

Abstract
Background:
Age-related cognitive decline has been suggested to result from an increase in the brain neuron loss, which is attributable to continued derangement of the brain’s oxidant/antioxidant balance. Increased oxidative stress and a concomitant decrease in the brain’s antioxidant defense system have been associated with functional senescence and organismal ageing. However, nature has configured certain foods to be rich sources of nootropic agents, with research showing that increased consumption of such foods or food ingredients may be protective against ageing-related memory decline. This knowledge is becoming increasingly valuable in an era when the boundary that separates food from medicine is becoming blurred. In this review, we examine extant literature dealing with the impact of ageing on brain structure and function, with an emphasis on the roles of oxidative stress. Secondly, we review the benefits of food-based antioxidants with nootropic effects and/or food-based nootropic agents in mitigating memory decline; with a view to improving our understanding of likely mechanisms. We also highlight some of the limitations to the use of food-based nootropics and suggest ways in which they can be better employed in the clinical management of age-related cognitive decline.
Conclusion:
While it is known that the human brain endures diverse insults in the process of ageing, food-based nootropics are likely to go a long way in mitigating the impacts of these insults. Further research is needed before we reach a point where food-based nootropics are routinely prescribed.
Keywords: Ageing, antioxidants, dementia, nutrition, nutraceuticals, nootropic
1. INTRODUCTION
Over the past few decades, available data suggest a global increase in life- expectancy with the global average life-expectancy increasing by 5.5 years between the year 2000 and 2016 [1]. However, with an increase in the ageing population comes an associated increase in the economic, social, and health burden of age-related disorders and diseases [2]. These include cardiovascular diseases, cancer, inflammatory diseases and neurodegenerative disorders [3]. Senescence, programmed/non-programmed cell death, oxidative stress, and immune/endocrine system dysfunction are all factors that have been implicated in the development of age-related disorders and also linked to a number of ageing theories [4-8]. Cognitive decline is a worrisome consequence of normal or pathologic ageing, having a high personal, economic and societal burden; also, it could herald the onset of dementia which is associated with significant morbidity and mortality [9, 10]. Also, in recent times, the use of non-invasive techniques such as transcranial magnetic stimulation has proven to be beneficial in the early diagnosis of dementia, and for improving our understanding of the electrophysiological and neurochemical mechanisms that underlie the transition from normal brain ageing to a degenerative and vascular-related cognitive decline [11].
However, with a general increase in life expectancy and a corresponding increase in the prevalence of age-related cognitive impairment, there have been concerted efforts towards the development and adoption of preventive strategies that would minimise the risk of developing dementia, or reduce the rate of cognitive decline with ageing. In the last few decades, certain compounds have been found useful in the management of cognitive decline. The term ‘nootropic’ has been used to define such substances with the capacity to enhance cognition. Nootropic drugs can enhance cognition through their effects on the brain dopaminergic (amphetamine, armodafinil, modafinil), glutamatergic/cholinergic (piracetam, oxiracetam, aniracetam) and serotonergic (5-hydroxy-tryptophan) systems. Also, in recent times, there have been suggestions of associations between cognition and diet, with reports that poor nutrition could hasten the cognitive decline [12-15]. While the beneficial effects of synthetic nootropic agents like piracetam [16, 17] and related compounds [18, 19] in age-related cognitive decline have been demonstrated, the cost and side-effects of these drugs may limit their sustained use in clinical practice. Also, there are indications that they may not directly influence the pathologic processes that drive cognitive decline. Hence, they might only succeed in alleviating the symptoms.
Currently, the impacts of food-based nootropic agents are being evaluated, with a view to improving their quality/delivery (e.g nanoencapsulation and nanosuspensions) [20], or developing a drug which is a hybrid of synthetic and naturally-occurring nootropic agents, with possibly fewer side-effects, lesser cost, and higher efficacy [20, 21]. Also, the development and clinical adoption of a regimen consisting solely of food-based agents that mitigate or possibly reverse cognitive decline is worthy of consideration because it offers more options to patients who may be of poor socioeconomic backgrounds, or are residents of poorer nations.
In this review, we examine extant literature dealing with the topic of food-based agents with experimental or clinically- reported possible nootropic effects. This is with a view to improving our understanding of the different mechanisms by which they may improve memory function, and suggest ways in which they can be better used in the clinical management of age-related cognitive decline.
1.1. The Ageing Brain
Ageing has been described as a physiological process involving all body tissues. Over the years, a constant question that has continued to be raised by neuroscientists has been, at what age does the brain begin to age? While our perspectives of what can be considered normal ageing have continued to evolve over time, interventions are usually targeted at adults who are 60 years of age and older [22]. However, data from recent studies have suggested that contrary to previously-held beliefs that brain ageing does not begin until after middle age (45 years and above), a gradual deterioration of cognitive and intellectual ability (first observed as a cognitive-motor decline in complex tasks) begins at about the age of 24 years [23].
In the human brain, the effects of ageing have been studied extensively, with suggestions that ageing alters the brain‘s cellular, vascular and structural characteristics [24, 25], resulting in a decline in cognitive function, attention, and memory; and a delay in cognitive information processing. Structurally, brain volume and/or brain weight have been shown to decrease at a rate of about 5% per decade beginning from the age of 40 years [26]. Research also suggests an acceleration of the actual rate of brain atrophy with increasing age, especially from the age of 70 years and above [27]. Apart from the reduction in brain volume, there is also a decrease in neuronal volume and neuronal metabolic activity [24, 25].
Ageing has also been considered an important risk factor for the development of Alzheimer’s Disease (AD), and studies examining the differences between brain changes in physiological and pathological (patients with AD) ageing have been used to understand the effects of pathological ageing on the brain electrophysiology, morphology and neurochemistry. Pathological brain ageing is exemplified by conditions such as AD and vascular dementia whose electrophysiological platform is related to morphological changes such as progressive neuronal loss and ischaemic disruption, respectively. However, despite disease progression, an accompanying cortical/sub-cortical neuroplastic change might allow the preservation of motor programming and execution [28]. The disruption and loss of synaptic connectivity result in progressive cognitive decline due to white-matter abnormalities and anatomical/functional deficits [29]. Also associated with pathological ageing is an increase in the distribution and magnitude of extracellular deposition of Amyloid beta (Aβ) peptides and the intracellular aggregation of tau protein filaments [30].
Generally, studies have shown that a number of the structural and functional changes observed in the ageing brain can be attributed to increased brain oxidative stress. There have been suggestions that a progressive imbalance occurs between the intracellular concentration of reactive oxygen species and the brain’s antioxidant defense system [30]. Increased brain oxidative stress results not only in neuronal injury but also in the alteration of redox-sensitive signalling processes [30]. There have been suggestions that increased oxidative stress and decreased antioxidant defense system are important factors in functional senescence and organismal ageing [31]. Also, there are reports suggesting a decline in the normal antioxidant defense system, resulting in an increase in the vulnerability of the brain to the effects of oxidative stress. A number of preclinical [31-33] and clinical [34, 35] studies have evaluated the effects of oxidative stress on brain ageing, age-related memory decline, and Alzheimer’s disease associated with memory impairment. Haddadi et al. [31], using the Drosophila model reported an evidence of age-related decline in short-term and long-term consolidated forms of olfactory memory in old flies. In addition to morphological evidence demonstrating a decrease in the number of synaptic/architectural mitochondrial defects and an increase in apoptotic neurones in Kenyon cells; there was also a biochemical evidence of a decrease in the activity of antioxidant enzymes, and an increase in the level of reactive oxygen species and lipid peroxidation in the brain [31]. Fukui et al. [32, 33] reported an evidence of neuronal apoptosis in the hippocampus, and an immunohistochemical//biochemical evidence of oxidative stress including increased lipid peroxidation in the synaptic plasma membranes in aged rats [32, 33].
In healthy humans, the impact of increased oxidative stress on the development of age-related memory decline has also been studied. The results of a longitudinal cohort study in a healthy population of University staff revealed that increased oxidative stress was associated with a decline in the executive function [35]. Finally, when immunohistochemical methods and two-dimensional fingerprinting of oxidatively modified proteins were used to examine the impact of oxidative stress on the brain in pathological brain ageing (Alzheimer’s disease); results revealed increased protein oxidation in the hippocampus, with superior and middle temporal gyri, but not in the cerebellum [34].
1.2. Cognition and Ageing
There are reports suggesting that cognitive impairment is fast becoming a public health problem worldwide [36, 37], with projections suggesting that the prevalence will double every 20 years. This would correspond to about 65.7 million individuals diagnosed and living with dementia in the year 2030, and over 115.4 million persons worldwide by the year 2050 [38]. The World Health Organisation data for 2017 show that over 50 million people are presently living with dementia worldwide, with about 10 million new cases yearly [39]. Apart from the increasing prevalence of persons living with dementia, there are also large numbers of elderly people who have other variants of cognitive impairment. Conditions such as Cognitive Impairment with No Dementia (CIND) and Mild Cognitive Impairment (MCI) make up the grey zone between memory loss that occurs with normal ageing and dementia [40, 41].
It is generally accepted that a decline in some aspects of cognition occurs as part of physiological ageing [42]. The most commonly observed cognitive change associated with ageing is memory impairment (a phenomenon described as age-related memory impairment). Generally, memory function Fig. (1) can be classified into sensory, short-term or working memory, and long-term memory which could be explicit (declarative) memory or implicit (procedural memory). Declarative memory can be further divided into semantic and episodic memory, [43, 44] with semantic and episodic memory being of significance with regard to ageing. Semantic memory has been defined as that segment of long-term memory that processes concepts and ideas not related to personal experiences but rather includes pieces of information that are acquired over a lifetime such as names of different colours, names of countries and their capitals. However, episodic memory is memory that is specific to the individual, involving the recollection of biographical experiences as well as specific events that are personal to the subject. Examples of episodic memory would include recalling the first day of school or one’s wedding day. Procedural memory is defined as memory responsible for understanding how to do things; it is important in skills such as riding a bike. Memory is generally created and stored in all regions of the brain; however, some regions have been associated with specific memory domains. The temporal lobe is associated with sensory memory, while the frontal lobe is associated with both short- and long-term memory. Specifically, declarative memory is believed to depend primarily on medial temporal lobe structures, such as the hippocampus; however, non-declarative memory has been associated with brain regions such as the striatum, cerebellum, and cortical association areas [45].
Fig. (1).
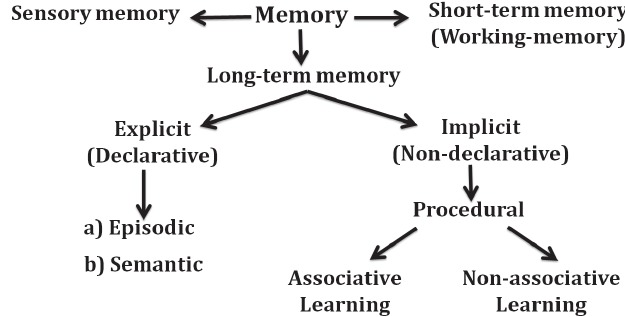
Classification of memory.
There are reports suggesting that different memory subtypes are affected by ageing. Episodic memory performances have been shown to decreases linearly with normal ageing [46-48]. Although conclusions arising from the results of longitudinal studies also suggest a relatively-stable episodic memory performance up to the middle age followed by a sharp decline thereafter [48]. Certain aspects of short-term, semantic and procedural memories have been suggested to have a fairly-constant performance level across the adult life span. There are reports that procedural memory is relatively stable with ageing [49] with little or no decline observed with ageing; although some cases of task-specific incidences of age-related decline in short-term memory have been reported [46]. Studies have reported reductions in executive functioning [50, 51] and mental speed [52], while semantic memory subtypes like naming category and verbal abilities are believed to improve with age [53, 54], although a decline is observed amongst the very old [55]. In pathological ageing (e.g. Alzheimer’s dementia), memory impairment initially affects short term memory; however, with the progression of disease, loss of declarative memory occurs with both episodic and semantic memory deficits observed in severe cases [56].
While there have been several reports suggesting age-dependent differences in cognitive functioning, considerable controversies still exist [22]. Although the specific changes associated with increasing age are still being examined, disputes regarding the reliability of longitudinal changes in older adults as being a true reflection of a continuum of alterations with onset in young adulthood or in the middle age still exist [44]. A diagnosis of dementia is necessitated by the presence of memory decline, loss of attention, and inability to perform activities of daily living [57]. MCI which is a transitional stage prior to the development of Alzheimer’s dementia (reported to primarily affect subjects 65 years and older) is characterised by the presence of mild to moderate memory loss and is associated with a risk of developing Alzheimer’s dementia [57].
2. FOOD AND BRAIN HEALTH
It is currently believed that the globally increased prevalence of dementia is mainly driven by population ageing, which would naturally increase the number of elderly people at risk of dementia [58-60]. In recent times, there have been suggestions that a reduction in the age-specific incidence of dementia could be driven by the modifications of exposures to presumed developmental, lifestyle and cardiovascular risk factors for the development of dementia [61]. Included in these lifestyle changes is dietary modification. The importance of diet in neurological development has been well-documented.
It had also been documented that dietary components such as gluten can trigger an immune response which may be associated with neurobehavioural or neuropsychiatric manifestation; also, gluten intolerance has been associated with changes in some transcranial magnetic stimulation measures of excitation and inhibition [62]. However, the potential role of gluten restriction in the management or prevention of subclinical neurological abnormalities, including “brain fog” that may be associated with dementia requires more investigation.
A number of studies have also demonstrated the beneficial effects of nutritive compounds on brain function. Some studies have shown that modifying lifestyle factors like diet may not only prevent cognitive decline that is associated with dementia but can also reduce the rate of decline in cognition that occurs with physiological ageing [63]. There have also been suggestions that a number of nutrients (that either occur naturally in food or are made available as dietary supplements) improve brain health and mental function via their effects on brain synaptic plasticity, neuronal function, and cognition [64, 65]. A number of studies have examined the effects of different dietary patterns on the age-related cognitive decline [14, 15]. The results generally point to the fact that an increase in the intake of fruits, vegetables, fish, nuts, and legumes (as observed with the Mediterranean diet, and a reduction in the intake of meat and high-fat dairy products (present in Western diets) is seemingly associated with better cognitive function and/or a reduced risk of developing dementia in later life [12, 66-69].
Some observational and interventional studies have also demonstrated that the diets which prevent or mitigate age-related cognitive decline contain high quantities of vitamins [70-74], omega-3 fatty acids [75, 76] and antioxidants [77]. These constituents might help to improve memory function by reducing inflammatory processes, amyloid deposition and oxidative stress within the brain, in addition to reducing the impact of cardiovascular morbidity. There have been reports that some of these foods may also contain natural nootropic factors like gingko biloba that directly improves cognition via its impact on specific brain regions including parietal, temporal, frontal, occipital lobes, and the hippocampus [78-81]. In rats, gingko biloba has been shown to increase brain acetylcholine levels, as well as the density of muscarinic cholinergic receptors in the hippocampus [79, 80].
2.1. Food-based Antioxidants and Diets with Nootropic Effects
In recent times, a number of studies have demonstrated the importance of antioxidants and vitamin supplements in slowing down the ageing process and stemming the progress of neurodegenerative disorders including Parkinson’s disease and Alzheimer’s disease [82]. There have also been suggestions that appropriate selection of whole foods or the inclusion of antioxidants in diet could be beneficial in increasing lifespan and mitigating age-related memory decline. The mechanisms involved in age-related memory decline continue to be studied. However, while the specific factors responsible for the structural and functional neuronal loss observed in normal ageing and the accelerated loss associated with pathological ageing are yet to be determined, oxidative stress has been shown to be important in ageing and age-related memory decline. While a number of preclinical and clinical studies have reported the beneficial effects of different food-based antioxidants and diets with nootropic effects [82-85], evidence from large human studies often fail to support the evidence garnered from preclinical studies. For example, there are reports that the antioxidant potential of vitamin E (as demonstrated by preclinical studies) has not been replicated in clinical trials [86].
2.1.1. Preclinical Studies
A number of preclinical studies have also demonstrated that the reduction of oxidative stress by the administration of antioxidants could mitigate age-related memory decline [82, 83]. Polyphenol-rich extracts of strawberry, blueberry and blackcurrant have been associated with antioxidant and anti-inflammatory properties that have been shown to mitigate age-related motor and cognitive disorders in rats [82, 87]. Joseph et al. [82] and Shukitt-Hale et al. [87] using aged Fischer 344 rats in separate studies reported that long-term supplementation with strawberry extract, spinach extract and vitamin E (in addition to improving antioxidant capacity) was associated with a reduction in age-related decline in memory and neuronal function [82, 87]. The nootropic effects of two antioxidants, alpha-lipoic acid and N-acetylcysteine, in rapidly ageing senescence-accelerated prone 8 (SAMP8) mice, which is a mouse model of sporadic Alzheimer’s Disease (AD) and dementia have also been reported [83]. Farr et al. [83] reported that in addition to reversing measured indices of oxidative stress, chronic administration of alpha-lipoic acid and N-acetylcysteine improved cognition in 12-month-old SAMP8 mice without altering motor activity, motivation to avoid shock, or body weight.
Studies evaluating the possible neuroprotective and cognition-enhancing effects of plant or diet-based flavonoids have demonstrated their effectiveness in enhancing cognitive function through the protection of vulnerable neurones, the enhancement of existing neuronal function and/or the stimulation of neurogenesis [88-91]. Animal studies in which diet was supplemented with flavonoid-rich foods such as pomegranate juice [91] or plant-derived flavonoids like quercetin, rutin [92] and fisetin [88, 90] have provided scientific evidence of the benefits of flavonoids in reversing the impact of ageing on brain structure and function. Currais et al. [90] evaluated the effects of fisetin supplementation in a mouse model of sporadic AD and reported a reduction in cognitive deficits in old SAMP8 mice, with the restoration of several markers of impaired synaptic function, stress and inflammation [90]. These beneficial effects have been attributed to fisetin’s direct antioxidant activity, increase in intracellular levels of glutathione, activation of key signalling pathways of a neurotrophic factor, as well as anti-inflammatory activity against glia [88].
2.1.2. Clinical Studies
In humans, a longitudinal population-based study carried out by Morris et al. [85] using three test batteries (mini-mental state examination, the East Boston Memory Test, and the Symbol Digit Modalities Test) reported that carotene, vitamin E and vitamin C intake from foods was associated with a decrease in memory decline with advancing age [84]. However, results from randomised controlled studies have failed to replicate this beneficial effect [85]. There have also been reports that the Mediterranean diet especially when supplemented with antioxidant-rich foods improves cognition and decreases cognitive decline when compared to the control diet [85]. The results of a clinical trial involving 447 volunteers (without cognitive deficits, and having a mean age of 66.9 years) who were randomly assigned to three groups based on the diet type (Mediterranean diet with extra-virgin olive oil, Mediterranean diet with mixed nuts, or control diet) showed that in the aged participants, a Mediterranean diet either supplemented with olive oil or nuts was associated with improved cognitive function [85]. A longitudinal study carried out by Solfrizzi et al. [93] reported that in an Italian population of elderly subjects on a typical Mediterranean diet, high mono-unsaturated fatty acid intakes could be protective against age-related memory decline [93]. Compounds with both antioxidant and anti-inflammatory potential like anthocyanins have also been shown to be of benefit in mitigating age-related cognitive decline, effects which have been attributed to their ability to increase neuronal signalling in brain regions that mediate memory function [94]. Krikorian et al. [94] reported that daily supplementation of wild blueberry juice (which is rich in anthocyanins) in a group of elderly subjects with early memory decline was associated with improvement in paired associative learning and word list recall [94].
2.1.3. Nootropic Agents
Nootropics, otherwise commonly-known as ‘smart drugs,’ are compounds that are known or believed to have the capacity to enhance the cognitive parameters such as memory, creativity, motivation, or attention. A number of them do so by modulating the activities of specific brain pathways involving neurotransmitters and neuromodulators that have distinct roles in the cognitive processes [95, 96]. Nootropics can be classified into two broad groups including synthetic compounds and natural or herbal nootropic agents [21]. Synthetic nootropic agents can be further classified into: a) agents that act at dopamine receptors (direct agonists and indirect agonists at Dopamine (D1) receptor), b) agents that act at adrenergic receptors such as the direct and indirect agonist of alpha (α2) adrenoceptor. The clinical efficacy of most of the synthetic nootropics that activate dopamine D1- and norepinephrine α2 receptors at low doses including amphetamines, methylphenidate, caffeine, armodafinil and modafinil is their ability to increase catecholamine levels in the prefrontal cortex [97-100], Other synthetic nootropic agents include; c) agent such as racetams that act at acetylcholine (Muscarinic (M1) agonist) and glutamate (AMPA (α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid) receptors [101-103].
While research has led to the synthesis of several drugs with nootropic effects, attention is now being shifted to the discovery, characterisation, and utilisation of nootropics from natural sources for the prevention and management of age-related cognitive decline. In the elderly, studies have demonstrated that some herbal or natural nootropic agents like gingko biloba improve cognition via their ability to induce vasodilatation in the small arteries and veins of the brain [95]. Advocates for the use of natural nootropic agents are also of the opinion that natural nootropic agents (e.g. bacopa monnieri and panax ginseng) act by enhancing hippocampal protein kinase activity [104], improve brain antioxidant activity and prevent neuronal apoptosis [105, 106] There have also been reports of the neuroprotective and adaptogenic properties of some herbal cognitive enhancers like Rhodiola rosea [107].
2.1.4. Natural Nootropics
The desire for drugs with fewer side-effects and similar (if not better) efficacy as standard drugs has continued to drive the search for newer natural or herbal nootropic agents [108]. In recent times, a number of herbs and plants with nootropic effects [21, 109] have been discovered, these include Ginkgo biloba [106, 110-112] Panax quinquefolius [113], Panax ginseng [114-116], Bacopa monnieri [117], Nicotiana tabacum, Nicotiana rustica [118, 119], Duboisia hopwoodii, Asclepias syriaca and Rhodiola rosea [107, 120]. Other plants with demonstrated or probable enhancement, preservative or restorative effects on memory include gotu kola (Centella asiatica) [121], lion's mane (Hericium erinaceus) [122], kapikacchu (Mucuna pruriens) [123], ashwagandha (Withania somnifera) [124, 125] and montmorency cherry [126]. In addition to these, natural compounds like huperzine A [127], galanthamine and vinpocetine [128] have been used for their nootropic effects. L-theanine [129, 130], which is an amino-acid that is present in green tea (Camelia sinensis), L-tyrosine [131] (an amino acid precursor to catecholamine neurotransmitters such as dopamine and noradrenaline), L-taurine [132, 133], a semi-essential, sulphur-containing amino acid and acetyl-l-carnitine [134], an endogenous antioxidant which also acts as a cellular energy carrier at the level of the mitochondrion are also notable compounds. Table 1 shows the specific mechanisms of action of synthetic nootropics as well as herbal and natural nootropic agents that have similar mechanisms, while Table 2 is a tabular presentation of the different preclinical and clinical studies that have examined the probable or confirmed roles and mechanisms Fig. (2) of a number of herbal and food-derived nootropic agents. Some of the herbs and food-derived nootropic agents that are shown in Table 2 are worthy of serious consideration, since they are already of some proven benefit in the clinical management of cognitive decline, or in its prevention. However, for many of the plants, only experimental evidence supported the empirical claims, thereby, pointing to the fact that there we are still a long way from confirming and establishing their clinical benefits and impacts of their long-term usage on general health.
Table 1. Mechanisms of action of synthetic nootropics and herbal/natural nootropic with similar mechanisms.
| Specific Mechanism | Synthetic Nootropics | Herbal/Natural Nootropic |
|---|---|---|
| Increased dopamine (D1) and adrenergic (α2) receptor activity, inhibition of norepinephrine uptake | Amphetamines, methylphenidate, modafinil, caffeine [97-100] |
Gingko biloba [112] Nicotine [118, 119], Kapikacchu (Mucuna pruriens) [123] Tyrosine [131] |
| Increased acetylcholine and/or glutamate receptor activity, acetylcholinesterase inhibiton | Racetams including piracetam, nefiracetam, nebracetam [101-103] | Panax ginseng [114-116], Bacopa monnieri [117], Huperzine A [127], Theanine [129, 130], Taurine [132, 133], Acetyl-l-carnitine [134] |
Table 2. Roles and efficacy of herbal and other food-derived nootropic agents.
| Natural Nootropic | Subject | Putative/Confirmed Roles and Mechanisms | References | |||
|---|---|---|---|---|---|---|
| Ginkgo biloba | Human (Clinical trial), rodent | Ginkgolides, bilobalide and proanthocyanidins are potent free radical scavengers/antioxidants which confer neuroprotection. Enhanced monoaminergic transmission. Increased cerebral blood flow via nitric oxide. Reduced amyloid neurotoxicity. | [111, 112] | |||
| Panax quinquefolius | Rodent | Increased choline uptake in synaptosomal preparations. Other putative mechanisms including hypoglycaemic action and effect of ginsenoside Rb1. | [113] | |||
| Panax ginseng | Human (Clinical trial), | Ginsenosides minimise inhibition of cholinergic transmission by amyloid-β protein in Alzheimer disease. Ginsenoside Rb1 increases cell survival in the hippocampus cornus ammonis 3 and dentate gyrus. |
[114, 115, 116] | |||
|
Bacopa
monnieri |
Rodent | Acetylcholinesterase inhibition, choline acetyltransferase activation, β-amyloid reduction, increased cerebral blood flow, monoamine potentiation and antioxidant neuroprotection. |
[117] | |||
| Nicotiana tabacum | Human (case study), rodent | Nicotine enhances efficacy of the cholinergic system, facilitating memory consolidation. Restoration of normal dendritic connections and the balance between hippocampal excitatory/inhibitory signals. |
[118, 119] | |||
| Nicotiana rustica | - | Nicotine content may facilitate memory during short-term use, as above. | - | |||
| Rhodiola rosea | - | Acts as an adaptogen due to the glycoside salidroside. | [120] | |||
| Gotu kola (Centella asiatica) | Rodent | Asiatic acid, asiaticoside, caffeoylquinic acids an madecassoside have neurotropic effects, including increased dendritic arborisation and synaptogenesis; probably due to modulations of signal transduction pathways. | [121] | |||
| Lion's mane (Hericium erinaceus) | Rodent | In mouse hippocampal slices, it causes an increase in spontaneous/evoked excitatory synaptic current in mossy fibre-Cornus ammonis-3synapse. Hericenones and erinacines increase levels of Nerve Growth Factor (NGF) in the brain, leading to increased neurite outgrowth and differentiation. | [122] | |||
| Kapikacchu (Mucuna pruriens) | Rodent | Contains L-Dopa which is a precursor of brain monoamines and neuromelanins. May contain other compounds with neuroprotective effects. | [123] | |||
| Ashwagandha (Withania somnifera) |
Guinea-pig, Rodent | Contains withanolides A to Y, dehydrowithanolide R, withasomniferin A, withasomidienone, withasomniferols A to C, withaferin A, and withanone. Also contains sitoindosides and beta-sitosterol. Restores levels of BDNF and regulates synaptic plasticity. Can trigger pathways for neural cell survival and plasticity. | [ 124. 125] | |||
| Montmorency cherry | Rodent | Reduced age-associated inflammatory (GFAP, NOX-2, COX-2)/autophagy (phosphorylated mTOR, Beclin 1, and p62/SQSTM) markers; and promotion of protein/cellular homeostasis in the hippocampus. | [126] | |||
| Huperzine A (Huperzia serrata) | Maacaques | Huperzine-A is a reversible acetylcholinesterase (AChE) inhibitor, and probable modulator of glutamatergic transmission. Also regulates brain cell mitochondria energetics and works as an antioxidant. | [127] | |||
| Vinpocetine | Rodent | Inhibits phosphodiesterase (PDE) enzyme to enhance second messenger-mediated signalling in pathways involved in learning and memory. Inhibits κB kinase (IKK)/Nuclear Factor-kappa B (NF-κB), and extracellular signal-regulated kinase (ERK) 1/2 to reduce inflammation. Enhances the structural dynamics of dendritic spines. | [128] | |||
| L-theanine | Human (Clinical trial), Rodent |
Improved cognitive function possibly via a decrease in NMDA-dependent CA1 long-term potentiation (LTP) and increase in NMDA-independent CA1-LTP. Antioxidant effect. Enhanced hippocampl neurogenesis. | [129, 130] | |||
| L-tyrosine | Rodent | Enhanced central catecholamine synthesis in rodents. | [131] | |||
| Taurine (l-taurine) | Rodent | Restores AChE and ChAT balance which are critical for the regulation of acetylcholine. Can decrease the insoluble fraction of amyloid beta in Alzheimer disease. | [132, 133] | |||
| Acetyl-l-carnitine | Rodent | Enhancement of high-affinity choline uptake, acetylcholine (ACh) synthesis, and depolarisation-evoked ACh release. Increased excitatory postsynaptic potential slope and population spike size in rat hippocampal slices. | [134] | |||
Fig. (2).
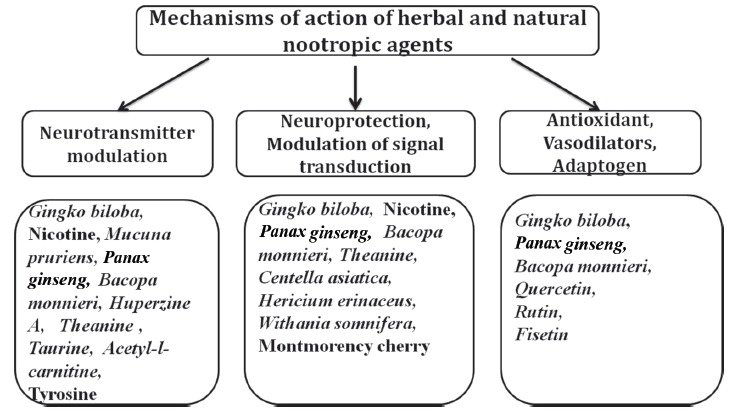
Mechanisms of action of herbal and natural nootropic agents.
2.1.4.1. Specific Herbs and Natural Compounds with Nootropic Activity
a). Ginkgo Biloba
For centuries, ginkgo biloba has been consumed as a source of food as well as for its medicinal value [111, 112, 135, 136]. In the last few decades or more, there has been extensive research into its possible nootropic and neuroprotective effects in humans and animals [111, 112, 136]. There are reports that ginkgo biloba is a widely consumed herbal supplement for the management of age-related memory decline [111]. In Europe and the United States of America, extracts of ginkgo biloba have been used increasingly to alleviate symptoms associated with age-related cognitive decline as well as Alzheimer's type and vascular dementia [111]. It had been reported that Ginkgo Biloba leaf Extract (GBE) retards production and aggregation of amyloid-β fibrils, reduces mitochondria-induced apoptosis, and down-regulates the activity of caspase-3 [81]. However, a few studies have demonstrated that administration of ginkgo biloba has no demonstrable effects on mitigating Alzheimer’s dementia or dementia due to other causes [78, 80]. Others have also suggested that its effects on cognitive change may be subtle, preventing or delaying memory decline in normal ageing and subjects with mild cognitive impairment [78]. Fehske et al. [112] using mice reported the cognition-enhancing effects of subchronic administration of standardised gingko biloba, which was linked to its ability to inhibit the activity of norepinephrine, serotonin and dopamine uptake transporters as well as the activity of monoamine oxidase in vitro [112]. In a randomised double-blind crossover study, subchronic administration of GBE was associated with improvement of working memory tasks in a cohort of healthy middle-aged (50-61-year-old) men [111], there was also associated increase in the amplitude if the Steady-State Visually Evoked Potential (SSVEP) amplitude at occipital and frontal sites, and increase SSVEP latency during the hold component of the working memory task [112]. Ginkgo biloba has also been reported to have antiapoptotic [137] and antioxidant [138] properties that are beneficial in ageing. The active compounds found in ginkgo biloba are terpenoids including ginkgolide and bilobalide which are believed to aid in the downregulation of caspase-3 activity [81], while the memory enhancing effects have been attributed to the presence of ginkgolides [135]. Ginkgolides have also been associated with terminating the effects of the amyloid peptide on long-term potentiation. In summary, putative mechanisms of ginkgo biloba activity on the brain include cerebral vasorelaxation [139, 140], modulation of brain neurotransmitters, and decrease in the production of oxygen free radicals [111, 112, 141, 142] Ginkgo biloba has also been suggested to improve memory by inhibiting the aggregation of amyloid [81].
b). Asian or American Ginseng
Ginseng has been used by Asians for at least 2000 years as a general body tonic [143]. Worldwide, it is one of the most widely-sold medicinal herbs [144]. Panax ginseng, also known as Asian ginseng occupies a very important place in Chinese traditional medicine [145] and has been used extensively for its cognition-enhancing and antioxidant properties [146]. In healthy subjects, panax ginseng has been associated with improvement of memory performance [106]. In patients with Alzheimer’s dementia, panax ginseng has also been shown to mitigate memory loss [114, 115, 146]. A 12-week randomised clinical trial carried out by Heo et al. [114] and Lee et al. [115], examining the effects of Panax ginseng on cognitive and functional performance (assessed with the mini mental scale examination, clinical dementia rating and Alzheimer disease assessment scale) in a cohort of Alzheimer’s disease patients reported significant improvement in cognitive functioning with increasing doses of Panax ginseng [114, 116]. Also, in rodents, it has been reported to improve memory in aged mice [147]. Biologically-active constituents of Panax ginseng are mainly ginsenoside saponins which can be divided into three groups namely panaxatriol, panaxadiol, and oleanolic acid. Other active constituents include peptides, polysaccharides, fatty acids and polyacetylenic alcohols [148]. The modulation of brain neurotransmitters by Panax ginseng has been attributed to panaxadiol and panaxatriol group of saponins [149]. Ginsenosides also affect the uptake of neurotransmitters like norepinephrine, serotonin, γ-amino butyric acid and dopamine [150]. Studies have also attributed the cognition-enhancing effects of panax ginseng to its high panaxatriol to panaxadiol ratio [151].
Panax quinquefolius also known as American ginseng is another prominent ginsenoside- containing herb, with a ginsenoside profile which is distinct from Panax ginseng. Studies have also shown that compared to Panax ginseng, panax quinquefolius has a lower ratio of panaxatriol to panaxadiol [152]. However, another study by Zhu et al. [153] reported that the ginsenoside content of panax quinquefolius was significantly higher compared to Panax ginseng [153]. While there is a dearth of scientific information on the cognition-enhancing properties of American ginseng in age-related memory impairment or dementia, the results of a randomised double blind placebo controlled study in young adults evaluating the neurocognitive effects of panax quinquefolius reported improvements in the cognitive performance in this cohort of young adults [154].
c). Nootropic Agents from the Indian Medicinal System
A number of herbs which had been in use in the Indian medicine system (Ayuverda) have in recent times been examined scientifically for documentation of their nootropic activity. These plants (Centella asiatica Linn, Glycirrhiza glabra Linn, Tinospora cordifolia and Convolvulus pleuricaulis Chois) which can be used either singly or in combination are collectively known as the Medhya Rasayanas [155]. In addition to their nootropic effects, they have also been used for their ability to nourish the body, improve immunity and enhance longevity. Other herbs include Celastrus panniculata, Bacopa monniera, Acorus calamus Benincasa hispida, and Nardostachys jatamamsi [155]. A number of these plants including Centella asiatica (which can be consumed as whole plant juices) contain saponins including medacoside, Asiatic acid, asiaticoside and triterpenic acid [156]. Studies using a mouse model of memory-impairment demonstrated that Centella asiatica mitigates scopolamine-induced memory loss through the inhibition of acetylcholinesterase [155]. Another herb Glycirrhiza glabra Linn whose active ingredients include flavonones, glycyrrhizine, isoflavones, glycyrrhetenic acid and phenolic compounds [155, 157] can be consumed as a liquorice and has been shown to be effective in mitigating memory loss in mice [158].
Bacopa monnieri is a small creeping plant indigenous to India which is best administered as a juice made from the maceration of whole parts of the plant. It has been shown to have in addition to its cognition-enhancing properties sedative, antioxidant and neuroprotective effects [155, 159, 160]. The nootropic effects have been attributed to a) its ability to scavenge reactive oxygen species [104] and enhance antioxidant activity [161], b) the enhancement of protein kinase activity in the hippocampus [162]. Bacopa monnieri is rich in saponins including bacosides which has been described as the active nootropic principle, especially in the alcoholic extract of the plant [163].
d). Other Plants
Other plants such as Moringa oleifera [164], Foeniculum vulgare Linn [165] and Nardostachys jatamansi [166] have been reported to have the ability to mitigate age-related memory decline in rodents by using various mechanisms, including the modulation of the cholinergic system [165], mitigation of lipid peroxidation, and improvement of antioxidant status [164, 166]. The plant alkaloid nicotine which is found in high very high concentrations in the leaves of the tobacco plant (Nicotiana tabacum) and other related species of plants including Nicotiana rustica, Duboisia hopwoodii, and Asclepias syriaca [167] have also been studied for its memory-enhancing properties specifically in age-related memory decline and Alzheimer’s dementia. Despite its potential for addiction and other undesirable effects [168], the use of nicotine has been associated with improvement of learning and memory indices in healthy cohorts and the mitigation of memory impairment in ageing. These benefits have been attributed to its ability to downregulate the expression of the nicotinic receptors in subjects with Alzheimer's disease [169].
CONCLUSION
While it is a known fact that the human brain endures diverse insults in the process of ageing, food-based nootropics are likely to go a long way in mitigating the impacts of these insults. However, before we can get to the point where food-based nootropics are ‘routinely prescribed’ for the prevention or management of age-related cognitive decline, issues that need to be addressed include those of safety, standardisation of active constituents, and ascertaining which combinations work best for different conditions. Also, the details of the interactions that may occur between food-based nootropics and orthodox medications must be known.
ACKNOWLEDGEMENTS
Declared none.
AUTHOR CONTRIBUTION
Onaolapo AY, Obelawo AY and Onaolapo OJ: All authors contributed equally to the writing of this manuscript.
CONSENT FOR PUBLICATION
Not applicable.
FUNDING
None.
CONFLICT OF INTEREST
The authors declare no conflict of interest, financial or otherwise.
REFERENCES
- 1.European Commission Eurostat Active Ageing and Solidarity Between Generations. A Statistical Portrait of the European Union. 2012. [Google Scholar]
- 2.Kelley A.S., McGarry K., Gorges R., Skinner J.S. The burden of health care costs for patients with dementia in the last 5 years of life. Ann. Intern. Med. 2015;163:729–736. doi: 10.7326/M15-0381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Joseph J., Cole G., Head E., Ingram D. Nutrition, brain aging, and neurodegeneration. J. Neurosci. 2009;29:12795–12801. doi: 10.1523/JNEUROSCI.3520-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Cornelius E. Increased incidence of lymphomas in thymectomized mice-evidence for an immunological theory of aging. Experientia. 1972;28:459. doi: 10.1007/BF02008340. [DOI] [PubMed] [Google Scholar]
- 5.Davidovic M., Sevo G., Svorcan P., Milosevic D.P., Despotovic N., Erceg P. Old age as a privilege of the “selfish ones”. Aging Dis. 2010;1:139–146. [PMC free article] [PubMed] [Google Scholar]
- 6.Jin K. Modern biological theories of aging. Aging Dis. 2010;1:72–74. [PMC free article] [PubMed] [Google Scholar]
- 7.van Heemst D. Insulin, IGF-1 and longevity. Aging Dis. 2010;1:147–157. [PMC free article] [PubMed] [Google Scholar]
- 8.Sergiev P.V., Dontsova O.A., Berezkin G.V. Theories of aging: An ever-evolving field. J Nature. 2015;7:9–18. [PMC free article] [PubMed] [Google Scholar]
- 9.Brayne C. The elephant in the room: Healthy brains in later life, epidemiology and public health. Nat. Rev. Neurosci. 2007;8:233–239. doi: 10.1038/nrn2091. [DOI] [PubMed] [Google Scholar]
- 10.Deary I.J., Corley J., Gow A.J., Harris S.E., Houlihan L.M., Marioni R.E., et al. Age-associated cognitive decline. Br. Med. Bull. 2009;92:135–152. doi: 10.1093/bmb/ldp033. [DOI] [PubMed] [Google Scholar]
- 11.Pennisi G., Bella R., Lanza G. Motor cortex plasticity in subcortical ischemic vascular dementia: What can TMS say? Clin. Neurophysiol. 2015;126:851–852. doi: 10.1016/j.clinph.2014.09.001. [DOI] [PubMed] [Google Scholar]
- 12.Psaltopoulou T., Sergentanis T.N., Panagiotakos D.B., Sergentanis I.N., Kosti R., Scarmeas N. Mediterranean diet, stroke, cognitive impairment, and depression: A meta-analysis. Ann. Neurol. 2013;74:580–591. doi: 10.1002/ana.23944. [DOI] [PubMed] [Google Scholar]
- 13.Kuczmarski M.F., Allegro D., Stave E. The association of healthful diets and cognitive function: A review. J. Nutr. Gerontol. Geriatr. 2014;33:69–90. doi: 10.1080/21551197.2014.907101. [DOI] [PubMed] [Google Scholar]
- 14.Allès B., Samieri C., Féart C., Jutand M.A., Laurin D., Barberger-Gateau P. Dietary patterns: A novel approach to examine the link between nutrition and cognitive function in older individuals. Nutr. Res. Rev. 2015;25:207–222. doi: 10.1017/S0954422412000133. [DOI] [PubMed] [Google Scholar]
- 15.van de Rest O., Berendsen A.A., Haveman-Nies A., de Groot L.C. Dietary patterns, cognitive decline, and dementia: A systematic review. Adv. Nutr. 2015;6:154–168. doi: 10.3945/an.114.007617. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Gouliaev A.H., Senning A. Piracetam and other structurally related nootropics. Brain Res. Brain Res. Rev. 1994;19:180–222. doi: 10.1016/0165-0173(94)90011-6. [DOI] [PubMed] [Google Scholar]
- 17.Noble S., Benfield P. Piracetam: A review of its clinical potential in the management of patients with stroke. CNS Drugs. 1998;9:497–511. doi: 10.2165/00023210-199809060-00006. [DOI] [Google Scholar]
- 18.Giovannini M.G., Rodinò P., Mutolo D., Pepeu G. Oxiracetam and aniracetam increase acetylcholine release from the rat hippocampus in vivo. Drug Dev. Res. 1993;28:503–509. doi: 10.1002/ddr.43028040. [DOI] [Google Scholar]
- 19.Rao Y., Xiao P., Xu S.T. Effects of intrahippocampal aniracetam treatment on Y-maze avoidance learning performance and behavioral long-term potentiation in dentate gyrus in rat. Neurosci. Lett. 2001;298:183–186. doi: 10.1016/S0304-3940(00)01744-4. [DOI] [PubMed] [Google Scholar]
- 20.Dahiya S., Rani R., Dhingra D., Kumar S., Dilbaghi N. Potentiation of nootropic activity of EGCG loaded nanosuspension by piperine in swiss male albino mice. J. Pharm. Sci. 2018;4:296–302. [Google Scholar]
- 21.Suliman N.A., Mat T.C.N., Mohd M.M.A., Adenan M.I., Hidayat B.M.T., Basir R. Establishing natural nootropics: Recent molecular enhancement influenced by natural nootropic. Evid. Based Complement. Alternat. Med. 2016;2016:4391375. doi: 10.1155/2016/4391375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Salthouse TA. When does age-related cognitive decline begin? Neurobiol Aging. 2009;30:507–14. doi: 10.1016/j.neurobiolaging.2008.09.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Thompson J.J., Blair M.R., Henrey A.J. Over the hill at 24: Persistent age-related cognitive-motor decline in reaction times in an ecologically valid video game task begins in early adulthood. PLoS One. 2014;9:e94215. doi: 10.1371/journal.pone.0094215. p. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Peters R. Ageing and the brain. Postgrad. Med. J. 2006;82:84–88. doi: 10.1136/pgmj.2005.036665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Schmitz B., Wang X., Barker P.B., Pilatus U., Bronzlik P., Dadak M., et al. Effects of aging on the human brain: A Proton and Phosphorus MR spectroscopy study at 3T. J. Neuroimaging. 2018;28:416–421. doi: 10.1111/jon.12514. [DOI] [PubMed] [Google Scholar]
- 26.Svennerholm L., Boström K., Jungbjer B. Changes in weight and compositions of major membrane components of human brain during the span of adult human life of Swedes. Acta Neuropathol. 1997;94:345–352. doi: 10.1007/s004010050717. [DOI] [PubMed] [Google Scholar]
- 27.Scahill R.I., Frost C., Jenkins R., Whitwell J.L., Rossor M.N., Fox N.C. A longitudinal study of brain volume changes in normal aging using serial registered magnetic resonance imaging. Arch. Neurol. 2003;60:989–994. doi: 10.1001/archneur.60.7.989. [DOI] [PubMed] [Google Scholar]
- 28.Lanza G., Bramanti P., Cantone M., Pennisi M., Pennisi G., Bella R. Vascular cognitive impairment through the looking glass of transcranial magnetic stimulation. Behav. Neurol. 2017;2017:1421326. doi: 10.1155/2017/1421326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Ishii R., Canuet L., Aoki Y., Hata M., Iwase M., Ikeda S., et al. Healthy and pathological brain aging: From the perspective of oscillations, functional connectivity, and signal complexity. Neuropsychobiology. 2017;75:151–161. doi: 10.1159/000486870. [DOI] [PubMed] [Google Scholar]
- 30.Dröge W., Schipper H.M. Oxidative stress and aberrant signalling in aging and cognitive decline. Aging Cell. 2007;6:361–370. doi: 10.1111/j.1474-9726.2007.00294.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Haddadi M., Jahromi S.R., Sagar B.K., Patil R.K., Shivanandappa T., Ramesh S.R. Brain aging, memory impairment and oxidative stress: A study in Drosophila melanogaster. Behav. Brain Res. 2014;259:60–69. doi: 10.1016/j.bbr.2013.10.036. [DOI] [PubMed] [Google Scholar]
- 32.Fukui K., Onodera K., Shinkai T., Suzuki S., Urano S. Impairment of learning and memory in rats caused by oxidative stress and aging, and changes in antioxidative defense systems. Ann. N. Y. Acad. Sci. 2001;928:168–175. doi: 10.1111/j.1749-6632.2001.tb05646.x. [DOI] [PubMed] [Google Scholar]
- 33.Fukui K., Omoi N.O., Hayasaka T., Shinnkai T., Suzuki S., Abe K., et al. Cognitive impairment of rats caused by oxidative stress and aging, and its prevention by vitamin E. Ann. N. Y. Acad. Sci. 2002;959:275–284. doi: 10.1111/j.1749-6632.2002.tb02099.x. [DOI] [PubMed] [Google Scholar]
- 34.Aksenov M.Y., Aksenova M.V., Butterfield D.A., Geddes J.W., Markesbery W.R. Protein oxidation in the brain in Alzheimer’s disease. Neuroscience. 2001;103:373–383. doi: 10.1016/s0306-4522(00)00580-7. [DOI] [PubMed] [Google Scholar]
- 35.Hajjar I., Hayek S.S., Goldstein F.C., Martin G., Jones D.P., Quyyumi A. Oxidative stress predicts cognitive decline with aging in healthy adults: An observational study. J. Neuroinflammation. 2018;15:17. doi: 10.1186/s12974-017-1026-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Alzheimer’s Association. Alzheimer’s Facts and Figures. 2014 http://www.alz.org/alzheimers_disease_ facts_and_figures.asp
- 37.Alzheimer’s Association Alzheimer’s Facts and Figures. 2018;14:367–429. [Google Scholar]
- 38.World Health Organization. Dementia: A public health priority. Executive Summary; 2012. [Google Scholar]
- 39.World Health Organisation. Towards a dementia plan. 2018. A WHO guide. [Google Scholar]
- 40.Plassman B.L., Langa K.M., Fisher G.G., Heeringa S.G., Weir D.R., Ofstedal M.B., et al. Prevalence of cognitive impairment without dementia in the United States. Ann. Intern. Med. 2008;148:427–434. doi: 10.7326/0003-4819-148-6-200803180-00005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Unverzagt F.W., Ogunniyi A., Taler V., Gao S., Lane K.A., Baiyewu O., et al. Incidence and risk factors for cognitive impairment no dementia and mild cognitive impairment in African Americans. Alzheimer Dis. Assoc. Disord. 2011;25:4–10. doi: 10.1097/WAD.0b013e3181f1c8b1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Parikh P.K., Troyer A.K., Maione A.M., Murphy K.J. The impact of memory change on daily life in normal aging and mild cognitive impairment. Gerontologist. 2016;56:877-85. doi: 10.1093/geront/gnv030. https://doi.org/10 [DOI] [PubMed] [Google Scholar]
- 43.Parkin A. Memory and amnesia. Oxford: Blackwall; 1997. [Google Scholar]
- 44.Fjell A.M., McEvoy L., Holland D., Dale A.M., Walhovd K.B. Alzheimer’s Disease Neuroimaging Initiative. What is normal in normal aging? Effects of aging, amyloid and Alzheimer’s disease on the cerebral cortex and the hippocampus. Prog. Neurobiol. 2014;117:20–40. doi: 10.1016/j.pneurobio.2014.02.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Brem A.K., Ran K., Pascual-Leone A. Learning and memory. Handb. Clin. Neurol. 2013;116:693–737. doi: 10.1016/B978-0-444-53497-2.00055-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Connelly S.L., Hasher L., Zacks R.T. Age and reading: The impact of distraction. Psychol. Buckner RL Memory and executive function in aging and AD: Multiple factors that cause decline and reserve factors that compensate. Neuron. 2004;44:195–208. doi: 10.1016/j.neuron.2004.09.006. [DOI] [PubMed] [Google Scholar]
- 47.Nyberg L., Lovden M., Riklund K., Lindenberger U., Backman L. Memory aging and brain maintenance. Trends Cogn. Sci. 2012;16:292–305. doi: 10.1016/j.tics.2012.04.005. [DOI] [PubMed] [Google Scholar]
- 48.Nilsson L.G. Memory function in normal aging. Acta Neurol. Scand. Suppl. 2003;179:7–13. doi: 10.1034/j.1600-0404.107.s179.5.x. [DOI] [PubMed] [Google Scholar]
- 49.Fleischman D.A., Wilson R.S., Gabrieli J.D., Bienias J.L., Bennett D.A. A longitudinal study of implicit and explicit memory in old persons. Psychol. Aging. 2004;19:617–625. doi: 10.1037/0882-7974.19.4.617. [DOI] [PubMed] [Google Scholar]
- 50.Rabbitt P., Lowe C., Shilling V. Frontal tests and models for cognitive ageing. Eur. J. Cogn. Psychol. 2001;13:5–28. [Google Scholar]
- 51.Salthouse T.A. The processing-speed theory of adult age differences in cognition. Psychol. Rev. 1996;103:403–428. doi: 10.1037/0033-295x.103.3.403. [DOI] [PubMed] [Google Scholar]
- 52.Park D.C., Reuter-Lorenz P. The adaptive brain: Aging and neurocognitive scaffolding. Annu. Rev. Psychol. 2009;60:173–196. doi: 10.1146/annurev.psych.59.103006.093656. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Dillon C., Serrano C.M., Castro D., Leguizamón P.P., Heisecke S.L., Taragano F.E. Behavioral symptoms related to cognitive impairment. Neuropsychiatr. Dis. Treat. 2013;9:1443–1455. doi: 10.2147/NDT.S47133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Verhaeghen P. Aging and vocabulary scores: A meta-analysis. Psychol. Aging. 2003;18:332–339. doi: 10.1037/0882-7974.18.2.332. [DOI] [PubMed] [Google Scholar]
- 55.Kent P.S., Luszcz M.A. A review of the Boston naming test and multiple-occasion normative data for older adults on 15-item versions. Clin. Neuropsychol. 2002;16:555–574. doi: 10.1076/clin.16.4.555.13916. [DOI] [PubMed] [Google Scholar]
- 56.Rogers S.L., Friedman R.B. The underlying mechanisms of semantic memory loss in Alzheimer’s disease and semantic dementia. Neuropsychologia. 2008;46:12–21. doi: 10.1016/j.neuropsychologia.2007.08.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.ADI. Alzheimer’s Disease International. Policy brief for G8 heads of government: the global impact of dementia 2013-2050. London: Alzheimer’s Disease International; 2013. [Google Scholar]
- 58.Prince M., Bryce R., Albanese E., Wimo A., Ribeiro W., Ferri C.P. The global prevalence of dementia: A systematic review and metaanalysis. Alzheimers Dement. 2013;9:63–75. doi: 10.1016/j.jalz.2012.11.007. [DOI] [PubMed] [Google Scholar]
- 59.Prince M., Ali G.C., Guerchet M., Prina A.M., Albanese E., Wu Y.T. Recent global trends in the prevalence and incidence of dementia, and survival with dementia. Alzheimers Res. Ther. 2016;8:23. doi: 10.1186/s13195-016-0188-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Langa K.M. Is the risk of Alzheimer’s disease and dementia declining? Alzheimers Res. Ther. 2015;7:34. doi: 10.1186/s13195-015-0118-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Smith P.J., Blumenthal J.A. Dietary Factors and Cognitive Decline. J. Prev. Alzheimers Dis. 2016;3:53–64. doi: 10.14283/jpad.2015.71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Pennisi G., Lanza G., Giuffrida S., Vinciguerra L., Puglisi V., Cantone M., et al. Excitability of the Motor Cortex in De Novo Patients with Celiac Disease. PLoS One. 2014;9:e102790. doi: 10.1371/journal.pone.0102790. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Gómez-Pinilla F. Brain foods: The effects of nutrients on brain function. Nat. Rev. Neurosci. 2008;9:568–578. doi: 10.1038/nrn2421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Spencer S.J., Korosi A., Layé S., Shukitt-Hale B., Barrientos R.M. Food for thought: How nutrition impacts cognition and emotion. NPJ Sci Food. 2017;1:7. doi: 10.1038/s41538-017-0008-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Lourida I., Soni M., Thompson-Coon J., Purandare N., Lang I.A., Ukoumunne O.C., et al. Mediterranean diet, cognitive function, and dementia: A systematic review. Epidemiology. 2013;24:479–489. doi: 10.1097/EDE.0b013e3182944410. [DOI] [PubMed] [Google Scholar]
- 66.Beilharz J.E., Maniam J., Morris M.J. Diet-Induced cognitive deficits: The role of fat and sugar, potential mechanisms and nutritional interventions. Nutrients. 2015;7:6719–6738. doi: 10.3390/nu7085307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Petersson S.D., Philippou E. Mediterranean diet, cognitive function, and dementia: A systematic review of the evidence. Adv. Nutr. 2016;7:889–904. doi: 10.3945/an.116.012138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Shakersain B., Santoni G., Larsson S.C., Faxén-Irving G., Fastbom J., Fratiglioni L., et al. Prudent diet may attenuate the adverse effects of Western diet on cognitive decline. Alzheimers Dement. 2016;12:100–109. doi: 10.1016/j.jalz.2015.08.002. [DOI] [PubMed] [Google Scholar]
- 69.Malouf R., Grimley E.J. The effect of vitamin B6 on cognition. Cochrane Database Syst. Rev. 2003;4:CD004393. doi: 10.1002/14651858.CD004393. [DOI] [PubMed] [Google Scholar]
- 70.Quadri P., Fragiacomo C., Pezzati R., Zanda E., Forloni G., Tettamanti M., et al. Homocysteine, folate, and vitamin B-12 in mild cognitive impairment, Alzheimer disease, and vascular dementia. Am. J. Clin. Nutr. 2004;80:114–122. doi: 10.1093/ajcn/80.1.114. [DOI] [PubMed] [Google Scholar]
- 71.Quadri P., Fragiacomo C., Pezzati R., Zanda E., Tettamanti M., Lucca U. Homocysteine and B vitamins in mild cognitive impairment and dementia. Clin. Chem. Lab. Med. 2005;43:1096–1100. doi: 10.1515/CCLM.2005.191. [DOI] [PubMed] [Google Scholar]
- 72.Malouf R., Grimley E.J. Folic acid with or without vitamin B12 for the prevention and treatment of healthy elderly and demented people. Cochrane Database Syst. Rev. 2008 doi: 10.1002/14651858.CD004514.pub2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Wald D.S., Kasturiratne A., Simmonds M. Effect of folic acid, with or without other B vitamins, on cognitive decline: meta-analysis of randomized trials. Am. J. Med. 2010;123:522–527. doi: 10.1016/j.amjmed.2010.01.017. [DOI] [PubMed] [Google Scholar]
- 74.Muskiet F.A., van Goor S.A., Kuipers R.S., Velzing-Aarts F.V., Smit E.N., Bouwstra H., et al. Long-chain polyunsaturated fatty acids in maternal and infant nutrition. Prostaglandins Leukot. Essent. Fatty Acids. 2006;75:135–144. doi: 10.1016/j.plefa.2006.05.010. [DOI] [PubMed] [Google Scholar]
- 75.Boudrault C., Bazinet R.P., Ma D.W. Experimental models and mechanisms underlying the protective effects of n-3 polyunsaturated fatty acids in Alzheimer’s disease. J. Nutr. Biochem. 2009;20:1–10. doi: 10.1016/j.jnutbio.2008.05.016. [DOI] [PubMed] [Google Scholar]
- 76.Letenneur L., Proust-Lima C., Le Gouge A., Dartigues J.F., Barberger-Gateau P. Flavonoid intake and cognitive decline over a 10-year period. Am. J. Epidemiol. 2007;165:1364–1371. doi: 10.1093/aje/kwm036. [DOI] [PubMed] [Google Scholar]
- 77.Giudetti A.M., Salzet M., Cassano T. Oxidative Stress in Aging Brain: Nutritional and Pharmacological Interventions for Neurodegenerative Disorders. Oxid. Med. Cell. Longev. 2018;2018:3416028. doi: 10.1155/2018/3416028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.DeKosky S.T., Williamson J.D., Fitzpatrick A.L., Kronmal R.A., Ives D.G., Saxton J.A., et al. Ginkgo Evaluation of Memory (GEM) Study Investigators JAMA. 2008;300:2253–2262. [Google Scholar]
- 79.Taylor J.E. Neuromediator binding to receptors in the rat brain. The effect of chronic administration of Ginkgo biloba extract. Presse Med. 1986;15(31):1491–1493. [PubMed] [Google Scholar]
- 80.Luo Y., Smith J.V., Paramasivam V., Burdick A., Curry K.J., Buford J.P., et al. Inhibition of amyloid-β aggregation and caspase-3 activation by the Ginkgo biloba extract EGb761. Proc. Natl. Acad. Sci. USA. 2002;99:12197–12202. doi: 10.1073/pnas.182425199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Christen Y. Ginkgo biloba and neurodegenerative disorders. Front. Biosci. 2004;9:3091–3104. doi: 10.2741/1462. [DOI] [PubMed] [Google Scholar]
- 82.Joseph J.A., Shukitt-Hale B., Denisova N.A., Prior R.L., Cao G., Martin A., et al. Long-term dietary strawberry, spinach, or vitamin E supplementation retards the onset of age-related neuronal signal-transduction and cognitive behavioral deficits. J. Neurosci. 1998;18:8047–8055. doi: 10.1523/JNEUROSCI.18-19-08047.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Farr S.A., Poon H.F., Dogrukol-Ak D., Drake J., Banks W.A., Eyerman E., et al. The antioxidants alpha-lipoic acid and N-acetyl cysteine reverse memory impairment and brain oxidative stress in aged SAMP8 mice. J. Neurochem. 2003;84:1173–1183. doi: 10.1046/j.1471-4159.2003.01580.x. [DOI] [PubMed] [Google Scholar]
- 84.Morris M.C., Evans D.A., Bienias J.L., Tangney C.C., Wilson R.S. Vitamin E and cognitive decline in older persons. Arch. Neurol. 2002;59:1125–1132. doi: 10.1001/archneur.59.7.1125. [DOI] [PubMed] [Google Scholar]
- 85.Valls-Pedret C., Sala-Vila A., Serra-Mir M., Corella D., de la Torre R., Martínez-González M.Á., et al. Mediterranean diet and age-related cognitive decline: A randomized clinical trial. JAMA Intern. Med. 2015;175:1094–1103. doi: 10.1001/jamainternmed.2015.1668. [DOI] [PubMed] [Google Scholar]
- 86.Corbett A., Ballard C. The value of vitamin E as a treatment for Alzheimer’s disease remains unproven despite functional improvement, due to a lack of established effect on cognition or other outcomes from RCTs. Evid. Based Med. 2014;19:140. doi: 10.1136/eb-2014-101741. [DOI] [PubMed] [Google Scholar]
- 87.Shukitt-Hale B., Cheng V., Joseph J.A. Effects of blackberries on motor and cognitive function in aged rats. Nutr. Neurosci. 2009;12(3):135–140. doi: 10.1179/147683009X423292. [DOI] [PubMed] [Google Scholar]
- 88.Maher P. How fisetin reduces the impact of age and disease on CNS function. Front. Biosci. (Schol. Ed.) 2015;7:58–82. doi: 10.2741/s425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Rendeiro C., Spencer J.P., Vauzour D., Butler L.T., Ellis J.A., Williams C.M. The impact of flavonoids on spatial memory in rodents: From behaviour to underlying hippocampal mechanisms. Genes Nutr. 2009;4(4):251–270. doi: 10.1007/s12263-009-0137-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Currais A., Farrokhi C., Dargusch R., Armando A., Quehenberger O., Schubert D., et al. Fisetin reduces the impact of aging on behavior and physiology in the rapidly agingsamp8 mouse. J. Gerontol. A Biol. Sci. Med. Sci. 2018;73:299–307. doi: 10.1093/gerona/glx104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Hartman R.E., Shah A., Fagan A.M., Schwetye K.E., Parsadanian M., Schulman R.N., et al. Pomegranate juice decreases amyloid load and improves behavior in a mouse model of Alzheimer’s disease. Neurobiol. Dis. 2006;24:506–515. doi: 10.1016/j.nbd.2006.08.006. [DOI] [PubMed] [Google Scholar]
- 92.Pu F., Mishima K., Irie K., Motohashi K., Tanaka Y., Orito K., et al. Neuroprotective effects of quercetin and rutin on spatial memory impairment in an 8-arm radial maze task and neuronal death induced by repeated cerebral ischemia in rats. J. Pharmacol. Sci. 2007;104:329–334. doi: 10.1254/jphs.fp0070247. [DOI] [PubMed] [Google Scholar]
- 93.Solfrizzi V., Panza F., Torres F., Mastroianni F., Del Parigi A., Venezia A., et al. High monounsaturated fatty acids intake protects against age-related cognitive decline. Neurology. 1999;52:1563–1569. doi: 10.1212/wnl.52.8.1563. [DOI] [PubMed] [Google Scholar]
- 94.Krikorian R., Shidler M.D., Nash T.A., Kalt W., Vinqvist-Tymchuk M.R., Shukitt-Hale B., et al. Blueberry supplementation improves memory in older adults. J. Agric. Food Chem. 2010;58:3996–4000. doi: 10.1021/jf9029332. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Colucci L., Bosco M., Rosario Z.A., Rea R., Amenta F., Fasanaro A.M. Effectiveness of nootropic drugs with cholinergic activity in treatment of cognitive deficit: A review. J. Exp. Pharmacol. 2012;4:163–172. doi: 10.2147/JEP.S35326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Dartigues J.F., Carcaillon L., Helmer C., Lechevallier N., Lafuma A., Khoshnood B. Vasodilators and nootropics as predictors of dementia and mortality in the PAQUID cohort. J. Am. Geriatr. Soc. 2007;55:395–399. doi: 10.1111/j.1532-5415.2007.01084.x. [DOI] [PubMed] [Google Scholar]
- 97.Ilieva I.P., Hook C.J., Farah M.J. Prescription stimulants’ effects on healthy inhibitory control, working memory, and episodic memory: A Meta-analysis. J. Cogn. Neurosci. 2015;27:1069–1089. doi: 10.1162/jocn_a_00776. [DOI] [PubMed] [Google Scholar]
- 98.Spencer R.C., Devilbiss D.M., Berridge C.W. The cognition-enhancing effects of psychostimulants involve direct action in the prefrontal cortex. Biol. Psychiatry. 2015;77:940–950. doi: 10.1016/j.biopsych.2014.09.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Camfield D.A., Stough C., Farrimond J., Scholey A.B. Acute effects of tea constituents L-theanine, caffeine, and epigallocatechin gallate on cognitive function and mood: A systematic review and meta-analysis. Nutr. Rev. 2014;72:507–522. doi: 10.1111/nure.12120. [DOI] [PubMed] [Google Scholar]
- 100.Battleday R.M., Brem A.K. Modafinil for cognitive neuroenhancement in healthy non-sleep-deprived subjects: A systematic review. Eur. Neuropsychopharmacol. 2015;25:1865–1881. doi: 10.1016/j.euroneuro.2015.07.028. [DOI] [PubMed] [Google Scholar]
- 101.Ahmed A.H., Oswald R.E. Piracetam defines a new binding site for allosteric modulators of alpha-amino-3-hydroxy-5-methyl-4-isoxazole-propionic acid (AMPA) receptors. J. Med. Chem. 2010;53:2197–2203. doi: 10.1021/jm901905j. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Takeo S., Hayashi H., Miyake K., Takagi K., Tadokoro M., Takagi N., et al. Effects of delayed treatment with nebracetam on neurotransmitters in brain regions after microsphere embolism in rats. Br. J. Pharmacol. 1997;121:477–48. doi: 10.1038/sj.bjp.0701161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Oyaizu M., Narahashi T. Modulation of the neuronal nicotinic acetylcholine receptor-channel by the nootropic drug nefiracetam. Brain Res. 1999;822:72–79. doi: 10.1016/s0006-8993(99)01077-x. [DOI] [PubMed] [Google Scholar]
- 104.Singh H.K., Dhawan B.N. Neuropsychopharmacological effects of the Ayurvedic nootropic Bacopa monniera Linn. (Brahmi). Int. J. Pharmacol. 1997;29:359–s65. [Google Scholar]
- 105.Anand T., Naika M., Swamy M.S.L., Khanum F. Antioxidant and DNA damage preventive properties of Bacopa monniera (L) wettst. Free Radic. Antioxid. 2011;1:84–90. [Google Scholar]
- 106.Kennedy D.O., Scholey A.B. Ginseng: Potential for the enhancement of cognitive performance and mood. Pharmacol. Biochem. Behav. 2003;75:687–700. doi: 10.1016/s0091-3057(03)00126-6. [DOI] [PubMed] [Google Scholar]
- 107.Qu Z.Q., Zhou Y., Zeng Y.S., Li Y., Chung P. Pretreatment with Rhodiola rosea extract reduces cognitive impairment induced by intracerebroventricular streptozotocin in rats: Implication of anti-oxidative and neuroprotective effects. Biomed. Environ. Sci. 2009;22(4):318–326. doi: 10.1016/S0895-3988(09)60062-3. [DOI] [PubMed] [Google Scholar]
- 108.Cervenka F., Jahodár L. Plant metabolites as nootropics and cognitives. Ceska Slov. Farm. 2006;55:219–229. [PubMed] [Google Scholar]
- 109.Roodenrys S., Booth D., Bulzomi S., Phipps A., Micallef C., Smoker J. Chronic effects of Brahmi (Bacopa monnieri) on human memory. Neuropsychopharmacology. 2002;27:279–281. doi: 10.1016/s0893-133x(01)00419-5. [DOI] [PubMed] [Google Scholar]
- 110.Onaolapo O.J., Onaolapo A.Y. Effects of Ginsomin® on Selected Behaviours in Mice. Int J Neurosci Behav Sci. 2017;5:45–54. doi: 10.13189/ijnbs.2017.050301. [DOI] [Google Scholar]
- 111.Silberstein R.B., Pipingas A., Song J., Camfield D.A., Nathan P.J., Stough C. Examining brain-cognition effects of Ginkgo biloba extract: Brain activation in the left temporal and left prefrontal cortex in an object working memory task evidence-based complement. Alt Med. 2011;2011:164139. doi: 10.1155/2011/164139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Fehske C.J., Leuner K., Müller W.E. Ginkgo biloba extract (EGb761®) influences monoaminergic neurotransmission via inhibition of NE uptake, but not MAO activity after chronic treatment. Pharmacol. Res. 2009;60:68–73. doi: 10.1016/j.phrs.2009.02.012. [DOI] [PubMed] [Google Scholar]
- 113.Sloley B., Pang P., Huang B., Ba F., Li F., Benishin C., et al. American ginseng extract reduces scopolamine-induced amnesia in a spatial learning task. J. Psychiatry Neurosci. 1999;24:442. [PMC free article] [PubMed] [Google Scholar]
- 114.Heo J.H., Lee S.T., Chu K., Oh M.J., Park H.J., Shim J.Y., et al. An open-label trial of Korean red ginseng as an adjuvant treatment for cognitive impairment in patients with Alzheimer’s disease. Eur. J. Neurol. 2008;15:865–868. doi: 10.1111/j.1468-1331.2008.02157.x. [DOI] [PubMed] [Google Scholar]
- 115.Lee S.T., Chu K., Sim J.Y., et al. Panax ginseng enhances cognitive performance in Alzheimer disease. Alzheimer Dis. Assoc. Disord. 2008;22:222–226. doi: 10.1097/WAD.0b013e31816c92e6. [DOI] [PubMed] [Google Scholar]
- 116.Liu L., Hoang-Gia T., Wu H., Lee M.R., Gu L., Wang C., et al. Ginsenoside Rb1 improves spatial learning and memory by regulation of cell genesis in the hippocampal subregions of rats. Brain Res. 2001;25(1382):147–154. doi: 10.1016/j.brainres.2011.01.051. [DOI] [PubMed] [Google Scholar]
- 117.Aguiar S., Borowski T. Neuropharmacological review of the nootropic herb bacopa monnieri. Rejuvenation Res. 2013;16:313–326. doi: 10.1089/rej.2013.1431. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Beer A.L., Vartak D., Greenlee M.W. Nicotine facilitates memory consolidation in perceptual learning. Neuropharmacology. 2013;64:443–451. doi: 10.1016/j.neuropharm.2012.06.019. [DOI] [PubMed] [Google Scholar]
- 119.Molas S., Gener T., Güell J., Martín M., Ballesteros-Yáñez I., Sanchez-Vives M.V., et al. Hippocampal changes produced by overexpression of the human CHRNA5/A3/B4 gene cluster may underlie cognitive deficits rescued by nicotine in transgenic mice. Acta Neuropathol. Commun. 2014;11:147. doi: 10.1186/s40478-014-0147-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Palmeri A., Mammana L., Tropea M.R., Gulisano W., Puzzo D. Salidroside, a bioactive compound of Rhodiola Rosea, ameliorates memory and emotional behavior in adult mice. J. Alzheimers Dis. 2016;52:65–75. doi: 10.3233/JAD-151159. [DOI] [PubMed] [Google Scholar]
- 121.Gray N.E., Magana A.A., Lak P., Wright K.M., Quinn J., Stevens J.F., et al. Centella asiatica: Phytochemistry and mechanisms of neuroprotection and cognitive enhancement. Phytochem. Rev. 2018;17:161–194. doi: 10.1007/s11101-017-9528-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Brandalise F., Cesaroni V., Gregori A., Repetti M., Romano C., Orrù G., et al. Dietary supplementation of Hericium erinaceus increases mossy Fiber-CA3 hippocampal neurotransmission and recognition memory in wild-type mice evidence-based complement. Alt Med. 2017;2017:3864340. doi: 10.1155/2017/3864340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Johnson S.L., Park H.Y., DaSilva N.A., Vattem D.A., Ma H., Seeram N.P. Levodopa-reduced Mucuna pruriens seed extract shows neuroprotective effects against Parkinson’s disease in murine microglia and human neuroblastoma cells, caenorhabditis elegans, and Drosophila melanogaster. Nutrients. 2018;10:E1139. doi: 10.3390/nu10091139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Matsuda H., Murakami T., Kishi A., Yoshikawa M. Structures of withanosides I, II, III, IV, V, VI, and VII, new withanolide glycosides, from the roots of Indian Withania somnifera DUNAL and inhibitory activity for tachyphylaxis to clonidine in isolated guinea-pig ileum. Bioorg. Med. Chem. 2001;9:1499–1507. doi: 10.1016/s0968-0896(01)00024-4. [DOI] [PubMed] [Google Scholar]
- 125.Manchanda S., Kaur G. Withania somnifera leaf alleviates cognitive dysfunction by enhancing hippocampal plasticity in high fat diet induced obesity model. BMC Complement. Altern. Med. 2017;17:136. doi: 10.1186/s12906-017-1652-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Thangthaeng N., Poulose S.M., Gomes S.M., Miller M.G., Bielinski D.F., Shukitt-Hale B. Tart cherry supplementation improves working memory, hippocampal inflammation, and autophagy in aged rats. Age (Dordr.) 2016;38:393–404. doi: 10.1007/s11357-016-9945-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Malkova L, Kozikowski AP, Gale K. The effects of huperzine A and IDRA 21 on visual recognition memory in young macaques. Neuropharmacology. 2011;60:1262–8. doi: 10.1016/j.neuropharm.2010.12.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Shang Y., Wang L., Li Y., Gu P.F. Vinpocetine improves scopolamine induced learning and memory dysfunction in C57 BL/6J mice. Biol. Pharm. Bull. 2016;39:1412–1418. doi: 10.1248/bpb.b15-00881. [DOI] [PubMed] [Google Scholar]
- 129.Lardner A.L. Neurobiological effects of the green tea constituent theanine and its potential role in the treatment of psychiatric and neurodegenerative disorders. Nutr. Neurosci. 2014;17:145–155. doi: 10.1179/1476830513Y.0000000079. [DOI] [PubMed] [Google Scholar]
- 130.Takeda A., Sakamoto K., Tamano H., Fukura K., Inui N., Suh S.W., et al. Facilitated neurogenesis in the developing hippocampus after intake of theanine, an amino acid in tea leaves, and object recognition memory. Cell. Mol. Neurobiol. 2011;31:1079–1088. doi: 10.1007/s10571-011-9707-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Van de Rest O., Bloemendaal M., De Heus R., Aarts E. Dose-Dependent effects of oral tyrosine administration on plasma tyrosine levels and cognition in aging. Nutrients. 2017;9:1279. doi: 10.3390/nu9121279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Lu C.L., Tang S., Meng Z.J., He Y.Y., Song L.Y., Liu Y.P., et al. Taurine improves the spatial learning and memory ability impaired by sub-chronic manganese exposure. J. Biomed. Sci. 2014;21:51. doi: 10.1186/1423-0127-21-51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Kim H.Y., Kim H.V., Yoon J.H., Kang B.R., Cho S.M., Lee S., et al. Taurine in drinking water recovers learning and memory in the adult APP/PS1 mouse model of Alzheimer’s disease. Sci. Rep. 2014;4:7467. doi: 10.1038/srep07467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Kobayashi S., Iwamoto M., Kon K., Waki H., Ando S., Tanaka Y. Acetyl-L-carnitine improves aged brain function. Geriatr. Gerontol. Int. 2010;10:S99–S106. doi: 10.1111/j.1447-0594.2010.00595.x. [DOI] [PubMed] [Google Scholar]
- 135.Coombes A.J. Dictionary of Plant Names. London, UK: Hamlyn Books; 1994. [Google Scholar]
- 136.Nakanishi K. Terpene trilactones from Gingko biloba: From ancient times to the 21st century. Bioorg. Med. Chem. 2005;13:4987–5000. doi: 10.1016/j.bmc.2005.06.014. [DOI] [PubMed] [Google Scholar]
- 137.Serrano-García N., Pedraza-Chaverri J., Mares-Sámano J.J., Orozco-Ibarra M., Cruz-Salgado A., Jiménez-Anguiano A., et al. Antiapoptotic effects of EGb 761. Evid. Based Complement. Alternat. Med. 2013;2013:495703. doi: 10.1155/2013/495703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Droy-Lefaix M.T. Effect of the antioxidant action of Ginkgo biloba extract (EGb 761) on aging and oxidative stress. Age (Omaha) 1997;20(3):141–149. doi: 10.1007/s11357-997-0013-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Snitz B.E., O’Meara E.S., Carlson M.C., Arnold A.M., Ives D.G., Rapp S.R., et al. Ginkgo Evaluation of Memory (GEM) study investigators Ginkgo biloba for preventing cognitive decline in older adults: A randomized trial. JAMA. 2009;302:2663–2670. doi: 10.1001/jama.2009.1913. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Chen X., Salwinski S., Lee T.J. Extracts of Ginkgo biloba and ginsenosides exert cerebral vasorelaxation via a nitric oxide pathway. Clin. Exp. Pharmacol. Physiol. 1997;24:958–959. doi: 10.1111/j.1440-1681.1997.tb02727.x. [DOI] [PubMed] [Google Scholar]
- 141.Huang S.Y., Jeng C., Kao S.C., Yu J.J., Liu D.Z. Improved haemorrheological properties by Ginkgo biloba extract (Egb 761) in type 2 diabetes mellitus complicated with retinopathy. Clin. Nutr. 2004;23:615–621. doi: 10.1016/j.clnu.2003.10.010. [DOI] [PubMed] [Google Scholar]
- 142.Huguet F., Tarrade T. Alpha 2-adrenoceptor changes during cerebral ageing. The effect of Ginkgo biloba extract. J. Pharm. Pharmacol. 1992;44:24–27. doi: 10.1111/j.2042-7158.1992.tb14357.x. [DOI] [PubMed] [Google Scholar]
- 143.Hadjiivanova C.I., Petkov V.V. Effect of Ginkgo biloba extract on beta-adrenergic receptors in different rat brain regions. Phytother. Res. 2002;16:488–490. doi: 10.1002/ptr.933. [DOI] [PubMed] [Google Scholar]
- 144.Baeg I.H., So S.H. The world ginseng market and the ginseng (Korea). J. Ginseng Res. 2013;37:1–7. doi: 10.5142/jgr.2013.37.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Geng J, Dong J, Ni H, Lee MS, Wu T, Jiang K, et al. Ginseng for cognition. (2:CD007769. Cochrane Database Syst Rev. 2010 doi: 10.1002/14651858.CD007769.pub2.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Xie J.T., Mchendale S., Yuan C.S. Ginseng and diabetes. Am. J. Chin. Med. 2005;33:397–404. doi: 10.1142/S0192415X05003004. [DOI] [PubMed] [Google Scholar]
- 147.Lee Y., Oh S. Administration of red ginseng ameliorates memory decline in aged mice. J. Ginseng Res. 2015;39:250–256. doi: 10.1016/j.jgr.2015.01.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Attele A.S., Wu J.A., Yuan C.S. Ginseng pharmacology: Multiple constituents and multiple actions. Biochem. Pharmacol. 1999;58:1685–1693. doi: 10.1016/s0006-2952(99)00212-9. [DOI] [PubMed] [Google Scholar]
- 149.Zhang J.T., Qu Z.W., Liu Y., Deng H.L. Preliminary study on the antiamnestic mechanism of ginsenoside Rg1 and Rb1. Eur. J. Pharmacol. 1990;183:1460–1461. [PubMed] [Google Scholar]
- 150.Tsang D., Yeung H.W., Tso W.W., Peck H. Ginseng saponins: Influence on neurotransmitter uptake in rat brain synaptosomes. Planta Med. 1985;3:221–224. doi: 10.1055/s-2007-969463. [DOI] [PubMed] [Google Scholar]
- 151.Jin S.H., Park J.K., Nam K.Y., Park S.N., Jung N.P. Korean red ginseng saponins with low ratios of protopanaxadiol and protopanaxatriol saponin improve scopolamine-induced learning disability and spatial working memory in mice. J. Ethnopharmacol. 1999;66(2):123–129. doi: 10.1016/s0378-8741(98)00190-1. [DOI] [PubMed] [Google Scholar]
- 152.Li W., Fitzloff J.F. HPLC determination of ginsenosides content in ginseng dietary supplements using ultraviolet detection. J. Liq. Chromatogr. Relat. Technol. 2002;25:2485–2500. doi: 10.1081/JLC-120014269. [DOI] [Google Scholar]
- 153.Zhu S., Zou K., Fushimi H., Cai S., Komatsu K. Comparative study on triterpene saponins of Ginseng drugs. Planta Med. 2004;70:666–677. doi: 10.1055/s-2004-827192. [DOI] [PubMed] [Google Scholar]
- 154.Scholey A., Ossoukhova A., Owen L., Ibarra A., Pipingas A., He K., et al. Effects of American ginseng (Panax quinquefolius) on neurocognitive function: An acute, randomised, double-blind, placebo-controlled, crossover study. Psychopharmacology (Berl.) 2010;212:345–356. doi: 10.1007/s00213-010-1964-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Kulkarni R., Girish K.J., Kumar A. Nootropic herbs (medhya rasayana) in ayurveda: An update. Pharmacogn. Rev. 2012;6:147–153. doi: 10.4103/0973-7847.99949. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Sharma P.C., Yelne M.B., Dennis T.J. Database on Medicinal plants used in Ayurveda and Sidha. Vol. 1. New Delhi: CCRAS, Dept. of AYUSH, Ministry of Health and Family Welfare, Govt. of India; 2005. pp. 26–6. [Google Scholar]
- 157.Kuroda M., Mimaki Y., Sashida Y., Mae T., Kishida H., Nishiyama T., et al. Phenolics with PPAR-gamma ligand-binding activity obtained from licorice (Glycyrrhiza uralensis roots) and ameliorative effects of glycyrin on genetically diabetic KK-A(y) mice. Bioorg. Med. Chem. Lett. 2003;13:4267–4272. doi: 10.1016/j.bmcl.2003.09.052. [DOI] [PubMed] [Google Scholar]
- 158.Dhingra D., Parle M., Kulkarni S.K. Memory enhancing activity of Glycyrrhiza glabra in mice. J. Ethnopharmacol. 2004;91:361–365. doi: 10.1016/j.jep.2004.01.016. [DOI] [PubMed] [Google Scholar]
- 159.Tripathi Y.B., Chaurasia S., Tripathi E., Upadhyay A., Dubey G.P. Bacopa monniera Linn as an antioxidant: Mechanism of action. Indian J. Exp. Biol. 1996;34:523–526. [PubMed] [Google Scholar]
- 160.Bhattacharya S.K., Bhattacharya A., Kumar A., Ghosal S. Antioxidant activity of Bacopa monniera in rat frontal cortex, striatum and hippocampus. Phytother. Res. 2000;14:174–179. doi: 10.1002/(sici)1099-1573(200005)14:3<174::aid-ptr624>3.0.co;2-o. [DOI] [PubMed] [Google Scholar]
- 161.Rehni A.K., Pantlya H.S., Shri R., Singh M. Effect of chlorophyll and aqueous extracts of Bacopa monniera and Valeriana wallichii on ischaemia and reperfusion-induced cerebral injury in mice. Indian J. Exp. Biol. 2007;45:764–769. [PubMed] [Google Scholar]
- 162.Anand T., Naika M., Swamy M.S.L., Khanum F. Antioxidant and DNA damage preventive properties of Bacopa monniera (L) wettst. Free Radic. Antioxid. 2011;1:84–90. [Google Scholar]
- 163.Girish S.A., Barabde U., Wadodkar S., Dorle A. Effect of Bramhi Ghrita, an polyherbal formulation on learning and memory paradigms in experimental animals. Indian J. Pharmacol. 2004;36:159–162. [Google Scholar]
- 164.Sutalangka C., Wattanathorn J., Muchimapura S., Thukham-mee W. Moringa oleifera mitigates memory impairment and neurodegeneration in animal model of age-related dementia oxidative. Med Cell Longev. 2013;2013:695936. doi: 10.1155/2013/695936. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Joshi H., Parle M. Cholinergic basis of memory-strengthening effect of Foeniculum vulgare linn. J. Med. Food. 2006;9:413–417. doi: 10.1089/jmf.2006.9.413. [DOI] [PubMed] [Google Scholar]
- 166.Joshi H., Parle M. Nardostachys jatamansiImproves learning and memory in mice. J Med Food. 2006;9:113–8. doi: 10.1089/jmf.2006.9.113. 10.1089/jmf.2006.9.113 [DOI] [PubMed] [Google Scholar]
- 167.Metcalf R.L. Ullmann’s Encyclopedia of Industrial Chemistry. 1989. Insect control. [Google Scholar]
- 168.Lloyd G.K., Williams M. Neuronal nicotinic acetylcholine receptors as novel drug targets. J. Pharmacol. Exp. Ther. 2000;292:461–467. [PubMed] [Google Scholar]
- 169.Levin E.D. Nicotinic systems and cognitive function. Psychopharmacology (Berl.) 1992;108:417–431. doi: 10.1007/BF02247415. [DOI] [PubMed] [Google Scholar]