

Abstract
Decades of research suggest that the mesocortical dopamine system exerts powerful control over mPFC physiology and function. Indeed, dopamine signaling in the medial prefrontal cortex (mPFC) is implicated in a vast array of processes, including working memory, stimulus discrimination, stress responses, and emotional and behavioral control. Consequently, even slight perturbations within this delicate system result in profound disruptions of mPFC-mediated processes. Many neuropsychiatric disorders are associated with dysregulation of mesocortical dopamine, including schizophrenia, depression, attention deficit hyperactivity disorder, post-traumatic stress disorder, among others. Here, we review the anatomy and functions of the mesocortical dopamine system. In contrast to the canonical role of striatal dopamine in reward-related functions, recent work has revealed that mesocortical dopamine fine-tunes distinct efferent projection populations in a manner that biases subsequent behavior towards responding to stimuli associated with potentially aversive outcomes. We propose a framework wherein dopamine can serve as a signal for switching mPFC states by orchestrating how information is routed to the rest of the brain.
Keywords: Valence, Signal-to-noise ratio, Mesocortical, Aversion, Mesolimbic
1. Introduction to mesocortical dopamine
Dopamine is one of the most well-studied neurotransmitters in the brain and is primarily found in the nigrostriatal, tuberoinfundibular, mesolimbic, and mesocortical pathways (Björklund and Dunnett, 2007; Ikemoto, 2010). Mesencephalic, or midbrain, dopamine neurons innervate limbic and cortical areas through the mesolimbic and mesocortical pathways, respectively. A rich literature demonstrates the critical role of the mesolimbic pathway in reward-seeking, learning, and motoric function (Berridge and Robinson, 1998; Ikemoto, 2007; Wise, 2008). In contrast, the mesocortical dopamine system, which has been long-known to be a distinct and important population, has gained far less attention. Despite a relative paucity of investigations into mesocortical dopamine system function, studies consistently report that dysregulation of mesocortical dopamine underlies a wide array of pathologies found in neuropsychiatric disorders — including schizophrenia, depression, attention deficit hyperactivity disorder, and post-traumatic stress disorder (Arnsten and Dudley, 2005; Deutch and Young, 1995; Grace, 2016; Granon et al., 2000; Howes and Kapur, 2009; Lee et al., 2016; Lindström et al., 1999; Okubo et al., 1997; Pitman et al., 2012).
In preclinical literature, evidence supporting the idea that mesocortical dopamine is functionally distinct from mesolimbic dopamine dates back more than 40 years (Thierry et al., 1976). Early studies proposed the controversial theory that, in stark contrast to the well-documented role of mesolimbic dopamine in reward-related processes, mesocortical dopamine is primarily involved in encoding aversive events (Abercrombie et al., 1989; Mantz et al., 1989; Thierry et al., 1976). Over the last two decades, technical advances have greatly expanded our ability to assess the function of mesocortical dopamine circuits, and several groundbreaking studies definitively demonstrating a role of mesocortical dopamine in aversive processing have reinvigorated interest in this system (Gunaydin et al., 2014; Horvitz, 2000; Lammel et al., 2011, 2012).
Here, we review the evidence for mesocortical dopamine in aversion and highlight recent work that has begun to map out the complex actions of dopamine on diverse mPFC circuits. We outline a framework whereby mesocortical dopamine acts to amplify or dampen information streams within the mPFC to select the most appropriate information to be routed to downstream circuits.
2. Functions of the prefrontal cortex
To understand the activities of mesocortical dopamine, it is imperative to appreciate the functional and anatomical landscape in which it operates. The mPFC is a heterogeneous brain region implicated in a diverse range of cognitive and behavioral functions— including attention, decision-making, working memory, long-term memory, emotional control, inhibitory control, motivation, among many others (Bechara et al., 2000; Euston et al., 2012; Laubach et al., 2015; Miller, 1999; Ridderinkhof et al., 2004; Sawaguchi and Goldman-Rakic, 1991). While these functions may appear disparate, they all are related to cognitive control, or the ability to coordinate emotions and actions to support internally held goals (Miller, 2000; Miller and Cohen, 2001). Lesions of mPFC result in robust deficits in performance in tasks that require subjects to adapt their behavior across varying contexts, or tasks with dynamic or unexpected rules (Dias et al., 1996a, 1996b, 1997; Gehring and Knight, 2000; Gregoriou et al., 2014; Koenigs and Grafman, 2009; Milner, 1963; Rossi et al., 2007, 2009; Wilkinson et al., 1997). Thus, the mPFC appears to play a significant role in simultaneously inhibiting inappropriate behaviors, and promoting task- and contextually-relevant ones (Aron et al., 2004; Buschman and Miller, 2007; Dias et al., 1997; Thompson-Schill et al., 2002). Though many brain regions, neurotransmitter systems, and neural circuits support goal-directed behavior, the mPFC is anatomically well-positioned to act as a hub that coordinates sensory inputs with internal states and goals to support adaptive behavioral outputs.
Successfully seeking rewards and avoiding punishments is critical for survival, and a defining characteristic of adaptive behavior. For these behaviors to occur optimally, neural systems must integrate internal motivational states with external information to orchestrate approach and avoidance of rewarding and aversive stimuli, respectively. Here we use rewarding or aversive stimuli as umbrella terms for stimuli that typically act as reinforcers or punishments in operant or place conditioning tasks, though we acknowledge that the response to many of these stimuli are dependent on a wide array of environmental factors. For example, an animal engaging in foraging behavior for food must constantly evaluate its environment for potential threats, which necessitate a transition from reward-seeking to threat avoidance. While the mPFC is not critical for basic stimulus-driven unconditioned behaviors, which are mediated by subcortical structures like the periaqueductal gray (Assareh et al., 2016; Bandler and Depaulis, 1988; Deng et al., 2016; Tovote et al., 2016) and hypothalamus (Anand and Brobeck, 1951; Bains et al., 2015; Betley et al., 2015; Burton et al., 1976; Jennings et al., 2015; Lin et al., 2011; Nieh et al., 2015, 2016), it plays a substantial role in deciphering ambiguous situations. In the example above, the foraging animal must not only perceive the environmental threat, but also evaluate its importance in relation to its ongoing efforts (e.g., How far away is the threat? How severe is the threat? How hungry am I? When will I be able to forage again?). In other words, the mPFC integrates sensory information with the perceived value of that information (i.e., how “good” or “bad” a stimulus is) together with internal belief states to provide top-down management of motivated behaviors.
There is substantial evidence supporting a crucial role of the mPFC in decision-making in the context of risk and reward, particularly in situations involving competition of appetitive and aversive drives — when both rewarding and aversive stimuli are simultaneously present (Bechara et al., 1994, 2005; Botvinick et al., 2004; Burgos-Robles et al., 2017; Mansouri et al., 2009; Milham et al., 2001). In rodents, pharmacological inactivation of the mPFC produces deficits in the coordination of reward-seeking and threat avoidance (Sangha et al., 2014; Sierra-Mercado et al., 2011). In humans, mPFC activity is modulated by the proximity of threat (Mobbs et al., 2007) suggesting a role in coordinating behavioral coping strategies. Not surprisingly, mPFC neuronal activity is robustly modulated by motivationally-relevant stimuli; including reward-delivery, reward-expectation, omission of reward, pain, and pain-predictive cues (Euston et al., 2012). Further, mPFC activity is highly correlated with aspects of reward- and fear-motivated behavioral output (Burgos-Robles et al., 2009, 2013). Thus, the mPFC has been metaphorically compared to a “switch operator” for sending trains down different tracks (Miller and Cohen, 2001), but the rules determining where information is sent, and the mechanisms for parsing information are still being discovered.
3. Anatomy of the prefrontal cortex
Consistent with the idea that the mPFC is crucial for cognitive control, this cortical region is most developed in primates and animals that have elaborate behavioral repertoires (Adolphs, 2009; Dunbar, 2009; Kerney et al., 2017; Noonan et al., 2018). In the rodent, the mPFC is often subdivided into four subregions – the secondary motor area (M2), the anterior cingulate cortex (ACC), the prelimbic cortex (PL), and infralimbic cortex (IL) – based on connectivity, cytoarchitectural, and functional differences (Heidbreder and Groenewegen, 2003; Ongür and Price, 2000) (Figure 1). A functional gradient along the dorsal-ventral axis of the mPFC (M2 > ACC > PL > IL) has been proposed, where more dorsal components control behavioral action and the more ventral components regulate emotions (Euston et al., 2012; Groenewegen and Uylings, 2000). M2 is the most dorsal of the mPFC regions and influences downstream circuits through inputs to motor regions and the spinal cord (Donoghue and Wise, 1982; Gabbott et al., 2005). While there is debate regarding the inclusion of M2 as a member of the mPFC family (Laubach et al., 2018), this premotor region is thought to link the evaluation of sensory information to motor outcomes (Barthas and Kwan, 2017). Adjacent to and ventral of M2 is the ACC, which is implicated in the error detection and evaluation of negative outcomes (Botvinick et al., 2004; Johansen and Fields, 2004; Shackman et al., 2011). In line with these theories, the ACC is robustly activated in response to pain and pain-predictive stimuli (Allsop et al., 2018; Frankland et al., 2004; Vogt, 2005). Finally, the PL and IL areas comprise the ventral medial aspects of the mPFC and are strongly interconnected to both limbic and autonomic control centers (Gabbott et al., 2005; Heidbreder and Groenewegen, 2003; Vertes, 2004). Further, these regions appear to be crucial for the detection, monitoring, and updating of instrumental contingencies (Cardinal et al., 2002). Together, this collection of mPFC nuclei access motivationally-relevant and homeostatic information to guide adaptive, goal-directed behaviors via input to emotional, autonomic, and motor loci.
Figure 1.
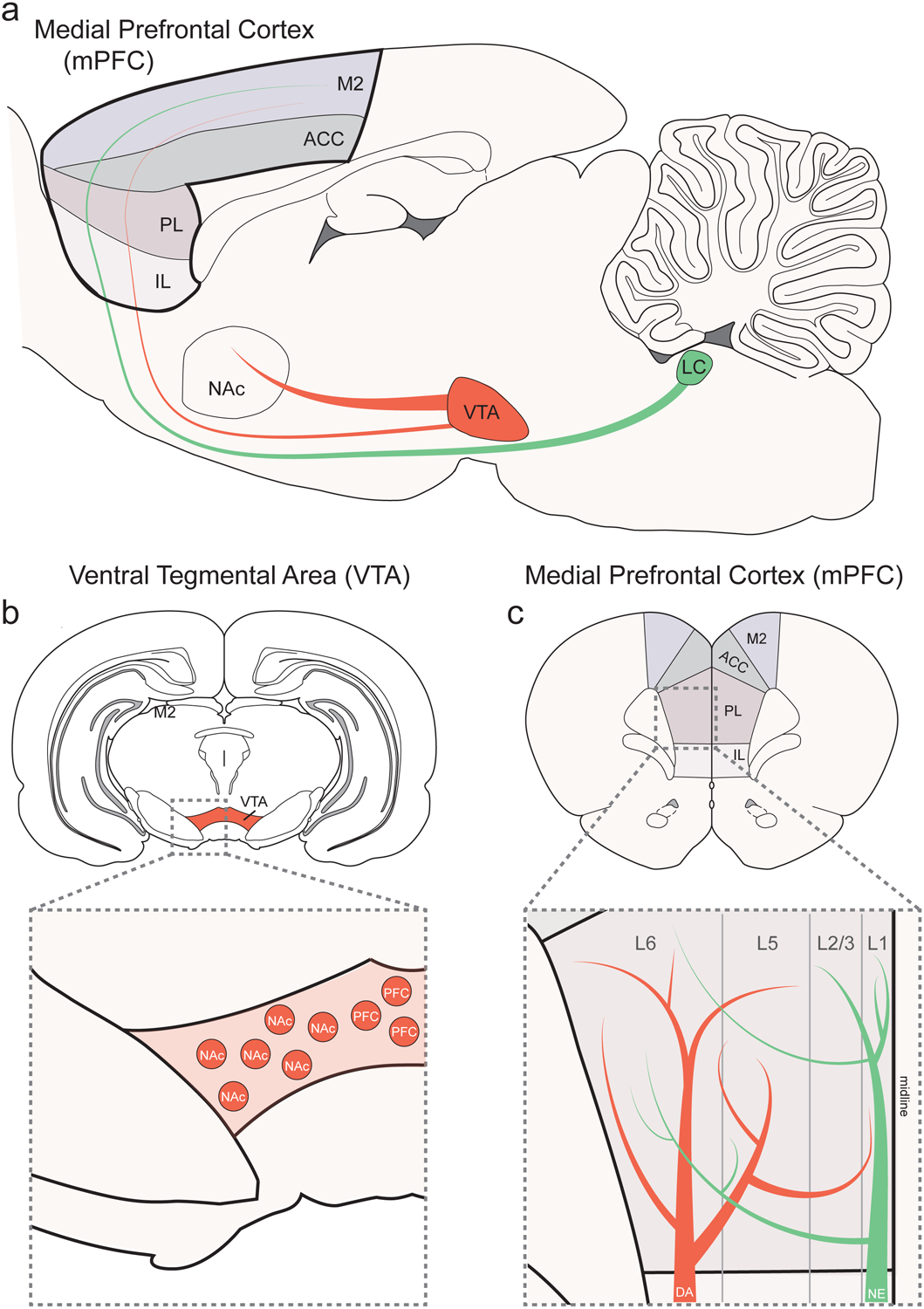
Catecholaminergic innervation of the corticolimbic system. a. Sagittal section of the rat brain showing catecholaminergic innervation of the medial prefrontal cortext (mPFC) and its four major subdivisions: secondary motor cortex (M2), anterior cingulate cortex (ACC), prelimbic cortex (PL), and infralimbic cortex (IL). Dopamine neurons originate from the ventral tegmental area (VTA) and project to the mPFC and ventral striatum / nucleus accumbens (NAc). Norepinephrine neurons in the locus coeruleus (LC) travel below the VTA to the mPFC an other corticolimbic regions. b. Coronal section of the rat brain showing the location of the VTA on an anterior-posterior axis. Inset shows the medial-lateral distribution of projection defined VTA dopamine neurons. Mesocortical dopamine neurons reside in the more medial aspects of the VTA and mesolimbic dopamine neurons reside in the more lateral aspects. c. Coronal section of the rat brain showing the mPFC and its major subregions on an anterior-posterior axis. Inset shows catecholaminergic innervation of mPFC layers. Dopaminergic axon terminals preferentially innervate the deep layers of the mPFC (Layers 5–6), while noreadrenergic axon terminals preferentially innervate the superficial layers (Layers 1–2/3). Abbreviations: DA = dopamine; NE = norepinephrine.
The mPFC consists of diverse cells types. The neuronal cell types are broadly split into excitatory, glutamatergic pyramidal cells, and inhibitory GABAergic interneurons, which represent ~80–90% and 10–20% of the population, respectively (Gabbott et al., 1997, 2005). Traditionally, it has been thought that pyramidal neurons comprise the entirety of the long-range projection neurons in the mPFC while GABAergic interneurons powerfully regulate their activity via dense local connectivity (Isaacson and Scanziani, 2011). However, recently long-range GABAergic projections targeting the nucleus accumbens (NAc) have also been described (Lee et al., 2014). Both pyramidal and interneuron populations contain many subclasses distinguished by their functional, laminar, morphological, electrophysiological, and molecular properties (Gabbott et al., 1997, 2005; Kvitsiani et al., 2013; Markram et al., 2004). Overall, the delicate balance of these excitatory and inhibitory forces (i.e., “E/I balance”) within the mPFC are crucial for optimal cognitive performance (Rubenstein and Merzenich, 2003; Yizhar et al., 2011).
Similar to other cortical structures, the mPFC is laminarly organized (Douglas and Martin, 2004; Gabbott et al., 2005; Kritzer and Goldman-Rakic, 1995). The mPFC receives diverse sensory and limbic afferent connections (Hoover and Vertes, 2007) arriving predominantly in superficial Layers 1 and 2/3 (Douglas and Martin, 2004; Romanski et al., 1999a, 1999b). mPFC efferent connections are equally robust, projecting to cortical and subcortical brain regions (Gabbott et al., 2005; Groenewegen et al., 1997; Heidbreder and Groenewegen, 2003; Sesack et al., 1989; Vertes, 2004). Long-range efferent cortical-subcortical projections originate mainly from the deep layers (Layer 5 and Layer 6), while cortical-cortical projections arise primarily from superficial Layer 2/3 (DeFelipe and Fariñas, 1992; Douglas and Martin, 2004; Gabbott et al., 2005). Notably, efferent connections appear to be densest to regions involved in emotional and autonomic control— including the amygdala, ventral striatum, insula, hypothalamus, periaqueductal gray, habenula, and midline thalamic structures (Gabbott et al., 2005; Groenewegen et al., 1997; Heidbreder and Groenewegen, 2003; Sesack et al., 1989; Vertes, 2004). Through this dense interconnectivity, the mPFC is thought to exert executive, top-down control over processing of rewarding and aversive events, as well as subsequent emotional regulation and control of actions. However precise, causal investigations into the potentially distinct functions of mPFC output populations are still lacking. It is unclear how mPFC parses incoming information, how this information is divided between outputs, and how these projections are influenced by dopamine.
4. Midbrain dopamine neurons
The mPFC has reciprocal connectivity with several neuromodulatory systems known to be involved in adaptive responses to rewarding and aversive events— including the midbrain dopamine system (Christie et al., 1985; Gabbott et al., 1997; Heidbreder and Groenewegen, 2003; Tye et al., 2013; Warden et al., 2012). The two largest dopaminergic nuclei are the ventral tegmental area (VTA, also known as A10) and substrantia nigra pars compacta (SNc, also known as A9) (Björklund and Dunnett, 2007), which are both anatomically and functionally distinct populations. While VTA dopamine neurons regulate motivation and encode reward expectation, substantia nigra neurons are more involved in motor functions via connections to the dorsal striatum / caudate putamen (Berridge and Robinson, 1998; Björklund and Dunnett, 2007; Michel et al., 2016; Phillips et al., 2003; York, 1973). In the rodent brain, VTA dopamine neurons send dense projections to the ventral striatum (including the NAc) and sparser projections to the mPFC, amygdala, and hippocampus (Haber and Fudge, 1997) (Figure 1a). However, it is important to note that there is considerable variation across mammalian species. For example, dopaminergic innervation of the cortex is much denser in primates (Bentivoglio and Morelli, 2005; Descarries et al., 1987; Lewis and Sesack, 1997; Thierry et al., 1973) and appears to be more homogenous across cortical layers (Berger et al., 1991; Williams and Goldman-Rakic, 1993; Vander Weele et al., 2018).
VTA dopamine neurons transmit signal in two modes: 1) “Tonic” mode is characterized by a consistent, pacemaker-like 2–5 Hz firing rate, and 2) “Phasic” mode is characterized transient bursts of high frequency 10–20+ Hz firing lasting 100–500 ms (Bunney et al., 1991; Grace and Bunney, 1984a, 1984b; Grace et al., 2007). The net effect of tonic firing/release, reuptake, diffusion and degradation mechanisms result in a relatively stable basal, extracellular level of dopamine, or tonic dopamine, in downstream targets (Cragg and Rice, 2004; Ferris et al., 2014). Phasic VTA dopamine neuron firing rapidly and transiently increases dopamine levels, estimated to reach levels of ~100 μM in striatal synapses (Garris and Wightman, 1994; Grace et al., 2007) (Garris and Wightman, 1994; Grace et al., 2007), termed ‘phasic dopamine release’ or dopamine ‘transients’ (Robinson et al., 2003). Both tonic dopamine level and phasic release modulate the activity of specific populations of neurons expressing postsynaptic dopamine receptors (see below for more detailed discussion) (Dreyer et al., 2010, 2016; Owesson-White et al., 2016). It is thought that the impact of phasic dopamine transients are determined by the size of the transient relative to the basal level of dopamine present in that microdomain (Wanat et al., 2009), making the interplay between tonic and phasic dopamine a critical determinant of the impact of dopamine cell firing. To make matters more complicated, action potential bursts at the dopamine cell bodies do not always translate to phasic release events downstream. Indeed, dopamine levels in striatum and cell firing of dopamine neurons in the VTA are often decoupled (Mohebi et al., 2018; Trulson, 1985), which is a result of the fact that dopamine terminal activity is heavily influenced by a number of mechanisms including input from local interneurons and a wide array of presynaptic modulatory receptors, such as dopamine autoreceptors (Cachope and Cheer, 2014; Cachope et al., 2012; Cragg and Rice, 2004; Dreyer and Hounsgaard, 2013; Zhang and Sulzer, 2004). As such, it is important to use multidisciplinary approaches to appreciate the complex and dynamic interactions between somatic activity, dopamine release, reuptake, and local modulatory mechanisms.
Dopamine release and reuptake dynamics and their postsynaptic actions have been studied in detail within the striatum, but the extent to which these principles translate to other brain regions which receive dopaminergic afferents, like the mPFC, is unclear. Certainly there are some significant differences. For example, in contrast to tight reuptake regulation in the striatum, dopamine neurotransmission in the mPFC is not as temporally controlled. In the striatum, the dopamine transporter (DAT) dictates the duration of dopamine’s actions in the synapse and the extent to which it diffuses (Cragg and Rice, 2004). In the mPFC, dopamine axons largely lack DAT immunoreactivity (Sesack et al., 1998) which results in reduced reuptake and greater extracellular diffusion distances (Cass and Gerhardt, 1995; Garris and Wightman, 1994; Garris et al., 1993; Vander Weele et al., 2018). Evidence suggests that dopamine clearance is mediated in part by the norepinephrine transporter (NET) (Carboni et al., 1990; Miner et al., 2003; Sesack et al., 1998; Tanda et al., 1994). However, NET immunoreactivity is sparse in the deep layers of the mPFC and recordings of dopamine neurotransmission show temporally prolonged signals indicative of reduced capacity for clearance (Garris and Wightman, 1994; Garris et al., 1993; Vander Weele et al., 2018). Thus dopamine release in the deep layers of the mPFC appears to be primarily regulated by extraneuronal metabolism (Karoum et al., 1994; Maisonneuve et al., 1990; Sharp et al., 1986), an enzymatic degradation process that is slower than the fast protein-mediated reuptake. Considering this, caution should be used when comparing dopamine dynamics and functions between the striatum and the mPFC as the results may not be generalizable.
Phasic dopamine neuron activity and phasic dopamine release have been studied extensively in the context of reward (Covey et al., 2014; Tsai et al., 2009; Wanat et al., 2009; Wickham et al., 2013), and to a lesser extent, aversion (Badrinarayan et al., 2012; Budygin et al., 2012; Wenzel et al., 2015). In a seminal study by Schultz and colleagues (1997), dopamine neurons were found to change their activity based on reward expectation: dopamine neurons increased their firing to unexpected reward delivery, but shifted this response to cues that predict reward delivery after pairing— which is reflected in ventral striatal phasic dopamine release (Day et al., 2007). VTA dopamine neurons also exhibited a decrease in firing if an expected reward (i.e., predicted by a conditioned stimulus) was omitted (Schultz et al., 1997). Further, some studies report inhibition of dopamine activity in response to aversive events or conditioned stimuli that predict them (Badrinarayan et al., 2012; Oleson et al., 2012; Roitman et al., 2008; Twining et al., 2015; Ungless et al., 2004; Wheeler et al., 2011). Finally, by optogenetically activating dopamine neurons at the time of reward receipt, motivational value can be attributed to neutral cues, even in the absence of error, demonstrating causality between dopamine neuron activation and Pavlovian conditioning of antecedent cues (Steinberg et al., 2013). Together, these neurons have been described as encoding “reward prediction error” (RPE) by signaling discrepancies between expected and actual outcomes. As such, RPEs provide a mechanism for learning the outcomes of predictive stimuli and updating their internal representations when the magnitude, valence, or timing of predicted outcomes differs from expectation.
RPE theory is largely based on observing the endogenous action potential activity of midbrain dopamine neurons, many of which project to the ventral striatum, and by observing dopamine transients in the ventral striatum. While the majority of dopamine neurons in the midbrain have activity profiles that appear consistent with RPE theory, there are also small subpopulations that are not consistent, such as cells that show phasic responses to aversive stimuli (Brischoux et al., 2009, 2009; Mantz et al., 1989; Matsumoto and Hikosaka, 2009; Matsumoto et al.). These findings have sparked considerable debate over dopamine’s role in processing of aversive events. As previously mentioned, several theories have suggested that phasic dopamine responses primarily encode reward-related events— including food reward, water reward, social targets, sex, and drugs of abuse (Covey et al., 2014; Robinson et al., 2003). However, dopamine responses to various salient and aversive stimuli — including surprising events, stress, pain, and fear-predictive cues have also been reported (Abercrombie et al., 1989; Badrinarayan et al., 2012; Budygin et al., 2012; Matsumoto and Hikosaka, 2009; Matsumoto et al.; McCutcheon et al., 2012; Salamone, 1994; Wenzel et al., 2015). As a result, new theories of dopamine function have suggested roles in alerting (Bromberg-Martin et al., 2010), invigoration of ongoing behaviors (Niv, 2007; Salamone et al., 2007; da Silva et al., 2018), or encoding the value of work (Berke, 2018; Hamid et al., 2016).
In addition to the VTA and SNc, there are several other dopaminergic nuclei scattered throughout the brain. One population of interest is dopamine neurons located in the dorsal raphe nucleus (DRN), an area traditionally known for being the primary source of serotonin in the brain, which recently have also been implicated in the processing of salient stimuli (Cho et al., 2017; Li et al., 2016; Matthews et al., 2016). Activation of DRN dopamine neurons promotes avoidance in real-time and conditioned place-preference assays suggesting that activity within this population is aversive (Matthews et al., 2016). DRN dopamine neurons send ascending projections primarily to the central nucleus of the amygdala (CeA) and the bed nucleus of the stria terminalis (BNST) (Li et al., 2016; Matthews et al., 2016). Additionally, functionally distinct populations within the VTA and substantia nigra continue to be discovered. For example, a recent set of studies identified a group of dopamine neurons projecting from the substantia nigra pars lateralis to a long-overlooked region in the caudal most extent of the striatum, called the tail of the striatum (Menegas et al., 2015). This population encodes the perceived threat of stimuli, and reinforces retreat behaviors (Menegas et al., 2015). The increasing evidence for heterogeneity within and between dopamine neuron populations in the processing of rewarding and aversive stimuli call for more precise interrogations into their cognitive and behavioral operations.
5. Characteristics of the mesocortical dopamine pathway
How is dopamine involved in so many seemingly disparate processes? Over the last two decades it has become increasingly clear that VTA dopamine neurons form subpopulations with heterogeneous functions, which are, at least in part, defined by their disparate projection targets (Bromberg-Martin et al., 2010; Kim et al., 2016a; Lammel et al., 2014). For example, VTA dopamine neurons that project to the NAc appear to be involved in different operations than those projecting to the mPFC, amygdala, and hippocampus. This notion of defining subpopulations of dopamine neurons by their anatomy (e.g., “projection-defined”) is supported by the finding that the majority of VTA dopamine neurons do not collateralize. Current evidence suggests that they send independent, non-overlapping ascending projections to target regions (Albanese and Minciacchi, 1983; Breton et al., 2018; Chandler et al., 2013; Fallon, 1981). Further, these projection-defined VTA dopamine subpopulations maintain several distinct features, including receptor expression (Bannon and Roth, 1983; Bannon et al., 1981; Lammel et al., 2008), electrophysiological properties (Chiodo et al., 1984; Lammel et al., 2008), anatomical location (Lammel et al., 2012) (Figure 1b), afferent connectivity (Lammel et al., 2012), and responses to stimuli with positive or negative valence (Abercrombie et al., 1989; Bassareo et al., 2002; Kim et al., 2016a; Lammel et al., 2011; Mantz et al., 1989). (Figure 2). Accordingly, growing evidence supports distinct behavioral roles of VTA dopamine subpopulations depending on their forebrain projection target.
Figure 2.
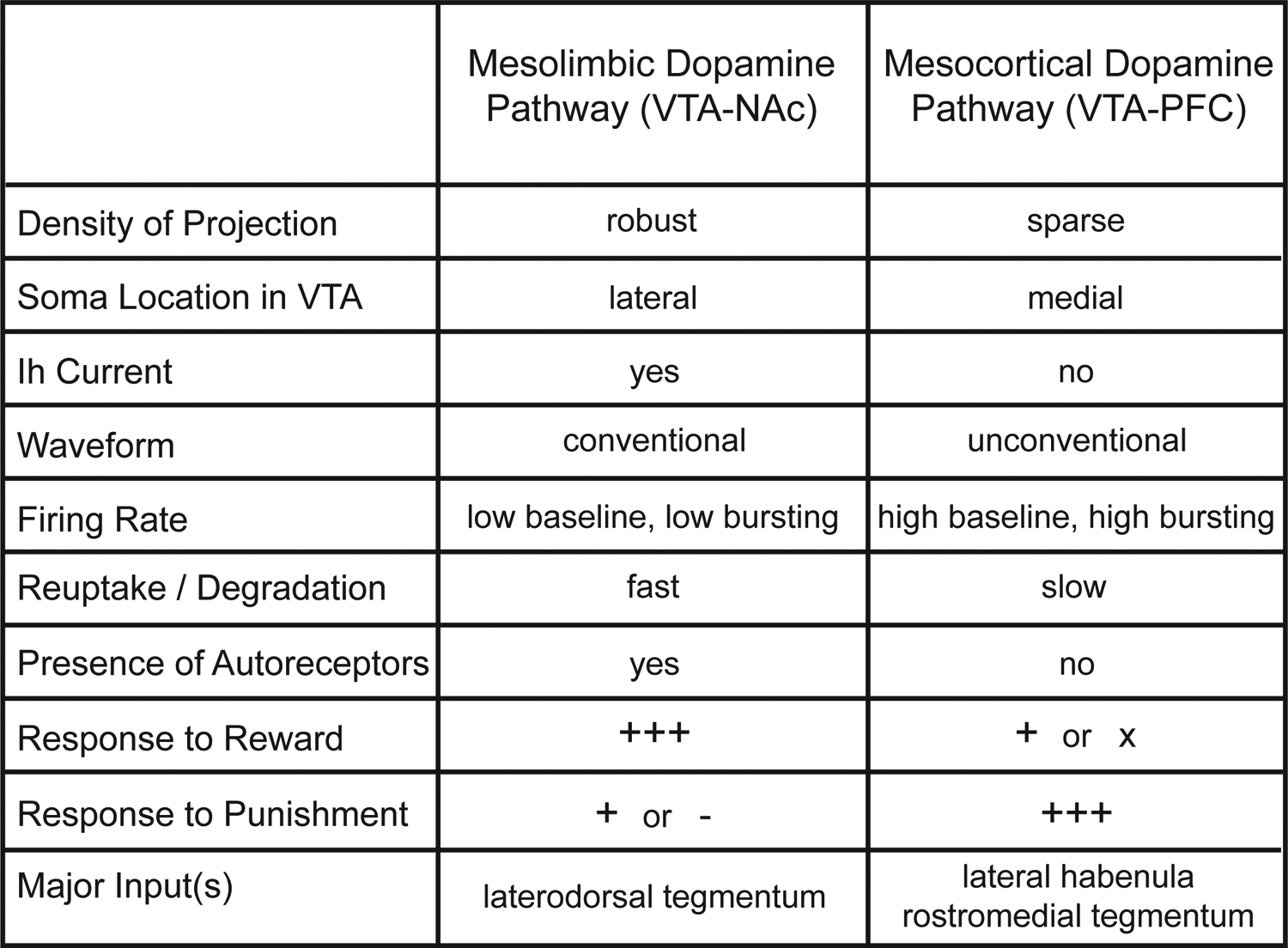
Abridged summary of differences between the mesolimbic and mesocortical dopamine pathways.
In rodents, although dopaminergic terminals in the mPFC are relatively sparse (Descarries et al., 1987) and correspondingly, dopamine release is lower compared to other forebrain regions, such as the NAc (Garris and Wightman, 1994; Garris et al., 1993), motivationally-relevant stimuli robustly modulate dopamine neurotransmission in the mPFC (Abercrombie et al., 1989; Bassareo et al., 2002; Finlay et al., 1995; Hernandez and Hoebel, 1990), which profoundly impacts mPFC activity (Arnsten, 1997; Berridge and Arnsten, 2013; Decot et al., 2017; Seamans and Yang, 2004; Sesack and Bunney, 1989). Measurements of neurotransmitter release and neuronal activity show the mPFC dopamine pathway is activated by both rewarding (Ahn and Phillips, 1999; Bassareo et al., 2002; Ellwood et al., 2017; St Onge et al., 2012) and aversive stimuli (Finlay et al., 1995, 1995; Kim et al., 2016a; Lammel et al., 2011; Mantz et al., 1989; Thierry et al., 1976). However, several studies have suggested that the mPFC dopamine system is preferentially sensitive to aversive events.
An important series of studies by Lammel and colleagues demonstrated that projection-defined VTA dopamine neurons differentially respond to rewarding and aversive stimuli. More specifically, these studies have suggested that VTA dopamine neurons projecting to the NAc encode aspects of reward or reinforcement, while VTA dopamine neurons projecting to the mPFC may encode aspects of aversion (Lammel et al., 2011, 2012). First, Lammel and colleagues (2011) showed that enhanced synaptic plasticity onto VTA dopamine neurons following rewarding and aversive events depends on projection-defined subpopulations. In vitro AMPA/NMDA ratios, as a measure of synaptic strength, are enhanced in VTA dopamine neurons projecting to the NAc medial shell after cocaine experience – a highly rewarding and reinforcing experience (Deroche et al., 1999; Nomikos and Spyraki, 1988; Roberts et al., 1977). In contrast, aversive stimulus experience — formalin injection into the paw (Dubuisson and Dennis, 1977) – did not change AMPA/NMDA in NAc medial shell-projecting VTA dopamine neurons. The opposite pattern of results was observed in VTA dopamine neurons projecting to the mPFC: synaptic plasticity was enhanced following aversive experience but not after rewarding experience (Lammel et al., 2011).
Direct measurements of dopamine activity have also revealed activation of the mesocortical pathway in response to noxious stimuli. In an early, influential in vivo microdialysis study, Abercrombie and colleagues showed that dopamine in the mPFC is robustly elevated following tail pinch compared to dopamine levels in the dorsal and ventral portions of the striatum, despite significantly denser striatal innervation (Abercrombie et al., 1989). Within the VTA, in vivo electrophysiological recordings of putative mPFC- and NAc-projecting dopamine neurons revealed preferential responsivity of the mesocortical pathway to tail pinch – with 65% exhibiting excitation and 25% inhibition – whereas NAc-projecting units were largely non-responsive (Mantz et al., 1989). More recently, bulk calcium signals from dopaminergic axon terminals recorded simultaneously in the mPFC and NAc with in vivo fiber photometry show divergent responsivity to rewarding and aversive stimuli (Kim et al., 2016). Here, VTA dopamine terminals in the NAc were activated by water reward and inhibited by tail shock, whereas terminals in the mPFC were activated by tail shock and not robustly modulated by water reward (Kim et al., 2016).
While microdialysis and photometry studies have been used to make critical discoveries regarding the function of mesocortical dopamine, these methods are limited in several important ways (for review, Wickham et al., 2013). Microdialysis provides a direct measure of dopamine release, but is limited in temporal resolution as samples are collected every on the order of minutes. Photometry has temporal resolution in the low seconds range, closer to that of real-time dopamine signaling, but is an indirect, proxy measure of dopamine release. The most direct way of monitoring real-time phasic dopamine release is fast-scan cyclic voltammetry (FSCV), which has been used with great success in dopamine-rich areas such as the striatum, but has proven difficult to implement in the mPFC. Indeed, while FSCV offers a direct measurement of catecholamine neurotransmission with precise temporal resolution, it is rarely used outside the NAc (but see: Garris and Wightman, 1994; Garris et al., 1993; Matthews et al., 2016) due to difficulty in discriminating between norepinephrine and dopamine (Heien et al., 2003) as well as relatively poor sensitivity – which has limited its application in brain regions like the mPFC which receive both dopaminergic and noradrenergic innervation (Figure 1a), and also have much lower magnitude dopamine release events. Thus, the time course and source of dopamine release in the mPFC upon aversive stimuli presentation has been difficult to parse with this method, and instead has been inferred from less direct approaches. Recently however, we used optogenetic and pharmacological methods to identify the source of dopamine release in the mPFC in response to tail pinch. Consistent with previous studies, we found that tail pinch produces rapid elevations in catecholamine release that are time-locked to its onset (Vander Weele et al., 2018), particularly in the deep layers of the mPFC where dopamine terminals are the densest relative to norepinephrine terminals (Figure 1c). Considering optogenetic inhibition of VTA dopamine neurons attenuated tail pinch evoked release, and that pharmacological inactivation of norepinephrine neurons in the locus coeruleus did effect responses to tail pinch (Vander Weele et al., 2018), these findings strongly suggests that this catecholamine signal is dopaminergic and arises from VTA dopamine neurons.
While several reports have convincingly demonstrated that mesocortical dopamine is preferentially responsive to aversive stimuli, this conclusion does not resolve all of the empirical data. Indeed, several microdialysis studies have reported enhancements in extracellular dopamine release in the mPFC following exposure to rewarding stimuli, specifically food consumption (Ahn and Phillips, 1999; Bassareo et al., 2002; Hernandez and Hoebel, 1990; St. Onge et al., 2012). Recently, phasic dopamine responses were reported from VTA dopamine terminals in the mPFC in response to reward, measured by in vivo fiber photometry (Ellwood et al., 2017). Here they observed an increase in axonal calcium signals from VTA dopamine terminals in the mPFC in response to reward-predictive cues and reward-delivery (Ellwood et al., 2017), reminiscent of dopamine transients observed in the NAc using in vivo fast-scan cyclic voltammetry during a similar Pavlovian conditioning paradigm (Day et al., 2007). These data suggest that dopamine neurotransmission in the mPFC may play a role in reward-related processes. However, non-psychostimulant addictive drugs (e.g., morphine, ethanol, nicotine) do not enhance dopamine release in the mPFC (Bassareo et al., 1996), despite their rewarding and reinforcing properties. Further, drugs that enhance dopamine concentrations in the mPFC (e.g., antidepressants) are not typically substances of abuse (Tanda et al., 1994), suggesting that dopamine in mPFC has a differential role in reward and reinforcement as compared to accumbal dopamine. While distinct from mesolimbic dopamine, mesocortical dopamine still plays an important role in self-administration of abused compounds. Indeed, destruction of dopamine terminals in the mPFC increases motivation to self-administer cocaine in rats (McGregor et al., 1996), and micro-infusions of cocaine into the mPFC are capable of maintaining self-administration (Goeders and Smith, 1983). Reconciling these findings is difficult for several reasons including the disparate time scales over which dopamine has been recorded (e.g. FSCV versus microdialysis), and the fact that many studies only assessed the effects of either an aversive or rewarding stimuli rather than comparing between the two. Indeed Ellwood and colleagues (2017) did not also investigate dopamine responsivity to aversive stimuli, omission of reward, or under competitive situations. It remains possible that dopamine optimizes prefrontal functions in each given context and that aversive stimuli represent a robust strategy-switching signal that is reflected in the mesocortical dopamine pathway’s sensitivity to aversive events. Another potential explanation is that mesocortical dopamine is preferentially responsive to aversive stimuli on a short, second to second timescale, but is elevated by rewarding stimuli over longer periods of time. Additional studies will be needed to fully resolve this important discrepancy in the extant literature.
The idea that mesocortical dopamine is preferentially sensitive to aversive stimuli has been proposed several times to be a unique feature of this population, but largely has not been incorporated into overarching models and theories of mesocortical dopamine function. In a second study by Lammel and colleagues (2012) they show that activation of inputs onto VTA neurons projecting to either NAc or mPFC with optogenetics resulted in reinforcement and aversion, respectively. Further, direct activation of VTA dopamine terminals has been shown to support conditioned place avoidance and promote anxiety-related behavior (Gunaydin et al., 2014), while destruction of dopaminergic innervation of the mPFC disrupts escape behavior (Sokolowski et al., 1994). Yet the role of mesocortical dopamine in aversive motivation remains controversial. For example, in stark contrast to the initial reports by Gunaydin and colleagues (2014), two recent studies showed no effect of optical activation of VTA dopamine axon terminals on place preference (Ellwood et al., 2017, Vander Weele et al., 2018). These seemingly contradictory reports fail to provide a cohesive understanding of the functional role of dopamine in the mPFC.
Positive and negative cues that guide motivated behaviors rarely exist in isolation outside of the laboratory. In naturalistic settings, stimuli that predict rewards and punishments coexist and the mPFC evaluates the probability of their outcomes in order to guide adaptive behavior. However, few preclinical studies have explored dopaminergic modulation of competitive situations. Dopaminergic modulation of mPFC activity may have profound behavioral impact considering the importance of mPFC in higher-order cognition, decision-making and evaluation of uncertain situations (Floresco, 2013; Floresco and Magyar, 2006; Ragozzino, 2002). Recently, a behavioral assay was developed to examine the role of mesocortical dopamine during conflicting Pavlovian associations (Burgos-Robles et al., 2017). In this assay, rats learned to discriminate between sucrose- and footshock-predictive cues, resulting in approach to the reward delivery port and freezing, respectively. After learning, conflict was introduced by simultaneously presenting the two stimuli to induce an intermediate behavioral phenotype – characterized by within- and between-subject individual differences in sucrose port approach and freezing behavior. Optogenetic activation of VTA dopamine terminals in the mPFC during these competition trials biased behavior towards freezing at the expense of port approach and sucrose consumption (Vander Weele et al., 2018). These data suggest that under certain conditions, the sensitivity of the mesocortical system to aversive stimuli may tune mPFC circuits to promote avoidance or escape behaviors, and may help to explain the variability across studies in the ability of dopamine in the mPFC to directly condition aversive responses.
6. Valence routing by prefrontal projections
It is still unknown if neural responses in mPFC carrying information of competing valences can be mapped onto specific output circuits, what modulates their competitive interactions, and how competing information streams ultimately win control of behavioral outputs. Further, considering mPFC neurons are responsive to both rewarding and aversive stimuli (Burgos-Robles et al., 2009, 2013; Euston et al., 2012), it is possible that mPFC neuronal circuits encoding aversive or rewarding events are differentially modulated by dopamine, resulting in a vast array of computations and behavioral outcomes. As previously mentioned, subcortical projection predominately reside in output Layers 5 and 6 (Gabbott et al., 2005), which also express dopamine receptors (Gaspar et al., 1995). Prominent projections from the mPFC include the: basolateral amygdala, NAc / ventral striatum, lateral hypothalamus, periaqueductal gray, medial dorsal thalamus, and paraventricular nucleus of the thalamus (Gabbott et al., 2005; Vertes 2004). Here we highlight a subset of projection-defined mPFC subpopulations implicated in motivated behaviors in response to rewarding and aversive stimuli.
mPFC-NAc
The NAc is historically viewed as a reward center because of its involvement in reward-learning (Day and Carelli, 2007; Day et al., 2007), reward-seeking (Phillips et al., 2003) and hedonic pleasure (Castro and Berridge, 2014). mPFC projections to the NAc comprise ~27% of Layer 5 pyramidal neurons and ~13% of superficial Layer 2/3 (Gabbott et al., 2005). Optogenetic stimulation of mPFC terminals in the NAc has been shown to support place preference and operant reinforcement in some (Britt et al., 2012), but not in all studies (Otis et al., 2017; Stuber et al., 2011, Vander Weele et al., 2018) – perhaps due to location of stimulation (i.e., soma versus terminal) or stimulation parameters (i.e., frequency, light power). Recently, in vivo cellular resolution calcium imaging of mPFC-NAc neurons during reward-learning has revealed enhanced responses to reward-predictive cues during conditioning in this pathway (Otis et al., 2017), consistent with previous reports (Ishikawa et al., 2008). Given that optogenetic stimulation promotes the expression of conditioned-reward-seeking (Otis et al., 2017), current evidence ties the excitatory mPFC-NAc pathway to orchestrating components of reward-learning and seeking. Recently, the mPFC-NAc pathway has proven to demonstrate combined coding for social rewards, representing both social and spatial coding (Murugan et al., 2017). These studies suggest that the mPFC-NAc projection may represent diverse features about cues and contexts related to reward.
mPFC-PAG
The PAG is also innervated by mPFC, but this population of mPFC output neurons is functionally and anatomically distinct (Vander Weele et al., 2018) from those that project to the NAc. The PAG is a midbrain structure considered an important relay center for information regarding threatening stimuli and subsequent defensive responses (Bandler and Carrive, 1988; Bandler and Depaulis, 1988; Bandler et al., 1985; Fanselow, 1991; Tovote et al., 2016). In humans, the PAG is implicated in mediating negative affective states and processing of aversive events (Buhle et al., 2013; Satpute et al., 2013; Young et al., 1985), threat proximity (Canteras and Graeff, 2014; Mobbs et al., 2007, 2010), threat anticipation (Meyer et al., 2017), and escape (Evans et al., 2018). In non-human primates, local electric and glutamate stimulation of the PAG elicits a diverse range of aversive and defensive behavioral reactions, which are topographically segregated in subcolumns along both medial-lateral and dorsal-ventral axes (Bandler and Depaulis, 1988; Deng et al., 2016). mPFC projections to the PAG comprise ~3% of Layer 5 pyramidal neurons (Gabbott et al., 2005) and innervate both dorsal (dPAG) and ventral (vPAG) subregions. mPFC projections to the dPAG are comprised entirely of glutamatergic neurons, and synapse on glutamatergic neurons in the dPAG (Franklin et al., 2017). Although the genetic identity of mPFC projections to the vPAG have not been explored, the ventral and dorsal subregions are known to regulate distinct fear-related operations. Indeed, while the dPAG orchestrates active threat avoidance strategies (i.e., escape), the vPAG promotes passive coping strategies (i.e., freezing) (Bandler and Carrive, 1988; Deng et al., 2016; Halladay and Blair, 2015; Tovote et al., 2016). These functional distinctions also appear to be reflected in cortical input to PAG subregions along the dorsal-ventral axis. Optogenetic activation of the mPFC-dPAG pathway specifically supports conditioned and real-time place aversion / avoidance and promotes marble burying (Vander Weele et al., 2018), a defensive behavior in rodents in response to localizable threats (De Boer and Koolhaas, 2003). In contrast, the mPFC projections to vPAG mediate aspects of contextual fear conditioning and discrimination (Rozeske et al., 2018). Together these data strongly suggest that the mPFC projections to the PAG mediated various coping strategies in response to aversive stimuli.
mPFC-thalamus
The mPFC also has dense reciprocal connectivity with medial dorsal thalamic nuclei (Collins et al., 2018; Gabbott et al., 2005; Ray and Price, 1993; Schmitt et al., 2017), which are anatomically distinct from other mPFC outputs (Otis et al., 2017). Thalamic projections arise predominately from deep layer 6, comprising almost half of Layer 6 pyramidal neurons (Gabbott et al., 2005). While mPFC neurons innervate various thalamic structures, two downstream targets of interest are the mediodorsal thalamus (MD) and periventricular nucleus of the thalamus (PVT). Similar to projections to NAc and PAG, mPFC projections to the PVT have been implicated in valence processing. Do-Monte and colleagues (2015) have shown that during late-stage fear memory retrieval, mPFC projections to PVT are activated, and that this activity is necessary for fear memory retrieval (Do-Monte et al., 2015). Complimentary findings have demonstrated that this circuit is tuned during reward learning such that reward-predictive cues elicit inhibitory responses, which may be permissive to the expression of cued reward seeking (Otis et al., 2017). Accordingly, optogenetic activation of mPFC projections to the PVT time-locked to the presentation of a reward-predictive cue blocks both the acquisition and expression of conditioned reward-seeking. Given that activation of mPFC-PVT neurons does not to support real-time place preference or aversion (Otis et al., 2017), these data implicate this pathway in mediating conditioned/learned, rather than unconditioned/innate motivated behaviors. Connections between the mPFC and the thalamic nucleus reuniens (RE) are involved in the extinction of Pavlovian fear memories and appear to inhibit the expression of fear (Ramanathan et al., 2018). While there is rich preclinical literature exploring MD inputs to the mPFC, particularly in the context of working memory and behavioral flexibility (Bolkan et al., 2017; Schmitt et al., 2017; Wimmer et al., 2015), there have been considerably less functional investigations in the reciprocal direction (but see: Marton et al., 2018). Considering the density of the mPFC-MD pathway and its association with neuropsychiatric disorders (George et al., 2001; Greicius et al., 2007), exploration into its role in processing of rewarding and aversive stimuli could be an interesting avenue for future research.
mPFC-BLA
Connections between the mPFC and BLA have received significant attention (Adhikari et al., 2015; McGarry and Carter, 2017; Murugan et al., 2017; Sotres-Bayon and Quirk, 2010; Yizhar and Klavir, 2018), given the BLA’s role in processing salient and valance-defined stimuli (Janak and Tye, 2015; Namburi et al., 2015; Shabel and Janak, 2009). mPFC neurons projecting to the BLA originate in both Layer 2/3 (10% of Layer 2/3 pyramidal neurons) and Layer 5 (8% of Layer 5 pyramidal neurons (Gabbot et al., 2005). mPFC-BLA projectors appear to be anatomically distinct from other projector populations (Murugan et al., 2017), but there is evidence of collateralization (Vander Weele et al., 2018). This circuit has been implicated in mediating components of aversive behaviors, including forms of fear-learning (Allsop et al., 2018; Karalis et al., 2016), fear-extinction (Adhikari et al., 2015; Bukalo et al., 2015), anxiety (Adhikari et al., 2015), and innate threat responses (Jhang et al., 2018) (for review, Rozeske and Herry, 2018; Sotres-Bayon and Quirk, 2010). Indeed, theta oscillations in this pathway have been casually linked to the expression of fear-behavior (Karalis et al., 2016). However, it is important to appreciate the heterogeneity across prefrontal subregions in regard to the function of their projections to the BLA. For example, Adhikari and colleagues (2015) show that dorsal and ventral mPFC neurons projecting to BLA differentially regulate conditioned fear responses and anxiety. Here they demonstrate that ventral mPFC-BLA inhibits fear- and anxiety-related behaviors, while dorsal mPFC-BLA promotes anxiety (Adhikari et al., 2015). Although beyond the scope of this review, reciprocal projections also exist from the BLA to the mPFC (Burgos-Robles et al., 2017; Felix-Ortiz et al., 2016; McGarry and Carter, 2017).
Diversity also exists within other prefrontal circuits – ACC projections to the BLA are implicated in both innate threat responses and socially transmitted fear learning, but seem to be less involved in classically conditioned fear responses (Burgos-Robles et al., 2017; Felix-Ortiz et al., 2016; McGarry and Carter, 2017)j. Functional heterogeneity based on projection origin within the mPFC subregions likely extend to other mPFC output populations (i.e., mPFC-PAG, mPFC-NAc), but largely have not been parsed to this level of granularity. Overlaying the functional aspects of defined mPFC circuits on the dorsal-ventral axis of the mPFC will be an important future direction.
Other mPFC circuits
The mPFC contains several other projection-defined and local subpopulations that may be responsible for differential routing of reward and aversive information. For example, the pathway between the mPFC and dorsal raphe nucleus (DRN) is involved in the selection and activation of coping behaviors in the face of threat (Baratta et al., 2009; Warden et al., 2012), but further investigation of this pathway in both reward and aversive contexts is required. Additionally, while the lateral hypothalamus has been implicated in both aversive and appetitive behaviors, the role of input from the mPFC has not yet been thoroughly interrogated, despite relatively dense connectivity (Gabbott et al., 2005). Finally, active avoidance requires inhibitory signaling in the mPFC (Diehl et al., 2018) and a subsets of local inhibitory cells have also been shown to be important for fear responses (Courtin et al., 2014; Sparta et al., 2014). Yet understanding how these local microcircuits interface with specific projections, and how dopamine may regulate these interactions to modulate their behavioral outcomes remains an ongoing effort.
7. Dopaminergic mechanisms of action and circuit modulation in the mPFC
One difficulty in unraveling the mystery of dopamine’s actions in the prefrontal cortex is in simply determining the post-synaptic actions of dopamine on mPFC neurons. This is a particularly complex issue compared to the well-documented effects of dopamine on principle neurons in the striatum. In the striatum, D1 and D2-type dopamine receptors (based on mRNA expression) are expressed on almost entirely non-overlapping populations of medium spiny neurons, the primary output neuron of the striatum which account for more than 95% of total striatal neurons (Gerfen and Surmeier, 2011; Tepper et al., 2008). Dopamine receptors are densely expressed in striatal medium spiny neurons, but very few, co-express D1 and D2 receptors (Valjent et al., 2009); although they may express other dopamine receptor subtypes. Striatal D1- and D2-expressing medium spiny neurons are anatomically distinct, as they have non-overlapping projection targets (though this distinction may be less clear in the ventral striatum, see Kupchik et al., 2015), and activation of these two populations produces appetitive and aversive behavioral responses, respectively (Kravitz et al., 2012). While dopamine action in the striatum is by no means simple, the relatively clear-cut mRNA expression patterns of dopamine receptors in populations with diametrically opposed functions lend itself well to parsing the physiological and behavioral actions of dopamine in this area.
Indeed, controversy has long surrounded the topic of how dopamine alters the firing of mPFC neurons, and which types of neurons are affected. Seemingly contradictory results from different groups performing whole-cell patch-clamp recordings from pyramidal neurons in Layer 5 of mPFC have reported dopamine-induced increases (Penit-Soria et al., 1987; Yang and Seamans, 1996) or decreases (Bunney and Aghajanian, 1976; Gulledge and Jaffe, 1998) in activity. One possible explanation is that because the neurons were randomly sampled from Layer 5, different studies may have recorded from different projection and/or genetic subpopulations due to sampling bias. This notion is supported by more recent studies demonstrating that different cell types in mPFC display differential responses to dopamine (Gee et al., 2012; Tritsch and Sabatini, 2012, Vander Weele et al., 2018)
In the mPFC, dopaminergic terminals synapse on both pyramidal neurons (Goldman-Rakic et al., 1989; Séguéla et al., 1988; Zhang et al., 2010) and GABAergic interneurons (Verney et al., 1990). Canonically dopamine exerts its postsynaptic effects on neuronal activity via two main types of receptors— D1-type (including D1 and D5) and D2-type (including D2, D3, and D4) (Gaspar et al., 1995; Santana et al., 2009; Seamans and Yang, 2004; Sesack and Bunney, 1989). Both D1 and D2 sub-types are G-protein-coupled receptors, exerting slow, metabotropic modulation of postsynaptic cells (for review, see: Seamans and Yang et al., 2004). Activation of D1 receptors in the mPFC produces excitatory effects through modulation of sodium (Na+), potassium (K+), and calcium (Ca2+) currents (González-Burgos et al., 2002; Gorelova and Yang, 2000; Henze et al., 2000; Yang and Seamans, 1996). In contrast, activation of mPFC D2 receptors exerts an inhibitory effect through the modulation of glutamatergic receptors and Na+ conductances (Gorelova and Yang, 2000; Gulledge and Jaffe, 1998, 2001). However, receptor signaling and effects are complex in the mPFC (For extensive review, Seamans & Yang, 2004). For example, in both cases, activation of receptors can produce long-lasting (~30 min) or biphasic changes in excitability (Gorelova and Yang, 2000; Gulledge and Jaffe, 1998, 2001; Seamans et al., 2001a, 2001b; Semans and Yang, 2004). Recent studies have demonstrated that D2 receptor signaling in the mPFC is often excitatory (Robinson and Sohal, 2017; Urs et al., 2016, 2017), which may also be developmentally dependent (Tseng and O’Donnell, 2007). Further, dopamine can also have domain-specific physiological effects. Dopamine gates neuronal excitability at the axon initial segment (Clarkson et al., 2017) which may be a mechanism to control action potential initiation (Bender et al., 2010; Kole and Stuart, 2012). In this manner, dopamine may enhance signal-to-noise ratio within mPFC neurons and modulate their responses to presynaptic input. Finer investigations of dopamine’s actions in the mPFC is warranted, particularly in vivo where inputs and endogenous circuitry are intact.
In the mouse mPFC, D1 and D2 receptors are expressed in both pyramidal neurons and GABAergic interneurons (Gaspar et al., 1995; Vincent et al., 1993). D1 receptors are primarily located on the dendritic spines and shafts of pyramidal neurons and on the axon terminals of putative GABAergic interneurons (Bergson et al., 1995a, 1995b; Muly et al., 1998). Further, D1 immunoreactivity is present on presynaptic axon terminals that form asymmetric (i.e., putative glutamatergic) synapses with mPFC dendrites (Paspalas and Goldman-Rakic, 2005), suggesting that dopamine may influence the excitability of mPFC neurons both directly, and indirectly through modulation of excitatory inputs. In pyramidal neurons, D1 and D2 receptors are expressed in Layer 5 (Gaspar et al., 1995; Gee et al., 2012; Santana et al., 2009; Seong and Carter, 2012) – one of the major output layers of the mPFC. D1 receptor mRNA is expressed in 20% Layer 5 neurons, whereas 25% express D2 (Santana et al., 2009). Using pharmacological approaches, dopamine has been shown to increase (Henze et al., 2000; Penit-Soria et al., 1987; Yang and Seamans, 1996) and decrease (Bunney and Aghajanian, 1976; Gulledge and Jaffe, 1998; Sesack and Bunney, 1989) the excitability mPFC neurons – suggesting differential modulation by dopamine depending on mPFC cell-type or projection target. However, the relatively low expression level of dopamine receptors in mPFC projection neurons suggest that a larger portion of Layer 5 pyramidal neurons are not directly modulated by dopamine, but may be subject to dopaminergic modulation of presynaptic inputs (Seamans et al., 2001b; Tritsch and Sabatini, 2012).
The mPFC also expresses D3-, D4-, and D5-receptors. In mPFC pyramidal neurons, the cellular distribution of D1- and D5-receptors overlap with each other, but have distinct ultra-structural targeting: D1-receptor immunoreactivity is enriched on dendritic spines, whereas D5-receptor immunoreactivity is more prominent on dendritic shafts (Bergson et al., 1995c). Bilateral activation of D1- and D5-receptors in the mPFC (via pharmacological SKF-38393 infusion) promotes the incubation of aversive memories (Castillo Díaz et al., 2017) suggesting that this population is involved in the regulation of emotional behaviors. While it has been suggested that D3- and D4-receptors are largely located on GABAergic interneurons (Khan et al., 1998), D3-expressing mPFC projection neurons have been identified (Clarkson et al., 2017). Pharmacological investigations implicate D4-receptors in the encoding of fear (Laviolette et al., 2005), but not reward-related (Woolley et al., 2008) memories, but its still unknown whether D4-receptors are expressed on any specific mPFC projection pathway. Considering there is emerging evidence that suggests that dopamine may differentially modulate specific mPFC projection pathways to optimize circuit computations that result in adaptive behavioral outcomes (Vander Weele et al., 2018), systematic investigation of anatomically distinct mPFC subpopulation could be the catalyst for a new age in understanding mesocortical dopamine (Figure 3).
Figure 3.
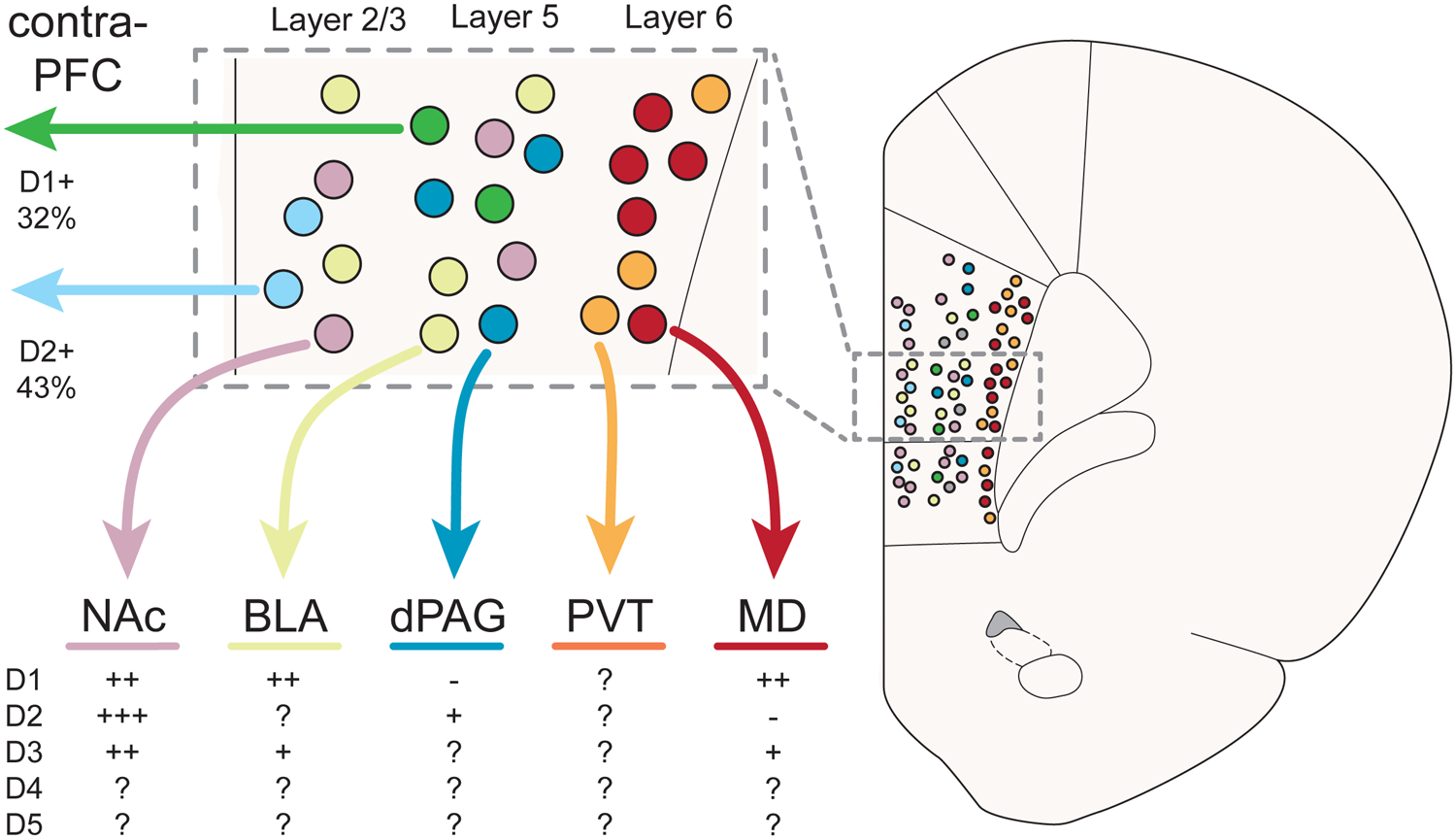
Laminar origins of select mPFC projection populations and their dopamine receptor expression. Abbreviations: BLA = basolateral amygdala; contra-PFC = contralater prefrontal cortex; dPAG = dorsal periaqueductal gray; MD = mediodorsal thalamus; NAc = nucleus accumbens; PVT = periventricular nucleus of the thalamus.
Dopamine on mPFC-NAc
The heterogeneous effects of dopamine on the activity of post-synaptic cells in the mPFC, discussed above, has led to investigations into how dopamine may be differentially affecting projection-defined mPFC neurons. For example, VTA dopamine terminals have been shown to target mPFC neurons that project to the NAc (Carr et al., 1999). Not surprisingly, NAc-projecting mPFC neurons express D1 (Gaspar et al., 1995; Han et al., 2017, Vander Weele et al., 2018), D2 (Gaspar et al., 1995, Vander Weele et al., 2018), and D3 receptors (Clarkson et al., 2017), and their activity is modulated by dopamine (Brady and O’Donnell, 2004; Buchta et al., 2017, Vander Weele et al., 2018). Initial inquiry into the exact distribution and proportion of dopamine receptor expression in striatum-projecting neurons found that ~20% express D1 and 60% express D2 – however this investigation did not isolate neurons projecting to the NAc from neurons projecting to other striatal regions (e.g., dorsal striatum) (Gaspar et al., 1995). Recently, we employed transgenic D1- and D2-Cre mouse driver lines and retrograde labeling to specifically examine dopamine receptor expression on NAc-projecting mPFC neurons, and found slightly higher D1 (~30%) and D2 (~85%) receptor expression (Vander Weele et al., 2018). These difference could be due to the developmental caveats of using transgenic reporter lines (Chan et al., 2012; Tritsch and Sabatini, 2012) or the differentiation of the NAc from other striatal regions. Despite these differences, based on the predominance of D2 expression in both studies, we predicted that the mPFC-NAc circuit would be inhibited by dopamine. Indeed, we found that Layer 5 neurons projecting to the NAc were inhibited by activation of VTA dopamine terminal in ex vivo brain slices, an effect that was dependent on D2-receptors (Vander Weele et al., 2018). These data are consistent with in vivo interrogations of this pathway, which report similar inhibitory actions by dopamine via D2 receptors (Brady and O’Donnell, 2004). Together, it appears that dopamine largely inhibits the mPFC to NAc pathway (but see: Buchta et al., 2017). Given the role of mPFC projections to the NAc in promoting reward-seeking, discussed above, we hypothesize that dopamine-induced inhibition of this circuit acts as a permissive signal for aversive information to be routed downstream through complimentary circuits.
Dopamine on mPFC-PAG
In contrast to the role of mPFC neurons projecting to the NAc in reward seeking, projections to the PAG regulate aversive behavioral responses; however, this pathway does not appear to robustly express dopamine receptors. Indeed, only sparse labeling of terminals is observed in the PAG of D1 transgenic mice (Han et al., 2017). Further, we have investigated this question using D1- and D2-Cre driver lines combined with retrograde labeling and found that only ~5% of mPFC-dPAG projectors express D1 receptors, and ~20% express D2 receptors (Vander Weele et al., 2018). This was corroborated by functional studies showing that VTA dopamine terminal stimulation does not affect excitability of this circuit in ex vivo brain slices (Vander Weele et al., 2018). However, evidence supports indirect modulation of mPFC pyramidal neurons by dopamine — either via interactions with local GABAergic interneurons or modulation of other inputs (Paspalas and Goldman-Rakic, 2005; Seamans et al., 2001b; Tritsch and Sabatini, 2012), which motivated us to test the impact of dopamine on this projection in vivo where the circuitry required for indirect modulatory mechanisms is still intact. In in vivo preparations we have found that dopamine increases signal-to-noise ratio in mPFC-dPAG neurons during processing of aversive stimuli (Vander Weele et al., 2018). Specifically, VTA dopamine terminal stimulation enhanced neural responses to airpuff in photoidentified mPFC-dPAG neurons, but did not effect responses to sucrose delivery. In the homecage where no aversive stimuli were present, single-cell calcium imaging recordings revealed that VTA dopamine terminal stimulation enhanced the amplitude of individual calcium transients in mPFC-dPAG neurons but decreased there overall frequency – perhaps by filtering relevant inputs or changing the pattern of firing. Indeed, we found that VTA dopamine terminal stimulation selectively increased the average frequency within a burst firing event within mPFC-dPAG (Vander Weele et al., 2018), effectively increasing the signal-to-noise ratio between burst and baseline firing. Considering calcium transient amplitude linearly related to the number of spikes within a burst event (Chen et al., 2013; Otis et al., 2017), this pattern of firing may underlie changes in calcium event amplitude. These data suggest that dopamine in the mPFC may promote the encoding and subsequent behavioral responses to aversive stimuli by modulating incoming inputs impinging upon the mPFC-dPAG circuit.
Dopamine on mPFC-BLA
Several studies suggest that mPFC-BLA neurons express D1 (Han et al., 2017; Land et al., 2014; Rey et al., 2014) and perhaps to a lesser degree D2 (Pinto and Sesack, 2008; Rosenkranz and Grace, 2001), and D3 (Clarkson et al., 2017) – although the exact proportions are still unclear. Unlike projections to the NAc and dPAG (Vander Weele et al., 2018), anatomical investigations using retrograde tracing and dopamine receptor labeling in the mPFC-BLA have yet to be completed. Although the mPFC-BLA connection has largely been studied in the context for fear and aversive responses, specific activation of mPFC D1-expressing terminals in the BLA promotes feeding (Land et al., 2014), a behavior typically regarded as reward-related. However, functional heterogeneity also exists within the BLA (Beyeler et al., 2016; Kim et al., 2016b; Namburi et al., 2015). Given that D1 and D2 receptor activation differentially influence BLA neuronal responses to mPFC input (Rosenkranz and Grace, 2002), it is tempting to speculate that via this mechanism dopamine could route information about positive and negative stimuli to specific subpopulations of neurons responsible for valence-encoding within the BLA. Future studies dissecting the precise connectivity and behavioral operations of mPFC D1- and D2-expressing inputs to the BLA could yield exciting new insights. To date, there is not enough evidence to form conclusions regarding how dopamine directs the flow of information about motivationally-relevant stimuli within the mPFC-BLA circuit.
Dopamine on mPFC-thalamus and other circuits
Even less is known about dopaminergic modulation of other mPFC projections, including those to the MD and PVT. Although we’ve known that ~25% of mPFC-MD neurons express D1 mRNA for some time (Gaspar et al., 1995; Han et al., 2017), little is known about their cognitive of behavioral functions. Interestingly, while most mPFC-MD projectors reside in Layer 6 and do not express D2 receptor (Gaspar et al., 1995), a small population of MD projectors reside in Layer 5 and almost exclusively express D2 (Gee et al., 2012). Given this stark contrast, interesting functional differences between Layer 5 and Layer 6 mPFC-MD pathways may be revealed in the future. Dopamine receptor localization on other projectors, including the PVT is still unclear; however, a recent anatomical characterization D1 transgenic driver line suggest that mPFC D1 neurons may also project to the dorsal striatum, claustrum, and ventral thalamus (Han et al., 2017). D3-expressing mPFC neurons have been identified within mPFC-NAc, mPFC-BLA, and mPFC-MD projection pathways (Clarkson et al., 2017), as discussed in the sections above. However, the precise behavioral function of these specific subpopulations has yet to be determined.
8. Dopamine in the mPFC sets the tone for aversive circuit computations
In summary, midbrain dopamine neurons are critically involved in modulating neural circuits responsible for appetitive and aversive motivation. While the impact of dopamine release within the striatum is studied extensively, particularly in the context of reward-related behaviors, the mesocortical pathway’s role in motivation has been relatively understudied. Mounting evidence suggests that under certain conditions, the responsivity of the mesocortical system to aversive stimuli may promoting behavioral coping strategies in response to aversive outcomes and threatening stimuli (Lammel et al., 2011, 2012; Vander Weele et al., 2018). Further, considering mPFC neurons are responsive to both rewarding and aversive stimuli (Burgos-Robles et al., 2009, 2013; Euston et al., 2012), it is possible that mPFC neuronal circuits encoding aversive or rewarding events are differentially modulated by dopamine – a theory supported by emerging results (Vander Weele et al., 2018) (Figure 4). This flexibility in function can result in a vast array of computations and behavioral outcomes that could highly depend on environmental context. For example, in environments where only reward-related stimuli are present, perhaps dopamine in the mPFC functions to optimize reward-learning and acquisition. However, when threatening stimuli are present, an organism must rapidly and efficiently respond to avoid harm. Perhaps in these situations, the sensitivity of mesocortical dopamine to aversive stimuli serves as a robust strategy-switching signal to aid in threat avoidance. In these situations, dopamine would set the stage for aversive processing by amplifying inputs onto cortical pathways essential for escape and avoidance, while suppressing information that does not require immediate action.
Figure 4.
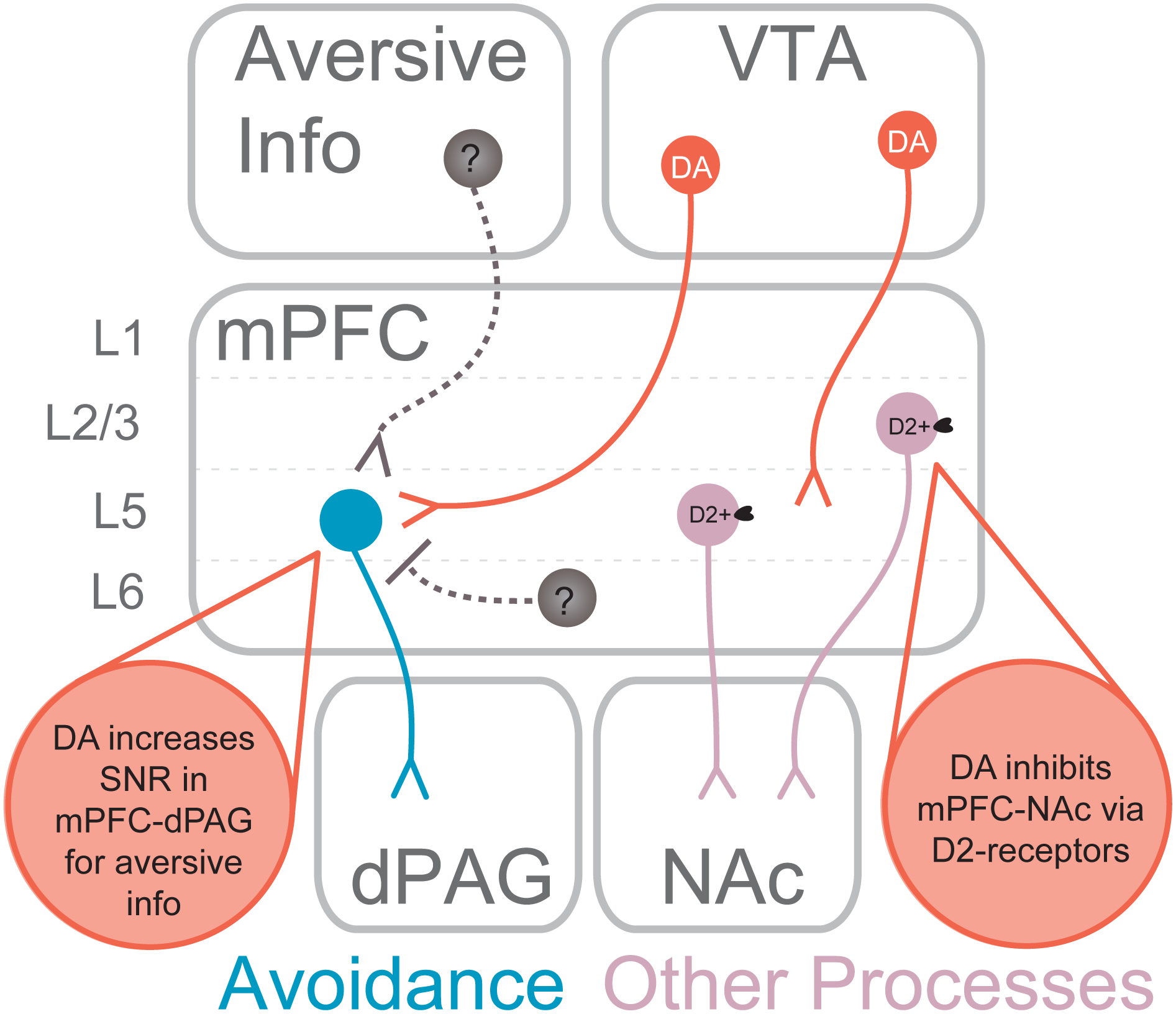
Summary of findings found in Vander Weele et al., 2018. Threatening events evoke dopamine release in the mPFC which enhances the signal-to-noise ratio of responses to aversive stimuli within the mPFC-dPAG pathway. Dopamine suppresses excitability of neurons in the mPFC-NAc pathway via D2-receptors. These actions support adaptive behavioral responses to threat, such as avoidance.
The notion that dopamine can alter signal-to-noise is attractive, as dopamine-mediated increases in signal-to-noise ratio could explain its diverse behavioral and cognitive functions. Despite the popularity of the signal-to-noise model for mPFC dopamine in computational and theoretical neuroscience (Cohen and Servan-Schreiber, 1992; Cohen et al., 2002; Durstewitz and Seamans, 2008; Rolls et al., 2008), the degree to which it translates across brain functions is unknown. Support for dopamine-mediated signal-to-noise ratio modulations has been found ex vivo (Kroener et al., 2009; Seamans et al., 2001b), and in vivo during auditory stimulus discrimination (Popescu et al., 2016), working memory (Williams and Goldman-Rakic, 1995), and visual cortex orientation selectivity (Noudoost and Moore, 2011). Dopamine may function similarly within the mPFC to optimize motivated behaviors. Our recent findings show that dopamine release serves to sharpen responses to motivationally significant stimuli by simultaneously enhancing the signal-to-noise in aversion-encoding pathways and inhibiting pathways that contribute to other behaviors. Therefore, while it is still debated whether mesocortical dopamine release is innately aversive (Ellwood et al., 2017; Gunaydin et al., 2014; Popescu et al., 2016, Vander Weele et al., 2018), we propose that mesocortical dopamine promotes responding to aversive stimuli when necessary via the fine-tuning of cortical subpopulations carrying information of positive or negative valence.
HIGHLIGHTS.
Dopamine has diverse actions on prefrontal cortex physiology and function.
The mesocortical dopamine pathway is uniquely sensitive to aversive stimuli.
Dopamine differentially influences projection-defined prefrontal subpopulations.
By calibrating distinct prefrontal outputs, dopamine sets the stage for aversive processing.
FUNDING AND ACKNOWLEDGEMENTS
K.M.T. is a New York Stem Cell Foundation - Robertson Investigator, a McKnight Scholar and this work was supported by funding from the JPB Foundation, PIIF, PNDRF, JFDP, Klingenstein Foundation, NARSAD Young Investigator Award, New York Stem Cell Foundation, NIH R01-MH102441-01 (NIMH), NIH Director’s New Innovator Award DP2-DK-102256-01 (NIDDK), and Pioneer Award DP1-AT009925 (NCCIH). C.M.V.W. was supported by the NSF Graduate Research Fellowship and the Integrative Neuronal Systems Training Fellowship (T32 GM007484). C.A.S. is supported by NIH grants F32 MH111216 (NIMH) and K99 DA045103 (NIDA), and a NARSAD Young Investigator Award from the Brain and Behavior Research Foundation.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
REFERENCES
- Abercrombie ED, Keefe KA, DiFrischia DS, and Zigmond MJ (1989). Differential effect of stress on in vivo dopamine release in striatum, nucleus accumbens, and medial frontal cortex. J. Neurochem 52, 1655–1658. [DOI] [PubMed] [Google Scholar]
- Adhikari A, Lerner TN, Finkelstein J, Pak S, Jennings JH, Davidson TJ, Ferenczi E, Gunaydin LA, Mirzabekov JJ, Ye L, et al. (2015). Basomedial amygdala mediates top-down control of anxiety and fear. Nature 527, 179–185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Adolphs R (2009). The Social Brain: Neural Basis of Social Knowledge. Annu. Rev. Psychol 60, 693–716. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ahn S, and Phillips AG (1999). Dopaminergic correlates of sensory-specific satiety in the medial prefrontal cortex and nucleus accumbens of the rat. J. Neurosci. Off. J. Soc. Neurosci 19, RC29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Albanese A, and Minciacchi D (1983). Organization of the ascending projections from the ventral tegmental area: a multiple fluorescent retrograde tracer study in the rat. J. Comp. Neurol 216, 406–420. [DOI] [PubMed] [Google Scholar]
- Allsop SA, Wichmann R, Mills F, Burgos-Robles A, Chang C-J, Felix-Ortiz AC, Vienne A, Beyeler A, Izadmehr EM, Glober G, et al. (2018). Corticoamygdala Transfer of Socially Derived Information Gates Observational Learning. Cell 173, 1329–1342.e18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anand BK, and Brobeck JR (1951). Hypothalamic control of food intake in rats and cats. Yale J. Biol. Med 24, 123–140. [PMC free article] [PubMed] [Google Scholar]
- Arnsten AFT (1997). Catecholamine regulation of the prefrontal cortex, Catecholamine regulation of the prefrontal cortex. J. Psychopharmacol. (Oxf.) 11, 151–162. [DOI] [PubMed] [Google Scholar]
- Arnsten AF, and Dudley AG (2005). Methylphenidate improves prefrontal cortical cognitive function through α2 adrenoceptor and dopamine D1 receptor actions: Relevance to therapeutic effects in Attention Deficit Hyperactivity Disorder. Behav. Brain Funct 1, 2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aron AR, Robbins TW, and Poldrack RA (2004). Inhibition and the right inferior frontal cortex. Trends Cogn. Sci 8, 170–177. [DOI] [PubMed] [Google Scholar]
- Assareh N, Sarrami M, Carrive P, and McNally GP (2016). The organization of defensive behavior elicited by optogenetic excitation of rat lateral or ventrolateral periaqueductal gray. Behav. Neurosci 130, 406–414. [DOI] [PubMed] [Google Scholar]
- Badrinarayan A, Wescott SA, Vander Weele CM, Saunders BT, Couturier BE, Maren S, and Aragona BJ (2012). Aversive stimuli differentially modulate real-time dopamine transmission dynamics within the nucleus accumbens core and shell. J. Neurosci. Off. J. Soc. Neurosci 32, 15779–15790. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bains JS, Cusulin JIW, and Inoue W (2015). Stress-related synaptic plasticity in the hypothalamus. Nat. Rev. Neurosci 16, 377–388. [DOI] [PubMed] [Google Scholar]
- Bandler R, and Carrive P (1988). Integrated defence reaction elicited by excitatory amino acid microinjection in the midbrain periaqueductal grey region of the unrestrained cat. Brain Res. 439, 95–106. [DOI] [PubMed] [Google Scholar]
- Bandler R, and Depaulis A (1988). Elicitation of intraspecific defence reactions in the rat from midbrain periaqueductal grey by microinjection of kainic acid, without neurotoxic effects. Neurosci. Lett 88, 291–296. [DOI] [PubMed] [Google Scholar]
- Bandler R, Depaulis A, and Vergnes M (1985). Identification of midbrain neurones mediating defensive behaviour in the rat by microinjections of excitatory amino acids. Behav. Brain Res 15, 107–119. [DOI] [PubMed] [Google Scholar]
- Bannon MJ, and Roth RH (1983). Pharmacology of mesocortical dopamine neurons. Pharmacol. Rev 35, 53–68. [PubMed] [Google Scholar]
- Bannon MJ, Michaud RL, and Roth RH (1981). Mesocortical dopamine neurons. Lack of autoreceptors modulating dopamine synthesis. Mol. Pharmacol 19, 270–275. [PubMed] [Google Scholar]
- Baratta MV, Zarza CM, Gomez DM, Campeau S, Watkins LR, and Maier SF (2009). Selective activation of dorsal raphe nucleus-projecting neurons in the ventral medial prefrontal cortex by controllable stress. Eur. J. Neurosci 30, 1111–1116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barthas F, and Kwan AC (2017). Secondary Motor Cortex: Where ‘Sensory’ Meets ‘Motor’ in the Rodent Frontal Cortex. Trends Neurosci. 40, 181–193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bassareo V, Tanda G, Petromilli P, Giua C, and Chiara GD (1996). Non-psychostimulant drugs of abuse and anxiogenic drugs activate with differential selectivity dopamine transmission in the nucleus accumbens and in the medial prefrontal cortex of the rat. Psychopharmacology (Berl.) 124, 293–299. [DOI] [PubMed] [Google Scholar]
- Bassareo V, De Luca MA, and Di Chiara G (2002). Differential Expression of Motivational Stimulus Properties by Dopamine in Nucleus Accumbens Shell versus Core and Prefrontal Cortex. J. Neurosci. Off. J. Soc. Neurosci 22, 4709–4719. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bechara A, Damasio AR, Damasio H, and Anderson SW (1994). Insensitivity to future consequences following damage to human prefrontal cortex. Cognition 50, 7–15. [DOI] [PubMed] [Google Scholar]
- Bechara A, Tranel D, and Damasio H (2000). Characterization of the decision-making deficit of patients with ventromedial prefrontal cortex lesions. Brain 123, 2189–2202. [DOI] [PubMed] [Google Scholar]
- Bechara A, Damasio H, Tranel D, and Damasio AR (2005). The Iowa Gambling Task and the somatic marker hypothesis: some questions and answers. Trends Cogn. Sci 9, 159–162; discussion 162–164. [DOI] [PubMed] [Google Scholar]
- Bender KJ, Ford CP, and Trussell LO (2010). Dopaminergic Modulation of Axon Initial Segment Calcium Channels Regulates Action Potential Initiation. Neuron 68, 500–511. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bentivoglio M, and Morelli M (2005). Chapter I The organization and circuits of mesencephalic dopaminergic neurons and the distribution of dopamine receptors in the brain In Handbook of Chemical Neuroanatomy, Dunnett SB, Bentivoglio M, Björklund A, and Hökfelt T, eds. (Elsevier; ), pp. 1–107. [Google Scholar]
- Berger B, Gaspar P, and Verney C (1991). Dopaminergic innervation of the cerebral cortex: unexpected differences between rodents and primates. Trends Neurosci. 14, 21–27. [DOI] [PubMed] [Google Scholar]
- Bergson C, Mrzljak L, Lidow MS, Goldman-Rakic PS, and Levenson R (1995a). Characterization of subtype-specific antibodies to the human D5 dopamine receptor: studies in primate brain and transfected mammalian cells. Proc. Natl. Acad. Sci. U. S. A 92, 3468–3472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bergson C, Mrzljak L, Smiley JF, Pappy M, Levenson R, and Goldman-Rakic PS (1995b). Regional, cellular, and subcellular variations in the distribution of D1 and D5 dopamine receptors in primate brain. J. Neurosci. Off. J. Soc. Neurosci 15, 7821–7836. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bergson C, Mrzljak L, Smiley JF, Pappy M, Levenson R, and Goldman-Rakic PS (1995c). Regional, cellular, and subcellular variations in the distribution of D1 and D5 dopamine receptors in primate brain. J. Neurosci. Off. J. Soc. Neurosci 15, 7821–7836. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berke JD (2018). What does dopamine mean? Nat. Neurosci 21, 787–793. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berridge CW, and Arnsten AFT (2013). Psychostimulants and motivated behavior: arousal and cognition. Neurosci. Biobehav. Rev 37, 1976–1984. [DOI] [PubMed] [Google Scholar]
- Berridge KC, and Robinson TE (1998). What is the role of dopamine in reward: hedonic impact, reward learning, or incentive salience? Brain Res. Brain Res. Rev 28, 309–369. [DOI] [PubMed] [Google Scholar]
- Betley JN, Xu S, Cao ZFH, Gong R, Magnus CJ, Yu Y, and Sternson SM (2015). Neurons for hunger and thirst transmit a negative-valence teaching signal. Nature 521, 180–185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beyeler A, Namburi P, Glober GF, Simonnet C, Calhoon GG, Conyers GF, Luck R, Wildes CP, and Tye KM (2016). Divergent Routing of Positive and Negative Information from the Amygdala during Memory Retrieval. Neuron 90, 348–361. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Björklund A, and Dunnett SB (2007). Dopamine neuron systems in the brain: an update. Trends Neurosci. 30, 194–202. [DOI] [PubMed] [Google Scholar]
- Bolkan SS, Stujenske JM, Parnaudeau S, Spellman TJ, Rauffenbart C, Abbas AI, Harris AZ, Gordon JA, and Kellendonk C (2017). Thalamic projections sustain prefrontal activity during working memory maintenance. Nat. Neurosci 20, 987–996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Botvinick MM, Cohen JD, and Carter CS (2004). Conflict monitoring and anterior cingulate cortex: an update. Trends Cogn. Sci 8, 539–546. [DOI] [PubMed] [Google Scholar]
- Brady AM, and O’Donnell P (2004). Dopaminergic modulation of prefrontal cortical input to nucleus accumbens neurons in vivo. J. Neurosci. Off. J. Soc. Neurosci 24, 1040–1049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Breton JM, Charbit AR, Snyder BJ, Fong PTK, Dias EV, Himmels P, Lock H, and Margolis EB (2018). Relative Contributions and Mapping of Ventral Tegmental Area Dopamine and GABA Neurons by Projection Target in the Rat. J. Comp. Neurol [DOI] [PMC free article] [PubMed]
- Brischoux F, Chakraborty S, Brierley DI, and Ungless MA (2009). Phasic excitation of dopamine neurons in ventral VTA by noxious stimuli. Proc. Natl. Acad. Sci 106, 4894–4899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Britt JP, Benaliouad F, McDevitt RA, Stuber GD, Wise RA, and Bonci A (2012). Synaptic and behavioral profile of multiple glutamatergic inputs to the nucleus accumbens. Neuron 76, 790–803. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bromberg-Martin ES, Matsumoto M, and Hikosaka O (2010). Dopamine in motivational control: rewarding, aversive, and alerting. Neuron 68, 815–834. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buchta WC, Mahler SV, Harlan B, Aston-Jones GS, and Riegel AC (2017). Dopamine terminals from the ventral tegmental area gate intrinsic inhibition in the prefrontal cortex. Physiol. Rep 5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Budygin EA, Park J, Bass CE, Grinevich VP, Bonin KD, and Wightman RM (2012). Aversive stimulus differentially triggers subsecond dopamine release in reward regions. Neuroscience 201, 331–337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buhle JT, Kober H, Ochsner KN, Mende-Siedlecki P, Weber J, Hughes BL, Kross E, Atlas LY, McRae K, and Wager TD (2013). Common representation of pain and negative emotion in the midbrain periaqueductal gray. Soc. Cogn. Affect. Neurosci 8, 609–616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bukalo O, Pinard CR, Silverstein S, Brehm C, Hartley ND, Whittle N, Colacicco G, Busch E, Patel S, Singewald N, et al. (2015). Prefrontal inputs to the amygdala instruct fear extinction memory formation. Sci. Adv 1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bunney BS, and Aghajanian GK (1976). Dopamine and norepinephrine innervated cells in the rat prefrontal cortex: pharmacological differentiation using microiontophoretic techniques. Life Sci. 19, 1783–1789. [DOI] [PubMed] [Google Scholar]
- Bunney BS, Chiodo LA, and Grace AA (1991). Midbrain dopamine system electrophysiological functioning: a review and new hypothesis. Synap. N. Y. N 9, 79–94. [DOI] [PubMed] [Google Scholar]
- Burgos-Robles A, Vidal-Gonzalez I, and Quirk GJ (2009). Sustained conditioned responses in prelimbic prefrontal neurons are correlated with fear expression and extinction failure. J. Neurosci. Off. J. Soc. Neurosci 29, 8474–8482. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burgos-Robles A, Bravo-Rivera H, and Quirk GJ (2013). Prelimbic and infralimbic neurons signal distinct aspects of appetitive instrumental behavior. PloS One 8, e57575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burgos-Robles A, Kimchi EY, Izadmehr EM, Porzenheim MJ, Ramos-Guasp WA, Nieh EH, Felix-Ortiz AC, Namburi P, Leppla CA, Presbrey KN, et al. (2017). Amygdala inputs to prefrontal cortex guide behavior amid conflicting cues of reward and punishment. Nat. Neurosci 20, 824–835. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burton MJ, Rolls ET, and Mora F (1976). Effects of hunger on the responses of neurons in the lateral hypothalamus to the sight and taste of food. Exp. Neurol 51, 668–677. [DOI] [PubMed] [Google Scholar]
- Buschman TJ, and Miller EK (2007). Top-Down Versus Bottom-Up Control of Attention in the Prefrontal and Posterior Parietal Cortices. Science 315, 1860–1862. [DOI] [PubMed] [Google Scholar]
- Cachope R, and Cheer JF (2014). Local control of striatal dopamine release. Front. Behav. Neurosci 8, 188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cachope R, Mateo Y, Mathur BN, Irving J, Wang H-L, Morales M, Lovinger DM, and Cheer JF (2012). Selective activation of cholinergic interneurons enhances accumbal phasic dopamine release: setting the tone for reward processing. Cell Rep. 2, 33–41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Canteras NS, and Graeff FG (2014). Executive and modulatory neural circuits of defensive reactions: Implications for panic disorder. Neurosci. Biobehav. Rev 46, 352–364. [DOI] [PubMed] [Google Scholar]
- Carboni E, Tanda GL, Frau R, and Chiara GD (1990). Blockade of the Noradrenaline Carrier Increases Extracellular Dopamine Concentrations in the Prefrontal Cortex: Evidence that Dopamine Is Taken up In Vivo by Noradrenergic Terminals. J. Neurochem 55, 1067–1070. [DOI] [PubMed] [Google Scholar]
- Cardinal RN, Parkinson JA, Hall J, and Everitt BJ (2002). Emotion and motivation: the role of the amygdala, ventral striatum, and prefrontal cortex. Neurosci. Biobehav. Rev 26, 321–352. [DOI] [PubMed] [Google Scholar]
- Carr DB, O’Donnell P, Card JP, and Sesack SR (1999). Dopamine terminals in the rat prefrontal cortex synapse on pyramidal cells that project to the nucleus accumbens. J. Neurosci. Off. J. Soc. Neurosci 19, 11049–11060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cass WA, and Gerhardt GA (1995). In vivo assessment of dopamine uptake in rat medial prefrontal cortex: comparison with dorsal striatum and nucleus accumbens. J. Neurochem 65, 201–207. [DOI] [PubMed] [Google Scholar]
- Castillo Díaz F, Kramar CP, Hernandez MA, and Medina JH (2017). Activation of D1/5 Dopamine Receptors in the Dorsal Medial Prefrontal Cortex Promotes Incubated-Like Aversive Responses. Front. Behav. Neurosci 11, 209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Castro DC, and Berridge KC (2014). Opioid hedonic hotspot in nucleus accumbens shell: mu, delta, and kappa maps for enhancement of sweetness “liking” and “wanting.” J. Neurosci. Off. J. Soc. Neurosci 34, 4239–4250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chan CS, Peterson JD, Gertler TS, Glajch KE, Quintana RE, Cui Q, Sebel LE, Plotkin JL, Shen W, Heiman M, et al. (2012). Strain-specific regulation of striatal phenotype in Drd2-eGFP BAC transgenic mice. J. Neurosci. Off. J. Soc. Neurosci 32, 9124–9132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chandler DJ, Lamperski CS, and Waterhouse BD (2013). Identification and distribution of projections from monoaminergic and cholinergic nuclei to functionally differentiated subregions of prefrontal cortex. Brain Res. 1522, 38–58. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen T-W, Wardill TJ, Sun Y, Pulver SR, Renninger SL, Baohan A, Schreiter ER, Kerr RA, Orger MB, Jayaraman V, et al. (2013). Ultrasensitive fluorescent proteins for imaging neuronal activity. Nature 499, 295–300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chiodo LA, Bannon MJ, Grace AA, Roth RH, and Bunney BS (1984). Evidence for the absence of impulse-regulating somatodendritic and synthesis-modulating nerve terminal autoreceptors on subpopulations of mesocortical dopamine neurons. Neuroscience 12, 1–16. [DOI] [PubMed] [Google Scholar]
- Cho JR, Treweek JB, Robinson JE, Xiao C, Bremner LR, Greenbaum A, and Gradinaru V (2017). Dorsal Raphe Dopamine Neurons Modulate Arousal and Promote Wakefulness by Salient Stimuli. Neuron 94, 1205–1219.e8. [DOI] [PubMed] [Google Scholar]
- Christie MJ, Bridge S, James LB, and Beart PM (1985). Excitotoxin lesions suggest an aspartatergic projection from rat medial prefrontal cortex to ventral tegmental area. Brain Res. 333, 169–172. [DOI] [PubMed] [Google Scholar]
- Clarkson RL, Liptak AT, Gee SM, Sohal VS, and Bender KJ (2017). D3 Receptors Regulate Excitability in a Unique Class of Prefrontal Pyramidal Cells. J. Neurosci. Off. J. Soc. Neurosci 37, 5846–5860. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cohen JD, and Servan-Schreiber D (1992). Context, cortex, and dopamine: A connectionist approach to behavior and biology in schizophrenia. Psychol. Rev 99, 45–77. [DOI] [PubMed] [Google Scholar]
- Cohen JD, Braver TS, and Brown JW (2002). Computational perspectives on dopamine function in prefrontal cortex. Curr. Opin. Neurobiol 12, 223–229. [DOI] [PubMed] [Google Scholar]
- Collins DP, Anastasiades PG, Marlin JJ, and Carter AG (2018). Reciprocal Circuits Linking the Prefrontal Cortex with Dorsal and Ventral Thalamic Nuclei. Neuron 98, 366–379.e4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Courtin J, Chaudun F, Rozeske RR, Karalis N, Gonzalez-Campo C, Wurtz H, Abdi A, Baufreton J, Bienvenu TCM, and Herry C (2014). Prefrontal parvalbumin interneurons shape neuronal activity to drive fear expression. Nature 505, 92–96. [DOI] [PubMed] [Google Scholar]
- Covey DP, Roitman MF, and Garris PA (2014). Illicit dopamine transients: reconciling actions of abused drugs. Trends Neurosci. 37, 200–210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cragg SJ, and Rice ME (2004). DAncing past the DAT at a DA synapse. Trends Neurosci. 27, 270–277. [DOI] [PubMed] [Google Scholar]
- Day JJ, and Carelli RM (2007). The nucleus accumbens and Pavlovian reward learning. Neurosci. Rev. J. Bringing Neurobiol. Neurol. Psychiatry 13, 148–159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Day JJ, Roitman MF, Wightman RM, and Carelli RM (2007). Associative learning mediates dynamic shifts in dopamine signaling in the nucleus accumbens. Nat. Neurosci 10, 1020–1028. [DOI] [PubMed] [Google Scholar]
- De Boer SF, and Koolhaas JM (2003). Defensive burying in rodents: ethology, neurobiology and psychopharmacology. Eur. J. Pharmacol 463, 145–161. [DOI] [PubMed] [Google Scholar]
- Decot HK, Namboodiri VMK, Gao W, McHenry JA, Jennings JH, Lee S-H, Kantak PA, Jill Kao Y-C, Das M, Witten IB, et al. (2017). Coordination of Brain-Wide Activity Dynamics by Dopaminergic Neurons. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol 42, 615–627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- DeFelipe J, and Fariñas I (1992). The pyramidal neuron of the cerebral cortex: morphological and chemical characteristics of the synaptic inputs. Prog. Neurobiol 39, 563–607. [DOI] [PubMed] [Google Scholar]
- Deng H, Xiao X, and Wang Z (2016). Periaqueductal Gray Neuronal Activities Underlie Different Aspects of Defensive Behaviors. J. Neurosci. Off. J. Soc. Neurosci 36, 7580–7588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deroche V, Le Moal M, and Piazza PV (1999). Cocaine self-administration increases the incentive motivational properties of the drug in rats. Eur. J. Neurosci 11, 2731–2736. [DOI] [PubMed] [Google Scholar]
- Descarries L, Lemay B, Doucet G, and Berger B (1987). Regional and laminar density of the dopamine innervation in adult rat cerebral cortex. Neuroscience 21, 807–824. [DOI] [PubMed] [Google Scholar]
- Deutch AY, and Young CD (1995). A model of the stress-induced activation of prefrontal cortical dopamine systems: Coping and the development of post-traumatic stress disorder In Neurobiological and Clinical Consequences of Stress: From Normal Adaptation to Post-Traumatic Stress Disorder, (Philadelphia, PA, US: Lippincott Williams & Wilkins Publishers; ), pp. 163–175. [Google Scholar]
- Dias R, Robbins TW, and Roberts AC (1996a). Dissociation in prefrontal cortex of affective and attentional shifts. Nature 380, 69–72. [DOI] [PubMed] [Google Scholar]
- Dias R, Robbins TW, and Roberts AC (1996b). Primate analogue of the Wisconsin Card Sorting Test: effects of excitotoxic lesions of the prefrontal cortex in the marmoset. Behav. Neurosci 110, 872–886. [DOI] [PubMed] [Google Scholar]
- Dias R, Robbins TW, and Roberts AC (1997). Dissociable forms of inhibitory control within prefrontal cortex with an analog of the Wisconsin Card Sort Test: restriction to novel situations and independence from “on-line” processing. J. Neurosci. Off. J. Soc. Neurosci 17, 9285–9297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Diehl MM, Bravo-Rivera C, Rodriguez-Romaguera J, Pagan-Rivera PA, Burgos-Robles A, Roman-Ortiz C, and Quirk GJ (2018). Active avoidance requires inhibitory signaling in the rodent prelimbic prefrontal cortex. ELife 7, e34657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Do-Monte FH, Quiñones-Laracuente K, and Quirk GJ (2015). A temporal shift in the circuits mediating retrieval of fear memory. Nature 519, 460–463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Donoghue JP, and Wise SP (1982). The motor cortex of the rat: Cytoarchitecture and microstimulation mapping. J. Comp. Neurol 212, 76–88. [DOI] [PubMed] [Google Scholar]
- Douglas RJ, and Martin KAC (2004). Neuronal circuits of the neocortex. Annu. Rev. Neurosci 27, 419–451. [DOI] [PubMed] [Google Scholar]
- Dreyer JK, and Hounsgaard J (2013). Mathematical model of dopamine autoreceptors and uptake inhibitors and their influence on tonic and phasic dopamine signaling. J. Neurophysiol 109, 171–182. [DOI] [PubMed] [Google Scholar]
- Dreyer JK, Herrik KF, Berg RW, and Hounsgaard JD (2010). Influence of phasic and tonic dopamine release on receptor activation. J. Neurosci. Off. J. Soc. Neurosci 30, 14273–14283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dreyer JK, Vander Weele CM, Lovic V, and Aragona BJ (2016). Functionally Distinct Dopamine Signals in Nucleus Accumbens Core and Shell in the Freely Moving Rat. J. Neurosci. Off. J. Soc. Neurosci 36, 98–112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dubuisson D, and Dennis SG (1977). The formalin test: a quantitative study of the analgesic effects of morphine, meperidine, and brain stem stimulation in rats and cats. Pain 4, 161–174. [DOI] [PubMed] [Google Scholar]
- Dunbar RIM (2009). The social brain hypothesis and its implications for social evolution. Ann. Hum. Biol 36, 562–572. [DOI] [PubMed] [Google Scholar]
- Durstewitz D, and Seamans JK (2008). The dual-state theory of prefrontal cortex dopamine function with relevance to catechol-o-methyltransferase genotypes and schizophrenia. Biol. Psychiatry 64, 739–749. [DOI] [PubMed] [Google Scholar]
- Ellwood IT, Patel T, Wadia V, Lee AT, Liptak AT, Bender KJ, and Sohal VS (2017). Tonic or Phasic Stimulation of Dopaminergic Projections to Prefrontal Cortex Causes Mice to Maintain or Deviate from Previously Learned Behavioral Strategies. J. Neurosci. Off. J. Soc. Neurosci 37, 8315–8329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Euston DR, Gruber AJ, and McNaughton BL (2012). The role of medial prefrontal cortex in memory and decision making. Neuron 76, 1057–1070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Evans DA, Stempel AV, Vale R, Ruehle S, Lefler Y, and Branco T (2018). A synaptic threshold mechanism for computing escape decisions. Nature 1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fallon JH (1981). Collateralization of monoamine neurons: mesotelencephalic dopamine projections to caudate, septum, and frontal cortex. J. Neurosci. Off. J. Soc. Neurosci 1, 1361–1368. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fanselow MS (1991). The Midbrain Periaqueductal Gray as a Coordinator of Action in Response to Fear and Anxiety In The Midbrain Periaqueductal Gray Matter, (Springer, Boston, MA: ), pp. 151–173. [Google Scholar]
- Felix-Ortiz AC, Burgos-Robles A, Bhagat ND, Leppla CA, and Tye KM (2016). Bidirectional modulation of anxiety-related and social behaviors by amygdala projections to the medial prefrontal cortex. Neuroscience 321, 197–209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ferris MJ, España RA, Locke JL, Konstantopoulos JK, Rose JH, Chen R, and Jones SR (2014). Dopamine transporters govern diurnal variation in extracellular dopamine tone. Proc. Natl. Acad. Sci 201407935. [DOI] [PMC free article] [PubMed]
- Finlay JM, Zigmond MJ, and Abercrombie ED (1995). Increased dopamine and norepinephrine release in medial prefrontal cortex induced by acute and chronic stress: effects of diazepam. Neuroscience 64, 619–628. [DOI] [PubMed] [Google Scholar]
- Floresco SB (2013). Prefrontal dopamine and behavioral flexibility: shifting from an “inverted-U” toward a family of functions. Front. Neurosci 7, 62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Floresco SB, and Magyar O (2006). Mesocortical dopamine modulation of executive functions: beyond working memory. Psychopharmacology (Berl.) 188, 567–585. [DOI] [PubMed] [Google Scholar]
- Frankland PW, Bontempi B, Talton LE, Kaczmarek L, and Silva AJ (2004). The Involvement of the Anterior Cingulate Cortex in Remote Contextual Fear Memory. Science 304, 881–883. [DOI] [PubMed] [Google Scholar]
- Franklin TB, Silva BA, Perova Z, Marrone L, Masferrer ME, Zhan Y, Kaplan A, Greetham L, Verrechia V, Halman A, et al. (2017). Prefrontal cortical control of a brainstem social behavior circuit. Nat. Neurosci 20, 260–270. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gabbott PL, Dickie BG, Vaid RR, Headlam AJ, and Bacon SJ (1997). Local-circuit neurones in the medial prefrontal cortex (areas 25, 32 and 24b) in the rat: morphology and quantitative distribution. J. Comp. Neurol 377, 465–499. [DOI] [PubMed] [Google Scholar]
- Gabbott PLA, Warner TA, Jays PRL, Salway P, and Busby SJ (2005). Prefrontal cortex in the rat: projections to subcortical autonomic, motor, and limbic centers. J. Comp. Neurol 492, 145–177. [DOI] [PubMed] [Google Scholar]
- Garris PA, and Wightman RM (1994). Different kinetics govern dopaminergic transmission in the amygdala, prefrontal cortex, and striatum: an in vivo voltammetric study. J. Neurosci 14, 442–450. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garris PA, Collins LB, Jones SR, and Wightman RM (1993). Evoked extracellular dopamine in vivo in the medial prefrontal cortex. J. Neurochem 61, 637–647. [DOI] [PubMed] [Google Scholar]
- Gaspar P, Bloch B, and Le Moine C (1995). D1 and D2 receptor gene expression in the rat frontal cortex: cellular localization in different classes of efferent neurons. Eur. J. Neurosci 7, 1050–1063. [DOI] [PubMed] [Google Scholar]
- Gee S, Ellwood I, Patel T, Luongo F, Deisseroth K, and Sohal VS (2012). Synaptic activity unmasks dopamine D2 receptor modulation of a specific class of layer V pyramidal neurons in prefrontal cortex. J. Neurosci. Off. J. Soc. Neurosci 32, 4959–4971. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gehring WJ, and Knight RT (2000). Prefrontal–cingulate interactions in action monitoring. Nat. Neurosci 3, 516–520. [DOI] [PubMed] [Google Scholar]
- George MS, Anton RF, Bloomer C, Teneback C, Drobes DJ, Lorberbaum JP, Nahas Z, and Vincent DJ (2001). Activation of prefrontal cortex and anterior thalamus in alcoholic subjects on exposure to alcohol-specific cues. Arch. Gen. Psychiatry 58, 345–352. [DOI] [PubMed] [Google Scholar]
- Gerfen CR, and Surmeier DJ (2011). Modulation of striatal projection systems by dopamine. Annu. Rev. Neurosci 34, 441–466. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goeders NE, and Smith JE (1983). Cortical dopaminergic involvement in cocaine reinforcement. Science 221, 773–775. [DOI] [PubMed] [Google Scholar]
- Goldman-Rakic PS, Leranth C, Williams SM, Mons N, and Geffard M (1989). Dopamine synaptic complex with pyramidal neurons in primate cerebral cortex. Proc. Natl. Acad. Sci. U. S. A 86, 9015–9019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- González-Burgos G, Kröner S, Krimer LS, Seamans JK, Urban NN, Henze DA, Lewis DA, and Barrionuevo G (2002). Dopamine modulation of neuronal function in the monkey prefrontal cortex. Physiol. Behav 77, 537–543. [DOI] [PubMed] [Google Scholar]
- Gorelova NA, and Yang CR (2000). Dopamine D1/D5 receptor activation modulates a persistent sodium current in rat prefrontal cortical neurons in vitro. J. Neurophysiol 84, 75–87. [DOI] [PubMed] [Google Scholar]
- Grace AA (2016). Dysregulation of the dopamine system in the pathophysiology of schizophrenia and depression. Nat. Rev. Neurosci 17, 524–532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grace AA, and Bunney BS (1984a). The control of firing pattern in nigral dopamine neurons: burst firing. J. Neurosci. Off. J. Soc. Neurosci 4, 2877–2890. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grace AA, and Bunney BS (1984b). The control of firing pattern in nigral dopamine neurons: single spike firing. J. Neurosci. Off. J. Soc. Neurosci 4, 2866–2876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grace AA, Floresco SB, Goto Y, and Lodge DJ (2007). Regulation of firing of dopaminergic neurons and control of goal-directed behaviors. Trends Neurosci. 30, 220–227. [DOI] [PubMed] [Google Scholar]
- Granon S, Passetti F, Thomas KL, Dalley JW, Everitt BJ, and Robbins TW (2000). Enhanced and impaired attentional performance after infusion of D1 dopaminergic receptor agents into rat prefrontal cortex. J. Neurosci. Off. J. Soc. Neurosci 20, 1208–1215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gregoriou GG, Rossi AF, Ungerleider LG, and Desimone R (2014). Lesions of prefrontal cortex reduce attentional modulation of neuronal responses and synchrony in V4. Nat. Neurosci 17, 1003–1011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Greicius MD, Flores BH, Menon V, Glover GH, Solvason HB, Kenna H, Reiss AL, and Schatzberg AF (2007). Resting-State Functional Connectivity in Major Depression: Abnormally Increased Contributions from Subgenual Cingulate Cortex and Thalamus. Biol. Psychiatry 62, 429–437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Groenewegen HJ, and Uylings HB (2000). The prefrontal cortex and the integration of sensory, limbic and autonomic information. Prog. Brain Res 126, 3–28. [DOI] [PubMed] [Google Scholar]
- Groenewegen HJ, Wright CI, and Uylings HB (1997). The anatomical relationships of the prefrontal cortex with limbic structures and the basal ganglia. J. Psychopharmacol. Oxf. Engl 11, 99–106. [DOI] [PubMed] [Google Scholar]
- Gulledge AT, and Jaffe DB (1998). Dopamine decreases the excitability of layer V pyramidal cells in the rat prefrontal cortex. J. Neurosci. Off. J. Soc. Neurosci 18, 9139–9151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gulledge AT, and Jaffe DB (2001). Multiple effects of dopamine on layer V pyramidal cell excitability in rat prefrontal cortex. J. Neurophysiol 86, 586–595. [DOI] [PubMed] [Google Scholar]
- Gunaydin LA, Grosenick L, Finkelstein JC, Kauvar IV, Fenno LE, Adhikari A, Lammel S, Mirzabekov JJ, Airan RD, Zalocusky KA, et al. (2014). Natural neural projection dynamics underlying social behavior. Cell 157, 1535–1551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haber SN, and Fudge JL (1997). The primate substantia nigra and VTA: integrative circuitry and function. Crit. Rev. Neurobiol 11, 323–342. [DOI] [PubMed] [Google Scholar]
- Halladay LR, and Blair HT (2015). Distinct ensembles of medial prefrontal cortex neurons are activated by threatening stimuli that elicit excitation vs. inhibition of movement. J. Neurophysiol 114, 793–807. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hamid AA, Pettibone JR, Mabrouk OS, Hetrick VL, Schmidt R, Vander Weele CM, Kennedy RT, Aragona BJ, and Berke JD (2016). Mesolimbic dopamine signals the value of work. Nat. Neurosci 19, 117–126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Han S-W, Kim Y-C, and Narayanan NS (2017). Projection targets of medial frontal D1DR-expressing neurons. Neurosci. Lett 655, 166–171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heidbreder CA, and Groenewegen HJ (2003). The medial prefrontal cortex in the rat: evidence for a dorso-ventral distinction based upon functional and anatomical characteristics. Neurosci. Biobehav. Rev 27, 555–579. [DOI] [PubMed] [Google Scholar]
- Heien MLAV, Phillips PEM, Stuber GD, Seipel AT, and Wightman RM (2003). Overoxidation of carbon-fiber microelectrodes enhances dopamine adsorption and increases sensitivity. The Analyst 128, 1413–1419. [DOI] [PubMed] [Google Scholar]
- Henze DA, González-Burgos GR, Urban NN, Lewis DA, and Barrionuevo G (2000). Dopamine increases excitability of pyramidal neurons in primate prefrontal cortex. J. Neurophysiol 84, 2799–2809. [DOI] [PubMed] [Google Scholar]
- Hernandez L, and Hoebel BG (1990). Feeding can enhance dopamine turnover in the prefrontal cortex. Brain Res. Bull 25, 975–979. [DOI] [PubMed] [Google Scholar]
- Hoover WB, and Vertes RP (2007). Anatomical analysis of afferent projections to the medial prefrontal cortex in the rat. Brain Struct. Funct 212, 149–179. [DOI] [PubMed] [Google Scholar]
- Horvitz JC (2000). Mesolimbocortical and nigrostriatal dopamine responses to salient non-reward events. Neuroscience 96, 651–656. [DOI] [PubMed] [Google Scholar]
- Howes OD, and Kapur S (2009). The dopamine hypothesis of schizophrenia: version III--the final common pathway. Schizophr. Bull 35, 549–562. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ikemoto S (2007). Dopamine reward circuitry: two projection systems from the ventral midbrain to the nucleus accumbens-olfactory tubercle complex. Brain Res. Rev 56, 27–78. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ikemoto S (2010). Brain reward circuitry beyond the mesolimbic dopamine system: A neurobiological theory. Neurosci. Biobehav. Rev 35, 129–150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Isaacson JS, and Scanziani M (2011). How inhibition shapes cortical activity. Neuron 72, 231–243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ishikawa A, Ambroggi F, Nicola SM, and Fields HL (2008). Dorsomedial prefrontal cortex contribution to behavioral and nucleus accumbens neuronal responses to incentive cues. J. Neurosci. Off. J. Soc. Neurosci 28, 5088–5098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Janak PH, and Tye KM (2015). From circuits to behaviour in the amygdala. Nature 517, 284–292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jennings JH, Ung RL, Resendez SL, Stamatakis AM, Taylor JG, Huang J, Veleta K, Kantak PA, Aita M, Shilling-Scrivo K, et al. (2015). Visualizing hypothalamic network dynamics for appetitive and consummatory behaviors. Cell 160, 516–527. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jhang J, Lee H, Kang MS, Lee H-S, Park H, and Han J-H (2018). Anterior cingulate cortex and its input to the basolateral amygdala control innate fear response. Nat. Commun 9, 2744. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johansen JP, and Fields HL (2004). Glutamatergic activation of anterior cingulate cortex produces an aversive teaching signal. Nat. Neurosci 7, 398–403. [DOI] [PubMed] [Google Scholar]
- Karalis N, Dejean C, Chaudun F, Khoder S, Rozeske RR, Wurtz H, Bagur S, Benchenane K, Sirota A, Courtin J, et al. (2016). 4-Hz oscillations synchronize prefrontal–amygdala circuits during fear behavior. Nat. Neurosci 19, 605–612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karoum F, Chrapusta SJ, and Egan MF (1994). 3-Methoxytyramine Is the Major Metabolite of Released Dopamine in the Rat Frontal Cortex: Reassessment of the Effects of Antipsychotics on the Dynamics of Dopamine Release and Metabolism in the Frontal Cortex, Nucleus Accumbens, and Striatum by a Simple Two Pool Model. J. Neurochem 63, 972–979. [DOI] [PubMed] [Google Scholar]
- Kerney M, Smaers JB, Schoenemann PT, and Dunn JC (2017). The coevolution of play and the cortico-cerebellar system in primates. Primates J. Primatol 58, 485–491. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khan ZU, Gutiérrez A, Martín R, Peñafiel A, Rivera A, and De La Calle A (1998). Differential regional and cellular distribution of dopamine D2-like receptors: an immunocytochemical study of subtype-specific antibodies in rat and human brain. J. Comp. Neurol 402, 353–371. [DOI] [PubMed] [Google Scholar]
- Kim CK, Yang SJ, Pichamoorthy N, Young NP, Kauvar I, Jennings JH, Lerner TN, Berndt A, Lee SY, Ramakrishnan C, et al. (2016a). Simultaneous fast measurement of circuit dynamics at multiple sites across the mammalian brain. Nat. Methods 13, 325–328. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim J, Pignatelli M, Xu S, Itohara S, and Tonegawa S (2016b). Antagonistic negative and positive neurons of the basolateral amygdala. Nat. Neurosci 19, 1636–1646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koenigs M, and Grafman J (2009). The functional neuroanatomy of depression: Distinct roles for ventromedial and dorsolateral prefrontal cortex. Behav. Brain Res 201, 239–243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kole MHP, and Stuart GJ (2012). Signal processing in the axon initial segment. Neuron 73, 235–247. [DOI] [PubMed] [Google Scholar]
- Kravitz AV, Tye LD, and Kreitzer AC (2012). Distinct roles for direct and indirect pathway striatal neurons in reinforcement. Nat. Neurosci 15, 816–818. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kritzer MF, and Goldman-Rakic PS (1995). Intrinsic circuit organization of the major layers and sublayers of the dorsolateral prefrontal cortex in the rhesus monkey. J. Comp. Neurol 359, 131–143. [DOI] [PubMed] [Google Scholar]
- Kroener S, Chandler LJ, Phillips PEM, and Seamans JK (2009). Dopamine modulates persistent synaptic activity and enhances the signal-to-noise ratio in the prefrontal cortex. PloS One 4, e6507. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kupchik YM, Brown RM, Heinsbroek JA, Lobo MK, Schwartz DJ, and Kalivas PW (2015). Coding the direct/indirect pathways by D1 and D2 receptors is not valid for accumbens projections. Nat. Neurosci 18, 1230–1232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kvitsiani D, Ranade S, Hangya B, Taniguchi H, Huang JZ, and Kepecs A (2013). Distinct behavioural and network correlates of two interneuron types in prefrontal cortex. Nature 498, 363–366. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lammel S, Hetzel A, Häckel O, Jones I, Liss B, and Roeper J (2008). Unique properties of mesoprefrontal neurons within a dual mesocorticolimbic dopamine system. Neuron 57, 760–773. [DOI] [PubMed] [Google Scholar]
- Lammel S, Ion DI, Roeper J, and Malenka RC (2011). Projection-specific modulation of dopamine neuron synapses by aversive and rewarding stimuli. Neuron 70, 855–862. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lammel S, Lim BK, Ran C, Huang KW, Betley MJ, Tye KM, Deisseroth K, and Malenka RC (2012). Input-specific control of reward and aversion in the ventral tegmental area. Nature 491, 212–217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lammel S, Lim BK, and Malenka RC (2014). Reward and aversion in a heterogeneous midbrain dopamine system. Neuropharmacology 76 Pt B, 351–359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Land BB, Narayanan NS, Liu R-J, Gianessi CA, Brayton CE, Grimaldi DM, Sarhan M, Guarnieri DJ, Deisseroth K, Aghajanian GK, et al. (2014). Medial prefrontal D1 dopamine neurons control food intake. Nat. Neurosci 17, 248–253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laubach M, Caetano MS, and Narayanan NS (2015). Mistakes were made: Neural mechanisms for the adaptive control of action initiation by the medial prefrontal cortex. J. Physiol.-Paris 109, 104–117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laubach M, Amarante L, Swanson K, and White SR (2018). What, if anything, is rodent prefrontal cortex? PsyArXiv. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee AT, Vogt D, Rubenstein JL, and Sohal VS (2014). A class of GABAergic neurons in the prefrontal cortex sends long-range projections to the nucleus accumbens and elicits acute avoidance behavior. J. Neurosci. Off. J. Soc. Neurosci 34, 11519–11525. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee JC, Wang LP, and Tsien JZ (2016). Dopamine Rebound-Excitation Theory: Putting Brakes on PTSD. Front. Psychiatry 7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lewis DA, and Sesack SR (1997). Chapter VI Dopamine systems in the primate brain In Handbook of Chemical Neuroanatomy, Bloom FE, Björklund A, and Hökfelt T, eds. (Elsevier; ), pp. 263–375. [Google Scholar]
- Li C, Sugam JA, Lowery-Gionta EG, McElligott ZA, McCall NM, Lopez AJ, McKlveen JM, Pleil KE, and Kash TL (2016). Mu Opioid Receptor Modulation of Dopamine Neurons in the Periaqueductal Gray/Dorsal Raphe: A Role in Regulation of Pain. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol 41, 2122–2132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin D, Boyle MP, Dollar P, Lee H, Lein ES, Perona P, and Anderson DJ (2011). Functional identification of an aggression locus in the mouse hypothalamus. Nature 470, 221–226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lindström LH, Gefvert O, Hagberg G, Lundberg T, Bergström M, Hartvig P, and Långström B (1999). Increased dopamine synthesis rate in medial prefrontal cortex and striatum in schizophrenia indicated by L-(β−11C) DOPA and PET. Biol. Psychiatry 46, 681–688. [DOI] [PubMed] [Google Scholar]
- Maisonneuve IM, Keller RW, and Glick SD (1990). Similar effects ofd-amphetamine and cocaine on extracellular dopamine levels in medial prefrontal cortex of rats. Brain Res. 535, 221–226. [DOI] [PubMed] [Google Scholar]
- Mansouri FA, Tanaka K, and Buckley MJ (2009). Conflict-induced behavioural adjustment: a clue to the executive functions of the prefrontal cortex. Nat. Rev. Neurosci 10, 141–152. [DOI] [PubMed] [Google Scholar]
- Mantz J, Thierry AM, and Glowinski J (1989). Effect of noxious tail pinch on the discharge rate of mesocortical and mesolimbic dopamine neurons: selective activation of the mesocortical system. Brain Res. 476, 377–381. [DOI] [PubMed] [Google Scholar]
- Markram H, Toledo-Rodriguez M, Wang Y, Gupta A, Silberberg G, and Wu C (2004). Interneurons of the neocortical inhibitory system. Nat. Rev. Neurosci 5, 793–807. [DOI] [PubMed] [Google Scholar]
- Marton T, Seifikar H, Luongo FJ, Lee AT, and Sohal VS (2018). Roles of prefrontal cortex and mediodorsal thalamus in task engagement and behavioral flexibility. J. Neurosci. Off. J. Soc. Neurosci [DOI] [PMC free article] [PubMed]
- Matsumoto M, and Hikosaka O (2009). Two types of dopamine neuron distinctly convey positive and negative motivational signals. Nature 459, 837–841. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matsumoto H, Tian J, Uchida N, and Watabe-Uchida M Midbrain dopamine neurons signal aversion in a reward-context-dependent manner. ELife 5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matthews GA, Nieh EH, Vander Weele CM, Halbert SA, Pradhan RV, Yosafat AS, Glober GF, Izadmehr EM, Thomas RE, Lacy GD, et al. (2016). Dorsal Raphe Dopamine Neurons Represent the Experience of Social Isolation. Cell 164, 617–631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McCutcheon JE, Ebner SR, Loriaux AL, and Roitman MF (2012). Encoding of aversion by dopamine and the nucleus accumbens. Front. Neurosci 6, 137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McGarry LM, and Carter AG (2017). Prefrontal Cortex Drives Distinct Projection Neurons in the Basolateral Amygdala. Cell Rep. 21, 1426–1433. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McGregor A, Baker G, and Roberts DCS (1996). Effect of 6-hydroxydopamine lesions of the medial prefrontal cortex on intravenous cocaine self-administration under a progressive ratio schedule of reinforcement. Pharmacol. Biochem. Behav 53, 5–9. [DOI] [PubMed] [Google Scholar]
- Menegas W, Bergan JF, Ogawa SK, Isogai Y, Umadevi Venkataraju K, Osten P, Uchida N, and Watabe-Uchida M (2015). Dopamine neurons projecting to the posterior striatum form an anatomically distinct subclass. ELife 4, e10032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meyer CT, Padmala S, and Pessoa L (2017). Tracking dynamic threat imminence. BioRxiv 183798. [Google Scholar]
- Michel PP, Hirsch EC, and Hunot S (2016). Understanding Dopaminergic Cell Death Pathways in Parkinson Disease. Neuron 90, 675–691. [DOI] [PubMed] [Google Scholar]
- Milham MP, Banich MT, Webb A, Barad V, Cohen NJ, Wszalek T, and Kramer AF (2001). The relative involvement of anterior cingulate and prefrontal cortex in attentional control depends on nature of conflict. Brain Res. Cogn. Brain Res 12, 467–473. [DOI] [PubMed] [Google Scholar]
- Miller EK (1999). The Prefrontal Cortex: Complex Neural Properties for Complex Behavior. Neuron 22, 15–17. [DOI] [PubMed] [Google Scholar]
- Miller EK (2000). The prefontral cortex and cognitive control. Nat. Rev. Neurosci 1, 59–65. [DOI] [PubMed] [Google Scholar]
- Miller EK, and Cohen JD (2001). An Integrative Theory of Prefrontal Cortex Function. Annu. Rev. Neurosci 24, 167–202. [DOI] [PubMed] [Google Scholar]
- Milner B (1963). Effects of Different Brain Lesions on Card Sorting: The Role of the Frontal Lobes. Arch. Neurol 9, 90–100. [Google Scholar]
- Miner LH, Schroeter S, Blakely RD, and Sesack SR (2003). Ultrastructural localization of the norepinephrine transporter in superficial and deep layers of the rat prelimbic prefrontal cortex and its spatial relationship to probable dopamine terminals. J. Comp. Neurol 466, 478–494. [DOI] [PubMed] [Google Scholar]
- Mobbs D, Petrovic P, Marchant JL, Hassabis D, Weiskopf N, Seymour B, Dolan RJ, and Frith CD (2007). When fear is near: threat imminence elicits prefrontal-periaqueductal gray shifts in humans. Science 317, 1079–1083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mobbs D, Yu R, Rowe JB, Eich H, FeldmanHall O, and Dalgleish T (2010). Neural activity associated with monitoring the oscillating threat value of a tarantula. Proc. Natl. Acad. Sci. U. S. A 107, 20582–20586. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mohebi A, Pettibone J, Hamid A, Wong J-M, Kennedy R, and Berke J (2018). Forebrain dopamine value signals arise independently from midbrain dopamine cell firing. BioRxiv 334060. [Google Scholar]
- Muly EC, Szigeti K, and Goldman-Rakic PS (1998). D1 receptor in interneurons of macaque prefrontal cortex: distribution and subcellular localization. J. Neurosci. Off. J. Soc. Neurosci 18, 10553–10565. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murugan M, Jang HJ, Park M, Miller EM, Cox J, Taliaferro JP, Parker NF, Bhave V, Hur H, Liang Y, et al. (2017). Combined Social and Spatial Coding in a Descending Projection from the Prefrontal Cortex. Cell 171, 1663–1677.e16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Namburi P, Beyeler A, Yorozu S, Calhoon GG, Halbert SA, Wichmann R, Holden SS, Mertens KL, Anahtar M, Felix-Ortiz AC, et al. (2015). A circuit mechanism for differentiating positive and negative associations. Nature 520, 675–678. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nieh EH, Matthews GA, Allsop SA, Presbrey KN, Leppla CA, Wichmann R, Neve R, Wildes CP, and Tye KM (2015). Decoding neural circuits that control compulsive sucrose seeking. Cell 160, 528–541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nieh EH, Vander Weele CM, Matthews GA, Presbrey KN, Wichmann R, Leppla CA, Izadmehr EM, and Tye KM (2016). Inhibitory Input from the Lateral Hypothalamus to the Ventral Tegmental Area Disinhibits Dopamine Neurons and Promotes Behavioral Activation. Neuron 90, 1286–1298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Niv Y (2007). Cost, benefit, tonic, phasic: what do response rates tell us about dopamine and motivation? Ann. N. Y. Acad. Sci 1104, 357–376. [DOI] [PubMed] [Google Scholar]
- Nomikos GG, and Spyraki C (1988). Cocaine-induced place conditioning: importance of route of administration and other procedural variables. Psychopharmacology (Berl.) 94, 119–125. [DOI] [PubMed] [Google Scholar]
- Noonan MP, Mars RB, Sallet J, Dunbar RIM, and Fellows LK (2018). The structural and functional brain networks that support human social networks. Behav. Brain Res [DOI] [PMC free article] [PubMed]
- Noudoost B, and Moore T (2011). Control of visual cortical signals by prefrontal dopamine. Nature 474, 372–375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Okubo Y, Suhara T, Suzuki K, Kobayashi K, Inoue O, Terasaki O, Someya Y, Sassa T, Sudo Y, Matsushima E, et al. (1997). Decreased prefrontal dopamine D1 receptors in schizophrenia revealed by PET. Nature 385, 634–636. [DOI] [PubMed] [Google Scholar]
- Oleson EB, Gentry RN, Chioma VC, and Cheer JF (2012). Subsecond dopamine release in the nucleus accumbens predicts conditioned punishment and its successful avoidance. J. Neurosci. Off. J. Soc. Neurosci 32, 14804–14808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ongür D, and Price JL (2000). The organization of networks within the orbital and medial prefrontal cortex of rats, monkeys and humans. Cereb. Cortex N. Y. N 1991 10, 206–219. [DOI] [PubMed] [Google Scholar]
- Otis JM, Namboodiri VMK, Matan AM, Voets ES, Mohorn EP, Kosyk O, McHenry JA, Robinson JE, Resendez SL, Rossi MA, et al. (2017). Prefrontal cortex output circuits guide reward seeking through divergent cue encoding. Nature 543, 103–107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Owesson-White C, Belle AM, Herr NR, Peele JL, Gowrishankar P, Carelli RM, and Wightman RM (2016). Cue-Evoked Dopamine Release Rapidly Modulates D2 Neurons in the Nucleus Accumbens During Motivated Behavior. J. Neurosci. Off. J. Soc. Neurosci 36, 6011–6021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paspalas CD, and Goldman-Rakic PS (2005). Presynaptic D1 dopamine receptors in primate prefrontal cortex: target-specific expression in the glutamatergic synapse. J. Neurosci. Off. J. Soc. Neurosci 25, 1260–1267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Penit-Soria J, Audinat E, and Crepel F (1987). Excitation of rat prefrontal cortical neurons by dopamine: an in vitro electrophysiological study. Brain Res. 425, 263–274. [DOI] [PubMed] [Google Scholar]
- Phillips PEM, Stuber GD, Heien MLAV, Wightman RM, and Carelli RM (2003). Subsecond dopamine release promotes cocaine seeking. Nature 422, 614–618. [DOI] [PubMed] [Google Scholar]
- Pinto A, and Sesack SR (2008). Ultrastructural analysis of prefrontal cortical inputs to the rat amygdala: spatial relationships to presumed dopamine axons and D1 and D2 receptors. Brain Struct. Funct 213, 159–175. [DOI] [PubMed] [Google Scholar]
- Pitman RK, Rasmusson AM, Koenen KC, Shin LM, Orr SP, Gilbertson MW, Milad MR, and Liberzon I (2012). Biological studies of post-traumatic stress disorder. Nat. Rev. Neurosci 13, 769–787. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Popescu AT, Zhou MR, and Poo M-M (2016). Phasic dopamine release in the medial prefrontal cortex enhances stimulus discrimination. Proc. Natl. Acad. Sci. U. S. A 113, E3169–3176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ragozzino ME (2002). The Effects of Dopamine D1 Receptor Blockade in the Prelimbic–Infralimbic Areas on Behavioral Flexibility. Learn. Mem 9, 18–28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ramanathan KR, Jin J, Giustino TF, Payne MR, and Maren S (2018). Prefrontal projections to the thalamic nucleus reuniens mediate fear extinction. Nat. Commun 9, 4527. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ray JP, and Price JL (1993). The organization of projections from the mediodorsal nucleus of the thalamus to orbital and medial prefrontal cortex in macaque monkeys. J. Comp. Neurol 337, 1–31. [DOI] [PubMed] [Google Scholar]
- Rey CD, Lipps J, and Shansky RM (2014). Dopamine D1 Receptor Activation Rescues Extinction Impairments in Low-Estrogen Female Rats and Induces Cortical Layer-Specific Activation Changes in Prefrontal–Amygdala Circuits. Neuropsychopharmacology 39, 1282–1289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ridderinkhof KR, van den Wildenberg WPM, Segalowitz SJ, and Carter CS (2004). Neurocognitive mechanisms of cognitive control: The role of prefrontal cortex in action selection, response inhibition, performance monitoring, and reward-based learning. Brain Cogn. 56, 129–140. [DOI] [PubMed] [Google Scholar]
- Roberts DC, Corcoran ME, and Fibiger HC (1977). On the role of ascending catecholaminergic systems in intravenous self-administration of cocaine. Pharmacol. Biochem. Behav 6, 615–620. [DOI] [PubMed] [Google Scholar]
- Robinson SE, and Sohal VS (2017). Dopamine D2 receptors modulate pyramidal neurons in mouse medial prefrontal cortex through a stimulatory G protein pathway. J. Neurosci 1893–17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robinson DL, Venton BJ, Heien MLAV, and Wightman RM (2003). Detecting Subsecond Dopamine Release with Fast-Scan Cyclic Voltammetry in Vivo. Clin. Chem 49, 1763–1773. [DOI] [PubMed] [Google Scholar]
- Roitman MF, Wheeler RA, Wightman RM, and Carelli RM (2008). Real-time chemical responses in the nucleus accumbens differentiate rewarding and aversive stimuli. Nat. Neurosci 11, 1376–1377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rolls ET, Loh M, Deco G, and Winterer G (2008). Computational models of schizophrenia and dopamine modulation in the prefrontal cortex. Nat. Rev. Neurosci 9, 696–709. [DOI] [PubMed] [Google Scholar]
- Romanski LM, Bates JF, and Goldman-Rakic PS (1999a). Auditory belt and parabelt projections to the prefrontal cortex in the rhesus monkey. J. Comp. Neurol 403, 141–157. [DOI] [PubMed] [Google Scholar]
- Romanski LM, Tian B, Fritz J, Mishkin M, Goldman-Rakic PS, and Rauschecker JP (1999b). Dual streams of auditory afferents target multiple domains in the primate prefrontal cortex. Nat. Neurosci 2, 1131–1136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosenkranz JA, and Grace AA (2001). Dopamine Attenuates Prefrontal Cortical Suppression of Sensory Inputs to the Basolateral Amygdala of Rats. J. Neurosci 21, 4090–4103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosenkranz JA, and Grace AA (2002). Cellular Mechanisms of Infralimbic and Prelimbic Prefrontal Cortical Inhibition and Dopaminergic Modulation of Basolateral Amygdala Neurons In Vivo. J. Neurosci 22, 324–337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rossi AF, Bichot NP, Desimone R, and Ungerleider LG (2007). Top down attentional deficits in macaques with lesions of lateral prefrontal cortex. J. Neurosci. Off. J. Soc. Neurosci 27, 11306–11314. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rossi AF, Pessoa L, Desimone R, and Ungerleider LG (2009). The prefrontal cortex and the executive control of attention. Exp. Brain Res 192, 489–497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rozeske RR, and Herry C (2018). Neuronal coding mechanisms mediating fear behavior. Curr. Opin. Neurobiol 52, 60–64. [DOI] [PubMed] [Google Scholar]
- Rozeske RR, Jercog D, Karalis N, Chaudun F, Khoder S, Girard D, Winke N, and Herry C (2018). Prefrontal-Periaqueductal Gray-Projecting Neurons Mediate Context Fear Discrimination. Neuron 97, 898–910.e6. [DOI] [PubMed] [Google Scholar]
- Rubenstein JLR, and Merzenich MM (2003). Model of autism: increased ratio of excitation/inhibition in key neural systems. Genes Brain Behav. 2, 255–267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Salamone JD (1994). The involvement of nucleus accumbens dopamine in appetitive and aversive motivation. Behav. Brain Res 61, 117–133. [DOI] [PubMed] [Google Scholar]
- Salamone JD, Correa M, Farrar A, and Mingote SM (2007). Effort-related functions of nucleus accumbens dopamine and associated forebrain circuits. Psychopharmacology (Berl.) 191, 461–482. [DOI] [PubMed] [Google Scholar]
- Sangha S, Robinson PD, Greba Q, Davies DA, and Howland JG (2014). Alterations in reward, fear and safety cue discrimination after inactivation of the rat prelimbic and infralimbic cortices. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol 39, 2405–2413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Santana N, Mengod G, and Artigas F (2009). Quantitative analysis of the expression of dopamine D1 and D2 receptors in pyramidal and GABAergic neurons of the rat prefrontal cortex. Cereb. Cortex N. Y. N 1991 19, 849–860. [DOI] [PubMed] [Google Scholar]
- Satpute AB, Wager TD, Cohen-Adad J, Bianciardi M, Choi J-K, Buhle JT, Wald LL, and Barrett LF (2013). Identification of discrete functional subregions of the human periaqueductal gray. Proc. Natl. Acad. Sci. U. S. A 110, 17101–17106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sawaguchi T, and Goldman-Rakic PS (1991). D1 dopamine receptors in prefrontal cortex: involvement in working memory. Science 251, 947–950. [DOI] [PubMed] [Google Scholar]
- Schmitt LI, Wimmer RD, Nakajima M, Happ M, Mofakham S, and Halassa MM (2017). Thalamic amplification of cortical connectivity sustains attentional control. Nature 545, 219–223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schultz W, Dayan P, and Montague PR (1997). A Neural Substrate of Prediction and Reward. Science 275, 1593–1599. [DOI] [PubMed] [Google Scholar]
- Seamans JK, and Yang CR (2004). The principal features and mechanisms of dopamine modulation in the prefrontal cortex. Prog. Neurobiol 74, 1–58. [DOI] [PubMed] [Google Scholar]
- Seamans JK, Gorelova N, Durstewitz D, and Yang CR (2001a). Bidirectional dopamine modulation of GABAergic inhibition in prefrontal cortical pyramidal neurons. J. Neurosci. Off. J. Soc. Neurosci 21, 3628–3638. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seamans JK, Durstewitz D, Christie BR, Stevens CF, and Sejnowski TJ (2001b). Dopamine D1/D5 receptor modulation of excitatory synaptic inputs to layer V prefrontal cortex neurons. Proc. Natl. Acad. Sci. U. S. A 98, 301–306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Séguéla P, Watkins KC, and Descarries L (1988). Ultrastructural features of dopamine axon terminals in the anteromedial and the suprarhinal cortex of adult rat. Brain Res. 442, 11–22. [DOI] [PubMed] [Google Scholar]
- Seong HJ, and Carter AG (2012). D1 receptor modulation of action potential firing in a subpopulation of layer 5 pyramidal neurons in the prefrontal cortex. J. Neurosci. Off. J. Soc. Neurosci 32, 10516–10521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sesack SR, and Bunney BS (1989). Pharmacological characterization of the receptor mediating electrophysiological responses to dopamine in the rat medial prefrontal cortex: a microiontophoretic study. J. Pharmacol. Exp. Ther 248, 1323–1333. [PubMed] [Google Scholar]
- Sesack SR, Deutch AY, Roth RH, and Bunney BS (1989). Topographical organization of the efferent projections of the medial prefrontal cortex in the rat: an anterograde tract-tracing study with Phaseolus vulgaris leucoagglutinin. J. Comp. Neurol 290, 213–242. [DOI] [PubMed] [Google Scholar]
- Sesack SR, Hawrylak VA, Matus C, Guido MA, and Levey AI (1998). Dopamine Axon Varicosities in the Prelimbic Division of the Rat Prefrontal Cortex Exhibit Sparse Immunoreactivity for the Dopamine Transporter. J. Neurosci 18, 2697–2708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shabel SJ, and Janak PH (2009). Substantial similarity in amygdala neuronal activity during conditioned appetitive and aversive emotional arousal. Proc. Natl. Acad. Sci pnas.0905580106. [DOI] [PMC free article] [PubMed]
- Shackman AJ, Salomons TV, Slagter HA, Fox AS, Winter JJ, and Davidson RJ (2011). The integration of negative affect, pain and cognitive control in the cingulate cortex. Nat. Rev. Neurosci 12, 154–167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sharp T, Zetterström T, and Ungerstedt U (1986). An in vivo study of dopamine release and metabolism in rat brain regions using intracerebral dialysis. J. Neurochem 47, 113–122. [DOI] [PubMed] [Google Scholar]
- Sierra-Mercado D, Padilla-Coreano N, and Quirk GJ (2011). Dissociable roles of prelimbic and infralimbic cortices, ventral hippocampus, and basolateral amygdala in the expression and extinction of conditioned fear. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol 36, 529–538. [DOI] [PMC free article] [PubMed] [Google Scholar]
- da Silva JA, Tecuapetla F, Paixão V, and Costa RM (2018). Dopamine neuron activity before action initiation gates and invigorates future movements. Nature 554, 244–248. [DOI] [PubMed] [Google Scholar]
- Sokolowski JD, McCullough LD, and Salamone JD (1994). Effects of dopamine depletions in the medial prefrontal cortex on active avoidance and escape in the rat. Brain Res. 651, 293–299. [DOI] [PubMed] [Google Scholar]
- Sotres-Bayon F, and Quirk GJ (2010). Prefrontal control of fear: more than just extinction. Curr. Opin. Neurobiol 20, 231–235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sparta DR, Hovelsø N, Mason AO, Kantak PA, Ung RL, Decot HK, and Stuber GD (2014). Activation of Prefrontal Cortical Parvalbumin Interneurons Facilitates Extinction of Reward-Seeking Behavior. J. Neurosci 34, 3699–3705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- St Onge JR, Ahn S, Phillips AG, and Floresco SB (2012). Dynamic fluctuations in dopamine efflux in the prefrontal cortex and nucleus accumbens during risk-based decision making. J. Neurosci. Off. J. Soc. Neurosci 32, 16880–16891. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Steinberg EE, Keiflin R, Boivin JR, Witten IB, Deisseroth K, and Janak PH (2013). A causal link between prediction errors, dopamine neurons and learning. Nat. Neurosci 16, 966–973. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stuber GD, Sparta DR, Stamatakis AM, van Leeuwen WA, Hardjoprajitno JE, Cho S, Tye KM, Kempadoo KA, Zhang F, Deisseroth K, et al. (2011). Excitatory transmission from the amygdala to nucleus accumbens facilitates reward seeking. Nature 475, 377–380. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tanda G, Carboni E, Frau R, and Di Chiara G (1994). Increase of extracellular dopamine in the prefrontal cortex: a trait of drugs with antidepressant potential? Psychopharmacology (Berl.) 115, 285–288. [DOI] [PubMed] [Google Scholar]
- Tepper JM, Wilson CJ, and Koós T (2008). Feedforward and feedback inhibition in neostriatal GABAergic spiny neurons. Brain Res. Rev 58, 272–281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thierry AM, Blanc G, Sobel A, Stinus L, and Glowinski J (1973). Dopaminergic Terminals in the Rat Cortex. Science 182, 499–501. [DOI] [PubMed] [Google Scholar]
- Thierry AM, Tassin JP, Blanc G, and Glowinski J (1976). Selective activation of mesocortical DA system by stress. Nature 263, 242–244. [DOI] [PubMed] [Google Scholar]
- Thompson-Schill SL, Jonides J, Marshuetz C, Smith EE, D’Esposito M, Kan IP, Knight RT, and Swick D (2002). Effects of frontal lobe damage on interference effects in working memory. Cogn. Affect. Behav. Neurosci 2, 109–120. [DOI] [PubMed] [Google Scholar]
- Tovote P, Esposito MS, Botta P, Chaudun F, Fadok JP, Markovic M, Wolff SBE, Ramakrishnan C, Fenno L, Deisseroth K, et al. (2016). Midbrain circuits for defensive behaviour. Nature 534, 206–212. [DOI] [PubMed] [Google Scholar]
- Tritsch NX, and Sabatini BL (2012). Dopaminergic modulation of synaptic transmission in cortex and striatum. Neuron 76, 33–50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Trulson ME (1985). Simultaneous recording of substantia nigra neurons and voltammetric release of dopamine in the caudate of behaving cats. Brain Res. Bull 15, 221–223. [DOI] [PubMed] [Google Scholar]
- Tsai H-C, Zhang F, Adamantidis A, Stuber GD, Bonci A, Lecea L. de, and Deisseroth K (2009). Phasic Firing in Dopaminergic Neurons Is Sufficient for Behavioral Conditioning. Science 324, 1080–1084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tseng K-Y, and O’Donnell P (2007). Dopamine Modulation of Prefrontal Cortical Interneurons Changes during Adolescence. Cereb. Cortex 17, 1235–1240. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Twining RC, Wheeler DS, Ebben AL, Jacobsen AJ, Robble MA, Mantsch JR, and Wheeler RA (2015). Aversive stimuli drive drug seeking in a state of low dopamine tone. Biol. Psychiatry 77, 895–902. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tye KM, Mirzabekov JJ, Warden MR, Ferenczi EA, Tsai H-C, Finkelstein J, Kim S-Y, Adhikari A, Thompson KR, Andalman AS, et al. (2013). Dopamine neurons modulate neural encoding and expression of depression-related behaviour. Nature 493, 537–541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ungless MA, Magill PJ, and Bolam JP (2004). Uniform inhibition of dopamine neurons in the ventral tegmental area by aversive stimuli. Science 303, 2040–2042. [DOI] [PubMed] [Google Scholar]
- Urs NM, Gee SM, Pack TF, McCorvy JD, Evron T, Snyder JC, Yang X, Rodriguiz RM, Borrelli E, Wetsel WC, et al. (2016). Distinct cortical and striatal actions of a β-arrestin–biased dopamine D2 receptor ligand reveal unique antipsychotic-like properties. Proc. Natl. Acad. Sci 113, E8178–E8186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Urs NM, Peterson SM, and Caron MG (2017). New Concepts in Dopamine D2 Receptor Biased Signaling and Implications for Schizophrenia Therapy. Biol. Psychiatry 81, 78–85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Valjent E, Bertran-Gonzalez J, Hervé D, Fisone G, and Girault J-A (2009). Looking BAC at striatal signaling: cell-specific analysis in new transgenic mice. Trends Neurosci. 32, 538–547. [DOI] [PubMed] [Google Scholar]
- Verney C, Alvarez C, Geffard M, and Berger B (1990). Ultrastructural Double-Labelling Study of Dopamine Terminals and GABA-Containing Neurons in Rat Anteromedial Cerebral Cortex. Eur. J. Neurosci 2, 960–972. [DOI] [PubMed] [Google Scholar]
- Vertes RP (2004). Differential projections of the infralimbic and prelimbic cortex in the rat. Synap. N. Y. N 51, 32–58. [DOI] [PubMed] [Google Scholar]
- Vincent SL, Khan Y, and Benes FM (1993). Cellular distribution of dopamine D1 and D2 receptors in rat medial prefrontal cortex. J. Neurosci. Off. J. Soc. Neurosci 13, 2551–2564. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vogt BA (2005). Pain and emotion interactions in subregions of the cingulate gyrus. Nat. Rev. Neurosci 6, 533–544. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wanat MJ, Willuhn I, Clark JJ, and Phillips PEM (2009). Phasic dopamine release in appetitive behaviors and drug addiction. Curr. Drug Abuse Rev 2, 195–213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Warden MR, Selimbeyoglu A, Mirzabekov JJ, Lo M, Thompson KR, Kim S-Y, Adhikari A, Tye KM, Frank LM, and Deisseroth K (2012). A prefrontal cortex-brainstem neuronal projection that controls response to behavioural challenge. Nature 492, 428–432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weele CMV, Siciliano CA, Matthews GA, Namburi P, Izadmehr EM, Espinel IC, Nieh EH, Schut EHS, Padilla-Coreano N, Burgos-Robles A, et al. (2018). Dopamine enhances signal-to-noise ratio in cortical-brainstem encoding of aversive stimuli. Nature 563, 397. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wenzel JM, Rauscher NA, Cheer JF, and Oleson EB (2015). A Role for Phasic Dopamine Release within the Nucleus Accumbens in Encoding Aversion: A Review of the Neurochemical Literature. ACS Chem. Neurosci 6, 16–26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wheeler RA, Aragona BJ, Fuhrmann KA, Jones JL, Day JJ, Cacciapaglia F, Wightman RM, and Carelli RM (2011). Cocaine cues drive opposing context-dependent shifts in reward processing and emotional state. Biol. Psychiatry 69, 1067–1074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wickham RJ, Solecki W, Rathbun LR, Neugebauer NM, Wightman RM, and Addy NA (2013). Advances in studying phasic dopamine signaling in brain reward mechanisms. Front. Biosci. Elite Ed 5, 982–999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilkinson LS, Dias R, Thomas KL, Augood SJ, Everitt BJ, Robbins TW, and Roberts AC (1997). Contrasting effects of excitotoxic lesions of the prefrontal cortex on the behavioural response to D-amphetamine and presynaptic and postsynaptic measures of striatal dopamine function in monkeys. Neuroscience 80, 717–730. [DOI] [PubMed] [Google Scholar]
- Williams GV, and Goldman-Rakic PS (1995). Modulation of memory fields by dopamine D1 receptors in prefrontal cortex. Nature 376, 572–575. [DOI] [PubMed] [Google Scholar]
- Williams SM, and Goldman-Rakic PS (1993). Characterization of the Dopaminergic Innervation of the Primate Frontal Cortex Using a Dopamine-specific Antibody. Cereb. Cortex 3, 199–222. [DOI] [PubMed] [Google Scholar]
- Wimmer RD, Schmitt LI, Davidson TJ, Nakajima M, Deisseroth K, and Halassa MM (2015). Thalamic control of sensory selection in divided attention. Nature 526, 705–709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wise RA (2008). Dopamine and reward: the anhedonia hypothesis 30 years on. Neurotox. Res 14, 169–183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woolley ML, Waters KA, Reavill C, Bull S, Lacroix LP, Martyn AJ, Hutcheson DM, Valerio E, Bate S, Jones DNC, et al. (2008). Selective dopamine D4 receptor agonist (A-412997) improves cognitive performance and stimulates motor activity without influencing reward-related behaviour in rat. Behav. Pharmacol 19, 765–776. [DOI] [PubMed] [Google Scholar]
- Yang CR, and Seamans JK (1996). Dopamine D1 receptor actions in layers V-VI rat prefrontal cortex neurons in vitro: modulation of dendritic-somatic signal integration. J. Neurosci. Off. J. Soc. Neurosci 16, 1922–1935. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yizhar O, and Klavir O (2018). Reciprocal amygdala–prefrontal interactions in learning. Curr. Opin. Neurobiol 52, 149–155. [DOI] [PubMed] [Google Scholar]
- Yizhar O, Fenno LE, Prigge M, Schneider F, Davidson TJ, O’Shea DJ, Sohal VS, Goshen I, Finkelstein J, Paz JT, et al. (2011). Neocortical excitation/inhibition balance in information processing and social dysfunction. Nature 477, 171–178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- York DH (1973). Motor responses induced by stimulation of the substantia nigra. Exp. Neurol 41, 323–330. [DOI] [PubMed] [Google Scholar]
- Young RF, Kroening R, Fulton W, Feldman RA, and Chambi I (1985). Electrical stimulation of the brain in treatment of chronic pain. Experience over 5 years. J. Neurosurg 62, 389–396. [DOI] [PubMed] [Google Scholar]
- Zhang H, and Sulzer D (2004). Frequency-dependent modulation of dopamine release by nicotine. Nat. Neurosci 7, 581–582. [DOI] [PubMed] [Google Scholar]
- Zhang Z-W, Burke MW, Calakos N, Beaulieu J-M, and Vaucher E (2010). Confocal Analysis of Cholinergic and Dopaminergic Inputs onto Pyramidal Cells in the Prefrontal Cortex of Rodents. Front. Neuroanat 4, 21. [DOI] [PMC free article] [PubMed] [Google Scholar]