

Abstract
Despite constant exposure to various environmental stimuli, the ocular surface remains intact and uninflamed while maintaining the transparency of the cornea and its visual function. This ‘immune privilege’ of the ocular surface is not simply a result of the physical barrier function of the mucosal lining but, more importantly, is actively maintained through a variety of immunoregulatory mechanisms that prevent the disruption of immune homeostasis. In this review, we focus on essential molecular and cellular players that promote immune quiescence in steady-state conditions and suppress inflammation in disease-states. Specifically, we examine the interactions between the ocular surface and its local draining lymphoid compartment, by encompassing the corneal epithelium, corneal nerves and cornea-resident myeloid cells, conjunctival goblet cells, and regulatory T cells (Treg) in the context of ocular surface autoimmune inflammation (dry eye disease) and alloimmunity (corneal transplantation). A better understanding of the immunoregulatory mechanisms will facilitate the development of novel, targeted immunomodulatory strategies for a broad range of ocular surface inflammatory disorders.
Keywords: Immune regulation, Ocular surface, Inflammation, Autoimmunity, Alloimmunity
1. Introduction
The ocular surface consists of the corneal and conjunctival mucosal linings extending to the mucocutaneous junctions of the lid margins. The tear film - an aqueous medium enriched with lipids and mucins from meibomian glands and goblet cells, respectively, maintains the ocular interface with the external environment [1]. The ocular surface is known as an “immune-privileged” site: It holds the capability to mount effective immune defense, but it employs tight regulatory measures to prevent unnecessary local inflammatory responses to preserve corneal integrity and function. In this review, we discuss the following regulatory mechanisms that are critical to maintaining immune homeostasis of the ocular surface: 1) Cornea avascularity: The cornea must stay transparent and, therefore, be avascular to function normally. The lack of blood vessels limits the ingress of circulating blood leukocytes, while lack of lymphatic vessels impedes leukocytes from migrating out of the tissue. Thus, corneal avascularity essentially contributes to the immune privilege of the cornea [2]. 2) The healthy cornea is devoid of ‘mature’ leukocytes: The healthy cornea is devoid of lymphoid leukocytes but is endowed with a sizable number of myeloid leukocytes [3–8], which function in both innate and adaptive immunity in response to external insults. These resident leukocytes are phenotypically “immature”; they express limited pro-inflammatory cytokines and are limited in their ability to sensitize lymphoid cells and provoke effector T cell responses. Rather, they promote induction of immune tolerance at the ocular surface [5,6,9,10]. 3) Regulatory role of the corneal epithelium: The corneal epithelium prevents invasion of immunogenic molecules through its barrier function and actively contributes to immune quiescence regulation; this layer secretes various immunoregulatory factors that sustain the cornea’s avascularity and preserve the “immature” status of resident leukocytes at the ocular surface [11–14]. 4) Neuroimmune cross-talk at the ocular surface: More recently, immunoregulatory function of corneal nerves and their cross-talk with immune cells has drawn significant interest. Indeed, the cornea is the most densely innervated tissue in the human body, and proper corneal innervation with the physiological levels of neuropeptides at the ocular surface has been demonstrated essential to promote immune homeostasis in addition to regulating tear secretion and the blink reflex [15–17]. 5) Conjunctiva-associated lymphoid tissue (CALT) and goblet cells: Unlike the cornea, the conjunctiva harbors a diverse group of immune cells with dominant lymphoid cells (primarily T cells) during the steady-state conditions [18]. Unique to the conjunctiva, some resident T cells aggregate in dense foci in a form of mucosa-associated lymphoid tissue (MALT) and are specifically termed conjunctiva-associated lymphoid tissue (CALT) [19]. CALT increases in size and number after ocular antigen stimulation and is implicated in the induction of mucosal tolerance and protection from infectious and non-infectious insults [20]. In addition, a special type of epithelial cell within the conjunctiva is the goblet cell, which serves as the primary source for mucins that exert antimicrobial functions and induce tolerogenic dendritic cells (DC) [21,22]. 6) Regulatory T cells (Treg) in the regional lymph nodes: ‘Intraocular’ immune quiescence is primarily maintained through a phenomenon called anterior chamber-associated immune deviation (ACAID) that involves the spleen. However, the adaptive T cell immunity at the ‘ocular surface’ is mainly regulated within regional lymph nodes [23,24]. Indeed, suppression of effector T cells in the local lymphoid compartment is an essential mechanism maintaining the integrity of the ocular surface. In this regard, regulatory T cells (Treg) in the lymph nodes draining the ocular surface have been shown to potently suppress sensitization of naive T cells and function of activated T cells, thus preventing the loss of ocular surface immune quiescence [23,24]. Key cellular and molecular components participating in the immune regulation of the ocular surface are illustrated in Fig. 1.
Figure 1. The major cells and molecules promoting ocular surface immune homeostasis.
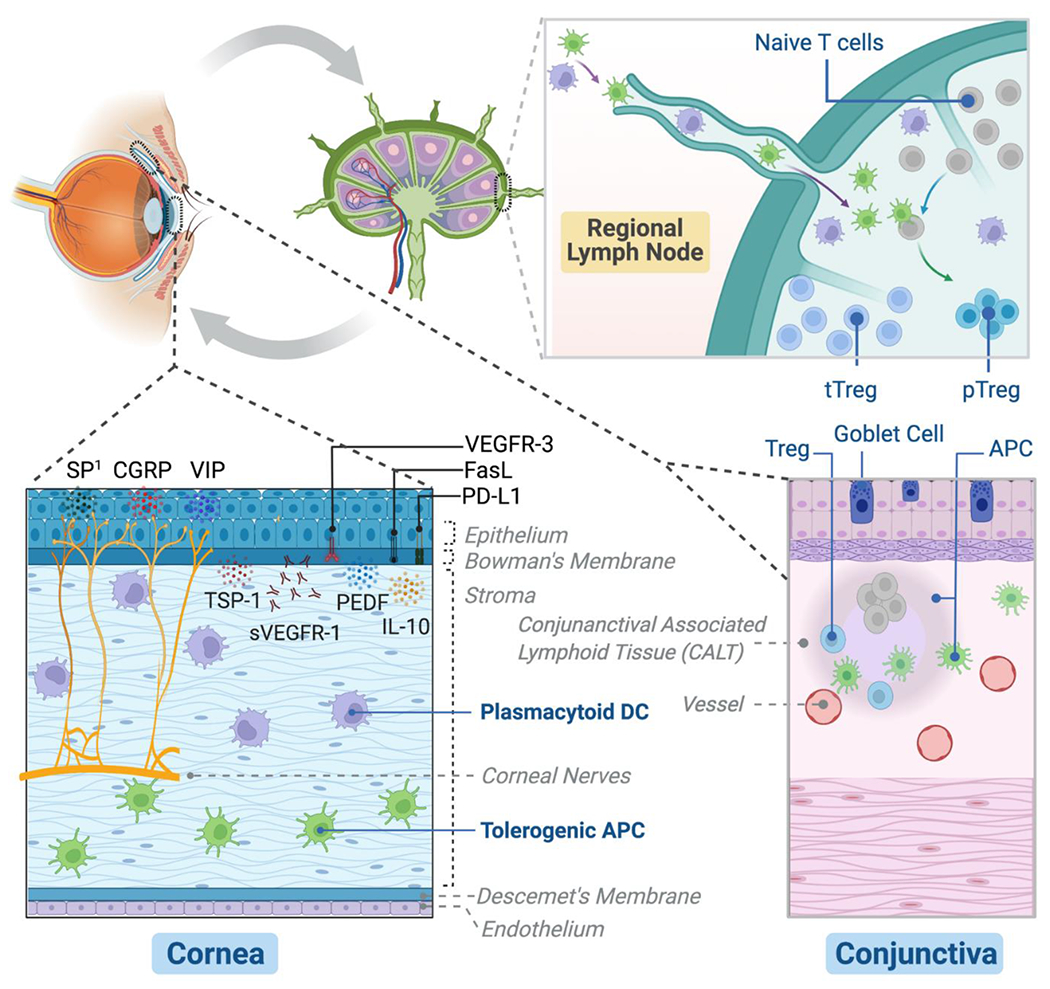
(1) Cornea: Epithelium constitutively expresses an array of immunoregulatory factors to suppress inflammatory cells and inhibit angiogenesis, including membrane-bound programmed death ligand-1 (PD-L1), Fas ligand (FasL), and vascular endothelial growth factor receptor-3 (VEGFR-3), as well as secreted pigment epithelial-derived factor (PEDF), thrombospondin-1 (TSP-1), soluble VEGFR-1 (sVEFGR-1), and IL-10. The tolerogenic antigen-presenting cells (APC) and plasmacytoid dendritic cells (DC) primarily residing in the stroma are critically involved in the induction of immune tolerance at both ocular surface and regional lymph nodes, and plasmacytoid DC additionally contribute to corneal nerve integrity and avascularity. The intense innervation of the cornea is essential for ocular surface barrier function and epithelial health with the support from the physiological level of substance P (SP) expressed by the nerves (1Excessive SP, on the contrary, promotes ocular surface inflammation and tissue damage). Other cornea nerve-derived neuropeptides, including vasoactive intestinal peptide (VIP) and calcitonin gene-related peptide (CGRP) exert important anti-inflammatory functions. (2) Conjunctiva: The specialized goblet cells in the epithelium are critical to maintaining tear film stability by secreting soluble mucins. It promotes ocular surface immune tolerance by inducing tolerogenic APC. In addition, conjunctiva-associated lymphoid tissue (CALT), which harbors regulatory T cells (Treg) is involved in the protective mucosal immune regulation. (3) Regional lymph nodes: Ocular surface adaptive immunity is primarily regulated in the local eye-draining lymph nodes by the specialized immunoregulatory cells – Treg, which include the larger thymus-derived Treg (tTreg) population and the smaller but more potent peripheral induced Treg (pTreg) population after antigen stimulation by tolerogenic APC or plasmacytoid DC.
2. Immunoregulatory function of corneal epithelium
The corneal epithelium is the frontline of innate ocular immunity. It forms a physical barrier against micro-organisms and environmental insults, maintained firmly by cellular structures including desmosomal junctions, hemidesmosomes, and basement membrane [25,26]. Additionally, there is increasing evidence demonstrating that the corneal epithelium has a critical immunoregulatory role in the eye. These cells constitutively express an array of immunoregulatory factors, including programmed death ligand-1 (PD-L1). Fas ligand (FasL), pigment epithelial-derived factor (PEDF), and thrombospondin-1 (TSP-1), among others [11–14].
PD-L1 binds to its receptor programmed death (PD)-1 on activated effector T cells, triggering their cell death and thereby suppressing T cell effector response [27]. PD-L1 is constitutively expressed in high levels by the corneal epithelium [11,28], but a variety of acute inflammatory insults can also transiently elevate its expression [11]. For instance, in the setting of corneal transplantation, grafts derived from allogeneic PD-L1 knockout (KO) mice are shown to be significantly more susceptible to immune rejection compared to the grafts from wild-type mice, primarily as a result of increased infiltration of effector T cells at the graft site [11]. PD-L1 KO mice show spontaneous T cell infiltration in the cornea, and these mice are prone to develop more severe dry eye disease (DED) [28]. Wild-type mice with DED show reduced expression of PD-L1 in their corneal epithelium, and blockade of PD-L1 further exacerbates ocular surface inflammation in DED [28]. These findings highlight the critical role of PD-L1 expressed by the corneal epithelium in inhibiting the ocular surface adaptive T cell immunity. Furthermore, PD-L1 expressed by the corneal epithelium binds to its other receptor, CD80, on microvascular endothelial cells and inhibits abnormal corneal blood vessel growth (hemangiogenesis), thereby maintaining the corneal avascularity [29].
FasL is a member of the tumor necrosis factor (TNF) family and functions as a negative regulator of T effectors and neovascularization. FasL mediates such roles by inducing apoptotic cell death through binding to its receptor, Fas, expressed by a variety of cells and tissues, including immune cells [30,31]. There are two forms of FasL: 1) The pro-inflammatory, membrane-bound form (mFasL); and 2) the anti-inflammatory, soluble form (sFasL) which antagonizes the function of mFasL [32]. The balance of sFasL:mFasL is a critical factor regulating their function: for instance, sFasL is the predominant form in the normal retina [33]. Although FasL is uniformly expressed by normal corneal epithelium and endothelium, it remains unclear which form(s) of FasL is predominant in the cornea [34].
PEDF belongs to the serine protease inhibitor family and is widely expressed by various ocular tissues, including the retina, choroid, ciliary body, and corneal epithelium/endothelium [14,35,36]. PEDF expressed by corneal epithelium has been shown to suppress the maturation and activation of corneal resident leucocytes effectively. Topical or systemic administration of recombinant PEDF significantly alleviates DED severity by suppressing innate inflammation, inhibiting adaptive effector T cell response, and promoting immunosuppressive functions of Treg in the local draining lymph nodes [14,37]. In mechanical and chemical corneal injury, topical application of PEDF or its derivatives effectively promotes wound healing through enhancing limbal stem cell proliferation and inhibits corneal neovascularization through suppressing vascular endothelial growth factor (VEGF) expression [38,39].
TSP-1 is an extracellular matrix protein constitutively expressed by corneal epithelium and endothelium [13,40,41]. It regulates ocular surface homeostasis by interacting with an array of receptors, growth factors, cytokines, and proteases. TSP-1 mediates its immunoregulatory function principally by transforming the latent form of transforming growth factor-β (TGF-β) to its functional activated form, which serves as a critical anti-inflammatory cytokine [42,43]. Reactive up-regulation of TSP-1 in corneal epithelium has been observed in DED, which plays a critical role in suppressing corneal resident leukocyte activation. Furthermore, topical application of recombinant TSP-1 or its peptide derivative has been demonstrated to suppress the immune-mediated ocular surface inflammation in both DED and allergic eye disease [13,44,45]. Studies employing exogenous TSP-1 supplementation or animal models with a deletion in endogenous TSP-1 implicate the critical role of TSP-1 in the induction of Treg and their functional maintenance, which is disrupted Treg in DED [44,46]. TSP-1 mainly engages Treg indirectly through activation of TGF-β, which potently modulates Treg differentiation. Similar to PEDF, TSP-1 also serves as a potent endogenous anti-angiogenic factor. Experimental data from TSP-1 null mice show that TSP-1 suppresses both corneal hemangiogenesis and lymphangiogenesis (abnormal lymphatic vessels growth) [47,48], the pathological processes promoting DED inflammation and endangering corneal graft rejection [49,50], by binding to CD36 on monocytes regulating their expressions of VEGF [48,51].
Emerging evidence has demonstrated that corneal epithelium is also the primary source of endogenous polyunsaturated fatty acids (PUFA)-derived specialized pro-resolving mediators (SPM) at the ocular surface, including lipoxin A4 (LXA4), neuroprotectin D1 (NPD1), and resolvin D1 (RvD1) [52,53]. SPM is a class of bioactive lipids metabolized from the substrates of essential PUFA, including arachidonic acid (ω-6 AA), docosahexaenoic acid (ω-3 DHA), eicosapentaenoic acid (ω-3 EPA), and docosapentaenoic acid (ω-3 DPA), with specialized functions of limiting inflammation and promoting tissue homeostasis primarily through autocrine and paracrine actions on target cells that encompass ocular surface epithelial and stromal cells as well as various types of immune cells such as macrophages, DC, and T cells [54,55]. Elimination of endogenous LXA4 has been shown to increase corneal neovascularization, and treatment with LXA4, its analogue, or RvD1 can effectively suppress corneal hemangiogenesis [54,56]. It has also been demonstrated that SPM promotes corneal epithelial cell proliferation and migration in vitro, and facilitates corneal wound healing and nerve regeneration in vivo [52,57–59]. As SPM are rapidly inactivated by cells in vitro, structural analogues of SPM have been designed to increase their stability and bioavailability for therapeutic studies. Topical treatment with resolvin E1 (RvE1) analogues or α-linoleic acid (ALA, a metabolic precursor for EPA and DHA) in pre-clinical models of DED has been demonstrated to effectively ameliorate corneal epitheliopathy, reduce goblet cell loss, and suppress inflammatory cell activation [60–62]. Pre-treatment of human donor corneas with a stable LXA4 analogue during storage significantly increases endothelial viability [63], and treatment of hosts with a stable RvD1 analogue in a murine model of corneal transplantation effectively enhances graft survival by suppressing conventional DC activation and allosensitization [64].
The normal cornea is free of both blood vessels and lymphatic vessels, which is critical to maintaining its optical function and immune quiescence. Abnormal hemangiogenesis in host cornea, often induced by inflammation, has been determined a high-risk factor for the survival of corneal transplantation [65]. The neovascularized host bed promotes the maturation of resident myeloid cells and increases the recruitment of peripheral innate immune cells, thus enhancing the sensitization of T cells, and supporting the migration of activated T cells to the graft site inducing graft rejection [24]. Corneal transplantation also induces the formation of corneal lymphangiogenesis [50], which can provide access for activated myeloid cells at the ocular surface to regional lymphoid compartments where they prime and activate adaptive T cells, thus critically contributing to immune rejection of corneal grafts [66]. In addition, selective lymphangiogenesis has been observed in the DED cornea that starts from the peripheral cornea and advances into the central cornea with the progression of the disease, dependent on the interaction of VEGF-C and VEGF-D and their receptor VEGFR-3 [49,67,68]. These lymphatic vessels serve as an afferent pathway for activated myeloid cells to relocate to the local draining lymph nodes mediating T cell activation. Corneal epithelium produces a variety of factors, including the aforementioned, to work in concert to maintain corneal avascularity and clarity. Normal corneal epithelium constitutively expresses soluble vascular endothelial growth factor receptor-1 (sVEGFR-1, also known as sflt-1) to inhibit VEGF-A mediated new blood vessel formation by serving as an endogenous VEGF-A trap [69]. In addition, strong expression of VEGFR-3 by healthy corneal epithelium functions as a “sink” for VEGF-C and VEGF-D to prevent their interaction with VEGFR-3 on vascular endothelium, thus preventing corneal new blood and lymphatic vessels formation [70].
More recently, the immunoregulatory functions of limbal melanocytes, an essential component of the corneal epithelial stem cell niche, have been explored. In addition to their well-known protective roles in preventing limbal epithelial stem/progenitor cells from UV damages, limbal melanocytes have been shown in a series of in vitro studies to effectively suppress T cell proliferation and cytokine production primarily through cell contact-dependent mechanisms. They additionally have been shown to inhibit angiogenesis via preventing vascular endothelial cell proliferation and capillary formation [71]. These findings suggest important immunoregulatory contributions of the limbal melanocytes to limbal niche homeostasis.
Notably, the anti-inflammatory properties of corneal epithelium can be overturned in a variety of inflammatory conditions, such as DED, leading to the production and release of pro-inflammatory cytokines by the cornea, including TNF-α, IL-1β, and IL-6 [72] that lead to the disruption of immune quiescence of ocular surface.
3. Immunoregulatory function of corneal resident myeloid leukocytes
The normal cornea has no T or B lymphocytes but is endowed with bone marrow-derived principally myeloid immune cells characterized by a CD11b+CD3−CD19− phenotype, distributed predominantly at the periphery and decreasing gradually toward the center [6,9]. These resident myeloid cells comprise a major population of CD11b+CD11c− macrophages/monocytes in the deep stroma and CD11b+CD11c+ dendritic cells (DC) in the anterior stroma, along with a few CD11blo/− CD11c+ Langerhans cells in the epithelium [3–8]. These tissue-resident cells play essential roles in activating adaptive immune response by functioning as antigen-presenting cells (APC). Corneal APC are sentinels of the immune system at the interface of the ocular surface and outside environment, and the preponderance of them display an “immature” status characterized by low expression levels of MHC class II (MHC-II) and the absence of co-stimulatory molecules B7 (CD80 and CD86) and CD40, especially amongst those residing in the central corneal stroma [6,9], rendering them immune quiescent and even maturation-resistant thus playing regulatory functions in the setting of inflammatory insults. Those cells that possess diminished antigen presentation, production of regulatory cytokines, and generation and expansion of Treg are defined as tolerogenic APC, which are critically involved in promoting transplant tolerance and allograft survival [73,74]. Expression of TSP-1 by corneal APC has been identified as one of the mechanisms rendering these cells as tolerogenic, via preventing these cells from maturation and impairing their capacity to migrate, thus inhibiting T cell allosensitization [75]. Enrichment of tolerogenic APC in donor corneas by ex vivo treatment of corneal buttons with IL-10 and TGF-β has been successfully demonstrated to suppress allosensitization and improve the corneal allograft survival in high-risk transplantation (neovascularized host bed) [76]. However, on the other hand, immature corneal resident APC can be stimulated by a variety of up-regulated innate factors including mast cells [77] to acquire a highly mature phenotype and mobilize to regional lymphoid tissues where they prime T cells and initiate adaptive immunity, promoting the disruption of immune homeostasis at the ocular surface, in particular leading to DED and corneal graft rejection [62,78,79].
In contrast to conventional DC (cDC), plasmacytoid dendritic cells (pDC), characterized by CD45+ PDCA-1+ CD45R/B220+ CD11clow CD11b− in murine and CD45+ BDCA-2+ BDCA-4+ CD11clow in humans, are a special type of innate cell residing in the anterior stroma of cornea and limbus. pDC are primarily present in the peripheral blood and secondary lymphoid tissues, and although they are a rare population, they constitute about 15-25% of total corneal CD45+ cells with immature phenotypes during steady state [80,81]. Originally identified as a critical source of type I IFN, pDC have been shown to play critical immunoregulatory roles. In vitro co-culture of pDC with Treg effectively sustains Treg expression of the function marker Foxp3, demonstrating that pDC are a key type of cell capable of inducing tolerance by promoting generation of Treg and preventing their dysfunction, probably through IFN-α independent mechanisms [81]. pDC also express the co-inhibitory molecule PD-L1 [81,82], which can limit effector T cell responses via interacting with PD-1 on T cells. In fact, depletion of corneal pDC in the setting of corneal transplantation has been demonstrated to increase corneal allograft opacity and effector T cell infiltration, as well as decrease Foxp3 expression by Treg in the draining lymph nodes [81]. In addition, corneal pDC contribute to neuroimmune cross-talk by maintaining the homeostasis of corneal nerves and promoting nerve regeneration, at least partially through secretion of the neurotrophic nerve growth factor (NGF) [81]. pDC residing in the corneal limbus also play significant roles in sustaining corneal angiogenic privilege through secreting a variety of anti-angiogenic factors such as TSP-1 [81].
4. Immunoregulatory function of corneal nerves and neuropeptides
The cornea is densely innervated by the ophthalmic branch of the trigeminal nerve, which enters the corneal stroma and further branches into the epithelium [83]. The integrity of corneal nerves is essential for relaying the sensory signals and providing epithelial-trophic factors to maintain ocular surface homeostasis, including critical regulation of corneal epithelial cell and limbal stem cell survival [84–86]. Corneal neuropathy has been reported to contribute to corneal epitheliopathy in DED [87–90]. In addition, normal corneal innervation is required to suppress corneal neovascularization as corneal denervation leads to significant corneal angiogenesis and inflammation [91].
Importantly, nerve-derived neuropeptides play an important role in regulating immunity and angiogenesis. Substance P (SP) is one of the major neuropeptides constitutively secreted by nerve endings in the normal cornea, and a physiological level of SP is required for corneal epithelial homeostasis [15–17]. Complete abrogation of SP function by genetically knocked-out NK1R, the preferred SP receptor, has been shown to cause loss of corneal epithelial cells [92], and topical treatment with SP-derived peptide in neurotrophic keratopathy, a corneal disease characterized by impaired corneal nerve function which may lead to decreased SP production at the ocular surface, has been shown to promote the closure of the corneal epithelial defect [93]. However, the protective roles of SP can be overturned when there is an excessive amount of SP at the ocular surface that is higher than the physiological level, primarily through promoting corneal resident APC maturation and mobilization, recruiting peripheral leukocytes to infiltrate the cornea, and inducing corneal neovascularization directly on parent vessels and indirectly through activating innate immune cells including mast cells to secret VEGF-A, all of which finally leads to neurogenic inflammation and propagation of an inflammatory cascade, as reported in DED and corneal injuries [94–102]. Heightened expression of SP subsequent to the severance of corneal nerves during surgery has been reported to disrupt ocular immune privilege and promotes corneal allograft rejection [103].
Vasoactive intestinal peptide (VIP) is another important neuropeptide constitutively present in the cornea that exerts neuroimmunomodulatory function [104]. In addition, VIP has been shown effective to maintain the corneal endothelial cell integrity during donor cornea tissue storage, through preventing oxidative stress- or inflammatory cytokines-induced apoptosis, which is critical for a successful corneal transplantation. Further, intracameral administration of VIP has been demonstrated to preserve corneal endothelial cells and improve the allograft survival after corneal transplantation [105]. Calcitonin gene-related peptide (CGRP) is also abundantly expressed by corneal nerves, primarily by sensory nerves [106,107]. Reduced CGRP-expressing corneal nerves has been associated with DED progression [108].
It is well-known that α-melanocyte-stimulating hormone (α-MSH) present in the aqueous humor plays essential roles in modulating intraocular immunity [109,110]. Recently, β-MSH has been detected in healthy human tears at a level about 100-fold lower than that in the aqueous humor [111,112], suggesting a role of α-MSH in regulating ocular surface immune homeostasis. In vitro treatment of peripheral blood mononuclear cells from patients with perennial allergic conjunctivitis with α-MSH significantly decreased the activated effector T cells and Th2-associated cytokines while increasing the Treg population [111]. Topical application of α-MSH in a DED model has been shown to effectively improve corneal integrity and tear secretion, preserve conjunctival goblet cells, as well as suppress the expression of inflammatory cytokines [113]. In addition, our group has shown that addition of α-MSH to standard human donor cornea storage medium effectively protects against corneal endothelial loss secondary to oxidative and pro-inflammatory cytokine-induced stresses [114]. Furthermore, subconjunctival injection of α-MSH to mice undergoing corneal transplantation significantly improves their graft survival, suppresses allosensitization of their T cells, and reduces the graft infiltration with inflammatory cells [115].
5. Immunoregulatory function of conjunctival goblet cells and CALT
Goblet cells are specialized epithelial cells in the conjunctiva serving as the primary source of mucins for tear film that covers the ocular surface [116]. Disrupted corneal barrier function is a major pathological feature in DED visualized by corneal fluorescein staining in the clinic, and decreased numbers and atrophy of conjunctival goblet cells have been consistently observed in DED [117–119]. More recently, conjunctival goblet cells have been shown to importantly contribute to local immune tolerance to foreign antigen stimulation, presumably via conditioning conjunctival APC to acquire tolerogenic properties [22]. The underlying mechanisms by which goblet cells interact with APC have yet to be determined, although studies from intestinal goblet cells suggest that goblet cell-associated antigen passages (GAPs) mediate local antigen delivery by goblet cells to APC, promoting tolerogenic tissue cDC1 [120,121]; similar GAPs have recently been identified in the conjunctiva as well [122]. In addition, MUC2, which is expressed by goblet cells in both the gut and conjunctiva [123,124] has been shown to be uptaken by intestinal DC, leading to inhibition of pro-inflammatory cytokine production and induction of tolerogenic properties of the DC [123]. Conjunctival goblet cells can produce active form of TGF-β, a critical immunoregulatory cytokine that may allow goblet cells to imprint a tolerogenic phenotype onto APC [125]. Another immunoregulatory factor secreted by conjunctival goblet cells is retinoic acid, which inhibits maturation of bone marrow-derived DC and their production of the inflammatory cytokine IL-12, through binding to the retinoid receptor X (RXR) [126,127]. Additional putative functions of conjunctival goblet cells include promotion of CALT function [128]. CALT organization consists of a lymphoid follicle with a T-zone containing CD4+ cells and a B-zone with B cells and follicular dendritic cells, as well as adjacent high-endothelial venules (HEVs) and lymphatics [20]. CALT is believed to be a normal and noninflammatory component of the ocular surface [129], and is involved in the maintenance of protective mucosal immune regulation and contributes to immune tolerance at ocular surface [130]. The presence of CD4+CD25+Foxp3+ Treg within CALT supports an immune regulatory function of CALT [20]. Experimental studies have shown an increase in number and size of CALT in ocular allergy but a decrease in DED [131]. In contrast, human studies show increased CALT in both DED and ocular allergy patients, although these in vivo examinations need to be validated by immunohistology [132]. The specific functions of CLAT in ocular surface disease and immune regulation deserve further investigations.
6. Immunoregulatory function of Treg
Regional eye-draining lymph nodes are the primary site where adaptive immunity is activated and regulated in ocular surface inflammation. Once the immature status of ocular surface resident APC is overridden by inflammatory stimulus, these APC become activated and mobilized, migrating to the local draining lymph nodes and priming naive CD4+ T cells through providing antigenic and co-stimulatory signals, as well as secreting T cell-polarizing cytokines [133–138], leading to generation of effector Th1, Th2 or Th17 cells as dominant type of central pathogenic cells mediating corneal allograft rejection, ocular allergy or DED, respectively [23,139–146]. The principal immunoregulatory mechanism that restricts and suppresses pro-inflammatory effector T cell response is mediated by the specialized CD4+ regulatory T cells (Treg), characterized by high expression of CD25 (the high affinity IL-2 receptor to maintain survival and proliferation of Treg) and Foxp3 (the lineage transcriptional factor for the development and function of Treg) [147,148]. The predominant Treg population residing in the lymphoid tissues is developed during the normal process of T cell maturation in the thymus, and thus is termed thymus-derived Treg (tTreg) or “natural Treg” (nTreg). In addition, a smaller but antigen-specific population that is induced in the periphery from naive T cells in the presence of IL-2 and TGF-β or after encounters with foreign antigens, termed iTreg for in vitro generated or pTreg for in vivo generated, may play more robust regulatory functions [149–151].
Reduced number and impaired function of Treg, especially pTreg, has been reported in corneal allograft rejection, characterized by defects in both contact-dependent (decreased expression of CTLA-4) and –independent suppression (reduced secretion of IL-10 and TGF-β) [152]. In fact, hosts with accepted grafts show enhanced Treg suppressive function [153], and adoptive transfer of pTreg from these hosts has been shown to effectively prevent graft rejection in high-risk setting [152]. In addition, local subconjunctival injection of nTreg has been demonstrated to significantly suppress Th1 response and improve corneal graft survival, and the injected Treg migrate to ipsilateral cornea, conjunctiva, and draining lymph nodes, suggesting that Treg suppress both effector T cell induction in regional lymphoid tissues and function of activated Th1 cells at the ocular surface [154]. Alternatively, systemic administration of low-dose IL-2 is another promising therapeutic strategy to effectively expand Treg and enhance their function (increased CTLA-4 and IL-10 expressions), and thus improve corneal allograft survival by inhibiting effector Th1-mediated graft attack [155].
In mouse models of ocular allergy, depletion of Treg by anti-CD25 antibody treatment has been demonstrated to exacerbate the allergic disease [156,157]. On the other hand, the high dose antigen sensitization-achieved suppression of allergic conjunctivitis is dependent on the induced Treg expansion and their augmented function in suppressing Th2 response [157]. Consistently, decreased expression of Foxp3 in CD4+ T cells has been reported in patients with allergic conjunctivitis, and these Treg are defective in suppressing effector Th2 response [158,159].
Treg play an essential role in suppressing Th17-mediated inflammation and maintaining immune homeostasis. Dysfunction of Treg in suppressing effector Th17 cells critically contributes to induction of ocular surface inflammation in DED [143,160]. Increased expression of SP in draining lymph nodes has been shown as one of underlying mechanisms directly suppressing Treg function, and in DED there is increased NK1R+ Treg that is functionally significantly compromised as compared to NK1R− Treg [161]. Depletion of Treg using an anti-CD25 antibody or CD25 knockout mice leads to worsened DED, while reconstitution of nTreg results in resistance to disease induction [162–165]. However, it remains to be determined whether Treg dysfunction persists over the long term (i.e. is it spontaneously reversible through dynamic microenvironmental cues) and contributes to dysregulated T cell memory that mediates the chronicity of ocular surface inflammation in DED [166]. Evidence from human study suggests a positive answer, as disruption of Th17/Treg balance in Sjogren’s syndrome patients has been reported [167,168]. Further, low-dose IL-2 treatment has been demonstrated in a pilot study to successfully restore the Th17/Treg balance and Treg function, along with reduced use of glucocorticoid and disease-modifying anti-rheumatic drugs in the patients [168].
7. Conclusions and perspectives
The maintenance of ocular surface homeostasis by a variety of immunoregulatory mechanisms is critical to avoid excessive inflammation that may cause tissue damage and impairment of vision. Novel therapeutic strategies harnessing these critical mechanisms can be designed to enhance the immune regulation resulting in restoration of ocular surface homeostasis in a myriad of ocular surface diseases. In addition to the mainstay of the ocular surface-regional lymphoid compartment axis we have systemically reviewed, the potential immune regulation in lacrimal glands that contribute to tear film stability and ocular surface integrity has yet to be studied. Outside local mechanisms, systemic modulation of ocular surface health has been increasingly recognized. For example, age-related “immunosenescence” is associated with dysregulated Treg population and function that may contribute to the higher susceptibility to DED in the elderly [169]. Additionally, although the ocular surface is a relatively sterile site with low bacterial load, normal gut microbiota has been demonstrated to provide important protection in ocular surface homeostasis, and fecal microbiota transplant has been shown to dampen DED severity [170]. The understanding of systemic regulatory mechanisms is rapidly evolving, and these may provide added benefits to local immunotherapy for ocular surface disorders in translational and clinical studies.
Highlights.
Active regulation operates within the ocular surface – regional lymph nodes axis.
Corneal epithelium-derived factors promote avascularity and immune quiescence.
Corneal resident ‘immature’ myeloid cells promote induction of immune tolerance.
Corneal nerves and conjunctival goblet cells modulate ocular surface immunity.
Regulatory T cells in regional lymph nodes critically suppress adaptive immunity.
Funding source:
This work was supported by NIH grants EY20889 (RD) and EY012963 (RD) and NIH core grant P30EY003790.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Conflict of interest:
R.D. is consultant to Novartis, GSK, and Kala and holds equity in Claris Biotherapeutics, Aramis Biosciences, GelMEDIX, and Kera Therapeutics. Massachusetts Eye and Ear owns intellectual property related to blockade of substance P in ocular immunoinflammatory diseases.
References
- [1].Bron AJ, de Paiva CS, Chauhan SK, Bonini S, Gabison EE, Jain S, et al. TFOS DEWS II pathophysiology report. Ocul Surf 2017;15:438–510. 10.1016/j.jtos.2017.05.011. [DOI] [PubMed] [Google Scholar]
- [2].Cursiefen C Immune privilege and angiogenic privilege of the cornea. Chem Immunol Allergy 2007;92:50–7. 10.1159/000099253. [DOI] [PubMed] [Google Scholar]
- [3].Hamrah P, Liu Y, Zhang Q, Dana MR. The corneal stroma is endowed with a significant number of resident dendritic cells. Investig Ophthalmol Vis Sci 2003;44:581–9. 10.1167/iovs.02-0838. [DOI] [PubMed] [Google Scholar]
- [4].Hattori T, Chauhan SK, Lee H, Ueno H, Dana R, Kaplan DH, et al. Characterization of langerin-expressing dendritic cell subsets in the normal cornea. Investig Ophthalmol Vis Sci 2011;52:4598–604. 10.1167/iovs.10-6741. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [5].Hattori T, Takahashi H, Dana R. Novel insights into the immunoregulatory function and localization of dendritic cells. Cornea 2016;35:S49–54. 10.1097/ICO.0000000000001005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [6].Hamrah P, Zhang Q, Liu Y, Dana MR. Novel characterization of MHC class II-negative population of resident corneal Langerhans cell-type dendritic cells. Investig Ophthalmol Vis Sci 2002;43:639–46. [PubMed] [Google Scholar]
- [7].Yamagami S, Ebihara N, Usui T, Yokoo S, Amano S. Bone marrow-derived cells in normal human corneal stroma. Arch Ophthalmol 2006;124:62–9. 10.1001/archopht.124.1.62. [DOI] [PubMed] [Google Scholar]
- [8].Yamagami S, Yokoo S, Usui T, Yamagami H, Amano S, Ebihara N. Distinct populations of dendritic cells in the normal human donor corneal epithelium. Investig Ophthalmol Vis Sci 2005;46:4489–94. 10.1167/iovs.05-0054. [DOI] [PubMed] [Google Scholar]
- [9].Barabino S, Chen Y, Chauhan S, Dana R. Ocular surface immunity: Homeostatic mechanisms and their disruption in dry eye disease. Prog Retin Eye Res 2012;31:271–85. 10.1016/j.preteyeres.2012.02.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [10].Lutz MB, Schuler G. Immature, semi-mature and fully mature dendritic cells: Which signals induce tolerance or immunity? Trends Immunol 2002;23:445–9. 10.1016/S1471-4906(02)02281-0. [DOI] [PubMed] [Google Scholar]
- [11].Shen L, Jin Y, Freeman GJ, Sharpe AH, Dana MR. The Function of Donor versus Recipient Programmed Death-Ligand 1 in Corneal Allograft Survival. J Immunol 2007;179:3672–9. 10.4049/jimmunol.179.6.3672. [DOI] [PubMed] [Google Scholar]
- [12].Griffith TS, Brunner T, Fletcher SM, Green DR, Ferguson TA. Fas ligand-induced apoptosis as a mechanism of immune privilege. Science (80-.) 1995;270:1189–92. 10.1126/science.270.5239.1189. [DOI] [PubMed] [Google Scholar]
- [13].Tan X, Chen Y, Foulsham W, Amouzegar A, Inomata T, Liu Y, et al. The immunoregulatory role of corneal epithelium-derived thrombospondin-1 in dry eye disease. Ocul Surf 2018;16:470–7. 10.1016/j.jtos.2018.07.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [14].Singh RB, Blanco T, Mittal SK, Taketani Y, Chauhan SK, Chen Y, et al. Pigment Epithelium-derived Factor secreted by corneal epithelial cells regulates dendritic cell maturation in dry eye disease. Ocul Surf 2020;18:460–9. 10.1016/j.jtos.2020.05.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [15].Yamada M, Ogata M, Kawai M, Mashima Y, Nishida T. Substance P and its metabolites in normal human tears. Investig Ophthalmol Vis Sci 2002;43:2622–5. [PubMed] [Google Scholar]
- [16].Yamada M, Ogata M, Kawai M, Mashima Y, Nishida T. Substance P in human tears. Cornea, vol. 22, 2003. 10.1097/00003226-200310001-00007. [DOI] [PubMed] [Google Scholar]
- [17].Suvas S Role of Substance P Neuropeptide in Inflammation, Wound Healing, and Tissue Homeostasis. J Immunol 2017;199:1543–52. 10.4049/jimmunol.1601751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [18].Hingorani M, Metz D, Lightman SL. Characterisation of the normal conjunctival leukocyte population. Exp Eye Res 1997;64:905–12. 10.1006/exer.1996.0280. [DOI] [PubMed] [Google Scholar]
- [19].Knop N, Knop E. Conjunctiva-associated lymphoid tissue in the human eye. Investig Ophthalmol Vis Sci 2000;41:1270–9. [PubMed] [Google Scholar]
- [20].Siebelmann S, Gehlsen U, Hüttmann G, Koop N, Bölke T, Gebert A, et al. Development, alteration and real time dynamics of conjunctiva-associated lymphoid tissue. PLoS One 2013;8. 10.1371/journal.pone.0082355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [21].Gordon YJ, Romanowski EG, McDermott AM. Mini review: A review of antimicrobial peptides and their therapeutic potential as anti-infective drugs. Curr Eye Res 2005;30:505–15. 10.1080/02713680590968637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [22].Ko BY, Xiao Y, Barbosa FL, de Paiva CS, Pflugfelder SC. Goblet cell loss abrogates ocular surface immune tolerance. JCI Insight 2018;3. 10.1172/jci.insight.98222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [23].Chauhan SK, Dana R. Role of Th17 cells in the immunopathogenesis of dry eye disease. Mucosal Immunol 2009;2:375–6. 10.1038/mi.2009.21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [24].Amouzegar A, Chauhan SK, Dana R. Alloimmunity and Tolerance in Corneal Transplantation. J Immunol 2016;196:3983–91. 10.4049/jimmunol.1600251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [25].Henriksson JT, McDermott AM, Bergmanson JPG. Dimensions and morphology of the cornea in three strains of mice. Investig Ophthalmol Vis Sci 2009;50:3648–54. 10.1167/iovs.08-2941. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [26].Bergmanson JPG, Texas Eye Research and Technology Center. Clinical ocular anatomy and physiology. 15th ed. Houston: Texas Eye Research and Technology Center; 2009. [Google Scholar]
- [27].Freeman GJ, Long AJ, Iwai Y, Bourque K, Chernova T, Nishimura H, et al. Engagement of the PD-1 immunoinhibitory receptor by a novel B7 family member leads to negative regulation of lymphocyte activation. J Exp Med 2000;192:1027–34. 10.1084/jem.192.7.1027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [28].El-Annan J, Goyal S, Zhang Q, Freeman GJ, Sharpe AH, Dana R. Regulation of T-cell chemotaxis by programmed Death-Ligand 1 (PD-L1) in dry eye-associated corneal inflammation. Investig Ophthalmol Vis Sci 2010;51:3418–23. 10.1167/iovs.09-3684. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [29].Jin Y, Chauhan SK, Annan JEI, Sage PT, Sharpe AH, Dana R. A novel function for programmed death ligand-1 regulation of angiogenesis. Am J Pathol 2011;178:1922–9. 10.1016/j.ajpath.2010.12.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [30].Suda T, Takahashi T, Golstein P, Nagata S. Molecular cloning and expression of the fas ligand, a novel member of the tumor necrosis factor family. Cell 1993. ;75:1169–78. 10.1016/0092-8674(93)90326-L. [DOI] [PubMed] [Google Scholar]
- [31].Ferguson TA, Griffith TS. A vision of cell death: Fas ligand and immune privilege 10 years later. Immunol Rev 2006;213:228–38. 10.1111/j.1600-065X.2006.00430.x. [DOI] [PubMed] [Google Scholar]
- [32].Hohlbaum AM, Moe S, Marshak-Rothstein A. Opposing effects of transmembrane and soluble Fas ligand expression on inflammation and tumor cell survival. J Exp Med 2000;191:1209–19. 10.1084/jem.191.7.1209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [33].Krishnan A, Fei F, Jones A, Busto P, Marshak-Rothstein A, Ksander BR, et al. Overexpression of Soluble Fas Ligand following Adeno-Associated Virus Gene Therapy Prevents Retinal Ganglion Cell Death in Chronic and Acute Murine Models of Glaucoma. J Immunol 2016;197:4626–38. 10.4049/jimmunol.1601488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [34].Podskochy A, Fagerholm P. The expression of fas ligand protein in ultraviolet-exposed rabbit corneas. Cornea 2002;21:91–4. 10.1097/00003226-200201000-00019. [DOI] [PubMed] [Google Scholar]
- [35].Dawson DW, Volpert OV, Gillis P, Crawford SE, Xu HJ, Benedict W, et al. Pigment epithelium-derived factor: A potent inhibitor of angiogenesis. Science (80-) 1999;285:245–8. 10.1126/science.285.5425.245. [DOI] [PubMed] [Google Scholar]
- [36].Karakousis PC, John SK, Behling KC, Surace EM, Smith JE, Hendrickson A, et al. Localization of pigment epithelium derived factor (PEDF) in developing and adult human ocular tissues. Mol Vis 2001;7:154–63. [PubMed] [Google Scholar]
- [37].Singh RB, Blanco T, Mittal SK, Alemi H, Chauhan SK, Chen Y, et al. Pigment Epithelium–Derived Factor Enhances the Suppressive Phenotype of Regulatory T Cells in a Murine Model of Dry Eye Disease. Am J Pathol 2021;191:720–9. 10.1016/j.ajpath.2021.01.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [38].Ho TC, Chen SL, Wu JY, Ho MY, Chen LJ, Hsieh JW, et al. PEDF promotes self-renewal of limbal stem cell and accelerates corneal epithelial wound healing. Stem Cells 2013;31:1775–84. 10.1002/stem.1393. [DOI] [PubMed] [Google Scholar]
- [39].Matsui T, Nishino Y, Maeda S, ichi Yamagishi S. PEDF-derived peptide inhibits corneal angiogenesis by suppressing VEGF expression. Microvasc Res 2012;84:105–8. 10.1016/j.mvr.2012.02.006. [DOI] [PubMed] [Google Scholar]
- [40].Sekiyama E, Nakamura T, Cooper LJ, Kawasaki S, Hamuro J, Fullwood NJ, et al. Unique distribution of thrombospondin-1 in human ocular surface epithelium. Investig Ophthalmol Vis Sci 2006;47:1352–8. 10.1167/iovs.05-1305. [DOI] [PubMed] [Google Scholar]
- [41].Hiscott P, Seitz B, Schlötzer-Schrehardt U, Naumann GOH. Immunolocalisation of thrombospondin 1 in human, bovine and rabbit cornea. Cell Tissue Res 1997;289:307–10. 10.1007/s004410050877. [DOI] [PubMed] [Google Scholar]
- [42].Crawford SE, Stellmach V, Murphy-Ullrich JE, Ribeiro SMF, Lawler J, Hynes RO, et al. Thrombospondin-1 is a major activator of TGF-β1 in vivo. Cell 1998;93:1159–70. 10.1016/S0092-8674(00)81460-9. [DOI] [PubMed] [Google Scholar]
- [43].Ribeiro SMF, Poczatek M, Schultz-Cherry S, Villain M, Murphy-Ullrich JE. The activation sequence of thrombospondin-1 interacts with the latency- associated peptide to regulate activation of latent transforming growth factor-β. J Biol Chem 1999;274:13586–93. 10.1074/jbc.274.19.13586. [DOI] [PubMed] [Google Scholar]
- [44].Contreras Ruiz L, Mir FA, Turpie B, Masli S. Thrombospondin-derived peptide attenuates Sjögren’s syndrome-associated ocular surface inflammation in mice. Clin Exp Immunol 2017;188:86–95. 10.1111/cei.12919. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [45].Smith RE, Reyes NJ, Khandelwal P, Schlereth SL, Lee HS, Masli S, et al. Secondary allergic T cell responses are regulated by dendritic cell-derived thrombospondin-1 in the setting of allergic eye disease. J Leukoc Biol 2016;100:371–80. 10.1189/jlb.3a0815-357rr. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [46].Turpie B, Yoshimura T, Gulati A, Rios JD, Dartt DA, Masli S. Sjögren’s syndrome-like ocular surface disease in thrombospondin-1 deficient mice. Am J Pathol 2009;175:1136–47. 10.2353/ajpath.2009.081058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [47].Cursiefen C, Masli S, Ng TF, Dana MR, Bornstein P, Lawler J, et al. Roles of thrombospondin-1 and -2 in regulating corneal and iris angiogenesis. Investig Ophthalmol Vis Sci 2004;45:1117–24. 10.1167/iovs.03-0940. [DOI] [PubMed] [Google Scholar]
- [48].Cursiefen C, Maruyama K, Bock F, Saban D, Sadrai Z, Lawler J, et al. Thrombospondin 1 inhibits inflammatory lymphangiogenesis by CD36 ligation on monocytes. J Exp Med 2011;208:1083–92. 10.1084/jem.20092277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [49].Goyal S, Chauhan SK, El Annan J, Nallasamy N, Zhang Q, Dana R. Evidence of corneal lymphangiogenesis in dry eye disease: A potential link to adaptive immunity? Arch Ophthalmol 2010;128:819–24. 10.1001/archophthalmol.2010.124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [50].Cursiefen C, Cao J, Chen L, Liu Y, Maruyama K, Jackson D, et al. Inhibition of hemangiogenesis and lymphangiogenesis after normal-risk corneal transplantation by neutralizing VEGF promotes graft survival. Investig Ophthalmol Vis Sci 2004;45:2666–73. 10.1167/iovs.03-1380. [DOI] [PubMed] [Google Scholar]
- [51].Jimönez B, Volpert OV, Crawford SE, Febbraio M, Silverstein RL, Bouck N. Signals leading to apoptosis-dependent inhibition of neovascularization by thrombospondin-1. Nat Med 2000;6:41–8. 10.1038/71517. [DOI] [PubMed] [Google Scholar]
- [52].Gronert K, Maheshwari N, Khan N, Hassan IR, Dunn M, Schwartzman ML. A role for the mouse 12/15-lipoxygenase pathway in promoting epithelial wound healing and host defense. J Biol Chem 2005. 10.1074/jbc.M410638200. [DOI] [PubMed] [Google Scholar]
- [53].English JT, Norris PC, Hodges RR, Dartt DA, Serhan CN. Identification and Profiling of Specialized Pro-Resolving Mediators in Human Tears by Lipid Mediator Metabolomics. Prostaglandins Leukot Essent Fat Acids 2017. 10.1016/j.plefa.2017.01.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [54].Jin Y, Arita M, Zhang Q, Saban DR, Chauhan SK, Chiang N, et al. Anti-angiogenesis effect of the novel anti-inflammatory and pro-resolving lipid mediators. Investig Ophthalmol Vis Sci 2009;50:4743–52. 10.1167/iovs.08-2462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [55].Flitter BA, Fang X, Matthay MA, Gronert K. The potential of lipid mediator networks as ocular surface therapeutics and biomarkers. Ocul Surf 2021. 10.1016/j.jtos.2020.04.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [56].Leedom AJ, Sullivan AB, Dong B, Lau D, Gronert K. Endogenous LXA4 circuits are determinants of pathological angiogenesis in response to chronic injury. Am J Pathol 2010. 10.2353/ajpath.2010.090678. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [57].Zhang F, Yang H, Pan Z, Wang Z, Wolosin JM, Gjorstrup P, et al. Dependence of resolvin-induced increases in corneal epithelial cell migration on EGF receptor transactivation. Investig Ophthalmol Vis Sci 2010. 10.1167/iovs.09-4468. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [58].Wang SB, Hu KM, Seamon KJ, Mani V, Chen Y, Gronert K. Estrogen negatively regulates epithelial wound healing and protective lipid mediator circuits in the cornea. FASEB J 2012. 10.1096/fi.11-198036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [59].Kenchegowda S, He J, Bazan HEP. Involvement of pigment epithelium-derived factor, docosahexaenoic acid and neuroprotectin D1 in corneal inflammation and nerve integrity after refractive surgery. Prostaglandins Leukot Essent Fat Acids 2013. 10.1016/j.plefa.2012.03.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [60].De Paiva CS, Schwartz CE, Gjörstrup P, Pflugfelder SC. Resolvin E1 (RX-10001) reduces corneal epithelial barrier disruption and protects against goblet cell loss in a murine model of dry eye. Cornea 2012. 10.1097/ICO.0b013e31823f789e. [DOI] [PubMed] [Google Scholar]
- [61].Li N, He J, Schwartz CE, Gjorstrup P, Bazan HEP. Resolvin E1 improves tear production and decreases inflammation in a dry eye mouse model. J Ocul Pharmacol Ther 2010. 10.1089/jop.2010.0019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [62].Rashid S, Jin Y, Ecoiffier T, Barabino S, Schaumberg DA, Dana MR. Topical omega-3 and omega-6 fatty acids for treatment of dry eye. Arch Ophthalmol 2008;126:219–25. 10.1001/archophthalmol.2007.61. [DOI] [PubMed] [Google Scholar]
- [63].He J, Kakazu AH, Bazan NG, Bazan HEP. Aspirin-triggered lipoxin A4 (15-epi-LXA4) increases the endothelial viability of human corneas storage in Optisol-GS. J Ocul Pharmacol Ther 2011. 10.1089/jop.2010.0187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [64].Hua J, Jin Y, Chen Y, Inomata T, Lee HS, Chauhan SK, et al. The resolvin D1 analogue controls maturation of dendritic cells and suppresses alloimmunity in corneal transplantation. Investig Ophthalmol Vis Sci 2014. 10.1167/iovs.14-14356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [65].Dana MR, Qian Y, Hamrah P. Twenty-five-year panorama of corneal immunology: Emerging concepts in the immunopathogenesis of microbial keratitis, peripheral ulcerative keratitis, and corneal transplant rejection. Cornea 2000;19:625–43. 10.1097/00003226-200009000-00008. [DOI] [PubMed] [Google Scholar]
- [66].Dietrich T, Bock F, Yuen D, Hos D, Bachmann BO, Zahn G, et al. Cutting Edge: Lymphatic Vessels, Not Blood Vessels, Primarily Mediate Immune Rejections After Transplantation. J Immunol 2010;184:535–9. 10.4049/jimmunol.0903180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [67].Chauhan SK, Jin Y, Goyal S, Lee HSK, Fuchsluger TA, Lee HSK, et al. A novel pro-lymphangiogenic function for Th17/IL-17. Blood 2011;118:4630–4. 10.1182/blood-2011-01-332049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [68].Goyal S, Chauhan SK, Dana R. Blockade of prolymphangiogenic vascular endothelial growth factor C in dry eye disease. Arch Ophthalmol 2012;130:84–9. 10.1001/archophthalmol.2011.266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [69].Ambati BK, Nozaki M, Singh N, Takeda A, Jani PD, Suthar T, et al. Corneal avascularity is due to soluble VEGF receptor-1. Nature 2006;443:993–7. 10.1038/nature05249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [70].Cursiefen C, Chen L, Saint-Geniez M, Hamrah P, Jin Y, Rashid S, et al. Nonvascular VEGF receptor 3 expression by corneal epithelium maintains avascularity and vision. Proc Natl Acad Sci U S A 2006;103:11405–10. 10.1073/pnas.0506112103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [71].Polisetti N, Gießl A, Zenkel M, Heger L, Dudziak D, Naschberger E, et al. Melanocytes as emerging key players in niche regulation of limbal epithelial stem cells. Ocul Surf 2021. 10.1016/j.jtos.2021.08.006. [DOI] [PubMed] [Google Scholar]
- [72].Luo L, Li DQ, Doshi A, Farley W, Corrales RM, Pflugfelder SC. Experimental dry eye stimulates production of inflammatory cytokines and MMP-9 and activates MAPK signaling pathways on the ocular surface. Investig Ophthalmol Vis Sci 2004;45:4293–301. 10.1167/iovs.03-1145. [DOI] [PubMed] [Google Scholar]
- [73].Morelli AE, Thomson AW. Tolerogenic dendritic cells and the quest for transplant tolerance. Nat Rev Immunol 2007;7:610–21. 10.1038/nri2132. [DOI] [PubMed] [Google Scholar]
- [74].Ezzelarab M, Thomson AW. Tolerogenic dendritic cells and their role in transplantation. Semin Immunol 2011;23:252–63. 10.1016/j.smim.2011.06.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [75].Saban DR, Bock F, Chauhan SK, Masli S, Dana R. Thrombospondin-1 Derived from APCs Regulates Their Capacity for Allosensitization. J Immunol 2010;185:4691–7. 10.4049/jimmunol.1001133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [76].Tahvildari M, Emami-Naeini P, Omoto M, Mashaghi A, Chauhan SK, Dana R. Treatment of donor corneal tissue with immunomodulatory cytokines: A novel strategy to promote graft survival in high-risk corneal transplantation. Sci Rep 2017;7. 10.1038/s41598-017-01065-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [77].Li M, Mittal SK, Foulsham W, Amouzegar A, Sahu SK, Chauhan SK. Mast cells contribute to the induction of ocular mucosal alloimmunity. Am J Transplant 2019. 10.1111/ajt.15084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [78].Schaumburg CS, Siemasko KF, De Paiva CS, Wheeler LA, Niederkom JY, Pflugfelder SC, et al. Ocular Surface APCs Are Necessary for Autoreactive T Cell-Mediated Experimental Autoimmune Lacrimal Keratoconjunctivitis. J Immunol 2011;187:3653–62. 10.4049/jimmunol.1101442. [DOI] [PubMed] [Google Scholar]
- [79].Huq S, Liu Y, Benichou G, Dana MR. Relevance of the Direct Pathway of Sensitization in Corneal Transplantation Is Dictated by the Graft Bed Microenvironment. J Immunol 2004;173:4464–9. 10.4049/jimmunol.173.7.4464. [DOI] [PubMed] [Google Scholar]
- [80].Jamali A, Hu K, Sendra VG, Blanco T, Lopez MJ, Ortiz G, et al. Characterization of Resident Corneal Plasmacytoid Dendritic Cells and Their Pivotal Role in Herpes Simplex Keratitis. Cell Rep 2020;32. 10.1016/j.celrep.2020.108099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [81].Jamali A, Kenyon B, Ortiz G, Abou-Slaybi A, Sendra VG, Harris DL, et al. Plasmacytoid dendritic cells in the eye. Prog Retin Eye Res 2021;80. 10.1016/j.preteyeres.2020.100877. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [82].Jamali A, Harris DL, Blanco T, Lopez MJ, Hamrah P. Resident plasmacytoid dendritic cells patrol vessels in the naïve limbus and conjunctiva. Ocul Surf 2020. 10.1016/j.jtos.2020.02.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [83].Müller LJ, Pels L, Vrensen GFJM. Ultrastructural organization of human corneal nerves. Investig Ophthalmol Vis Sci 1996;37:476–88. [PubMed] [Google Scholar]
- [84].Dastjerdi MH, Dana R. Corneal nerve alterations in dry eye-associated ocular surface disease. Int Ophthalmol Clin 2009;49:11–20. 10.1097/IIO.0b013e31819242c9. [DOI] [PubMed] [Google Scholar]
- [85].Ferrari G, Chauhan SK, Ueno H, Nallasamy N, Gandolfi S, Borges L, et al. A novel mouse model for neurotrophic keratopathy: Trigeminal nerve stereotactic electrolysis through the brain. Investig Ophthalmol Vis Sci 2011;52:2532–9. 10.1167/iovs.10-5688. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [86].Ueno H, Ferrari G, Hattori T, Saban DR, Katikireddy KR, Chauhan SK, et al. Dependence of corneal stem/progenitor cells on ocular surface innervation. Investig Ophthalmol Vis Sci 2012;53:867–72. 10.1167/iovs.11-8438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [87].Kheirkhah A, Dohlman TH, Amparo F, Arnoldner MA, Jamali A, Hamrah P, et al. Effects of corneal nerve density on the response to treatment in dry eye disease. Ophthalmology 2015;122:662–8. 10.1016/j.ophtha.2014.11.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [88].Benítez Del Castillo JM, Wasfy MAS, Fernandez C, Garcia-Sanchez J. An in vivo confocal masked study on corneal epithelium and subbasal nerves in patients with dry eye. Investig Ophthalmol Vis Sci 2004;45:3030–5. 10.1167/iovs.04-0251. [DOI] [PubMed] [Google Scholar]
- [89].Stepp MA, Pal-Ghosh S, Tadvalkar G, Williams A, Pflugfelder SC, de Paiva CS. Reduced intraepithelial corneal nerve density and sensitivity accompany desiccating stress and aging in C57BL/6 mice. Exp Eye Res 2018;169:91–8. 10.1016/j.exer.2018.01.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [90].Guzmán M, Miglio M, Keitelman I, Shiromizu CM, Sabbione F, Fuentes F, et al. Transient tear hyperosmolarity disrupts the neuroimmune homeostasis of the ocular surface and facilitates dry eye onset. Immunology 2020;161:148–61. 10.1111/imm.13243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [91].Ferrari G, Hajrasouliha AR, Sadrai Z, Ueno H, Chauhan SK, Dana R. Nerves and neovessels inhibit each other in the cornea. Investig Ophthalmol Vis Sci 2013;54:813–20. 10.1167/iovs.11-8379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [92].Gaddipati S, Rao P, Jerome AD, Burugula BB, Gerard NP, Suvas S. Loss of Neurokinin-1 Receptor Alters Ocular Surface Homeostasis and Promotes an Early Development of Herpes Stromal Keratitis. J Immunol 2016;197:4021–33. 10.4049/jimmunol.1600836. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [93].Chikama TI, Fukuda K, Morishige N, Nishida T. Treatment of neurotrophic keratopathy with substance-P-derived peptide (FGLM) and insulin-like growth factor I. Lancet 1998;351:1783–4. 10.1016/S0140-6736(98)24024-4. [DOI] [PubMed] [Google Scholar]
- [94].Yu M, Lee SM, Lee H, Amouzegar A, Nakao T, Chen Y, et al. Neurokinin-1 Receptor Antagonism Ameliorates Dry Eye Disease by Inhibiting Antigen-Presenting Cell Maturation and T Helper 17 Cell Activation. Am J Pathol 2020;190:125–33. 10.1016/j.ajpath.2019.09.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [95].Chao C, Stapleton F, Zhou X, Chen S, Zhou S, Golebiowski B. Structural and functional changes in corneal innervation after laser in situ keratomileusis and their relationship with dry eye. Graefe’s Arch Clin Exp Ophthalmol 2015;253:2029–39. 10.1007/s00417-015-3120-1. [DOI] [PubMed] [Google Scholar]
- [96].Ziche M, Morbidelli L, Pacini M, Geppetti P, Alessandri G, Maggi CA. Substance P stimulates neovascularization in vivo and proliferation of cultured endothelial cells. Microvasc Res 1990;40:264–78. 10.1016/0026-2862(90)90024-L. [DOI] [PubMed] [Google Scholar]
- [97].Bignami F, Giacomini C, Lorusso A, Aramini A, Rama P, Ferrari G. NK1 receptor antagonists as a new treatment for corneal neovascularization. Invest Ophthalmol Vis Sci 2014;55:6783–94. 10.1167/iovs.14-14553. [DOI] [PubMed] [Google Scholar]
- [98].Green DP, Limjunyawong N, Gour N, Pundir P, Dong X. A Mast-Cell-Specific Receptor Mediates Neurogenic Inflammation and Pain. Neuron 2019. 10.1016/j.neuron.2019.01.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [99].Liu L, Dana R, Yin J. Sensory neurons directly promote angiogenesis in response to inflammation via substance P signaling. FASEB J 2020;34:6229–43. 10.1096/fj.201903236R. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [100].Barbariga M, Fonteyne P, Ostadreza M, Bignami F, Rama P, Ferrari G. Substance p modulation of human and murine corneal neovascularization. Investig Ophthalmol Vis Sci 2018. 10.1167/iovs.17-23198. [DOI] [PubMed] [Google Scholar]
- [101].Cho WK, Mittal SK, Elbasiony E, Chauhan SK. Activation of ocular surface mast cells promotes corneal neovascularization. Ocul Surf 2020. 10.1016/j.jtos.2020.09.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [102].Sahu SK, Mittal SK, Foulsham W, Li M, Sangwan VS, Chauhan SK. Mast cells initiate the recruitment of neutrophils following ocular surface injury. Investig Ophthalmol Vis Sci 2018. 10.1167/iovs.17-23398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [103].Paunicka KJ, Mellon J, Robertson D, Petroll M, Brown JR, Niederkorn JY. Severing corneal nerves in one eye induces sympathetic loss of immune privilege and promotes rejection of future corneal allografts placed in either eye. Am J Transplant 2015;15:1490–501. 10.Ill1/ajt.13240. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [104].Szliter EA, Lighvani S, Barrett RP, Hazlett LD. Vasoactive Intestinal Peptide Balances Pro- and Anti-Inflammatory Cytokines in the Pseudomonas aeruginosa -Infected Cornea and Protects against Corneal Perforation. J Immunol 2007;178:1105–14. 10.4049/jimmunol.178.2.1105. [DOI] [PubMed] [Google Scholar]
- [105].Satitpitakul V, Sun Z, Suri K, Amouzegar A, Katikireddy KR, Jurkunas UV., et al. Vasoactive Intestinal Peptide Promotes Corneal Allograft Survival. Am J Pathol 2018;188:2016–24. 10.1016/j.ajpath.2018.05.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [106].Hegarty DM, Tonsfeldt K, Hermes SM, Helfand H, Aicher SA. Differential localization of vesicular glutamate transporters and peptides in corneal afferents to trigeminal nucleus caudalis. J Comp Neurol 2010;518:3557–69. 10.1002/cne.22414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [107].Jones MA, Marfurt CF. Peptidergic innervation of the rat cornea. Exp Eye Res 1998;66:421–35. 10.1006/exer.1997.0446. [DOI] [PubMed] [Google Scholar]
- [108].Tatematsu Y, Khan Q, Blanco T, Bair JA, Hodges RR, Masli S, et al. Thrombospondin-1 is necessary for the development and repair of corneal nerves. Int J Mol Sci 2018;19. 10.3390/ijms19103191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [109].Nishida T, Taylor AW. Specific aqueous humor factors induce activation of regulatory T cells. Investig Ophthalmol Vis Sci 1999. [PubMed] [Google Scholar]
- [110].Taylor AW, Namba K. In vitro induction of CD25+ CD4+ regulatory T cells by the neuropeptide alpha-melanocyte stimulating hormone (α-MSH). Immunol Cell Biol 2001. 10.1046/j.1440-1711.2001.01022.x. [DOI] [PubMed] [Google Scholar]
- [111].Nieto JE, Casanova I, Sema-Ojeda JC, Graue-Hernández EO, Quintana G, Salazar A, et al. Increased expression of tlr4 in circulating cd4+t cells in patients with allergic conjunctivitis and in vitro attenuation of th2 inflammatory response by alpha-msh. Int J Mol Sci 2020. 10.3390/ijms21217861. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [112].Taylor AW, Streilein JW, Cousins SW. Identification of alpha-melanocyte stimulating hormone as a potential immunosuppressive factor in aqueous humor. Curr Eye Res 1992. 10.3109/02713689208999545. [DOI] [PubMed] [Google Scholar]
- [113].Ru Y, Huang Y, Liu H, Du J, Meng Z, Dou Z, et al. α-Melanocyte-stimulating hormone ameliorates ocular surface dysfunctions and lesions in a scopolamine-induced dry eye model via PKA-CREB and MEK-Erk pathways. Sci Rep 2015. 10.1038/srep18619. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [114].Lužnik Z, Sun Z, Nakagawa H, Taylor AW, Jurkunas UV., Yin J, et al. Association of α-melanocyte-stimulating hormone with corneal endothelial cell survival during oxidative stress and inflammation-induced cell loss in donor tissue. JAMA Ophthalmol 2020. 10.1001/jamaophthalmol.2020.3413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [115].Hamrah P, Haskova Z, Taylor AW, Zhang Q, Ksander BR, Dana MR. Local treatment with alpha-melanocyte stimulating hormone reduces corneal allorejection. Transplantation 2009. 10.1097/TP.0b013e3181ac11ea. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [116].Floyd AM, Zhou X, Evans C, Rompala OJ, Zhu L, Wang M, et al. Mucin Deficiency Causes Functional and Structural Changes of the Ocular Surface. PLoS One 2012;7. 10.1371/journal.pone.0050704. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [117].Chen Y, Chauhan SK, Shao C, Omoto M, Inomata T, Dana R. IFN-γ–Expressing Th17 Cells Are Required for Development of Severe Ocular Surface Autoimmunity. J Immunol 2017;199:1163–9. 10.4049/jimmunol.1602144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [118].Fabiani C, Barabino S, Rashid S, Dana MR. Corneal epithelial proliferation and thickness in a mouse model of dry eye. Exp Eye Res 2009;89:166–71. 10.1016/j.exer.2009.03.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [119].Ouyang W, Wu Y, Lin X, Wang S, Yang Y, Tang L, et al. Role of CD4+ T helper cells in the development of BAC-induced dry eye syndrome in mice. Investig Ophthalmol Vis Sci 2021;62:25. 10.1167/IOVS.62.1.25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [120].Knoop KA, McDonald KG, McCrate S, McDole JR, Newberry RD. Microbial sensing by goblet cells controls immune surveillance of luminal antigens in the colon. Mucosal Immunol 2015;8:198–210. 10.1038/mi.2014.58. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [121].McDole JR, Wheeler LW, McDonald KG, Wang B, Konjufca V, Knoop KA, et al. Goblet cells deliver luminal antigen to CD103 + dendritic cells in the small intestine. Nature 2012;483:345–9. 10.1038/nature10863. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [122].Barbosa FL, Xiao Y, Bian F, Coursey TG, Ko BY, Clevers H, et al. Goblet cells contribute to ocular surface immune tolerance—implications for dry eye disease. Int J Mol Sci 2017; 18. 10.3390/ijms18050978. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [123].Shan M, Gentile M, Yeiser JR, Walland AC, Bornstein VU, Chen K, et al. Mucus enhances gut homeostasis and oral tolerance by delivering immunoregulatory signals. Science (80-) 2013. 10.1126/science.1237910. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [124].McKenzie RW, Jumblatt JE, Jumblatt MM. Quantification of MUC2 and MUC5AC transcripts in human conjunctiva. Investig Ophthalmol Vis Sci 2000. [PubMed] [Google Scholar]
- [125].Contreras-Ruiz L, Masli S. Immunomodulatory cross-talk between conjunctival goblet cells and dendritic cells. PLoS One 2015;10:e0120284. 10.1371/journal.pone.0120284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [126].Xiao Y, De Paiva CS, Yu Z, De Souza RG, Li DQ, Pflugfelder SC. Goblet cell-produced retinoic acid suppresses CD86 expression and IL-12 production in bone marrow-derived cells. Int Immunol 2018. 10.1093/intimm/dxy045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [127].Alam J, de Paiva CS, Pflugfelder SC. Immune - Goblet cell interaction in the conjunctiva. Ocul Surf 2020. 10.1016/j.jtos.2019.12.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [128].Swamynathan SK, Wells A. Conjunctival goblet cells: Ocular surface functions, disorders that affect them, and the potential for their regeneration. Ocul Surf 2020; 18:19–26. 10.1016/j.jtos.2019.11.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [129].Knop E, Knop N. The role of eye-associated lymphoid tissue in corneal immune protection. J Anat 2005;206:271–85. 10.1111/j.1469-7580.2005.00394.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [130].Knop N, Knop E. Regulation of the inflammatory component in chronic dry eye disease by the eye-associated lymphoid tissue (EALT). Dev Ophthalmol 2010;45:23–39. 10.1159/000315017. [DOI] [PubMed] [Google Scholar]
- [131].Steven P, Schwab S, Kiesewetter A, Saban DR, Stern ME, Gehlsen U. Disease- specific expression of conjunctiva associated lymphoid tissue (CALT) in mouse models of dry eye disease and ocular allergy. Int J Mol Sci 2020;21:1–14. 10.3390/ijms21207514. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [132].Mastropasqua R, Agnifili L, Fasanella V, Nubile M, Gnama AA, Falconio G, et al. The Conjunctiva-Associated Lymphoid Tissue in Chronic Ocular Surface Diseases. Microsc Microanal 2017;23:697–707. 10.1017/S1431927617000538. [DOI] [PubMed] [Google Scholar]
- [133].Kodati S, Chauhan SK, Chen Y, Dohlman TH, Karimian P, Saban D, et al. CCR7 is critical for the induction and maintenance of Th17 immunity in dry eye disease. Investig Ophthalmol Vis Sci 2014;55:5871–7. 10.1167/iovs.14-14481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [134].Chen Y, Chauhan SK, Saban DR, Sadrai Z, Okanobo A, Dana R. Interferon-γ-secreting NK cells promote induction of dry eye disease. J Leukoc Biol 2011;89:965–72. 10.1189/jlb.1110611. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [135].Ecoiffier T, El Annan J, Rashid S, Schaumberg D, Dana R. Modulation of integrin α4 β1 (VLA-4) in dry eye disease. Arch Ophthalmol 2008;126:1695–9. 10.1001/archopht.126.12.1695. [DOI] [PubMed] [Google Scholar]
- [136].Grewal IS, Flavell RA. CD40 and CD154 in cell-mediated immunity. Annu Rev Immunol 1998; 16:111–35. 10.1146/annurev.immunol.16.1.111. [DOI] [PubMed] [Google Scholar]
- [137].Stockinger B, Veldhoen M. Differentiation and function of Th17 T cells. Curr Opin Immunol 2007;19:281–6. 10.1016/j.coi.2007.04.005. [DOI] [PubMed] [Google Scholar]
- [138].Bettelli E, Carrier Y, Gao W, Korn T, Strom TB, Oukka M, et al. Reciprocal developmental pathways for the generation of pathogenic effector TH17 and regulatory T cells. Nature 2006;441:235–8. 10.1038/nature04753. [DOI] [PubMed] [Google Scholar]
- [139].Hegde S, Beauregard C, Mayhew E, Niederkorn JY. CD4+ T-cell-mediated mechanisms of corneal allograft rejection: Role of Fas-induced apoptosis. Transplantation 2005;79:23–31. 10.1097/01.TP.0000147196.79546.69. [DOI] [PubMed] [Google Scholar]
- [140].Yamada J, Kurimoto I, Streilein JW. Role of CD4+ T cells in immunobiology of orthotopic corneal transplants in mice. Investig Ophthalmol Vis Sci 1999;40:2614–21. [PubMed] [Google Scholar]
- [141].Reyes NJ, Saban DR. T helper subsets in allergic eye disease. Curr Opin Allergy Clin Immunol 2014;14:477–84. 10.1097/ACI.0000000000000088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [142].De Paiva CS, Chotikavanich S, Pangelinan SB, Pitcher JD, Fang B, Zheng X, et al. IL-17 disrupts corneal barrier following desiccating stress. Mucosal Immunol 2009;2:243–53. 10.1038/mi.2009.5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [143].Chauhan SK, El Annan J, Ecoiffier T, Goyal S, Zhang Q, Saban DR, et al. Autoimmunity in Dry Eye Is Due to Resistance of Th17 to Treg Suppression. J Immunol 2009;182:1247–52. 10.4049/jimmunol.182.3.1247. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [144].Lin X, Rui K, Deng J, Tian J, Wang X, Wang S, et al. Th17 cells play a critical role in the development of experimental Sjogren’s syndrome. Ann Rheum Dis 2015;74:1302–10. 10.1136/annrheumdis-2013-204584. [DOI] [PubMed] [Google Scholar]
- [145].Oh JY, Kim MK, Choi HJ, Ko JH, Kang EJ, Lee HJ, et al. Investigating the relationship between serum interleukin-17 levels and systemic immune-mediated disease in patients with dry eye syndrome. Korean J Ophthalmol 2011;25:73–6. 10.3341/kjo.2011.25.2.73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [146].Garbutcheon-Singh KB, Carnt N, Pattamatta U, Samarawickrama C, White A, Calder V. A Review of the Cytokine IL-17 in Ocular Surface and Corneal Disease. Curr Eye Res 2019;44:1–10. 10.1080/02713683.2018.1519834. [DOI] [PubMed] [Google Scholar]
- [147].Sakaguchi S, Yamaguchi T, Nomura T, Ono M. Regulatory T Cells and Immune Tolerance. Cell 2008;133:775–87. 10.1016/j.cell.2008.05.009. [DOI] [PubMed] [Google Scholar]
- [148].Foulsham W, Marmalidou A, Amouzegar A, Coco G, Chen Y, Dana R. Review: The function of regulatory T cells at the ocular surface. Ocul Surf 2017;15:652–9. 10.1016/j.jtos.2017.05.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [149].Piccirillo CA, Shevach EM. Naturally-occurring CD4+CD25+ immunoregulatory T cells: Central players in the arena of peripheral tolerance. Semin Immunol 2004;16:81–8. 10.1016/j.smim.2003.12.003. [DOI] [PubMed] [Google Scholar]
- [150].Kretschmer K, Apostolou I, Hawiger D, Khazaie K, Nussenzweig MC, von Boehmer H. Inducing and expanding regulatory T cell populations by foreign antigen. Nat Immunol 2005;6:1219–27. 10.1038/ni1265. [DOI] [PubMed] [Google Scholar]
- [151].Shevach EM. Foxp3+ T regulatory cells: Still many unanswered Questions-A perspective after 20 years of study. Front Immunol 2018;9. 10.3389/fimmu.2018.01048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [152].Inomata T, Hua J, Di Zazzo A, Dana R. Impaired Function of Peripherally Induced Regulatory T Cells in Hosts at High Risk of Graft Rejection. Sci Rep 2016;6. 10.1038/srep39924. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [153].Chauhan SK, Saban DR, Lee HK, Dana R. Levels of Foxp3 in Regulatory T Cells Reflect Their Functional Status in Transplantation. J Immunol 2009;182:148–53. 10.4049/jimmunol.182.1.148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [154].Shao C, Chen Y, Nakao T, Amouzegar A, Yin J, Tahvildari M, et al. Local Delivery of Regulatory T Cells Promotes Corneal Allograft Survival. Transplantation 2019;103:182–90. 10.1097/TP.0000000000002442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [155].Tahvildari M, Omoto M, Chen Y, Emami-Naeini P, Inomata T, Dohlman TH, et al. In vivo expansion of regulatory T cells by low-dose interleukin-2 treatment increases allograft survival in corneal transplantation. Transplantation 2016;100:525–32. 10.1097/TP.0000000000001044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [156].Sumi T, Fukushima A, Fukuda K, Kumagai N, Nishida T, Takeuchi M, et al. Thymus-derived CD4+ CD25+ T cells suppress the development of murine allergic conjunctivitis. Int Arch Allergy Immunol 2007;143:276–81. 10.1159/000100573. [DOI] [PubMed] [Google Scholar]
- [157].Lee HS, Schlereth S, Khandelwal P, Saban DR. Ocular Allergy Modulation to Hi-Dose Antigen Sensitization Is a Treg-Dependent Process. PLoS One 2013;8. 10.1371/journal.pone.0075769. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [158].Galicia-Carreón J, Santacruz C, Ayala-Balboa J, Robles-Contreras A, Perez-Tapia SM, Garfias Y, et al. An imbalance between frequency of CD4+CD25+FOXP3+ regulatory t cells and CCR4+ and CCR9+ circulating helper T cells is associated with active perennial allergic conjunctivitis. Clin Dev Immunol 2013;2013. 10.1155/2013/919742. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [159].Grindebacke H, Wing K, Andersson AC, Suri-Payer E, Rak S, Rudin A. Defective suppression of Th2 cytokines by CD4+CD25+ regulatory T cells in birch allergies during birch pollen season. Clin Exp Allergy 2004;34:1364–72. 10.1111/j.1365-2222.2004.02067.x. [DOI] [PubMed] [Google Scholar]
- [160].Chen Y, Chauhan SK, Lee HS, Stevenson W, Schaumburg CS, Sadrai Z, et al. Effect of desiccating environmental stress versus systemic muscarinic AChR blockade on dry eye immunopathogenesis. Investig Ophthalmol Vis Sci 2013;54:2457–64. 10.1167/iovs.12-11121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [161].Taketani Y, Marmalidou A, Dohlman TH, Singh RB, Amouzegar A, Chauhan SK, et al. Restoration of Regulatory T-Cell Function in Dry Eye Disease by Antagonizing Substance P/Neurokinin-1 Receptor. Am J Pathol 2020;190:1859–66. 10.1016/j.ajpath.2020.05.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [162].Niederkorn JY, Stern ME, Pflugfelder SC, De Paiva CS, Corrales RM, Gao J, et al. Desiccating Stress Induces T Cell-Mediated Sjögren’s Syndrome-Like Lacrimal Keratoconjunctivitis. J Immunol 2006;176:3950–7. 10.4049/jimmunol.176.7.3950. [DOI] [PubMed] [Google Scholar]
- [163].Rahimy E, Pitcher JD, Pangelinan SB, Chen W, Farley WJ, Niederkorn JY, et al. Spontaneous autoimmune dacryoadenitis in aged CD25KO mice. Am J Pathol 2010;177:744–53. 10.2353/ajpath.2010.091116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [164].de Paiva CS, Hwang CS, Pitcher JD, Pangelinan SB, Rahimy E, Chen W, et al. Age-related T-cell cytokine profile parallels corneal disease severity in Sjögren’s syndrome-like keratoconjunctivitis sicca in CD25KO mice. Rheumatology 2010;49:246–58. 10.1093/rheumatology/kep357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [165].Trujillo-Vargas CM, Schaefer L, Alam J, Pflugfelder SC, Britton RA, de Paiva CS. The gut-eye-lacrimal gland-microbiome axis in Sjögren Syndrome. Ocul Surf 2020;18:335–44. 10.1016/j.jtos.2019.10.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [166].Chen Y, Chauhan SK, Soo Lee H, Saban DR, Dana R. Chronic dry eye disease is principally mediated by effector memory Th17 cells. Mucosal Immunol 2014;7:38–45. 10.1038/mi.2013.20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [167].Hao LR, Li XF, Gao C, Cao L, Han ZY, Gao H, et al. Th17/Treg cell level and clinical characteristics of peripheral blood of patients with Sjogren’s syndrome complicated with primary biliary cirrhosis. Med (United States) 2019;98. 10.1097/MD.0000000000015952. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [168].Miao M, Hao Z, Guo Y, Zhang X, Zhang S, Luo J, et al. Short-term and low-dose IL-2 therapy restores the Th17/Treg balance in the peripheral blood of patients with primary Sjögren’s syndrome. Ann Rheum Dis 2018;77:1838–40. 10.1136/annrheumdis-2018-213036. [DOI] [PubMed] [Google Scholar]
- [169].Farrand KF, Fridman M, Stillman IÖ, Schaumberg DA. Prevalence of Diagnosed Dry Eye Disease in the United States Among Adults Aged 18 Years and Older. Am J Ophthalmol 2017;182:90–8. 10.1016/j.ajo.2017.06.033. [DOI] [PubMed] [Google Scholar]
- [170].Wang C, Zaheer M, Bian F, Quach D, Swennes AG, Britton RA, et al. Sjögren-like lacrimal Keratoconjunctivitis in germ-free mice. Int J Mol Sci 2018; 19. 10.3390/ijms19020565. [DOI] [PMC free article] [PubMed] [Google Scholar]