G-protein, mGluR6, metabotropic glutamate receptor, retina, bipolar cells, electroretinogram, rod bipolar cells, second messenger cascade
ON bipolar neurons comprise three-quarters of all bipolar cells in mammalian retina, including all rod bipolar and one-half of the cone bipolar cells (Sterling et al., 1988; Cohen and Sterling, 1990; Martin and Grünert, 1992;Strettoi and Masland, 1995). These neurons detect glutamate released by rods and cones by means of metabotropic glutamate receptor 6 (mGluR6) (Nakajima et al., 1993; Nomura et al., 1994). Initially, mGluR6 was thought to trigger a cascade resembling that for phototransduction (Nawy and Jahr, 1990; Shiells and Falk, 1990). However, the main components of the phototransduction cascade (transducin, phosphodiesterase, and a cGMP-gated cation channel) were not found in the ON bipolar dendrites (Wässle et al., 1992; Vardi et al., 1993). Instead, a different trimeric G-protein was found: Go (Vardi et al., 1993; Vardi, 1998).
Go has been suggested as the second step in the ON bipolar cascade because of the following: (1) it colocalizes with mGluR6 in the dendrites (Vardi, 1998; Vardi et al., 2000); (2) agonist-bound mGluR6 can activate its α subunit (Gαo) in vitro (Weng et al., 1998); and (3) Gαo dialyzed into ON bipolar cells reduces the response to glutamate (Nawy, 1999). However, doubt persists regarding the role of Go because, rather than coupling mGluR6 to the effector, it might merely modulate the cascade, e.g., by affecting channel phosphorylation. If Gomerely modulates the cascade, a retina lacking Gαo might have an altered ON response (Greif et al., 2000), but if Gαoactually couples mGluR6 to an effector, a null retina would have no ON response. Here we show that the retina in a Gαo−/− mouse appears normal, but the ON response to rod and cone stimuli, as measured by the b-wave of an electroretinogram (ERG), is completely absent.
MATERIALS AND METHODS
Mice null for Gαo (both splice variants) rarely reach adulthood (Valenzuela et al., 1997; Jiang et al., 1998), so the colony was maintained by breeding heterozygotes. Heterozygous animals were received from the late Eva J. Neer (Harvard Medical School, Boston, MA). All experiments involving animals were done in compliance with federal regulations and University of Pennsylvania policy.
Genotyping. The tip of a mouse tail (0.5 cm) was incubated in lysis buffer containing 100 mm Tris-HCl, pH 8.5, 5 mm EDTA, 0.2% SDS, 200 mm NaCl, and 100 μg/ml proteinase K at 55°C overnight. After centrifugation, the supernatant was decanted into 500 μl of isopropanol to precipitate the DNA. PCR primers used to amplify Gαo were as follows: 5′-AGG GGA TGA GAG CCG CCT GCA GTC-3′ and 5′-ATG ATG GCC GTG ACA TCC TCG AAG CA-3′. Theneo gene was amplified using 5′-ACC TGG TCA TAG CCG CTG AGT-3′ and 5′-TGC CGA GAA AGT ATC CAT CAT G-3′ (Jiang et al., 1998). PCR conditions were as follows: an initial cycle at 96°C (3 min), 63°C (5 min), and 72°C (45 sec); then 30 cycles at 94°C (30 sec), 63°C (30 sec), and 72°C (45 sec); and a final hold at 72°C (5 min).
Electroretinographic recordings. The experimental apparatus, methods of light stimulation and quantification, ERG recording, and cone signal isolation have been described in detail previously (Lyubarsky et al., 1999). Briefly, a mouse was dark-adapted for 2 hr; then, under dim red light, it was deeply anesthetized by injecting intraperitoneally ketamine (20 μg/gm) plus xylazine (8 μg/gm). The animal was immobilized in a holder with the right eye pointing upward; then the pupil was dilated with 1% tropicamide, and the eye was protected with a drop of methylcellulose. A platinum recording electrode contacted the cornea, and a tungsten reference electrode was inserted subcutaneously on the forehead. The animal in its holder was then placed inside a light-proof Faraday cage, and light stimuli were delivered through several ports. Light stimuli were 1 msec flashes generated with xenon flash tubes. The intensity and spectral composition were controlled with neutral density and bandpass interference filters. Light intensities were calibrated and converted to estimated number of photoisomerization per photoreceptor (R*) as described previously (Lyubarsky et al., 1999). For dim flashes, 10- 40 responses were averaged for each experimental condition; for intense flashes, two to five responses were averaged.
Immunocytochemistry. Eyes were enucleated from an anesthetized mouse (85 μg/gm ketamine and 13 μg/gm xylazine), and the animal was killed by anesthetic overdose. A small cut was made through the lens, after which the eyeball was immersion-fixed in 4% paraformaldehyde plus 0.01% glutaraldehyde buffered at pH 7.3 [room temperature (RT), 1 hr]. The eye was then rinsed in buffer, soaked overnight in 30% buffered sucrose, embedded in a mixture of 20% buffered sucrose and tissue freezing medium (2:1), and cryosectioned at 10 μm. Sections were soaked in 0.1 m phosphate buffer containing 10% normal goat serum, 5% sucrose, and 0.3% Triton X-100 (diluent) (RT, 1 hr), incubated in primary antibody (in diluent) (4°C, overnight), washed and incubated in secondary antibody conjugated to a fluorescent marker (3 hr) (Jackson ImmunoResearch, West Grove, PA), and mounted in Vectashield (Vector Laboratories, Burlingame, CA). Digital images were acquired by a confocal microscope (Leica, Exton, PA). Images were cropped, enlarged, and contrast-enhanced with Adobe Photoshop (Adobe Systems, San Jose, CA). Final resolution was ∼300 dots per inch.
Antibodies. Two antibodies for Gαo were used: rabbit polyclonal directed against the specific peptide ANNLRGCGLY located at the C terminus of the α subunit (gift from Dr. D. Manning, University of Pennsylvania, Philadelphia, PA) (Carlson et al., 1989; Law et al., 1991; Vardi et al., 1993) and a mouse monoclonal against the purified bovine protein (MAB 3071; Chemicon, Temecula, CA) (Li et al., 1995). Anti-mGluR6 was directed against the C terminus of rat mGluR6 (gift from Dr. S. Nakanishi, Kyoto University, Kyoto, Japan). Anti-recoverin was directed against bovine recoverin (gift from Dr. A. M. Dizhoor, University of Washington, Seattle, WA). Anti-calbindin D28 was a rabbit polyclonal (Swant, Bellinzona, Switzerland), and anti-protein kinase C (PKC) was a mouse monoclonal (Sigma, St. Louis, MO and Amersham Pharmacia Biotech, Little Chalfont, UK).
Electron microscopy. Eyes were fixed in phosphate buffer (0.12 m, pH 7.3) containing 2% glutaraldehyde, 2% paraformaldehyde, and 5% glucose. Eyes were rinsed in phosphate buffer, and then a small piece of retina was taken (∼1 × 1 mm2, with the optic disk in the center). Tissue was osmicated (1%, 1 hr), partially dehydrated in 50% and 70% ethanol, stained en bloc in 1% ethanolic uranyl acetate (1 hr), dehydrated, and embedded in Epon. Ultrathin sections (80–90 nm) were counterstained with uranyl acetate and lead citrate and photographed in an electron microscope. Semi-thin sections (1 μm) were stained with toluidine blue.
RESULTS
Gαo−/− retina appears normal
To test whether the absence of Gαo affects retinal development, we compared retinas from 3–8 postnatal week null mice with wild-type littermates. The null retinas were identical in thickness to wild-type retinas of the same age. Photoreceptors, including outer and inner segments, axon, soma, and terminal, appeared fully developed, and all layers were present at normal thickness (Fig.1).
Fig. 1.

Null retina has normal thickness.A, One micrometer radial Epon sections (stained with toluidine blue) show that all the layers in the Gαo−/− retina are present at normal morphology (6 postnatal weeks).B, Measurements of individual retinal layers show no difference between wild type and null (2 retinas at ∼4 postnatal weeks and 2 at 6 postnatal weeks; measured from Epon sections).OS, Outer segment; IS, inner segment;ONL, outer nuclear layer; OPL, outer plexiform layer; INL, inner nuclear layer;IPL, inner plexiform layer; GCL, ganglion cell layer; PR, photoreceptor outer and inner segments;WR, whole retina.
We probed the morphology of specific cell types by staining with established immunomarkers: anti-PKC for rod bipolar cells; anti-recoverin for OFF cone bipolar cells; and anti-calbindin for horizontal and amacrine cells. These cell types were indistinguishable in null and wild-type retinas (Fig. 2). Thus, rod bipolar somas were positioned normally, high in the inner nuclear layer (INL), with dendrites arborizing in the outer plexiform layer (OPL) and axons arborizing in the deepest stratum of the inner plexiform layer (IPL) (Fig. 2A,B). The OFF cone bipolar cell stained for recoverin (probably CB2) (Euler and Wässle, 1995) arborized diffusely in the OFF sublamina of the IPL (Fig. 2C,D). Horizontal cell processes invaginated rod and cone terminals, and amacrine processes formed three sharp bands in the IPL (Fig.2E,F). These experiments were performed on 3- and 7-week-old animals with same results.
Fig. 2.

Specific cell types in the null retina express standard chemical markers and normal morphology (confocal images; 3 postnatal weeks). A, B, Rod bipolar cells (RB) express PKC. C, D, OFF bipolar cells (probably type CB2) express recoverin.E, F, Horizontal (HC) and amacrine (AC) cells express calbindin. G,H, Wild-type retina expresses Gαo in ON bipolar cells (On-BP) and IPL, but null retina does not.bv, Blood vessel.
Wild-type retinas already expressed Gαo by 3 postnatal weeks, showing the adult pattern (Fig. 2G). Thus, immunostain was present in bipolar somas high in the INL, in bipolar dendrites, and in the IPL. Null retinas were immunonegative for Gαo (Fig. 2H). In short, except for the absence of Gαo, the null retina seemed completely normal.
Photoreceptor synaptic complex appears normal
Conceiveably, the absence of Gαo from the ON bipolar dendrite might affect the expression of mGluR6, or there might be trans-synaptic effects on the structure of the rod and cone terminal. If so, a primary defect of bipolar cell transduction would not be distinguishable from such second-order effects. However, both null and wild-type retinas stained identically for mGluR6; puncta were stained in OPL and somas in the upper tiers of the INL (Fig.3, left). Also, detailed examination by electron microscopy of the rod and cone terminals (tissue was examined in two wild types at 3.5 and 6 weeks old and two nulls at 4 and 6 weeks old) showed similar synaptic ribbons, synaptic vesicles, invaginating processes, “synaptic triad” (two horizontal cell processes flanking a central bipolar dendrite), electron-dense apposition between photoreceptor plasma membrane and bipolar dendrite, and “basal contact” between cone basal surface and bipolar dendrite (Fig. 3). Thus, in the null retina, all of the key ultrastructural features of the photoreceptor synaptic complex seemed normal.
Fig. 3.
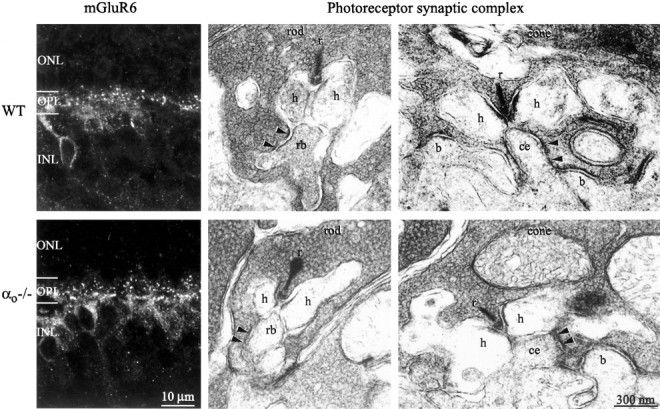
Synaptic apparatus of the null retina is normal.Left, Confocal image of immunostain for mGluR6. Stain concentrates in the outer plexiform layer in dendritic tips of ON bipolar cells and shows no difference between wild type and null (7 postnatal weeks). Middle, Electromicrographs of rod terminal. Right, Electromicrographs of cone terminals. Presynaptic elements: synaptic ribbon (r), synaptic vesicles, and dense photoreceptor membrane (arrowheads) are intact in the null retina. Postsynaptic elements: appear at their normal positions, lateral for horizontal cell processes (h), central for rod bipolar dendrite (rb) and ON cone bipolar dendrite (ce), and basal for OFF bipolar dendrites (b).
Gαo null mice lack the b-wave of the electroretinogram but retain the a-wave
Most of the components of the ERG can be recorded from a wild-type mouse 2 weeks postnatally, after which the ERG components rapidly reach mature proportions (see below). We recorded ERGs from postnatal day 21 (P21) to P60 mice.
A normal, dark-adapted mouse responds to a dim flash [510 nm; ∼20 photoisomerizations (R*) rod−1] with a “scotopic b-wave” (100–200 μV), generated by the transduction current of the depolarizing rod bipolar cells (Robson and Frishman, 1995). We observed the scotopic b-wave in both wild-type and heterozygous mice. However, in Gαo null mice, the same flash evoked no b-wave (Figs. 4A,5A). Of course, the b-wave would fail if the rods were themselves impaired, so we examined rod function by stimulating dark-adapted mice with an intense white flash that isomerized ∼1% of the rhodopsin. This strong stimulus causes rapid closure of the cGMP-activated channels and completely suppresses the rod circulating current. Suppression of the circulating current produces an ERG field potential, the a-wave, of saturating amplitude (Hagins et al., 1970; Hood and Birch, 1993; Breton et al., 1994;Lyubarsky and Pugh, 1996; Pugh et al., 1998). The saturating a-wave amplitudes in the wild-type, heterozygous, and null mice were indistinguishable (Figs. 4B, 5B). Thus, we conclude that the rods have normal cGMP-activated currents and normal phototransduction. Because rods are functional, the absence of the b-wave in the null mice in response to both the dim and bright flashes indicates failure of signal transduction in the rod bipolar cell.
Fig. 4.

Rod- and cone-driven b-waves are absent from the electroretinogram of the null mouse. A, Animals dark-adapted for 2 hr were stimulated with dim flashes. Such flashes elicited a rod-driven, corneal-positive b-wave in the heterozygotes but no positive-going responses in the null mice. The estimated flash intensities in photoisomerizations per rod (Φ) and the number of responses (n) averaged for eachtrace shown were as follows: for the P21 and P31 mice, Φ = 20, n = 11; for the P44 mouse, Φ = 3, n = 40. B, Dark-adapted animals were stimulated with an intense flash (isomerizing ∼1% of the rhodopsin). This elicited in the heterozygote a negative a-wave (shading), followed by a positive-going b-wave. In the null mice, the a-wave was normal, but the b-wave was absent. The flash intensities (Φ) and the number of responses (n) averaged were as follows: for the P21 and P31 mice, Φ = 106, n = 2–4. For the P44 mouse responses to three intensities are shown: Φ = 20,n = 20; Φ = 500, n = 16; Φ = 106, n = 2.C, Mice were adapted to a bright background (540 nm; 20,000 R* rod−1s−1) that completely suppressed the cGMP-activated current of the rods. They were then stimulated with an intense white flash that isomerizes ∼1% of the M-cone pigment and 0.1% of the UV-cone pigment in adult mice. The cone-driven a-wave was not visible in the P21 animals but was pronounced in P31 and P44 animals (both Gαo+/− and Gαo−/−). A typical cone-driven b-wave (positive-going response with superimposed oscillations, peaking ∼70–90 msec after the flash) was observed in the Gαo+/− mice of all age groups but was absent in the Gαo −/− mice. For P21 and P31 Gαo+/− mice, n = 10; for P21 Gαo−/−,n = 20; for P31 and P44 Gαo−/−,n = 40. The slow positive-going potential in P21 mouse is probably an artifact attributable to movement of the lightly anesthetized mouse.
Fig. 5.

ON components of the ERG remain absent from Gαo−/− mice during development. The peak amplitude of the ERG a- and b-waves is variable because of variable contact with the electrode and because of rapid growth between P21 and P30. However, at all ages, the rod- and cone-driven b-waves were missing from the Gαo−/− mouse.
Mouse retina contains a substantial population of ON bipolar cells driven by cones (Jeon et al., 1998). Because dendrites of these cells express Go, it was of interest to examine cone-driven responses. To isolate the cone-driven components of the ERG, we presented a bright steady background (540 nm; ∼20,000R* rod−1s−1), which completely suppresses the rod circulating current and permits measurement of purely cone-driven a- and b-waves (Pugh et al., 1998; Lyubarsky et al., 1999, 2000). The wild-type and heterozygous mice exhibited robust cone b-waves, but the null showed no reliable positive-going responses (Figs. 4C,5C). All mice groups showed normal cone a-waves (Figs.4C, 5D). Previous work has attributed the cone a-wave mostly to the suppression of the cone circulating current; however, it may also contain a component contributed by the activation of cone-driven OFF bipolar cells.
Because the Gαo−/− mice have a low survival rate, we took most of our data points from the early age of 21–30 d. However, to test whether the lack of a b-wave might be attributable to a lag in development, we also tested older mice of up to 60 d. In the wild type, all of the ERG components grew rapidly between P21 and P25. In the null mouse, the rod- and cone-driven a-waves were similar to those of the wild type, but the b-waves were always missing (Fig. 5).
DISCUSSION
Gαo is necessary for the ON bipolar response
A mouse lacking the α subunit of Go fails to produce either a scotopic or a photopic b-wave. The b-wave is elicited when the glutamate release from a photoreceptor is suppressed by light increment (Pugh et al., 1998; Green and Kapousta-Bruneau, 1999; Robson and Frishman, 1999; Shiells and Falk, 1999). This opens cation channels in the ON bipolar dendrite that had been closed in darkness by tonic activation of the mGluR6 cascade (Stockton and Slaughter, 1989; Nawy and Jahr, 1990; Shiells and Falk, 1990; Yamashita and Wässle, 1991; de la Villa et al., 1995; Euler et al., 1996). Thus, absence of the b-wave reflects loss of function in ON bipolar cells. Consistent with this, a mouse lacking the mGluR6 receptor also lacks the b-wave (Masu et al., 1995).
Failure of the ON response in the Gαo−/− mouse cannot be attributed to a gross deficit in retinal organization: (1) rods and cones respond normally (a-wave); (2) rod and cone synaptic terminals show normal ultrastructure (ribbons, vesicles, triad, etc.); (3) mGluR6 is expressed at the ON bipolar dendritic tip; and (4) ON bipolar cells show normal morphology and spatial density. It is possible that lack of Gαo affects expression of other proteins whose importance is currently unknown. However, it is more likely that the b-wave is impaired because Go crucially links mGluR6 to an effector that closes the cation channel.
This conclusion is further supported by a considerable body of circumstantial evidence: (1) Gαo colocalizes with mGluR6 in the dendritic tips of all ON bipolar types (Vardi et al., 1993, 2000; Vardi and Morigiwa, 1997; Vardi, 1998); (2) purified mGluR6 stimulated by its specific agonist,l-2-amino-4-phosphonobutyrate (l-AP-4) activates Gαo 18-fold more strongly than Gt (transducin) (Weng et al., 1998); (3)l-AP-4 in a retinal homogenate suppresses ADP-ribosylation of a G-protein by pertussis toxin but not cholera toxin (Kikkawa et al., 1993), consistent with the sensitivity of Goto pertussis but not cholera toxin (Gilman, 1987); and (4) dialysis of Gαo or antibody to Gαointo ON bipolar cells reduces the response to glutamate (Nawy, 1999).
Gαo-mediated cascades are poorly understood
Go is the most abundant G-protein in the brain (Sternweis and Robishaw, 1984; Huff et al., 1985; Terashima et al., 1987; Asano et al., 1988; Li et al., 2000), and it appears to be involved in a large repertoire of signal transduction cascades. For example, Go regulates several types of voltage-gated Ca2+ channels (Kleuss et al., 1991; Valenzuela et al., 1997; Jiang et al., 1998), K+ channels (VanDongen et al., 1988), a cGMP-dependent channel (in scallop ciliary photoreceptors; Kojima et al., 1997), possibly a cAMP-dependent channel (in vomeronasal organ;Berghard and Buck, 1996), and a variety of serotonin-controlled behaviors in Caenorhabditis elegans (Mendel et al., 1995;Ségalat et al., 1995; Nurrish et al., 1999). In cases where Go gates the N-type Ca2+ channel, the Gβγ of Go complex directly binds to the α subunit of the channel (Dolphin, 1998), but this cannot account for the diverse effects of Go, so there are probably other mechanisms.
Initially the G-protein of the ON bipolar cell was thought to activate a cGMP phosphodiesterase (Nawy and Jahr, 1990; Shiells and Falk, 1990). However, recent experiments using a nonhydrolyzable analog of cGMP, or various concentrations of glutamate plus phosphodiesterase inhibitor, failed to prevent glutamate from closing the channel. This suggested that the channel might be gated by something other than cGMP (Nawy, 1999). Conceivably, because the βγ subunits of Go can interact directly with Q- and N-type voltage-gated calcium channels, these subunits could also act on the ON bipolar cell cation channel. However, to the contrary, dialyzing the α subunit of Go into the ON bipolar cell mimics the effect of activated mGluR6, i.e., the cell hyperpolarizes (Nawy, 1999). If the Gβγ subunits served as linkers, dialyzing Gαo should have bound the Gβγ subunits, prevented their activation, and thus depolarized the cell. In contrast, if the dialyzed free Gαo subunit were activated by GTP, it would mimic mGluR6. These experiments seem to rule out the βγ subunits and support a role for Gαo.
The targets of Gαo remain obscure, and this seems astonishing given that Go is the most abundant protein in the brain. However, the present study establishing a firm requirement for Go for signal transmission at a specific synapse may be an important step toward identifying the downstream targets in ON bipolar cells and in other parts of the nervous system.
Footnotes
This work was supported by National Eye Institute Grants EY11105, EY00828, EY02660, and DK19318. We thank Eva Neer for providing us with the initial pairs of Gαo+/− mice; Dave Manning, Alexander Dizhoor, and Shigetada Nakanishi for providing antibodies; Yi-Jun Shi, Sally Shrom, and Jian Li for technical assistance; and Sharron Fina for preparing this manuscript.
Correspondence should be addressed to Dr. Noga Vardi, Department of Neuroscience, University of Pennsylvania, Philadelphia, PA 19104. E-mail: noga@retina.anatomy.upenn.edu.
REFERENCES
- 1.Asano T, Semba R, Kamiya N, Ogasawara N, Kato K. Go, a GTP-binding protein: immunochemical and immunohistochemical localization in the rat. J Neurochem. 1988;50:1164–1169. doi: 10.1111/j.1471-4159.1988.tb10588.x. [DOI] [PubMed] [Google Scholar]
- 2.Berghard A, Buck LB. Sensory transduction in vomeronasal neurons: evidence for Gαo, Gαi2, and adenylyl cyclase II as major components of a pheromone signaling cascade. J Neurosci. 1996;16:909–918. doi: 10.1523/JNEUROSCI.16-03-00909.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Breton ME, Schueller AW, Lamb TD, Pugh EN., Jr Analysis of ERG a-wave amplification and kinetics in terms of the G-protein cascade of phototransduction. Invest Ophthalmol Vis Sci. 1994;35:295–309. [PubMed] [Google Scholar]
- 4.Carlson KE, Brass LF, Manning DR. Thrombin and phorbol esters cause the selective phosphorylation of a G-protein other than Gi in human platelets. J Biol Chem. 1989;264:13298–13305. [PubMed] [Google Scholar]
- 5.Cohen E, Sterling P. Convergence and divergence of cones onto bipolar cells in the central area of cat retina. Philos Trans R Soc Lond B Biol Sci. 1990;330:323–328. doi: 10.1098/rstb.1990.0202. [DOI] [PubMed] [Google Scholar]
- 6.de la Villa P, Kurahashi T, Kaneko A. l-glutamate-induced responses and cGMP-activated channels in three subtypes of retinal bipolar cells dissociated from the cat. J Neurosci. 1995;15:3571–3582. doi: 10.1523/JNEUROSCI.15-05-03571.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Dolphin AC. Mechanisms of modulation of voltage-dependent calcium channels by G-proteins. J Physiol (Lond) 1998;506:3–11. doi: 10.1111/j.1469-7793.1998.003bx.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Euler T, Wässle H. Immunocytochemical identification of cone bipolar cells in the rat retina. J Comp Neurol. 1995;361:461–478. doi: 10.1002/cne.903610310. [DOI] [PubMed] [Google Scholar]
- 9.Euler T, Schneider H, Wässle H. Glutamate responses of bipolar cells in a slice preparation of the rat retina. J Neurosci. 1996;16:2934–2944. doi: 10.1523/JNEUROSCI.16-09-02934.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Gilman AG. G proteins: transducers of receptor-generated signals. Annu Rev Biochem. 1987;56:615–649. doi: 10.1146/annurev.bi.56.070187.003151. [DOI] [PubMed] [Google Scholar]
- 11.Green DG, Kapousta-Bruneau NV. A dissection of the electroretinogram from the isolated rat retina with microelectrodes and drugs. Vis Neurosci. 1999;16:727–741. doi: 10.1017/s0952523899164125. [DOI] [PubMed] [Google Scholar]
- 12.Greif GJ, Sodickson DL, Bean BP, Neer EJ, Mende U. Altered regulation of potassium and calcium channels by GABA(B) and adenosine receptors in hippocampal neurons from mice lacking Galpha(o). J Neurophysiol. 2000;83:1010–1018. doi: 10.1152/jn.2000.83.2.1010. [DOI] [PubMed] [Google Scholar]
- 13.Hagins WA, Penn RD, Yoshikami Y. Dark current and photocurrent in retinal rods. Biophys J. 1970;10:380–412. doi: 10.1016/S0006-3495(70)86308-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Hood DC, Birch DG. Human cone receptor activity: the leading edge of the a-wave and models of receptor activity. Vis Neurosci. 1993;10:857–871. doi: 10.1017/s0952523800006076. [DOI] [PubMed] [Google Scholar]
- 15.Huff RM, Axton JM, Neer EJ. Physical and immunological characterization of a guanine nucleotide-binding protein purified from bovine cerebral cortex. J Biol Chem. 1985;260:10864–10871. [PubMed] [Google Scholar]
- 16.Jeon C-J, Strettoi E, Masland RH. The major cell populations of the mouse retina. J Neurosci. 1998;18:8936–8946. doi: 10.1523/JNEUROSCI.18-21-08936.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Jiang M, Gold MS, Boulay G, Spicher K, Peyton M, Brabet P, Srinivasan Y, Rudolph U, Ellison G, Birnbaumer L. Multiple neurological abnormalities in mice deficient in the G protein Go. Proc Natl Acad Sci USA. 1998;95:3269–3274. doi: 10.1073/pnas.95.6.3269. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Kikkawa S, Nakagawa M, Iwasa T, Kaneko A, Tusda M. GTP-binding protein couples with metabotropic glutamate receptor in bovine retinal ON-bipolar cell. Biochem Biophys Res Commun. 1993;195:374–379. doi: 10.1006/bbrc.1993.2053. [DOI] [PubMed] [Google Scholar]
- 19.Kleuss C, Hescheler J, Ewel C, Rosenthal W, Schultz G, Wittig B. Assignment of G-protein subtypes to specific receptors inducing inhibition of calcium currents. Nature. 1991;353:43–48. doi: 10.1038/353043a0. [DOI] [PubMed] [Google Scholar]
- 20.Kojima D, Terakita A, Ishikawa T, Tsukahara Y, Maeda A, Shichida Y. A novel Go-mediated phototransduction cascade in scallop visual cells. J Biol Chem. 1997;272:22979–22982. doi: 10.1074/jbc.272.37.22979. [DOI] [PubMed] [Google Scholar]
- 21.Law SF, Manning D, Reisine T. Identification of subunits of GTP binding proteins coupled to somatostatin receptors. J Biol Chem. 1991;266:17885–17897. [PubMed] [Google Scholar]
- 22.Li X, Mumby S, Greenwood A, Jope R. Pertussis toxin-sensitive G-protein α-subunits: production of monoclonal antibodies and detection of differential increases upon differentiation of PC12 and LA-N-5 cells. J Neurochem. 1995;64:1107–1117. doi: 10.1046/j.1471-4159.1995.64031107.x. [DOI] [PubMed] [Google Scholar]
- 23.Li Y, Mende U, Lewis C, Neer EJ. Maintenance of cellular levels of G-proteins: different efficiencies of alpha s and alpha o synthesis in GH3 cells. Biochem J. 2000;318:1071–1077. doi: 10.1042/bj3181071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Lyubarsky AL, Pugh EN., Jr Recovery phase of the murine rod photoresponse reconstructed from electroretinographic recordings. J Neurosci. 1996;16:563–571. doi: 10.1523/JNEUROSCI.16-02-00563.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Lyubarsky AL, Falsini B, Pennesi ME, Valentini P, Pugh EN., Jr UV- and midwave-sensitive cone-driven retinal responses of the mouse: a possible phenotype for coexpression of cone photopigments. J Neurosci. 1999;19:442–455. doi: 10.1523/JNEUROSCI.19-01-00442.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Lyubarsky AL, Chen C-K, Simon MI, Pugh EN., Jr Mice lacking G-protein receptor kinase 1 have profoundly slowed recovery of cone-driven retinal responses. J Neurosci. 2000;20:2209–2217. doi: 10.1523/JNEUROSCI.20-06-02209.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Martin PR, Grünert U. Spatial density and immunoreactivity of bipolar cells in the Macaque monkey retina. J Comp Neurol. 1992;323:269–287. doi: 10.1002/cne.903230210. [DOI] [PubMed] [Google Scholar]
- 28.Masu M, Iwakabe H, Tagawa Y, Miyoshi T, Yamashita M, Fukuda Y, Sasaki H, Hiroi K, Nakamura Y, Shigemoto R. Specific deficit on the ON response in visual transmission by targeted disruption of the mGluR6 gene. Cell. 1995;80:757–765. doi: 10.1016/0092-8674(95)90354-2. [DOI] [PubMed] [Google Scholar]
- 29.Mendel JE, Korswagen HC, Liu KS, Hajdu-Cronin YM, Simon MI, Plasterk RHA, Sternberg PW. Participation of the protein Go in multiple aspects of behavior in C. elegans. Science. 1995;267:1652–1655. doi: 10.1126/science.7886455. [DOI] [PubMed] [Google Scholar]
- 30.Nakajima Y, Iwakabe H, Akazawa C, Nawa H, Shigemoto R, Mizuno N, Nakanishi S. Molecular characterization of a novel retinal metabotropic glutamate receptor mGluR6 with a high agonist selectivity for l-2-amino-4-phosphonobutyrate. J Biol Chem. 1993;268:11868–11873. [PubMed] [Google Scholar]
- 31.Nawy S. The metabotropic receptor mGluR6 may signal through Go, but not phosphodiesterase, in retinal bipolar cells. J Neurosci. 1999;19:2938–2944. doi: 10.1523/JNEUROSCI.19-08-02938.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Nawy S, Jahr CE. Suppression by glutamate of cGMP-activated conductance in retinal bipolar cells. Nature. 1990;346:269–271. doi: 10.1038/346269a0. [DOI] [PubMed] [Google Scholar]
- 33.Nomura A, Shigemoto R, Nakamura Y, Okamoto N, Mizuno N, Nakanishi S. Developmentally-regulated postsynaptic localization of a metabotropic glutamate-receptor in rat rod bipolar cells. Cell. 1994;77:361–369. doi: 10.1016/0092-8674(94)90151-1. [DOI] [PubMed] [Google Scholar]
- 34.Nurrish S, Ségalat L, Kaplan JM. Serotonin inhibition of synaptic transmission: Gαo decreases the abundance of UNC-13 at release sites. Neuron. 1999;24:231–242. doi: 10.1016/s0896-6273(00)80835-1. [DOI] [PubMed] [Google Scholar]
- 35.Pugh EN, Jr, Falsini B, Lyubarsky A. The origin of the major rod- and cone-driven components of the rodent electroretinogram and the effect of age and light-rearing history on the magnitude of these components. In: Williams T, Thistle A, editors. Photostasis and related phenomena. Plenum; New York: 1998. pp. 93–128. [Google Scholar]
- 36.Robson JG, Frishman LJ. Response linearity and kinetics of the cat retina: the bipolar cell component of the dark-adapted electroretinogram. Vis Neurosci. 1995;12:837–850. doi: 10.1017/s0952523800009408. [DOI] [PubMed] [Google Scholar]
- 37.Robson JG, Frishman LJ. Dissecting the dark-adapted electroretinogram. Doc Ophthalmol. 1999;95:187–215. doi: 10.1023/a:1001891904176. [DOI] [PubMed] [Google Scholar]
- 38.Ségalat L, Elkes DA, Kaplan JM. Modulation of serotonin-controlled behaviors by Go in Caenorhabditis elegans. Science. 1995;267:1648–1651. doi: 10.1126/science.7886454. [DOI] [PubMed] [Google Scholar]
- 39.Shiells RA, Falk G. Glutamate receptors of rod bipolar cells are linked to a cyclic GMP cascade via a G-protein. Proc R Soc Lond B Biol Sci. 1990;242:91–94. doi: 10.1098/rspb.1990.0109. [DOI] [PubMed] [Google Scholar]
- 40.Shiells RA, Falk G. Contribution of rod, on-bipolar, and horizontal cell light responses to the ERG of dogfish retina. Vis Neurosci. 1999;16:503–511. doi: 10.1017/s0952523899163119. [DOI] [PubMed] [Google Scholar]
- 41.Sterling P, Freed MA, Smith RG. Architecture of the rod and cone circuits to the On-beta ganglion cell. J Neurosci. 1988;8:623–642. doi: 10.1523/JNEUROSCI.08-02-00623.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Sternweis PC, Robishaw JD. Isolation of two proteins with high affinity for guanine nucleotides from membranes of bovine brain. J Biol Chem. 1984;259:13806–13813. [PubMed] [Google Scholar]
- 43.Stockton RA, Slaughter MM. B-wave of the electroretinogram. A reflection of ON bipolar cell activity. J Gen Physiol. 1989;93:101–122. doi: 10.1085/jgp.93.1.101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Strettoi E, Masland R. The organization of the inner nuclear layer of the rabbit retina. J Neurosci. 1995;15:875–888. doi: 10.1523/JNEUROSCI.15-01-00875.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Terashima T, Katada T, Okada E, Ui M, Inoue Y. Light microscopy of GTP-binding protein (Go) immunoreactivity within the retina of different vertebrates. Brain Res. 1987;436:384–389. doi: 10.1016/0006-8993(87)91685-4. [DOI] [PubMed] [Google Scholar]
- 46.Valenzuela D, Han X, Mende U, Fankhauser C, Mashimo H, Huang P, Pfeffer J, Neer EJ, Fishman MC. G alphao is necessary for muscarinic regulation of Ca2+ channels in mouse heart. Proc Natl Acad Sci USA. 1997;94:1727–1732. doi: 10.1073/pnas.94.5.1727. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.VanDongen AM, Codina J, Olate J, Mattera R, Joho R, Birnbaumer L, Brown AM. Newly identified brain potassium channels gated by the guanine nucleotide binding protein Go. Science. 1988;242:1433–1437. doi: 10.1126/science.3144040. [DOI] [PubMed] [Google Scholar]
- 48.Vardi N. Alpha subunit of Go localizes in the dendritic tips of ON bipolar cells. J Comp Neurol. 1998;395:43–52. [PubMed] [Google Scholar]
- 49.Vardi N, Morigiwa K. ON cone bipolar cells in rat express the metabotropic receptor mGluR6. Vis Neurosci. 1997;14:789–794. doi: 10.1017/s0952523800012736. [DOI] [PubMed] [Google Scholar]
- 50.Vardi N, Matesic DF, Manning DR, Liebman PA, Sterling P. Identification of a G-protein in depolarizing rod bipolar cells. Vis Neurosci. 1993;10:473–478. doi: 10.1017/s0952523800004697. [DOI] [PubMed] [Google Scholar]
- 51.Vardi N, Duvoisin RM, Wu G, Sterling P. Localization of mGluR6 to dendrites of ON bipolar cells in primate retina. J Comp Neurol. 2000;423:402–412. doi: 10.1002/1096-9861(20000731)423:3<402::aid-cne4>3.0.co;2-e. [DOI] [PubMed] [Google Scholar]
- 52.Wässle H, Grünert U, Cook NJ, Molday RS. The cGMP-gated channel of rod outer segments is not localized in bipolar cells of the mammalian retina. Neurosci Lett. 1992;134:199–202. doi: 10.1016/0304-3940(92)90516-a. [DOI] [PubMed] [Google Scholar]
- 53.Weng K, Lu C-C, Daggett LP, Kuhn R, Flor PJ, Johnson ED, Robinson PR. Functional coupling of a human retinal metabotropic glutamate receptor (hmGluR6) to bovine rod transducin and rat Go in an in vitro reconstruction system. J Biol Chem. 1998;272:33100–33104. doi: 10.1074/jbc.272.52.33100. [DOI] [PubMed] [Google Scholar]
- 54.Yamashita M, Wässle H. Responses of rod bipolar cells isolated from the rat retina to the glutamate agonist 2-amino-4-phosphonobutyric acid (APB). J Neurosci. 1991;11:2372–2382. doi: 10.1523/JNEUROSCI.11-08-02372.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]