

Abstract
Most genes in photosynthetic organisms remain functionally uncharacterized. Here, using a barcoded mutant library of the model eukaryotic alga Chlamydomonas reinhardtii, we determined the phenotypes of more than 58,000 mutants under more than 121 different environmental growth conditions and chemical treatments. A total of 59% of genes are represented by at least one mutant that showed a phenotype, providing clues to the functions of thousands of genes. Mutant phenotypic profiles place uncharacterized genes into functional pathways such as DNA repair, photosynthesis, the CO2-concentrating mechanism and ciliogenesis. We illustrate the value of this resource by validating phenotypes and gene functions, including three new components of an actin cytoskeleton defense pathway. The data also inform phenotype discovery in land plants; mutants in Arabidopsis thaliana genes exhibit phenotypes similar to those we observed in their Chlamydomonas homologs. We anticipate that this resource will guide the functional characterization of genes across the tree of life.
Subject terms: Plant genetics, High-throughput screening
Systematic phenotyping of 58,101 mutants of the model eukaryotic alga Chlamydomonas reinhardtii under 121 environmental and chemical stress conditions provides a large resource for characterizing gene function.
Main
Major contributions to our understanding of gene functions in photosynthetic organisms have been made by studying microbial models, including the discovery and characterization of the Calvin–Benson–Bassham CO2 fixation cycle1 as well as the structures2, order3 and cloning4 of complexes in the photosynthetic electron transport chain. Advances in technology now provide opportunities for microbes to serve as powerful complements to land plants in the characterization of gene functions by enabling substantially higher experimental throughput5.
The single-celled green alga Chlamydomonas (Chlamydomonas reinhardtii) is a well-established model system for studies of key pathways, including photosynthesis, primary metabolism, interorganelle communication and stress response6. Furthermore, amenability to microscopy and biochemical purifications have made Chlamydomonas a leading model system for studies of cilia7–9. Despite promising progress with the development of clustered regularly interspaced short palindromic repeats (CRISPR)-based reagents to generate targeted mutants10,11, low editing efficiencies currently prevent large-scale CRISPR single guide RNA library screens in Chlamydomonas. The recent generation of a barcoded Chlamydomonas mutant collection facilitates the study of individual genes and enables forward genetic screens12. In the present work, we leverage the amenability of Chlamydomonas to high-throughput methods to connect genotypes to phenotypes on a massive scale, allowing placement of genes into pathways and discovery of conserved gene functions in land plants.
Results
Systematic genome-scale phenotyping
To connect genotypes to phenotypes, we measured the growth of 58,101 Chlamydomonas mutants representing 14,695 genes (83% of all genes encoded in the Chlamydomonas genome, based on the current genome annotation, v5.6) under 121 environmental and chemical stress conditions (both control and experimental conditions are given in Supplementary Tables 1 and 2). We pooled the entire Chlamydomonas mutant collection from plates into a liquid culture and used molecular barcodes to quantify the relative abundance of each mutant after competitive growth (Fig. 1a–f). Growth conditions included heterotrophic, mixotrophic and photoautotrophic growth under different photon flux densities and CO2 concentrations, as well as abiotic stress conditions such as various pH and temperatures. We also subjected the library to known chemical stressors, including DNA-damaging agents, reactive oxygen species, antimicrobial drugs such as paromomycin and spectinomycin and the actin-depolymerizing drug latrunculin B (LatB). To further expand the range of stressors in the dataset, we identified 1,222 small molecules from the Library of AcTive Compounds on Arabidopsis (LATCA)13 that negatively influence Chlamydomonas growth (Extended Data Fig. 1, Supplementary Table 3 and Supplementary Data 1) and performed competitive growth experiments in the presence of 52 of the most potent compounds. We chose to screen the LATCA library for active compounds in Chlamydomonas because we believed that these compounds would be more likely to impact pathways both in Chlamydomonas and in plants, thus providing more general insights into gene functions in the green lineage. Taken together, this effort represents, to the best of our knowledge, the largest mutant-by-phenotype dataset to date for any photosynthetic organism, with 16.8 million data points (Supplementary Table 4).
Fig. 1. Pooled phenotyping enables the large-scale discovery of genotype-phenotype relationships in a unicellular photosynthetic eukaryote.

a, The Chlamydomonas mutant library was pooled and used to prepare a liquid starting culture of 58,101 mutants. b, Aliquots of the starting culture were used to inoculate pooled growth experiments to assess the fitness of each mutant under a variety of environmental and chemical stress treatments. c, The relative abundance of each mutant was quantified via polymerase chain reaction (PCR)-based amplification of individual mutant barcodes and subsequent Illumina sequencing. d, Mutants negatively affected by the treatment have a lower barcode read count compared to the control. e, Many genes were represented by multiple mutants, which allowed the identification of high-confidence gene phenotypes. We then clustered genes based on their phenotypic profile to place genes into pathways and predict the functions of previously uncharacterized genes. f, The data predict gene function in Chlamydomonas and land plants. WT, wild type.
Extended Data Fig. 1. A screen of the chemical library ‘LATCA’ identified 1,222 inhibitors of Chlamydomonas growth, 136 of which are active at 2 μM or less.
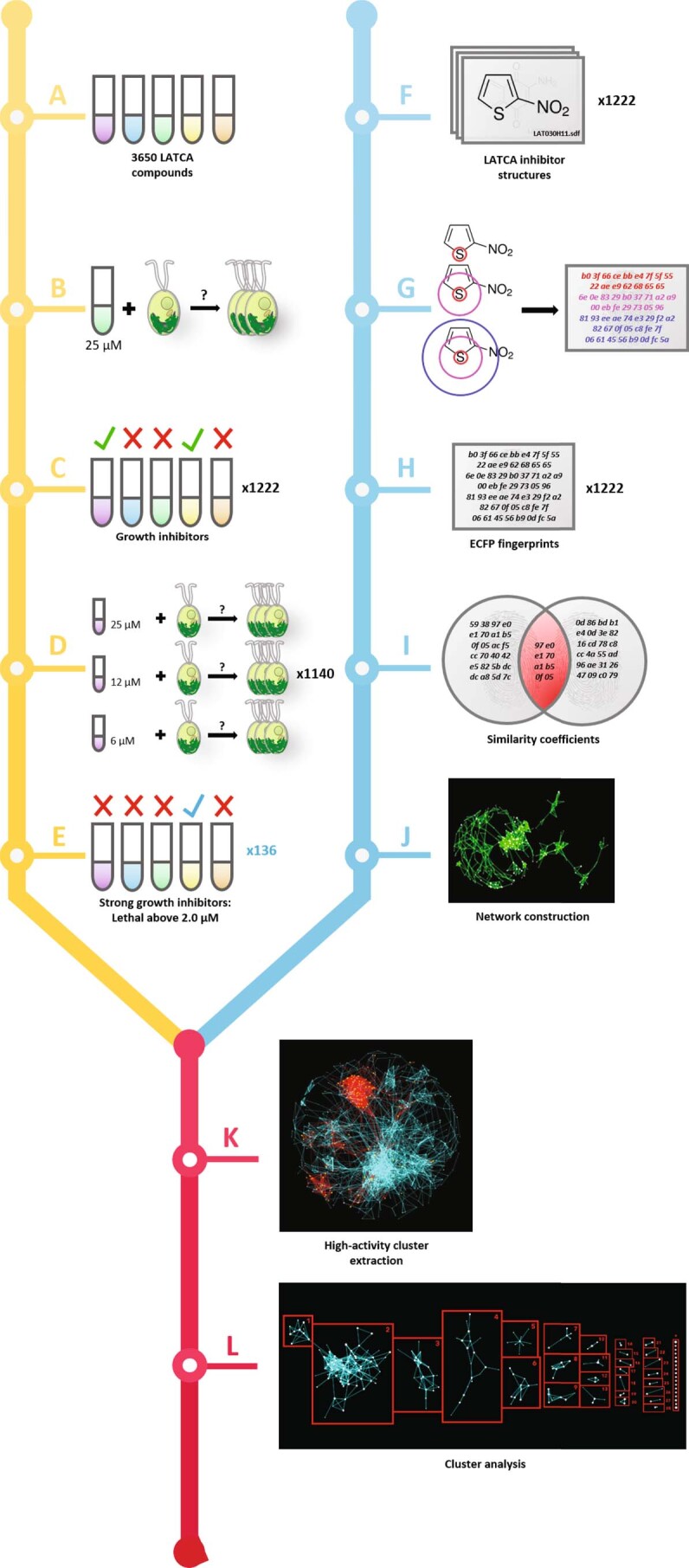
Phase 1 of the LATCA screen is depicted in A–E: The growth rate of wild-type Chlamydomonas (cMJ030) was evaluated in TAP and TP in the presence of 3,650 LATCA compounds. 1,222 out of the 3,650 LATCA compounds reduced growth by 90% or more at 25 μM (a–c). Dose-response experiments were performed in TAP medium with 1,140 out of the 1,222 highly active compounds that reduced growth at 2 μM or less (d,e). Phase 2 of the LATCA screen is depicted in F–J: Structural data files (SDFs) were acquired for all LATCA inhibitors (f) and converted into numerical fingerprints (extended-connectivity fingerprints; ECFPs) (g, h). ECFPs were then used to compute the structural similarity of pairs of compounds using Tanimoto coefficients (i). The set of Tanimoto coefficients between all pairs of inhibitors was condensed into a usable network (j). Phase 3 of the LATCA screen is depicted in K and L: Data from A–E was used to further reduce the similarity network from J to 28 clusters of structures exhibiting high levels of growth inhibition along with a group of singleton structures (*) that did not cluster. Supplementary Table 3 summarizes data A–E and shows cluster annotations from L; see also Extended Data File 1 for all chemical structures from L.
Mutants show genotype–phenotype specificity
To identify mutants with growth defects or enhancements due to a specific treatment, we compared the abundance of each mutant after growth under the treatment condition to its abundance after growth under a control condition (Fig. 2a). We called this comparison a screen and the ratio of these abundances the mutant phenotype (Fig. 2b,c). Mutant phenotypes were reproducible between independent replicates of a screen (Fig. 2c,d).
Fig. 2. We determined the fitness of 58,101 Chlamydomonas mutants under 121 growth conditions.

a, The phenotype of each mutant was determined by comparing its molecular barcode read count under a treatment and control condition. As an example, results from a screen using the drug cisplatin are shown. b, A hierarchically clustered heatmap shows the phenotype [log2(treatment reads/control reads)] of mutants across 212 screens representing 121 growth conditions. c, The typical reproducibility is illustrated with two replicate cisplatin screens. d, Mutants show screen-specific phenotypes. e, GO term analysis reveals enrichment of biological functions associated with specific screens. f, Most genes are represented by at least one mutant that shows a phenotype in at least one treatment condition. DSB, double-strand break; FDR, false discovery rate.
Individual mutants exhibited genotype–phenotype specificity. For example, mutants disrupted in the DNA repair gene POLYMERASE ZETA (POLZ, encoded by Cre09.g387400) exhibited growth defects in the presence of the DNA crosslinker cisplatin, and these mutants did not show growth defects in unrelated screens (Fig. 2d). We observed similar genotype–phenotype specificity for other genes and phenotypes, including sensitivity to low CO2, ciliogenesis and LatB sensitivity (Fig. 2d).
In many screens, mutants that exhibited phenotypes were enriched for disruptions in genes with expected function. In 46 out of 223 screens, at least one Gene Ontology (GO)14 term was enriched (FDR < 0.05) in the genes disrupted in mutants whose growth was perturbed in the screen (Fig. 2e, Extended Data Fig. 2 and Supplementary Table 5). These enriched GO terms corresponded to functions known to be required for survival under the respective treatments. For example, screens with DNA-damaging agents resulted in GO term enrichments such as ‘DNA replication’, ‘Nucleotide binding’ or ‘Damaged DNA binding’. These GO term enrichments suggest that the phenotypic screens are correctly identifying genes required for growth under the corresponding treatments.
Extended Data Fig. 2. Full GO term enrichments.

Gene Ontology term analysis reveals enrichment of biological functions observed for specific screens. GO; FDR < 0.05.
In total, 10,380 genes (59% of all Chlamydomonas genes) are represented by one or more 5′ untranslated region (UTR), coding DNA sequence (CDS) or intron insertion mutant that showed a phenotype (decreased abundance below our detection limit) in at least one screen (Fig. 2f). Although a lone mutant showing a phenotype is not sufficient evidence to conclusively establish a gene–phenotype relationship, we anticipate that these data will be useful to the research community in at least three ways. First, they can help prioritize the characterization of candidate genes identified by other means, such as transcriptomics or protein–protein interactions. Second, they facilitate the generation of hypotheses about the functions of poorly characterized genes. Third, they enable prioritization of available mutant alleles for further studies, including to establish a gene–phenotype relationship by complementation and/or backcrossing. The genotype–phenotype specificity of individual mutants and the enrichment of expected functions suggest that our data can serve as a guide for understanding the functions of thousands of poorly characterized genes.
High-confidence gene–phenotype relationships
The availability of multiple independent mutant alleles for individual genes allowed us to identify high-confidence gene–phenotype relationships. When multiple independent mutant alleles for the same gene show the same phenotype, the confidence in a gene lesion–phenotype relationship increases, because it is less likely that the phenotype is due to a mutation elsewhere in the genome (on average, there are 1.2 integration events per mutant, and the mutants can also carry other mutations such as point mutations) or that there was an error in mapping of the mutation12. Using a statistical framework that leverages multiple independent mutations in the same gene (Methods), we identified 1,218 high-confidence (FDR < 0.3) gene–phenotype relationships involving 684 genes (Fig. 3a and Supplementary Tables 6 and 7), including hundreds of genes with no functional annotation in the green lineage (Supplementary Table 8). Our gene–phenotype relationships include 302 high-confidence (FDR < 0.3) interactions involving 195 genes and 39 LATCA drugs, providing clues to the drugs’ targets and improving the value of these drugs as tools for perturbing specific pathways (Supplementary Table 7). Based on the highest-confidence (FDR < 0.05) phenotypes, we suggest names for 89 previously unnamed genes (Supplementary Table 9).
Fig. 3. Multiple alleles provide high confidence and reveal new phenotypes.

a, The number of genes with significant phenotypes in each class of screen is shown for two FDR thresholds. b, FDR is plotted against log2 median phenotype for all genes in the MMS screen. c, Growth assay of WT, mars1, mars1:MARS1 and mars1:MARS1-KD cells after 48 h in the presence or absence of MMS. Three biological replicates were used for each strain. For more details, Methods. d, Average chlorophyll concentrations of the liquid cultures shown in Fig. 3c. Data are presented as mean values ± s.d. n = 3 independent experiments. e, Immunoblot analysis of VESICLE-INDUCING PROTEIN IN PLASTIDS (VIPP2), a downstream target of MARS1, in WT and mars1 cells in the presence or absence of MMS. Immunoblot representative of n = 2 independent experiments.
As an example of how individual gene–phenotype relationships advance our understanding, we made the unexpected observation that mutants in the gene encoding the chloroplast unfolded protein response (cpUPR) kinase, MUTANT AFFECTED IN CHLOROPLAST-TO-NUCLEUS RETROGRADE SIGNALING (MARS1)15, were sensitive (FDR < 10−9) to the DNA-damaging agent methyl methanesulfonate (MMS) (Fig. 3b). We validated this phenotype in a separate growth assay and showed that the MMS sensitivity of these mutants is rescued by complementation with a wild-type copy of MARS1, but not by a kinase-dead version (Fig. 3c,d). We also determined that treatment with MMS led to induction of VESICLE-INDUCING PROTEIN IN PLASTIDS 2 (VIPP2), a highly selective cpUPR marker, in wild-type cells but not in mutants lacking MARS1 (Fig. 3e). These results illustrate the value of our high-throughput data and suggest the intriguing possibility that the cpUPR is activated via MARS1 upon DNA damage or protein alkylation and has a protective role against these stressors.
From phenotypes to pathways
To facilitate data visualization and predict the functions of poorly characterized genes in our dataset, we used the principle that mutant alleles with similar phenotypes tend to occur in genes that function in the same pathway5. We clustered the 684 genes with high-confidence phenotypes based on the similarity of their phenotypes across different treatments (Fig. 4a and Supplementary Data 2). The correlation of phenotypes was largely unrelated to transcriptional expression correlation16, suggesting that the two approaches provide complementary information (Extended Data Fig. 3 and Supplementary Table 10). We named some of our gene clusters based on the presence of previously characterized genes or based on the conditions that produced the most dramatic phenotypes in a cluster (Fig. 4b–f and Supplementary Table 9). Below, we provide examples of how the data recapitulate known genetic relationships and provide insights into the functions of poorly characterized genes.
Fig. 4. Similarity of mutant phenotypes places genes into pathways and reveals new players.

a, A total of 684 genes were clustered based on the similarity of their phenotypes across 120 screens. b–f, Examples of how subclusters enriched in specific pathways predict new genes in these pathways: nonphotoautotrophic light insensitive (b), nonphotoautotrophic light sensitive (c), cilia (d), DNA damage sensitive (e) and LatB sensitive (f).
Extended Data Fig. 3. Comparison of phenotypic and transcriptomic correlations.

a, 684 genes were clustered based on the similarity of their phenotypes across 120 screens. b, Spearman correlation matrix of phenotypes (FDR < 0.3). c, Transcriptome correlation of gene with phenotype (FDR < 0.3). d, 194 genes were clustered based on the similarity of their phenotype across 120 screens. e, Spearman correlation matrix of phenotypes (FDR < 0.05). f, Transcriptome correlation of gene with phenotype (FDR < 0.05). Data can be found in Supplementary Table 10.
Essential DNA repair pathways are conserved in green algae
DNA damage repair pathways are among the best-characterized and most highly conserved across all organisms17,18; thus, they serve as a useful test case of the quality of our data. In our dataset, homologues of known DNA repair proteins are present in a large cluster (Fig. 4e), demonstrating the quality of our phenotypic data, validating our ability to identify that these genes work in a common pathway and extending the conservation of their functions to green algae.
Mutants for various DNA repair genes exhibit expected differences in their sensitivities to different types of DNA damage: (1) DNA double-strand breaks (zeocin and bleomycin), (2) DNA crosslinks (mitomycin C and cisplatin) and (3) DNA alkylation (MMS). For example, mutants exhibiting sensitivity to all DNA damage conditions included mutants lacking upstream DNA damage-sensing kinase ATAXIA TELANGIECTASIA AND RAD3-related protein (ATR, encoded by Cre10.g467200) (ref. 19), as well as mutants lacking the cell cycle checkpoint control protein RADIATION SENSITIVE 9 (RAD9, encoded by Cre16.g682950) or its binding partner HYDROXYUREA-SENSITIVE 1 (HUS1, encoded by Cre12.g524350) (ref. 20). Mutants specifically sensitive to the double-strand break-inducing agents zeocin and bleomycin included the upstream sensor of double-strand breaks, the kinase ATAXIA-TELANGIECTASIA MUTATED (ATM, encoded by Cre13.g564350) (ref. 21) (Supplementary Table 6) and DNA POLYMERASE THETA (POLQ, encoded by Cre16.g664301), which facilitates error-prone double-strand break repair and can maintain genome integrity when other repair pathways are insufficient22,23 (Supplementary Table 6). Mutants specifically sensitive to the DNA crosslinker cisplatin included cells with genetic lesions in the helicases REGULATOR OF TELOMERE ELONGATION HELICASE 1 (RTEL1, encoded by Cre02.g089608) (ref. 24) and FANCONI ANEMIA COMPLEMENTATION GROUP M (FANCM, encoded by Cre03.g208833) and in the crossover junction endonuclease METHANSULFONATE UV SENSITIVE 81 (MUS81, encoded by Cre12.g555050).
Our data suggest several instances where a given factor is required for the repair of a specific class of DNA damage in Chlamydomonas, but not in Arabidopsis, or vice versa, suggesting lineage-specific differences in how DNA damage is repaired. For example, Chlamydomonas fancm mutants are sensitive to the DNA crosslinker cisplatin, whereas Arabidopsis fancm mutants are not25. Conversely, Arabidopsis mus81 mutants are sensitive to the alkylating agent MMS and the DNA crosslinker mitomycin C26, whereas Chlamydomonas mus81 mutants were not.
Taken together, our data suggest that the core eukaryotic DNA repair machinery defined in other systems is generally conserved in green algae. Moreover, the observation of expected phenotypes illustrates the quality of the presented data and the utility of the platform for chemical genomic studies.
Classification of genes based on photosynthesis phenotypes
Our data allowed the classification of 38 genes whose disruption leads to a photoautotrophic growth defect into two clusters. One cluster consisted of genes whose disruption confers sensitivity to light when grown on medium supplemented with acetate, whereas the other contained genes whose disruption does not (Fig. 4b,c and Supplementary Data 2).
The light-sensitive cluster (Fig. 4c) included genes encoding core photosynthesis components and biogenesis factors such as the mRNA trans-splicing factors RNA MATURATION Of PSAA (RAA1)27, RAA328, OCTOTRICOPEPTIDE REPEAT 120 (OPR120) and OPR10429; photosystem II biogenesis factor CONSERVED IN PLANT LINEAGE AND DIATOMS 10 (CPLD10)29,30; the chlorophyll biogenesis factor Mg-CHELATASE SUBUNIT D (CHLD)31; the ATP synthase translation factor TRANSLATION DEFICIENT ATPase 1 (TDA1)32; the Rubisco mRNA stabilization factor MATURATION OF RBCL 1 (MRL1)33; and the Calvin–Benson–Bassham cycle enzymes SEDOHEPTULOSE-BISPHOSPHATASE 1 (SEBP1)34 and PHOSPHORIBULOKINASE 1 (PRK1)35. Several highly conserved but poorly characterized genes are also found in this cluster, including the putative Rubisco methyltransferase encoded by Cre12.g52450036, the putative thioredoxin Cre01.g037800, the predicted protein with a domain of unknown function (DUF1995) Cre06.g281800 (which we named LIGHT SENSITIVE AND/OR ACETATE-REQUIRING 4 (LSAR4)) and Cre13.g572100 (which we named LIGHT GROWTH SENSITIVE 4 (LGS4)), as well as four Chlorophyta-specific genes. The mutant phenotypes of these poorly characterized genes and their presence in this light-sensitive cluster together suggest that their products could mediate the biogenesis, function or regulation of core components of the photosynthetic machinery.
The low CO2-sensitive cluster (Fig. 4b) contains known and new components of the algal CO2-concentrating mechanism (CCM), as detailed below.
New CCM components
The CCM increases the CO2 concentration around the CO2-fixing enzyme Rubisco, thus enhancing the rate of carbon uptake. The mechanism uses carbonic anhydrases in the chloroplast stroma to convert CO2 to HCO3−, which is transported into the lumen of the thylakoid membranes that traverse a Rubisco-containing structure called the pyrenoid37. There, the lower pH drives the conversion of HCO3− back into concentrated CO2 that feeds Rubisco37. Mutants deficient in the CCM are unable to grow photoautotrophically in air, but their photoautotrophic growth is rescued in 3% CO2 (ref. 37). We observed this phenotype for one or more alleles of genes whose disruption was previously shown to disrupt the CCM (Supplementary Table 4), including genes encoding the chloroplast envelope HCO3− transporter LOW CO2 INDUCIBLE GENE A (LCIA)38, and the thylakoid lumen CARBONIC ANHYDRASE 3 (CAH3) (ref. 39), the stromal carbonic anhydrase LOW CO2 INDUCIBLE GENE B (LCIB)40, the master transcriptional regulator CCM1/CIA5 (refs. 41,42) and the pyrenoid structural protein STARCH GRANULES ABNORMAL 1 (SAGA1) (ref. 43) (Supplementary Table 6).
We observed similar high CO2 rescue of photoautotrophic growth defects for mutants in multiple poorly characterized genes in the light-insensitive cluster, suggesting that many of these genes are new components in the CCM. These genes formed a cluster with SAGA1 (ref. 43), the only previously known CCM gene with enough alleles to be present in the cluster. We named one of these genes, Cre06.g259100, SAGA3 (STARCH GRANULES ABNORMAL FAMILY MEMBER 3) because its protein product shows homology to the two pyrenoid structural proteins SAGA1 and SAGA2 (ref. 44) (Extended Data Fig. 4). Consistent with a role in the CCM, SAGA3 localizes to the pyrenoid45. We also observed this phenotype in mutants lacking the pyrenoid starch sheath-localized protein STARCH BRANCHING ENZYME 3 (SBE3) (ref. 46), suggesting that this enzyme plays a key role in the biogenesis of the pyrenoid starch sheath, a structure surrounding the pyrenoid that was recently shown to be important for pyrenoid function under some conditions47. Our cluster also contains the gene encoding FUZZY ONIONS (FZO)-like (FZL), a dynamin-related membrane remodeling protein involved in thylakoid fusion and light stress; mutants in this gene have pyrenoid shape defects48. Our results suggest that thylakoid organization influences pyrenoid function. The cluster additionally includes genes encoding CLV1 (Cre13.g574000), a predicted voltage-gated chloride channel that we hypothesize is important for regulating the ion balance in support of the CCM or, alternatively, may directly mediate HCO3− transport; a protein containing a Rubisco-binding motif44 (Cre12.g528300, which we named LOW CO2 SENSITIVE 1 (LOCO1)); and a predicted Ser-Thr kinase HIGH LEAF TEMPERATURE 1 (HT1) (Cre02.g111550). The kinase is a promising candidate for a regulator in the CCM, as multiple CCM components are known to be phosphorylated49–51, but no kinase had previously been shown to have a CCM phenotype.
Extended Data Fig. 4. SAGA protein alignments.

a, Alignments of SAGA1, SAGA2, and SAGA3. Domain annotation was based on three different tools under the Geneious visualization platform. b, BLOSUM90 alignments between SAGA proteins.
Also in this cluster are genes encoding the predicted PYRUVATE DEHYDROGENASE 2 (PDH2) (Cre03.g194200) and the predicted DIHYDROLIPOYL DEHYDROGENASE (DLD2) (Cre01.g016514). We hypothesize that these proteins are part of a glycine decarboxylase complex that functions in photorespiration, a pathway that recovers carbon from the products of the Rubisco oxygenation reaction. PDH2 was found in the pyrenoid proteome52, suggesting the intriguing possibility that glycine decarboxylation may be localized to the pyrenoid, where the released CO2 could be recaptured by Rubisco.
New genes with roles in cilia function
Chlamydomonas cells swim using two motile cilia. To identify mutants with abnormal cilia function, we separated cells based on swimming ability by placing the mutant pool in a vertical column and collecting the supernatant and pellet. In this assay, mutants with altered swimming behavior were enriched in GO terms such as ‘dynein complex’, which comprises motor proteins involved in ciliary motility (Fig. 2e). Eighteen genes were represented by enough alleles to provide high confidence (FDR < 0.3) that their disruption produces a defect in swimming (Fig. 4d). These genes were enriched (P = 0.0075, Fisher’s exact test) in genes encoding proteins found in the Chlamydomonas flagella proteome53. Half of these genes or their orthologs have previously been associated with a cilia-related phenotype in Chlamydomonas and/or mice (Supplementary Table 11).
In our analysis, these 18 genes formed four clusters that appeared to subclassify their function (Fig. 4d). The first cluster is enriched in known regulators of ciliary membrane composition and includes the gene encoding NEPHROCYSTIN-4-LIKE PROTEIN (NPHP4)54; the gene encoding its physical interactor TRANSMEMBRANE PROTEIN 67 (TMEM67, also named MECKEL SYNDROME TYPE 3 (MKS3) in mammals), which has been implicated in photoreceptor intraciliary transport55; and the gene encoding CENTRIOLE PROTEOME PROTEIN 290 (CEP290) (ref. 56). We validated the swimming defect of tmem67 and observed that the mutant has shorter cilia (Extended Data Fig. 5). The poorly annotated gene Cre15.g638551 clusters with these genes, suggesting that it may also regulate ciliary membrane composition.
Extended Data Fig. 5. Validation of cilia mutant phenotypes.

a, Bright-field microscopy microscope images of cilia mutants show defects in ciliary length. Scale bar: 10 µm. b, Quantification of cilia length. n =number of samples (WT n = 60, tmem67-1 n = 112, tmem67-2 n = 46, fap81-1 n = 139). c, Swimming behavior of mutants, as determined by growth on TAP medium solidified with 0.15% agar. Scale bars: 5 cm.
The second cluster contains genes encoding BARDET-BIEDL SYNDROME 1 PROTEIN 1 (BBS1) and BBS9, components of the Bardet–Biedl syndrome-associated complex that regulates targeting of proteins to cilia57. The poorly annotated gene Cre15.g640502 clustered with these genes, suggesting that it may also play a role in targeting proteins to cilia.
The third cluster contains eight genes, four of which relate to the dynein complex, including the ciliary dynein assembly factor DYNEIN ASSEMBLY LEUCINE-RICH REPEAT PROTEIN (DAU1) (ref. 58,59), OUTER DYNEIN ARM (ODA), DYNEIN ARM INTERMEDIATE CHAIN 1 (DIC1) (ref. 60), DYNEIN HEAVY CHAIN 1 (DHC1) (ref. 61) and TUBULIN-TYROSINE LIGASE 9 (TTLL9), which modulates ciliary beating through the addition of polyglutamate chains to alpha-tubulin62. The predicted thioredoxin peroxidase gene Cre04.g218750 and three poorly annotated genes (Cre07.g338850, Cre01.g012900 and Cre16.g675600) clustered with these genes, suggesting possible roles in dynein assembly or regulation.
The fourth cluster contains three poorly characterized genes, FLAGELLA ASSOCIATED PROTEIN2 (FAP2), FLAGELLA ASSOCIATED PROTEIN 81 (FAP81) and TEF24. The protein encoded by FAP81 (Cre06.g296850) was identified in the Chlamydomonas cilia proteome53, and its human homologue DELETED IN LUNG AND ESOPHAGEAL CANCER PROTEIN 1 (DLEC1) localizes to motile cilia63. We validated the swimming defect of the fap81 mutant and established that it has shorter cilia (Extended Data Fig. 5). The localization to motile cilia in humans and our finding that mutating the encoding gene leads to a ciliary motility defect together suggest the intriguing possibility that impaired cilia motility contributes to certain lung and esophageal cancers.
New genes required for actin cytoskeleton integrity
Our analysis revealed a group of genes that render cells sensitive to LatB when any are mutated (Fig. 4f). LatB binds to monomers of actin, one of the most abundant and conserved proteins in eukaryotic cells, and prevents actin polymerization64 (Fig. 5a). LatB was first discovered as a small molecule that protects the sea sponge Latrunculina magnifica from predation by fish65 and is an example of the chemical warfare that organisms use to defend themselves and compete in nature (Fig. 5b).
Fig. 5. The approach revealed new conserved components of a defense mechanism against cytoskeleton inhibitors.

a, LatB interferes with actin polymerization. b, Soil microorganisms deploy (arrows) actin inhibitors (blue circles) for a competitive advantage in their environment. c, Chlamydomonas responds to actin inhibition by degrading its conventional actin, IDA5, and upregulating an alternative actin, NAP1. d, Growth of new lat mutants identified in this study (lat5-1, lat6-1 and lat7-2) was compared to previously isolated lat1-5, lat2-1, lat3-1 and nap1-1 mutants66 in the absence (control) and presence (LatB) of 3 µM LatB. e, Immunoblot of conventional (IDA5) and alternative (NAP1) actins shows that lat5-1, lat6-1 and lat7-2 are deficient in actin degradation. Immunoblot representative of n = 3 independent experiments. f, The F-actin homeostasis pathway is conserved between green algae and plants. Mutants in Arabidopsis genes homologous to Chlamydomonas lat3, lat5 and lat6 are sensitive to LatB, as evidenced by decreased root length. g, Quantification of root length in Arabidopsis mutants. Asterisks mark significant changes relative to wild type under the same condition based on two-way analysis of variance. The exact value of P = 2.4 × 10–47 (Ler versus lat3), P = 1.4 × 10–62 (Col-0 versus lat5), P = 6.8 × 10–23 (Col-0 versus lat6). n = 26 roots examined over three independent experiments.
Chlamydomonas protects itself against LatB-mediated inhibition of its conventional actin INNER DYNEIN ARM5 (IDA5) by upregulating the highly divergent actin homologue NOVEL ACTIN-LIKE PROTEIN 1 (NAP1), which appears to perform most of the same functions as actin but is resistant to inhibition by LatB66. Upon inhibition of IDA5 by LatB, IDA5 is degraded and the divergent actin NAP1 is expressed66. The expression of NAP1 is dependent on three other known genes, LatB-SENSITIVE1-3 (LAT1–LAT3) (Fig. 5c); thus, mutants lacking any of these four genes are highly sensitive to LatB66.
Our phenotype data revealed three new components of this F-actin homeostasis pathway, which we named LAT5 (encoded by Cre17.g721950), LAT6 (encoded by Cre15.g640101) and LAT7 (encoded by Cre11.g482750). LAT5 and LAT6 clustered together with three previously known components of the pathway (NAP1, LAT2 and LAT3), and disruption of all six genes rendered cells sensitive to LatB (Supplementary Table 6). Mutants in all three new components show a relatively mild phenotype when compared to those mutants in LAT1–LAT3 (Fig. 5d), illustrating the sensitivity of our phenotyping platform.
Ubiquitin proteasome-mediated proteolysis of IDA5 has been hypothesized to drive the degradation of IDA5 and promote the formation of F-NAP1 (ref. 67), but the factors involved were unknown. LAT5 and LAT6 encode predicted subunits of a SKP1, CDC53/CULLIN, F-BOX RECEPTOR (SCF) E3 ubiquitin ligase complex, whose homologues promote the degradation of target proteins68. The disruption of LAT5 and LAT6 impaired degradation of IDA5 upon LatB treatment, suggesting that LAT5 and LAT6 mediate IDA5 degradation (Fig. 5e). LAT7 encodes a predicted importin, and its disruption impairs IDA5 degradation after LatB treatment (Fig. 5e), suggesting that nuclear import is required for IDA5 degradation.
It was previously not clear how broadly conserved this F-actin homeostasis pathway is. We found that the land plant model Arabidopsis has homologs of IDA5, NAP1, LAT3, LAT5, LAT6 and LAT7. We observed that Arabidopsis mutants disrupted in LAT3, LAT5 and LAT6 are sensitive to LatB treatment (Fig. 5f,g), which was not expected a priori, suggesting that this pathway for actin cytoskeleton integrity and the gene functions identified here are conserved in land plants.
Discussion
In this work, we determined the phenotypes of 58,101 Chlamydomonas mutants across a broad variety of growth conditions. We observed a phenotype for mutants representing 10,380 genes, providing a valuable starting point for characterizing the functions of thousands of genes. Mutant phenotypes are searchable at chlamylibrary.org, and individual mutants can be ordered from the Chlamydomonas Resource Center.
We provided several examples of how the data enable discovery of gene functions and phenotypes in algae and plants. We validated our discovery of three new genes in the actin cytoskeleton integrity pathway, obtained insights into their molecular functions and found that this pathway appears to be conserved in land plants. We validated our discovery of cilia function defects for two new genes and our observation of an unexpected sensitivity of the chloroplast unfolded protein response to the alkylating agent MMS. We also discussed how our data provide insights and candidate genes in other pathways, including DNA damage repair, photosynthesis and the CCM.
Altogether, 58% of the high-confidence gene–phenotype interactions involve a Chlamydomonas gene with a predicted Arabidopsis homologue (Supplementary Table 8); for approximately 79% of the corresponding Arabidopsis homologues, our data predict a new gene–phenotype relationship. This work illustrates the value of using a microbial photosynthetic organism for discovering gene functions on a large scale. We hope that the genotype–phenotype relationships identified here will guide the characterization of thousands of genes, with potential applications in agriculture, the global carbon cycle and our basic understanding of cell biology.
Methods
Library maintenance
The Chlamydomonas mutant collection12 was maintained by robotically passaging 384-colony arrays to fresh medium using a Singer RoToR robot (Singer Instruments, 704). The mutant collection was grown on 1.5% agar Tris-acetate-phosphate (TAP) medium with modified trace elements69 in complete darkness at room temperature. The routine passaging interval of 4 weeks for library maintenance was shortened to 2 weeks during the time period of pooled screens to increase cell viability.
Screening of LATCA to identify Chlamydomonas growth inhibitors
LATCA13 was used to identify molecules capable of inhibiting growth in wild-type Chlamydomonas (cMJ030). We found that 1,222 of these 3,650 LATCA compounds reduce growth by 90% at 25 μM (Supplementary Table 3). Due to resource limitations, we could not perform competitive growth experiments with all 1,222 active chemicals. Hence, we further selected the most active compounds and analyzed their structural similarity to identify the most diverse set of compounds for the competitive growth experiments. Dose–response experiments with 1,140 compounds validated activity for 954 compounds, and identified 136 chemicals that reduce growth at 2 µM or less (Supplementary Table 3). We then used the extended-connectivity fingerprint algorithm70 to convert all LATCA compound structures into numerical fingerprints. Extended-connectivity fingerprints were then used to compute structural similarity of pairs of compounds on a scale of 0 to 1 using Tanimoto coefficients71. The set of Tanimoto coefficients between all pairs of inhibitors was visualized using Cytoscape72. We then used the most active inhibitors to further reduce the similarity network to 28 clusters of structures exhibiting high levels of biological activity and selected 52 of these chemicals for subsequent treatment of the Chlamydomonas mutant library (Extended Data Fig. 1, Supplementary Table 3 and Supplementary Data 1).
Library pooling and competitive growth experiments
The first two rounds of mutant library screening (R1 and R2) were performed with the entire mutant collection (550 384-mutant array plates) in 20-liter carboys (Supplementary Table 1 and Supplementary Table 2). Mutants were pooled from 5-day-old 384-colony array plates into liquid TAP medium at room temperature and low light. In R1, the pool included nine additional copies for each of three plates (668–670) in the collection to test how quantitatively we can track the relative abundance of mutants in the starting population. In R2, we pooled a subset of the mutant collection (plates 597–670) from 384-colony array plates and another subset from 1,536-colony array plates (101–596) to test the performance of denser colony arrays for pooled screens.
Subsequent rounds of mutant library screening (R3–R6) were performed on the rearrayed library (245 384-mutant array plates) in 2-liter bottles. Mutants were pooled from 5-day-old 1,536-colony array plates. Condensing the library from 384 to 1,536-colony array plates helped to both homogenize colony growth and reduce the laborious pooling procedure.
We produced subpools each containing cells from eight 384 or 1,535-colony array plates by using sterile glass spreaders to pool cells from the plates into 50-ml conical tubes containing 40 ml TAP medium. These subpools were mixed by pipetting to break cell clumps using a 10-ml serological pipette with a P200 tip attached to it. Then, all subpools were combined into the final mutant collection pool by pipetting the subpools through a 100-µm cell strainer (VWR, 10054-458). The final pool was mixed using a magnetic stir bar, and the cell density was measured (Invitrogen, Countess) and adjusted to 1 × 105 cells ml−1. For experiments not performed in TAP medium, cells were pelleted (1,000× g, 5 min, room temperature) and washed twice with the actual medium used for the pooled growth experiment.
Aliquots of 2 × 108 cells were pelleted (1,000× g, 5 min, room temperature) by centrifugation and frozen to determine the relative abundance of each mutant in the starting population. These samples are denoted as ‘initial’.
Cultures were inoculated with 2 × 104 cells ml−1 in transparent 20-liter carboy tanks (R1 and R2) or standard 2-liter bottles (R3–R6) using aliquots of the final mutant pool. Cultures were inoculated with 2 × 104 cells ml−1, and most experiments were performed in 2-liter vessels (4 × 107 cells total) with ~58,000 mutants (Supplementary Tables 1 and 2), resulting in ~700 cells per mutant on average in the 2-liter competitive growth experiments. Cultures were grown under a broad variety of conditions (Supplementary Table 2) of which 49 had two or more replicates. Unless otherwise indicated, cells were grown in TAP medium with modified trace elements at pH 7.5 under constant light (100 µmol photons m−2 s−1 using Lumigrow Lumibar lights, catalog number 8100-5502; equal levels of red, blue and white light) at 22 °C, aerated with air and mixed using a conventional magnetic stirrer at 200 rpm. The cell density of competitive growth experiments was tracked, and aliquots of 2 × 108 cells were pelleted by centrifugation after approximately seven doublings, when the culture reached approximately 2 × 106 cells ml−1. We sought to avoid letting the cultures reach stationary phase, where experiments are less reproducible. At seven divisions, the mutant pool was typically in the late exponential growth phase. Cell pellets were frozen for subsequent DNA extraction and barcode quantification. Algal predator experiments were performed with Daphnia magna, Philodina sp. (Rotifer) and Hypsibius exemplaris (Tardigrade) purchased from Carolina Biological Supply.
DNA extraction
Total genomic DNA was extracted from frozen cell pellets representing 2 × 108 cells of each sample (initial, control and treatment).
First, frozen pellets were thawed at room temperature and resuspended in 1.6 ml resuspension buffer (1% SDS, 200 mM NaCl, 20 mM EDTA and 50 mM Tris-HCl, pH 8.0).
Second, 2 ml phenol/chloroform/isoamyl alcohol (25:24:1) was added to each sample and mixed by vortexing. This solution was then transferred into 15-ml Qiagen MaXtract High Density tubes (catalog number 129065) and centrifuged at 3,500 rpm for 5 min. Subsequently, the aqueous phase was transferred to a new 15-ml conical tube, 6.4 µl RNase A was added and the solution was incubated at 37 °C for 30 min. The phenol/chloroform/isoamyl alcohol extraction was then repeated, and the aqueous phase was transferred into a new 15-ml Qiagen MaXtract High Density tube before adding 2 ml phenol/chloroform/isoamyl alcohol (25:24:1). This solution was mixed by vortexing and centrifuged at 3,500 rpm for 5 min; then, 400-µl aliquots of the aqueous phase were transferred to 1.5-ml reaction tubes for DNA precipitation (typically four aliquots per sample).
Third, 1 ml ice-cold 100% ethanol was added to the solution to precipitate DNA. The tubes were gently mixed and incubated at –20 °C overnight. The DNA was pelleted at 13,200 rpm and 4 °C. The supernatant was discarded and the pellet washed in 1 ml 70% ethanol. The supernatant was discarded again, and the pellet was air-dried before resuspension in 50 µl water. Subsequently, the elution fractions of each sample were pooled and the DNA concentration was measured using a Qubit fluorometer (Invitrogen).
Internal barcode amplification and Illumina library preparation
Internal barcodes were amplified using Phusion Hot Start II (HSII) DNA Polymerase (Thermo Fisher, F549L) using previously described primers12.
The 50-µl PCR mixture for 5′ barcode amplification contained 125 ng genomic DNA, 10 µl GC buffer, 5 µl DMSO, 1 µl dNTPs at 10 mM, 1 µl MgCl2 at 50 mM, 2.5 µl of each primer at 10 µM and 1 µl Phusion HSII polymerase. Eight tubes of the PCR mixture were processed per sample and incubated at 98 °C for 3 min, followed by 10 three-step cycles (98 °C for 10 s, 58 °C for 25 s and 72 °C for 15 s) and then 11 two-step cycles (98 °C for 10 s and 72 °C for 40 s).
The 50-µl PCR mixture for 3′ barcode amplification contained 125 ng genomic DNA, 10 µl GC buffer, 5 µl DMSO, 1 µl dNTPs at 10 mM, 2 µl MgCl2 at 50 mM, 2.5 µl of each primer at 10 µM and 1 µl Phusion HSII polymerase. Eight tubes of the PCR mixture were processed per sample and incubated at 98 °C for 3 min, followed by 10 three-step cycles (98 °C for 10 s, 63 °C for 25 s and 72 °C for 15 s) and then 11 two-step cycles (98 °C for 10 s and 72 °C for 40 s).
The PCR products of each sample were pooled for further processing. First, successful PCR was confirmed on a TBE 8% agarose gel in 1x Tris Borate EDTA before concentrating the PCR products on a Qiagen MinElute column and measuring the DNA concentration on a Qubit fluorometer. Second, 200–250 ng of up to 16 3′ or 5′ PCR products was combined into an Illumina HiSeq2000 library. Third, the internal barcode bands of the Illumina HiSeq2000 libraries were gel-purified and subjected to quality control on an Agilent Bioanalyzer. In addition, DNA concentration was determined on a Qubit fluorometer. Fourth, HiSeq2000 libraries were sequenced at the Genome Sequencing Service Center at Stanford University (Palo Alto, CA).
Data analysis
Initial reads were trimmed using cutadapt version 1.7.1 (ref. 73) using the command ‘cutadapt -a <seq > -e 0.1 -m 21 -M 23 input_file.gz -o output_file.fastq’, where <seq> is GGCAAGCTAGAGA for 5′ data and TAGCGCGGGGCGT for 3′ data. Barcodes were counted by collapsing identical sequences using ‘fastx_collapser’ (http://hannonlab.cshl.edu/fastx_toolkit) and denoted as ‘_read_count’. Across all experiments conducted, ~62 million barcode read counts were determined. Barcode read counts for each dataset were normalized to a total of 100 million and denoted as ‘_normalized_reads’ (Supplementary Table 12). Replicate control treatments performed in the same screening round were averaged by taking the mean of the normalized read counts to generate the average normalized read count (denoted as ‘_average_normalized_reads’). To calculate a ‘_read_count’ (nonnormalized) for the averaged samples, the read counts for all of the averaged samples were summed and denoted as the ‘_average_read_count’. Control treatments that were averaged are denoted with ‘average’ and can be found in Supplementary Table 13.
Mutants in the library contain on average 1.2 insertions12, each of which may contain a 5′ barcode, a 3′ barcode, both barcodes or potentially more than two barcodes if multiple cassettes were inserted at the loci. To represent a given insertion within a mutant, we selected a single barcode to represent it. All barcodes associated with the same gene and deconvoluted to the same library well and plate position were assumed to be from the same insertion and were then compared to identify the barcode with the highest read counts in the initial samples (R2–R6) to serve as the representative barcode.
To identify mutants with growth defects or enhancements due to a specific treatment, we compared the abundance of each mutant after growth under the treatment condition to its abundance after growth under a control condition. We called this comparison a ‘screen’ and the ratio of these abundances the ‘mutant phenotype’. In order for a phenotype to be calculated, we required the control treatment to have a read count above 50, which allowed for 16.8 million phenotypes to be determined.
To identify high-confidence gene–phenotype relationships we developed a statistical framework that leverages multiple independent mutant alleles. For each gene, we generated a contingency table of the phenotypes, Φ, by counting the number of alleles that met the following thresholds: [Φ < 0.0625, 0.0625 ≤ Φ < 0.125, 0.125 ≤ Φ < 0.25, 0.25 ≤ Φ < 0.5, 0.5 ≤ Φ < 2.0, 2.0 ≤ Φ < 4.0, 4.0 ≤ Φ < 8.0, 8.0 ≤ Φ < 16.0]. Only alleles that were mapped with confidence level 4 or less (corresponding to a likelihood of correct mapping of 58% or higher)12 had an insertion in CDS/intron/5′ UTR feature, and had greater than 50 reads in the control condition were included in the analysis. The frequency of cassette insertion location based on gene feature was intron 25%, 3′ UTR 23%, CDS 19%, not mapped 14%, intergenic 6%, 5′ UTR 5% and multiple or others 8%. The insertion cassette used to generate the mutants contains two transcriptional terminators; thus, we reasoned that insertions in 5′ UTRs, introns and exons will lead to transcriptional disruption and loss-of-function mutants. Mutants with cassette integrations in the 3′ UTR were not expected to result in transcriptional disruption so were excluded from our statistical framework. A P value was generated for each gene by using Fisher’s exact test to compare a gene’s phenotype contingency table to a phenotype contingency table for all insertions in the screen. An FDR was performed on the P values of genes with more than two alleles using the Benjamini–Hochberg method74.
To determine a representative phenotype for a gene, the median phenotype for all alleles of that gene that were included in the Fisher’s exact test was used. For clustering, these gene phenotypes were normalized by setting the median value of all gene phenotypes in a screen to zero. Clustering was performed with Python (2.7.11) packages SciPy (0.17.0) (ref. 75) and visualized with Seaborn (0.7.1). To generate the hierarchical cluster in Fig. 4a, the pairwise distance was calculated using the ‘correlation’ metric, which calculates the correlation (Pearson) distance. The linkage matrix was calculated using the ‘average’ method. Pairwise Pearson correlation coefficients between gene phenotypes (Extended Data Fig. 3 and Supplementary Table 10) were calculated in Pandas (0.18.1). Transcriptome correlation data was collected, curated and analyzed in the Merchant laboratory16. Data were plotted and visualized with the Python packages Matplotlib (1.5.1) and Seaborn (0.7.1).
To determine if biological functions were associated with specific screens, we performed a GO term enrichment analysis. Using the same approach as with genes, we generated contingency tables of mutant phenotypes for each GO term. If a mutant’s insertion is within a gene that had multiple GO term annotations, the mutant’s phenotype data was added to each GO term’s contingency table. A P value was generated for each GO term by using Fisher’s exact test to compare a GO term’s phenotype contingency table to a phenotype contingency table for all GO terms in the screen. An FDR was performed on the P values using the Benjamini–Hochberg method74. Clustering was performed (Extended Data Fig. 2) in Seaborn using the ‘Euclidean’ metric to calculate the pairwise distance and the ‘average’ method to calculate the linkage.
All analysis was performed using JGI Phytozome release v5.0 of the Chlamydomonas assembly and v5.6 of the Chlamydomonas annotation76. Gene identifiers (CreXX.gXXXXXX) can be used to link data found in the supplemental tables to gene annotation updates. All data have been deposited in Dryad Digital Repository (10.6086/D1Q96Z). Custom code used for data analysis has been deposited in Zenodo (10.5281/zenodo.6340170) (ref. 77).
MMS growth assays and VIPP2 immunoblot analysis
The following strains were used15: WT = CC-4533; mars1 = mars1-3; mars1:MARS1-D = mars1-3 transformed with the MARS1-D transgene containing a 3×-Flag epitope after Met139; and mars1:MARS1-D KD = mars1-3 transformed with a catalytically-inactive MARS1-D bearing the kinase active site D1871A mutation. Before starting liquid cultures in TAP medium, all strains were restreaked in fresh TAP plates and grown in similar light conditions (i.e., ~50–70 µmol photons m−2 s−1, ~22 °C) for about 5–6 days. Before starting the MMS treatment, all strains were preconditioned in liquid cultures for 3 or 4 days. Next, cell cultures were equally diluted to ~5 µg chlorophyll ml−1 and incubated in the presence or absence of MMS for 48 h. A 1% (vol/vol) MMS stock solution (Sigma-Aldrich, 129925) was freshly prepared in double-distilled H2O at the beginning of each experiment. This MMS stock solution was further diluted 200 times directly into TAP medium to a final concentration of 0.05% (vol/vol). All chlorophyll concentration measurements were performed using a previously described methanol extraction method78.
VIPP2 and alpha-TUBULIN immunoblot analyses were carried out as described previously15 using denatured total protein samples prepared from liquid cultures incubated for 27 h in the presence or absence of 0.05% (vol/vol) MMS.
Cilia-related mutant phenotyping
Cilia mutants were grown in liquid TAP medium until they reached exponential phase. Cells were then mounted in u-Slide 8-well chambers (Ibidi, 80826) with 2% low-melting-point agarose (Sigma-Aldrich, A9414). Cilia defects were scored using a Leica DMi8 inverted microscope. Cilia length was measured using Fiji. Cilia swimming behavior was scored using TAP agar plates with 0.15% agar.
Chlamydomonaslat mutant phenotyping
Mutants used in this study are listed in Supplementary Table 14, and sequence information for all genotyping primers is summarized in Supplementary Table 15. Individual mutants were grown with gentle agitation at 100 µmol photons m−2 s−1. Disruption of LAT5, LAT6 and LAT7 genes (Cre17.g721950, Cre15.g640101 and Cre11.g482750) in the original isolates of lat5-1, lat6-1 and lat7-2 were confirmed by PCR. These mutants were then backcrossed with CC-124 or CC-125 three times, with perfect linkage of paromomycin resistance and LatB sensitivity in at least 10 tetrads observed after each round. The backcrossed strains and the previously established lat1-5, lat2-1, lat3-1 and nap1-1 mutants in the CC-124 background66 were spotted on TAP agar containing 0.1% DMSO with or without 3 µM LatB (Adipogen, AG-CN2-0031, lot A00143/J) as 5× serial dilutions.
Immunoblot materials IDA5 and NAP1 immunoblot analyses
Cells were grown in liquid TAP medium at 21 °C with gentle agitation under 100 µmol photons m−2 s−1 and collected by centrifugation. Pellets were frozen in liquid nitrogen and subsequently resuspended in 100 µl ice-cold PNE buffer (10 mM phosphate, pH 7.0, 150 mM NaCl2 and 2 mM EDTA) supplemented with 2× concentration of complete protease-inhibitor cocktail (Roche, 11697498001) and disrupted by vortexing with acid-washed glass beads. These samples were mixed directly with SDS–PAGE sample buffer, boiled for 3 min and cleared of debris by centrifugation at 12,000× g for 10 min at 4 °C before electrophoresis. SDS–PAGE was performed using 11% Tris-glycine. Blots were stained using a mouse monoclonal anti-actin antibody (EMD Millipore, clone C4, MAB1501), which recognizes IDA5, but not NAP1, and a rabbit anti-NAP1 antibody (generous gift from R. Kamiya and T. Kato-Minoura), which recognizes NAP1, but not IDA5. Horseradish peroxidase-conjugated anti-mouse IgG (ICN Pharmaceuticals, 55564) or anti-rabbit IgG (Southern Biotech, 4050-05) were used as secondary antibodies, respectively.
Arabidopsislat mutant phenotyping
Mutants used in this study are listed in Supplementary Table 14 and sequence information for all genotyping primers is summarized in Supplementary Table 15. Seeds were surface-sterilized in 20% bleach for 5 min. Seeds were then rinsed with sterile water four times and stored at 4 °C for 3 days in the dark. After stratification, seeds were sown into square 10 cm × 10 cm petri plates containing full-strength Murashige and Skoog medium (MSP01-50LT), 1% agar (Duchefa, 9002-18-0), 1% sucrose and 0.05% MES and adjusted to pH 5.7 with 1 M KOH. Seedlings were grown in the presence of LatB (Sigma, L5288) or mock control containing an equivalent volume of the LatB solvent, DMSO. Plates were imaged using a CanonScan 9000 flatbed scanner. Root lengths were quantified using Fiji. Two-way analysis of variance and data visualization were done using Python.
Reporting Summary
Further information on research design is available in the Nature Research Reporting Summary linked to this article.
Online content
Any methods, additional references, Nature Research reporting summaries, source data, extended data, supplementary information, acknowledgements, peer review information; details of author contributions and competing interests; and statements of data and code availability are available at 10.1038/s41588-022-01052-9.
Supplementary information
List of supplementary materials
Spreadsheet with Supplementary Tables 1–3, 5–11 and 13–15
Zip file that contains the chemical structure of all LATCA compounds used in this study.
Zip file that contains the Java TreeView files of the FDR < 0.3 gene cluster.
Acknowledgements
We thank M. Cahn for developing and improving the CLiP website; X. Ji at the Stanford Functional Genomics Facility and Z. Weng at the Stanford Center for Genomics and Personalized Medicine for deep-sequencing services; K. Barton, W. Briggs and Z.-Y. Wang for providing lab space; T. Raab for providing lab equipment; J. Robertson and N. Ivanova for helping with library maintenance; M. Sison-Mangus for advice on Daphnia experiments; T. Kato-Minoura and R. Kamiya for the anti-NAP1 antibody; J. R. Pringle for supporting M.O.; J. Whitney for assistance in figure preparation; and members of the Dinneny, Jonikas and Jinkerson Labs and T. Xiang for constructive suggestions on the manuscript. This project was supported by grants from the National Institutes of Health (DP2-GM-119137), the National Science Foundation (MCB-1914989 and MCB-1146621) and the Simons Foundation and Howard Hughes Medical Institute (55108535) awarded to M.C.J.; a German Academic Exchange Service (DAAD) research fellowship awarded to F.F.; Simons Foundation fellowships of the Life Sciences Research Foundation awarded to R.E.J. and J.V.-B.; an EMBO long-term fellowship (ALTF 1450-2014 and ALTF 563-2013) awarded to J.V.-B. and S.R.; an National Science Foundation MCB Grant (1818383) awarded to M.O.; a Swiss National Science Foundation Advanced PostDoc Mobility Fellowship (P2GEP3_148531) awarded to S.R.; and a Simons Foundation and Howard Hughes Medical Institute (55108515) Faculty Scholars grant awarded to J.R.D. S.W. was supported by the U.S. Department of Energy, Office of Science, Basic Energy Sciences, Chemical Sciences, Geosciences, and Biosciences Division under field work proposal 449B. K.K.N. is an investigator at the Howard Hughes Medical Institute. Work in the Merchant laboratory is supported by a cooperative agreement with the US Department of Energy Office of Science, Office of Biological and Environmental Research program under award DE-FC02-02ER63421.
Extended data
Source data
Unprocessed western blots.
Unprocessed western blots.
Author contributions
S.W. and K.K.N. performed rose bengal treatments; R.G.K., Y.K. and A.R.G. performed anoxia and high light treatments; J.V.-B., F.F. and R.E.J. prepared mutant pools, performed all other treatments and processed all samples; S.R.C. provided the LATCA compounds; M.M., J.O., C.P. and M.N. validated the active LATCA compounds; M.M. performed chemoinformatics analysis; M.O. validated LatB phenotypes in Chlamydomonas and performed immunoblots; S.R. and P.W. validated the mars1 phenotype; W.P. guided statistical analysis and website development; P.A.S. and S.S.M. provided transcriptomics data; X.L. provided early access to the Chlamydomonas mutant library; J.V.-B. and J.R.D. confirmed Arabidopsis phenotypes; J.V.-B., F.F., M.C.J. and R.E.J. designed experiments and analyzed and interpreted the data; and J.V.-B., F.F., M.C.J. and R.E.J. wrote the manuscript, with input from all authors.
Peer review
Peer review information
Nature Genetics thanks Jun Minagawa, Jean-David Rochaix and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Peer reviewer reports are available.
Data availability
Mutant barcode read count data (Supplementary Table 12) and mutant phenotypes across all screens (Supplementary Table 4) can be found at 10.6086/D1Q96Z in the Dryad Digital Repository. Source data are provided with this paper.
Code availability
Custom code used for data analysis has been deposited in Zenodo (10.5281/zenodo.6340170)77. Source data are provided with this paper.
Competing interests
J.V.-B., F.F., M.C.J. and R.E.J. note that a provisional patent application (US 63/123,422) on aspects of these findings has been submitted to the USPTO. The other authors declare no competing interests.
Footnotes
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
These authors contributed equally: Friedrich Fauser, Josep Vilarrasa-Blasi.
Contributor Information
José R. Dinneny, Email: dinneny@stanford.edu
Martin C. Jonikas, Email: mjonikas@princeton.edu
Robert E. Jinkerson, Email: robert.jinkerson@ucr.edu
Extended data
is available for this paper at 10.1038/s41588-022-01052-9.
Supplementary information
The online version contains supplementary material available at 10.1038/s41588-022-01052-9.
References
- 1.Bassham JA, et al. The path of carbon in photosynthesis. XXI. The cyclic regeneration of carbon dioxide acceptor. J. Am. Chem. Soc. 1954;76:1760–1770. doi: 10.1021/ja01636a012. [DOI] [Google Scholar]
- 2.Deisenhofer J, Epp O, Miki K, Huber R, Michel H. X-ray structure analysis of a membrane protein complex. Electron density map at 3 A resolution and a model of the chromophores of the photosynthetic reaction center from Rhodopseudomonas viridis. J. Mol. Biol. 1984;180:385–398. doi: 10.1016/S0022-2836(84)80011-X. [DOI] [PubMed] [Google Scholar]
- 3.Levine RP. Genetic dissection of photosynthesis. Science. 1968;162:768–771. doi: 10.1126/science.162.3855.768. [DOI] [PubMed] [Google Scholar]
- 4.Mayfield SP, Rahire M, Frank G, Zuber H, Rochaix JD. Expression of the nuclear gene encoding oxygen-evolving enhancer protein 2 is required for high levels of photosynthetic oxygen evolution in Chlamydomonas reinhardtii. Proc. Natl Acad. Sci. USA. 1987;84:749–753. doi: 10.1073/pnas.84.3.749. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Hillenmeyer ME, et al. The chemical genomic portrait of yeast: uncovering a phenotype for all genes. Science. 2008;320:362–365. doi: 10.1126/science.1150021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Harris, E. H., Stern, D. B. & Witman, G. B. The Chlamydomonas Sourcebook: Introduction to Chlamydomonas and Its Laboratory Use (Elsevier Science, 2009).
- 7.Ostrowski LE, Dutcher SK, Lo CW. Cilia and models for studying structure and function. Proc. Am. Thorac. Soc. 2011;8:423–429. doi: 10.1513/pats.201103-027SD. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Lefebvre PA, Rosenbaum JL. Regulation of the synthesis and assembly of ciliary and flagellar proteins during regeneration. Annu. Rev. Cell Biol. 1986;2:517–546. doi: 10.1146/annurev.cb.02.110186.002505. [DOI] [PubMed] [Google Scholar]
- 9.Luck DJ. Genetic and biochemical dissection of the eucaryotic flagellum. J. Cell Biol. 1984;98:789–794. doi: 10.1083/jcb.98.3.789. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Park RV, Asbury H, Miller SM. Modification of a Chlamydomonas reinhardtii CRISPR/Cas9 transformation protocol for use with widely available electroporation equipment. MethodsX. 2020;7:100855. doi: 10.1016/j.mex.2020.100855. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Greiner A, et al. Targeting of photoreceptor genes in Chlamydomonas reinhardtii via zinc-finger nucleases and CRISPR/Cas9. Plant Cell. 2017;29:2498–2518. doi: 10.1105/tpc.17.00659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Li X, et al. A genome-wide algal mutant library and functional screen identifies genes required for eukaryotic photosynthesis. Nat. Genet. 2019;51:627–635. doi: 10.1038/s41588-019-0370-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Zhao Y, et al. Chemical genetic interrogation of natural variation uncovers a molecule that is glycoactivated. Nat. Chem. Biol. 2007;3:716–721. doi: 10.1038/nchembio.2007.32. [DOI] [PubMed] [Google Scholar]
- 14.Ashburner M, et al. Gene ontology: tool for the unification of biology. The Gene Ontology Consortium. Nat. Genet. 2000;25:25–29. doi: 10.1038/75556. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Perlaza K, et al. The Mars1 kinase confers photoprotection through signaling in the chloroplast unfolded protein response. Elife. 2019;8:e49577. doi: 10.7554/eLife.49577. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Salomé PA, Merchant SS. Co-expression networks in Chlamydomonas reveal significant rhythmicity in batch cultures and empower gene function discovery. Plant Cell. 2021;33:1058–1082. doi: 10.1093/plcell/koab042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Hakem R. DNA-damage repair; the good, the bad, and the ugly. EMBO J. 2008;27:589–605. doi: 10.1038/emboj.2008.15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Vlcek D, Sevcovicova A, Sviezena B, Galova E, Miadokova E. Chlamydomonas reinhardtii: a convenient model system for the study of DNA repair in photoautotrophic eukaryotes. Curr. Genet. 2008;53:1–22. doi: 10.1007/s00294-007-0163-9. [DOI] [PubMed] [Google Scholar]
- 19.Culligan K, Tissier A, Britt A. ATR regulates a G2-phase cell-cycle checkpoint in Arabidopsis thaliana. Plant Cell. 2004;16:1091–1104. doi: 10.1105/tpc.018903. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Heitzeberg F, et al. The Rad17 homologue of Arabidopsis is involved in the regulation of DNA damage repair and homologous recombination. Plant J. 2004;38:954–968. doi: 10.1111/j.1365-313X.2004.02097.x. [DOI] [PubMed] [Google Scholar]
- 21.Garcia V, Salanoubat M, Choisne N, Tissier A. An ATM homologue from Arabidopsis thaliana: complete genomic organisation and expression analysis. Nucleic Acids Res. 2000;28:1692–1699. doi: 10.1093/nar/28.8.1692. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Feng W, et al. Genetic determinants of cellular addiction to DNA polymerase theta. Nat. Commun. 2019;10:4286. doi: 10.1038/s41467-019-12234-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Plecenikova A, Slaninova M, Riha K. Characterization of DNA repair deficient strains of Chlamydomonas reinhardtii generated by insertional mutagenesis. PLoS One. 2014;9:e105482. doi: 10.1371/journal.pone.0105482. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Recker J, Knoll A, Puchta H. The Arabidopsis thaliana homolog of the helicase RTEL1 plays multiple roles in preserving genome stability. Plant Cell. 2014;26:4889–4902. doi: 10.1105/tpc.114.132472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Knoll A, et al. The Fanconi anemia ortholog FANCM ensures ordered homologous recombination in both somatic and meiotic cells in Arabidopsis. Plant Cell. 2012;24:1448–1464. doi: 10.1105/tpc.112.096644. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Hartung F, Suer S, Bergmann T, Puchta H. The role of AtMUS81 in DNA repair and its genetic interaction with the helicase AtRecQ4A. Nucleic Acids Res. 2006;34:4438–4448. doi: 10.1093/nar/gkl576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Perron K, Goldschmidt-Clermont M, Rochaix JD. A factor related to pseudouridine synthases is required for chloroplast group II intron trans-splicing in Chlamydomonas reinhardtii. EMBO J. 1999;18:6481–6490. doi: 10.1093/emboj/18.22.6481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Jacobs J, et al. Identification of a chloroplast ribonucleoprotein complex containing trans-splicing factors, intron RNA, and novel components. Mol. Cell. Proteom. 2013;12:1912–1925. doi: 10.1074/mcp.M112.026583. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Marx C, Wünsch C, Kück U. The octatricopeptide repeat protein Raa8 is required for chloroplast trans splicing. Eukaryot. Cell. 2015;14:998–1005. doi: 10.1128/EC.00096-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Schult K, et al. The nuclear-encoded factor HCF173 is involved in the initiation of translation of the psbA mRNA in Arabidopsis thaliana. Plant Cell. 2007;19:1329–1346. doi: 10.1105/tpc.106.042895. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Brzezowski P, et al. Mg chelatase in chlorophyll synthesis and retrograde signaling in Chlamydomonas reinhardtii: CHLI2 cannot substitute for CHLI1. J. Exp. Bot. 2016;67:3925–3938. doi: 10.1093/jxb/erw004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Eberhard S, et al. Dual functions of the nucleus-encoded factor TDA1 in trapping and translation activation of atpA transcripts in Chlamydomonas reinhardtii chloroplasts. Plant J. 2011;67:1055–1066. doi: 10.1111/j.1365-313X.2011.04657.x. [DOI] [PubMed] [Google Scholar]
- 33.Johnson X, et al. MRL1, a conserved Pentatricopeptide repeat protein, is required for stabilization of rbcL mRNA in Chlamydomonas and Arabidopsis. Plant Cell. 2010;22:234–248. doi: 10.1105/tpc.109.066266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Liu XL, Yu HD, Guan Y, Li JK, Guo FQ. Carbonylation and loss-of-function analyses of SBPase reveal its metabolic interface role in oxidative stress, carbon assimilation, and multiple aspects of growth and development in Arabidopsis. Mol. Plant. 2012;5:1082–1099. doi: 10.1093/mp/sss012. [DOI] [PubMed] [Google Scholar]
- 35.Brandes HK, Larimer FW, Hartman FC. The molecular pathway for the regulation of phosphoribulokinase by thioredoxin f. J. Biol. Chem. 1996;271:3333–3335. doi: 10.1074/jbc.271.7.3333. [DOI] [PubMed] [Google Scholar]
- 36.Klein RR, Houtz RL. Cloning and developmental expression of pea ribulose-1,5-bisphosphate carboxylase/oxygenase large subunit N-methyltransferase. Plant Mol. Biol. 1995;27:249–261. doi: 10.1007/BF00020181. [DOI] [PubMed] [Google Scholar]
- 37.Wang Y, Stessman DJ, Spalding MH. The CO2 concentrating mechanism and photosynthetic carbon assimilation in limiting CO2: how Chlamydomonas works against the gradient. Plant J. 2015;82:429–448. doi: 10.1111/tpj.12829. [DOI] [PubMed] [Google Scholar]
- 38.Wang Y, Spalding MH. Acclimation to very low CO2: contribution of limiting CO2 inducible proteins, LCIB and LCIA, to inorganic carbon uptake in Chlamydomonas reinhardtii. Plant Physiol. 2014;166:2040–2050. doi: 10.1104/pp.114.248294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Moroney JV, et al. The carbonic anhydrase isoforms of Chlamydomonas reinhardtii: intracellular location, expression, and physiological roles. Photosynth. Res. 2011;109:133–149. doi: 10.1007/s11120-011-9635-3. [DOI] [PubMed] [Google Scholar]
- 40.Wang Y, Spalding MH. An inorganic carbon transport system responsible for acclimation specific to air levels of CO2 in Chlamydomonas reinhardtii. Proc. Natl Acad. Sci. USA. 2006;103:10110–10115. doi: 10.1073/pnas.0603402103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Fukuzawa H, et al. Ccm1, a regulatory gene controlling the induction of a carbon-concentrating mechanism in Chlamydomonas reinhardtii by sensing CO2 availability. Proc. Natl Acad. Sci. USA. 2001;98:5347–5352. doi: 10.1073/pnas.081593498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Xiang Y, Zhang J, Weeks DP. The Cia5 gene controls formation of the carbon concentrating mechanism in Chlamydomonas reinhardtii. Proc. Natl Acad. Sci. USA. 2001;98:5341–5346. doi: 10.1073/pnas.101534498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Itakura AK, et al. A Rubisco-binding protein is required for normal pyrenoid number and starch sheath morphology in Chlamydomonas reinhardtii. Proc. Natl Acad. Sci. USA. 2019;116:18445–18454. doi: 10.1073/pnas.1904587116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Meyer MT, et al. Assembly of the algal CO2-fixing organelle, the pyrenoid, is guided by a Rubisco-binding motif. Sci. Adv. 2020;6:eabd2408. doi: 10.1126/sciadv.abd2408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Kobayashi Y, et al. Eukaryotic components remodeled chloroplast nucleoid organization during the green plant evolution. Genome Biol. Evol. 2015;8:1–16. doi: 10.1093/gbe/evv233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Mackinder LCM, et al. A spatial interactome reveals the protein organization of the algal CO2-concentrating mechanism. Cell. 2017;171:133–147.e114. doi: 10.1016/j.cell.2017.08.044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Toyokawa C, Yamano T, Fukuzawa H. Pyrenoid starch sheath is required for LCIB localization and the CO2-concentrating mechanism in green algae. Plant Physiol. 2020;182:1883–1893. doi: 10.1104/pp.19.01587. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Findinier J, Delevoye C, Cohen MM. The dynamin-like protein Fzl promotes thylakoid fusion and resistance to light stress in Chlamydomonas reinhardtii. PLoS Genet. 2019;15:e1008047. doi: 10.1371/journal.pgen.1008047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Mackinder LCM, et al. A repeat protein links Rubisco to form the eukaryotic carbon-concentrating organelle. Proc. Natl Acad. Sci. USA. 2016;113:5958–5963. doi: 10.1073/pnas.1522866113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Yamano T, et al. Light and low-CO2-dependent LCIB–LCIC complex localization in the chloroplast supports the carbon-concentrating mechanism in Chlamydomonas reinhardtii. Plant Cell Physiol. 2010;51:1453–1468. doi: 10.1093/pcp/pcq105. [DOI] [PubMed] [Google Scholar]
- 51.Turkina MV, Blanco-Rivero A, Vainonen JP, Vener AV, Villarejo A. CO2 limitation induces specific redox-dependent protein phosphorylation in Chlamydomonas reinhardtii. Proteomics. 2006;6:2693–2704. doi: 10.1002/pmic.200500461. [DOI] [PubMed] [Google Scholar]
- 52.Zhan Y, et al. Pyrenoid functions revealed by proteomics in Chlamydomonas reinhardtii. PLoS One. 2018;13:e0185039. doi: 10.1371/journal.pone.0185039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Pazour GJ, Agrin N, Leszyk J, Witman GB. Proteomic analysis of a eukaryotic cilium. J. Cell Biol. 2005;170:103–113. doi: 10.1083/jcb.200504008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Awata J, et al. NPHP4 controls ciliary trafficking of membrane proteins and large soluble proteins at the transition zone. J. Cell Sci. 2014;127:4714–4727. doi: 10.1242/jcs.155275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Collin GB, et al. Meckelin is necessary for photoreceptor intraciliary transport and outer segment morphogenesis. Invest. Ophthalmol. Vis. Sci. 2012;53:967–974. doi: 10.1167/iovs.11-8766. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Craige B, et al. CEP290 tethers flagellar transition zone microtubules to the membrane and regulates flagellar protein content. J. Cell Biol. 2010;190:927–940. doi: 10.1083/jcb.201006105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Liu P, Lechtreck KF. The Bardet-Biedl syndrome protein complex is an adapter expanding the cargo range of intraflagellar transport trains for ciliary export. Proc. Natl Acad. Sci. USA. 2018;115:E934–E943. doi: 10.1073/pnas.1713226115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Mitchison HM, et al. Mutations in axonemal dynein assembly factor DNAAF3 cause primary ciliary dyskinesia. Nat. Genet. 2012;44:381–389. doi: 10.1038/ng.1106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Hom EF, et al. A unified taxonomy for ciliary dyneins. Cytoskeleton (Hoboken) 2011;68:555–565. doi: 10.1002/cm.20533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Kamiya R. Mutations at twelve independent loci result in absence of outer dynein arms in Chylamydomonas reinhardtii. J. Cell Biol. 1988;107:2253–2258. doi: 10.1083/jcb.107.6.2253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Myster SH, Knott JA, O’Toole E, Porter ME. The Chlamydomonas Dhc1 gene encodes a dynein heavy chain subunit required for assembly of the I1 inner arm complex. Mol. Biol. Cell. 1997;8:607–620. doi: 10.1091/mbc.8.4.607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Kubo T, Yanagisawa H-A, Yagi T, Hirono M, Kamiya R. Tubulin polyglutamylation regulates axonemal motility by modulating activities of inner-arm dyneins. Curr. Biol. 2010;20:441–445. doi: 10.1016/j.cub.2009.12.058. [DOI] [PubMed] [Google Scholar]
- 63.Verdier P, Morthorst SK, Pedersen LB. Targeting of ASH domain-containing proteins to the centrosome. Methods Mol. Biol. 2016;1454:15–33. doi: 10.1007/978-1-4939-3789-9_2. [DOI] [PubMed] [Google Scholar]
- 64.Morton WM, Ayscough KR, McLaughlin PJ. Latrunculin alters the actin-monomer subunit interface to prevent polymerization. Nat. Cell Biol. 2000;2:376–378. doi: 10.1038/35014075. [DOI] [PubMed] [Google Scholar]
- 65.Spector I, Shochet NR, Kashman Y, Groweiss A. Latrunculins: novel marine toxins that disrupt microfilament organization in cultured cells. Science. 1983;219:493–495. doi: 10.1126/science.6681676. [DOI] [PubMed] [Google Scholar]
- 66.Onishi M, Pringle JR, Cross FR. Evidence that an unconventional actin can provide essential F-actin function and that a surveillance system monitors F-actin integrity in Chlamydomonas. Genetics. 2016;202:977–996. doi: 10.1534/genetics.115.184663. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Onishi M, Pecani K, Jones TT, Pringle JR, Cross FR. F-actin homeostasis through transcriptional regulation and proteasome-mediated proteolysis. Proc. Natl Acad. Sci. USA. 2018;115:E6487–E6496. doi: 10.1073/pnas.1721935115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Feldman RM, Correll CC, Kaplan KB, Deshaies RJ. A complex of Cdc4p, Skp1p, and Cdc53p/cullin catalyzes ubiquitination of the phosphorylated CDK inhibitor Sic1p. Cell. 1997;91:221–230. doi: 10.1016/S0092-8674(00)80404-3. [DOI] [PubMed] [Google Scholar]
- 69.Kropat J, et al. A revised mineral nutrient supplement increases biomass and growth rate in Chlamydomonas reinhardtii. Plant J. 2011;66:770–780. doi: 10.1111/j.1365-313X.2011.04537.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Rogers D, Hahn M. Extended-connectivity fingerprints. J. Chem. Inf. Model. 2010;50:742–754. doi: 10.1021/ci100050t. [DOI] [PubMed] [Google Scholar]
- 71.Bajusz D, Racz A, Heberger K. Why is Tanimoto index an appropriate choice for fingerprint-based similarity calculations? J. Cheminform. 2015;7:20. doi: 10.1186/s13321-015-0069-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Shannon P, et al. Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. 2003;13:2498–2504. doi: 10.1101/gr.1239303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Martin M. Cutadapt removes adapter sequences high-throughput sequencing reads. EMBnet.journal. 2011;17:10–12. doi: 10.14806/ej.17.1.200. [DOI] [Google Scholar]
- 74.Benjamini Y, Hochberg Y. Controlling the false discovery rate: a practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B Methodol. 1995;57:289–300. [Google Scholar]
- 75.Virtanen P, et al. SciPy 1.0: fundamental algorithms for scientific computing in Python. Nat. Methods. 2020;17:261–272. doi: 10.1038/s41592-019-0686-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Merchant SS, et al. The Chlamydomonas genome reveals the evolution of key animal and plant functions. Science. 2007;318:245–250. doi: 10.1126/science.1143609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.JinkersonLab. JinkersonLab/chlamy-screen: v1.0.0 first release. Zenodo https://zenodo.org/record/6340170#.YkMCYOfMKUk (2022).
- 78.Porra RJ, Thompson WA, Kriedemann PE. Determination of accurate extinction coefficients and simultaneous equations for assaying chlorophylls a and b extracted with four different solvents: verification of the concentration of chlorophyll standards by atomic absorption spectroscopy. Biochim. Biophys. Acta Bioenerg. 1989;975:384–394. doi: 10.1016/S0005-2728(89)80347-0. [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
List of supplementary materials
Spreadsheet with Supplementary Tables 1–3, 5–11 and 13–15
Zip file that contains the chemical structure of all LATCA compounds used in this study.
Zip file that contains the Java TreeView files of the FDR < 0.3 gene cluster.
Data Availability Statement
Mutant barcode read count data (Supplementary Table 12) and mutant phenotypes across all screens (Supplementary Table 4) can be found at 10.6086/D1Q96Z in the Dryad Digital Repository. Source data are provided with this paper.
Custom code used for data analysis has been deposited in Zenodo (10.5281/zenodo.6340170)77. Source data are provided with this paper.