

Abstract
Insulin-like 3 peptide (INSL3) is a member of the insulin-like peptide superfamily and is the only known physiological ligand of relaxin family peptide receptor 2 (RXFP2), a G protein-coupled receptor (GPCR). In mammals INSL3 is primarily produced both in testicular Leydig cells and in ovarian theca cells, but circulating levels of the hormone are much higher in males than in females. The INSL3/RXFP2 system has an essential role in the development of the gubernaculum for the initial transabdominal descent of the testis and in maintaining proper reproductive health in men. Although its function in female physiology has been less well-characterized, it was reported that INSL3 deletion affects antral follicle development during the follicular phase of the menstrual cycle and uterus function. Since the discovery of its role in the reproductive system, the study of INSL3/RXFP2 has expanded to others organs such as skeletal muscle, bone, kidney, thyroid, brain, and eye. This review aims to summarize the various advances in understanding the physiological function of this ligand-receptor pair since its first discovery and elucidate its future therapeutic potential in the management of various diseases.
Keywords: INSL3, RXFP2, GPCR, reproduction, bone development
Introduction
RXFP2 (previously known as GREAT or LGR8) is a GPCR from the relaxin family peptide receptors, which also contains 3 other members: RXFP1, RXFP3, and RXFP4 (Halls et al., 2007b). Relaxin is the cognate ligand for RXFP1 and is the most studied ligand-receptor pair of the family, being well-known for its vasodilator and antifibrotic properties. The neuropeptide Relaxin-3, the cognate ligand for RXFP3, has a role in stress and feeding responses, while INSL5 is the cognate ligand for RXFP4 and is involved in gut contractility. RXFP1 and RXFP2 are structurally highly similar and share 60% amino acid sequence identity (Halls et al., 2007b). The RXFP2 cognate ligand is INSL3, which is produced as a prepro-hormone which, after removal of the signal peptide and cleavage of the C-peptide, gives rise to the mature active hormone consisting of A and B chains linked by 2 disulfide bonds and an additional disulfide bond within the A chain. INSL3 activation of the RXFP2 receptor causes an increase in cAMP production (Kumagai et al., 2002, Rosengren et al., 2006). Relaxin peptides from some species, like porcine relaxin (Lin et al., 2004) and human H2 relaxin (Halls et al., 2005), are capable of activating this receptor as shown in cAMP assays in vitro, but only at concentrations far above physiological levels. Interestingly, mouse and rat relaxin do not activate RXFP2 (Bathgate et al., 2006, Halls et al., 2005).
The effect of the INSL3/RXFP2 system on male reproductive tract development was first discovered two decades ago in gene-deficient male mice with cryptorchidism or undescended testes (Nef and Parada, 1999, Zimmermann et al., 1999, Overbeek et al., 2001). In humans this genital malformation has an overall incidence of around 1-3% in newborn boys and is more common in those who are born prematurely (Kurz, 2016). Spontaneous resolution of this abnormality by one year of age was reported in less than 10% to more than 50% of affected boys in different studies (Wenzler et al., 2004, Berkowitz et al., 1993). For others, the most common treatment is an orchiopexy, a surgical procedure where the undescended testes are brought into the scrotum. If left untreated, cryptorchidism can lead to infertility and testicular cancer (Kurz, 2016, Ferguson and Agoulnik, 2013). In females, INSL3 is involved in ovarian follicle maturation and could play a role in the pathology of Polycystic Ovary Syndrome (PCOS) (Pelusi et al., 2013). In addition, a recent line of investigation has elucidated the anabolic functions of the INSL3/RXFP2 system in osteoblasts, skeletal muscles, and other organs, highlighting the role of this ligand-receptor pair outside reproductive physiology (De Toni et al., 2019). The unique RXFP2 structural features, cell surface expression, and confined expression pattern make it a potentially desirable pharmacological target for various diseases including osteoporosis and hypogonadism, although more work has to be done to further characterize INSL3/RXFP2 signaling and functions.
Mechanisms of INSL3 binding and activation of RXFP2
INSL3 treatment of HEK293T cells transfected with RXFP2 causes activation of the classical adenylyl cyclase pathway via activation of Gαs, resulting in increased cAMP production. At its N-terminus, RXFP2 contains a large extracellular domain with a low-density lipoprotein domain (LDLa) joined with a linker to a ten leucine rich repeat domain (LRR), followed by a seven transmembrane helical domain (TM) (Figure 1). Within the receptor family, the LDLa module is only present in RXFP1 and RXFP2 (Halls et al., 2007b). Mutagenesis studies have shown the importance of the module for receptor expession in the cell surface and cell signaling (Bogatcheva et al., 2007, Scott et al., 2006). Mutation of amino acid residues C71 and D70, which are involved in Ca-stabilization of the LDLa molecule, have been shown to decrease cell surface expression of the RXFP2 receptor and abrogate cAMP production in response to INSL3 (Bogatcheva et al., 2007). Moreover, a RXFP2 splice variant expressed in human uterine tissue that lacks the LDLa module had intact ligand binding affinity but an impaired cAMP signaling when tested in HEK293-RXFP2 cells (Scott et al., 2006). Recent studies have shown that upon receptor binding to INSL3, the RXFP2 linker region located between the LDLa and LRRs may help to direct the LDLa domain to interact with the TM and activate the receptor together with the INSL3 A-chain (Bruell et al., 2017).
Figure 1. INSL3/RXFP2 binding-activation model and signaling mechanisms.
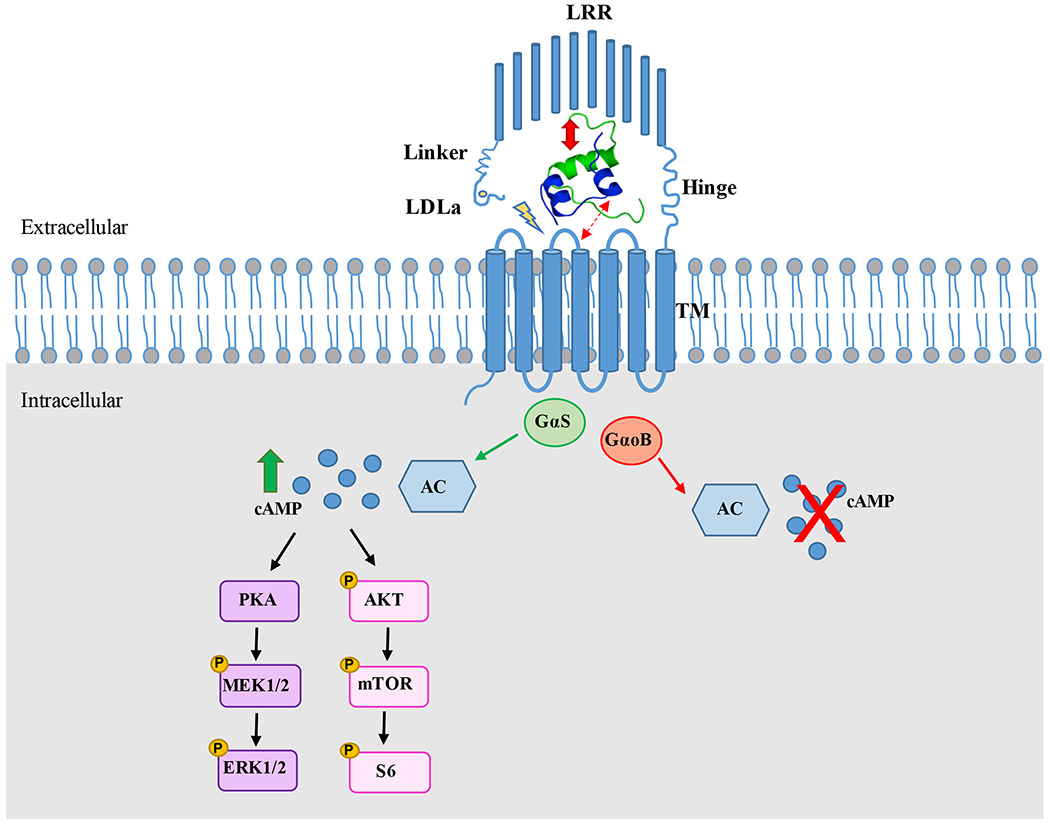
The RXFP2 receptor consists of a low-density lipoprotein (LDLa), 10 leucine rich repeat domains (LRR) and 7 transmembrane helical domains (TM). The mature INSL3 hormone is formed by a B-chain and A-chain joined by disulfide bonds. The predicted INSL3/RXFP2 binding model shows a B-chain high affinity-binding site in the LRR (bold arrow), as well as an A-chain low affinity-binding site in the extracellular loops of the TM (dotted arrow). After binding, the Linker region helps direct the LDLa module towards the extracellular loops of the TM to activate the receptor, together with the N-terminal region of the INSL3 A-chain (lightning symbol). INSL3 activates the RXFP2 receptor causing coupling to the Gαs subunit, which activates adenylyl cyclase and increases cAMP levels. This signaling mechanism has been proven in mouse Leydig and C2C12 cells, rat gubernacular cells, MG63 cells and primary human osteoblasts. cAMP downstream signaling through phosphorylation of AKT, mTOR, and S6 has been found in C2C12 cells, as well as activation of the MAPK/ERK pathway in primary human osteoblasts. In rat female and male germ cells, INSL3 activation of RXFP2 has been shown to cause coupling to the GαoB subunit, which inhibits adenylyl cyclase and decreases cAMP levels. INSL3 structure is from PDBe (entry 2H8B).
Several structure-activity relationship studies (SAR) based on homology models have characterized the RXFP2 structure and interactions with its ligand INSL3, however no high-resolution crystal structure of the receptor has been solved to date. A model was proposed where the B chain of INSL3 binds to high-affinity binding sites in the LRRs of RXFP2, while the A chain of INSL3 binds to low-affinity binding sites in the TM (Halls et al., 2005) (Figure 1). Mutation analysis has shown that specific residues from the LRR domain are essential for receptor cell surface expression and binding of the receptor with the B-chain of INSL3 (Scott et al., 2007). A human uterine splice variant of RXFP2 lacking one LRR repeat was found incapable of binding INSL3 and H2 relaxin (Muda et al., 2005), emphasizing the crucial role of the LRR domains in ligand binding. A study using chimeric RXFP2 receptors also demonstrated that there is a high-affinity INSL3 binding site in the LRR domain (Halls et al., 2005). When the LRR of RXFP2 was replaced with the RXFP1 LRR, the affinity of the receptor was lower than the wild-type (WT) RXFP2 receptor (Halls et al., 2005). Studies using RXFP2 chimeric receptors also suggested that there is a low-affinity INSL3 binding site in the extracellular loops of the TM domain, since RXFP2 chimeras with the LRR and extracellular loops replaced with RXFP1 were unable to bind INSL3 or induce cAMP production (Sudo et al., 2003).
In INSL3, H36, R40, V43, R44, and T53 located within the B-chain form a receptor binding motif, with T53 appearing to be the most crucial (Rosengren et al., 2006, Bullesbach and Schwabe, 1999, Shabanpoor et al., 2010). Shortened mimetics of the B-chain demonstrated micromolar range binding affinity to RXFP2 and antagonistic activity when used in combination with INSL3 (Del Borgo et al., 2006, Shabanpoor et al., 2007). More potent antagonists based on the B-chain sequence have been designed through systematic efforts to induce α-helicity in the isolated INSL3 B-chain by constraining the B-chain with a short region of A-chain or dimerizing the B-chain (Shabanpoor et al., 2010).
RXFP2 cellular signaling
Signal transduction has been primarily studied in HEK293T cells transfected with RXFP2. Stimulation of RXFP2 in these cells results in its coupling to Gαs, which leads to cAMP production and further activates CRE-dependent gene transcription (Halls et al., 2007a). On the other hand, coupling of GαoB also occurs, which is involved in negative modulation of cAMP accumulation in HEK293T cells (Halls et al., 2006). Unlike in relaxin/RXFP1 signaling, there is no evidence of RXFP2 signaling in HEK293T cells through the Gαi3 isoform, which inhibits cAMP production (Halls et al., 2009) (Figure 1).
Additional studies performed on cells that endogenously express RXFP2 have provided insight into cell-dependent responses to INSL3 treatment. Rat gubernacular and mouse Leydig cells showed increased production of cAMP upon stimulation with INSL3 (Kumagai et al., 2002, Pathirana et al., 2012) while the opposite effect was reported in rat female and male primary germ cells (Kawamura et al., 2004). Stimulation with INSL3 resulted in an inhibition of intraoocyte cAMP levels and forskolin-induced cAMP in seminiferous tubular cells. Studies using myotubes derived from C2C12 cells have shown increased phosphorylation of AKT, mTOR, and S6 after INSL3 treatment, identifying a possible downstream cAMP pathway activated by INSL3/RXFP2 (Ferlin et al., 2018). In the human osteoblast cell line MG-63, INSL3 stimulation also increased intracellular cAMP production (Ferlin et al., 2008). Further investigations of INSL3/RXFP2 signaling in primary human osteoblasts have shown increased MEK and ERK1/2 phosphorylation via the AC/cAMP/PKA pathway (Ferlin et al., 2011). Surprisingly, a classical regulatory mechanism of GPCR signaling, beta-arrestin-induced internalization and desensitization, does not appear to be a major factor in RXFP2 regulation. RXFP2 has a sustained cAMP response when stimulated with INSL3, which may be due to reduced recruitment of beta-arrestin to the cell surface and attenuated internalization of the receptor (Callander et al., 2009).
To date, only a few studies have been performed to analyze whole transcriptome changes in response to INSL3. To identify downstream pathways activated by INSL3/RXFP2 signaling, rat gubernacular bulb cells isolated from embryos on day 17 were treated with INSL3 and analyzed using Affymetrix microarrays (Johnson et al., 2010). Male gubernacular bulb cells responded to INSL3 with changes in expression of genes involved in cAMP, BMP, WNT/β-catenin, pluripotency and several other signaling pathways, including genes playing a role in osteoblasts, osteoclasts, and chondrocyte participation in rheumatoid arthritis. As expected, Gene Ontology (GO) analysis revealed changes in genes involved in GPCR signaling, extracellular, and plasma membrane functions. In addition, several GO related to neurogenesis were altered by INSL3 (Johnson et al., 2010).
More studies need to be conducted to fully understand the signaling mechanisms of INSL3/RXFP2 in various cells and their biological significance. This is also important for efficient targeting of this receptor in future therapeutic applications, providing for specificity of response while avoiding potential harmful off-target effects.
INSL3 in male reproductive physiology
In mammalian male reproductive organs, INSL3 is produced mainly in the testicular Leydig cells (Balvers et al., 1998, Zarreh-Hoshyari-Khah et al., 1999, Ivell et al., 1997, Pitia et al., 2017) and RXFP2 is expressed in testis in Leydig cells as well as in germ cells, especially at the post-meiotic stage (Feng et al., 2007, Pitia et al., 2017, Huang et al., 2012). INSL3 is considered an indicator of normal Leydig cell function and overall reproductive health in men (Ivell et al., 2013). Human male fetuses produce INSL3 during gestation, measured at approximately 0.12 ng/ml in amniotic fluid (Bay et al., 2008). This level of circulating INSL3 is maintained from birth until the age of 3 months, at which point it begins to decrease (Bay et al., 2007). The INSL3 level rises again through puberty and is sustained between 0.5-1 ng/ml in adult men (Bay et al., 2005). During puberty, luteinizing hormone (LH) drives Leydig cell maturation, which coincides with a spike in INSL3 production that positively correlates with an increase of sex hormones such as follicle stimulating hormone (FSH), LH, and testosterone (Ferlin et al., 2006). Low INSL3 serum levels are characterisitic of orchidectomized and infertile men, individuals with Klinefelter’s syndrome, hypogonadotropic hypogonadism, and cryptorchidism (Ferlin and Foresta, 2005, Bay et al., 2005). Males with anorchism have no detectable levels of INSL3 (Bay et al., 2005). To further investigate the correlation between INSL3 and LH, subjects with unilateral orchiectomy were treated with a single dose of human chorionic gonadotropin (hCG), resulting in elevated testosterone levels around 3 days after treatment but no impact on INSL3 levels (Bay et al., 2005). However, when hypogonadotropic hypogonadism subjects were treated with repeated doses of hCG over several months, INSL3 levels increased (Bay et al., 2005). In agreement with these findings, normal human male subjects who were gonadotropin deprived had decreased INSL3 levels. Such patients had spontaneous partial recovery of INSL3 to 38.9% of baseline as opposed to an almost complete recovery of testosterone at 80.2% (Bay et al., 2006). It was also shown that direct hCG stimulation of Leydig cells in vitro does not increase INSL3 expression (Sadeghian et al., 2005). These findings suggest that long-term LH stimulation of Leydig cells is required for INSL3 expression but testosterone production is not co-regulated with INSL3 in mature Leydig cells.
Testicular descent during embryogenesis consists of a transabdominal phase followed by an inguinoscrotal phase. The INSL3/RXFP2 system has a determining role in the development of the gubernaculum ligament and testicular descent during the transabdominal phase (Huang et al., 2012, Nef and Parada, 1999, Overbeek et al., 2001, Zimmermann et al., 1999). The inguinoscrotal phase of testes descent is known to be androgen mediated, but studies have also shown the possible synergistic involvement of INSL3/RXFP2 during this phase (Yuan et al., 2010). Genetic ablation of Insl3 in mice has resulted in various degrees of intraabdominal cryptorchidism, the most common congenital birth defect in newborn boys. Insl3-/- adult mice showed decreased testes size, lesions in the seminiferous tubules, absence of spermatid and mature sperm, and gubernaculum underdevelopment (Nef and Parada, 1999, Zimmermann et al., 1999). Interestingly, Insl3-/- males had normal copulatory behavior, normal seminal vesicles and prostate weights, which suggests that the mutants had normal androgen production and that this form of cryptorchidism is androgen independent (Nef and Parada, 1999, Zimmermann et al., 1999). On the other hand, transgenic female mice overexpressing Insl3 develop bilateral inguinal hernias due to the descent of the ovaries into the processus vaginalis via outgrowth of the gubernaculum (Adham et al., 2002).
Rxfp2 involvement in cryptorchidism was discovered in mutant mice with a 550kb deletion containing the gene in chromosome 5 (Overbeek et al., 2001). Breeding of these mutant mice, known as crsp, resulted in bilateral intraabdominal cryptorchidism in male homozygotes. All crsp/crsp adult males had small testes size and epididymis, absence of spermatogenesis, vacuolization of Sertoli cells, lesions of the seminiferous tubules and were infertile (Overbeek et al., 2001). Recently, more sophisticated genetic tools were used to generate transgenic mouse lines to further prove the involvement of Insl3 and Rxfp2 in testicular descent. Rxfp2 with LacZ reporter allowed for detection of Rxfp2 expression in gubernaculum, Leydig cells, and postmeiotic spermatogenic cells in testis (Huang et al., 2012). Cryptorchidism was also observed when Rxfp2 was deleted specifically in gubernacular embryonic mesenchymal tissue using a Cre/loxP approach, but the abnormal phenotype was not found after deletion of Rxfp2 in gubernacular striated or smooth muscle cells (Huang et al., 2012). This finding further demonstrates the importance of Rxfp2 for testicular transabdominal descent via regulation of the outgrowth of the gubernaculum in mice.
INSL3/RXFP2 signalling induces WNT and BMP developmental pathways and drives morphogenetic changes in gubernaculum (Johnson et al., 2010). It was shown that Rxfp2-/- male mice embryos experienced a dramatic decrease in expression of developmental signaling molecules β-catenin, NOTCH1, and WNT1 in the gubernaculum (Kaftanovskaya et al., 2011). The disruption of normal RXFP2 expression resulted in the failure of the testes to descend away from the caudal pole of the kidney at day E16.5, as well as an underdeveloped gubernaculum (Kaftanovskaya et al., 2011). To further understand the pathways leading to the failure of gubernacular development, β-catenin or Notch1 was ablated in the gubernacular ligament. In these males, the gubernacular ligament had a lack of muscle layers, which was consistent with previous observations in Rxfp2-deficient mice (Kaftanovskaya et al., 2011).
Recently, a mammalian phylogenetic study in Afrotherians revealed that 4 testicond species from this lineage had loss of functional RXFP2 and INSL3, which correlates with a lack of gubernaculum and testes descent, demonstrating the highly evolutionarily conserved function of this ligand-receptor pair (Sharma et al., 2018).
Orchidopexy in Insl3−/− and crsp/crsp male mice recovered spermatogenesis at least partially, suggesting that the increased testicular temperature as a result of cryptorchidism may be a contributing factor for the defects observed in mouse male germ cell differentiation (Zimmermann et al., 1999, Overbeek et al., 2001). Additionally, conditional deletion of Rxfp2 in germ cells beginning from premeiotic stages did not affect spermatogenesis, fertility, or germ cell survival in adult male mice (Huang et al., 2012).
Although studies in mice showed that deletion of Insl3 or Rxfp2 does not appear to have a direct impact on spermatogenesis or germ cell survival, other studies have drawn links between INSL3 levels and improved survivability of germ cells. Studies in gonadotropin compromised male rats showed the ability of INSL3 to suppress germ cells apoptosis (Kawamura et al., 2004). In a human contraceptive study, subjects who were given a combination of testosterone and progesterone as contraceptive treatment for 24 weeks but did not display azoospermia had higher serum INSL3 levels compared to those who were azoospermic (Amory et al., 2007). This suggests that INSL3 may prevent apoptosis in male germ cells as non-azoospermic subjects also have higher sperm concentration, which positively correlated with INSL3 serum levels (Amory et al., 2007). Treatment of boar with neutralizing antibody against INSL3 also showed similar results, in that lower levels of available INSL3 reduced anti-apoptotic XIAP and BCL2 levels and sperm concentration (Minagawa et al., 2018). Taken together, these studies demonstrate that INSL3 may play a role in regulating apoptosis and turnover of male germ cells, especially when spermatogenesis is under stress conditions.
Clinical case studies in cryptorchid patients have identified dozens of mutations in INSL3 and RXFP2 genes (El Houate et al., 2007, Bogatcheva et al., 2003, Bogatcheva et al., 2007, Gorlov et al., 2002, Ayers et al., 2019) (Figure 2). Functional analysis of these INSL3 mutants in cells transfected with the RXFP2 receptor showed a decreased cAMP response for V18M, P49S, R73X and R102C mutants, suggesting potential clinical relevance (El Houate et al., 2007, Bogatcheva et al., 2003). A recent genetic analysis in patients with testicular torsion has also described mutations in the INSL3 gene, including a T60A polymorphism in the C-peptide region (Capra et al., 2018), a common polymorphism that was previously identified with the same frequency in cryptorchid and control patients (Lim et al., 2001). A missense mutation variant T222P in the LRR of RXFP2 impairs receptor expression on the cell surface and has been strongly associated with cryptorchidism (Bogatcheva et al., 2007, Gorlov et al., 2002). However, a more recent screen of cryptorchidic subjects in Spanish and Italian populations revealed that carrying this mutation may lead to increased risk of cryptorchidism only in the Italian population (Ars et al., 2011). Another missense mutation, G499E, was identified in the third TM domain of RXFP2 by conducting whole exome sequencing of 4 brothers who presented with bilateral cryptorchidism (Ayers et al., 2019). A mutagenesis study further validated that G499E is a loss of function mutation. The G499E RXFP2 receptor showed no ligand binding or activation when stimulated with INSL3, and the mutant receptor expression on the cell surface was about 12% of the WT RXFP2 (Ayers et al., 2019). Significantly, in all studies only heterozygous mutant carriers were found. Thus, while mutation analysis in cryptorchid patients suggests a possible contribution of INSL3 and RXFP2 in this abnormality in men, further studies are necessary to prove a direct cause-effect relationship.
Figure 2. Mutation sites in INSL3 and RXFP2 genes found in cryptorchid patients.
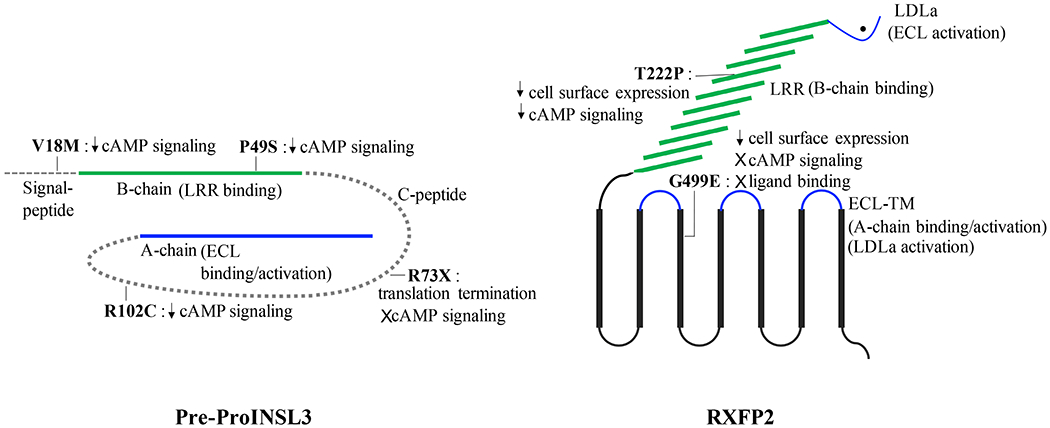
Several mutations in INSL3 and RXFP2 have been found clinically relevant based on mutagenesis studies. INSL3 mutants V18M, P49S and R102C have decreased cAMP responses compared to WT INSL3. The mutant R73X produces a truncated INSL3 containing only the B-chain region of the peptide, which is insufficient to induce cAMP signaling. RXFP2 mutants T222P and G499E are not well expressed on the cell surface, which could explain the signaling impairment seen during functional analysis. Mutations INSL3-P49S and RXFP2-T222P are of great importance due to their location in the B-chain-LRR binding site. The newly discovered mutant RXFP2-G499E has void cAMP signaling and binding, suggesting its potential importance in the binding and activation site located in the extracellular loops of the transmembrane domain (ECL-TM).
INSL3 in female reproductive physiology
In females INSL3 is produced primarily in the follicular theca interna cells of the ovary, where RXFP2 expression is also found (Bamberger et al., 1999, Kawamura et al., 2004, Zarreh-Hoshyari-Khah et al., 1999, Dai et al., 2017b). Lower levels of INSL3 and RXFP2 expression have also been detected in the corpus luteum and uterus (Bamberger et al., 1999, Balvers et al., 1998, Zarreh-Hoshyari-Khah et al., 1999, Dai et al., 2017b, Li et al., 2011). INSL3 in healthy women is first detectable during the late stages of puberty (Hagen et al., 2015), and serum levels remain around 79 pg/ml, which is much lower than in men (Anand-Ivell et al., 2013). INSL3 levels in women fluctuate in a phasic manner throughout the menstrual cycle and become undetectable after menopause (Anand-Ivell et al., 2006). The secretion of INSL3 is at its lowest levels during menses and spikes to significantly higher levels during the follicular phase of the menstrual cycle, concordant with the recruitment of growing antral follicles (Anand-Ivell et al., 2013). The highest levels of INSL3 positively correlated with pre-ovulatory ovarian hormones such as Anti-Müllerian hormone (AMH) and Inhibin B (Anand-Ivell et al., 2013). The LH spike during ovulation negatively correlates with INSL3 and pre-ovulatory hormones and this is a causal relationship that was also described in studies on bovine theca cells (Anand-Ivell et al., 2013, Dai et al., 2017a). In primary bovine theca interna cells, LH at low doses stimulates production of INSL3, but higher LH levels comparable to those measured during ovulation had an inhibitory effect on INSL3 production (Dai et al., 2017a). Through further testing, it was revealed that INSL3 production is stimulated by LH and estradiol, acting through the protein kinase A pathway (Dai et al., 2017a). A feedback loop between INSL3 and steriodogenesis in theca cells was established when siRNA knockdown of RXFP2 sharply reduced CYP17A1 expression and androstenedione secretion (Glister et al., 2013). Further, the inhibition of CYP17A1 reduced androgen secretion and INSL3 and RXFP2 expression. Additionally, treatment of theca cells with BMP6 dramatically down-regulated expression of INSL3 along with CYP17A1 and several other key steroidogenesis genes. Taken together, these studies showed that estrogen-induced INSL3 production is a key mediator during follicular phase steroidogenesis, and levels of LH that peak at ovulation negatively regulate INSL3.
Abnormal levels of INSL3 in woman have been shown to correlate with Polycystic Ovary Syndrome, PCOS (Seyam and Hefzy, 2018, Shaikh et al., 2016, Pelusi et al., 2013). Patients with PCOS were categorized by their menstrual cycles into amenorrheic, eumenorrheic, or oligomenorrheic (Pelusi et al., 2013). Amenorrheic and oligomenorrheic groups had significantly higher levels of INSL3 and AMH compared to control women, which suggests a potential role in the follicle arrest and anovulation typically seen in this disease (Pelusi et al., 2013). Women with abnormal anatomical characteristics such as ectopic ovaries were also identified from a sample of PCOS women with positively correlated INSL3 and androgen levels. (Seyam and Hefzy, 2018). A recent study suggested that the common polymorphism T60A of the INSL3 gene could increase the risk of developing PCOS (Shaikh et al., 2016). Although this finding would need to be confirmed, it provides an argument towards a relationship between INSL3 and development of PCOS.
Rodent models have also proved useful in establishing the role of the INSL3/RXFP2 system in female reproduction (Nef and Parada, 1999, Li et al., 2011, Kawamura et al., 2004) . Deletion of the Insl3 gene resulted in abnormal estrous cycle and reduced fertility (Nef and Parada, 1999) further demonstrating the involvement of INSL3 in maintaining healthy reproductive functions. Rxfp2 and Caveolin 1 (Cav1) have been associated with epithelial and stromal cell growth and overall homeostasis in the mouse uterus, as the rate of uterine cyst development sharply increased when both Cav1 and Rxfp2 were deleted in females (Li et al., 2011). INSL3 induces oocyte maturation in rats, shown by a dose-dependent increase in germinal vesicle breakdown (GVBD) after INSL3 treatment (Kawamura et al., 2004).
Role of INSL3 in bone and skeletal muscle physiology
INSL3/RXFP2 signaling has been shown to play a role in maintaining normal bone characteristics in both mouse and human (Ferlin et al., 2011, Ferlin et al., 2008). Young men with cryptorchidism carrying the T222P mutation in the RXFP2 gene have significantly reduced bone density, resulting in osteopenia and osteoporosis (Ferlin et al., 2008). Expression of RXFP2 has been shown in human osteoblasts and osteocytes, as well as in mouse osteoblasts (Ferlin et al., 2008, Di Nisio et al., 2018) Treatment of primary human osteoblasts with INSL3 showed a dose-dependent increase in proliferation (Ferlin et al., 2008). Moreover, femurs from Rxfp2−/− mice had significantly decreased bone mass and trabecular number compared to WT mice. A histomorphometric analysis of the lumbar spine of mutant mice also revealed reduced bone formation rate and mineralization surface compared to WT (Ferlin et al., 2008). Furthermore, INSL3 has been shown to regulate the expression of genes involved in differentiation and maturation of primary human osteoblasts, such as ALP, COL1A1, COL6A1, and Osteonectin (Ferlin et al., 2011). Treatment of primary human osteoblasts with INSL3 improved mineralization of the bone matrix (Ferlin et al., 2011). Decreased INSL3 levels in Klinefelter’s syndrome patients are correlated with increased levels of serum sclerostin, which is involved in bone catabolism by inhibiting osteoblasts differentiation and stimulating osteoclasts activation (Di Nisio et al., 2018). This relationship was further explored in cultured osteocytes, which had reduced expression of sclerostin when treated with INSL3. The negative correlation between INSL3 and sclerostin provides insights into the impact of INSL3 on bone health and its potential therapeutic value (Di Nisio et al., 2018).
RXFP2 was also suggested to have an important role in maintaining proper function of muscular tissues (Ferlin et al., 2018). INSL3 treatment of myotubes differentiated from C2C12 skeletal muscle cells resulted in increased cell size compared to untreated control. Expression of myosin heavy chain can be induced by INSL3, which leads to increased protein synthesis in these cells (Ferlin et al., 2018). Denervated muscles in Rxfp2 -/- mice developed greater muscle loss compared to WT mice. Additionally, the tibialis anterior muscle had significantly decreased beta-oxidative fibers, while the soleus muscle showed a significant decrease in fast and slow fibers in these mice. The soleus muscle absolute force, when normalized to the muscle mass of Rxfp2−/− mice, was also decreased compared to WT mice (Ferlin et al., 2018).
The emerging role of INSL3/RXFP2 in the musculoskeletal system reveals potential new targets for INSL3 and synthetic RXFP2 agonists in the treatment of diseases associated with bone and muscle loss.
INSL3 role in other organs
In adult rat forebrain, high Rxfp2 gene expression has been found in the thalamus, frontal and motor cortices. Receptor autoradiography showed INSL3 radioligand binding of RXFP2 in the thalamus and striatum nucleus (Sedaghat et al., 2008), suggesting a potential role of the INSL3/RXFP2 system in motor and sensory brain functions. Additionally, INSL3 and RXFP2 in the eye may play a role in wound healing (Hampel et al., 2013). Protein expression has been found in the human and mouse ocular surface and tears. A scratch test assay performed on human conjunctival and corneal epithelial cells showed increased migration and proliferation when treated with INSL3 (Hampel et al., 2013). A mouse corneal ulcer model was used to test topical application of INSL3, which was determined to be effective in re-epithelialization and healing of corneal wounds (Hampel et al., 2013). INSL3 radioligand binding was also detected in the glomeruli of the renal cortex of post-natal and adult rats (Fu et al., 2006). It has been proposed that INSL3 in the kidney inhibits glomerular cell proliferation, which may be beneficial in targeting glomerular diseases that are associated with uncontrolled mesangial cell proliferation (Fu et al., 2006).
The INSL3/RXFP2 system has also been tested in cancer pathology (Klonisch et al., 2005, Hombach-Klonisch et al., 2010). INSL3 in situ hybridization and immunoreactivity was shown in benign prostate hyperplasia and neoplasia. Stimulation of human prostate carcinoma cell line PC-3 with INSL3 resulted in cAMP production and showed a positive correlation with increased cell migration (Klonisch et al., 2005). RXFP2 expression has also been detected in human thyroid carcinoma tissues (Hombach-Klonisch et al., 2010). Expression of INSL3 in xenotransplants with the FTC133 human thyroid carcinoma cell line showed increased tumor growth in nude mice compared to FTC133 cells not transfected with INSL3 (Hombach-Klonisch et al., 2010). Additionally, in vitro INSL3 treatment of these tumor cells led to increased motility, which is indicative of enhanced tumor metastatic capacity. Treatment of HUVECs with INSL3 also showed significant tube formation, similar to the VEGF control, which suggested promotion of angiogenesis (Hombach-Klonisch et al., 2010). As future studies elucidate the role of the INSL3/RXFP2 in these systems, there may be additional opportunities for therapeutic intervention with RXFP2 agonists or antagonists.
Conclusion and future directions
The role and involvement of the INSL3/RXFP2 system in health and disease has greatly expanded beyond the initial finding of its role in testicular descent (Table 1). Recent studies suggest the importance of this system for maintaining overall bone and skeletal muscle health, which may hold great potential for treating common age-related disorders associated with osteoporosis or muscle loss. In addition, links to PCOS, corneal healing and several cancers have also been described, further underscoring the importance of INSL3/RXFP2 signaling outside reproductive physiology. Therefore, further efforts should be focused on defining INSL3 functions in human physiology along with designing pharmaceuticals that target this signaling system.
Table 1.
Summary of the known physiological roles of the INSL3/RXFP2 system and related pathologies.
| Organ | Physiology | Pathology |
|---|---|---|
| Testis | - Transabdominal phase of testis descent - Gubernaculum development - Germ cell survival - Leydig cells maturation marker |
- Cryptorchidism |
| Ovary | - Recruitment of antral follicles - Oocyte maturation |
- PCOS |
| Bone | - Maintain bone density - Osteoblast maturation |
- Osteopenia/Osteoporosis |
| Skeletal Muscle | - Maintain skeletal muscle strength | - Muscle wasting |
| Eye | - Corneal healing | - Unknown |
| Kidney | - Inhibition of glomerular cell proliferation | - Unknown |
| Prostate | - Unknown | - Prostate carcinoma |
| Thyroid | - Unknown | - Thyroid carcinoma |
To date, no clinical studies have been conducted targeting INSL3 or RXFP2, nor has a systemic analysis of recombinant INSL3 stability in vivo been reported. Studies of the effect of INSL3 injections on testicular functions in rats have shown that INSL3 can pass through the blood-testis barrier (Anand-Ivell et al., 2009) and protect against GnRH antagonist-induced apoptosis in germ cells (Kawamura et al., 2004). Various approaches previously used to improve the stability of relaxin (Nagorniewicz et al., 2019, Muppidi et al., 2019, Sun et al., 2019) can be applied to INSL3 to design biologicals with the full spectrum of INSL3/RXFP2 downstream signaling. Alternatevely, small molecule agonists have proven to be an attractive alternative to therapies with peptide ligands due to improved stability and potential oral bioavailability. A high throughput screening of small molecules successfully identified the first RXFP1 agonist, compound ML290, which has antifibrotic effects recently demonstrated in a mouse model of liver fibrosis (Kaftanovskaya et al., 2019). Due to the structural similarities of RXFP1 and RXFP2, the same approach that was used for the selection and optimization of ML290 can be applied to identify a small molecule agonist that specifically targets RXFP2. The development of such stable agonists will provide not only novel insights into RXFP2 signaling mechanisms, but potentially also become useful pharmacological agents with various clinical applications.
Acknowledgements
The authors thank Ms Courtney Myhr for editorial comments.
Funding
This work was supported by the National Institute of Arthritis and Musculoskeletal and Skin Diseases/National Institutes of Health grant 1R01AR070093 to AIA.
Footnotes
Declaration of interest
The authors declare that there is no conflict of interest that could be perceived as prejudicing the impartiality of this review.
References
- ADHAM IM, STEDING G, THAMM T, BULLESBACH EE, SCHWABE C, PAPROTTA I & ENGEL W 2002. The overexpression of the insl3 in female mice causes descent of the ovaries. Mol Endocrinol, 16, 244–52. [DOI] [PubMed] [Google Scholar]
- AMORY JK, PAGE ST, ANAWALT BD, COVIELLO AD, MATSUMOTO AM & BREMNER WJ 2007. Elevated end-of-treatment serum INSL3 is associated with failure to completely suppress spermatogenesis in men receiving male hormonal contraception. J Androl, 28, 548–54. [DOI] [PubMed] [Google Scholar]
- ANAND-IVELL R, HENG K, HAFEN B, SETCHELL B & IVELL R 2009. Dynamics of INSL3 peptide expression in the rodent testis. Biol Reprod, 81, 480–7. [DOI] [PubMed] [Google Scholar]
- ANAND-IVELL R, TREMELLEN K, DAI Y, HENG K, YOSHIDA M, KNIGHT PG, HALE GE & IVELL R 2013. Circulating insulin-like factor 3 (INSL3) in healthy and infertile women. Hum Reprod, 28, 3093–102. [DOI] [PubMed] [Google Scholar]
- ANAND-IVELL R, WOHLGEMUTH J, HAREN MT, HOPE PJ, HATZINIKOLAS G, WITTERT G & IVELL R 2006. Peripheral INSL3 concentrations decline with age in a large population of Australian men. Int J Androl, 29, 618–26. [DOI] [PubMed] [Google Scholar]
- ARS E, LO GIACCO D, BASSAS L, NUTI F, RAJMIL O, RUIZ P, GARAT JM, RUIZ-CASTANE E & KRAUSZ C 2011. Further insights into the role of T222P variant of RXFP2 in non-syndromic cryptorchidism in two Mediterranean populations. Int J Androl, 34, 333–8. [DOI] [PubMed] [Google Scholar]
- AYERS K, KUMAR R, ROBEVSKA G, BRUELL S, BELL K, MALIK MA, BATHGATE RA & SINCLAIR A 2019. Familial bilateral cryptorchidism is caused by recessive variants in RXFP2. J Med Genet. [DOI] [PMC free article] [PubMed] [Google Scholar]
- BALVERS M, SPIESS AN, DOMAGALSKI R, HUNT N, KILIC E, MUKHOPADHYAY AK, HANKS E, CHARLTON HM & IVELL R 1998. Relaxin-like factor expression as a marker of differentiation in the mouse testis and ovary. Endocrinology, 139, 2960–70. [DOI] [PubMed] [Google Scholar]
- BAMBERGER AM, IVELL R, BALVERS M, KELP B, BAMBERGER CM, RIETHDORF L & LONING T 1999. Relaxin-like factor (RLF): a new specific marker for Leydig cells in the ovary. Int J Gynecol Pathol, 18, 163–8. [PubMed] [Google Scholar]
- BATHGATE RA, LIN F, HANSON NF, OTVOS L JR., GUIDOLIN A, GIANNAKIS C, BASTIRAS S, LAYFIELD SL, FERRARO T, MA S, et al. 2006. Relaxin-3: improved synthesis strategy and demonstration of its high-affinity interaction with the relaxin receptor LGR7 both in vitro and in vivo. Biochemistry, 45, 1043–53. [DOI] [PubMed] [Google Scholar]
- BAY K, COHEN AS, JORGENSEN FS, JORGENSEN C, LIND AM, SKAKKEBAEK NE & ANDERSSON AM 2008. Insulin-like factor 3 levels in second-trimester amniotic fluid. J Clin Endocrinol Metab, 93, 4048–51. [DOI] [PubMed] [Google Scholar]
- BAY K, HARTUNG S, IVELL R, SCHUMACHER M, JURGENSEN D, JORGENSEN N, HOLM M, SKAKKEBAEK NE & ANDERSSON AM 2005. Insulin-like factor 3 serum levels in 135 normal men and 85 men with testicular disorders: relationship to the luteinizing hormone-testosterone axis. J Clin Endocrinol Metab, 90, 3410–8. [DOI] [PubMed] [Google Scholar]
- BAY K, MATTHIESSON KL, MCLACHLAN RI & ANDERSSON AM 2006. The effects of gonadotropin suppression and selective replacement on insulin-like factor 3 secretion in normal adult men. J Clin Endocrinol Metab, 91, 1108–11. [DOI] [PubMed] [Google Scholar]
- BAY K, VIRTANEN HE, HARTUNG S, IVELL R, MAIN KM, SKAKKEBAEK NE, ANDERSSON AM & TOPPARI J 2007. Insulin-like factor 3 levels in cord blood and serum from children: effects of age, postnatal hypothalamic-pituitary-gonadal axis activation, and cryptorchidism. J Clin Endocrinol Metab, 92, 4020–7. [DOI] [PubMed] [Google Scholar]
- BERKOWITZ GS, LAPINSKI RH, DOLGIN SE, GAZELLA JG, BODIAN CA & HOLZMAN IR 1993. Prevalence and natural history of cryptorchidism. Pediatrics, 92, 44–9. [PubMed] [Google Scholar]
- BOGATCHEVA NV, FERLIN A, FENG S, TRUONG A, GIANESELLO L, FORESTA C & AGOULNIK AI 2007. T222P mutation of the insulin-like 3 hormone receptor LGR8 is associated with testicular maldescent and hinders receptor expression on the cell surface membrane. Am J Physiol Endocrinol Metab, 292, E138–44. [DOI] [PubMed] [Google Scholar]
- BOGATCHEVA NV, TRUONG A, FENG S, ENGEL W, ADHAM IM & AGOULNIK AI 2003. GREAT/LGR8 is the only receptor for insulin-like 3 peptide. Mol Endocrinol, 17, 2639–46. [DOI] [PubMed] [Google Scholar]
- BRUELL S, SETHI A, SMITH N, SCOTT DJ, HOSSAIN MA, WU QP, GUO ZY, PETRIE EJ, GOOLEY PR & BATHGATE RAD 2017. Distinct activation modes of the Relaxin Family Peptide Receptor 2 in response to insulin-like peptide 3 and relaxin. Sci Rep, 7, 3294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- BULLESBACH EE & SCHWABE C 1999. Tryptophan B27 in the relaxin-like factor (RLF) is crucial for RLF receptor-binding. Biochemistry, 38, 3073–8. [DOI] [PubMed] [Google Scholar]
- CALLANDER GE, THOMAS WG & BATHGATE RA 2009. Prolonged RXFP1 and RXFP2 signaling can be explained by poor internalization and a lack of beta-arrestin recruitment. Am J Physiol Cell Physiol, 296, C1058–66. [DOI] [PubMed] [Google Scholar]
- CAPRA AP, FERRO E, LA ROSA MA, BRIUGLIA S, RUSSO T, ARENA S, SALPIETRO DAMIANO C, ROMEO C & IMPELLIZZERI P 2018. Genetic analysis of the human insulin-like 3 gene in pediatric patients with testicular torsion. Pediatr Surg Int, 34, 807–812. [DOI] [PubMed] [Google Scholar]
- DAI Y, IVELL R & ANAND-IVELL R 2017a. Theca Cell INSL3 and Steroids Together Orchestrate the Growing Bovine Antral Follicle. Front Physiol, 8, 1033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- DAI Y, IVELL R, LIU X, JANOWSKI D & ANAND-IVELL R 2017b. Relaxin-Family Peptide Receptors 1 and 2 Are Fully Functional in the Bovine. Front Physiol, 8, 359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- DE TONI L, AGOULNIK AI, SANDRI M, FORESTA C & FERLIN A 2019. INSL3 in the muscolo-skeletal system. Mol Cell Endocrinol, 487, 12–17. [DOI] [PubMed] [Google Scholar]
- DEL BORGO MP, HUGHES RA, BATHGATE RA, LIN F, KAWAMURA K & WADE JD 2006. Analogs of insulin-like peptide 3 (INSL3) B-chain are LGR8 antagonists in vitro and in vivo. J Biol Chem, 281, 13068–74. [DOI] [PubMed] [Google Scholar]
- DI NISIO A, DE TONI L, ROCCA MS, GHEZZI M, SELICE R, TAGLIALAVORO G, FERLIN A & FORESTA C 2018. Negative Association Between Sclerostin and INSL3 in Isolated Human Osteocytes and in Klinefelter Syndrome: New Hints for Testis-Bone Crosstalk. J Clin Endocrinol Metab, 103, 2033–2041. [DOI] [PubMed] [Google Scholar]
- EL HOUATE B, ROUBA H, SIBAI H, BARAKAT A, CHAFIK A, CHADLI EL B, IMKEN L, BOGATCHEVA NV, FENG S, AGOULNIK AI & MCELREAVEY K 2007. Novel mutations involving the INSL3 gene associated with cryptorchidism. J Urol, 177, 1947–51. [DOI] [PubMed] [Google Scholar]
- FENG S, BOGATCHEVA NV, TRUONG A, KORCHIN B, BISHOP CE, KLONISCH T, AGOULNIK IU & AGOULNIK AI 2007. Developmental expression and gene regulation of insulin-like 3 receptor RXFP2 in mouse male reproductive organs. Biol Reprod, 77, 671–80. [DOI] [PubMed] [Google Scholar]
- FERGUSON L & AGOULNIK AI 2013. Testicular cancer and cryptorchidism. Front Endocrinol (Lausanne), 4, 32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- FERLIN A, DE TONI L, AGOULNIK AI, LUNARDON G, ARMANI A, BORTOLANZA S, BLAAUW B, SANDRI M & FORESTA C 2018. Protective Role of Testicular Hormone INSL3 From Atrophy and Weakness in Skeletal Muscle. Front Endocrinol (Lausanne), 9, 562. [DOI] [PMC free article] [PubMed] [Google Scholar]
- FERLIN A & FORESTA C 2005. Insulin-like factor 3: a novel circulating hormone of testicular origin in humans. Ann N Y Acad Sci, 1041, 497–505. [DOI] [PubMed] [Google Scholar]
- FERLIN A, GAROLLA A, RIGON F, RASI CALDOGNO L, LENZI A & FORESTA C 2006. Changes in serum insulin-like factor 3 during normal male puberty. J Clin Endocrinol Metab, 91, 3426–31. [DOI] [PubMed] [Google Scholar]
- FERLIN A, PEPE A, GIANESELLO L, GAROLLA A, FENG S, GIANNINI S, ZACCOLO M, FACCIOLLI A, MORELLO R, AGOULNIK AI, et al. 2008. Mutations in the insulin-like factor 3 receptor are associated with osteoporosis. J Bone Miner Res, 23, 683–93. [DOI] [PMC free article] [PubMed] [Google Scholar]
- FERLIN A, PERILLI L, GIANESELLO L, TAGLIALAVORO G & FORESTA C 2011. Profiling insulin like factor 3 (INSL3) signaling in human osteoblasts. PLoS One, 6, e29733. [DOI] [PMC free article] [PubMed] [Google Scholar]
- FU P, SHEN PJ, ZHAO CX, SCOTT DJ, SAMUEL CS, WADE JD, TREGEAR GW, BATHGATE RA & GUNDLACH AL 2006. Leucine-rich repeat-containing G-protein-coupled receptor 8 in mature glomeruli of developing and adult rat kidney and inhibition by insulin-like peptide-3 of glomerular cell proliferation. J Endocrinol, 189, 397–408. [DOI] [PubMed] [Google Scholar]
- GLISTER C, SATCHELL L, BATHGATE RA, WADE JD, DAI Y, IVELL R, ANAND-IVELL R, RODGERS RJ & KNIGHT PG 2013. Functional link between bone morphogenetic proteins and insulin-like peptide 3 signaling in modulating ovarian androgen production. Proc Natl Acad Sci U S A, 110, E1426–35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- GORLOV IP, KAMAT A, BOGATCHEVA NV, JONES E, LAMB DJ, TRUONG A, BISHOP CE, MCELREAVEY K & AGOULNIK AI 2002. Mutations of the GREAT gene cause cryptorchidism. Hum Mol Genet, 11, 2309–18. [DOI] [PubMed] [Google Scholar]
- HAGEN CP, MIERITZ MG, NIELSEN JE, ANAND-IVELL R, IVELL R & JUUL A 2015. Longitudinal assessment of circulating insulin-like peptide 3 levels in healthy peripubertal girls. Fertil Steril, 103, 780–6.e1. [DOI] [PubMed] [Google Scholar]
- HALLS ML, BATHGATE RA & SUMMERS RJ 2006. Relaxin family peptide receptors RXFP1 and RXFP2 modulate cAMP signaling by distinct mechanisms. Mol Pharmacol, 70, 214–26. [DOI] [PubMed] [Google Scholar]
- HALLS ML, BATHGATE RA & SUMMERS RJ 2007a. Comparison of signaling pathways activated by the relaxin family peptide receptors, RXFP1 and RXFP2, using reporter genes. J Pharmacol Exp Ther, 320, 281–90. [DOI] [PubMed] [Google Scholar]
- HALLS ML, BOND CP, SUDO S, KUMAGAI J, FERRARO T, LAYFIELD S, BATHGATE RA & SUMMERS RJ 2005. Multiple binding sites revealed by interaction of relaxin family peptides with native and chimeric relaxin family peptide receptors 1 and 2 (LGR7 and LGR8). J Pharmacol Exp Ther, 313, 677–87. [DOI] [PubMed] [Google Scholar]
- HALLS ML, VAN DER WESTHUIZEN ET, BATHGATE RA & SUMMERS RJ 2007b. Relaxin family peptide receptors--former orphans reunite with their parent ligands to activate multiple signalling pathways. Br J Pharmacol, 150, 677–91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- HALLS ML, VAN DER WESTHUIZEN ET, WADE JD, EVANS BA, BATHGATE RA & SUMMERS RJ 2009. Relaxin family peptide receptor (RXFP1) coupling to G(alpha)i3 involves the C-terminal Arg752 and localization within membrane Raft Microdomains. Mol Pharmacol, 75, 415–28. [DOI] [PubMed] [Google Scholar]
- HAMPEL U, KLONISCH T, SEL S, SCHULZE U, GARREIS F, SEITMANN H, ZOUBOULIS CC & PAULSEN FP 2013. Insulin-like factor 3 promotes wound healing at the ocular surface. Endocrinology, 154, 2034–45. [DOI] [PubMed] [Google Scholar]
- HOMBACH-KLONISCH S, BIALEK J, RADESTOCK Y, TRUONG A, AGOULNIK AI, FIEBIG B, WILLING C, WEBER E, HOANG-VU C & KLONISCH T 2010. INSL3 has tumor-promoting activity in thyroid cancer. Int J Cancer, 127, 521–31. [DOI] [PubMed] [Google Scholar]
- HUANG Z, RIVAS B & AGOULNIK AI 2012. Insulin-like 3 signaling is important for testicular descent but dispensable for spermatogenesis and germ cell survival in adult mice. Biol Reprod, 87, 143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- IVELL R, BALVERS M, DOMAGALSKI R, UNGEFROREN H, HUNT N & SCHULZE W 1997. Relaxin-like factor: a highly specific and constitutive new marker for Leydig cells in the human testis. Mol Hum Reprod, 3, 459–66. [DOI] [PubMed] [Google Scholar]
- IVELL R, WADE JD & ANAND-IVELL R 2013. INSL3 as a biomarker of Leydig cell functionality. Biol Reprod, 88, 147. [DOI] [PubMed] [Google Scholar]
- JOHNSON KJ, ROBBINS AK, WANG Y, MCCAHAN SM, CHACKO JK & BARTHOLD JS 2010. Insulin-like 3 exposure of the fetal rat gubernaculum modulates expression of genes involved in neural pathways. Biol Reprod, 83, 774–82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- KAFTANOVSKAYA EM, FENG S, HUANG Z, TAN Y, BARBARA AM, KAUR S, TRUONG A, GORLOV IP & AGOULNIK AI 2011. Suppression of insulin-like3 receptor reveals the role of beta-catenin and Notch signaling in gubernaculum development. Mol Endocrinol, 25, 170–83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- KAFTANOVSKAYA EM, NG HH, SOULA M, RIVAS B, MYHR C, HO BA, CERVANTES BA, SHUPE TD, DEVARASETTY M, HU X, et al. 2019. Therapeutic effects of a small molecule agonist of the relaxin receptor ML290 in liver fibrosis. FASEB J, fj201901046R. [DOI] [PMC free article] [PubMed] [Google Scholar]
- KAWAMURA K, KUMAGAI J, SUDO S, CHUN SY, PISARSKA M, MORITA H, TOPPARI J, FU P, WADE JD, BATHGATE RA, et al. 2004. Paracrine regulation of mammalian oocyte maturation and male germ cell survival. Proc Natl Acad Sci U S A, 101, 7323–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- KLONISCH T, MULLER-HUESMANN H, RIEDEL M, KEHLEN A, BIALEK J, RADESTOCK Y, HOLZHAUSEN HJ, STEGER K, LUDWIG M, WEIDNER W, et al. 2005. INSL3 in the benign hyperplastic and neoplastic human prostate gland. Int J Oncol, 27, 307–15. [PubMed] [Google Scholar]
- KUMAGAI J, HSU SY, MATSUMI H, ROH JS, FU P, WADE JD, BATHGATE RA & HSUEH AJ 2002. INSL3/Leydig insulin-like peptide activates the LGR8 receptor important in testis descent. J Biol Chem, 277, 31283–6. [DOI] [PubMed] [Google Scholar]
- KURZ D 2016. Current Management of Undescended Testes. Curr Treat Options Pediatr, 2, 43–51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- LI Z, FENG S, LOPEZ V, ELHAMMADY G, ANDERSON ML, KAFTANOVSKAYA EM & AGOULNIK AI 2011. Uterine cysts in female mice deficient for caveolin-1 and insulin-like 3 receptor RXFP2. Endocrinology, 152, 2474–82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- LIM HN, RAIPERT-DE MEYTS E, SKAKKEBAEK NE, HAWKINS JR & HUGHES IA 2001. Genetic analysis of the INSL3 gene in patients with maldescent of the testis. Eur J Endocrinol, 144, 129–37. [DOI] [PubMed] [Google Scholar]
- LIN F, OTVOS L JR., KUMAGAI J, TREGEAR GW, BATHGATE RA & WADE JD 2004. Synthetic human insulin 4 does not activate the G-protein-coupled receptors LGR7 or LGR8. J Pept Sci, 10, 257–64. [DOI] [PubMed] [Google Scholar]
- MINAGAWA I, MURATA Y, TERADA K, SHIBATA M, PARK EY, SASADA H & KOHSAKA T 2018. Evidence for the role of INSL3 on sperm production in boars by passive immunisation. Andrologia, 50, e13010. [DOI] [PubMed] [Google Scholar]
- MUDA M, HE C, MARTINI PG, FERRARO T, LAYFIELD S, TAYLOR D, CHEVRIER C, SCHWEICKHARDT R, KELTON C, RYAN PL, et al. 2005. Splice variants of the relaxin and INSL3 receptors reveal unanticipated molecular complexity. Mol Hum Reprod, 11, 591–600. [DOI] [PubMed] [Google Scholar]
- MUPPIDI A, LEE SJ, HSU CH, ZOU H, LEE C, PFLIMLIN E, MAHANKALI M, YANG P, CHAO E, AHMAD I, et al. 2019. Design and Synthesis of Potent, Long-Acting Lipidated Relaxin-2 Analogs. Bioconjug Chem, 30, 83–89. [DOI] [PubMed] [Google Scholar]
- NAGORNIEWICZ B, MARDHIAN DF, BOOIJINK R, STORM G, PRAKASH J & BANSAL R 2019. Engineered Relaxin as theranostic nanomedicine to diagnose and ameliorate liver cirrhosis. Nanomedicine, 17, 106–118. [DOI] [PubMed] [Google Scholar]
- NEF S & PARADA LF 1999. Cryptorchidism in mice mutant for Insl3. Nat Genet, 22, 295–9. [DOI] [PubMed] [Google Scholar]
- OVERBEEK PA, GORLOV IP, SUTHERLAND RW, HOUSTON JB, HARRISON WR, BOETTGER-TONG HL, BISHOP CE & AGOULNIK AI 2001. A transgenic insertion causing cryptorchidism in mice. Genesis, 30, 26–35. [DOI] [PubMed] [Google Scholar]
- PATHIRANA IN, KAWATE N, BULLESBACH EE, TAKAHASHI M, HATOYA S, INABA T & TAMADA H 2012. Insulin-like peptide 3 stimulates testosterone secretion in mouse Leydig cells via cAMP pathway. Regul Pept, 178, 102–6. [DOI] [PubMed] [Google Scholar]
- PELUSI C, FANELLI F, PARIALI M, ZANOTTI L, GAMBINERI A & PASQUALI R 2013. Parallel variations of insulin-like peptide 3 (INSL3) and antimullerian hormone (AMH) in women with the polycystic ovary syndrome according to menstrual cycle pattern. J Clin Endocrinol Metab, 98, E1575–82. [DOI] [PubMed] [Google Scholar]
- PITIA AM, UCHIYAMA K, SANO H, KINUKAWA M, MINATO Y, SASADA H & KOHSAKA T 2017. Functional insulin-like factor 3 (INSL3) hormone-receptor system in the testes and spermatozoa of domestic ruminants and its potential as a predictor of sire fertility. Anim Sci J, 88, 678–690. [DOI] [PubMed] [Google Scholar]
- ROSENGREN KJ, ZHANG S, LIN F, DALY NL, SCOTT DJ, HUGHES RA, BATHGATE RA, CRAIK DJ & WADE JD 2006. Solution structure and characterization of the LGR8 receptor binding surface of insulin-like peptide 3. J Biol Chem, 281, 28287–95. [DOI] [PubMed] [Google Scholar]
- SADEGHIAN H, ANAND-IVELL R, BALVERS M, RELAN V & IVELL R 2005. Constitutive regulation of the Insl3 gene in rat Leydig cells. Mol Cell Endocrinol, 241, 10–20. [DOI] [PubMed] [Google Scholar]
- SCOTT DJ, LAYFIELD S, YAN Y, SUDO S, HSUEH AJ, TREGEAR GW & BATHGATE RA 2006. Characterization of novel splice variants of LGR7 and LGR8 reveals that receptor signaling is mediated by their unique low density lipoprotein class A modules. J Biol Chem, 281, 34942–54. [DOI] [PubMed] [Google Scholar]
- SCOTT DJ, WILKINSON TN, ZHANG S, FERRARO T, WADE JD, TREGEAR GW & BATHGATE RA 2007. Defining the LGR8 residues involved in binding insulin-like peptide 3. Mol Endocrinol, 21, 1699–712. [DOI] [PubMed] [Google Scholar]
- SEDAGHAT K, SHEN PJ, FINKELSTEIN DI, HENDERSON JM & GUNDLACH AL 2008. Leucine-rich repeat-containing G-protein-coupled receptor 8 in the rat brain: Enrichment in thalamic neurons and their efferent projections. Neuroscience, 156, 319–33. [DOI] [PubMed] [Google Scholar]
- SEYAM E & HEFZY E 2018. Evaluation of the correlation between insulin like factor 3, polycystic ovary syndrome, and ovarian maldescent. Gynecol Endocrinol, 34, 481–488. [DOI] [PubMed] [Google Scholar]
- SHABANPOOR F, BATHGATE RA, HOSSAIN MA, GIANNAKIS E, WADE JD & HUGHES RA 2007. Design, synthesis and pharmacological evaluation of cyclic mimetics of the insulin-like peptide 3 (INSL3) B-chain. J Pept Sci, 13, 113–20. [DOI] [PubMed] [Google Scholar]
- SHABANPOOR F, HUGHES RA, ZHANG S, BATHGATE RA, LAYFIELD S, HOSSAIN MA, TREGEAR GW, SEPAROVIC F & WADE JD 2010. Effect of helix-promoting strategies on the biological activity of novel analogues of the B-chain of INSL3. Amino Acids, 38, 121–31. [DOI] [PubMed] [Google Scholar]
- SHAIKH N, DADACHANJI R, MEHERJI P, SHAH N & MUKHERJEE S 2016. Polymorphisms and haplotypes of insulin-like factor 3 gene are associated with risk of polycystic ovary syndrome in Indian women. Gene, 577, 180–6. [DOI] [PubMed] [Google Scholar]
- SHARMA V, LEHMANN T, STUCKAS H, FUNKE L & HILLER M 2018. Loss of RXFP2 and INSL3 genes in Afrotheria shows that testicular descent is the ancestral condition in placental mammals. PLoS Biol, 16, e2005293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- SUDO S, KUMAGAI J, NISHI S, LAYFIELD S, FERRARO T, BATHGATE RA & HSUEH AJ 2003. H3 relaxin is a specific ligand for LGR7 and activates the receptor by interacting with both the ectodomain and the exoloop 2. J Biol Chem, 278, 7855–62. [DOI] [PubMed] [Google Scholar]
- SUN J, HAO W, FILLMORE N, MA H, SPRINGER D, YU ZX, SADOWSKA A, GARCIA A, CHEN R, MUNIZ-MEDINA V, et al. 2019. Human Relaxin-2 Fusion Protein Treatment Prevents and Reverses Isoproterenol-Induced Hypertrophy and Fibrosis in Mouse Heart. J Am Heart Assoc, 8, e013465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- WENZLER DL, BLOOM DA & PARK JM 2004. What is the rate of spontaneous testicular descent in infants with cryptorchidism? J Urol, 171, 849–51. [DOI] [PubMed] [Google Scholar]
- YUAN FP, LI X, LIN J, SCHWABE C, BULLESBACH EE, RAO CV & LEI ZM 2010. The role of RXFP2 in mediating androgen-induced inguinoscrotal testis descent in LH receptor knockout mice. Reproduction, 139, 759–69. [DOI] [PMC free article] [PubMed] [Google Scholar]
- ZARREH-HOSHYARI-KHAH MR, EINSPANIER A & IVELL R 1999. Differential splicing and expression of the relaxin-like factor gene in reproductive tissues of the marmoset monkey (Callithrix jacchus). Biol Reprod, 60, 445–53. [DOI] [PubMed] [Google Scholar]
- ZIMMERMANN S, STEDING G, EMMEN JM, BRINKMANN AO, NAYERNIA K, HOLSTEIN AF, ENGEL W & ADHAM IM 1999. Targeted disruption of the Insl3 gene causes bilateral cryptorchidism. Mol Endocrinol, 13, 681–91. [DOI] [PubMed] [Google Scholar]