

Supplemental digital content is available in the text.
Key Words: EXERCISE, PERFORMANCE, PREDICTION, RUNNING
ABSTRACT
Introduction
Critical speed (CS) represents the highest intensity at which a physiological steady state may be reached. The aim of this study was to evaluate whether estimations of CS obtained from raw training data can predict performance and pacing in marathons.
Methods
We investigated running activities logged into an online fitness platform by >25,000 recreational athletes before big-city marathons. Each activity contained time, distance, and elevation every 100 m. We computed grade-adjusted pacing and the fastest pace recorded for a set of target distances (400, 800, 1000, 1500, 3000, and 5000 m). CS was determined as the slope of the distance–time relationship using all combinations of, at least, three target distances.
Results
The relationship between distance and time was linear, irrespective of the target distances used (pooled mean ± SD: R2 = 0.9999 ± 0.0001). The estimated values of CS from all models were not different (3.74 ± 0.08 m·s−1), and all models correlated with marathon performance (R2 = 0.672 ± 0.036, error = 8.01% ± 0.51%). CS from the model including 400, 800, and 5000 m best predicted performance (R2 = 0.695, error = 7.67%) and was used in further analysis. Runners completed the marathon at 84.8% ± 13.6% CS, with faster runners competing at speeds closer to CS (93.0% CS for 150 min marathon times vs 78.9% CS for 360 min marathon times). Runners who completed the first half of the marathon at >94% of their CS, and particularly faster than CS, were more likely to slowdown by more than 25% in the second half of race.
Conclusion
This study suggests that estimations of CS from raw training data can successfully predict marathon performance and provide useful pacing information.
Marathon performance is determined by several factors, including the physiological characteristics of the runner, race course, weather and ambient conditions, and pacing strategies (1–3). The physiological factors associated with endurance performance typically include the maximum oxygen consumption (V˙O2max), lactate threshold, and running economy (1,3). Alternatively, predictions of endurance performance can be derived from the relationship between the intensity of exercise (e.g., running speed) and the time such intensity can be sustained until task failure (Tlim) (4). Specifically, as running speed (S) increases, the duration of exercise until Tlim decreases, which forms a hyperbolic function (equation 1):
| [1] |
The asymptote of the hyperbola is known as critical speed (CS), and the curvature constant is termed as D′. Equation 1 can be rearranged to produce a linear function between distance (D) and Tlim (equation 2):
| [2] |
Physiologically, CS represents the highest intensity at which oxidative phosphorylation is sufficient to satisfy the energy demand (5,6), whereas D′ represents a finite work capacity above CS (7). A metabolic steady state, therefore, can be achieved during exercise below CS, but not above CS. For example, pulmonary oxygen uptake (V˙O2), the concentration of intramuscular metabolites such as inorganic phosphate, hydrogen ions, and phosphocreatine, and the concentration of lactate in blood reach a steady state below, but not above, CS (5,8–10). Exercise above CS is characterized by the progressive depletion of D′, which has been linked to substrate phosphorylation (11–13). Furthermore, exercise above CS also results in the inexorable increase in the concentration of intramuscular metabolites and continued increase of V˙O2 until a “critical” threshold of muscle substrate and metabolite concentrations is reached, V˙O2max is attained, and ultimately exercise is terminated (7–9,14).
Given the profound differences observed during exercise below versus above CS, there is a growing body of evidence suggesting that CS and D′ are strong predictors of endurance performance (e.g., [15–20]). Jones and Vanhatalo (19) conducted a recent analysis of elite marathon runners and observed that their best marathon performance has been achieved at speeds corresponding to ~96% of their CS. However, it remains unknown whether recreational runners also complete their marathon at speeds close to, but below, their CS. Furthermore, because an even pace seems to be the best strategy in a marathon (21), it can be hypothesized that runners who complete the first half of the marathon closer to, and particularly faster than their CS, would experience a significant slowdown in the final phase of the marathon.
The determination of CS is a relatively simple process but can be time consuming as it requires undertaking a series of maximal efforts (22). Furthermore, it has been suggested that the attainment of V˙O2max should be verified in trials intended to be used to calculate CS (22). Some authors have used personal best performance to determine CS in elite athletes (19), or to examine the evolution of human performance (through CS and D′) over the history of Olympic Games (23). Furthermore, critical power and W′, the cycling analogous of CS and D′, respectively, have been determined from “intentional” and “nonintentional” efforts recorded over 4 wk of training data (24). There has been an increase in online platforms where runners log their activities using devices such as smartphones or smartwatches. Such activities are likely to include both training sessions and races and may be used to estimate CS and D′.
The aims of the current study, therefore, were (i) to use a large set of raw training data logged into an online fitness platform to calculate CS and D′ and compare such estimates to marathon performance and (ii) to investigate whether the calculated CS and D′ can offer any insight into pacing during marathon performance. We hypothesized that (i) training data logged into an online fitness platform will allow determination of CS, as demonstrated by strong linearity of the distance–Tlim relationship and low standard error of estimate (SEE) in CS and D′; (ii) the calculated CS and D′ obtained from training data will be strongly associated with marathon time (MT) and, specifically, marathon speed (MS) will be close to, but lower than, CS; and (iii) CS will offer valuable information regarding pacing, so that athletes who complete the first half of the marathon faster than their CS were at a greater risk of experiencing a positive split.
METHODS
Data set
The data set for this study consisted of the activity logged to an online platform (Strava®) by 31,190 runners of the Dublin, London, and New York marathons from mid-2014 to late 2017 (Table 1). These data were, in part, provided by Strava® under limited research license. Each runner included in the data set registered a race time for one of these marathons, and the data set included all of their logged training activities for the 16 wk before that race. Each runner was characterized with gender and age information, and each activity included time, distance, and elevation data sampled every 100 m. Taking into consideration the different energy costs of uphill and downhill running (25), we computed pacing and grade-adjusted pacing at 100 m intervals. A detailed explanation of how grade-adjusted pacing was calculated is provided in the supplementary material (see Appendix 1, Supplemental Digital Content 1, Grade adjusted pace calculations, http://links.lww.com/MSS/C10).
TABLE 1.
Summary of the data set by city and year.
| City | Year | Sex (M/F) | Number of Runners | Age (yr) | Finishers (%) | Race Time (Min) | Frequency (Activities per Week) | Volume (km·wk−1) |
|---|---|---|---|---|---|---|---|---|
| Dublin | 2014 | F | 59 | 38 ± 7 | 90.1 | 258.4 ± 30.9 | 3.2 ± 1.1 | 33.8 ± 9.9 |
| M | 313 | 38 ± 7 | 97.0 | 221.0 ± 35.8 | 3.8 ± 1.7 | 42.9 ± 17.0 | ||
| 2015 | F | 91 | 39 ± 8 | 90.9 | 253.5 ± 33.9 | 3.5 ± 1.3 | 36.3 ± 12.3 | |
| M | 506 | 39 ± 8 | 96.0 | 217.8 ± 35.8 | 3.9 ± 1.8 | 42.5 ± 17.8 | ||
| 2016 | F | 220 | 39 ± 8 | 93.0 | 257.3 ± 40.9 | 3.6 ± 1.4 | 36.8 ± 13.6 | |
| M | 959 | 40 ± 8 | 97.2 | 220.2 ± 35.3 | 3.8 ± 1.7 | 42.1 ± 16.8 | ||
| London | 2015 | F | 641 | 39 ± 8 | 96.3 | 242.2 ± 42.4 | 3.7 ± 1.7 | 40.3 ± 14.9 |
| M | 2151 | 40 ± 8 | 97.1 | 202.5 ± 38.0 | 4.2 ± 2.2 | 47.1 ± 21.4 | ||
| 2016 | F | 1053 | 38 ± 9 | 96.2 | 242.3 ± 43.1 | 3.7 ± 1.5 | 39.3 ± 15.3 | |
| M | 3197 | 40 ± 8 | 97.3 | 203.0 ± 39.3 | 4.3 ± 2.3 | 47.3 ± 22.4 | ||
| 2017 | F | 1824 | 39 ± 8 | 96.5 | 251.4 ± 47.7 | 3.6 ± 1.7 | 37.8 ± 14.8 | |
| M | 4278 | 41 ± 8 | 97.6 | 207.1 ± 41.9 | 4.3 ± 2.3 | 47.3 ± 22.5 | ||
| New York | 2015 | F | 412 | 38 ± 9 | 87.8 | 253.2 ± 43.6 | 3.7 ± 1.4 | 39.5 ± 14.4 |
| M | 1460 | 40 ± 8 | 92.3 | 223.9 ± 42.8 | 3.8 ± 1.6 | 44.0 ± 19.2 | ||
| 2016 | F | 841 | 37 ± 9 | 87.9 | 250.3 ± 42.7 | 3.8 ± 1.6 | 39.6 ± 15.5 | |
| M | 2314 | 40 ± 9 | 92.7 | 224.0 ± 41.5 | 3.8 ± 1.7 | 43.4 ± 18.0 | ||
| 2017 | F | 1427 | 37 ± 9 | 90.6 | 250.3 ± 43.0 | 3.8 ± 1.5 | 40.0 ± 16.2 | |
| M | 3497 | 40 ± 9 | 93.0 | 223.7 ± 41.1 | 3.8 ± 1.6 | 43.5 ± 18.5 |
The study was reviewed by the ethics committees at University College Dublin and University of Hertfordshire and deemed to be exempt from ethical approval by both institutions since no data were collected. It is worth noting that, first, the data set was anonymized by assigning a unique code for each runner in each marathon. Therefore, it is not possible to identify whether a given runner has competed in multiple races. Second, the logged activities may not provide a complete record of all training activities because only logged running activities were included in our analysis.
Calculation of CS from raw training data
For each runner, the fastest time recorded at any time within the 16-wk period before the marathon was calculated for a range of target distances: 400, 800, 1000, 1500, 3000, and 5000 m. This range of target distances was selected because training programs may push runners to achieve fast times for these specific distances. Furthermore, athletes training for a marathon are likely to also participate in other races, such as 5000-m races, during their training, whereas the shorter distances are common in training programs with prescribed interval sessions or time trials. This range of distances is also similar to those used previously to determine CS in elite runners from performance data and from field studies (19,23,26).
Because the data were sampled at 100-m intervals, a rolling average of the grade-adjusted pacing was used to determine the fastest time recorded for each target distances and assumed to be Tlim. For example, the fastest 1000-m performance was calculated as the rolling average of 10 consecutive values of the grade-adjusted pace that resulted in the shortest time. CS and D′ were then calculated for each athlete using the relationship between distance and Tlim, where the slope of the line represents CS, and the y-intercept represents D′ (equation 2 [27]). For each athlete, 42 values of CS and D′ were calculated using all possible combinations that included, at least, three of the target distances: 400, 800, 1000, 1500, 3000, and 5000 m.
Statistical analysis
The ability of CS and D′ to predict marathon performance, defined as MT, was calculated using a linear regression. Specifically, we conducted 42 linear regressions, using all available values of CS and D′. However, for clarity, subsequent analyses were conducted only on the model using 400-, 800-, and 5000-m distances, which was selected as the best performing model (i.e., the model with the lowest SEE predicting MT). Briefly, to do this, we used each runner as the test runner and fit a linear regression model using the CS and D′ values of the remaining runners and their MT. Then, with the CS and D′ values of the test runner, we used the resulting linear regression model to predict the test runner’s MT. This was repeated for all runners, and the percentage error between the actual and the predicted MT was used to compare the accuracy of predictions.
A linear regression between relative MS and MT was conducted to investigate how runners paced their marathon relative to their CS. Relative MS was determined as average speed during the marathon (i.e., MS) relative to CS (equation 3):
| [3] |
In addition, base speed (BS) was calculated as the average speed from the 2 km to the 16 km during the marathon, as we observed that during this early portion of the race most runners were able to sustain a relative stable pace. Relative BS was subsequently calculated as BS relative to CS (equation 4):
| [4] |
A linear regression was conducted to investigate the relationship between relative BS and relative MS. In addition, a linear regression was conducted between relative BS and frequency of runners that experienced a significant reduction in speed in the last section of the race, defined as a slowing down during the last 12.2 km of the marathon by more than 25% relative to their BS (i.e., slowdown >1.25).
The athletes were categorized based on gender (male and female) and age, and all analyses were conducted for male and female runners and younger and older runners. For age, we selected an arbitrary threshold of 40 yr to distinguish between younger (<40 yr old) and older (40+ yr old) runners, as this age resulted in approximately the same number of younger and older runners. The significance of the linear regressions was tested using F-tests, and we used one-tailed Welch’s t-tests to assess the difference between groups (male vs female runners, younger vs older runners) for the variables above. We performed this analysis based on absolute MT, grouped in 10-min bins, and relative marathon performance (i.e., relative MS and relative BS), grouped in 2% bins. The SEE associated with CS and D′ was calculated and reported as coefficient of variation (CV%). All analyses were performed using Python. Results are presented as mean ± SD, unless otherwise stated. In all cases, a 99% confidence level (P < 0.01) was used as the threshold for significance.
RESULTS
Determination of CS and D′ from raw training data
The relationship between distance and duration was highly linear for all 42 combinations of target distances and Tlim used (pooled average from all models and all participants: R2 = 0.9999 ± 0.0001). The CS obtained from the 42 models was 3.74 ± 0.08 m·s−1, and the SEE associated with these estimates was 0.05 ± 0.02 m·s−1 (CV% = 1.3%). The corresponding value for D′ from the 42 models was 149 ± 24 m, and the SEE of these estimates was 28 ± 11 m (CV% = 19.44%).
The model using 400, 800, and 5000 m produced the lowest error to predict marathon performance and, therefore, was deemed as the best performing model. It resulted in a strong linear relationship between distance and Tlim for all participants (R2 = 0.999 ± 0.001), and the estimated mean values of CS and D′ for this model were 3.69 ± 0.57 m·s−1 and 136 ± 39 m, respectively. The corresponding CV% values associated with these estimations were 0.73% and 14.56%, respectively. For simplicity, all subsequent results refer to estimates of CS and D′ derived only from this best performing model.
The estimates of CS were ~16.4% greater for male runners compared with females (3.83 ± 0.54 vs 3.29 ± 0.47 m·s−1; t = 75.92, P < 0.01), whereas D′ estimates were ~7.8% lower for male runners compared with females (133 ± 37 vs 144 ± 41 m, respectively; t = −18.24, P < 0.01). With regard to age, estimates of CS for younger athletes were 8.8% higher than for older athletes (3.76 ± 0.59 vs 3.62 ± 0.54 m·s−1; t = 20.04, P < 0.01), whereas D′ was 1.6% greater in the younger compared with older age category (137 ± 38 m vs 135 ± 39 m; t = 4.57, P < 0.01).
CS and prediction of marathon performance
The best performing model was correlated with MT (R2 = 0.695, f = 2.88 × e04, P < 0.01) and was able to predict MT with a 7.67% error. In fact, all of 42 models of CS and D′ generated were successful at predicting MT (R2 = 0.67 ± 0.03; P < 0.01). The error associated with these predictions was 8.01% ± 0.51%. The single worst performing model used 400, 800, and 1000 m and produced an R2 of 0.54 and a 9.72% prediction error.
The mean percentage prediction error in marathon performance for the model used in this study, as a function of MT, is presented in Figure 1 for male and female athletes and for both age categories. The overall percentage error was not significantly different between male and female runners, 7.67% ± 6.63% and 7.67% ± 6.20%, respectively (t = 0.02, P = 0.98). However, for MT from 170 to 240 min, inclusive, the percentage error was significantly greater in female runners (P < 0.01). For slower runners with MT from 250 to 360 min, the percentage error was significantly greater among males compared with females (P < 0.01). There was a small, but significant, difference in the percentage error of the model between younger and older runners (7.56% ± 6.42% and 7.79% ± 6.63%, respectively; t = −2.84 and P = 0.004).
FIGURE 1.
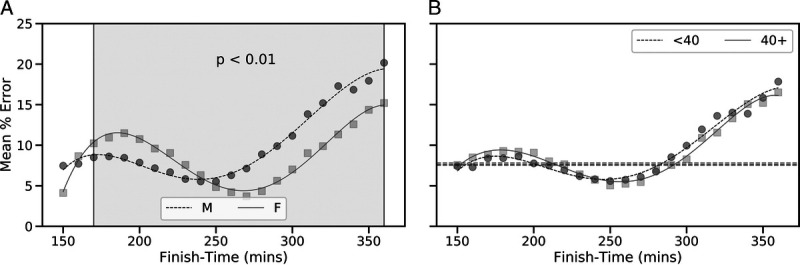
Mean percentage prediction error by finish time and based on gender (A) and age category (B) using a leave-one-out test to evaluate the best performing model at predicting MT. The shaded region in panel A indicates finish times associated with error differences between male and female runners that were found to be significant (P < 0.01) based on a one-tailed Welch’s t-test.
Marathon pacing relative to CS
Runners completed their marathon at an average speed that corresponded to 84.8% ± 13.6% of their CS (Fig. 2). Relative to CS, females completed the marathon slightly (~1%), but significantly, faster than males (85.5% ± 23.2% CS vs 84.6% ± 7.8%; t = −2.94, P = 0.003). Specifically, females run relatively faster (i.e., closer to CS) than males for MT from 170 to 360 min, inclusive (Fig. 3; P < 0.01). There were no differences in relative MS in both age categories (Fig. 2), with younger runners competing at 84.8% ± 17.4% of their CS and older runners at 84.9% ± 7.7% of their CS (t = −0.79, P = 0.42).
FIGURE 2.
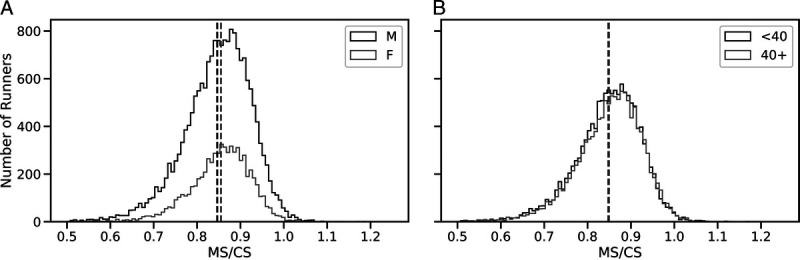
Distributions of relative MS (Rel MS), defined as CS relative to MS, for male and female (A) and younger and older (B) runners.
FIGURE 3.
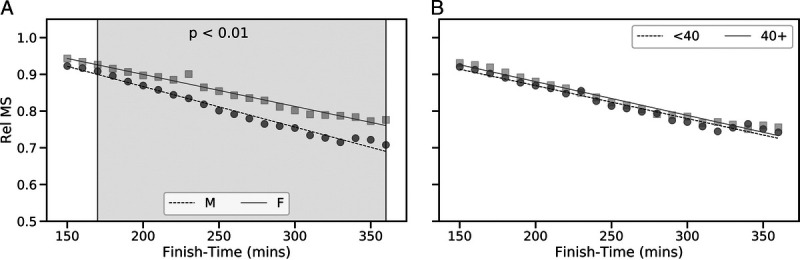
Relative MS (Rel MS), defined as CS relative to MS, of runners versus finish time. Results are presented based on gender (A) and age category (B). The shaded regions in panel A indicates finish times associated with relative MS differences between male and female runners that were found to be significant (P < 0.01) based on a one-tailed Welch’s t-test.
Faster runners completed the marathon at speeds closer to CS compared with slower runners, as denoted by the negative relationship between MS relative to CS (relative MS), and overall marathon performance (Fig. 3, all P < 0.01). This held true irrespective of gender (R2 of 0.99 and 0.98 for males and females, respectively) and age (R2 = 0.98 for both age-groups). Irrespective of gender or age, runners with finishing times of ~150 min completed the marathon at 93.0% of their CS, whereas runners with finishing times >300 min completed the marathon at 78.9% of their CS (Fig. 3).
The speed of runners from 2 to 16 km into the marathon (BS) was 3.24 ± 0.55 m·s−1, which corresponds to 87.6% ± 6.9% of CS. There was a small but significant difference in relative BS (BS relative to CS) between male and female runners (87.5% ± 7.0% vs 88.0% ± 6.5% of CS, respectively; t = −4.7 and P < 0.01). There was a similar small but significant difference in relative BS between younger and older runners (87.4% ± 6.8% vs 87.9% ± 6.9%, respectively; t = −5.3, P < 0.01).
Runners with a higher relative BS experienced a greater relative split, as denoted by the positive correlations between relative BS and relative split time (Fig. 4). This observation held true irrespective of gender (R2 = 0.72 and R2 = 0.69 for males and females, respectively) and age category (R2 = 0.73 and R2 = 0.77 for younger and older runners, respectively; all P < 0.01 except in the case of older runners). In other words, typically the faster an athlete ran the first part of the marathon, relative to their CS, the greater the slowdown they experienced in the second half of the marathon. Male runners had greater relative splits (slower second half) than female runners for relative BS values from 76% to 94% of CS (P < 0.01). There were no significant differences in relative BS and relative split between younger and older runners.
FIGURE 4.
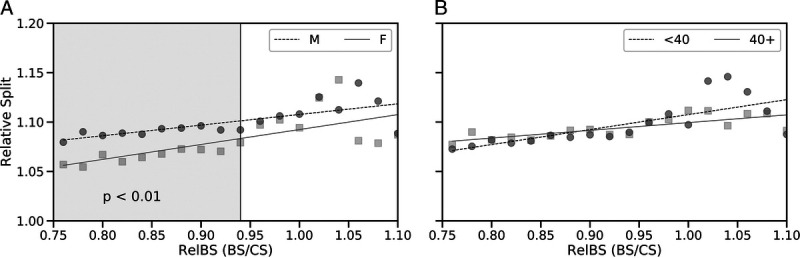
Relative split of runners (second-half time divided by first-half time) as a function of relative BS (relative BS, defined as average running speed 2–16 km as a fraction of CS). Results are presented based on gender (A) and age category (B). The shaded region in panel A indicates the relative BS range that was associated with relative split differences, between male and female runners, that were found to be significant (P < 0.01) based on a one-tailed Welch’s t-test.
The percentage of runners that exhibited a decrease in speed greater than 25% in the last 12.2 km of the marathon, relative to their BS (i.e., a relative split ≥1.25), was greater among males compared with females (20.5% vs 10.7%, respectively; t = 17.86, P < 0.01). The percentage of athletes with such slowdown was not statistically different with respect the age categories (17.9% vs 18.1% for young and old age category, respectively; t = −0.55, P = 0.57).
The relationship between relative BS and percentage of runners with a slowdown ≥1.25 is presented in Figure 5. There were positive correlations between relative BS and percentage of runners with a slowdown ≥1.25 for males and females (R2 = 0.62 with P = 0.005 and R2 = 0.36 with P = 0.13, respectively). Similarly, there was a positive correlation between relative BS and percentage in younger (R2 = 0.67, P = 0.002) but not older runners (R2 = 0.04, P = 0.7). It is worth noting that 23% of runners with relative BS > 1 (that is, running the first part of the race faster than their CS) experienced a significant slowdown, compared with 17% of runners who run the first half of the marathon at speeds slower than their CS, and suffered a comparable slowdown during the second half of the race (P < 0.01). There was evidence to suggest that a relative BS of 0.94 represents an important threshold for runners, with respect to their ability to avoid a significant slowdown (≥1.25) later in the race (Fig. 5). Specifically, the proportions of male and female athletes with a relative BS < 0.94 who go on slow significantly are 20.5% and 9.6%, respectively, compared with 26.0% and 15.6% for men and women with a relative BS > 0.94 (both P < 0.01).
FIGURE 5.
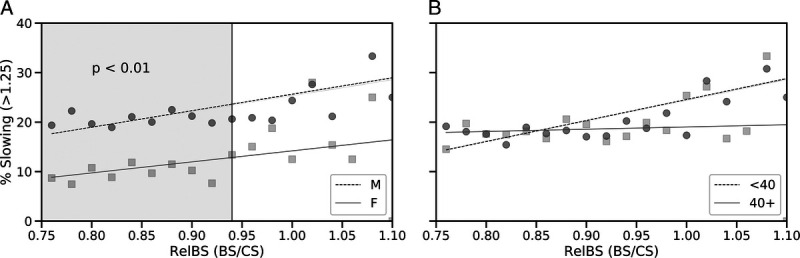
Percentage of runners experiencing a late-race (final 12 km) slowdown in excess of 25% (relative to BS) versus relative BS (Rel BS, defined as average running speed 2–16 km as a fraction of CS). Results are presented based on gender (A) and age category (B). The shaded region in panel A indicates the relative BS range that was associated with percentage slowing differences, between male and female runners, that were found to be significant (P < 0.01) based on a one-tailed Welch’s t-test.
DISCUSSION
CS is a physiological threshold with the potential to assess athletic ability. In this study, we investigated whether CS determined from raw training data logged into a fitness application can be used to predict performance and offer insight into pacing in a marathon race. The main findings from this study were as follows: (i) it seems feasible to estimate CS from raw training data, and the estimates were sufficiently accurate to be useful for predicting athletic performance; (ii) the average runner completed their marathon at ~85% of their estimated CS, with faster runners achieving speeds that were on average closer to, but still below, their estimated CS; and (iii) CS may be useful to monitor pacing during a marathon, as runners who perform the early part of the marathon (2–16 km) faster than 94% of their CS were associated with a slower second-half time and a greater likelihood of experiencing a slowdown greater than 25% in the final stages of the race (>30 km).
Determination of CS from raw training data
The conventional approach to determine CS and D′ requires a series of maximal efforts, either at constant intensity to task failure or as time trials (22). In the present study, we used raw training data to calculate CS and D′ and, therefore, did not have access to such information. Furthermore, we did not have access to physiological (e.g., heart rate) or psychobiological (e.g., rating of perceived exertion) data, which can be used as surrogate measures to evaluate whether a maximal effort has been performed. Instead we used the fastest pace recorded for six target distances, from 400 m to 5 km, during the weeks leading up to a marathon. This may impose a limitation on this study because using raw training data to calculate CS and D′ does not allow verification of maximal effort. We conducted some further analysis to develop an adjusted model in an effort to improve estimates of the fastest paces for runner, which form the basis of our model and estimates of CS and D′, in an effort to assess whether predictive trials were maximal. The adjusted model was developed with the assumption that when the fastest time for a target distance was recorded, the remaining of the activity should be considerably slower. The details of this adjustment are presented in the supplementary material accompanying this article (Supplemental Digital Content 2, http://links.lww.com/MSS/C11). In brief, as an example, if a runner ran their fastest 1 km over a 5-km session, but the remaining 4 km was not much slower, then it suggested that the fastest 1-km pace was not a maximal effort pace and, therefore, should be adjusted (i.e., increase pace). The adjusted model was complex, and the key results were similar to those obtained from the original analysis (see Appendix 2, Supplemental Digital Content 2, Adjusted Model, http://links.lww.com/MSS/C11) and, therefore, not included as part of the document. Further methodological considerations are discussed below. Nonetheless, the results presented herein suggest that raw training data have the potential to permit estimates of CS and D′ that are accurate and useful in practice. Further to its potential to assess endurance performance and inform pacing (see discussion below), CS may be seen as a tool to assess cardiorespiratory fitness because of its strong correlations with V˙O2max (5), the time constant of V˙O2 kinetics (28,29), the percentage of Type I muscle fibers (30,31), and muscle capillarity (31). Future studies may investigate whether raw training data can be used to assess improvement in fitness, by looking at the changes in CS and D′ with training.
The physiological basis of D′ is less well understood, and its precise etiology is complex and multifactorial (5,22). Nonetheless, there is some evidence suggesting that D′ may be linked to substrate phosphorylation and muscle mass and function (32,33). Therefore, we would expect a higher D′ in males compared with females, and in young compared with older runners. However, although we observed a greater D′ in the younger group compared with older runners, the estimated D′ was ~7.8% greater in females compared with males. The reason for this phenomenon is not clear, but it may be linked to greater error inherent in the estimation of D′.
CS and marathon performance
An important finding from this study was that marathon performance can be predicted using estimations of CS and D′ obtained from raw training data. Previous studies have already shown that CS determined in laboratory conditions can be a good predictor of endurance performance (r ≥ 0.87 [15–18,20]), including evidence that CS may be associated with marathon performance (19,20). The strength of the correlation between CS and marathon performance was remarkably similar in the study by Florence and Weir [20] (R2 = 0.76), where CS was determined in controlled laboratory conditions for 12 participants, and in the current study, where CS was determined using raw training data for >25,000 runners (R2 = 0.67). The best performing model (using 400-, 800-, and 5000-m distances) was able to predict marathon performance with 7.67% error. Indeed, most combinations of target distances (61%) used to calculate CS and D′ resulted in errors of <8%, and the worst performing model (using the distances 400, 800, and 1000 m) was associated with a prediction error of 9.7%. The error associated with these predictions was lower in males compared with females for fast runners, but the model was more accurate in females for slow marathon runners. It is plausible that predictions of fast marathon (~175 to 225 min) MT were more accurate in males compared with females because males also had faster MT (~215 min) compared with females (~250 min). Therefore, more data were available, and the models were able to produce more accurate predictions of marathon performance. Nonetheless, overall the predictions of MT had ~8% error, which corresponds to ~18 min for the average MT of 230 min. This value compares favorably with the error obtained from equations that aim to predict MT (34). However, there are multiple equations available to predict MT, which are based on several variables, such as anthropometric, physiological, and/or training history and previous performance, and it remains unclear which equations provide the best estimation (34). The present study demonstrates that CS determined from raw training data can be useful to predict MT in a large, heterogeneous sample.
The data reported in this study show that estimates of D′ exhibit a much greater CV% than that of CS (~16% vs ~1%, respectively), which is a common finding within the literature (22). For example, Black et al. (9) and Muniz-Pumares et al. (22) reported CV% of 2%–3% for critical power, but CV% of ~10% for W′. It should be noted that the relative contribution of D′ is small in endurance events such as the marathon, but it remains unclear whether CS and D′ determined from raw training data may be able to predict performance in events in which the relative contribution of D′ is larger, such as a 5-km race.
Marathon pacing relative to CS
It has long been appreciated that intensities corresponding to CS (or its cycling analogous critical power) can only be sustained for up to 1 h (22). In the present study, therefore, it was hypothesized that marathons would be performed at speed close to, but below, CS. The results showed that recreational athletes completed the marathon at ~85% CS (Fig. 2). Previous data on marathon performance relative to CS are limited to a small group of elite athletes (19), who were able to run the marathon at ~96% of their CS. It is not clear why elite athletes were able to sustain a higher fraction of their CS in the marathon. However, it has been recently shown that prolonged, submaximal exercise can affect both critical power and W′ (35–37). Specifically, Clark et al. (35–37) reported a ~10% reduction in critical power after 120 min of exercise below critical power. Assuming a similar decline in CS is also observed during prolonged exercise, CS may have been reduced during the marathon preventing participants to complete at a higher fraction of their CS. Clark et al. (35) suggested that the decline in critical power may occur as a function of time, as it was evident after 120 min of exercise but not after 40 and 80 min of exercise. In the present study, we analyzed data from recreational runners with an average MT of ~230 min, which may result in greater a decline in CS compared with elite athletes who completed the marathon in ~125 min (19). In the present study, we observed that more competitive athletes were able to maintain speeds of up to 93.0% of their CS, but somewhat slower athletes were only able to complete the marathon at speeds corresponding to ~80% CS. Indeed, there was a strong negative relationship between MS relative to CS and finish time (R2 ≥ 0.98; Fig. 3), which may be interpreted as (i) CS decreases as a function of time, and therefore faster athletes complete the marathon with a lower MT, which results in lower decrease in CS; (ii) faster athletes are fitter athletes, and fitness itself offers some protection to the deterioration of CS during exercise; or (iii) a combination of both. This remains speculative, and further research may investigate whether CS is also affected by prolonged running; its etiology and possible strategies to minimize such decline, including carbohydrate supplementation (37); and its implication for accuracy of MT predictions derived from CS and D′.
It is worth noting that a small number of runners (0.82%) completed the marathon at speeds faster than their estimated CS (i.e., MS relative to CS > 1 in Fig. 1). This result is difficult to reconcile with the current understanding of the CS as the upper limit of sustainable exercise. It is possible that available raw training data failed to capture maximal effort in some participants and resulted in the underestimation of CS. Then again, it is plausible that some runners benefited from a taper premarathon or experienced favorable conditions during the race compared with conditions they may have experienced during training. For instance, all marathons investigated where completed very close to sea level (Dublin, London, and New York). It is well known that altitude has a negative effect on endurance performance, which is evident from altitudes of only 150–300 m above sea level (38). Therefore, raw training data may underestimate CS in athletes that live at moderate altitudes and then compete at sea level. It is also plausible that in big-city marathons, with thousands of athletes competing simultaneously, some athletes benefit from drafting, which results in decreased oxygen cost of running (39), and therefore an increased performance (2). Nonetheless, the results from the current study show that, as hypothesized, >99% of 25,000 recreational runners performed the marathon at speeds close to but below their CS.
The results from this study suggest runners who completed the initial part of the marathon closer to their CS were more likely to experience difficulties later in the race, quantified as a positive relative split (i.e., slowing down in the second half of the race) and also as the percentage of athletes who experienced a significant slowdown in the final stages of the race. The results reported in the current study showed a positive correlation between BS relative to CS and subsequent impairment in performance, irrespective of gender and age category. Adopting an even split has been suggested as the best approach for endurance events (21), and large-scale studies have already suggested that an even split is optimal for endurance performance (40). The data presented in the current study suggest that such even pace should be close to, but below, the CS. For example, runners who complete an evenly paced race (defined as a race in which the first-half time is within 2%, faster or slower, than the second-half time) do so by running at a speed that is ~88% of their CS, whereas runners whose pacing falls outside of this range completed their race at ~84% of their CS. The results of the present study suggest that speed in the first half of the marathon should stay within ~94% of CS, or athletes risk having to disproportionately slowdown in the second half of the marathon.
The results also showed that although no differences were observed between younger and older athletes, males experienced a greater slowdown in the second half of the marathon, and severe slowdowns >25% occurred more frequently compared with females. This is consistent with research showing that female runners tend to pace their marathons more evenly than male runners and that they are less susceptible to severe slowdowns and hitting the wall when compared with male marathoners (40,41).
Methodological considerations
The determination of CS and D′ requires participants to perform a series of maximal efforts. When testing is conducted in a laboratory, it is possible to assess if participants have performed a maximal effort, for instance, by checking whether V˙O2max has been attained during predictive tests (22). In the current study, however, CS and D′ have been determined using raw training data, and it was not possible to verify whether predictive tests represented a maximal effort. Although this is a potential limitation for using raw training data to calculate CS and D′, it is worth noting that (i) when performing maximal efforts, the relationship between distance and time becomes linear. The data reported herein also show a very strong linear relationship between distance and time for all participants. (ii) We used data from a 16-wk period before the race and only included runners with at least 24 activities logged during this period. It is plausible that some of the activities in the data set correspond to shorter races (e.g., 5 km) and training activities performed maximally, e.g., high-intensity interval training. (iii) It has been shown that unintended efforts over a single training session can be used to estimate critical power and W′, and such estimations are similar to those obtained from field data do not have to perform predefined intentional efforts (24). However, Karsten et al. (24) included race data, whereas it was not possible to differentiate training or race data in the current study. (iv) CS and D′ were determined using an adjusted model, with the assumption that if runs included in the analysis of CS and D′ would approximate a maximal effort, these would be likely to occur early in a training session. The results from the adjusted model (presented in the Supplemental Digital Content) were similar to those presented herein. Therefore, although a degree of caution is warranted until a direct validation of CS and D′ determined from raw training data is performed against laboratory-based estimations of these parameters, the results suggest that it would be possible to calculate CS and D′ from raw training data and such estimates can be useful to estimate marathon performance and inform pacing.
CONCLUSIONS
In the current study, we aimed to use a large data set of raw training data to determine CS and D′ and to evaluate whether such predictions can be used to assess performance and pacing during a marathon. The main, original finding in the study is that CS and D′ determined from a large data set of raw training data may be useful to predict marathon performance and pacing. These results have the potential to enable fitness apps to provide more targeted advice to their users about training options and racing strategies, perhaps enabling more targeted, personalized training programs that are based on physiological thresholds (i.e., CS) and that can adapt to individual runners, thereby helping them to optimize their training and performance.
Acknowledgments
The authors declare no conflict of interest and do not have any financial disclosures. The results of the study are presented clearly, honestly, and without fabrication, falsification, or inappropriate data manipulation. The results of the present study do not constitute endorsement by the American College of Sports Medicine.
Footnotes
Supplemental digital content is available for this article. Direct URL citations appear in the printed text and are provided in the HTML and PDF versions of this article on the journal’s Web site (www.acsm-msse.org).
REFERENCES
- 1.Joyner MJ, Coyle EF. Endurance exercise performance: the physiology of champions. J Physiol. 2008;586(1):35–44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Hoogkamer W, Kram R, Arellano CJ. How biomechanical improvements in running economy could break the 2-hour marathon barrier. Sports Med. 2017;47(9):1739–50. [DOI] [PubMed] [Google Scholar]
- 3.Joyner MJ, Hunter SK, Lucia A, Jones AM. Physiology and fast marathons. J Appl Physiol. 2020;128(4):1065–8. [DOI] [PubMed] [Google Scholar]
- 4.Vanhatalo A, Jones AM, Burnley M. Application of critical power in sport. Int J Sports Physiol Perform. 2011;6(1):128–36. [DOI] [PubMed] [Google Scholar]
- 5.Poole DC, Burnley M, Vanhatalo A, Rossiter HB, Jones AM. Critical power: an important fatigue threshold in exercise physiology. Med Sci Sports Exerc. 2016;48(11):2320–34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Jones AM, Burnley M, Black MI, Poole DC, Vanhatalo A. The maximal metabolic steady state: redefining the ‘gold standard’. Physiol Rep. 2019;7:1–16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Chidnok W, Dimenna FJ, Bailey SJ, Wilkerson DP, Vanhatalo A, Jones AM. Effects of pacing strategy on work done above critical power during high-intensity exercise. Med Sci Sports Exerc. 2013;45(7):1377–85. [DOI] [PubMed] [Google Scholar]
- 8.Jones AM, Wilkerson DP, Dimenna FJ, Fulford J, Poole DC. Muscle metabolic responses to exercise above and below the “critical power” assessed using 31P-MRS. Am J Physiol Regul Integr Comp Physiol. 2008;294(2):R585–93. [DOI] [PubMed] [Google Scholar]
- 9.Black MI Jones AM Blackwell JR, et al. Muscle metabolic and neuromuscular determinants of fatigue during cycling in different exercise intensity domains. J Appl Physiol. 2017;122(3):446–59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Poole DC, Ward SA, Gardner GW, Whipp BJ. Metabolic and respiratory profile of the upper limit for prolonged exercise in man. Ergonomics. 1988;31(9):1265–79. [DOI] [PubMed] [Google Scholar]
- 11.Muniz-Pumares D, Pedlar CR, Godfrey R, Glaister M. Accumulated oxygen deficit during exercise to exhaustion determined at different supramaximal work rates. Int J Sports Physiol Perform. 2017;12(3):351–6. [DOI] [PubMed] [Google Scholar]
- 12.Muniz-Pumares D, Pedlar C, Godfrey R, Glaister M. A comparison of methods to estimate anaerobic capacity: accumulated oxygen deficit and W’ during constant and all-out work-rate profiles. J Sports Sci. 2017;35(23):2357–64. [DOI] [PubMed] [Google Scholar]
- 13.Morton RH. The critical power and related whole-body bioenergetic models. Eur J Appl Physiol. 2006;96(4):339–54. [DOI] [PubMed] [Google Scholar]
- 14.Vanhatalo A, Fulford J, DiMenna FJ, Jones AM. Influence of hyperoxia on muscle metabolic responses and the power–duration relationship during severe-intensity exercise in humans: a 31P magnetic resonance spectroscopy study. Exp Physiol. 2010;95(4):528–40. [DOI] [PubMed] [Google Scholar]
- 15.Black MI, Durant J, Jones AM, Vanhatalo A. Critical power derived from a 3-min all-out test predicts 16.1-km road time-trial performance. Eur J Sport Sci. 2014;14(3):217–23. [DOI] [PubMed] [Google Scholar]
- 16.Bosquet L, Duchene A, Lecot F, Dupont G, Leger L. Vmax estimate from three-parameter critical velocity models: validity and impact on 800 m running performance prediction. Eur J Appl Physiol. 2006;97(1):34–42. [DOI] [PubMed] [Google Scholar]
- 17.Kranenburg KJ, Smith DJ. Comparison of critical speed determined from track running and treadmill tests in elite runners. Med Sci Sports Exerc. 1999;28(5):614–8. [DOI] [PubMed] [Google Scholar]
- 18.Nimmerichter A, Novak N, Triska C, Prinz B, Breese BC. Validity of treadmill-derived critical speed on predicting 5000-meter track-running performance. J Strength Cond Res. 2017;31(3):706–14. [DOI] [PubMed] [Google Scholar]
- 19.Jones AM, Vanhatalo A. The ‘critical power’ concept: applications to sports performance with a focus on intermittent high-intensity exercise. Sports Med. 2017;47(1 Suppl):65–78. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Florence S, Weir JP. Relationship of critical velocity to marathon running performance. Eur J Appl Physiol. 1997;75(3):274–8. [DOI] [PubMed] [Google Scholar]
- 21.Atkinson G, Peacock O, St Clair Gibson A, Tucker R. Distribution of power output during cycling: impact and mechanisms. Sports Med. 2007;37(8):647–67. [DOI] [PubMed] [Google Scholar]
- 22.Muniz-Pumares D, Karsten B, Triska C, Glaister M. Methodological approaches and related challenges associated with the determination of critical power and curvature constant. J Strength Cond Res. 2019;33(2):584–96. [DOI] [PubMed] [Google Scholar]
- 23.Dekerle J, Nesi X, Carter H. The distance–time relationship over a century of running Olympic performances: a limit on the critical speed concept. J Sports Sci. 2006;24(11):1213–21. [DOI] [PubMed] [Google Scholar]
- 24.Karsten B, Jobson SA, Hopker JG. Validity and reliability of critical power field testing. Eur J Appl Physiol. 2015;115(1):197–204. [DOI] [PubMed] [Google Scholar]
- 25.Minetti AE, Moia C, Roi GS, Susta D, Ferretti G. Energy cost of walking and running at extreme uphill and downhill slopes. J Appl Physiol. 2002;93(3):1039–46. [DOI] [PubMed] [Google Scholar]
- 26.Galbraith A, Hopker JG, Lelliott S, Diddams L, Passfield L. A single-visit field test of critical speed. Int J Sports Physiol Perform. 2014;9(6):931–5. [DOI] [PubMed] [Google Scholar]
- 27.Scherrer J, Monod H. Local muscle work and fatigue in man. J Physiol (Paris). 1960;52:419–501. [PubMed] [Google Scholar]
- 28.Murgatroyd SR, Ferguson C, Ward SA, Whipp BJ, Rossiter HB. Pulmonary O2 uptake kinetics as a determinant of high-intensity exercise tolerance in humans. J Appl Physiol. 2011;110(6):1598–606. [DOI] [PubMed] [Google Scholar]
- 29.Goulding RP, Roche DM, Marwood S. Prior exercise speeds pulmonary oxygen uptake kinetics and increases critical power during supine but not upright cycling. Exp Physiol. 2017;102(9):1158–76. [DOI] [PubMed] [Google Scholar]
- 30.Vanhatalo A Black MI Dimenna FJ, et al. The mechanistic bases of the power–time relationship: muscle metabolic responses and relationships to muscle fibre type. J Physiol. 2016;594(15):4407–23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Mitchell EA, Martin NRW, Bailey SJ, Ferguson RA. Critical power is positively related to skeletal muscle capillarity and type I muscle fibers in endurance-trained individuals. J Appl Physiol. 2018;125(3):737–45. [DOI] [PubMed] [Google Scholar]
- 32.Kordi M, Menzies C, Parker Simpson L. Relationship between power–duration parameters and mechanical and anthropometric properties of the thigh in elite cyclists. Eur J Appl Physiol. 2018;118(3):637–45. [DOI] [PubMed] [Google Scholar]
- 33.Weber CL, Schneider DA. Maximal accumulated oxygen deficit expressed relative to the active muscle mass for cycling in untrained male and female subjects. Eur J Appl Physiol. 2000;82(4):255–61. [DOI] [PubMed] [Google Scholar]
- 34.Keogh A, Smyth B, Caulfield B, Lawlor A, Berndsen J, Doherty C. Prediction equations for marathon performance: a systematic review. Int J Sports Physiol Perform. 2019;14(9):1159–69. [DOI] [PubMed] [Google Scholar]
- 35.Clark IE Vanhatalo A Bailey SJ, et al. Effects of two hours of heavy-intensity exercise on the power–duration relationship. Med Sci Sports Exerc. 2018;50(8):1658–68. [DOI] [PubMed] [Google Scholar]
- 36.Clark IE Vanhatalo A Thompson C, et al. Changes in the power–duration relationship following prolonged exercise: estimation using conventional and all-out protocols and relationship with muscle glycogen. Am J Physiol Regul Integr Comp Physiol. 2019;317(1):R59–67. [DOI] [PubMed] [Google Scholar]
- 37.Clark IE Vanhatalo A Thompson C, et al. Dynamics of the power–duration relationship during prolonged endurance exercise and influence of carbohydrate ingestion. J Appl Physiol. 2019;127(3):726–36. [DOI] [PubMed] [Google Scholar]
- 38.Hamlin MJ, Hopkins WG, Hollings SC. Effects of altitude on performance of elite track-and-field athletes. Int J Sports Physiol Perform. 2015;10(7):881–7. [DOI] [PubMed] [Google Scholar]
- 39.Pugh LG. The influence of wind resistance in running and walking and the mechanical efficiency of work against horizontal or vertical forces. J Physiol. 1971;213(2):255–76. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Smyth B. Fast starters and slow finishers: a large-scale data analysis of pacing at the beginning and end of the marathon for recreational runners. J Sport Anal. 2018;4(3):229–42. [Google Scholar]
- 41.Deaner RO, Carter RE, Joyner MJ, Hunter SK. Men are more likely than women to slow in the marathon. Med Sci Sports Exerc. 2014;47(3):607–16. [DOI] [PMC free article] [PubMed] [Google Scholar]