

Abstract
According to the World Health Organization (WHO), the global nutrition report shows that whilst part of the world’s population starves, the other part suffers from obesity and associated complications. A balanced diet counterparts these extreme conditions with the proper proportion, composition, quantity, and presence of macronutrients, micronutrients, and bioactive compounds. However, little is known on the way these components exert any influence on our health. These nutrients aiming to feed our bodies, our tissues, and our cells, first need to reach mitochondria, where they are decomposed into CO2 and H2O to obtain energy. Mitochondria are the powerhouse of the cell and mainly responsible for nutrients metabolism, but they are also the main source of oxidative stress and cell death by apoptosis. Unappropriated nutrients may support mitochondrial to become the Trojan horse in the cell. This review aims to provide an approach to the role that some nutrients exert on mitochondria as a major contributor to high prevalent Western conditions including metabolic syndrome (MetS), a constellation of pathologic conditions which promotes type II diabetes and cardiovascular risk. Clinical and experimental data extracted from in vitro animal and cell models further demonstrated in patients, support the idea that a balanced diet, in a healthy lifestyle context, promotes proper bioenergetic and mitochondrial function, becoming the best medicine to prevent the onset and progression of MetS. Any advance in the prevention and management of these prevalent complications help to face these challenging global health problems, by ameliorating the quality of life of patients and reducing the associated sociosanitary burden.
Keywords: mitochondria, metabolic syndrome, mitochondrial dysfunction, balanced diet, nutrients, lifestyle
1. Introduction
In the last decades, global dietary patterns have experienced a transition towards increasingly Westernized and less healthier diets. This trend is linked to the substantial increase in the prevalence of non-communicable diseases (NCDs) such as type II diabetes (T2D) or cardiovascular disease (CVD) [1]. During the last 40 years, the global prevalence of T2D has almost doubled, from 4.7% in 1980 to 8.5% in 2014. Likewise, the number of deaths attributed to CVD (mainly coronary heart disease, stroke, and rheumatic heart disease) has increased from 14.4 million in 1990 to 17.5 million, being the number 1 cause of death globally [2].
The metabolic syndrome (MetS), a cluster of pathologic conditions that raise the risk of CVD and T2D [3], has been emerged as a health problem in modern society, associated with enormous social, personal and economic burden in both the developing and developed world [4]. Currently, MetS is calculated to affect 25% of the global population [5] and all the epidemiological studies have demonstrated its prevalence increases with age [6]. Additionally, the growing age of the population in first world societies will increase such trends in coming years.
International food organizations have traditionally focused on food security and micronutrient deficiency. However, diet-related health burdens contributing to the above-mentioned diseases are now surpassing those due to undernutrition in nearly every region of the world. Therefore, the establishment of healthy dietary patterns is a global priority to reduce the onset of these diseases [7].
There are plenty of studies establishing healthy dietary patterns [1]. However, scarce knowledge is available on the way these nutrients underscore or promote our health. First, because there is a wide panoply of nutrients and, second, because each one of them can trigger a multitude of effects, organ, tissue, cell, and even molecule specific. However, a universal fact is that all of them need to be processed into small subunits and metabolized by mitochondria to obtain energy. Thus, mitochondrial function is key to understand dietary effects in our organism and, at the same time, understanding how dietary patterns and nutrients influence mitochondrial function is essential to establish causal relationships between food and health.
The diet’s impact on mitochondrial function has been widely studied due to its role as a powerhouse of the cell and its involvement in nutrients’ metabolism. In fact, mitochondria are critical in the maintenance of metabolic flexibility, efficient switches in metabolism depending on the environmental demand (feeding/fasting cycles) [8]. Moreover, also the intake amount (p.e. high-fat and/or sugars diet) conditionate mitochondrial function [9]. Additionally, micronutrients (the ~40 essential vitamins, minerals, fatty acids, and amino acids) and bioactive compounds, must be considered in a balanced diet to get an optimal cell–mitochondrial–metabolic axis function [10].
These dietary considerations are closely related to the manifestation of the most common NCDs [11], including MetS [12], where the mitochondrial dys(function) can be considered a common hallmark.
Consequently, the aim of this work is to review the role of some nutrients, easily incorporable in a balanced diet, over mitochondrial (dys)function and its impact on MetS. For this purpose, we herein have reviewed data extracted from experimental and clinical studies published in PubMed and Google Scholar database, from inception to July 2020, to document the principal examples of nutrients’ effect on mitochondrial activity and their influence on MetS.
2. A Healthy Diet
Diet is the most significant single risk factor for disability and premature death [13]. Dietary choices contribute to the development of pathologic conditions such as hypertension, hypercholesterolemia, overweight/obesity, and inflammation, which in turn increase the risk for associated diseases, including CVD or T2D. Several studies have demonstrated the causal link between the global shift towards Westernized dietary patterns, characterized by high intakes of red and processed meat, pre-packaged foods, sweets, and refined grains, and lacking in fruits and vegetables, whole-grains, fish, and nuts, and the increased prevalence of these diet-related chronic diseases in the last decades [1,14].
Dietary changes recommended by the World Health Organization (WHO) include balancing energy intake, limiting saturated and trans fats, and shifting toward unsaturated fats consumption, increasing intake of fruits and vegetables, and limiting the consumption of sugar and salt. These dietary patterns are naturally occurring in certain world regions, as it is the case of the Mediterranean diet (MedDiet) and traditional Asian diets such as Korean, Chinese, and Japanese diets. Both the MedDiet (rich in fish, monounsaturated fats from olive oil, fruits, vegetables, whole grains, and legumes/nuts) and traditional Asian diets (composed of rice and other whole grains, fermented food, soy, fruits and vegetables, fish and seafood) [1] have moderate to strong evidence for preventing T2D and MetS, decreasing cancer and CVD incidence and mortality, decreasing overall mortality, and treating obesity [15,16,17,18]. Moreover, healthy dietary patterns have also been developed based on nutrient intake studies and their outcomes, as it is the case of DASH (Dietary Approaches to Stop Hypertension) [19] and MIND (Mediterranean-DASH Intervention for Neurodegenerative Delay) diets [20], among others.
At the molecular level, a healthy diet is characterized by the consumption of appropriate proportions of macronutrients (carbohydrates, proteins, and fats) to support energetic and physiologic functions, sufficient micronutrients (vitamins and mineral) and hydration, to meet the physiologic needs of the body [1]. Additionally, there is increasing evidence of the beneficial role of bioactive compounds, such as polyphenols or lycopene, in a healthy diet to prevent diseases. These molecules, generally natural and non-nutritive compounds, can be found in small quantities in plants and lipid-rich foods, and are essential to fight age-related and chronic diseases [21].
3. Metabolic Syndrome (MetS)
The MetS is not a disease per se, but a constellation of pathologic conditions that raises the risk of CVD by 1.5–2.5 fold [22] and T2D by 3–20 fold, depending on the numbers of pathologic factors [23]. To be diagnosed with MetS, at least three of the five deregulations shown in Table 1 must be present in the patient [24].
Table 1.
Risk factors associated with metabolic syndrome and definition of the parameters for its diagnosis.
| Risk Factor | Parameters | Definition (Male/Female) |
|---|---|---|
| Central obesity | Waist circumference 1 | America: ≥102 cm/≥88 cm Asia: ≥90 cm/≥80 cm Europe: ≥94 cm/≥80 cm Africa: ≥94 cm/≥80 cm Middle East: ≥94 cm/≥80 cm |
| Hypertriglyceridemia | Triglyceride concentrations | ≥150 mg/dL |
| Low HDL-cholesterol levels | HDL-cholesterol levels | ≤40 mg/dL/<50 mg/dL |
| Hypertension | Systolic/diastolic blood pressure | ≥130/85 mm Hg |
| Hyperglycemia | Fasting plasma glucose | ≥100 mg/dL |
1 The definition of this measure varies among regions. HDL = High density lipoprotein.
The determining factors for the development of MetS are, in order of relevance/influence: weight, genetics, aging, and lifestyle [25]. Based on that, risk factors associated with MetS can be reduced by the introduction of a balanced-diet and some regular exercise [26].
4. Mitochondria in Health and Disease
Mitochondria are unique structures in the cell, originally alpha-proteobacterium phagocyted by a eukaryotic progenitor [27]. Since then, proper mitochondrial activity is critical for the survival of the eukaryotic cell. In fact, mitochondrial deregulation has been described as a hallmark underlying several complex diseases, since mitochondrial dysfunction can trigger, by its own, oxidative stress, bioenergetic failure, inflammation, protein accumulation, and cell death [28], common aspects in metabolic disorders, such as MetS (Figure 1). That is why the nature of mitochondrial disorders underlies the numerous roles that mitochondria play.
Figure 1.

Mitochondrial (dys)function factors. Mitochondrial dysfunction is characterized by the combination of some of the processes described above and plays a central role in several diseases, such as metabolic syndrome. All these processes are closely related, usually affecting one each other and rarely having isolated effects. Oxidative stress leads to the accumulation of reactive oxygen species (ROS). In turn, high amount of ROS might trigger DNA mutations. Mitochondrial (dys)function can also be associated to reduced mitochondrial biogenesis through the decreased expression of mitochondrial genes. Moreover, altered proteostasis (biogenesis, folding, trafficking, and degradation of mitochondrial proteins) is common. Another frequent consequence is an energy imbalance limiting the ATP supply. Furthermore, mitochondrial (dys)function may condition mitochondrial fusion-fission processes compromising mitochondrial renewal and material exchange. Similarly, impaired mitophagy triggers the accumulation of non-functional mitochondria. Finally, mitochondrial altered Ca2+ flux and buffering, disrupt communication with other organelles such as endoplasmic reticulum and, in case of calcium release into the cytoplasm, the potential trigger of cell death.
4.1. Mitochondrial (Dys)Functions
The best-known mitochondrial function is energy production, through the generation of ATP molecules, via oxidative phosphorylation. The mitochondrial matrix houses the Krebs cycle (KC) enzymes, important producers of electron donors such as NADH and FADH2. These molecules bring electrons to the mitochondrial respiratory chain (MRC), formed by four complexes (I-IV), that pump protons from the mitochondrial matrix into the inner membrane space through sequential redox reactions that finally reduce O2 into H2O at complex IV and, the complex V or ATP synthase, uses the accumulated proto-gradient energy to phosphorylate ADP into ATP (chemiosmotic theory of Mitchell).
One of the major cell producers of NADH is the oxidation of fatty acids (FA) to acyl-CoA, a process called β-oxidation, which takes place in the mitochondrial matrix. The acetyl-CoA resulting from this process can enter the KC and get oxidized, coupled to the production of reducing power [29]. Furthermore, alterations in these metabolic routes have also been widely described in cell and animal models, as well as in patients suffering the most prevalent NCDs such as heart failure, T2D [30], or MetS [31,32].
Secondarily, these redox reactions generate reactive oxygen species (ROS) (Figure 2). Low ROS increase can trigger healthy physiological responses by activating complex transcriptional cascades of stress resistance, cell proliferation, and differentiation pathways [33]. However, when cells overproduce ROS, those can trigger pathologic consequences by promoting damage of cellular compounds (in sugars, lipids, proteins, or nucleic acids) [34,35], thus impairing cell proliferation, differentiation, and apoptosis, among others [36]. Additionally, mitochondria can trigger or amplify by their own cell death signals triggered by an alternative source of damage. Altogether, these effects may have a deep impact in mitochondrial deregulation and derived diseases, including MetS [37].
Figure 2.

Main sources of mitochondrial reactive oxygen species (ROS). Mitochondrial respiratory chain (MRC) is composed of four enzyme complexes (I–IV). Electrons donated from NADH and FADH2 in the Krebs cycle are transferred at complex I and II respectively, and consecutively to complex III and IV. This electrons transfer is coupled with protons transport across the inner membrane, generating a proton-gradient, allowing ADP phosphorylation through ATP synthase (complex V). Oxygen metabolism generates superoxide anion (·02−), which in turn is conversed into hydrogen peroxide (H2O2) by SOD (superoxide dismutase) and then converted into water thanks to the action of some of the antioxidant enzymatic defenses such as catalase (CAT) or the glutathione peroxidase (GPx) reductase (GPr) system. Excessive ROS production can oxidize proteins, lipids, or mitochondrial DNA (mtDNA), eventually leading to mitochondrial dysfunction and cell death.
To fight oxidative stress, understood as the imbalance between ROS production and antioxidants in favor of ROS species, cells have several molecules involved in the antioxidant defense system. The enzymatic barrier includes superoxide dismutase (SOD), catalase (CAT), glutathione reductase (GR), and glutathione peroxidase (GPx). Whilst other barriers, non-enzymatic antioxidants molecules, are vitamins E and C, glutathione (GSH), and various carotenoids and flavonoids [4], present in foods and beverages.
The proton leak generated in the MRC is mainly used to produce ATP but can also be used for thermogenesis purposes or to reduce the mitochondrial membrane potential, aiming to protect cells from oxidative stress [38]. The uncoupling proteins (UCPs) are a family of 5 proteins responsible for uncoupling and widely distributed in specific tissues. In fact, several studies in rodents and patients demonstrated abnormal expression or activity of UCPs related to obesity [39] and T2D [40].
Interestingly, mitochondrial functions reach far beyond bioenergetics. Mitochondria are the key organelles in iron metabolism, involved in heme and Fe-S clusters synthesis [41], essential structures in oxygen transport, MRC, and DNA repair machinery.
Moreover, mitochondria regulate Ca2+ homeostasis, a key factor regulating aerobic metabolism and one of the apoptosis triggering factors [42,43]. Alterations in these pathways are linked to NCDs [44,45].
For all the above mentioned, mitochondria are key organelles for cell survival and adaptation to changing environmental conditions. Thus, for instance, high energy requirements activate mitochondrial biogenesis, induce the generation of new MRC units, and trigger an integrated physiological response of metabolic and functional remodeling. Several molecules are involved in the detection of specific signals of mitochondrial activity, such as NAD+:NADH, AMP:ATP ratios, or acetyl-CoA levels. That is why alterations in this fragile equilibrium conduct to a dysregulated mitochondrial activity, leading to the altered pathophysiological response observed in, among others, MetS.
Similarly, mitochondrial activities depend on the abundance of these organelles, their morphology, and their functional properties, regulated, among others, by mitochondrial dynamics [46,47,48]. Mitochondria are permanently fusing and fissioning with each other, allowing the renewal of genetic, structural, and functional components. It is especially the mitochondrial fusion process, which has been reported to allow the exchange of mitochondrial content, including mitochondrial DNA (mtDNA), which may buffer partial defects and transient stresses [49,50]. The loss of fusion and fission abilities results in altered mitochondrial populations, which leads to the most common NCDs [51].
To summarize, mitochondrial dysfunction can be due to several levels of deficiency and lead to different pathological features. All their levels of function are closely related, affecting one each other, rarely having isolated effects. In the case of MetS, reduced biogenesis conducing to variations in mitochondrial number, altered membrane potential, leading to defective energy production and defects in the activity of oxidative proteins, has been reported, all together linked with the accumulation of ROS in cells and tissues [52].
4.2. Some Interactions with Other Cell Structures
Mitochondria are not isolated organelles inside the cell, so interactions with other structures must be considered in health and disease. For instance, interactions with lysosomes [53] allow the degradation and turnover of defective or unnecessary mitochondrial by mitophagy, important to maintain mitochondrial quality control (MQC). The failure of the MQC system has been described in NCDs patients [54,55,56], demonstrating the importance of mitochondrial interactions with other eukaryotic cell structures.
Another important interaction of mitochondria to support cell functionality is with the endoplasmic reticulum (ER) [57], also known as mitochondria-associated membranes, MAMs, which allows another level of organelle and cell regulation. Alterations in MAMs have been demonstrated in NCDs patients [58,59,60] due, in part, to increased ER stress but also to mitochondrial dysfunction.
4.3. Some Mitoregulators
4.3.1. Epigenetics and Sirtuins
Epigenetic modifications in DNA, which include acetylation, methylation, and ubiquitination, among others [61], determine the transcriptional machinery accessibility to the genome [62]. However, these regulatory modifications are not limited to DNA and located in the nucleus, they can also target RNA, proteins, and mitochondria, and be associated with disease [63,64].
One well-known epigenome effector, which work as cellular sensors of metabolic status, are the NAD+ dependent histone deacetylases sirtuins [65]. Sirtuins are a family of seven proteins involved in different biological functions [66] such as DNA repair [67], apoptosis control [68,69], ROS and oxidative stress regulation [70], together with their modulatory activity over enzymes from catabolism to anabolism [71]. Some of them have mitochondrial location or activity [66,70].
The most studied sirtuins are Sirt1 and Sirt3. Genetic or pharmacological Sirt1 inhibition can induce insulin resistance (IR) [72], and genetic Sirt1 variations have been associated with human energy expenditure and obesity [73,74,75]. Moreover, the fasting state increases NAD+ levels and activate metabolism through Sirt3 [76]. In the absence of Sirt3, cell and mitochondrial proteins become hyperacetylated, contributing to the progression of metabolic diseases such as MetS [77,78].
4.3.2. miRNAs, mitomiRs, mitoRNAs, and xenomiRs
Non-coding ribonucleic acids (ncRNAs) are a wide heterogenous group of regulatory molecules described in the last decades. Inside this group, there are micro-ribonucleic acids (miRNAS), short molecules that prevent mRNA translation or induce transcripts degradation, thus preventing the generation of the protein and associated function. These molecules are mostly located in the cytoplasm, but there is a subset of about 150 different miRNAs, named mitomiRs, that have been also found in the mitochondrial fraction [79]. Furthermore, the transcriptome of mitochondrial genome has also revealed that it encodes multiple non-coding RNAs, which are named mitoRNAs [80]. Furthermore, in the past few years, these molecules have been associated with several NCDs [81,82,83], including MetS [84]. Interestingly, dietary changes in MetS patients associated with clinical improvements have been linked to changes in miRNAs profile [85], as later depicted.
Nutrients can interplay at different levels with the human body. Their components can be the anabolic precursors for the synthesis of complex molecules needed for cell survival. They can also interplay at the chemical level with cell constituents; this is the case of the antioxidant effect. The last findings support the idea that they can also regulate the expression of our genes. This is the case with the xenomiRs, or food-derived miRNAs, exogenous miRs that emulate the function of our endogenous miRs, exerting an additional mechanism of regulation for the expression of our genes. Despite there being only few data available and plenty of polemics, some studies revealed an important concentration of rice [86] or milk [87] xenomiRs in the plasma, after a dietary intervention. On the contrary, other publications were unable to detect them [88,89]. Additionally, in case these xenomiRs were absorbed, there is the crucial need to understand if they exert any function in our cells, as endogenous miRs do. What is certain is that this field of study will be of growing interest in the next coming years.
4.4. Contribution of Mitochondrial (Dys)Function to MetS
The association between MetS and mitochondrial (dys)function has been suspected since long ago and its contribution to the deterioration of the metabolic risk factors involved in the disease has been studied.
Westernized diets, that usually include high caloric intake (high fat and/or sugars) might promote an increase in the MRC activity that could lead to the overproduction of mitochondrial ROS [90]. In turn, the oxidative stress leads to reduced ATP production and decrease of the overall metabolism in the cell, which is tightly associated with MetS [91,92,93]. Moreover, oxidative stress might contribute to the appearance of insulin resistance (IR), defined as a decreased cellular response to physiological insulin levels [94]. However, it remains unclear whether mitochondrial (dys)function is the main cause of IR or vice-versa [95,96].
Furthermore, oxidative stress directly interferes in lipid oxidation, which results in an increased cellular lipid accumulation (mainly diacylglycerols and ceramides), a situation that inhibits insulin signaling [94,97,98]. This situation affects the translocation of the glucose transporter type 4 (Glut-44) in insulin-sensitive tissues such muscle or fat [99], which leads to circulating glucose accumulation and consequently, to hyperglycemia [100].
Moreover, lipids and free fatty acids (FFA) also accumulate in the circulatory system, leading to hypertriglyceridemia [101], which in turn promotes the increase of visceral adiposity and a modification of fat distribution, and consequently the increase of waist circumference. This situation is caused by the inability of adipose tissue to store the energy excess, so the FFA overload is accumulated in the muscle, heart, liver, or as subcutaneous fat [102]. Under these conditions, fat redistribution is accompanied by macrophages infiltration, which secrete proinflammatory molecules such as interleukin-6 (IL-6), tumor necrosis factor (TNF), or C reactive protein (CRP), generating a low-grade chronic inflammation state in MetS patients [103,104]. Furthermore, abdominal obesity correlates with systemic levels of oxidative stress biomarkers [105,106] and defective mitochondrial biogenesis, manifested by impaired mitochondrial dysfunction, which includes alterations in oxidative metabolism, low mitochondrial gene expression, and reduced ATP generation [107,108]. Similarly, mitochondrial alterations, increased oxidative stress, and low-grade chronic inflammation contribute to the etiology of hypertension [109,110]. Finally, oxidative stress in also one of the main triggers of the formation and progression of the atheroma plaque responsible of cardiovascular events included in the MetS.
5. Nutrients (Well-Being Through Feeding)
One of the best options to avoid, delay, or minimize risk factors contributing to MetS is changing lifestyle routines, including a balanced diet.
5.1. Monounsaturated Fatty Acids (MUFAs)
MUFAs are fats with one unsaturation in their carbon chain. Dietary MUFAs can have different origin, thus in Western diets, the supply is through foods of animal origin, whilst the main MUFAs source in south European countries is extra virgin olive oil containing oleic acid [111]. Other sources of MUFAs are the rest of vegetables oils and nuts such as macadamia nuts, hazelnuts, or pecans [112].
5.1.1. Some Extra Virgin Olive Oil Everywhere. Oleic Acid
Some cellular model studies have reported that oleic acid can have a positive role in mitochondrial function. In fact, one study from in vitro modeling of β-pancreatic cells showed that oleic acid enhances antioxidative defense [113]. Moreover, alternative studies demonstrated that oleic acid supplementation reduces ROS generation and protects mitochondria from oxidative stress or apoptosis [114,115]. Furthermore, oleic acid has been also reported as an anti-inflammatory modulator through AMP-activated protein kinase (AMPK) activation and its activity among different targets like nuclear factor kB (NF-kB) inhibition, and consequently, decreasing cytokines secretion such as TNF [116,117,118].
In turn, oleic acid also enhances fatty acid β-oxidation through AMPK response and interaction with Sirt1 and the peroxisome proliferator-activated receptor gamma coactivator 1-alpha PGC-1α [119,120], the master gene regulator of mitochondrial remodeling and biogenesis. Finally, oleic acid has been demonstrated to increase the carnitine palmitoyl transferase 1 (CPT-1) level promoting the fatty acid transport into mitochondria for β-oxidation [121,122]. Of note, most of these mechanisms have been described using in vitro models responding to higher concentrations of oleic acid compared to those obtained by food intake. Interestingly, several clinical benefits have been described with lower doses (see Section 5.1.2). Further and deeper investigation is needed to validate the mechanisms and dose-response effects of oleic acid, often observed in proof-of-concept studies.
5.1.2. Extra Virgin Olive Oil Against MetS
Systematic review and meta-analysis of cohort studies have demonstrated the positive effects of oleic acid decreasing mortality risk (11%) and cardiovascular risk (12%) [123,124]. Focusing attention on MetS, dietary MUFAs showed similar results [112]. In fact, clinical trials have demonstrated a reduction of MetS risk factors such as blood pressure or total low-density lipoprotein (LDL) cholesterol [125]. Moreover, also an improvement on insulin sensitivity and inflammatory response have been demonstrated using oleic acid in cell models [126] and randomized clinical trials [127]. Finally, recent systematic review on human intervention studies with enriched oleic acid diets demonstrated that this intervention can reduce central obesity and abdominal fat [128].
5.2. Polyunsaturated Fatty Acids (PUFAs)
PUFAs are fats with more than two unsaturations in their carbon chain that can be divided into two main groups. Omega-3 fatty acids (α-linolenic acid, eicosapentaenoic acid and docosahexaenoic acid) mainly found in blue fish, eggs, and walnuts. In turn, the omega-6 group (linoleic acid, arachidonic acid, and docosapentaenoic acid) are mostly founds in vegetable oils, such as sunflower, soybean, corn, and canola oils.
5.2.1. More Blue Fish and Less Vegetal Oils. Omega-3 vs. Omega 6
Omega-3 PUFAs have been reported to act as mitochondrial protectors, as they reduce ROS production and inflammation through the activation of AMPK. In turn, AMPK activates PGC-1α which, together with the decrease in fission genes such as dynamin related protein 1 (DRP1), stimulates mitochondrial function and fusion processes [129]. PGC-1α triggers different signaling pathways that promote mitochondrial biogenesis, β-oxidation, glucose utilization, antioxidants detoxification, and the activation of uncoupling proteins, which leads to a decrease in lipid accumulation and a reduction of ROS [130,131,132,133]. It has also been described to attenuate ER-stress and the subsequent disruption of Ca2+ homoeostasis. All these actions decrease the expression of pro-inflammatory cytokines and attenuate the inflammasome, which improves insulin sensitivity [134]. Some animal studies and clinical trials demonstrate that these actions would be triggered by changes in miRNAs profiles, among others [135,136]. In addition, the reduction of the omega-6/omega-3 balance and the omega-3 incorporation into mitochondrial membranes increase its viscosity. This situation improves mitochondrial function and avoids cell dysfunction and cell death [137]. However, the disbalanced omega-3/omega-6 ratio in detriment of omega-6 may also lead to adverse consequences, since excessive omega-3 PUFAs may eventually trigger pro-oxidant and pro-inflammatory environments [138]. Thus, the conservation of the omega-3/omega-6 ratio balance is key to avoid detrimental effects (Figure 3).
Figure 3.

Principal nutrients with beneficial effects on mitochondrial function. Oleic acid, omega-3, selenium and zinc, resveratrol, lycopene, and the organosulfur compounds (OSCs) activate numerous pathways, among them, direct or indirectly, PGC-1α signaling, one of the main coactivators and master gene regulator of energy metabolism and mitochondrial biogenesis. The activation of the PGC-1α promotes mitogenesis, β-oxidation, glucose utilization, uncoupling, and antioxidant detoxification. Moreover, vitamin A and vitamins from the B group are essential for a correct functioning of the glycolytic energy system, and vitamin C and oleic acid for the functioning of β-oxidation, among others. One of the main effects of oleic acid, lycopene, organosulfur compounds (OSCs), vitamins B, C, D, selenium, and zinc, catechins and olive oil polyphenols (OOPs) in our cells is the decrease of oxidative stress by both promoting an antioxidant response and inhibiting pro-oxidant enzymes. Vitamin E also acts as an antioxidant and maintains a correct mitochondrial structure, while omega-3 inhibits mitochondrial fission. Omega-3 and vitamin D are involved in calcium homeostasis, among others. Vitamin C, lycopene, and oleic acid have been reported to inhibit apoptosis, while oleic acid, omega-3, and OSCs reduce inflammation.
5.2.2. Omega-3 PUFAs Against MetS
It is still controversial whether omega-6 PUFAs exert pro-inflammatory or anti-inflammatory effects by themselves [139]. However, it is known that an unbalanced omega-6/omega-3 ratio in favor of omega-6 PUFAs is highly prothrombotic and proinflammatory. The high amounts of omega-6 PUFAs present in Western diets, together with the very low amounts of omega-3 PUFAs, leads to an unhealthy omega-6/omega-3 ratio that can reach a 20:1 proportion (instead the expected 1:1) [140], contributing to atherosclerosis, obesity, and T2D [141] (Figure 4).
Figure 4.

Main nutrient impact on metabolic syndrome (MetS) components. The MetS is a constellation of pathologic conditions which includes hyperglycemia, insulin resistance, dyslipidemia (hypertriglyceridemia and low high density lipoprotein (HDL)-cholesterol levels), central obesity, and hypertension. Several studies have demonstrated that nutrients such as oleic acid monounsaturated fatty acid (MUFA), omega-3 polyunsaturated fatty acids (PUFAs), vitamins A, B, C, D, and E, selenium and zinc elements, lycopene, olive oil polyphenols, resveratrol, organosulfured compounds, and catechins have a positive impact on MEtS components, improving the onset and development of the disease.
Moreover, animal studies have shown that omega-3 improves insulin sensitivity and reduces visceral fat storage [142,143]. Different randomized clinical trials have proven that omega-3 supplementation produces favorable hypolipidemic effects, a reduction in pro-inflammatory cytokine levels and improvement in glycaemia [144]. Animal and patient-based studies have shown that high omega-3 intake and low omega-6/omega-3 ratio are associated with lower MetS risk [145,146].
5.3. Vitamins
Vitamins are essential micronutrients for the proper functioning of metabolism. However, vitamins cannot be synthesized by the organism, or not in enough amount, so they must be obtained through diet. Vitamins can be classified into hydrosoluble (vitamins B and C) and liposoluble (vitamins A, D, E, and K). Their biological properties will be discussed below.
5.3.1. A Varied Diet. Vitamins B
Vitamins B comprise a set of molecules found in a wide variety of food groups such as dairy products (B2, B3, B5), eggs (B1, B3, B7, B12), fish and meat (B1, B3, B5,B6, B12), or plant-based foods (B1, B5, B6, B9). Vitamins B are essential in the KC, so deficiencies in some of these vitamins will lead to impaired mitochondrial metabolism, increased levels of mitochondrial ROS production, and decreased overall energy production [147].
Vitamin B1 (thiamin) is a cofactor of several enzymes such as the cytosolic pyruvate dehydrogenase or the mitochondrial ketoglutarate dehydrogenase; in vitro models demonstrate that its deficiency is linked to increased ROS formation [148]. Vitamin B2 (riboflavin) is essential as a prosthetic group for several enzymes catalyzing redox reactions; different studies demonstrated that its deficiency is associated with a loss of mitochondrial complex IV and induced oxidative stress [149]. Vitamin B3 (niacin) is an important cofactor involved in mitochondrial respiration reactions, glycolysis, or lipid oxidation; B3 have also demonstrated to have an important antioxidant activity in different investigations [150]. Vitamin B5 (pantothenic acid) is an important prosthetic group (CoA) involved in KC and lipid metabolism; cellular models also demonstrated an important role in the enzymatic antioxidant defense system promoting CAT, GR, or GPx [151]. Vitamin B6 (pyridoxal) have several functions in different metabolic pathways such one-carbon reactions, gluconeogenesis, or lipid metabolism, among others; even have an important antioxidant role through its activity in glutathione pathways, demonstrated in cellular models [152]. Vitamin B7 (biotin) is an important cofactor for some carboxylase enzymes involved in lipid metabolism; decreased B7 levels have been linked to increased ROS formation and impaired mitochondrial respiration [153]. Vitamin B9 (folic acid) exerts different biological functions like nucleotide synthesis or mitochondrial tRNA modification, among others; folate contributes to cellular redox states preventing oxidative stress indirectly [154]. Finally, vitamin B12 (cobalamin) is essential for nucleotide synthesis or succinyl-CoA generation; B12 deficiency has been related to indirect increased ROS levels due to glutathione dysregulation in patients [155].
5.3.2. Orange Juice. Vitamin C
Vitamin C, or ascorbic acid, is principally found in pepper (and other vegetables) and fruits (mainly in kiwi, guava, and citrus). This molecule is a potent antioxidant that activates Sirt1, triggering a signaling cascade that decrease ROS and apoptosis and also acts as a ROS quencher. Moreover, it is also involved in carnitine biosynthesis, the key factor in β-oxidation. Thus, vitamin C deficiency will lead to impaired ATP production, β-oxidation deficiency, and ROS formation [156,157], and therefore, mitochondrial dysfunction.
5.3.3. Carrot Cream. Vitamin A
Vitamin A or retinol is mainly found in carrots, sweet potatoes, or spinaches, as well as pork and beef liver, which concentrate the highest amounts. This vitamin affects mitochondrial function as it enhances the levels of the mitochondrial transcription factor A (mtTFA), a key transcription factor for mitochondrial function, and plays a key role in glycolytic energy generation, as an essential cofactor for protein kinase C delta (PKCẟ), which signals the pyruvate dehydrogenase complex for an enhanced flux of pyruvate into the KC [158]. Thus, deficiency of vitamin A has been associated with decreased respiration and ATP synthesis [158,159,160].
5.3.4. Slight Daily Sunbathe. Vitamin D
Vitamin D is naturally present in blue fish and egg yolks, among other foods. Furthermore, vitamin D can be synthesized by the human body when ultraviolet rays from sunlight strike the skin. However, after its obtention, either from sun exposure or food, it needs two hydroxylations to be biologically active.
Many biological functions of vitamin D are mediated by the control of the vitamin D receptor (VDR) on nuclear transcription. The VDR main function is to regulate Ca2+ homeostasis through the upregulation of calcium transporters. Moreover, VDR protects from an excessive respiratory activity and limits ROS production by controlling mitochondrial and nuclear transcription of the proteins involved in MRC and ATP synthesis [161,162].
5.3.5. Avocado Is the Answer. Vitamin E
Vitamin E is mainly found in avocado, leafy vegetables such as spinach and broccoli, sunflower seeds, nuts, and olive oil. Vitamin E is the most important antioxidant in cell membranes. It protects mitochondrial structure and function by maintaining mitochondrial membrane integrity, allowing the recovery of respiratory function and reducing lipid and protein oxidation [163]. This protective function would be, in part, explained by the inhibition of lipid peroxidation and the increase in CAT, SOD, GR, and GPx activity demonstrated in T2D rats [164].
5.3.6. Vitamins Against MetS
The role of vitamins in mitochondrial metabolism and their antioxidant properties suggest that they might play a role in the prevention of MetS. Indeed, a case control-study showed that plasma levels of vitamins A, C, E, and D were significantly lower in MetS patients compared to healthy subjects [165,166].
Different animal studies have suggested that vitamin A supplementation ameliorates obesity through UPC1-mediated thermogenesis and delays the appearance of T2D by increasing mtTFA [158,167]. Vitamin B7, biotin deficiency has been linked to impaired glucose tolerance, hyperglycemia, and decreased glucose oxidation in animal studies [168] and a cross-sectional study showed that high levels of vitamin B12 are protective to MetS [169]. Regarding vitamin C, different patient-based studies have suggested that intake or supplementation decreases the risk of MetS and improves the quality of life of MetS patients [169,170], especially when combined with physical exercise [171,172]. Moreover, a cross-sectional study with more than 2000 patients demonstrated that deficiency of vitamin C was associated with an increased likelihood of MetS [169]. Several epidemiologic studies have established a strong association between vitamin D deficiency and hypertension, obesity, and dyslipidemia [173,174]. In a randomized clinical trial with MetS patients, vitamin E was seen to exert beneficial effects on cytokines and the lipid profile [175]. Moreover, different studies have found that vitamin E could ameliorate the pathologic conditions associated with MetS such as hyperglycemia or obesity, although more studies in patients are needed to prove the consistency of these findings [175,176].
Nevertheless, when it comes to vitamins, overconsumption may be just as bad as scarcity. For instance, an excess of vitamin A intake (with a tolerable upper intake level for adults of 3000 μg/day) [177] promotes oxidative stress and mitochondrial death [178] and it is associated with an increased likelihood of MetS [160]. A high intake of vitamin B3 (niacin), can result in niacin-induced IR [179], and when Vitamin C and E are consumed in excess, they promote prooxidants activity instead of antioxidant function [147].
5.4. Trace Elements
Trace elements are minerals present in small amounts in living tissues. Some of them are nutritionally essential and so need to be incorporated through diet, such as selenium or zinc, and others are considered nonessential (or there is not consistent evidence to regard them as essentials) [180].
5.4.1. A Handful of Nuts. Selenium and Zinc
Selenium and zinc are mainly present in nuts, mushrooms, fish, selfish, and meat, with antioxidant and mitochondriogenic properties. Selenium promotes mitochondrial biogenesis through the stimulation of PGC-1a, while the nuclear redox factor1 (NRF-1) generation exerts its antioxidant role activating both GPx and GR enzymes. Some of these positive effects would be modulated by selenium impact on miRNAs profile preventing oxidative stress and inflammation [181]. On the other hand, zinc exerts its antioxidant power by promoting SOD and CAT activation, inhibiting important pro-oxidant enzymes such as NADPH oxidase and competing with redox-active transition metals such as iron and copper for certain binding sites (cell membranes, proteins), thereby prohibiting them from catalyzing the formation of ROS and the initiation of lipid peroxidation [182].
5.4.2. Trace Elements Against MetS
Some studies have shown that adequate intake of these trace elements is associated with an attenuation of MetS risk factors. In vivo and patient-based studies have found that recommended doses of selenium (1–2 μg/kg/day) act as an insulin-mimetic, attenuating diabetes [183,184,185] and have cardioprotective effects (increase plasma antioxidant capacity and decrease lipid peroxidation and LDL-cholesterol) [186,187,188]. Additionally, a cross-sectional study including more than 2000 patients showed that dietary selenium intake was negatively associated with MetS [170]. Regarding zinc, it is involved in different protective mechanisms against obesity, IR, hypertension, and dyslipidemia, which suggests potential roles of zinc in both the prevention and treatment of MetS. However, the current scientific evidence in humans regarding the associations between zinc status and occurrence of MetS is still inconsistent [189].
Despite these healthful and wellness properties of minerals, as it happens with vitamins, excess of these elements is just as bad as a deficiency. Fortunately, severe impact on human health of vitamins and minerals is only reached at very high concentrations. For instance, doses of selenium 100 times higher than expected had adverse effects than at recommended doses by increasing the risk of T2D and promoting CVD [190,191,192].
5.5. Polyphenols
Polyphenols are bioactive compounds with antioxidant properties mostly found in plant-based foods [193]. According to their molecular structure, polyphenols can be categorized into two main groups: flavonoids (such as catechins) and non-flavonoids, including phenolic acids (such as ellagic acid), stilbenes (such as resveratrol), lignans (such as pinoresinol), and others (in which we find oleuropein and hydroxytyrosol). Many of them show mitochondrial effects and have been reported to modulate MetS [194].
5.5.1. A Cup of Green Tea. Catechins
Inside the flavonoids family, we find the catechins molecules, present mainly in green tea (with a total minimum content of catechins of 8%), but also in a wide variety of fruits (Table 2). It has been demonstrated that catechins can directly work as ROS scavengers and metal ions chelators and activate some antioxidant enzymes such as CAT and SOD. Moreover, they can also inhibit some pro-oxidant enzymes like NADPH-oxidase and suppress stress-related signaling pathways (as those mediated by TNF or the activator protein 1). Some in vitro studies also demonstrated an anti-inflammatory effect of catechins through changing miRNAs profile [195]. All together, these effects contribute to decrease oxidative stress and improve mitochondrial function [196].
Table 2.
Summary of the principal nutrients with beneficial mitochondrial effects influencing metabolic syndrome.
| Nutrient | Molecular Group and Structure | Main Food Sources | RDA [197] | Principal Role in Mitochondria | Main Effect against Metabolic Syndrome | Main LoE | Reported References |
|---|---|---|---|---|---|---|---|
| Oleic acid |
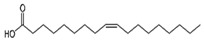 MUFA |
Olive oil Vegetables oils Nuts |
20 g of olive oil/day 1 | Antioxidant Antiapoptotic Enhances β-oxidation |
↓ Blood pressure | H | [125] |
| Improves lipid profile | H | [125] | |||||
| ↑ insulin sensitivity | H | [126,127] | |||||
| ↓ inflammation | H | [126,127] | |||||
| Reduces central adiposity | H | [128] | |||||
| Omega-3 |
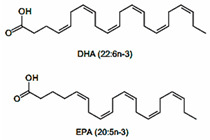 PUFA |
Fish and seafood Nuts and seeds |
1.1–1.6 g/day (ALA) | Uncoupling β-oxidation mitochondrial biogenesis |
hypolipidemic effects | H | [140,143,144] |
| ↑ insulin sensitivity | H | [142,144] | |||||
| ↓ inflammation | H | [140,144] | |||||
| ↓ MetS risk | H | [145,146] | |||||
| Vitamins B |
 Water-soluble vitamin 2 |
Dairy products (B2, B3, B5) Eggs (B1, B3, B7, B12) Fish and meat (B1, B3, B5, B12) Plant-based foods (B1, B5, B9) |
B1: 1.1–1.2 mg/day B2: 0.9–1.1 mg/day B3: 14–16 mg/day B5: 5 mg/day B6: 1.3 mg/day B7: 0.03 mg/day B9: 0.3–0.4 mg/day B12: 0.0024 mg/day |
Essential in Krebs Cycle Antioxidant |
Low levels are associated with hyperglycemia and insulin resistance | H | [167,179] |
| Protective to MetS | H | [168] | |||||
| Vitamin C |
 Water-soluble vitamin |
Guava Citrus fruit Kiwi Strawberries Peppers |
75–90 mg/day | Antioxidant Antiapoptotic Involved in β-oxidation |
↑ the effects of physical activity in the prevention of MetS and the quality of life of MetS patients | H | [168,169,170,171] |
| Vitamin A |
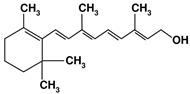 Fat-soluble vitamin |
Carrots Pork/beef liver Foie Spinach Sweet potato |
0.7–0.9 mg/day | Key role in mitochondrial respiration | Ameliorates obesity | A | [166] |
| Delays the appearance of diabetes | A | [158] | |||||
| Vitamin D |
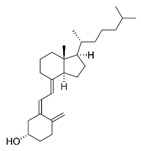 Fat-soluble vitamin 3 |
Blue fish Egg yolk Synthetized in the presence of sunlight exposure |
0.015 mg/day | Controls the respiratory activity and limits ROS production | Low levels are associated with hypertension, obesity and dyslipidemia | H | [161,162] |
| Vitamin E |
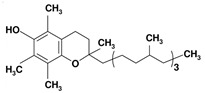 Fat-soluble vitamin |
Sunflower seeds Leafy vegetables Nuts Olive oil |
15 mg/day | Protects mitochondrial structure and function (antioxidant) | ↓ inflammation | H | [174] |
| Improves the lipid profile | H | [174] | |||||
| Selenium and Zinc 4 | NA Trace element |
Nuts Fish and selfish Meat |
Se: 0.055 mg/day Zn: 8–11 mg/day |
Antioxidant Mitochondrial biogenesis |
Insulin-mimetic | H | [183,184,185,189] |
| Improves lipid profile | A | [189,198] | |||||
| Cardioprotective | A | [186,187,188] | |||||
| Catechins |
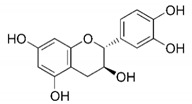 Flavonoid family Flavonoid family |
Green tea Broad beans Blueberries Black grapes Strawberries Apricots |
Up to 704 mg/day 1 | Antioxidants | ↓ Blood pressure | H | [199,200,201] |
| Improve lipid profile | H | [200,202] | |||||
| Insulin-like/-enhancing activities | H | [203,204] | |||||
| Resveratrol |
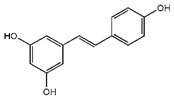 Polyphenols |
Grapes Apples Blueberries Pistachios Peanuts |
Up to 4 mg/day 1 | Mitochondrial protective agent (antioxidant) | ↓ body weight | H | [205,206] |
| ↓ waist circumference | H | [205,206] | |||||
| Improve lipid profile | H | [205] | |||||
| ↓ glucose levels | H | [205] | |||||
| Cardioprotective | H | [204,207,208] | |||||
| Oleuropein (OL), Hydroxytyrosol (HT), Pinoresinol |
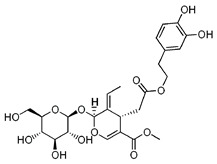 Oleuropein structure |
Olive oil | 20 g of olive oil/day 1 | Mitochondrial protective agent (antioxidant) | Cardioprotective | H | [204] |
| ↓ Insulin resistance | H | [204] | |||||
| ↓ Blood pressure | H | [209] | |||||
| Lycopene |
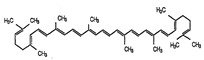 Carotenoid |
Tomato Watermelon Papaya Apricots Red grapefruit Carrots Pumpkin Sweet potato |
5.7–15 mg/day 1 | Antioxidant | ↓ Blood pressure | H | [210,211] |
| Anti-obesogenic | A | [212,213] | |||||
| improves the lipid profile | A | [214,215] | |||||
| ↑ insulin sensitivity and ↓ plasma glucose | H | [216,217] | |||||
| ↓ Risk of death in MetS patients | H | [211] | |||||
| Allicin |
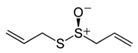 Organosulfur compounds |
Garlic | 1–2 g of raw garlic 1 | Antioxidant Uncoupling |
Improves obesity | H | [218,219,220,221,222] |
| Improves lipid profile | H | [223] | |||||
| ↓Hyperglycemia | H | [222] | |||||
| Cardioprotective | H | [224,225,226] | |||||
| ↓Hypertension | H | [223,227] |
1 Suggested recommended intake [228,229,230,231,232]. 2 Molecular structure from biotin (B7). 3 Molecular structure from cholecalciferol (D3). 4 The current scientific evidence in humans shows inconsistent results regarding the beneficial effects of zinc in MetS risk factors. RDA = Recommended dietary allowance; A = Animal studies; H = Human studies; NA = Not applicable.
5.5.2. Some Black Grapes. Resveratrol
Resveratrol is another antioxidant, mainly found in black grapes. Resveratrol is a mitochondrial protective agent. It activates Sirt1 which triggers different signaling pathways that promote antioxidant and anti-inflammatory effects, and parallelly activate PGC-1α, the master regulator of mitochondrial biogenesis. Sirt1 activation enhances β-oxidation and glucose utilization, promotes mitochondrial biogenesis, antioxidant detoxification, and the expression of uncoupling proteins [233,234]. Moreover, some of the anti-inflammatory effects of resveratrol are triggered by change in miRNAs profiles, as demonstrated in cell models [235] and clinical human trials [236]. Of note, resveratrol is one of those nutrients were herein reported beneficial in vitro effects have been observed with concentrations usually beyond those reached by food intake. Notwithstanding, and regardless the mechanism, clinically beneficial effects have also been reported [236]. Further studies are required to deepen in dose-response effects of resveratrol, their interaction with other nutrients, and host metabolism.
5.5.3. A Spoon of Olive Oil. Oleuropein, Hydroxytyrosol, and Pinoresinol
Olive oil have a great content of polyphenols such as oleuropein (OL), hydroxytyrosol (HT), and pinoresinol. Olive oil polyphenols (OOPs) are mitochondrial-protective agents for its antioxidant properties. It has lately emerged that OOPs, particularly HT, are able to induce Sirt1 expression. Moreover, Sirt1 interaction with nuclear redox factor 2 (NRF2) signaling, responsible for the transcriptional activation of antistress target genes, exerts a protective effect against oxidative stress. Furthermore, OL and HT can also scavenge ROS and OOPS have been demonstrated to activate antioxidant enzymes such as GPx and CAT. Finally, HT has also been reported to stabilize some mitochondrial complex-subunits and enhance the expression of PGC-1α and CPT-1 [204], with previously reported mitochondrial activity.
5.5.4. Polyphenols Against MetS
Different studies have proven the beneficial effects of polyphenols in the prevention of MetS. Animal studies have shown that green tea catechins lowers blood pressure by reducing ROS [199] and have lipid-lowering activities [202]. Moreover, catechins and OOPs have been demonstrated to enhance glucose tolerance and decrease events related to IR in both animal and human-based studies [203,204]. In addition, cardioprotective roles have been attributed to resveratrol and OOPs thanks to their ability to counteract inflammation and reduce LDL-oxidation [204,207,208].
In humans, a meta-analysis of 20 randomized clinical trials, with a total of 1536 participants who received green tea regularly, showed a slight decrease in systolic blood pressure and a moderate reduction of LDL cholesterol [200]. Another study with 48 MetS patients demonstrated that dietary achievable doses of blueberries significantly decreased blood pressure [201]. This blood pressure-reducing effect has also been attributed to OOPs by different intervention studies [209]. Regarding resveratrol, a meta-analysis including 16 studies, 10 human-based and 6 in vivo studies, showed that resveratrol intake meaningfully reduces the body weight, waist circumference, triglycerides, and glucose level [205]. Moreover, a meta-analysis including 28 randomized clinical trials also documented an improvement in obesity measures with resveratrol supplementation [206].
5.6. A Tomato Salad. Lycopene with Oleic Acid
Lycopene is a carotenoid found mainly in tomato, but also in other fruits such as watermelon, papaya, or red grapefruits. The lycopene content increases during different stages of fruit ripening, so the higher content of lycopene is found in ripe tomatoes [237].
However, lycopene undergoes photo-oxidation and degradation with light, which produces a decrease in bioavailability. The incorporation of lycopene in an oil phase overcomes photo-oxidation. An interesting option is the association of lycopene with olive oil, which prevents lycopene degradation and enriches its healthy properties with oleic acid, a monounsaturated fatty acid with antioxidant properties [238,239].
Lycopene is also a powerful antioxidant and anti-inflammatory molecule. It acts as both ROS and nitrogen species scavenger [240], decreases DNA damage [241,242,243], and modulates the production of SOD and GPx [244,245]. Moreover, it promotes the expression of constructs containing antioxidant response elements (ARE). Regarding its anti-inflammatory effects, lycopene reduces apoptosis as well as the expression of inflammatory cytokines [246]. Lycopene also exerts lipid-lowering properties through the induction of Sirt1 activity [247].
Lycopene Against MetS
Based on the potent antioxidant and lipid-lowering properties of lycopene, different studies have assessed it as a beneficial nutrient for the prevention and treatment of MetS. Animal and human-based studies have shown that lycopene reduces blood pressure [210,211], atherosclerotic burden [248] and improves blood antioxidant capacity [246,249], has an anti-obesity role [212,213], improves insulin sensitivity, reduces hyperglycemia [216,217], and improves the lipid profile [214,215]. Moreover, a retrospective study with 2500 patients with MetS showed that higher serum levels of lycopene are associated with a reduced risk of death [250].
5.7. Garlic Seasoning Is Always a Good Idea. Organosulfur Compounds (OSCs)
The OSCs comprise a group of molecules found in garlic that are accumulated as γ-glutamyl peptides. Depending on the processing status of the garlic, different OSCs can be present: alliin and sulfur amino acids from whole cloves, allicin from crushed cloves, ajoene and vinyldithiines from stir-fried garlic, and (poly)sulfides from steam-distilled garlic oil. However, the main active metabolite responsible for most of the biological activities of garlic is allicin [251].
The OSCs are potent antioxidant molecules, they decrease the activity of pro-oxidant molecules, such as NADPH oxidase, and increase the activity of antioxidant proteins, such as NRF2 [252,253]. Moreover, they also activate Sirt3, which prevents cardiac oxidative stress and mitochondrial dysfunction [224]. OSCs have also anti-inflammatory effects, as they suppress the activation of proinflammatory molecules such as NF-kB or TNF [254,255].
OSCs against MetS
In vitro and in vivo studies have highlighted the anti-adipogenic effect of OSCs through the promotion of mitochondrial uncoupling (increasing the expression of UPC-1) and the inhibition of adipocyte differentiation (through AMPK activation) [218,219,220,221], as well as their cardioprotective effects (decrease plasmatic insulin, total cholesterol, and oxidative stress levels) [224,225]. The cardioprotective effects of garlic have also been demonstrated in patient-based studies [226].
Moreover, different intervention studies have shown that garlic supplementation attenuates MetS abnormalities: reduces hypertension [222,227,256], improves lipid profile [222,223], decreases waist circumference and fasting blood glucose [222].
6. The Impact of Gut Microbiome on MetS. between Nutrients and Mitochondria
The human intestines are home to a vast number of bacteria, archaea, microbial eukaryotes, and viruses, collectively known as the gut microbiome [257]. Nutrients can modify microbiome composition (this is the case of yogurts and pickles) and, at the same time, microbiome can condition the absorption of some nutrients and, thus, our health. That is why gut microbiome can be considered another intrinsic factor contributing to MetS, and many other conditions.
Diet is accepted as the first driver of gut microbiome composition and not only the cause of preventable diseases. In 2007, it was demonstrated that a high-fat-diet affects the intestinal barrier leading the pass of microbiota products to systemic circulation. These molecules, such as lipopolysaccharide (LPS), showed deleterious effects on glycaemic function and activated the inflammatory response [258]. Recently, LPS has been also related to plasmatic decrease in high-density lipoprotein (HDL) cholesterol and increase in triglycerides [259,260].
In fact, the gut microbiome responds to diet composition [261], including changes depending on fat composition [262,263], fiber types [264,265], and food additives [266]. Moreov73er, some dietary interventions demonstrated changes in the microbiome population that negatively correlated with obesity and hypertension [267,268,269]. Some others do not. This is the case of dietary interventions with polyphenols in MetS patients, that despite decreasing the expression of MetS biomarkers, yielded to null differences in gut microbiota when compared to healthy subjects [270].
Furthermore, also gut microbiome metabolic end products like the short-chain fatty acids (SCFAs) that may play a role in MetS. Some SCFAs like butyrate appear diminished in inflammatory diseases [271,272], whilst another SCFAs study with propionate supplementation improved visceral adipose tissue levels and maintained insulin sensitivity [273]. Additionally, when produced in high amounts, these molecules have been also involved in obesity, among others [274].
Finally, gut microbiome can synthesize vitamin K, necessary for normal clotting activity, and some B vitamins, contributing to more than a quarter of the daily reference intake in the case of B3, B6, B9, and B12 [275]. All these vitamins are cofactors of some metabolic routes involved in proper mitochondrial function.
Beyond food and food compounds, some other interventions have an impact on gut microbiome and, consequently, on host metabolism, such as prebiotics, probiotics, and also fecal microbiome transplantation (FMT). Prebiotics are defined as non-digestible polysaccharides that promote “the selective stimulation of growth and/or activity(ies) of one or a limited number of microbial genus(era)/species in the gut microbiota that confer(s) health benefit to the host” [276]. In turn, probiotics are defined as “live microorganisms which when administered in adequate amounts, confer a beneficial health effect on the host” [277]. Several studies have been conducted in animal models [278,279,280] and humans [281,282,283,284], demonstrating benefits such as decrease in serum glucose, visceral fat area, waist circumference, or cholesterol levels. However, these results have not been always reproduced [285,286]. Finally, FMT has demonstrated that it can help reverse obesity in rodents, although there is no evidence in humans [287]. However, FMT has demonstrated to increase insulin sensitivity in MetS patients [288,289].
All these data suggest a role of gut microbiome in MetS, conditioning the interplay and effects of nutrient intake within the organism. However, this research area is quite novel and needs further assessment to understand the complex interactions between gut microbiome and its host.
7. Diet as a Therapy
7.1. Healthy Diet Base: Which Food and in Which Context
The nutrients and bioactive compounds reviewed above are just an example of the beneficial impact of some dietary patterns to avoid the onset and development of MetS and its associated complications. They exert a multitude of healthy actions, some of them through the modulation of mitochondrial functions.
For that purpose, dietary habits should focus on what and how we eat rather than on nutrients. This means predominating plant-based foods and fish, typical from the traditional MedDiet [290] rather than supplementation containing the nutrients or bioactive compounds (except for specific situations). Accordingly, a recent meta-analysis has shown that a prudent/healthy dietary pattern is a protective factor for MetS [291].
Nevertheless, multiple additional factors must be considered to achieve a health-promoting lifestyle. For instance, proper hydration it is very important. According to the European Food Safety Authority (EFSA) the adequate intake of daily total water for women and men is 2 and 2.5 L, respectively [292]. In fact, suboptimal hydration has an impact on health [293], metabolism [294] and T2D [295].
Additionally, a healthy lifestyle must minimize elements that have negative effect on health, and particularly on MetS such as tobacco [296], alcohol [297], or physical inactivity [298].
Moreover, related to the physical inactivity, some regular exercise has a positive impact on disease and on personal well-being [299]. Physical activity enhances energy consumption and reduces the risk of prevalent diseases such as T2D, CVD, and MetS [300]. Because of the increase of energy consumption, the metabolic disturbances associated with MetS such as increased blood glucose, triglycerides, or inflammation improve [300,301].
However, we must be careful with the intensity of physical exercise and avoid acute trainings, which has been related to an increase of ROS production [302]. For that purpose, the most beneficial practice will be moderate intensity training, promoting the proper balance between the ROS generation and the increase in the antioxidant defense [303].
7.2. Ketogenic Diet and Mitohormesis
Under some extreme situations, as primary mitochondrial diseases, changes in diet recommended by trained professionals may help to reconfigure mitochondrial activity and consequently, adverse clinical features [11,304].
This is the case of the ketogenic diet (KD), based on the intake of high-fat vs. low-carbohydrates content. This situation mimics starvation, forcing the body to use fat as energy source. Fatty acid oxidation results in ketogenesis [305] in the liver, with the consequent exportation of ketonic bodies throughout the rest of the body.
This diet must be prescribed and monitored by professionals to assess benefits and manage side effects [306]. Surprisingly, KD has shown anticonvulsant properties, at the clinical level, and the modulation of mitochondrial biogenesis [307] and the regulation of the levels of hormones or neurotransmitters [308], at the molecular level. These effects can be explained, at least in part, for an adaptive response called mitohormesis. Mitohormesis can be defined as the biological response where the induction of a reduced amount of mitochondrial stress leads to an increment in health and viability within a cell, tissue, or organism [309]. In this case, fat consumption would promote modest ROS increase, stimulating cell proliferation, differentiation, immunity, and adaptations that would eventually (and surprisingly) lead to enhancing resistance to oxidative stress in mitochondria [310,311].
About 25–30% of epilepsy patients worldwide show resistance to any pharmacological management (refractory epilepsy). Experiences on KD have shown a reduction in seizures greater than 50% [312,313], although the explanation for this effect remains unclear [314,315].
In inherited metabolic diseases such as Leigh syndrome, glucose transporter type 1 (Glut-1) and pyruvate dehydrogenase complex (PDH) deficiency and in a subset of patients with complex I deficiency [305,308], KD has also demonstrated to be useful. Under KD, most patients with epilepsy become seizure free and improve speech, motor, and mental development, and ability [316] despite its use is only partially able to reverse the clinical course of the neurodegenerative condition [317].
7.3. Enhancing MRC and Energy Buffering. Mitochondrial Burst
Some other mitochondrial diseases, contrarily, require the supplementation with CoQ10, riboflavin (vitamin B2), or thiamine (vitamin B1) to enhance ETC, or with creatine monohydrate, to increase energy buffering.
In some genetic diseases affecting enzymes involved in CoQ10 synthesis [318], supplementation can improve the clinical symptoms related to this deficiency [319]. Supplementation with riboflavin, a key building block in complex I and II, has particular usefulness in acyl-CoA dehydrogenase-9 (ACAD9) deficiency, resulting in increased complex I activity [320,321] and in the multiple acyl-CoA dehydrogenase deficiency caused by mutations in electron-transport flavoprotein dehydrogenase (ETFDH) [322,323]. Thiamine has been used, alone or in combination with other molecules, in the treatment of MELAS (mitochondrial myopathy, encephalopathy, lactic acidosis and stroke-like episodes), improving the symptoms of myopathy and lactic acidosis [324] and Leigh syndrome [325].
Finally, creatine binding to phosphate in the mitochondria is the main source of high energy in anaerobic metabolism. High energy demanding tissues like muscle or brain present the highest creatine levels [326]. In mitochondrial myopathy, the content of phosphocreatine is decreased [327], which is why supplementation with creatine monohydrate effectively improves the motor ability of patients with mitochondrial myopathy [328].
As explained, the complexity of dietary treatments for mitochondrial diseases requires the intervention of multidisciplinary and specialized units from clinical settings where both clinicians and dieticians design the proper treatment (including dietary interventions) for each affected patient. Although some recommendations may be similar, the dietetic support for mitochondrial diseases is set apart from the management of MetS.
8. Conclusions
The evidence herein discussed highlights the association of mitochondrial (dys)function with MetS. However, whether mitochondrial defects can be understood as a cause or consequence of MetS remains to be clarified. Similarly, whether all nutrients reach mitochondria and at which food dose do exert their beneficial effects is still controversial. In this sense, in vitro studies usually work with higher concentrations that significantly differ from those that are reached in human interventions and must be considered as models to understand molecular mechanisms difficult to approach in vivo. All approaches are needed to understand the interplay between nutrients, mitochondria, and MetS, or disease in general, where the effects of nutrients will differ also depending on their dose, their acute or chronic consumption, and their interaction with other food components or host characteristics. Despite growing interests, the complexity of reality is still far beyond current knowledge and information provided in the present review challenge new researchers to develop novel approaches. Whether nutrients exert their protective effect against MetS exclusively through mitochondria or by deploying a panoply of molecular effects beyond it, is also under consideration, at least for some of them.
However, some mitochondrial effects have been extensively documented in cell and animal models, as well as in patients. In particular, the above reviewed nutrients and bioactive compounds have been proved to improve mitochondrial function. In fact, oleic acid, omega-3, vitamins B and C, selenium/zinc, polyphenols, lycopene, and OSCs have been documented to improve mitochondrial antioxidant defense and reduce oxidative stress; oleic acid, lycopene, and OSCs have a potent anti-inflammatory role; vitamins A, B, and C are essential for mitochondrial energy generation; vitamin D and omega-3 regulate calcium homeostasis; vitamin E protects the mitochondrial structure; and selenium/zinc and some polyphenols stimulate mitochondrial biogenesis. At the same time, the consumption of all these elements has been associated with the reduction of MetS risk factors.
All this evidence confirms that a balanced diet represents a valuable therapeutic strategy to improve the global metabolic state, among others, through the enhancement of mitochondrial function. However, is not the intake of any of these isolated compounds by their own, as dietary supplements, but the synergic interplay of them all in foods and meals which makes the difference (for instance, lycopene of tomato with olive oil). Similarly, the excess of most of them in a diet is as bad as their absence. This is the case of the antioxidant paradox, since most of antioxidants, upon athreshold, become prooxidants compounds. In most cases, the balance is the key to obtain healthy effects within a diet. This is also the case of the intake of some foods such as fatty fish, with healthy effects for their composition in omega 3, but also prone to accumulate traces of some heavy metals and other pollutants with toxic effects at the nervous but also the cardiovascular system [329,330]. In these cases, moderate consumption is the best choice to promote healthy benefits.
Additionally, diet must not be the unique lifestyle intervention to ameliorate mitochondrial activity and MetS. In this regard, moderated physical activity has been also demonstrated to prevent and/or improve the metabolic alterations related with mitochondrial dys(function), as well as MetS, together with avoiding mitochondrial toxic drugs (as tobacco or alcohol).
In conclusion, following a balanced diet, like the MedDiet or traditional Asian diets, protects against the MetS (and many other diseases). The best medicine is in our hands; cheap, tasty, and close. However, the exact formula does not exist, maybe because it is specific for each subject. Despite all the advances in understanding the role of nutrition in mitochondrial function, the perfect dietary patterns and nutrients combination to preserve/enhance mitochondrial activity remains unclear, as well as how these dietary patterns get translated in enhanced metabolic health. For that purpose, further studies must be undertaken to fully understand the impact of a balanced diet on global/mitochondrial health. Moreover, a better comprehension of some novel intrinsic factors, such as gut microbiota interactions with nutrients and host metabolism, and extrinsic factors, such as the induced epigenetic response to dietary patterns, would contribute to the understanding of the complexity of the nutritional impact on health. Such findings should be effective for the development of the appropriate dietary interventions that may lead to avoid the onset and the development of one of the major health problems in the modern society, MetS. Interestingly, personalized medicine and individual dietary patterns will probably set the path for novel therapeutics and preventive medicine in coming years.
Acknowledgments
We are in debt to all the patients that participated in the reviewed papers, as well as to all the scientific staff that performed the studies, always devoted to their community.
Author Contributions
Conceptualization, F.J.G.-G. and G.G.; methodology all.; software F.J.G.-G. and A.M.-M.; validation all.; formal analysis all.; investigation F.J.G.-G. and A.M.-M.; resources F.C.; data curation F.J.G.-G. and A.M.-M.; writing—original draft preparation F.J.G.-G. and A.M.-M.; writing—review and editing, F.J.G.-G. and G.G.; visualization all; supervision G.G.; project administration all; funding acquisition F.C. and G.G. All authors have read and agreed to the published version of the manuscript.
Funding
The funders had no role in the study design, data collection and interpretation, or the decision to submit the work for publication, but contributed to support staff and material resources. This work was supported by funds from the Fondo de Investigaciones Sanitarias of the Instituto de Salud Carlos III (ISCIII) (grant numbers PI1800498 and PI1800451), the Centro de Investigación Biomédica en Red de Enfermedades Raras (CIBERER), all initiatives of ISCIII and FEDER (‘Una manera de hacer Europa’), as well as Fundació Privada Cellex and Suports a Grups de Recerca (grant code 2017 SGR 893) and CERCA programs from the Generalitat de Catalunya.
Conflicts of Interest
The authors declare no conflict of interest.
References
- 1.Cena H., Calder P.C. Defining a healthy diet: Evidence for the role of contemporary dietary patterns in health and disease. Nutrients. 2020;12:334. doi: 10.3390/nu12020334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.World Health Organization Healthy Diet. [(accessed on 4 May 2020)]; Available online: https://www.who.int/news-room/fact-sheets/detail/healthy-diet.
- 3.Kassi E., Pervanidou P., Kaltsas G., Chrousos G. Metabolic syndrome: Definitions and controversies. BMC Med. 2011;9:48. doi: 10.1186/1741-7015-9-48. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Bhatti J.S., Bhatti G.K., Reddy P.H. Mitochondrial dysfunction and oxidative stress in metabolic disorders—A step towards mitochondria based therapeutic strategies. Biochim. Biophys. Acta Mol. Basis Dis. 2016;1863:1066–1077. doi: 10.1016/j.bbadis.2016.11.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Bentley-Lewis R., Koruda K., Seely E.W. The metabolic syndrome in women. Nat. Clin. Pract. Endocrinol. Metab. 2007;3:696–704. doi: 10.1038/ncpendmet0616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Saklayen M.G. The global epidemic of the metabolic syndrome. Curr. Hypertens. Rep. 2018;20:1–8. doi: 10.1007/s11906-018-0812-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Imamura F., Micha R., Khatibzadeh S., Fahimi S., Shi P., Powles J., Mozaffarian D. Dietary quality among men and women in 187 countries in 1990 and 2010: A systematic assessment. Lancet Glob. Heal. 2015;3:e132–e142. doi: 10.1016/S2214-109X(14)70381-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Smith R.L., Soeters M.R., Wüst R.C.I., Houtkooper R.H. Metabolic flexibility as an adaptation to energy resources and requirements in health and disease. Endocr. Rev. 2018;39:489–517. doi: 10.1210/er.2017-00211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Leckey J.J., Hoffman N.J., Parr E.B., Devlin B.L., Trewin A.J., Stepto N.K., Morton J.P., Burke L.M., Hawley J.A. High dietary fat intake increases fat oxidation and reduces skeletal muscle mitochondrial respiration in trained humans. FASEB J. 2018;32:2979–2991. doi: 10.1096/fj.201700993R. [DOI] [PubMed] [Google Scholar]
- 10.Shenkin A. The key role of micronutrients. Clin. Nutr. 2006;25:1–13. doi: 10.1016/j.clnu.2005.11.006. [DOI] [PubMed] [Google Scholar]
- 11.Vidali S., Aminzadeh S., Lambert B., Rutherford T., Sperl W., Kofler B., Feichtinger R.G. Mitochondria: The ketogenic diet—A metabolism-based therapy. Int. J. Biochem. Cell Biol. 2015;63:55–59. doi: 10.1016/j.biocel.2015.01.022. [DOI] [PubMed] [Google Scholar]
- 12.Fabiani R., Naldini G., Chiavarini M. Dietary patterns and metabolic syndrome in adult subjects: A systematic review and meta-analysis. Nutrients. 2019;11:2056. doi: 10.3390/nu11092056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Devries S., Dalen J.E., Eisenberg D.M., Maizes V., Ornish D., Prasad A., Sierpina V., Weil A.T., Willett W. A deficiency of nutrition education in medical training. Am. J. Med. 2014;127:804–806. doi: 10.1016/j.amjmed.2014.04.003. [DOI] [PubMed] [Google Scholar]
- 14.Van Dam R.M., Rimm E.B., Willett W.C., Stampfer M.J., Hu F.B. Dietary patterns and risk for type 2 diabetes mellitus in U.S. men. Ann. Intern. Med. 2002;136:201–209. doi: 10.7326/0003-4819-136-3-200202050-00008. [DOI] [PubMed] [Google Scholar]
- 15.Locke A., Schneiderhan J., Zick S.M. Diets for health: Goals and guidelines. Am. Fam. Phys. 2018;97:721–728. [PubMed] [Google Scholar]
- 16.Lee K.W., Cho M.S. The traditional korean dietary pattern is associated with decreased risk of metabolic syndrome: Findings from the korean national health and nutrition examination survey, 1998–2009. J. Med. Food. 2014;17:43–56. doi: 10.1089/jmf.2013.3049. [DOI] [PubMed] [Google Scholar]
- 17.Liguori A., Petti F., Rughini S., Silli L., Asprino R., Maglio C., Leonetti F. Effect of a basic Chinese traditional diet in overweight patients. J. Tradit. Chin. Med. 2013;33:322–324. doi: 10.1016/S0254-6272(13)60173-9. [DOI] [PubMed] [Google Scholar]
- 18.Urpi-Sarda M., Casas R., Chiva-Blanch G., Romero-Mamani E.S., Valderas-Martínez P., Arranz S., Andres-Lacueva C., Llorach R., Medina-Remón A., Lamuela-Raventos R.M., et al. Virgin olive oil and nuts as key foods of the Mediterranean diet effects on inflammatory biomakers related to atherosclerosis. Pharmacol. Res. 2012;65:577–583. doi: 10.1016/j.phrs.2012.03.006. [DOI] [PubMed] [Google Scholar]
- 19.Campbell A.P. DASH eating plan: An eating pattern for diabetes management. Diabetes Spectr. 2017;30:76–81. doi: 10.2337/ds16-0084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Marcason W. What Are the components to the MIND diet? J. Acad. Nutr. Diet. 2015;115:1744. doi: 10.1016/j.jand.2015.08.002. [DOI] [PubMed] [Google Scholar]
- 21.Jayarathne S., Koboziev I., Park O.H., Oldewage-Theron W., Shen C.L., Moustaid-Moussa N. Anti-inflammatory and anti-obesity properties of food bioactive components: Effects on adipose tissue. Prev. Nutr. Food Sci. 2017;22:251–262. doi: 10.3746/pnf.2017.22.4.251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Cardinali D.P., Vigo D.E. Melatonin, mitochondria, and the metabolic syndrome. Cell. Mol. Life Sci. 2017;74:3941–3954. doi: 10.1007/s00018-017-2611-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Aschner P. Metabolic syndrome as a risk factor for diabetes. Expert Rev. Cardiovasc. Ther. 2010;8:407–412. doi: 10.1586/erc.10.13. [DOI] [PubMed] [Google Scholar]
- 24.Grundy S.M., Cleeman J.I., Daniels S.R., Donato K.A., Eckel R.H., Franklin B.A., Gordon D.J., Krauss R.M., Savage P.J., Smith S.C., et al. Diagnosis and management of the metabolic syndrome: An American Heart Association/National Heart, Lung, and Blood Institute scientific statement. Circulation. 2005;112:2735–2752. doi: 10.1161/CIRCULATIONAHA.105.169404. [DOI] [PubMed] [Google Scholar]
- 25.Katzmarzyk P.T., Leon A.S., Wilmore J.H., Skinner J.S., Rao D.C., Rankinen T., Bouchard C. Targeting the metabolic syndrome with exercise: Evidence from the HERITAGE family study. Med. Sci. Sports Exerc. 2003;35:1703–1709. doi: 10.1249/01.MSS.0000089337.73244.9B. [DOI] [PubMed] [Google Scholar]
- 26.Pérez-Martinez P., Mikhailidis D.P., Athyros V.G., Bullo M., Couture P., ıa Covas M.I., de Koning L., Delgado-Lista J., Diaz-Lopez A.D., Drevon C.A., et al. Lifestyle recommendations for the prevention and management of metabolic syndrome: An international panel recommendation. Nutr. Rev. 2017;75:307–326. doi: 10.1093/nutrit/nux014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Lane N., Martin W. The energetics of genome complexity. Nature. 2010;467:929–934. doi: 10.1038/nature09486. [DOI] [PubMed] [Google Scholar]
- 28.Bullon P., Marin-Aguilar F., Roman-Malo L. AMPK/Mitochondria in metabolic diseases. EXS. 2016;107:129–152. doi: 10.1007/978-3-319-43589-3_6. [DOI] [PubMed] [Google Scholar]
- 29.Spinelli J.B., Haigis M.C. The multifaceted contributions of mitochondria to cellular metabolism. Nat. Cell Biol. 2018;20:745–754. doi: 10.1038/s41556-018-0124-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Fillmore N., Mori J., Lopaschuk G.D. Mitochondrial fatty acid oxidation alterations in heart failure, ischaemic heart disease and diabetic cardiomyopathy. Br. J. Pharmacol. 2014;171:2080–2090. doi: 10.1111/bph.12475. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Karimi M., Pavlov V.I., Ziegler O., Sriram N., Yoon S.-Y., Agbortoko V., Alexandrova S., Asara J., Sellke F.W., Sturek M., et al. Robust effect of metabolic syndrome on major metabolic pathways in the myocardium. PLoS ONE. 2019;14:e0225857. doi: 10.1371/journal.pone.0225857. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Armani A., Berry A., Cirulli F., Caprio M. Molecular mechanisms underlying metabolic syndrome: The expanding role of the adipocyte. FASEB J. 2017;31:4240–4255. doi: 10.1096/fj.201601125RRR. [DOI] [PubMed] [Google Scholar]
- 33.Ristow M. Unraveling the truth about antioxidants. Nat. Med. 2014;20:709–711. doi: 10.1038/nm.3624. [DOI] [PubMed] [Google Scholar]
- 34.Muller F.L., Liu Y., van Remmen H. Complex III releases superoxide to both sides of the inner mitochondrial membrane. J. Biol. Chem. 2004;279:49064–49073. doi: 10.1074/jbc.M407715200. [DOI] [PubMed] [Google Scholar]
- 35.Murphy M.P. How mitochondria produce reactive oxygen species. Biochem. J. 2009;417:1–13. doi: 10.1042/BJ20081386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Hamanaka R.B., Chandel N.S. Mitochondrial reactive oxygen species regulate cellular signaling and dictate biological outcomes. Trends Biochem. Sci. 2010;35:505–513. doi: 10.1016/j.tibs.2010.04.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Aon M.A., Tocchetti C.G., Bhatt N., Paolocci N., Cortassa S. Protective mechanisms of mitochondria and heart function in diabetes. Antioxid. Redox Signal. 2015;22:1563–1586. doi: 10.1089/ars.2014.6123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Zhao R.Z., Jiang S., Zhang L., Yu Z. Bin Mitochondrial electron transport chain, ROS generation and uncoupling (Review) Int. J. Mol. Med. 2019;44:3–15. doi: 10.3892/ijmm.2019.4188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Pheiffer C., Jacobs C., Patel O., Ghoor S., Muller C., Louw J. Expression of UCP2 in Wistar rats varies according to age and the severity of obesity. J. Physiol. Biochem. 2016;72:25–32. doi: 10.1007/s13105-015-0454-4. [DOI] [PubMed] [Google Scholar]
- 40.González-Barroso M.M., Giurgea I., Bouillaud F., Anedda A., Bellanné-Chantelot C., Hubert L., de Keyzer Y., de Lonlay P., Ricquier D. Mutations in UCP2 in congenital hyperinsulinism reveal a role for regulation of insulin secretion. PLoS ONE. 2008;3:e3850. doi: 10.1371/journal.pone.0003850. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Lill R., Mühlenhoff U. Maturation of iron-sulfur proteins in Eukaryotes: Mechanisms, connected processes, and diseases. Annu. Rev. Biochem. 2008;77:669–700. doi: 10.1146/annurev.biochem.76.052705.162653. [DOI] [PubMed] [Google Scholar]
- 42.Baughman J.M., Perocchi F., Girgis H.S., Plovanich M., Belcher-Timme C.A., Sancak Y., Bao X.R., Strittmatter L., Goldberger O., Bogorad R.L., et al. Integrative genomics identifies MCU as an essential component of the mitochondrial calcium uniporter. Nature. 2011;476:341–345. doi: 10.1038/nature10234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Contreras L., Drago I., Zampese E., Pozzan T. Mitochondria: The calcium connection. Biochim. Biophys. Acta Bioenerg. 2010;1797:607–618. doi: 10.1016/j.bbabio.2010.05.005. [DOI] [PubMed] [Google Scholar]
- 44.Gazdar A.F., Bunn P.A., Minna J.D. Small-cell lung cancer: What we know, what we need to know and the path forward. Nat. Rev. Cancer. 2017;17:725–737. doi: 10.1038/nrc.2017.87. [DOI] [PubMed] [Google Scholar]
- 45.Monteith G.R., Prevarskaya N., Roberts-Thomson S.J. The calcium-cancer signalling nexus. Nat. Rev. Cancer. 2017;17:367–380. doi: 10.1038/nrc.2017.18. [DOI] [PubMed] [Google Scholar]
- 46.Meyer J.N., Leuthner T.C., Luz A.L. Mitochondrial fusion, fission, and mitochondrial toxicity. Toxicology. 2017;391:42–53. doi: 10.1016/j.tox.2017.07.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Nunnari J., Suomalainen A. Mitochondria: In sickness and in health. Cell. 2012;148:1145–1159. doi: 10.1016/j.cell.2012.02.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Gómez-Valadés A.G., Gonzalez-Franquesa A., Gama-Perez P., Claret M., Garcia-Roves P.M. Emerging concepts in diabetes: Mitochondrial dynamics and glucose homeostasis. Curr. Diabetes Rev. 2017;13 doi: 10.2174/1573399812666151012115229. [DOI] [PubMed] [Google Scholar]
- 49.Chen H., Vermulst M., Wang Y.E., Chomyn A., Prolla T.A., McCaffery J.M., Chan D.C. Mitochondrial fusion is required for mtdna stability in skeletal muscle and tolerance of mtDNA mutations. Cell. 2010;141:280–289. doi: 10.1016/j.cell.2010.02.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Chen H., McCaffery J.M., Chan D.C. Mitochondrial fusion protects against neurodegeneration in the cerebellum. Cell. 2007;130:548–562. doi: 10.1016/j.cell.2007.06.026. [DOI] [PubMed] [Google Scholar]
- 51.Wai T., Langer T. Mitochondrial dynamics and metabolic regulation. Trends Endocrinol. Metab. 2016;27:105–117. doi: 10.1016/j.tem.2015.12.001. [DOI] [PubMed] [Google Scholar]
- 52.Pieczenik S.R., Neustadt J. Mitochondrial dysfunction and molecular pathways of disease. Exp. Mol. Pathol. 2007;83:84–92. doi: 10.1016/j.yexmp.2006.09.008. [DOI] [PubMed] [Google Scholar]
- 53.Youle R.J., Narendra D.P. Mechanisms of mitophagy. Nat. Rev. Mol. Cell Biol. 2011;12:9–14. doi: 10.1038/nrm3028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Sliter D.A., Martinez J., Hao L., Chen X., Sun N., Fischer T.D., Burman J.L., Li Y., Zhang Z., Narendra D.P., et al. Parkin and PINK1 mitigate STING-induced inflammation. Nature. 2018;561:258–262. doi: 10.1038/s41586-018-0448-9. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 55.Kerr J.S., Adriaanse B.A., Greig N.H., Mattson M.P., Cader M.Z., Bohr V.A., Fang E.F. Mitophagy and Alzheimer’s Disease: Cellular and Molecular Mechanisms. Trends Neurosci. 2017;40:151–166. doi: 10.1016/j.tins.2017.01.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Fan H., He Z., Huang H., Zhuang H., Liu H., Liu X., Yang S., He P., Yang H., Feng D. Mitochondrial quality control in cardiomyocytes: A critical role in the progression of cardiovascular diseases. Front. Physiol. 2020;11:252. doi: 10.3389/fphys.2020.00252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Lee S., Min K.T. The interface between ER and mitochondria: Molecular compositions and functions. Mol. Cells. 2018;41:1000–1007. doi: 10.14348/molcells.2018.0438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Area-Gomez E., del Carmen Lara Castillo M., Tambini M.D., Guardia-Laguarta C., de Groof A.J.C., Madra M., Ikenouchi J., Umeda M., Bird T.D., Sturley S.L., et al. Upregulated function of mitochondria-associated ER membranes in Alzheimer disease. EMBO J. 2012;31:4106–4123. doi: 10.1038/emboj.2012.202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.McLelland G.L., Goiran T., Yi W., Dorval G., Chen C.X., Lauinger N.D., Krahn A.I., Valimehr S., Rakovic A., Rouiller I., et al. Mfn2 ubiquitination by PINK1/parkin gates the p97-dependent release of ER from mitochondria to drive mitophagy. Elife. 2018;7:e32866. doi: 10.7554/eLife.32866. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.López-Crisosto C., Bravo-Sagua R., Rodriguez-Peña M., Mera C., Castro P.F., Quest A.F.G., Rothermel B.A., Cifuentes M., Lavandero S. ER-to-mitochondria miscommunication and metabolic diseases. Biochim. Biophys. Acta Mol. Basis Dis. 2015;1852:2096–2105. doi: 10.1016/j.bbadis.2015.07.011. [DOI] [PubMed] [Google Scholar]
- 61.Lu C., Thompson C.B. Metabolic regulation of epigenetics. Cell Metab. 2012;16:9–17. doi: 10.1016/j.cmet.2012.06.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Suzuki M.M., Bird A. DNA methylation landscapes: Provocative insights from epigenomics. Nat. Rev. Genet. 2008;9:465–476. doi: 10.1038/nrg2341. [DOI] [PubMed] [Google Scholar]
- 63.Devall M., Mill J., Lunnon K. The mitochondrial epigenome: A role in Alzheimer’s disease? Epigenomics. 2014;6:665–675. doi: 10.2217/epi.14.50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Rocha M., Apostolova N., Herance J.R., Rovira-Llopis S., Hernandez-Mijares A., Victor V.M. Perspectives and potential applications of mitochondria-targeted antioxidants in cardiometabolic diseases and type 2 diabetes. Med. Res. Rev. 2014;34:160–189. doi: 10.1002/med.21285. [DOI] [PubMed] [Google Scholar]
- 65.North B.J., Verdin E. Sirtuins: Sir2-related NAD-dependent protein deacetylases. Genome Biol. 2004;5:224. doi: 10.1186/gb-2004-5-5-224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Finkel T., Deng C.X., Mostoslavsky R. Recent progress in the biology and physiology of sirtuins. Nature. 2009;460:587–591. doi: 10.1038/nature08197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Lombard D.B., Schwer B., Alt F.W., Mostoslavsky R. SIRT6 in DNA repair, metabolism and ageing. J. Intern. Med. 2008;263:128–141. doi: 10.1111/j.1365-2796.2007.01902.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Cohen H.Y., Miller C., Bitterman K.J., Wall N.R., Hekking B., Kessler B., Howitz K.T., Gorospe M., de Cabo R., Sinclair D.A. Calorie restriction promotes mammalian cell survival by inducing the SIRT1 deacetylase. Science. 2004;305:390–392. doi: 10.1126/science.1099196. [DOI] [PubMed] [Google Scholar]
- 69.Wang C., Chen L., Hou X., Li Z., Kabra N., Ma Y., Nemoto S., Finkel T., Gu W., Cress W.D., et al. Interactions between E2F1 and SirT1 regulate apoptotic response to DNA damage. Nat. Cell Biol. 2006;8:1025–1031. doi: 10.1038/ncb1468. [DOI] [PubMed] [Google Scholar]
- 70.Bell E.L., Guarente L. The SirT3 divining rod points to oxidative stress. Mol. Cell. 2011;42:561–568. doi: 10.1016/j.molcel.2011.05.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Schwer B., Verdin E. Conserved metabolic regulatory functions of sirtuins. Cell Metab. 2008;7:104–112. doi: 10.1016/j.cmet.2007.11.006. [DOI] [PubMed] [Google Scholar]
- 72.Sun C., Zhang F., Ge X., Yan T., Chen X., Shi X., Zhai Q. SIRT1 improves insulin sensitivity under insulin-resistant conditions by repressing PTP1B. Cell Metab. 2007;6:307–319. doi: 10.1016/j.cmet.2007.08.014. [DOI] [PubMed] [Google Scholar]
- 73.Weyrich P., Machicao F., Reinhardt J., Machann J., Schick F., Tschritter O., Stefan N., Fritsche A., Häring H.U. SIRT1 genetic variants associate with the metabolic response of Caucasians to a controlled lifestyle intervention-The TULIP Study. BMC Med. Genet. 2008;9:100. doi: 10.1186/1471-2350-9-100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Peeters A.V., Beckers S., Verrijken A., Mertens I., Roevens P., Peeters P.J., Hul W., Gaal L.F. Association of SIRT1 gene variation with visceral obesity. Hum. Genet. 2008;124:431–436. doi: 10.1007/s00439-008-0567-8. [DOI] [PubMed] [Google Scholar]
- 75.Obri A., Claret M. The role of epigenetics in hypothalamic energy balance control: Implications for obesity. Cell Stress. 2019;3:208–220. doi: 10.15698/cst2019.07.191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Schlicker C., Gertz M., Papatheodorou P., Kachholz B., Becker C.F.W., Steegborn C. Substrates and regulation mechanisms for the human mitochondrial sirtuins Sirt3 and Sirt5. J. Mol. Biol. 2008;382:790–801. doi: 10.1016/j.jmb.2008.07.048. [DOI] [PubMed] [Google Scholar]
- 77.Hirschey M.D., Shimazu T., Jing E., Grueter C.A., Collins A.M., Aouizerat B., Stančáková A., Goetzman E., Lam M.M., Schwer B., et al. SIRT3 deficiency and mitochondrial protein hyperacetylation accelerate the development of the metabolic syndrome. Mol. Cell. 2011;44:177–190. doi: 10.1016/j.molcel.2011.07.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Lombard D.B., Alt F.W., Cheng H.L., Bunkenborg J., Streeper R.S., Mostoslavsky R., Kim J., Yancopoulos G., Valenzuela D., Murphy A., et al. Mammalian Sir2 homolog SIRT3 regulates global Mitochondrial Lysine Acetylation. Mol. Cell. Biol. 2007;27:8807–8814. doi: 10.1128/MCB.01636-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Geiger J., Dalgaard L.T. Interplay of mitochondrial metabolism and microRNAs. Cell. Mol. Life Sci. 2017;74:631–646. doi: 10.1007/s00018-016-2342-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Ro S., Ma H.Y., Park C., Ortogero N., Song R., Hennig G.W., Zheng H., Lin Y.M., Moro L., Hsieh J.T., et al. The mitochondrial genome encodes abundant small noncoding RNAs. Cell Res. 2013;23:759–774. doi: 10.1038/cr.2013.37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Gallagher I.J., Scheele C., Keller P., Nielsen A.R., Remenyi J., Fischer C.P., Roder K., Babraj J., Wahlestedt C., Hutvagner G., et al. Integration of microRNA changes in vivo identifies novel molecular features of muscle insulin resistance in type 2 diabetes. Genome Med. 2010;2:9. doi: 10.1186/gm130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Piletič K., Kunej T. MicroRNA epigenetic signatures in human disease. Arch. Toxicol. 2016;90:2405–2419. doi: 10.1007/s00204-016-1815-7. [DOI] [PubMed] [Google Scholar]
- 83.Saeedi Borujeni M.J., Esfandiary E., Taheripak G., Codoñer-Franch P., Alonso-Iglesias E., Mirzaei H. Molecular aspects of diabetes mellitus: Resistin, microRNA, and exosome. J. Cell. Biochem. 2018;119:1257–1272. doi: 10.1002/jcb.26271. [DOI] [PubMed] [Google Scholar]
- 84.Bussler S., Penke M., Flemming G., Elhassan Y.S., Kratzsch J., Sergeyev E., Lipek T., Vogel M., Spielau U., Körner A., et al. Novel insights in the metabolic syndrome in childhood and adolescence. Horm. Res. Paediatr. 2017;88:181–193. doi: 10.1159/000479510. [DOI] [PubMed] [Google Scholar]
- 85.Marques-Rocha J.L., Milagro F.I., Mansego M.L., Zulet M.A., Bressan J., Martínez J.A. Expression of inflammation-related miRNAs in white blood cells from subjects with metabolic syndrome after 8 wk of following a Mediterranean diet-based weight loss program. Nutrition. 2016;32:48–55. doi: 10.1016/j.nut.2015.06.008. [DOI] [PubMed] [Google Scholar]
- 86.Zhang L., Hou D., Chen X., Li D., Zhu L., Zhang Y., Li J., Bian Z., Liang X., Cai X., et al. Exogenous plant MIR168a specifically targets mammalian LDLRAP1: Evidence of cross-kingdom regulation by microRNA. Cell Res. 2012;22:107–126. doi: 10.1038/cr.2011.158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Title A.C., Denzler R., Stoffel M. Uptake and function studies of maternal milk-derived MicroRNAs. J. Biol. Chem. 2015;290:23680–23691. doi: 10.1074/jbc.M115.676734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Snow J.W., Hale A.E., Isaacs S.K., Baggish A.L., Chan S.Y. Ineffective delivery of diet-derived microRNAs to recipient animal organisms. RNA Biol. 2013;10:1107–1116. doi: 10.4161/rna.24909. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Micó V., Martín R., Lasunción M.A., Ordovás J.M., Daimiel L. Unsuccessful detection of plant MicroRNAs in beer, extra virgin olive oil and human plasma after an acute ingestion of extra virgin olive oil. Plant Foods Hum. Nutr. 2016;71:102–108. doi: 10.1007/s11130-016-0534-9. [DOI] [PubMed] [Google Scholar]
- 90.Kowaltowski A.J., de Souza-Pinto N.C., Castilho R.F., Vercesi A.E. Mitochondria and reactive oxygen species. Free Radic. Biol. Med. 2009;47:333–343. doi: 10.1016/j.freeradbiomed.2009.05.004. [DOI] [PubMed] [Google Scholar]
- 91.Liesa M., Shirihai O.S. Mitochondrial dynamics in the regulation of nutrient utilization and energy expenditure. Cell Metab. 2013;17:491–506. doi: 10.1016/j.cmet.2013.03.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Rocha M., Rovira-Llopis S., Banuls C., Bellod L., Falcon R., Castello R., Morillas C., Herance J., Hernandez-Mijares A., Victor V. Mitochondrial dysfunction and oxidative stress in insulin resistance. Curr. Pharm. Des. 2013;19:5730–5741. doi: 10.2174/13816128113199990373. [DOI] [PubMed] [Google Scholar]
- 93.Parish R., Petersen K.F. Mitochondrial dysfunction and type 2 diabetes. Curr. Diabetes Rep. 2005;5:177–183. doi: 10.1007/s11892-005-0006-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Kim J.A., Wei Y., Sowers J.R. Role of mitochondrial dysfunction in insulin resistance. Circ. Res. 2008;102:401–414. doi: 10.1161/CIRCRESAHA.107.165472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.De Pauw A., Tejerina S., Raes M., Keijer J., Arnould T. Mitochondrial (dys)function in adipocyte (de)differentiation and systemic metabolic alterations. Am. J. Pathol. 2009;175:927–939. doi: 10.2353/ajpath.2009.081155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Kelley D.E., He J., Menshikova E.V., Ritov V.B. Dysfunction of mitochondria in human skeletal muscle in type 2 diabetes. Diabetes. 2002;51:2944–2950. doi: 10.2337/diabetes.51.10.2944. [DOI] [PubMed] [Google Scholar]
- 97.Krebs M., Roden M. Molecular mechanisms of lipid-induced insulin resistance in muscle, liver and vasculature. Diabetes Obes. Metab. 2005;7:621–632. doi: 10.1111/j.1463-1326.2004.00439.x. [DOI] [PubMed] [Google Scholar]
- 98.Sergi D., Naumovski N., Heilbronn L.K., Abeywardena M., O’Callaghan N., Lionetti L., Luscombe-Marsh N. Mitochondrial (dys)function and insulin resistance: From pathophysiological molecular mechanisms to the impact of diet. Front. Physiol. 2019;10:532. doi: 10.3389/fphys.2019.00532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Leguisamo N.M., Lehnen A.M., Machado U.F., Okamoto M.M., Markoski M.M., Pinto G.H., Schaan B.D. GLUT4 content decreases along with insulin resistance and high levels of inflammatory markers in rats with metabolic syndrome. Cardiovasc. Diabetol. 2012;11:100. doi: 10.1186/1475-2840-11-100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Pirola L., Johnston A.M., van Obberghen E. Modulation of insulin action. Diabetologia. 2004;47:170–184. doi: 10.1007/s00125-003-1313-3. [DOI] [PubMed] [Google Scholar]
- 101.Cooper S.A., Whaley-Connell A., Habibi J., Wei Y., Lastra G., Manrique C., Stas S., Sowers J.R. Renin-angiotensin-aldosterone system and oxidative stress in cardiovascular insulin resistance. Am. J. Physiol. Hear. Circ. Physiol. 2007;293:2009–2023. doi: 10.1152/ajpheart.00522.2007. [DOI] [PubMed] [Google Scholar]
- 102.Després J.P., Lemieux I. Abdominal obesity and metabolic syndrome. Nature. 2006;444:881–887. doi: 10.1038/nature05488. [DOI] [PubMed] [Google Scholar]
- 103.Berg A.H., Scherer P.E. Adipose tissue, inflammation, and cardiovascular disease. Circ. Res. 2005;96:939–949. doi: 10.1161/01.RES.0000163635.62927.34. [DOI] [PubMed] [Google Scholar]
- 104.Geto Z., Molla M.D., Challa F., Belay Y., Getahun T. Mitochondrial dynamic dysfunction as a main triggering factor for inflammation associated chronic non-communicable diseases. J. Inflamm. Res. 2020;13:97–107. doi: 10.2147/JIR.S232009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Couillard C., Ruel G., Archer W.R., Pomerleau S., Bergeron J., Couture P., Lamarche B., Bergeron N. Circulating levels of oxidative stress markers and endothelial adhesion molecules in men with abdominal obesity. J. Clin. Endocrinol. Metab. 2005;90:6454–6459. doi: 10.1210/jc.2004-2438. [DOI] [PubMed] [Google Scholar]
- 106.Fujita K., Nishizawa H., Funahashi T., Shimomura I., Shimabukuro M. Systemic Oxidative Stress is Associated With Visceral Fat Accumulation and the Metabolic Syndrome. Circ. J. 2006;70:1437–1442. doi: 10.1253/circj.70.1437. [DOI] [PubMed] [Google Scholar]
- 107.Nisoli E., Clementi E., Carruba M.O., Moncada S. Defective mitochondrial biogenesis: A hallmark of the high cardiovascular risk in the metabolic syndrome? Circ. Res. 2007;100:795–806. doi: 10.1161/01.RES.0000259591.97107.6c. [DOI] [PubMed] [Google Scholar]
- 108.Choo H.J., Kim J.H., Kwon O.B., Lee C.S., Mun J.Y., Han S.S., Yoon Y.S., Yoon G., Choi K.M., Ko Y.G., et al. Mitochondria are impaired in the adipocytes of type 2 diabetic mice. Diabetologia. 2006;49:784–791. doi: 10.1007/s00125-006-0170-2. [DOI] [PubMed] [Google Scholar]
- 109.Mansego M.L., Solar G.D.M., Alonso M.P., Martínez F., Sáez G.T., Escudero J.C.M., Redón J., Chaves F.J. Polymorphisms of antioxidant enzymes, blood pressure and risk of hypertension. J. Hypertens. 2011;29:492–500. doi: 10.1097/HJH.0b013e328341f1b2. [DOI] [PubMed] [Google Scholar]
- 110.Togliatto G., Lombardo G., Brizzi M.F. The future challenge of reactive oxygen species (ROS) in hypertension: From bench to bed side. Int. J. Mol. Sci. 2017;18:1988. doi: 10.3390/ijms18091988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Linseisen J., Welch A.A., Ocké M., Amiano P., Agnoli C., Ferrari P., Sonestedt E., Chajès V., Bueno-de-Mesquita H.B., Kaaks R., et al. Dietary fat intake in the european prospective investigation into cancer and nutrition: Results from the 24-h dietary recalls. Eur. J. Clin. Nutr. 2009;63:S61–S80. doi: 10.1038/ejcn.2009.75. [DOI] [PubMed] [Google Scholar]
- 112.Gillingham L.G., Harris-Janz S., Jones P.J.H. Dietary monounsaturated fatty acids are protective against metabolic syndrome and cardiovascular disease risk factors. Lipids. 2011;46:209–228. doi: 10.1007/s11745-010-3524-y. [DOI] [PubMed] [Google Scholar]
- 113.Sargsyan E., Artemenko K., Manukyan L., Bergquist J., Bergsten P. Oleate protects beta-cells from the toxic effect of palmitate by activating pro-survival pathways of the ER stress response. Biochim. Biophys. Acta Mol. Cell Biol. Lipids. 2016;1861:1151–1160. doi: 10.1016/j.bbalip.2016.06.012. [DOI] [PubMed] [Google Scholar]
- 114.Yuzefovych L., Wilson G., Rachek L. Different effects of oleate vs. palmitate on mitochondrial function, apoptosis, and insulin signaling in L6 skeletal muscle cells: Role of oxidative stress. Am. J. Physiol. Endocrinol. Metab. 2010;299 doi: 10.1152/ajpendo.00238.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Nakamura S., Takamura T., Matsuzawa-Nagata N., Takayama H., Misu H., Noda H., Nabemoto S., Kurita S., Ota T., Ando H., et al. Palmitate induces insulin resistance in H4IIEC3 hepatocytes through reactive oxygen species produced by mitochondria. J. Biol. Chem. 2009;284:14809–14818. doi: 10.1074/jbc.M901488200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Sacks D., Baxter B., Campbell B.C.V., Carpenter J.S., Cognard C., Dippel D., Eesa M., Fischer U., Hausegger K., Hirsch J.A., et al. Multisociety consensus quality improvement revised consensus statement for endovascular therapy of acute ischemic stroke. Int. J. Stroke. 2018;13:612–632. doi: 10.1016/j.jvir.2017.11.026. [DOI] [PubMed] [Google Scholar]
- 117.Castellano J.M., Garcia-Rodriguez S., Espinosa J.M., Millan-Linares M.C., Rada M., Perona J.S. Oleanolic acid exerts a neuroprotective effect against microglial cell activation by modulating cytokine release and antioxidant defense systems. Biomolecules. 2019;9:683. doi: 10.3390/biom9110683. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Palomer X., Pizarro-Delgado J., Barroso E., Vázquez-Carrera M. Palmitic and oleic acid: The yin and yang of fatty acids in type 2 diabetes mellitus. Trends Endocrinol. Metab. 2018;29:178–190. doi: 10.1016/j.tem.2017.11.009. [DOI] [PubMed] [Google Scholar]
- 119.Cantó C., Gerhart-Hines Z., Feige J.N., Lagouge M., Noriega L., Milne J.C., Elliott P.J., Puigserver P., Auwerx J. AMPK regulates energy expenditure by modulating NAD + metabolism and SIRT1 activity. Nature. 2009;458:1056–1060. doi: 10.1038/nature07813. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Lim J.H., Gerhart-Hines Z., Dominy J.E., Lee Y., Kim S., Tabata M., Xiang Y.K., Puigserver P. Oleic acid stimulates complete oxidation of fatty acids through protein kinase A-dependent activation of SIRT1-PGC1α complex. J. Biol. Chem. 2013;288:7117–7126. doi: 10.1074/jbc.M112.415729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Coll T., Eyre E., Rodríguez-Calvo R., Palomer X., Sánchez R.M., Merlos M., Laguna J.C., Vázquez-Carrera M. Oleate reverses palmitate-induced insulin resistance and inflammation in skeletal muscle cells. J. Biol. Chem. 2008;283:11107–11116. doi: 10.1074/jbc.M708700200. [DOI] [PubMed] [Google Scholar]
- 122.Henique C., Mansouri A., Fumey G., Lenoir V., Girard J., Bouillaud F., Prip-Buus C., Cohen I. Increased mitochondrial fatty acid oxidation is sufficient to protect skeletal muscle cells from palmitate-induced apoptosis. J. Biol. Chem. 2010;285:36818–36827. doi: 10.1074/jbc.M110.170431. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Schwingshackl L., Hoffmann G. Monounsaturated fatty acids, olive oil and health status: A systematic review and meta-analysis of cohort studies. Lipids Health Dis. 2014;13:154. doi: 10.1186/1476-511X-13-154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Estruch R., Ros E., Salas-Salvadó J., Covas M.I., Corella D., Arós F., Gómez-Gracia E., Ruiz-Gutiérrez V., Fiol M., Lapetra J., et al. Primary prevention of cardiovascular disease with a mediterranean diet. N. Engl. J. Med. 2013;368:1279–1290. doi: 10.1056/NEJMoa1200303. [DOI] [PubMed] [Google Scholar]
- 125.Vafeiadou K., Weech M., Altowaijri H., Todd S., Yaqoob P., Jackson K.G., Lovegrove J.A. Replacement of saturated with unsaturated fats had no impact on vascular function but beneficial effects on lipid biomarkers, E-selectin, and blood pressure: Results from the randomized, controlled Dietary Intervention and VAScular function (DIVAS) study. Am. J. Clin. Nutr. 2015;102:40–48. doi: 10.3945/ajcn.114.097089. [DOI] [PubMed] [Google Scholar]
- 126.Perdomo L., Beneit N., Otero Y.F., Escribano Ó., Díaz-Castroverde S., Gómez-Hernández A., Benito M. Protective role of oleic acid against cardiovascular insulin resistance and in the early and late cellular atherosclerotic process. Cardiovasc. Diabetol. 2015;14 doi: 10.1186/s12933-015-0237-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Due A., Larsen T.M., Hermansen K., Stender S., Holst J.J., Toubro S., Martinussen T., Astrup A. Comparison of the effects on insulin resistance and glucose tolerance of 6-mo high-monounsaturated-fat, low-fat, and control diets. Am. J. Clin. Nutr. 2008;87:1–3. doi: 10.1093/ajcn/87.4.855. [DOI] [PubMed] [Google Scholar]
- 128.Tutunchi H., Ostadrahimi A., Saghafi-Asl M. The effects of diets enriched in monounsaturated oleic acid on the management and prevention of obesity: A systematic review of human intervention studies. Adv. Nutr. 2020;11:864–877. doi: 10.1093/advances/nmaa013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Lanza I.R., Blachnio-Zabielska A., Johnson M.L., Schimke J.M., Jakaitis D.R., Lebrasseur N.K., Jensen M.D., Sreekumaran Nair K., Zabielski P. Influence of fish oil on skeletal muscle mitochondrial energetics and lipid metabolites during high-fat diet. Am. J. Physiol. Endocrinol. Metab. 2013;304:e1391–e1403. doi: 10.1152/ajpendo.00584.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Power G.W., Newsholme E.A. Dietary fatty acids influence the activity and metabolic control of mitochondrial carnitine palmitoyltransferase I in rat heart and skeletal muscle. J. Nutr. 1997;127:2142–2150. doi: 10.1093/jn/127.11.2142. [DOI] [PubMed] [Google Scholar]
- 131.Motawi T.M.K., Hashem R.M., Rashed L.A., El-Razek S.M.A. Comparative study between the effect of the peroxisome proliferator activated receptor-α ligands fenofibrate and n-3 polyunsaturated fatty acids on activation of 5′-AMP-activated protein kinase-α1 in high-fat fed rats. J. Pharm. Pharmacol. 2009;61:1339–1346. doi: 10.1211/jpp.61.10.0010. [DOI] [PubMed] [Google Scholar]
- 132.Lorrente-Cabrián S., Bustos M., Marti A., Martinez J.A., Moreno-Aliaga M.J. Eicosapentaenoic acid stimulates AMP-activated protein kinase and increses visfatin secretion in cultured murine adipocytes. Clin. Sci. 2009;117:243–249. doi: 10.1042/CS20090020. [DOI] [PubMed] [Google Scholar]
- 133.Baillie R.A., Takada R., Nakamura M., Clarke S.D. Coordinate induction of peroxisomal acyl-CoA oxidase and UCP-3 by dietary fish oil: A mechanism for decreased body fat deposition. Prostaglandins Leukot. Essent. Fat. Acids. 1999;60:351–356. doi: 10.1016/S0952-3278(99)80011-8. [DOI] [PubMed] [Google Scholar]
- 134.Lepretti M., Martucciello S., Aceves M.A.B., Putti R., Lionetti L. Omega-3 fatty acids and insulin resistance: Focus on the regulation of mitochondria and endoplasmic reticulum stress. Nutrients. 2018;10:350. doi: 10.3390/nu10030350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Zheng Z., Ge Y., Zhang J., Xue M., Li Q., Lin D., Ma W. PUFA diets alter the microRNA expression profiles in an inflammation rat model. Mol. Med. Rep. 2015;11:4149–4157. doi: 10.3892/mmr.2015.3318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Ortega F.J., Cardona-Alvarado M.I., Mercader J.M., Moreno-Navarrete J.M., Moreno M., Sabater M., Fuentes-Batllevell N., Ramírez-Chávez E., Ricart W., Molina-Torres J., et al. Circulating profiling reveals the effect of a polyunsaturated fatty acid-enriched diet on common microRNAs. J. Nutr. Biochem. 2015;26:1095–1101. doi: 10.1016/j.jnutbio.2015.05.001. [DOI] [PubMed] [Google Scholar]
- 137.Yin J., Peng M., Lin W. Visualization of mitochondrial viscosity in inflammation, fatty liver, and cancer living mice by a robust fluorescent probe. Anal. Chem. 2019;91:8415–8421. doi: 10.1021/acs.analchem.9b01293. [DOI] [PubMed] [Google Scholar]
- 138.Holm T., Berge R.K., Andreassen A.K., Ueland T., Kjekshus J., Simonsen S., Frøland S., Gullestad L., Aukrust P. Omega-3 fatty acids enhance tumor necrosis factor-α levels in heart transplant recipients. Transplantation. 2001;72:706–711. doi: 10.1097/00007890-200108270-00025. [DOI] [PubMed] [Google Scholar]
- 139.Tortosa-Caparrós E., Navas-Carrillo D., Marín F., Orenes-Piñero E. Anti-inflammatory effects of omega 3 and omega 6 polyunsaturated fatty acids in cardiovascular disease and metabolic syndrome. Crit. Rev. Food Sci. Nutr. 2017;57:3421–3429. doi: 10.1080/10408398.2015.1126549. [DOI] [PubMed] [Google Scholar]
- 140.Simopoulos A.P. An increase in the Omega-6/Omega-3 fatty acid ratio increases the risk for obesity. Nutrients. 2016;8:128. doi: 10.3390/nu8030128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Simopoulos A.P. Evolutionary aspects of diet, the omega-6/omega-3 ratio and genetic variation: Nutritional implications for chronic diseases. Biomed. Pharmacother. 2006;60:502–507. doi: 10.1016/j.biopha.2006.07.080. [DOI] [PubMed] [Google Scholar]
- 142.Lionetti L., Mollica M.P., Donizzetti I., Gifuni G., Sica R., Pignalosa A., Cavaliere G., Gaita M., De Filippo C., Zorzano A., et al. High-lard and high-fish-oil diets differ in their effects on function and dynamic behaviour of rat hepatic mitochondria. PLoS ONE. 2014;9:e92753. doi: 10.1371/journal.pone.0092753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Flachs P., Horakova O., Brauner P., Rossmeisl M., Pecina P., Franssen-Van Hal N., Ruzickova J., Sponarova J., Drahota Z., Vlcek C., et al. Polyunsaturated fatty acids of marine origin upregulate mitochondrial biogenesis and induce β-oxidation in white fat. Diabetologia. 2005;48:2365–2375. doi: 10.1007/s00125-005-1944-7. [DOI] [PubMed] [Google Scholar]
- 144.O’Mahoney L.L., Matu J., Price O.J., Birch K.M., Ajjan R.A., Farrar D., Tapp R., West D.J., Deighton K., Campbell M.D. Omega-3 polyunsaturated fatty acids favourably modulate cardiometabolic biomarkers in type 2 diabetes: A meta-analysis and meta-regression of randomized controlled trials. Cardiovasc. Diabetol. 2018;17:98. doi: 10.1186/s12933-018-0740-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Jang H., Park K. Omega-3 and omega-6 polyunsaturated fatty acids and metabolic syndrome: A systematic review and meta-analysis. Clin. Nutr. 2020;39:765–773. doi: 10.1016/j.clnu.2019.03.032. [DOI] [PubMed] [Google Scholar]
- 146.Liu R., Chen L., Wang Y., Zhang G., Cheng Y., Feng Z., Bai X., Liu J. High ratio of ω-3/ω-6 polyunsaturated fatty acids targets mTORC1 to prevent high-fat diet-induced metabolic syndrome and mitochondrial dysfunction in mice. J. Nutr. Biochem. 2020;9:e92753. doi: 10.1016/j.jnutbio.2019.108330. [DOI] [PubMed] [Google Scholar]
- 147.Wesselink E., Koekkoek W.A.C., Grefte S., Witkamp R.F., van Zanten A.R.H. Feeding mitochondria: Potential role of nutritional components to improve critical illness convalescence. Clin. Nutr. 2019;38:982–995. doi: 10.1016/j.clnu.2018.08.032. [DOI] [PubMed] [Google Scholar]
- 148.Manzetti S., Zhang J., van der Spoel D. Thiamin function, metabolism, uptake, and transport. Biochemistry. 2014;53:821–835. doi: 10.1021/bi401618y. [DOI] [PubMed] [Google Scholar]
- 149.Ashoori M., Saedisomeolia A. Riboflavin (vitamin B2) and oxidative stress: A review. Br. J. Nutr. 2014;111:1985–1991. doi: 10.1017/S0007114514000178. [DOI] [PubMed] [Google Scholar]
- 150.Depeint F., Bruce W.R., Shangari N., Mehta R., O’Brien P.J. Mitochondrial function and toxicity: Role of the B vitamin family on mitochondrial energy metabolism. Chem. Biol. Interact. 2006;163:94–112. doi: 10.1016/j.cbi.2006.04.014. [DOI] [PubMed] [Google Scholar]
- 151.Wojtczak L., Slyshenkov V.S. Protection by Pantothenic Acid Against Apoptosis and Cell Damage by Oxygen Free Radicals—The Role Of Glutathione. Volume 17. IOS Press; Amsterdam, The Netherlands: 2003. pp. 61–73. [DOI] [PubMed] [Google Scholar]
- 152.Roh T., De U., Lim S.K., Kim M.K., Choi S.M., Lim D.S., Yoon S., Kacew S., Kim H.S., Lee B.M. Detoxifying effect of pyridoxine on acetaminophen-induced hepatotoxicity via suppressing oxidative stress injury. Food Chem. Toxicol. 2018;114:11–22. doi: 10.1016/j.fct.2018.02.017. [DOI] [PubMed] [Google Scholar]
- 153.Moretti R., Peinkhofer C. B Vitamins and fatty acids: What do they share with small vessel disease-Related dementia? Int. J. Mol. Sci. 2019;20:5797. doi: 10.3390/ijms20225797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Fan J., Ye J., Kamphorst J.J., Shlomi T., Thompson C.B., Rabinowitz J.D. Quantitative flux analysis reveals folate-dependent NADPH production. Nature. 2014;510:298–302. doi: 10.1038/nature13236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Pastore A., Martinelli D., Piemonte F., Tozzi G., Boenzi S., Di Giovamberardino G., Petrillo S., Bertini E., Dionisi-Vici C. Glutathione metabolism in cobalamin deficiency type C (cblC) J. Inherit. Metab. Dis. 2014;37:125–129. doi: 10.1007/s10545-013-9605-3. [DOI] [PubMed] [Google Scholar]
- 156.Peng W., Xu S., Zhang J., Zhang Y. Vitamin C attenuates sodium fluoride-induced mitochondrial oxidative stress and apoptosis via Sirt1-SOD2 pathway in F9 cells. Biol. Trace Elem. Res. 2019;191:189–198. doi: 10.1007/s12011-018-1599-0. [DOI] [PubMed] [Google Scholar]
- 157.KC S., Càrcamo J.M., Golde D.W. Vitamin C enters mitochondria via facilitative glucose transporter 1 (Gluti) and confers mitochondrial protection against oxidative injury. FASEB J. 2005;19:1657–1667. doi: 10.1096/fj.05-4107com. [DOI] [PubMed] [Google Scholar]
- 158.Everts H.B., Claassen D.O., Hermoyian C.L., Berdanier C.D. Nutrient-gene interactions: Dietary vitamin A and mitochondrial gene expression. IUBMB Life. 2002;53:295–301. doi: 10.1080/15216540213465. [DOI] [PubMed] [Google Scholar]
- 159.Hammerling U. Vitamin A as PKC co-factor and regulator of mitochondrial energetics. Subcell. Biochem. 2016;81:201–230. doi: 10.1007/978-94-024-0945-1_8. [DOI] [PubMed] [Google Scholar]
- 160.Acin-Perez R., Hoyos B., Zhao F., Vinogradov V., Fischman D.A., Harris R.A., Leitges M., Wongsiriroj N., Blaner W.S., Manfredi G., et al. Control of oxidative phosphorylation by vitamin A illuminates a fundamental role in mitochondrial energy homoeostasis. FASEB J. 2010;24:627–636. doi: 10.1096/fj.09-142281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Silvagno F., Pescarmona G. Spotlight on vitamin D receptor, lipid metabolism and mitochondria: Some preliminary emerging issues. Mol. Cell. Endocrinol. 2017;450:24–31. doi: 10.1016/j.mce.2017.04.013. [DOI] [PubMed] [Google Scholar]
- 162.Ricca C., Aillon A., Bergandi L., Alotto D., Castagnoli C., Silvagno F. Vitamin D receptor is necessary for mitochondrial function and cell health. Int. J. Mol. Sci. 2018;19:1672. doi: 10.3390/ijms19061672. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Siddiqui S., Ahsan H., Khan M.R., Siddiqui W.A. Protective effects of tocotrienols against lipid-induced nephropathy in experimental type-2 diabetic rats by modulation in TGF-β expression. Toxicol. Appl. Pharmacol. 2013;273:314–324. doi: 10.1016/j.taap.2013.09.004. [DOI] [PubMed] [Google Scholar]
- 164.Zang Q.S., Sadek H., Maass D.L., Martinez B., Ma L., Kilgore J.A., Williams N.S., Frantz D.E., Wigginton J.G., Nwariaku F.E., et al. Specific inhibition of mitochondrial oxidative stress suppresses inflammation and improves cardiac function in a rat pneumonia-related sepsis model. Am. J. Physiol. Hear. Circ. Physiol. 2012;302:H1847–H1859. doi: 10.1152/ajpheart.00203.2011. [DOI] [PubMed] [Google Scholar]
- 165.Godala M., Materek-Kusmierkiewicz I., Moczulski D., Rutkowski M., Szatko F., Gaszyñska E., Tokarski S., Kowalski J. The risk of plasma Vitamin A, C, E and D deficiency in patients with metabolic syndrome: A case-control study. Adv. Clin. Exp. Med. 2017;26:581–586. doi: 10.17219/acem/62453. [DOI] [PubMed] [Google Scholar]
- 166.Prashanth A., Jeyakumar S.M., Singotamu L., Harishankar N., Giridharan N.V., Vajreswari A. Mitochondriogenesis and apoptosis: Possible cause of vitamin A-mediated adipose loss in WNIN/Ob-obese rats. Nutr. Metab. 2014;11:45. doi: 10.1186/1743-7075-11-45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Fernandez-Mejia C. Pharmacological effects of biotin. J. Nutr. Biochem. 2005;16:424–427. doi: 10.1016/j.jnutbio.2005.03.018. [DOI] [PubMed] [Google Scholar]
- 168.Kanagasabai T., Alkhalaqi K., Churilla J.R., Ardern C.I. The Association between metabolic syndrome and serum concentrations of micronutrients, inflammation, and oxidative stress outside of the clinical reference ranges: A cross-sectional study. Metab. Syndr. Relat. Disord. 2019;17:29–36. doi: 10.1089/met.2018.0080. [DOI] [PubMed] [Google Scholar]
- 169.Wei J., Zeng C., Gong Q.Y., Li X.X., Lei G.H., Yang T.B. Associations between dietary antioxidant intake and metabolic syndrome. PLoS ONE. 2015;10:e0130876. doi: 10.1371/journal.pone.0130876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Kim J., Choi Y.H. Physical activity, dietary vitamin C, and metabolic syndrome in the Korean adults: The Korea national health and nutrition examination survey 2008 to 2012. Public Health. 2016;135:30–37. doi: 10.1016/j.puhe.2016.01.002. [DOI] [PubMed] [Google Scholar]
- 171.Farag H.A.M., Hosseinzadeh-Attar M.J., Muhammad B.A., Esmaillzadeh A., Bilbeisi A.H. El Comparative effects of vitamin D and vitamin C supplementations with and without endurance physical activity on metabolic syndrome patients: A randomized controlled trial. Diabetol. Metab. Syndr. 2018;10:80. doi: 10.1186/s13098-018-0384-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Prasad P., Kochhar A. Interplay of Vitamin D and metabolic syndrome: A review. Diabetes Metab. Syndr. Clin. Res. Rev. 2016;10:105–112. doi: 10.1016/j.dsx.2015.02.014. [DOI] [PubMed] [Google Scholar]
- 173.Wimalawansa S.J. Associations of vitamin D with insulin resistance, obesity, type 2 diabetes, and metabolic syndrome. J. Steroid Biochem. Mol. Biol. 2018;175:177–189. doi: 10.1016/j.jsbmb.2016.09.017. [DOI] [PubMed] [Google Scholar]
- 174.Heng K.S., Hejar A.R., Johnson Stanslas J., Ooi C.F., Loh S.F. Potential of mixed tocotrienol supplementation to reduce cholesterol and cytokines level in adults with metabolic syndrome. Malays. J. Nutr. 2015;21:231–243. [Google Scholar]
- 175.Wong S.K., Chin K.Y., Suhaimi F.H., Ahmad F., Ima-Nirwana S. Vitamin E as a potential interventional treatment for metabolic syndrome: Evidence from animal and human studies. Front. Pharmacol. 2017;8:444. doi: 10.3389/fphar.2017.00444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Goncalves A., Amiot M.J. Fat-soluble micronutrients and metabolic syndrome. Curr. Opin. Clin. Nutr. Metab. Care. 2017;20:492–497. doi: 10.1097/MCO.0000000000000412. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Institute of Medicine (US) Dietary Reference Intakes: Vitamin A, Vitamin K, Arsenic, Boron, Chromium, Copper, Iodine, Iron, Manganese, Molybdenum, Nickel, Silicon, Vanadium, and Zinc. The National Academies Press; Washington, DC, USA: 2001. [DOI] [PubMed] [Google Scholar]
- 178.De Oliveira M.R. Vitamin A and retinoids as mitochondrial toxicants. Oxid. Med. Cell. Longev. 2015;2015:140267. doi: 10.1155/2015/140267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179.McCarty M.F. Co-administration of equimolar doses of betaine may alleviate the hepatotoxic risk associated with niacin therapy. Med. Hypotheses. 2000;55:189–194. doi: 10.1054/mehy.1999.1011. [DOI] [PubMed] [Google Scholar]
- 180.Chitturi R., Baddam V.R., Prasad L., Prashanth L., Kattapagari K. A review on role of essential trace elements in health and disease. J. Dr. NTR Univ. Heal. Sci. 2015;4:75. doi: 10.4103/2277-8632.158577. [DOI] [Google Scholar]
- 181.Alehagen U., Johansson P., Aaseth J., Alexander J., Wågsäter D. Significant changes in circulating microRNA by dietary supplementation of selenium and coenzyme Q10 in healthy elderly males. A subgroup analysis of a prospective randomized double-blind placebo-controlled trial among elderly Swedish citizens. PLoS ONE. 2017;12:e0174880. doi: 10.1371/journal.pone.0174880. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 182.Koekkoek W.A.C., van Zanten A.R.H. Antioxidant vitamins and trace elements in critical illness. Nutr. Clin. Pract. 2016;31:457–474. doi: 10.1177/0884533616653832. [DOI] [PubMed] [Google Scholar]
- 183.Mueller A.S., Pallauf J. Compendium of the antidiabetic effects of supranutritional selenate doses. In vivo and in vitro investigations with type II diabetic db/db mice. J. Nutr. Biochem. 2006 doi: 10.1016/j.jnutbio.2005.10.006. [DOI] [PubMed] [Google Scholar]
- 184.Zhou J., Huang K., Lei X.G. Selenium and diabetes-Evidence from animal studies. Free Radic. Biol. Med. 2013;65:1548–1556. doi: 10.1016/j.freeradbiomed.2013.07.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 185.Bahmani F., Kia M., Soleimani A., Mohammadi A.A., Asemi Z. The effects of selenium supplementation on biomarkers of inflammation and oxidative stress in patients with diabetic nephropathy: A randomised, double-blind, placebo-controlled trial. Br. J. Nutr. 2016;116:1222–1228. doi: 10.1017/S0007114516003251. [DOI] [PubMed] [Google Scholar]
- 186.Lymbury R.S., Marino M.J., Perkins A.V. Effect of dietary selenium on the progression of heart failure in the ageing spontaneously hypertensive rat. Mol. Nutr. Food Res. 2010;54:1436–1444. doi: 10.1002/mnfr.201000012. [DOI] [PubMed] [Google Scholar]
- 187.Gunes S., Sahinturk V., Karasati P., Sahin I.K., Ayhanci A. Cardioprotective effect of selenium against Cyclophosphamide-induced cardiotoxicity in rats. Biol. Trace Elem. Res. 2017;177:107–114. doi: 10.1007/s12011-016-0858-1. [DOI] [PubMed] [Google Scholar]
- 188.Chaâbane M., Tir M., Hamdi S., Boudawara O., Jamoussi K., Boudawara T., Ghorbel R.E., Zeghal N., Soudani N. Improvement of heart redox states contributes to the beneficial effects of selenium against penconazole-induced cardiotoxicity in adult rats. Biol. Trace Elem. Res. 2016;169:261–270. doi: 10.1007/s12011-015-0426-0. [DOI] [PubMed] [Google Scholar]
- 189.Ruz M., Carrasco F., Rojas P., Basfi-fer K., Hernández M.C., Pérez A. Nutritional effects of zinc on metabolic syndrome and type 2 diabetes: Mechanisms and main findings in human studies. Biol. Trace Elem. Res. 2019;188:177–188. doi: 10.1007/s12011-018-1611-8. [DOI] [PubMed] [Google Scholar]
- 190.Wang X., Zhang W., Chen H., Liao N., Wang Z., Zhang X., Hai C. High selenium impairs hepatic insulin sensitivity through opposite regulation of ROS. Toxicol. Lett. 2014;224:16–23. doi: 10.1016/j.toxlet.2013.10.005. [DOI] [PubMed] [Google Scholar]
- 191.Laclaustra M., Navas-Acien A., Stranges S., Ordovas J.M., Guallar E. Serum selenium concentrations and hypertension in the US population. Circ. Cardiovasc. Qual. Outcomes. 2009;2:369–376. doi: 10.1161/CIRCOUTCOMES.108.831552. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192.Berthold H.K., Michalke B., Krone W., Guallar E., Gouni-Berthold I. Influence of serum selenium concentrations on hypertension: The lipid analytic cologne cross-sectional study. J. Hypertens. 2012;30:1328–1335. doi: 10.1097/HJH.0b013e32835414df. [DOI] [PubMed] [Google Scholar]
- 193.Joseph S.V., Edirisinghe I., Burton-Freeman B.M. Fruit polyphenols: A review of anti-inflammatory effects in humans. Crit. Rev. Food Sci. Nutr. 2016;56:419–444. doi: 10.1080/10408398.2013.767221. [DOI] [PubMed] [Google Scholar]
- 194.Chiva-Blanch G., Badimon L. Effects of polyphenol intake on metabolic syndrome: Current evidences from human trials. Oxid. Med. Cell. Longvity. 2017 doi: 10.1155/2017/5812401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 195.Arola-Arnal A., Bladé C. Proanthocyanidins modulate microRNA expression in human HepG2 cells. PLoS ONE. 2011;6 doi: 10.1371/journal.pone.0025982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 196.Bernatoniene J., Kopustinskiene D.M. The role of catechins in cellular responses to oxidative stress. Molecules. 2018;23:965. doi: 10.3390/molecules23040965. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 197.NIH Office of Dietary Supplements National Institutes of Health (NIH) [(accessed on 20 June 2020)]; Available online: https://ods.od.nih.gov/factsheets.
- 198.Hasani M., Djalalinia S., Sharifi F., Varmaghani M., Zarei M., Abdar M.E., Asayesh H., Noroozi M., Kasaeian A., Gorabi A.M., et al. Effect of selenium supplementation on lipid profile: A systematic review and meta-analysis. Horm. Metab. Res. 2018;50:715–727. doi: 10.1055/a-0749-6655. [DOI] [PubMed] [Google Scholar]
- 199.Negishi H., Xu J.W., Ikeda K., Njelekela M., Nara Y., Yamori Y. Black and green tea Polyphenols attenuate blood pressure increases in stroke-prone spontaneously hypertensive rats. J. Nutr. 2004;134:38–42. doi: 10.1093/jn/134.1.38. [DOI] [PubMed] [Google Scholar]
- 200.Onakpoya I., Spencer E., Heneghan C., Thompson M. The effect of green tea on blood pressure and lipid profile: A systematic review and meta-analysis of randomized clinical trials. Nutr. Metab. Cardiovasc. Dis. 2014;24:823–836. doi: 10.1016/j.numecd.2014.01.016. [DOI] [PubMed] [Google Scholar]
- 201.Basu A., Du M., Leyva M.J., Sanchez K., Betts N.M., Wu M., Aston C.E., Lyons T.J. Blueberries decrease cardiovascular risk factors in obese men and women with metabolic syndrome. J. Nutr. 2010;140:1582–1587. doi: 10.3945/jn.110.124701. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 202.Yousaf S., Butt M.S., Suleria H.A.R., Iqbal M.J. The role of green tea extract and powder in mitigating metabolic syndromes with special reference to hyperglycemia and hypercholesterolemia. Food Funct. 2014;5:545–556. doi: 10.1039/c3fo60203f. [DOI] [PubMed] [Google Scholar]
- 203.Tsuneki H., Ishizuka M., Terasawa M., Wu J.B., Sasaoka T., Kimura I. Effect of green tea on blood glucose levels and serum proteomic patterns in diabetic (db/db) mice and on glucose metabolism in healthy humans. BMC Pharmacol. 2004;4:18. doi: 10.1186/1471-2210-4-18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 204.Finicelli M., Squillaro T., di Cristo F., di Salle A., Melone M.A.B., Galderisi U., Peluso G. Metabolic syndrome, Mediterranean diet, and polyphenols: Evidence and perspectives. J. Cell. Physiol. 2019;234:5807–5826. doi: 10.1002/jcp.27506. [DOI] [PubMed] [Google Scholar]
- 205.Asgary S., Karimi R., Momtaz S., Naseri R., Farzaei M.H. Effect of resveratrol on metabolic syndrome components: A systematic review and meta-analysis. Rev. Endocr. Metab. Disord. 2019;20:173–186. doi: 10.1007/s11154-019-09494-z. [DOI] [PubMed] [Google Scholar]
- 206.Mousavi S.M., Milajerdi A., Sheikhi A., Kord-Varkaneh H., Feinle-Bisset C., Larijani B., Esmaillzadeh A. Resveratrol supplementation significantly influences obesity measures: A systematic review and dose–response meta-analysis of randomized controlled trials. Obes. Rev. 2019;20:487–498. doi: 10.1111/obr.12775. [DOI] [PubMed] [Google Scholar]
- 207.Madreiter-Sokolowski C.T., Sokolowski A.A., Graier W.F. Dosis facit sanitatem—Concentration-dependent effects of resveratrol on mitochondria. Nutrients. 2017;9:1117. doi: 10.3390/nu9101117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 208.Sulaiman M., Matta M.J., Sunderesan N.R., Gupta M.P., Periasamy M., Gupta M. Resveratrol, an activator of SIRT1, upregulates sarcoplasmic calcium ATPase and improves cardiac function in diabetic cardiomyopathy. Am. J. Physiol. Hear. Circ. Physiol. 2010;298:H833–H843. doi: 10.1152/ajpheart.00418.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 209.Saibandith B., Spencer J.P.E., Rowland I.R., Commane D.M. Olive polyphenols and the metabolic syndrome. Molecules. 2017;22:1082. doi: 10.3390/molecules22071082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 210.Engelhard Y.N., Gazer B., Paran E. Natural antioxidants from tomato extract reduce blood pressure in patients with grade-1 hypertension: A double-blind, placebo-controlled pilot study. Am. Heart J. 2006;151:100. doi: 10.1016/j.ahj.2005.05.008. [DOI] [PubMed] [Google Scholar]
- 211.Han G.M., Liu P. Higher serum lycopene is associated with reduced prevalence of hypertension in overweight or obese adults. Eur. J. Integr. Med. 2017;13:34–40. doi: 10.1016/j.eujim.2017.07.002. [DOI] [Google Scholar]
- 212.Guerendiain M., Mayneris-Perxachs J., Montes R., López-Belmonte G., Martín-Matillas M., Castellote A.I., Martín-Bautista E., Martí A., Martínez J.A., Moreno L., et al. Relation between plasma antioxidant vitamin levels, adiposity and cardio-metabolic profile in adolescents: Effects of a multidisciplinary obesity programme. Clin. Nutr. 2017;36:209–217. doi: 10.1016/j.clnu.2015.11.001. [DOI] [PubMed] [Google Scholar]
- 213.Luvizotto R.A.M., Nascimento A.F., Miranda N.C.M., Wang X.D., Ferreira A.L.A. Lycopene-rich tomato oleoresin modulates plasma adiponectin concentration and mRNA levels of adiponectin, SIRT1, and FoxO1 in adipose tissue of obese rats. Hum. Exp. Toxicol. 2015;34:612–619. doi: 10.1177/0960327114551395. [DOI] [PubMed] [Google Scholar]
- 214.Jiang W., Guo M.H., Hai X. Hepatoprotective and antioxidant effects of lycopene on non-alcoholic fatty liver disease in rat. World J. Gastroenterol. 2016;22:10180–10188. doi: 10.3748/wjg.v22.i46.10180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 215.Zidani S., Benakmoum A., Ammouche A., Benali Y., Bouhadef A., Abbeddou S. Effect of dry tomato peel supplementation on glucose tolerance, insulin resistance, and hepatic markers in mice fed high-saturated-fat/high-cholesterol diets. J. Nutr. Biochem. 2017;40:164–171. doi: 10.1016/j.jnutbio.2016.11.001. [DOI] [PubMed] [Google Scholar]
- 216.Coyne T., Ibiebele T.I., Baade P.D., Dobson A., McClintock C., Dunn S., Leonard D., Shaw J. Diabetes mellitus and serum carotenoids: Findings of a population-based study in Queensland, Australia. Am. J. Clin. Nutr. 2005;82:685–693. doi: 10.1093/ajcn/82.3.685. [DOI] [PubMed] [Google Scholar]
- 217.Zeng Z., He W., Jia Z., Hao S. Lycopene improves insulin sensitivity through inhibition of STAT3/Srebp-1c-Mediated lipid accumulation and inflammation in mice fed a high-fat diet. Exp. Clin. Endocrinol. Diabetes. 2017;125:610–617. doi: 10.1055/s-0043-101919. [DOI] [PubMed] [Google Scholar]
- 218.Lee H.S., Lim W.C., Lee S.J., Lee S.H., Lee J.H., Cho H.Y. Antiobesity effect of garlic extract fermented by Lactobacillus plantarum BL2 in diet-induced obese mice. J. Med. Food. 2016;19:823–829. doi: 10.1089/jmf.2016.3674. [DOI] [PubMed] [Google Scholar]
- 219.Zhang Y., Xu L., Ding M., Su G., Zhao Y. Anti-obesity effect of garlic oil on obese rats via Shenque point administration. J. Ethnopharmacol. 2019;231:486–493. doi: 10.1016/j.jep.2018.11.030. [DOI] [PubMed] [Google Scholar]
- 220.Kim E.J., Lee D.H., Kim H.J., Lee S.J., Ban J.O., Cho M.C., Jeong H.S., Yang Y., Hong J.T., Yoon D.Y., et al. Thiacremonone, a sulfur compound isolated from garlic, attenuates lipid accumulation partially mediated via AMPK activation in 3T3-L1 adipocytes. J. Nutr. Biochem. 2012;23:1552–1558. doi: 10.1016/j.jnutbio.2011.10.008. [DOI] [PubMed] [Google Scholar]
- 221.Lee C.G., Rhee D.K., Kim B.O., Um S.H., Pyo S. Allicin induces beige-like adipocytes via KLF15 signal cascade. J. Nutr. Biochem. 2019;64:13–24. doi: 10.1016/j.jnutbio.2018.09.014. [DOI] [PubMed] [Google Scholar]
- 222.Choudhary P.R., Jani R.D., Sharma M.S. Effect of raw crushed garlic (Allium sativum L.) on components of metabolic syndrome. J. Diet. Suppl. 2018;15:499–506. doi: 10.1080/19390211.2017.1358233. [DOI] [PubMed] [Google Scholar]
- 223.Ried K., Travica N., Sali A. The Effect of kyolic aged garlic extract on gut microbiota, inflammation, and cardiovascular markers in hypertensives: The GarGIC trial. Front. Nutr. 2018;5:122. doi: 10.3389/fnut.2018.00122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 224.Sultana M.R., Bagul P.K., Katare P.B., Anwar Mohammed S., Padiya R., Banerjee S.K. Garlic activates SIRT-3 to prevent cardiac oxidative stress and mitochondrial dysfunction in diabetes. Life Sci. 2016;164:42–51. doi: 10.1016/j.lfs.2016.08.030. [DOI] [PubMed] [Google Scholar]
- 225.Supakul L., Pintana H., Apaijai N., Chattipakorn S., Shinlapawittayatorn K., Chattipakorn N. Protective effects of garlic extract on cardiac function, heart rate variability, and cardiac mitochondria in obese insulin-resistant rats. Eur. J. Nutr. 2014;53:919–928. doi: 10.1007/s00394-013-0595-6. [DOI] [PubMed] [Google Scholar]
- 226.Matsumoto S., Nakanishi R., Li D., Alani A., Rezaeian P., Prabhu S., Abraham J., Fahmy M.A., Dailing C., Flores F., et al. Aged garlic extract reduces low attenuation plaque in coronary arteries of patients with metabolic syndrome in a prospective randomized double-blind study. J. Nutr. 2016;146:427S–732S. doi: 10.3945/jn.114.202424. [DOI] [PubMed] [Google Scholar]
- 227.Zeb I., Ahmadi N., Flores F., Budoff M.J. Randomized trial evaluating the effect of aged garlic extract with supplements versus placebo on adipose tissue surrogates for coronary atherosclerosis progression. Coron. Artery Dis. 2018;29:325–328. doi: 10.1097/MCA.0000000000000587. [DOI] [PubMed] [Google Scholar]
- 228.Toti E., Oliver Chen C.Y., Palmery M., Valencia D.V., Peluso I. Non-provitamin A and provitamin A carotenoids as immunomodulators: Recommended dietary allowance, therapeutic index, or personalized nutrition? Oxid. Med. Cell. Longev. 2018;2018:4637861. doi: 10.1155/2018/4637861. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 229.Vang O., Ahmad N., Baile C.A., Baur J.A., Brown K., Csiszar A., Das D.K., Delmas D., Gottfried C., Lin H.Y., et al. What is new for an old molecule? systematic review and recommendations on the use of resveratrol. PLoS ONE. 2011;6:e19881. doi: 10.1371/journal.pone.0019881. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 230.Lawson L.D., Hunsaker S.M. Allicin bioavailability and bioequivalence from garlic supplements and garlic foods. Nutrients. 2018;10:812. doi: 10.3390/nu10070812. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 231.Food/FDA. [(accessed on 5 August 2020)]; Available online: https://www.fda.gov/food/
- 232.Hu J., Webster D., Cao J., Shao A. The safety of green tea and green tea extract consumption in adults–Results of a systematic review. Regul. Toxicol. Pharmacol. 2018;95:412–433. doi: 10.1016/j.yrtph.2018.03.019. [DOI] [PubMed] [Google Scholar]
- 233.Diaz-Gerevini G.T., Repossi G., Dain A., Tarres M.C., Das U.N., Eynard A.R. Beneficial action of resveratrol: How and why? Nutrition. 2016;32:174–178. doi: 10.1016/j.nut.2015.08.017. [DOI] [PubMed] [Google Scholar]
- 234.De Oliveira M.R., Nabavi S.F., Manayi A., Daglia M., Hajheydari Z., Nabavi S.M. Resveratrol and the mitochondria: From triggering the intrinsic apoptotic pathway to inducing mitochondrial biogenesis, a mechanistic view. Biochim. Biophys. Acta Gen. Subj. 2016;1860:727–745. doi: 10.1016/j.bbagen.2016.01.017. [DOI] [PubMed] [Google Scholar]
- 235.Bigagli E., Cinci L., Paccosi S., Parenti A., D’Ambrosio M., Luceri C. Nutritionally relevant concentrations of resveratrol and hydroxytyrosol mitigate oxidative burst of human granulocytes and monocytes and the production of pro-inflammatory mediators in LPS-stimulated RAW 264.7 macrophages. Int. Immunopharmacol. 2017;43:147–155. doi: 10.1016/j.intimp.2016.12.012. [DOI] [PubMed] [Google Scholar]
- 236.Tomé-Carneiro J., Larrosa M., Yáñez-Gascón M.J., Dávalos A., Gil-Zamorano J., Gonzálvez M., García-Almagro F.J., Ruiz Ros J.A., Tomás-Barberán F.A., Espín J.C., et al. One-year supplementation with a grape extract containing resveratrol modulates inflammatory-related microRNAs and cytokines expression in peripheral blood mononuclear cells of type 2 diabetes and hypertensive patients with coronary artery disease. Pharmacol. Res. 2013;72:69–82. doi: 10.1016/j.phrs.2013.03.011. [DOI] [PubMed] [Google Scholar]
- 237.Saini R.K., Zamany A.J., Keum Y.S. Ripening improves the content of carotenoid, α-tocopherol, and polyunsaturated fatty acids in tomato (Solanum lycopersicum L.) fruits. 3 Biotech. 2017;7:43. doi: 10.1007/s13205-017-0666-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 238.Mishra S., Chattopadhyay A., Naaz S., Ghosh A.K., Das A.R., Bandyopadhyay D. Oleic acid ameliorates adrenaline induced dysfunction of rat heart mitochondria by binding with adrenaline: An isothermal titration calorimetry study. Life Sci. 2019;218:96–111. doi: 10.1016/j.lfs.2018.12.035. [DOI] [PubMed] [Google Scholar]
- 239.Debbabi M., Nury T., Zarrouk A., Mekahli N., Bezine M., Sghaier R., Grégoire S., Martine L., Durand P., Camus E., et al. Protective effects of α-tocopherol, γ-tocopherol and oleic acid, three compounds of olive oils, and no effect of trolox, on 7-ketocholesterol-induced mitochondrial and peroxisomal dysfunction in microglial BV-2 cells. Int. J. Mol. Sci. 2016;17:1973. doi: 10.3390/ijms17121973. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 240.Kong K.W., Khoo H.E., Prasad K.N., Ismail A., Tan C.P., Rajab N.F. Revealing the power of the natural red pigment lycopene. Molecules. 2010;15:959–987. doi: 10.3390/molecules15020959. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 241.Devaraj S., Mathur S., Basu A., Meyers S., Jialal I., Aung H.H., Vasu V.T. A Dose-response study on the effects of purified lycopene supplementation on biomarkers of oxidative stress. J. Am. Coll. Nutr. 2008;27:267–273. doi: 10.1080/07315724.2008.10719699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 242.Matos H.R., Marques S.A., Gomes O.F., Silva A.A., Heimann J.C., Di Mascio P., Medeiros M.H.G. Lycopene and β-carotene protect in vivo iron-induced oxidative stress damage in rat prostate. Braz. J. Med. Biol. Res. 2006;39:203–210. doi: 10.1590/S0100-879X2006000200006. [DOI] [PubMed] [Google Scholar]
- 243.Duthie S.J., Ma A., Ross M.A., Collins A.R. Antioxidant supplementation decreases oxidative DNA damage in human lymphocytes. Cancer Res. 1996;56:1291–1295. [PubMed] [Google Scholar]
- 244.Zhao B., Ren B., Guo R., Zhang W., Ma S., Yao Y., Yuan T., Liu Z., Liu X. Supplementation of lycopene attenuates oxidative stress induced neuroinflammation and cognitive impairment via Nrf2/NF-κB transcriptional pathway. Food Chem. Toxicol. 2017;109:505–516. doi: 10.1016/j.fct.2017.09.050. [DOI] [PubMed] [Google Scholar]
- 245.Li W., Jiang B., Cao X., Xie Y., Huang T. Protective effect of lycopene on fluoride-induced ameloblasts apoptosis and dental fluorosis through oxidative stress-mediated caspase pathways. Chem. Biol. Interact. 2017;261:27–34. doi: 10.1016/j.cbi.2016.11.021. [DOI] [PubMed] [Google Scholar]
- 246.Agarwal S., Rao A.V. Tomato lycopene and low density lipoprotein oxidation: A human dietary intervention study. Lipids. 1998;33:981–984. doi: 10.1007/s11745-998-0295-6. [DOI] [PubMed] [Google Scholar]
- 247.Lomb D.J., Laurent G., Haigis M.C. Sirtuins regulate key aspects of lipid metabolism. Biochim. Biophys. Acta Proteins Proteomics. 2010;1804:1652–1657. doi: 10.1016/j.bbapap.2009.11.021. [DOI] [PubMed] [Google Scholar]
- 248.Chiva-Blanch G., Jiménez C., Pinyol M., Herreras Z., Catalán M., Martínez-Huélamo M., Lamuela-Raventos R.M., Sala-Vila A., Cofán M., Gilabert R., et al. 5-CIS-, trans-and total lycopene plasma concentrations inversely relate to atherosclerotic plaque burden in newly diagnosed type 2 diabetes subjects. Nutrients. 2020;12:1696. doi: 10.3390/nu12061696. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 249.Ghavipour M., Sotoudeh G., Ghorbani M. Tomato juice consumption improves blood antioxidative biomarkers in overweight and obese females. Clin. Nutr. 2015;34:805–809. doi: 10.1016/j.clnu.2014.10.012. [DOI] [PubMed] [Google Scholar]
- 250.Han G.M., Meza J.L., Soliman G.A., Islam K.M.M., Watanabe-Galloway S. Higher levels of serum lycopene are associated with reduced mortality in individuals with metabolic syndrome. Nutr. Res. 2016;36:402–407. doi: 10.1016/j.nutres.2016.01.003. [DOI] [PubMed] [Google Scholar]
- 251.Quesada I., de Paola M., Torres-Palazzolo C., Camargo A., Ferder L., Manucha W., Castro C. Effect of Garlic’s Active Constituents in Inflammation, Obesity and Cardiovascular Disease. Curr. Hypertens. Rep. 2020;22:6. doi: 10.1007/s11906-019-1009-9. [DOI] [PubMed] [Google Scholar]
- 252.Wang S., Ren D. Allicin protects traumatic spinal cord injury through regulating the HSP70/Akt/iNOS pathway in mice. Mol. Med. Rep. 2016;14:3086–3092. doi: 10.3892/mmr.2016.5651. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 253.Lee I.C., Kim S.H., Baek H.S., Moon C., Kang S.S., Kim S.H., Kim Y.B., Shin I.S., Kim J.C. The involvement of Nrf2 in the protective effects of diallyl disulfide on carbon tetrachloride-induced hepatic oxidative damage and inflammatory response in rats. Food Chem. Toxicol. 2014;63:174–185. doi: 10.1016/j.fct.2013.11.006. [DOI] [PubMed] [Google Scholar]
- 254.Shang A., Cao S.Y., Xu X.Y., Gan R.Y., Tang G.Y., Corke H., Mavumengwana V., Li H. Bin Bioactive compounds and biological functions of garlic (Allium sativum L.) Foods. 2019;8:246. doi: 10.3390/foods8070246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 255.Shin I.S., Hong J., Jeon C.M., Shin N.R., Kwon O.K., Kim H.S., Kim J.C., Oh S.R., Ahn K.S. Diallyl-disulfide, an organosulfur compound of garlic, attenuates airway inflammation via activation of the Nrf-2/HO-1 pathway and NF-kappaB suppression. Food Chem. Toxicol. 2013;62:506–513. doi: 10.1016/j.fct.2013.09.012. [DOI] [PubMed] [Google Scholar]
- 256.Ried K., Travica N., Sali A. The effect of aged garlic extract on blood pressure and other cardiovascular risk factors in uncontrolled hypertensives: The AGE at Heart trial. Integr. Blood Press. Control. 2016;9:9–21. doi: 10.2147/IBPC.S93335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 257.Hall A.B., Tolonen A.C., Xavier R.J. Human genetic variation and the gut microbiome in disease. Nat. Rev. Genet. 2017;18:690–699. doi: 10.1038/nrg.2017.63. [DOI] [PubMed] [Google Scholar]
- 258.Cani P.D., Amar J., Iglesias M.A., Poggi M., Knauf C., Bastelica D., Neyrinck A.M., Fava F., Tuohy K.M., Chabo C., et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes. 2007;56:1761–1772. doi: 10.2337/db06-1491. [DOI] [PubMed] [Google Scholar]
- 259.Xiao H.B., Sun Z.L., Zhang H.B., Zhang D.S. Berberine inhibits dyslipidemia in C57BL/6 mice with lipopolysaccharide induced inflammation. Pharmacol. Rep. 2012;64:889–895. doi: 10.1016/S1734-1140(12)70883-6. [DOI] [PubMed] [Google Scholar]
- 260.Lassenius M.I., Pietiläinen K.H., Kaartinen K., Pussinen P.J., Syrjänen J., Forsblom C., Pörsti I., Rissanen A., Kaprio J., Mustonen J., et al. Bacterial endotoxin activity in human serum is associated with dyslipidemia, insulin resistance, obesity, and chronic inflammation. Diabetes Care. 2011;34:1809–1815. doi: 10.2337/dc10-2197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 261.David L.A., Maurice C.F., Carmody R.N., Gootenberg D.B., Button J.E., Wolfe B.E., Ling A.V., Devlin A.S., Varma Y., Fischbach M.A., et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature. 2014;505:559–563. doi: 10.1038/nature12820. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 262.Dabke K., Hendrick G., Devkota S. The gut microbiome and metabolic syndrome. J. Clin. Investig. 2019;129:4050–4057. doi: 10.1172/JCI129194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 263.Wan Y., Wang F., Yuan J., Li J., Jiang D., Zhang J., Li H., Wang R., Tang J., Huang T., et al. Effects of dietary fat on gut microbiota and faecal metabolites, and their relationship with cardiometabolic risk factors: A 6-month randomised controlled-feeding trial. Gut. 2019;68:1417–1429. doi: 10.1136/gutjnl-2018-317609. [DOI] [PubMed] [Google Scholar]
- 264.Sonnenburg E.D., Smits S.A., Tikhonov M., Higginbottom S.K., Wingreen N.S., Sonnenburg J.L. Diet-induced extinctions in the gut microbiota compound over generations. Nature. 2016;529:212–215. doi: 10.1038/nature16504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 265.Desai M.S., Seekatz A.M., Koropatkin N.M., Kamada N., Hickey C.A., Wolter M., Pudlo N.A., Kitamoto S., Terrapon N., Muller A., et al. A dietary fiber-deprived gut microbiota degrades the colonic mucus barrier and enhances pathogen susceptibility. Cell. 2016;167:1339–1353. doi: 10.1016/j.cell.2016.10.043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 266.Chassaing B., van de Wiele T., de Bodt J., Marzorati M., Gewirtz A.T. Dietary emulsifiers directly alter human microbiota composition and gene expression ex vivo potentiating intestinal inflammation. Gut. 2017;66:1414–1427. doi: 10.1136/gutjnl-2016-313099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 267.Zhang X., Shen D., Fang Z., Jie Z., Qiu X., Zhang C., Chen Y., Ji L. Human gut microbiota changes reveal the progression of glucose intolerance. PLoS ONE. 2013;8:e71108. doi: 10.1371/journal.pone.0071108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 268.Li J., Zhao F., Wang Y., Chen J., Tao J., Tian G., Wu S., Liu W., Cui Q., Geng B., et al. Gut microbiota dysbiosis contributes to the development of hypertension. Microbiome. 2017;5:14. doi: 10.1186/s40168-016-0222-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 269.Yassour M., Lim M.Y., Yun H.S., Tickle T.L., Sung J., Song Y.M., Lee K., Franzosa E.A., Morgan X.C., Gevers D., et al. Sub-clinical detection of gut microbial biomarkers of obesity and type 2 diabetes. Genome Med. 2016;8:17. doi: 10.1186/s13073-016-0271-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 270.Moreno-Indias I., Sánchez-Alcoholado L., Pérez-Martínez P., Andrés-Lacueva C., Cardona F., Tinahones F., Queipo-Ortuño M.I. Red wine polyphenols modulate fecal microbiota and reduce markers of the metabolic syndrome in obese patients. Food Funct. 2016;7:1775–1787. doi: 10.1039/C5FO00886G. [DOI] [PubMed] [Google Scholar]
- 271.Mathewson N.D., Jenq R., Mathew A.V., Koenigsknecht M., Hanash A., Toubai T., Oravecz-Wilson K., Wu S.R., Sun Y., Rossi C., et al. Gut microbiome-derived metabolites modulate intestinal epithelial cell damage and mitigate graft-versus-host disease. Nat. Immunol. 2016;17:505–513. doi: 10.1038/ni.3400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 272.Dillon S.M., Kibbie J., Lee E.J., Guo K., Santiago M.L., Austin G.L., Gianella S., Landay A.L., Donovan A.M., Frank D.N., et al. Low abundance of colonic butyrate-producing bacteria in HIV infection is associated with microbial translocation and immune activation. AIDS. 2017;31:511–521. doi: 10.1097/QAD.0000000000001366. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 273.De Vadder F., Kovatcheva-Datchary P., Zitoun C., Duchampt A., Bäckhed F., Mithieux G. Microbiota-Produced Succinate Improves Glucose Homeostasis via Intestinal Gluconeogenesis. Cell Metab. 2016;24:151–157. doi: 10.1016/j.cmet.2016.06.013. [DOI] [PubMed] [Google Scholar]
- 274.Perry R.J., Peng L., Barry N.A., Cline G.W., Zhang D., Cardone R.L., Petersen K.F., Kibbey R.G., Goodman A.L., Shulman G.I., et al. Acetate mediates a microbiome-brain-β-cell axis to promote metabolic syndrome. Nature. 2016;534:213–217. doi: 10.1038/nature18309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 275.Magnúsdóttir S., Ravcheev D., de Crécy-Lagard V., Thiele I. Systematic genome assessment of B-vitamin biosynthesis suggests cooperation among gut microbes. Front. Genet. 2015;6 doi: 10.3389/fgene.2015.00148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 276.Roberfroid M., Gibson G.R., Hoyles L., McCartney A.L., Rastall R., Rowland I., Wolvers D., Watzl B., Szajewska H., Stahl B., et al. Prebiotic effects: Metabolic and health benefits. Br. J. Nutr. 2010;104:S1–S63. doi: 10.1017/S0007114510003363. [DOI] [PubMed] [Google Scholar]
- 277.Food and Agriculture Organization. World Health Organization . Probiotics in Food Health and Nutritional Properties and Guidelines for Evaluation. FAO; Rome, Italy: 2006. [Google Scholar]
- 278.Parnell J.A., Reimer R.A. Prebiotic fibres dose-dependently increase satiety hormones and alter Bacteroidetes and Firmicutes in lean and obese JCR:LA-cp rats. Br. J. Nutr. 2012;107:601–613. doi: 10.1017/S0007114511003163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 279.Cano P.G., Santacruz A., Trejo F.M., Sanz Y. Bifidobacterium CECT 7765 improves metabolic and immunological alterations associated with obesity in high-fat diet-fed mice. Obesity. 2013;21:2310–2321. doi: 10.1002/oby.20330. [DOI] [PubMed] [Google Scholar]
- 280.Tomaro-Duchesneau C., Saha S., Malhotra M., Jones M.L., Labbé A., Rodes L., Kahouli I., Prakash S. Effect of orally administered L. fermentum NCIMB 5221 on markers of metabolic syndrome: An in vivo analysis using ZDF rats. Appl. Microbiol. Biotechnol. 2014;98:115–126. doi: 10.1007/s00253-013-5252-8. [DOI] [PubMed] [Google Scholar]
- 281.Dewulf E.M., Cani P.D., Claus S.P., Fuentes S., Puylaert P.G.B., Neyrinck A.M., Bindels L.B., de Vos W.M., Gibson G.R., Thissen J.P., et al. Insight into the prebiotic concept: Lessons from an exploratory, double blind intervention study with inulin-type fructans in obese women. Gut. 2013;62:1112–1121. doi: 10.1136/gutjnl-2012-303304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 282.Barreto F.M., Colado Simão A.N., Morimoto H.K., Batisti Lozovoy M.A., Dichi I., Helena da Silva Miglioranza L. Beneficial effects of Lactobacillus plantarum on glycemia and homocysteine levels in postmenopausal women with metabolic syndrome. Nutrition. 2014;30:939–942. doi: 10.1016/j.nut.2013.12.004. [DOI] [PubMed] [Google Scholar]
- 283.Kadooka Y., Sato M., Ogawa A., Miyoshi M., Uenishi H., Ogawa H., Ikuyama K., Kagoshima M., Tsuchida T. Effect of Lactobacillus gasseri SBT2055 in fermented milk on abdominal adiposity in adults in a randomised controlled trial. Br. J. Nutr. 2013;110:1696–1703. doi: 10.1017/S0007114513001037. [DOI] [PubMed] [Google Scholar]
- 284.Gilijamse P.W., Hartstra A.V., Levin E., Wortelboer K., Serlie M.J., Ackermans M.T., Herrema H., Nederveen A.J., Imangaliyev S., Aalvink S., et al. Treatment with Anaerobutyricum soehngenii: A pilot study of safety and dose–response effects on glucose metabolism in human subjects with metabolic syndrome. NPJ Biofilms Microbiomes. 2020;6 doi: 10.1038/s41522-020-0127-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 285.Andreasen A.S., Larsen N., Pedersen-Skovsgaard T., Berg R.M.G., Mller K., Svendsen K.D., Jakobsen M., Pedersen B.K. Effects of Lactobacillus acidophilus NCFM on insulin sensitivity and the systemic inflammatory response in human subjects. Br. J. Nutr. 2010;104:1831–1838. doi: 10.1017/S0007114510002874. [DOI] [PubMed] [Google Scholar]
- 286.Kobyliak N., Falalyeyeva T., Mykhalchyshyn G., Kyriienko D., Komissarenko I. Effect of alive probiotic on insulin resistance in type 2 diabetes patients: Randomized clinical trial. Diabetes Metab. Syndr. Clin. Res. Rev. 2018;12:617–624. doi: 10.1016/j.dsx.2018.04.015. [DOI] [PubMed] [Google Scholar]
- 287.Peterson C.T., Sharma V., Elmén L., Peterson S.N. Immune homeostasis, dysbiosis and therapeutic modulation of the gut microbiota. Clin. Exp. Immunol. 2015;179:363–377. doi: 10.1111/cei.12474. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 288.Smits L.P., Bouter K.E.C., De Vos W.M., Borody T.J., Nieuwdorp M. Therapeutic potential of fecal microbiota transplantation. Gastroenterology. 2013;145:946–953. doi: 10.1053/j.gastro.2013.08.058. [DOI] [PubMed] [Google Scholar]
- 289.Aron-Wisnewsky J., Clément K., Nieuwdorp M. Fecal microbiota transplantation: A future therapeutic option for obesity/diabetes? Curr. Diab. Rep. 2019;19 doi: 10.1007/s11892-019-1180-z. [DOI] [PubMed] [Google Scholar]
- 290.Menotti A., Puddu P.E. How the Seven Countries Study contributed to the definition and development of the Mediterranean diet concept: A 50-year journey. Nutr. Metab. Cardiovasc. Dis. 2015;25:245–252. doi: 10.1016/j.numecd.2014.12.001. [DOI] [PubMed] [Google Scholar]
- 291.Rodríguez-Monforte M., Sánchez E., Barrio F., Costa B., Flores-Mateo G. Metabolic syndrome and dietary patterns: A systematic review and meta-analysis of observational studies. Eur. J. Nutr. 2017;56:925–947. doi: 10.1007/s00394-016-1305-y. [DOI] [PubMed] [Google Scholar]
- 292.EFSA Scientific opinion on dietary reference values for water. EFSA J. 2016;8 doi: 10.2903/j.efsa.2010.1459. [DOI] [Google Scholar]
- 293.Popkin B.M., D’Anci K.E., Rosenberg I.H. Water, hydration, and health. Nutr. Rev. 2010;68:439–458. doi: 10.1111/j.1753-4887.2010.00304.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 294.Allen M.D., Springer D.A., Burg M.B., Boehm M., Dmitrieva N.I. Suboptimal hydration remodels metabolism, promotes degenerative diseases, and shortens life. JCI Insight. 2019;4:e130949. doi: 10.1172/jci.insight.130949. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 295.Burge M.R., Garcia N., Qualls C.R., Schade D.S. Differential effects of fasting and dehydration in the pathogenesis of diabetic ketoacidosis. Metabolism. 2001;50:171–177. doi: 10.1053/meta.2001.20194. [DOI] [PubMed] [Google Scholar]
- 296.Sun K., Liu J., Ning G. Active smoking and risk of metabolic syndrome: A meta-analysis of prospective studies. PLoS ONE. 2012;7:e47791. doi: 10.1371/journal.pone.0047791. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 297.Fujita N., Takei Y. Alcohol consumption and metabolic syndrome. Hepatol. Res. 2011;41:287–295. doi: 10.1111/j.1872-034X.2011.00787.x. [DOI] [PubMed] [Google Scholar]
- 298.Mortality and Burden of Disease Attributable to Selected Major Risks. World Health Organization; Geneva, Switzerland: 2009. [Google Scholar]
- 299.Janiszewski P.M., Ross R. The utility of physical activity in the management of global cardiometabolic risk. Obesity. 2009;17:S3–S14. doi: 10.1038/oby.2009.382. [DOI] [PubMed] [Google Scholar]
- 300.Warburton D.E.R., Nicol C.W., Bredin S.S.D. Health benefits of physical activity: The evidence. CMAJ. 2006;174:801–809. doi: 10.1503/cmaj.051351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 301.Després J.-P., Lemieux I., Bergeron J., Pibarot P., Mathieu P., Larose E., Rodés-Cabau J., Bertrand O.F., Poirier P. Abdominal obesity and the metabolic syndrome: Contribution to global cardiometabolic risk. Arterioscler. Thromb. Vasc. Biol. 2008;28:1039–1049. doi: 10.1161/ATVBAHA.107.159228. [DOI] [PubMed] [Google Scholar]
- 302.Brioche T., Lemoine-Morel S. Oxidative stress, sarcopenia, antioxidant strategies and exercise: Molecular aspects. Curr. Pharm. Des. 2016;22:2664–2678. doi: 10.2174/1381612822666160219120531. [DOI] [PubMed] [Google Scholar]
- 303.Bouzid M.A., Filaire E., McCall A., Fabre C. Radical oxygen species, exercise and aging: An update. Sports Med. 2015;45:1245–1261. doi: 10.1007/s40279-015-0348-1. [DOI] [PubMed] [Google Scholar]
- 304.Zhang L., Zhang Z., Khan A., Zheng H., Yuan C., Jiang H. Advances in drug therapy for mitochondrial diseases. Ann. Transl. Med. 2020;8:17. doi: 10.21037/atm.2019.10.113. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 305.Veech R.L. The therapeutic implications of ketone bodies: The effects of ketone bodies in pathological conditions: Ketosis, ketogenic diet, redox states, insulin resistance, and mitochondrial metabolism. Prostaglandins Leukot. Essent. Fat. Acids. 2004;70:309–319. doi: 10.1016/j.plefa.2003.09.007. [DOI] [PubMed] [Google Scholar]
- 306.McDonald T.J.W., Cervenka M.C. Ketogenic diets for adults with highly refractory epilepsy. Epilepsy Curr. 2017;17:346–350. doi: 10.5698/1535-7597.17.6.346. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 307.Hughes S.D., Kanabus M., Anderson G., Hargreaves I.P., Rutherford T., Donnell M.O., Cross J.H., Rahman S., Eaton S., Heales S.J.R. The ketogenic diet component decanoic acid increases mitochondrial citrate synthase and complex I activity in neuronal cells. J. Neurochem. 2014;129:426–433. doi: 10.1111/jnc.12646. [DOI] [PubMed] [Google Scholar]
- 308.Giordano C., Marchiò M., Timofeeva E., Biagini G. Neuroactive peptides as putative mediators of antiepileptic ketogenic diets. Front. Neurol. 2014;5:63. doi: 10.3389/fneur.2014.00063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 309.Bárcena C., Mayoral P., Quirós P.M. Mitohormesis, an antiaging paradigm. Int. Rev. Cell Mol. Biol. 2018;340:35–77. doi: 10.1016/bs.ircmb.2018.05.002. [DOI] [PubMed] [Google Scholar]
- 310.Ristow M., Zarse K. How increased oxidative stress promotes longevity and metabolic health: The concept of mitochondrial hormesis (mitohormesis) Exp. Gerontol. 2010;45:410–418. doi: 10.1016/j.exger.2010.03.014. [DOI] [PubMed] [Google Scholar]
- 311.Cheng Z., Ristow M. Mitochondria and metabolic homeostasis. Antioxid. Redox Signal. 2013;19:240–242. doi: 10.1089/ars.2013.5255. [DOI] [PubMed] [Google Scholar]
- 312.Hemingway C., Freeman J.M., Pillas D.J., Pyzik P.L. The ketogenic diet: A 3- to 6-year follow-up of 150 children enrolled prospectively. Pediatrics. 2001;108:898–905. doi: 10.1542/peds.108.4.898. [DOI] [PubMed] [Google Scholar]
- 313.Neal E.G., Cross J.H. Efficacy of dietary treatments for epilepsy. J. Hum. Nutr. Diet. 2010;23:113–119. doi: 10.1111/j.1365-277X.2010.01043.x. [DOI] [PubMed] [Google Scholar]
- 314.Danial N.N., Hartman A.L., Stafstrom C.E., Thio L.L. How does the ketogenic diet work? Four potential mechanisms. J. Child Neurol. 2013;28:1027–1033. doi: 10.1177/0883073813487598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 315.Patel A., Pyzik P.L., Turner Z., Rubenstein J.E., Kossoff E.H. Long-term outcomes of children treated with the ketogenic diet in the past. Epilepsia. 2010;51:1277–1282. doi: 10.1111/j.1528-1167.2009.02488.x. [DOI] [PubMed] [Google Scholar]
- 316.De Giorgis V., Veggiotti P. GLUT1 deficiency syndrome 2013: Current state of the art. Seizure. 2013;22:803–811. doi: 10.1016/j.seizure.2013.07.003. [DOI] [PubMed] [Google Scholar]
- 317.Schoeler N.E., Simpson Z., Whiteley V.J., Nguyen P., Meskell R., Lightfoot K., Martin-McGill K.J., Olpin S., Ivison F. Biochemical assessment of patients following ketogenic diets for epilepsy: Current practice in the UK and Ireland. Epilepsia Open. 2020;5:73–79. doi: 10.1002/epi4.12371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 318.Taivassalo T., Shoubridge E.A., Chen J., Kennaway N.G., Dimauro S., Arnold D.L., Haller R.G. Aerobic conditioning in patients with mitochondrial myopathies: Physiological, biochemical, and genetic effects. Ann. Neurol. 2001;50:133–141. doi: 10.1002/ana.1050. [DOI] [PubMed] [Google Scholar]
- 319.Kang C., Li Ji L. Role of PGC-1α signaling in skeletal muscle health and disease. Ann. N. Y. Acad. Sci. 2012;1271:110–117. doi: 10.1111/j.1749-6632.2012.06738.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 320.Garone C., Donati M.A., Sacchini M., Garcia-Diaz B., Bruno C., Calvo S., Mootha V.K., DiMauro S. Mitochondrial encephalomyopathy due to a novel mutation in ACAD9. JAMA Neurol. 2013;70:1177–1179. doi: 10.1001/jamaneurol.2013.3197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 321.Haack T.B., Danhauser K., Haberberger B., Hoser J., Strecker V., Boehm D., Uziel G., Lamantea E., Invernizzi F., Poulton J., et al. Exome sequencing identifies ACAD9 mutations as a cause of complex i deficiency. Nat. Genet. 2010;42:1131–1134. doi: 10.1038/ng.706. [DOI] [PubMed] [Google Scholar]
- 322.Olsen R.K.J., Olpin S.E., Andresen B.S., Miedzybrodzka Z.H., Pourfarzam M., Merinero B., Frerman F.E., Beresford M.W., Dean J.C.S., Cornelius N., et al. ETFDH mutations as a major cause of riboflavin-responsive multiple acyl-CoA dehydrogenation deficiency. Brain. 2007;130:2045–2054. doi: 10.1093/brain/awm135. [DOI] [PubMed] [Google Scholar]
- 323.Bugiani M., Lamantea E., Invernizzi F., Moroni I., Bizzi A., Zeviani M., Uziel G. Effects of riboflavin in children with complex II deficiency. Brain Dev. 2006;28:576–581. doi: 10.1016/j.braindev.2006.04.001. [DOI] [PubMed] [Google Scholar]
- 324.Sato Y., Nakagawa M., Higuchi I., Osame M., Naito E., Oizumi K. Mitochondrial myopathy and familial thiamine deficiency. Muscle Nerve. 2000;23:1069–1075. doi: 10.1002/1097-4598(200007)23:7<1069::AID-MUS9>3.0.CO;2-0. [DOI] [PubMed] [Google Scholar]
- 325.Mermigkis C., Bouloukaki I., Mastorodemos V., Plaitakis A., Alogdianakis V., Siafakas N., Schiza S. Medical treatment with thiamine, coenzyme Q, vitamins e and C, and carnitine improved obstructive sleep apnea in an adult case of Leigh disease. Sleep Breath. 2013;17:1129–1135. doi: 10.1007/s11325-013-0816-5. [DOI] [PubMed] [Google Scholar]
- 326.Parikh S., Saneto R., Falk M.J., Anselm I., Cohen B.H., Haas R. A modern approach to the treatment of mitochondrial disease. Curr. Treat. Options Neurol. 2009;11:414–430. doi: 10.1007/s11940-009-0046-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 327.Kuszak A.J., Espey M.G., Falk M.J., Holmbeck M.A., Manfredi G., Shadel G.S., Vernon H.J., Zolkipli-Cunningham Z. Nutritional interventions for mitochondrial OXPHOS deficiencies: Mechanisms and model systems. Annu. Rev. Pathol. Mech. Dis. 2018;13:163–191. doi: 10.1146/annurev-pathol-020117-043644. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 328.Tarnopolsky M.A. Creatine as a therapeutic strategy for myopathies. Amino Acids. 2011;40:1397–1407. doi: 10.1007/s00726-011-0876-4. [DOI] [PubMed] [Google Scholar]
- 329.Raymond M.R., Christensen K.Y., Thompson B.A., Anderson H.A. Associations between fish consumption and contaminant biomarkers with cardiovascular conditions among older male anglers in Wisconsin. J. Occup. Environ. Med. 2016;58:676–682. doi: 10.1097/JOM.0000000000000757. [DOI] [PubMed] [Google Scholar]
- 330.Badimon L., Chagas P., Chiva-Blanch G. Diet and cardiovascular disease: Effects of foods and nutrients in classical and emerging cardiovascular risk factors. Curr. Med. Chem. 2017;26:3639–3651. doi: 10.2174/0929867324666170428103206. [DOI] [PubMed] [Google Scholar]